國立臺灣大學生命科學學院植物科學研究所 碩士論文
Institute of Plant Biology College of Life Science
National Taiwan University Master Thesis
鈣離子依賴磷酸激脢參與在植物先天性免疫反應以及 荷爾蒙相關防禦反應之功能性分析
Functional Characterization of CPK7 in Pattern-Triggered Immunity and Hormone-Related Plant Defense
陳太一 Tai-I Chen
指導教授:金洛仁 博士 Advisor: Laurent Zimmerli, Ph.D.
中華民國 105 年 1 月
January 2016
i
誌謝
真正完成口試之前,總好傻好天真的以為、當你/我看到誌謝這兩個字出現的 同時,一個嶄新的生命已經鋪天蓋地的在我面前展開!然而十分遺憾的(殘念!是 鼬!),心裡獲得的舒坦與成就感、至多像是下午一點二十清理完垃圾回收的職日 生,連插完 tip 的輪值都比不上──大概就是一種似甘蔗渣的無聊空虛吧,啊。不 就跟這個學位一樣嗎?開玩笑的ㄎㄎ。然而,就算主觀上最終綜合如此,實踐 上,屏除一些未被察覺目的以,客觀還是有許多值得感覺、或者感謝的事。
首先當然要感謝指導老師,金洛仁老師,非常非常感謝。如果沒有他,我們 怎麼可能知道這份學位真正的價值呢?這種獨一無二的經驗,貨真價實的人生歷 練,都是大學時期被專題老師各種呵護甚至吹捧的我無從知曉的。如果能夠與麥
克阿瑟分享這種種心路歷程,為子祈禱文大概就變成了:「主啊!請讓我的孩子
滿懷對生命科學的熱愛加入金洛仁老師的實驗室。」不論如何,身為敦厚老實的 呆灣郎,還是非常感謝老師許多方面的指導,尤其最後改論文以及人生方面;對 我而言,最後這份學位實質上更豐沛的、是管理學院的內涵。千言萬語,不及一 句感謝,就像老鼠愛大米、南無阿密陀佛。
在實驗室的兩年時間絕對是大學時難以想像的。前一年,敏原、子娟、
Cecile、郁欣、Jimmy、奕睿,還有鈺鴻、堯堯兩個神主牌,除了喜愛之外,更多 的是如火一般的尊敬。碩二開始覺得可以恣意對待他人了,小麥、貝爾、安博,
你們的出現實在恰如其分(就像你們在彼此的關係當中各司其職,科科),雖然對 我意義各有不同、但沒想到自己也可以這樣喜歡接近別人呢。
對於泰元、靜松、Eric,當然還有友慧,謝謝你們。因為我不是暴露狂,所 以一如往常,對於最重要的你們、這些最重要的點滴我會一字不提。封入箱子,
深深地埋在白茫茫的雲海裡。謝謝你們。
感謝總統卡神、James,還有口桀,有你們在身邊、讓我時時刻刻鞭策自 己、活著不僅僅是活著。家銘、大凱,堉嘉;資質、變態,猴子。這些奇妙的三 位一體,緊密連結著彼此,難道不就是足底根植的、無與倫比的蘭陽平原還有龜 山島?嘿!我們不需要說什麼謝不謝的吧!
在掙扎之中,我還是想感謝 Liebe Gloyia;還有一起走過、度過好多、最可 愛特別的時鐘發明者,願有一天都能光著腳、快樂的走跳於開滿小白花的草原 上。還有 CPK7,謝謝你的陪伴,也希望我夠謹慎堅強、沒有讓你因愚昧疏忽等 垃圾人類的垃圾理由被錯誤的理解。
Lieber Hermann Hesse、Heine、Kant、Beethoven、Bartok、Wagner、
Shostakovich、Furtwängler Coltrane und Schubert. Du hast mich in eine beßre Welt entrückt. Ich danke dir dafür.
還有我最最親愛的家人,媽媽、爸爸還有姊姊,辛苦了。沒有你們我甚至不 會是個完整的人。
ii 摘要
在植物面對各式生物性或非生物逆境時、細胞內鈣離子累積為普遍且廣泛的訊息
傳遞。在阿拉伯芥有一群包含 34 成員的鈣離子依賴蛋白磷酸激脢家族,同時具有
磷酸激脢以及與鈣離子結合的能力,以此參與鈣離子訊息傳遞。其中鈣離子依賴蛋
白磷酸激 7 與膜受體感受病原相關分子模式激活的免疫反應(PTI)正調控子 LecRK
VI.2 可能有共表現之現象,對於典型 PTI 機制如 MAPK 磷酸化以及癒傷葡聚醣
(callose)累積負調控。更多證據顯示,鈣離子依賴蛋白磷酸激 7 亦參與在荷爾蒙相
關的、水楊酸防禦反應、茉莉酸-乙烯防禦反應,例如突變株對於活體營養性病原
菌具有抗性但對於死體營養病原菌更感病,荷爾蒙標誌基因分析水楊酸途徑相對
應的 PR1 有更高的表現,而茉莉酸-乙烯途徑標誌基因 PDF1.2 則相對低,顯示了
此基因可能會影響兩個重要植物荷爾蒙平衡;另外,PEPR 抗病途徑亦於本基因的
突變株當中亦顯示出了更佳之活性。綜合以上所述,在植物抗病反應當中,CPK7
負向調控了 PTI 並且影響 PEPR 抗病途徑與荷爾蒙相關防禦反應。
關鍵字:鈣離子依賴蛋白磷酸激、 細菌性斑點病病原菌、細菌性軟腐病病原菌、
灰黴病病原菌、PAMP 誘發免疫反應、水楊酸防禦反應、茉莉酸-乙烯防禦反應、
PEPR 抗病途徑。
iii Abstract
Calcium ions (Ca2+) play an essential and general role as secondary messengers in many
cellular signaling pathways. In Arabidopsis, Ca2+-dependent protein kinases (CPKs), a
family of 34 members, are able to sense and to response to changes in calcium
concentrations through their calcium binding ability and kinase activity. Among these
CPKs, CPK7 was selected as a putative co-expressed gene with LecRK-VI.2, a positive
regulator of PAMP-triggered Immunity (PTI). In this work, we show that CPK7
negatively regulates typical PTI responses such as callose deposition and MAPK kinase
phosphorylation. Concomitantly, knock-out mutant lines of CPK7 were more resistant to
the hemibiotrophic pathogen Pseudomonas syringae pv. tomato DC3000 (Pst DC3000).
