Significant Association of Methylenetetrahydrofolate Reductase Single Nucleotide Polymorphisms with Prostate Cancer
Susceptibility in Taiwan
HSI-CHIN WU
1,4,*, CHAO-HSIANG CHANG
1,4*, RU-YIN TSAI
1*, CHIH-HSUEH LIN
5, ROU-FEN WANG
1,2, CHIA-WEN TSAI
1,2, KUEN-BAO CHEN
1, CHUN-HSU
YAO
3, CHANG-FANG CHIU
1, DA-TIAN BAU
1,2, and CHENG-CHIEH LIN
1,51
Terry Fox Cancer Research Laboratory, China Medical University Hospital, Taichung, Taiwan, ROC;
2
Graduate Institute of Basic Medical Science, and
3Department of Biomedical
Imaging and Radiological Science, China Medical University, Taichung, Taiwan, ROC;
4
Department of Urology, and
5Department of Family Medicine, China Medical University Hospital, Taichung, Taiwan, ROC
Running head: Bau et al: MTHFR Genotypes In Prostate Cancer
* These authors contributed equally to this work
Correspondence to: Da-Tian Bau and Cheng-Chieh Lin, Terry Fox Cancer Research Lab,
China Medical University Hospital, 2 Yuh-Der Road, Taichung, 404 Taiwan, P.R.C. Tel:
+886 422053366 ext 3312, Fax: +886 422053366 ext 1511, e-mail:
[email protected]; [email protected]
Key words: prostate cancer, methylenetetrahydrofolate reductase, polymorphism, and
carcinogenesis.
Abstract. Prostate cancer is the most common cause of cancer death in men and is a
major health problem worldwide. Methylene tetrahydrofolate reductase (MTHFR)
plays an important role for folate metabolism and is also an important source for DNA
methylation and DNA synthesis (nucleotide synthesis). To assess the association and
interaction of genotypic polymorphisms in MTHFR and lifestyle factors with prostate
cancer in Taiwan, we investigated two well-know polymorphic variants of MTHFR,
C677T (rs1801133) and A1298C (rs1801131), analyzed the association of specific
genotypes with prostate cancer susceptibility, and discussed their joint effects with
individual habits on prostate cancer risk. In total, 218 patients with prostate cancer
and 436 healthy controls recruited from the China Medical Hospital in central Taiwan
were genotyped investigating the association of these polymorphisms with prostate
cancer susceptibility. We found the MTHFR C677T but not the A1298C genotype,
was differently distributed between the prostate cancer and control groups. The T
allele of MTHFR C677T conferred a significantly (P = 0.0011) decreased risk of
prostate cancer. As for the A1298C polymorphism, there was no difference in
distribution between the prostate cancer and control groups. Gene interactions with
smoking were significant for MTHFR C677T polymorphism. The MTHFR C677T CT
and TT genotypes in association with smoking conferred a decreased risk of 0.501
(95% confidence interval=0.344-0.731) for prostate cancer. Our results provide the
first evidence that the C allele of MTHFR C677T may be associated with the
development of prostate cancer and may be a novel useful marker for primary
prevention and anticancer intervention.
Prostate cancer is one of the most important diseases in men all over the world. In the men of the United States and Western Europe, prostate cancer is a leading cause of illness and death (1), while the incidence of prostate cancer widely varies in different races. According to the literature, Asians have the lowest incidence among the major races, and African—American men have the greatest incidence in the world (2). In Taiwan, although the incidence of prostate cancer is much lower compared with other countries, it still takes the seventh place in the top ten cancer causes of death for male Taiwanese (3). The number of patients and the death rate have also been increasing during the two decades (3), and prostate cancer has become a serious public threaten of Taiwanese mature males. In the literature, some risk factors have been confirmed as being associated with prostate cancer, including age, race, and a family history of prostate cancer (2). Additionally, smoking, together with diet, androgens, occupational chemicals, inflammation and obesity, have been considered as secondary risk factors (2).
In recent years, environmental and genomic susceptibilities and interactions
among them have been used in evaluation of cancer risk. Primary candidates for gene—environment lifestyle interaction studies are those encoding enzymes related to
the metabolism of established risk factors of carcinogenesis. Methylene
tetrahydrofolate reductase (MTHFR) is a key enzyme in folate metabolism, which
catalyzes 5, 10-MTHF to 5-MTHF. The importance of MTHFR in cancer
susceptibility arises from its involvement in two pathways of folate metabolism. One leads to numerous methylation processes that are dependent on S-adenosyl-methionine (SAM), while the other, via thymidylate synthesis, contributes
to DNA replication and cell division. Reduced activity of MTHFR may decrease the methylation of homocysteine to methionine and in turn the level of SAM, resulting in DNA hypomethylation. On the other hand, the reduced level of MTHFR substrate, 5,10-MTHF, required for thymidylate synthesis could lead to uracil misincorporation into DNA, diminished DNA repair and increased frequency of chromosomal breaks and damage. Malignancies that are derived from rapidly proliferating tissues, which have a higher requirement for DNA synthesis, should be more susceptible to folate deficiency and resultant DNA damage. The DNA variants causing reduced MTHFR activity were found to be associated with reduced risk of leukemia, lymphoma and colorectal carcinoma. The mechanism proposed to explain these associations was the shunt of folate metabolism versus thymidine and purine synthesis, which would slow the incorporation of uracil into DNA and protect the cells against carcinogenesis (3).
Previous investigations of MTHFR genetic variations focused on the catalytic
domain and the two polymorphisms C677T and A1298C, which slightly change
enzymatic activity. In the case of C677T polymorphism, the cytosine base at position
number 677 changes to a thymidine base, which in turn affects the amino acid
sequence at position number 222 (alanine Æ valine). The MTHFR C677T variants are
MTHFR 677CC wild type (most common), MTHFR 677CT heterozygous genotype
and MTHFR 677TT homozygous genotype. The MTHFR enzymes with non-wild type polymorphic genotypes become thermo-labile, causing a loss of its activity with increased temperature. The modified protein loses its cofactor FAD more quickly and has a lower stability. The mutation effect can be suppressed by addition of folate, which causes a higher FAD affinity and an increase in MTHFR stability. The MTHFR A1298C polymorphism is localized in the coding regulatory region domain (4).
In 2003, the association between single nucleotide polymorphisms (SNPs) of MTHFR and lung cancer susceptibility was firstly examined in a Taiwan population,
indicating that C677T is not associated with lung cancer risk (5). However, the sample
size was rather small (control/case=232/59), and only one SNP was investigated in the
study. In the present work, we analyzed the genetic polymorphisms of both MTHFR
C677T and A1298C in a more representative population (controls/cases=436/218) in
Taiwan, and investigated the interaction of MTHFR genotypes and smoking habits in
a Taiwanese prostate cancer population.
Materials