By contrast, cpk7 mutants were more susceptible to the necrotrophic pathogens
Pectobacterium carotovorum ssp. carotovorum (Pcc) and Botrytis cinerea. The cpk7
mutants also demonstrated a potentiated accumulation of PR1 mRNA upon Pst DC3000
infection and less PDF1.2 up-regulation after Pcc inoculation, indicating that CPK7 may
also affect hormone responses of plant defense. Furthermore, CPK7 also negatively
regulates the (full name needed here) PEPR pathway. These results suggest that in
response to calcium signaling triggered by PTI, CPK7 modulates hormone homeostasis
and PEPR pathway activity, affecting plant defense to biotic stresses.
iv
Keywords: Ca2+-dependent protein kinases, Pseudomonas syringae, Pectobacterium
carotovorum ssp. carotovorum, Botrytis cinerea, PAMP-triggered immunity (PTI), SA-
dependent pathway, JA/ET-dependent pathway, PEPR pathway.
v Abbreviations
CPK Calcium-dependent Protein Kinase
PAMPs Pathogen-associated molecular patterns
PRRs Pattern-recognition receptors
PTI PAMPs-triggered immunity
SA Salicylic acid
JA Jasmonic acid
ET Ethylene
DAMP Danger-Associated Molecular Pattern
DPI DAMPs-triggered immunity
FRK1 FLG22-INDUCED RECEPTOR-LIKE 1
NHL10 NDR1/HIN1-like 10
PR1 PATHOGENESIS-RELATED PROTEIN 1
PDF1.2 PLANT DEFENSIN1.2
PROPEP1 PRECURSOR OF PEPTIDE 1
FLS2 FLAGELLIN-SENSITIVE 2
BAK1 BRI1-ASSOCIATED LINASE 1
BIK1 BOTRYTIS-INDUCED KINASE1
PEPR1 PEP receptor 1
Pst DC3000 Pseudomonas syringae pv. tomato DC3000
Pcc Pectobacterium carotovorum ssp. carotovorum
6
Content
誌謝 ... i
摘要 ... ii
Abstract ... iii
Abbreviations ... v
Introduction ... 8
Pattern-Triggered Immunity ... 8
SA-, JA/ET-dependent defense and PEPR pathway ... 10
CPKs and CPK7 ... 12
Material and Methods ... 15
Plant Materials and Growth Conditions ... 15
Pathogen Infection Assay ... 15
Oxidative Burst Kinetic Assay ... 16
Stomatal Assay ... 17
Callose Staining Assay ... 17
MAPK Phosphorylation Assay ... 18
PAMP Treatment ... 19
RNA Extraction and qRT-PCR ... 20
Subcellular Localization ... 21
Bimolecular Fluorescence Complementation Assay in Arabidopsis Protoplast... 22
Results ... 24
Enhanced resistance to hemi-biotrophic bacteria in Arabidopsis cpk7 mutants... 24
The cpk7 mutants generate WT-level of reactive oxygen species burst upon flg22 perception ... 24
CPK7 negatively modulates PTI-mediated callose deposition ... 25
CPK7 is not critical for stomatal innate immunity ... 25
Higher MPK3/MPK6 phosphorylation level is observed in cpk7 mutants after flg22 treatment ... 26
PTI marker genes up-regulation is comparable between WT and cpk7 mutants ... 27
cpk7 mutants demonstrate a susceptible phenotype to necrotrophic Pcc and B. cinerea infections .. 27
Expression of PR1 is potentiated in cpk7 mutants ... 28
cpk7 mutants accumulate less PDF1.2 transcripts upon Pcc infiltration but normal level upon B. cinerea infection ... 29
CPK7 negatively regulates FRK1 expression upon pep1 treatment but not PROPEP1 expression .. 30 CPK7 is localized on the plasma membrane and nucleus and localization is affected by flg22
7
treatment ... 31
CPK7 does not associate with FLS2, BAK1, BIK1 and PEPR1 ... 31
Discussion ... 33
CPK7 plays a role in the Arabidopsis PTI as a negative regulator. ... 33
CPK7 modulates SA-dependent and JA/SA-dependent defense and CPK7 negatively regulates PEPR pathway. ... 34
CPK7 acts downstream of PTI, modulating several defense pathways ... 35
Conclusion and future perspectives ... 37
Figure ... 38
Figure 1: CPK family in Arabidopsis... 38
Figure 2: The putative structure of CPK7 and cpk7 mutants. ... 39
Figure 3. Disease symptoms and bacterial titers of Pst DC3000 infected Col-0 and cpk7 mutant lines ... 41
Figure 4. ROS production after flg22 treatment. ... 42
Figure 5. Visualizations and quantifications of callose deposits upon flg22 treatment ... 45
Figure 6: CPK7 in stomatal innate immunity. ... 46
Figure 7: MAPK kinase Assay ... 48
Figure 8. Transcriptional expression of PTI-responsive genes FRK1 and NHL10 after flg22 treatment. ... 49
Figure 9. Disease symptoms and bacterial titers of Pcc infected Col-0 and cpk7 mutant lines ... 51
Figure 10: Disease symptoms and lesion perimeter of B. cinerea infected Col-0 and cpk7 mutant lines ... 53
Figure 11: Transcriptional expression of the SA-dependent pathway marker gene PR1 after Pst DC3000 infiltration. ... 55
Figure 12: Transcriptional expression of the JA/ET-dependent pathway marker gene PDF1.2 after Pcc and B. cinerea infiltration. ... 57
Figure 13: Transcriptional expression of FRK1 and PROPEP1 upon pep1 treatment. ... 59
Figure 14: Subcellular localization of CPK7. ... 60
Figure 15: Association of CPK7 with FLS2, BAK1, BIK1 and PEPR1 could not be observed. ... 62
Supplemental table S1. Putative Co-expressing Gene of LecRK VI.2 ... 63
Supplemental table S2. List of Primers ... 63
Supplemental Figure S1: Transcriptional expression of ERF1 upon ACC treatment. ... 64
References ... 65
8 Introduction
Pattern-Triggered Immunity
To survive, plants had to adapt to be able to resist to numerous stresses from the
harsh environment they are living in. To resist biotic stresses such as bacterial or fungal
pathogens attack, plants has developed several adaptive mechanisms (Van Verk et al.,
2009). Among these molecular defense mechanisms, the very first one during plant-
pathogen interaction is plant innate immunity. Like those of animal immune systems,
plant innate immunity is defined as recognition and reaction to pathogenic or non-
pathogenic microbes (Boller, 2009). In more details, when microbes approach a plant
cell, some molecular patterns such as flagellin and elongation factor Tu are passively
released. These microbial signatures, termed pathogen-associated molecular patterns
(PAMPs), are then captured and recognized by plant protein receptors localized on the
surface of plant cells. Recognition will lead to the activation of downstream defence
signaling events (Jones and Dangl, 2006). Plant receptor proteins are termed pattern-
recognition receptors (PRRs) and the whole pattern-recognition phenomenon is further
defined as pattern triggered immunity (PTI) (Ausubel, 2005; Boller and Felix, 2009).
PTI events include accumulation of reactive oxygen species (ROS), stomatal
closure and callose deposition (Boller and Felix, 2009). These defense responses either
create chemical stresses or physical barriers, enabling host to successfully generate a
9
basal resistance against invading pathogens such as hemibiotrophic bacterial pathogen
Pseudomonas syringae pv. tomato (Pst) and necrotrophic Pectobacterium carotovorum
ssp. carotovorum (Pcc) (Singh et al., 2012). Furthermore, PTI is also accompanied with
an intensive transcriptional change. Former studies have found two pathways mediating
near membrane signals and transcriptome. First, mitogen-activated protein kinase
(MAPK) cascade, which takes advantage on phosphate groups transferring through
proteins with different energy levels (Dong et al., 2002). The other one is ion influx
(Boudsocq et al., 2010), such as calcium oscillation, created by membrane-localized
calcium channel, sensed by cytosolic calcium sensing proteins, such as calcium
dependent protein kinases (CPKs) (DeFalco et al., 2010; Sanders et al., 2002). PTI-
responsive genes such as FRK1 and NHL10 will be up-regulated, depending on these
underlying signal transductions after PRRs recognition of PAMPs (Zipfel et al. 2004).
In Arabidopsis thaliana, a well-studied model for PTI includes the PRR
FLAGELLIN-SENSITIVE 2 (FLS2) and the co-receptor BRI1-ASSOCIATED
RECEPTOR KINASE 1 (BAK1). In this model, the PRR FLS2 perceives PAMPs such
as bacterial flagellin fragments and then associates with BAK1, another membrane
LRR-PK, then with cytosolic cofactor such as BRASSINOSTEROID INSENSITIVE1
(BIK1), this kinase complex is able to amplify the PTI signals and triggers downstream
PTI responses (Gomez-Gomez, 2001; Chinchilla et al., 2009). However, besides FLS2
10
and BAK1, in a previous study, the Zimmerli laboratory discovered the Lectin Receptor
Kinase-VI.2 (LecRK-VI.2), which serves as a positive regulator of PTI (Singh et al.,
2012). Over-expression lines of LecRK-VI.2 show constitutive PTI gene activation and
callose deposition with higher resistance to pathogen in comparison to wild-type. These
evidences suggest that in addition of the recognition ofPAMPs by PRRs, there should
be more players with more different kinds of patterns, involved in the PTI response.
SA-, JA/ET-dependent defense and PEPR pathway
Given the fact that the infection of pathogen occurs during a long period of time,
on several tissues and with multiple infectious strategies, a great challenge to plants is
to maintain the energy homeostasis during activating defense responses (Glazebrook,
2005; Pieterse et al., 2009). With this aspect, signaling crosstalk between major defense-
related hormones is necessary. Current thinking is that salicylic Acid (SA) induces
resistance against biotrophic pathogens due to programmed cell death in the host
(Pieterse et al., 2009). However, since necrotrophic pathogens even benefit from host
cell death, defense responses requiring jasmonic acid (JA) and ethylene (ET) signaling
will be activated in this situation (Bari and Jones, 2009). For SA-dependent signaling
pathway, in response to pathogen attack, SA INDUCTION DEFICIENT 2 (SID2) and
PHYTOALEXIN DEFICIENT 4 (PAD4) act upstream of SA to promote SA
accumulation (Feys et al., 2001). These SA signals will be perceived by the SA receptor
11
NON-EXPRESSOR OF PR1 (NPR1) to further promote the expression of downstream
associated genes such as PR1 and other R-genes (Pieterse et al., 2012). On the other
hand, after detecting pathogen infections and tissue damage, JA levels increase locally,
and its chemical derivate JA-Isoleucine (JA-Ile) is recognized by CORONATINE
INSENSITIVE1 (COI1), enhancing the expression of PLANT DEFENSIN1.2 (PDF1.2).
(rewrite this sentence as it is not possible to understand what you want to say here…),
suggesting that JA and ET contribute simultaneously to this pathway (Feys et al., 1994;
Yan et al., 2009).
Based on several pathological and molecular evidences, SA and JA/ET signaling
pathways are believed to be antagonistic (de Torres et al., 2009). For example, mpk4
mutant has strong deficiency of PDF1.2 expression while having constitutive activation
of SA-dependent pathway (Peterson, 2000). However, several studies suggest that the
regulation between these two pathways is actually more complex than just simply
antagonistic. In leaves of (abbreviation defined before?)WT Arabidopsis applied with
Arabidopsis danger peptides (AtPeps), both transcriptional levels of PR1 and PDF1.2
will be induced. This evidence suggests the existence of at least one circumstance when
SA-dependent and JA/ET dependent pathway can be equally activated (Ross et al.,
2014). Recently, more information about these endogenous peptides and their functions
in plant defense were revealed. Similar as many other endogenous danger-associated
12
molecular patterns (DAMPs), AtPeps are passively induced under conditions of biotic
stress, such as microbial infections, the detection of MAMPs such as flg22, MeJA and
ET treatment, then recognized by some leucine-rich repeat (LRR) receptor kinases such
as Pep Receptors (PEPRs), to initiate intracellular signaling and later activating the
defense responsible genes and expression of propeptides themselves (Yamaguchi et al.,
2010; Huffaker and Ryan, 2007). This mechanism is defined as the PEPR pathway. The
PEPR pathway mainly leads to two different categories of defense systems. The
application of AtPEPs triggers PTI-like responses, such as callose deposition and ion
influx (Bartels et al., 2013). Moreover, application of AtPeps will cause the
accumulation of JA and induce both SA-dependent and JA/ET dependent pathway and
consequently enhances plant resistance to pathogen (Ross et al., 2014).
CPKs and CPK7
In plant, calcium is a ubiquitous second messenger,that mediates many different
environmental stress responses (Boudsocq et al., 2010). In normal condition, calcium
is maintained in the cytosol at very low level (100-200 nM) and accumulates in a very
high level during stimulation (Downie, 2014). To decipher the amplitude, duration and
frequency of cytosolic calcium oscillation created by different stresses, calcium sensors,
such as calcium-dependent protein kinases (CPKs) are able to react in response to
calcium concentration by direct binding to calcium ion (Romeis and Herde, 2014). The
13
basic structure of CPKs consists of a variable N-terminal domain, a conserved
serine/threonine kinase domain, an autoinhibitory junction region and a C-terminal
regulatory CaM-like domain (CamLD) (Christodoulou et al., 2004). N-terminal domain
often contains predicted acylation sites, and autoinhibition inactivates kinase activity
by a pseudosubstrate association. The CamLD domain contains four EF-hand calcium
binding motifs. Recent study on protein structure demonstrates the activation of CPKs.
In normal condition, autoinhibition domain attaches to the kinase activation site as a
pseudosubstrate thus inactivated the kinase. Upon rising Ca2+ level, autoinhibition
region releases from kinase active site through conformational change by Ca2+ binding
(Christodoulou et al., 2004; Weljie and Vogel, 2004). In Arabidopsis, CPKs consists of
a large family of 34 genes and are divided into four groups (Fig. 1) (Boudsocq and
Sheen, 2013; Chen, et al., 2002). Many of the members from the CPK family have
already been identified to be involved in biotic stress signaling pathways, either with
stimulus-dependent biochemical reactions on transcriptome alternation or alternation
of cellular phosphorylation pattern (Boudsocq and Sheen, 2013). For example,
overexpression of CPK1 induces a broad-spectrum Arabidopsis resistance to bacteria
and fungi by triggering SA accumulation through the induction of SA regulatory and
biosynthesis genes such as PAD4 and SID2 (Coca, 2010). And for Ca2+ spiking upon
herbivore attack AtCPK3 and AtCPK13 serve as positive regulators of PDF1.2
14
induction upon caterpillars attack (Kanchiswamy, 2010). CPKs are also known to
function in PTI signaling (Gao et al., 2014). Notably, CPKs together with MAPKs are
critical for the activation of targeted transcription factors and PTI-responsive genes. For
instance, AtCPK4, AtCPK5, AtCPK6 and AtCPK11 positively regulate NHL10
synergistically with MAPK cascade upon flg22 treatment (reviewed by Boudsocq and
Sheen, 2013).
Members of Zimmerli laboratory have identified several genes that are putatively
co-expressed with the positive regulator of the PTI response LecRK-VI.2. Among them
(Table S1), CPK7 was selected for further research. Some in vitro biochemical assays
have already been performed on CPK7. With these evidences, CPK7 is proposed to be
an active kinase and is capable of Ca2+ binding. However its kinase activity is
independent of calcium concentration (Boudsocq, 2012). Further studies with a T-DNA
insertion CPK7 knock-out indicates that CPK7 negatively regulates aquaporin activity
by repressing cellular PIP2 expression levels in root (Li et al., 2014). In addition, CPK7
along with BIK1 is reported to be phosphorylated by flg22 treatment in vivo (Li et al.,
2015). Here, by applying reverse-genetic approach with two knockout mutant lines
termed cpk7 36-1 and cpk7 36-2, we suggest that CPK7 acts downstream of PTI,
modulating SA-dependent, JA/ET dependent plant defense and PEPR pathway, but the
certain role of how CPK7 plays, is remain unclear.
15 Material and Methods
Plant Materials and Growth Conditions
Arabidopsis ecotype Col-0 plants were grown in commercial potting soil/perlite
(3:2) at 22°C to 24°C day and 17°C to 19°C night temperature under a short day (9-h- light/15-h-dark) photoperiod. The lighting is supplied at an intensity of ∼100 μE m−2s−1
by fluorescence tubes. T-DNA insertion mutant cpk7 36-1 (SALK_035601) and cpk7
36-2 (SALK_127223) were obtained from the Arabidopsis Biological Resource Center
(ABRC). Bacterial Pathogens Pst DC3000 and Pcc were cultivate at 28◦C and 220 rpm
in King’s B medium containing 50mg/mL rifampicin or 100 mg/mL ampicillin,
respectively. The fungus B. cinerea was grown at room temperature (18°C ~ 25°C) with
high humidity on PDB-agar plates for 2 to 3 weeks as described (Zimmerli et al., 2001).
Pathogen Infection Assay
For bacterial pathogen, five-week-old Arabidopsis plants were dipped in a
bacterial suspension of 2 x 107 colony-forming units (CFU)/mL Pst DC3000 or 106
CFU/mL Pcc in 10 mM MgSO4 containing 0.01% Silwet L-77 (Lehle Seeds) for 15
min. After dipping, plants were kept at 100% relative humidity overnight. Disease
symptoms were evaluated at 3 days post inoculation (dpi). For bacterial titers, leaf discs
16
collected at 2 dpi were washed twice with sterile water and homogenized in 10 mM
MgSO4. Quantification was done by plating appropriate dilutions on King’s B agar
containing rifampicin (50mg/liter) as previously described (Zimmerli et al., 2000).
Each biological repeat represents nine leaf discs (0.5 cm diameter) from three different
plants.
For fungal pathogen infection, spores of B. cinerea were suspended in PDB at a
final concentration of 105 spores/mL. Eight μL droplets of inoculum were deposited on
4 leaves of 5-week-old plants (8 plants for each genotypes). Pictures of lesions were
captured 2 to 3 days after inoculation, and lesion perimeters were evaluated by using
the ImageJ software (http://rsb.info.nih.gov/ij/).
Oxidative Burst Kinetic Assay
Reactive Oxygen Species (ROS) evaluation was performed as previously
described (Keppler et al., 1989). Eight leaf discs of 0.25 cm2 from three 5-week-old
Arabidopsis plants were incubated in 100 μL ddH2O for 16 h in 96-well plates. Water
was then replaced by 100 μL of reaction solution containing 50 μM of luminol and 10
μg/mL of horseradish peroxidase (Sigma) supplemented with 10 nM of flg22 or water
only for the mock controls. ROS measurements expressed as means of RLU (Relative
Light Units) were measured immediately after adding the solution with a Centro
17
LIApcLB 692 plate luminometer (Berthold Technologies, Bad Wildbad, Germany).
ROS evaluation was performed at 2 min interval reading times for a period of 34 min.
Stomatal Assay
Plants were kept under light for at least 3 h to open stomata. Leaf peels were
collected from the abaxial side of fully expanded leaves and floated in stomatal buffer (10 mM MES-KOH, 30 mM KCl, pH 6.15) for 2.5 h under light (∼100 μE m−2 s−1).
After treatment with mock control (10 mM MgSO4) or 108 CFU/mL Pst DC3000 in 10
mM MgSO4, pictures of stomata were taken of random regions at various time points
using an Olympus BX51 microscope digital camera and application software DP2-
BSW. Stomatal apertures were measured using the “measure” function of ImageJ
(http://rsb.info.nih.gov/ij/). Three plants were used per biological replicates.
Callose Staining Assay
Leaves of 5-week-old Arabidopsis were syringe infiltrated 500 nM flg22 dissolved
in H2O. Control, mock-treated plants were infiltrated with H2O only. Twelve leaf discs
from three different plants were selected for analyses. Harvested leaf samples were
cleared overnight by incubation in 95% ethanol at room temperature and then washed
18
three times (0.5 h for each time) with sterilized H2O. Transparent leaves were stained
with 0.01% aniline blue in 0.15 M phosphate buffer, pH 8, for 1 h. Callose deposits
were visualized under UV illumination using an Olympus BX51 micro-scope digital
camera and application software DP2-BSW. Callose deposition was evaluated using the
“analyze particles” function of ImageJ (http://rsb.info.nih.gov/ij/).
MAPK Phosphorylation Assay
Nine leaves of 5-week-old Arabidopsis were syringe infiltrated with 100 nM flg22
dissolved in H2O. Control plants were infiltrated with H2O only. Samples were
collected after 10 min and immediately snap frozen in liquid nitrogen. The protein
concentration was determined using the BCA protein assay kit (Bio-red) with BSA as
a standard. Protein crude extracts were separated on 10% SDS gel, and proteins were
transferred to Polyvinylidene fluoride membrane (PVDF) by immunoblotting (Mini-
Protean II system; Bio-Rad). Activated MAPK3/MAPK6 were detected by overnight
incubation with anti-p42/44 MPK primary antibodies (1:3000 dilution, Cell Signaling
Technology), followed by incubation with anti-rabbit-HRP secondary antibodies (Sig-
ma-Aldrich) for 1 h. The signals were visualized using an enhanced chemiluminescence
system (Western Lightning Plus-ECL kit; Perkin-Elmer) following the manufacturer’s
instructions and LAS-3000 (Fujifilm).
19 PAMP Treatment
For determining FRK1 and NHL10 response to PAMPs, leaves of 5-week-old
Arabidopsis were syringe-infiltrated with 100 nM flg22 dissolved in H2O or ddH2O as
Mock control. Samples were collected at 5 h post infiltration. For PR1 expression
analysis, leaves were syringe-infiltrated with 108 CFU/mL Pst DC3000 or ddH2O only
as Mock control. Infiltrated leaves were collected at 5 h and 9 h post inoculation. For
PDF1.2 expression, leaves were syringe-infiltrated with Pcc 5 x 106 CFU/mL or
sprayed with spores from B. cinerea at a concentration of 105 spores/mL in PDB.
Infiltration of ddH2O or spraying with PDB was used as mock control respectively.
Plants were kept in high humidity sealed boxes for 24 h before samples collection. For
FRK1 and PROPEP1 expression, 10 of 10-day-old seedlings were growth on ½ MS
agar plates, and then moved to 6-well plate where each well contains ½ MS 6 mL for
recovering from possible wounding effect overnight, then for the experiment, directly
add additional pep1 to 50 nM each well and collect samples snap frozen in liquid
nitrogen every 1 h until 6 h.
20 RNA Extraction and qRT-PCR
Total RNA was extracted and purified using the MaestroZol reagent according to
the manufacturer’s instructions (Omics Biotechnology Co., Ltd) with the addition of
PLUS reagent for polysaccharides and proteoglycans elimination. Genomic DNA
contaminations were removed using Qiagen RNase-Free DNase Set. For cDNA
synthesis, 2 μg of total RNA were prepared in a volume of 22 μL DEPC-treated H2O
and denatured at 65°C for 5 min. Eighteen point five mL of master mix (16 μL of M-
MLV buffer, 1 μL of 10 mM dNTP, 1 μL of 100 mM OligoT, 0.5 μL of 100 U M-MLV
reverse transcriptase, [Invi-trogen]) was added into each tube and then incubated at
37°C for 1 h, and 70°C for 10 min. The cDNA was then diluted 5-times before
quantitative RT-PCR (qRT-PCR) analyses. The cycling conditions were 94°C for 3 min
for one initial step followed by 94°C for 30 s, 58°C for 30 s, and 72°C for 1 min, for 35
cycles. The PCR was terminated with one extra step at 72°C for 10 min. qRT-PCRs
were conducted on a CFX Real-Time PCR Detection System (Bio-Rad). SYBR Green
fast qPCR master mix (Bio-Rad; 1 μL of cDNA, 5 μL SYBR Green supermix, 5 μL
filtered sterile H2O, 0.5 μL of 10 mM forward and reverse primers, in a total volume
of 12 μL per well) was employed for the analysis. The cycling conditions were
composed of an initial 3 min denaturation step at 95°C, followed by 40 cycles at 95°C
for 3 s and 60°C for 30 s. Melting curves were run from 65°C to 95°C with 0.5-s time
21
interval to ensure the specificity of product. Data were analyzed using Bio-Rad CFX
manager software. UBQ10 was used as a reference gene for normalization of gene
expression levels in all samples. The wild-type (WT) without any treatment or mock
treatment were considered as controls in each experiment. All primers used for qRT-
PCR amplification are listed in supplemental table S1.
Subcellular Localization
The full-length CPK7 cDNA was PCR amplified from wide type Col-0 cDNAs
using the primers listed in Table S2, The PCR product was cloned into
pCR8/GW/TOPO vector (Invitrogen). The TOPO vector with full length genomic
LRR50 was then sequenced to ensure fidelity (Tri-I Biotech). After confirmation by
sequencing, TOPO-LRR50 was recombined into the Gateway-compatible destination
vector pEarlyGate 103 (obtained from ABRC) containing the 35S promoter and GFP-
6x His tag through LR reaction according to the manufacturer’s instructions
(Invitrogen). The recombination plasmid after the LR reaction was transformed into E.
coli strain DH5α competent cells (Omics Bio). The DH5α competent cells were then
incubated at 37℃ on LB agar plate with 100 ng/mL Kanamycin for selection. Colony
PCR was performed to screen for positive colonies using the forward primer 5'-
AAGGGTCTTGCGAAGGATAG -3' from the 35S promoter region and the reverse
22
primer 5'- CCGAGCCGTTGGGCTCAGT -3' from the LRR50 region. The positive E.
coli colonies were picked up and grown overnight in LB medium containing 100 ng/mL
kanamycin at 37˚C. The plasmids DNA were extracted by plasmid Mini kit (GeneMark).
Arabidopsis protoplasts were isolated and transfected by a polyethylene glycol method
as described previously (Yoo et al., 2007). Briefly, 0.1 mL of protoplast suspension
(200,000 cells mL-1) was transfected with a mixture of 4 μg of plasmid. After
transfection, protoplasts were incubated 16 h, and then samples were visualized using
a TCS SP5 confocal spectral microscope imaging system (Leica)
Bimolecular Fluorescence Complementation Assay in Arabidopsis Protoplast
For Bimolecular Fluorescence Complementation (BiFC) assay, plasmids
containing CPK7-YN, PEPR1-YN, YC-CPK7, YC-PEPR1 were constructed. PEPR1
CDS was PCR amplified from wide type Col-0 cDNA using the primers listed in Table
1 (name needed) and PCR products were then cloned into the pCR8 TOPO vector.
TOPO-cDNA plasmids were finally recombined into the Gateway-compatible
expression vectors pSAT4 (A)-DEST-n (1-174) EYFP-N1 (pE3134) (obtained from
Stanton B. Gelvin’s lab, Purdue University). Plasmids of CPK7-YN, CPK7-YC, FLS2-
YC, BAK1-YC, BAK1-Yn, BIK1-YC, PEPR1-YC or PEPR1-YN with different
combinations were transformed into Arabidopsis protoplasts by polyethylene glycol for
23
transient expression (Yoo et al., 2007). After 16 h incubation, samples were visualized
using a TCS SP5 confocal spectral microscope imaging system (Leica).
24 Results
Enhanced resistance to hemi-biotrophic bacteria in Arabidopsis cpk7 mutants
To evaluate whether CPK7, a gene co-expressed with the positive regulator of PTI
LecRK VI.2, the resistance response of cpk7 mutants to hemi-biotrophic bacteria was
evaluated. In comparison with WT Col-0, on the third day after challenged with Pst
DC3000, two cpk7 mutant alleles developed less necroses and bacteria-mediated leaf
yellowing (Fig. 3a). Similarly, bacterial titers were significantly lower in cpk7 mutants
than in Col-0 (Fig. 3b), suggesting that CPK7 acts as a negative regulator of
Arabidopsis defense to hemi-biotrophic pathogens such as Pst DC3000.
The cpk7 mutants generate WT-level of reactive oxygen species burst upon flg22
perception
To further evaluate the role of CPK7 in Arabidopsis defense, we tested PTI-
mediated ROS production in both cpk7 alleles. The oxidative burst is indeed considered
as a common PTI response that is rapidly induced after PAMP treatment and creates a
chemical stress to pathogen (Boller and Felix, 2009). ROS production of leaf discs from
WT and cpk7 mutants after treatment with 100 nM flg22 was evaluated by detecting
hydrogen peroxide (H2O2)-dependent luminescence of luminol. Within 34 min, cpk7
25
mutants have the same ROS productivity as WT, both in amplitude and sensitivity (Fig.
4). These data suggest that CPK7 does not play a role in the PTI-mediated ROS burst.
CPK7 negatively modulates PTI-mediated callose deposition
Callose is a polysaccharide in the form of β-1, 3-glucan and accumulates on the
cell wall of plants leading to mechanical reinforcement against pathogens invasion
(Chen and Kim, 2009). Since PAMP treatment causes callose accumulation in plant
tissues, callose deposition is thought be a biological signature of PTI activation (Boller
and Felix, 2009). After 100 nM flg22 infiltrations, leaves of both cpk7 mutant alleles
produced significantly more callose deposits at 6 h and 9 h post infiltration when
analyzed by the aniline blue staining method (Fig. 5a -5d). These results suggest that
CPK7 acts as a negative regulator of PTI-mediated callose deposition.
CPK7 is not critical for stomatal innate immunity
Stomatal movement is controlled by multiple factors such as circadian clock,
evapotranspiration and also critical for plant-microbe interaction (Misra et al., 2015).
During bacterial pathogen infection, guard cells close to form a physical barrier
blocking invading entrance (Melotto et al., 2006). This procedure is regarded as a part
26
of the plant innate immunity response. To test whether CPK7 participates in stomatal
immunity, stomatal apertures in epidermal peels of Col-0, cpk7 36-1 and cpk7 36-2
were measured upon Pst DC3000 inoculation. In normal condition, one and half hour
after Pst DC3000 infection, stomata close for limiting bacteria entry. However, the
virulent Pst DC3000 secretes the virulence factor coronatine (COR) to actively reopen
stomata at 3 hpi. In epidermis peels of the two cpk7 mutants, WT stomatal closure was
observed after Pst DC3000 inoculation (Fig. 6). Hence, CPK7 may not play a critical
role in Arabidopsis stomatal innate immunity.
Higher MPK3/MPK6 phosphorylation level is observed in cpk7 mutants after
flg22 treatment
Downstream of PRRs-mediated recognition of PAMPs, a complete MAPK
cascade including MEKK1-MKK4/5-MPK3/6 transfer PTI signaling through a serie of
transphosphorylation events (Asai et al., 2002). Hence, phosphorylation of
MPK3/MPK6 is regarded as a maker for PTI activation. To determine whether CPK7
affects the MAPK cascade, leaves of Col-0, cpk7 36-1 and cpk7 36-2 mutants were
infiltrated with 1 μm flg22. MPK3/MPK6 phosphorylation levels were higher in the
cpk7 mutants than in Col-0 WT (Fig. 7), indicating that MAPK cascade is more active
27
in the absence of CPK7. Hence, CPK7 may negatively regulate the PAMP-mediated
MAPK cascade.
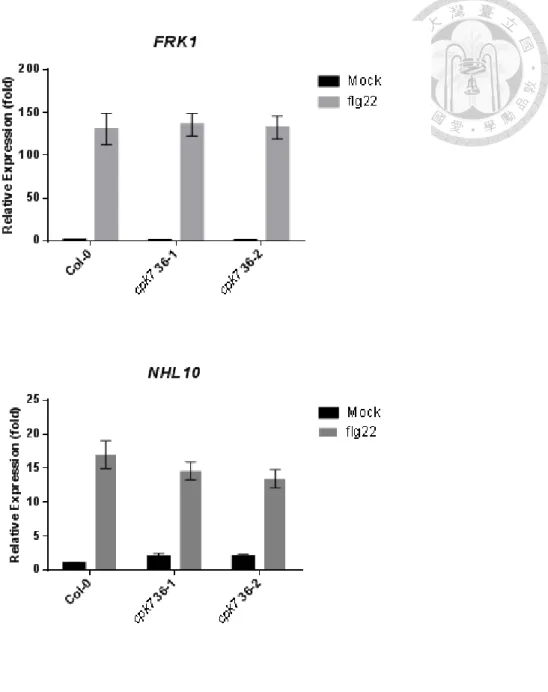
PTI marker genes up-regulation is comparable between WT and cpk7 mutants
As well as other defense mechanisms, activation of PTI is accompanied by
tremendous transcriptional alterations (Zipfel et al., 2004). Since cpk7 mutants appear
to have a stronger activation of MPK3/MPK6, we hypothesized that PTI maker genes
downstream of the MAPK cascade should be up-regulated. To test this hypothesis, 500
nM of flg22 were infiltrated into leaves of Col-0, cpk7 36-1 and cpk7 36-2, and the
expression levels of the PTI marker genes FRK1 and NHL10 were evaluated at 5 h after
infiltration. Surprisingly, up-regulation of both FRK1 and NHL10 was at WT levels in
the cpk7 mutants (Fig 8a and b). This result indicates that, although MPK3/MPK6 is
more activated in cpk7 mutants, the transcriptional level of downstream FRK1 and
NHL10 may not be affected, at least in the condition tested in our experiment.
cpk7 mutants demonstrate a susceptible phenotype to necrotrophic Pcc and B.
cinerea infections
Overexpression of LecRK VI.2 in Arabidopsis enhances resistance to both
28
biotrophic Pst DC3000 and necrotrophic Pcc pathogens, (Singh et al., 2012). To further
investigate the function of CPK7 in plant defense, cpk7 mutants were challenged with
necrotrophic Pcc bacteria. Observing symptoms on the 2nd day after dip-inoculation,
more bacteria-mediated lesions were produced on leaf areas of cpk7 36-1 and cpk7 36-
2 then on Col-0 WT (Fig. 9a). By measuring bacterial titers, similar with symptom
observation, both cpk7 mutant alleles were significantly more susceptible to Pcc than
to Col-0 WT (Fig. 9b). These results suggest that CPK7 may positively regulate the
Arabidopsis defense responses to Pcc.
To determine whether the susceptibility in cpk7 mutants is due to necrotrophic
nutrition, we then performed disease assay with B. cinerea, a necrotrophic fungi. After
3.5 days from inoculation, lesion circles on cpk7 36-1 and cpk7 36-2 were larger than
those on Col-0 (Fig. 10a). This result was also quantified by measuring lesion perimeter,
also showed that the symptom of cpk7 mutants is much more severe than Col-0 (Fig
10b). Together these observations indicate that mutations in CPK7 cause a loss of
resistance to necrotrophic pathogen.
Expression of PR1 is potentiated in cpk7 mutants
Plants take advantage of SA-dependent defense upon biotrophic pathogen attack
and JA/ET-dependent defense for necrotrophic pathway (Pieterse et al., 2009; Bari and
29
Jones, 2009). Since cpk7 mutants were resistant to hemi-biotrophic Pst but susceptible
to necrotrophic pathogens, we hypothesized that CPK7 could be involved in either one
of the two hormone-related defenses. Determination of PR1 expression level is a
common method to evaluate the activation of the SA-dependent pathway (Bari and
Jones, 2009). Leaves of Col-0, cpk7 36-1 and cpk7 36-2 were syringe-infiltrated with
107 cfu/mL of Pst DC3000, after 5 h, PR1 expression in cpk7 36-1 and cpk7 36-2 was
more up-regulated then in Col-0 WT (Fig. 11a), in which up-regulation only started at
9 hpi when in cpk7 mutants, PR1 expression levels were already decreased (Fig. 11b).
Consequently, by monitoring expression of PR1, we suggest that SA-dependent
pathway in cpk7 mutants is more active.
cpk7 mutants accumulate less PDF1.2 transcripts upon Pcc infiltration but
normal level upon B. cinerea infection
Like PR1 for SA-dependent defense, PDF1.2 serves as the marker gene for JA/ET-
dependent pathway (de Torres et al., 2009 ). Leaves of Col-0, cpk7 36-1 and cpk7 36-2
were syringe-infiltrated with 106 cfu/mL of Pcc. After 1 day, up-regulation of PDF1.2
in cpk7 36-1 and cpk7 36-2 was weaker than in Col-0 WT (Fig. 12a). For measuring
PDF1.2 expression upon B. cinerea infection, Col-0 and cpk7 mutants were uniformly
30
sprayed with 5 x 106 spores/mL from B. cinerea. Surprisingly, after 1 day, PDF1.2 is
fully up-regulated and the expression levels are similar between Col-0 and the two cpk7
mutants (Fig. 12b), indicating that there exists at least one circumstance in which cpk7
mutants is able to activate WT levels of the JA/ET responses. Results of these
experiments on hormone-related defenses suggest that CPK7 might be required for
signal crosstalk between SA-dependent and JA/ET- dependent pathways.
CPK7 negatively regulates FRK1 expression upon pep1 treatment but not
PROPEP1 expression
Recently, PEPR pathway was shown to act downstream of PTI and to regulate
hormone related-defenses (Ross et al., 2014). Activation of PEPR pathway is
accompanied with FRK1 up-regulation. To determine whether CPK7 take parts in the
PEPR signaling pathway, FRK1 expression levels after 50 nM pep1 treatment was
monitored for 6 hours. Interestingly, expression of FRK1 in Col-0 was low and rapidly
decreased at 4 h. Meanwhile in the two cpk7 mutants, FRK1 expression was higher and
did not decrease until 5 h (Fig. 13a). This prolonged signal implies that CPK7 might
serve as a negative regulator of the PEPR pathway.
Besides FRK1, it is also known that PEPR pathway forms a positive loop itself, so
31
induction of transcriptional expression of PROPEPs is another marker of the PEPR
pathway (Huffaker and Ryan, 2007)). No significant differences of PROPEP1
expression levels between Col-0 and cpk7 mutants upon pep1 treatment were observed
(Fig. 13b), suggesting that the amplification of FRK1 in cpk7 36-1 and cpk7 36-2 is not
a consequence of intrinsic pep1 production.
CPK7 is localized on the plasma membrane and nucleus and localization is
affected by flg22 treatment
To further investigate the characteristic of CPK7, subcellular localization of
CPK7 was analyzed by transient assay system with GFP fused proteins. Interestingly,
the fluorescence signals could be detected in nucleus and on membrane
simultaneously but not in cytosol (Fig. 14). Nonetheless, addition of flg22 arouse
changes to localization of CPK7, after treatment, fluorescence signals on membrane
were stronger while weaker in nucleus. This result in advanced suggests that CPK7
participates in the downstream of PTI.
CPK7 does not associate with FLS2, BAK1, BIK1 and PEPR1
This work proposes several evidences linking CPK7 to the PTI response, further
32
influencing hormone-related defenses and the PEPR pathway. FLS2, BAK1 and BIK1
act as core proteins of one of the PRR involved in PTI (Chinchilla, 2007). On the other
hand, PEPR1 is the receptor for pep1, therefore is critical for the PEPR pathway
(Yamaguchi et al., 2010). We thus used the BiFC assay to determine whether CPK7
associates with these proteins. None of these well-studied components of PTI (Fig. 15a)
or PEPR pathway (Fig. 15b) associated with CPK7, indicating that there must be other
way for CPK7 to influence plant defenses.
33 Discussion
CPK7 plays a role in the Arabidopsis PTI as a negative regulator.
In accordance of being activated by stresses triggering Ca2+ oscillation, members
in CPK family are often thought to play a positive role upon stresses (Boudsocq and
Sheen, 2013). Recently, study on CPK28 clearly demonstrated that CPK28 actively
suppresses PTI responses by directly phosphorylating RBOHD (Monaghan et al., 2015).
This report reveals a new characteristic of CPKs, CPKs can indeed also act as a negative
regulator to certain stress responses. Here, we find a novel negative regulator of PTI
from the CPK family, CPK7. cpk7 mutants were more resistant to hemi-biotrophic
bacterial pathogen, Pst DC3000, and were more active on MAPK phosphorylation and
callose deposition, suggesting a negative effect of CPK7 to plant innate immunity. In
comparison with CPK28, which is shown to rapidly repress the PTI response, notably
the oxidative burst, the regulatory function carried by CPK7 is rather at a later stage of
the PTI response, such as callose deposition. Confusingly, although activities of
MPK3/MPK6 were inhibited by CPK7, gene expression of downstream gene FRK1 and
NHL10 were not increased under our condition. Possible explanation to this
contradiction is, first, methodologically, we did miss the proper time point at which
expression levels of mutants and Col-0 differentiated. In other words, the effect from
the absence of CPK7 is covered by other mechanism.
34
CPK7 modulates SA-dependent and JA/SA-dependent defense and CPK7
negatively regulates PEPR pathway.
Typical PTI responses conduct a wide-range of resistance to both necrotrophic and
biotrophic pathogens (Singh et al., 2012). However, our results show that cpk7 mutants
are more susceptible to necrotrophic pathogens. Research on SA-dependent defense
indicates that, during biotrophic pathogen attack, SA quickly accumulates in local
invading region, triggering programmed cell death, and soon being transported to
protect adjacent cells (Greenberg and Yao, 2004). Consistent with this information, PR1
expression in cpk7 mutants is earlier induced and decreased, inferring a sensitive SA-
dependent pathway. On the other hand, our results show that PDF1.2 expression is
impaired after Pcc treatment but not after B. cinerea in cpk7 mutants. In comparison
with SA, JA/ET-dependent pathway underlies several feed-back loops and seems to be
more complex (Bostock, 2005), several hypothesizes could be provided. Perhaps from
the beginning, PAMPs from bacteria or from fungus have different effects of CPK7,
leading to the different pattern of JA/ET regulation. Based on these evidences, the
regulatory mechanism of CPK7 to hormone-related defense network is much more
likely to be temporary and buffering rather than constitutive. Confronted with the
traditional mutual antagonism between SA and JA/ET (de Torres et al., 2009), PEPR
35
pathway activates SA-dependent and JA/ET-dependent defenses simultaneously, and is
also regulated by PTI (Ross, et al., 2014; Huffaker and Ryan, 2007). We then assume
that CPK7 regulates SA-JA/ET crosstalk through PEPR pathway. Experiment on FRK1
and PROPEP1 upon pep1 treatment suggest that CPK7 negatively regulator to PEPR
pathway but not due to up regulation of PROPEP1 expression. Yet hardly could we
deduce to our hypothesis that CPK7 acts upstream of PEPR pathway to regulate SA-
JA/ET, more evidences are needed. Finally, note that the induced resistance to Pst
DC3000 and susceptibility to Pcc and B. cinerea are in cpk7 mutants are consist with
those of ein2 knock-out plant (Thomma et al., 1998), with the same phenotype as cpk7
mutants, giving us a clue that CPK7 might also involve in ethylene signaling pathway.
A pilot exam of ERF1 expression upon ACC treatment was performed showing
deficiency on two cpk7 mutants (see Fig. S2).
CPK7 acts downstream of PTI, modulating several defense pathways
Since no clear association between CPK7 and the core components of PTI and
PEPR pathway could be observed, the direct function of CPK7 still remains unknown.
A HPLC-MASS analysis on cpk7 mutants has already be done (Li et al., 2014).
However, without any elicitor treatment, no well-known defense-related genes could
be identified. Thus for surveying over possible interactome of CPK7, and again, with
36
the fact that CPK7 is a constitutive active kinase which localized on nucleus and
membrane, phosphoproteomic-based screening and with PAMP treatment might be a
feasible way. In conclusion, on the basis of our results, CPK7 acts downstream of PTI,
negatively regulate SA-dependent and PEPR pathway, but might be critical for full
activation of the JA/ET pathway.
37 Conclusion and future perspectives
CPK7 serves as a negative regulator of plant resistance to Pst DC3000 and
suppresses some of the PTI responses such as MPK3/MPK6 phosphorylation and
callose deposition. Nonetheless, CPK7 also modulates SA-dependent and JA/ET-
dependent pathways by fast accumulation of PR1 mRNAs and less PDF1.2 up-
regulation upon bacterial pathogen attack and thus having a susceptible phenotype to
necrotrophic pathogens such as Pcc and B. cinerea. CPK7 has also a long-term negative
effect on FRK1 expression upon pep1 treatment. With these evidences, we suggest that
CPK7 plays a complex role downstream of PTI and maintains balance with different
defense signals. Although no clear association between CPK7 and several critical PTI
components when tested with the BiFC assay, other approaches, such as yeast-two
hybrid, for the sake of interactome of CPK7, are necessary for identifying possible
direct relationship of CPK7 with key PTI players. For more details of the biochemical
and physical function of CPK7, generation of overexpression lines and kinase-dead,
EF-hand mutated complementary lines are on-going. By accomplishing these,
hopefully we will be able to sketch a fine graph to better define the function of CPK7
in the Arabidopsis defence signaling.
38 Figure
Figure 1: CPK family in Arabidopsis.
CPK family contains 34 members in 4 groups in Arabidopsis. Adopted from Chen, et
al., 2002.
39 (a)
(b)
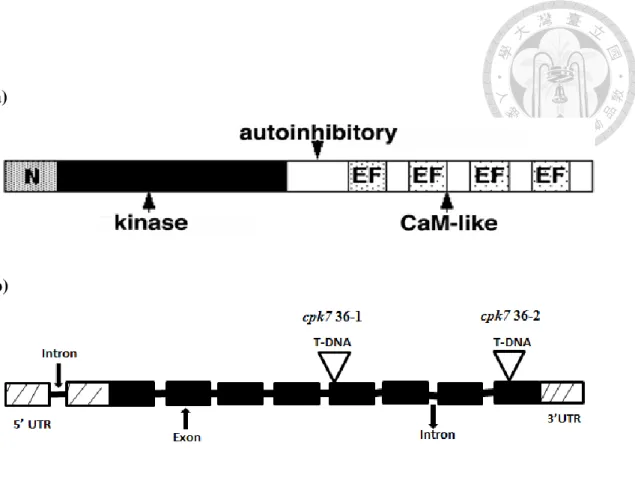
Figure 2: The putative structure of CPK7 and cpk7 mutants.
(a) CPK7 gene encodes a 535 amino acid residues polypeptide, containing a signal
peptide, a N-terminal domain (gray), a conserved serine/threonine kinase domain
(black), an autoinhibitory junction region (white) and a C-terminal CamLD (spotted
region).
(b) The T-DNA insertion sites for cpk7 36-1 and cpk7 36-2 in At5g12480. Plant
materials were well characterized to be homozygous and could not generate full-
length CDS of CPK7 by former member Shweta (Data not shown).
40 (a)
(b)
Log(CFU/cm2 )
Col-0
cpk7 36-1
cpk7 36-2
4.04.55.05.56.0
C o l- 0 c p k 7 3 6 - 1 c p k 7 3 6 - 2
*
*
41
Figure 3. Disease symptoms and bacterial titers of Pst DC3000 infected Col-0
and cpk7 mutant lines
(a) Five-week-old Arabidopsis (Col-0, cpk7 36-1 and cpk7 36-2) were dip-inoculated
with 2 × 107 cfu/mL Pst DC3000. Photos were taken after 3 days. This experiment
was repeated at least 3 times with similar results.
(b) Col-0, cpk7 36-1 and cpk7 36-2 were dip-inoculated with 1 × 106 cfu/mL Pst
DC3000 and bacterial titers were evaluated 2 days later. Data represent 3
independent biological replicates each with 4 technical repeats (n = 12). Asterisks
indicate a significant difference to WT control on a t test (P < 0.001).
42 Figure 4. ROS production after flg22 treatment.
Production of ROS in 5-week-old Col-0, cpk7 36-1 and cpk7 36-2 leaf discs after
treatment with 100 nM flg22 (F) as relative light units (RLU). Mock controls (M) were
treated with sterilized water. Values represent averages ± SE (n = 8 for F and 4 for M).
Experiments were repeated at least 3 times with similar results.
43 (a)
(b)
44 (c)
(d)
count/area(mm2 )
Col-0 M
Col-0 F
cpk7 36-1 M
cpk7 36-1 F
cpk7 36-2 M
cpk7 36-2 F 0
5 0 1 0 0 1 5 0
*
*
* *
* *
* *
count/area(mm2 )
Col-0 M
Col-0 F
cpk7 36-1 M
cpk7 36-1 F
cpk7 36-2 M
cpk7 36-2 F 0
1 0 0 2 0 0 3 0 0
* *
* *
* *
* *
* *
45
Figure 5. Visualizations and quantifications of callose deposits upon flg22
treatment
Five-week-old Col-0, cpk7 36-1 and cpk7 36-2 leaves were syringe-infiltrated with
100 nM flg22, and sterilized water were used as mock controls. Samples were harvested
6 h (a) or 9 h (b) later for aniline blue staining. These experiments were repeated 3 times
with similar results. Graph represents the average number of callose deposits observed
per square millimeter ± SD (n = 12) at 6 h (c) and 9 h (d). Asterisks indicate a significant
difference to WT Col-0 treated with flg22 based on a t test (*: P < 0.005 and **:
P<0.001).
46 Figure 6: CPK7 in stomatal innate immunity.
Stomatal apertures in leaf epidermal peels from 5-week-old Col-0, cpk7 36-1 and cpk7
36-2 exposed to MES buffer (control) or 1 × 108 cfu/mL Pst DC3000 for 1 and 3 h.
Values are shown as means ± SE (n = 80 stomata) of 3 independent biological replicates.
Asterisks indicate a significant difference to Col-0 treatment based on a t test analysis
(P < 0.05).
47 (a)
(b)
48 Figure 7: MAPK kinase Assay
(a) Nine leaves from 5-week-old Col-0, cpk7 36-1 and cpk7 36-2 were syringe-
infiltrated with 100 nM flg22 (+) and sterilized water (-) for 10 min as shown by
immunoblot analysis using phosphor-p44/42 MAK kinase antibody. Immunoblot,
top panel; Ponceu S-stained membrane is used to estimate equal loading in each
lane, bottom panel. This experiment is one of 2 independent replicates.
(b) Quantification of immunoblot signals to background. Relative densities are average
form 2 independent replicates. Asterisks indicate significant differences by t-test
(p<0.05).
49 (a)
(b)
Figure 8. Transcriptional expression of PTI-responsive genes FRK1 and NHL10
after flg22 treatment.
Relative expression level of FRK1 (a) or NHL10 (b) in 5-week-old Col-0, cpk7 36-1
and cpk7 36-2 were analyzed at 5 h after infiltrated with 100 nM flg22 by qRT-PCR.
UBQ10 was used for normalization. The values are the means ± SE of 3 biological
replicates (n = 9).
50 (a)
(b)
Log(CFU/cm2 )
Col-0
cpk7 36-1
cpk7 36-2 3
4 5 6
C o l- 0
c p k 7 3 6 - 1 c p k 7 3 6 - 2
* *
51
Figure 9. Disease symptoms and bacterial titers of Pcc infected Col-0 and cpk7
mutant lines
(c) Five-week-old Arabidopsis (Col-0, cpk7 36-1 and cpk7 36-2) were dip-inoculated
with 106 cfu/mL Pcc. Photos were taken after 2 days. This experiment was repeated
at least 3 times with similar results.
(d) Col-0, cpk7 36-1 and cpk7 36-2 were dip-inoculated with 106 cfu/mL Pcc and
bacterial titers were evaluated 2 days later. Data represent 3 independent biological
replicates each with 4 technical repeats (n = 12). Asterisks indicate a significant
difference to WT control on a t test (P < 0.001).
52 (a)
(b)
53
Figure 10: Disease symptoms and lesion perimeter of B. cinerea infected Col-0
and cpk7 mutant lines
(a) Disease phenotype. B. cinerea inoculation was performed on Five-week-old Col-
0, cpk7 36-1 and cpk7 36-2. Plants were inoculated with 8 µL droplets of B. cinerea
spores (105 spores/mL in PDB). Photos were taken at 3 dpi. This experiment was
repeated 3 times with similar results.
(b) Lesion perimeter. Measurements were realized at 3 dpi and quantified by Image J.
Values are the means ± SE of 3 biological replicates each consisting of 12 technical
repeats (n = 36). Asterisks indicate a significant difference to Col-0 control based on a
Student’s t test (p<0.001).
54 (a)
(b)
P R 1
Relative Expression (fold)
Col-0
cpk7 36-1
cpk7 36-2 0
5 1 0 1 5
*
* M o c k
P s t D C 3 0 0 0
P R 1
Relative Expression (fold)
Col-0
cpk7 36-1
cpk7 36-2 0
2 0 0 4 0 0 6 0 0
M o c k
P s t D C 3 0 0 0
* *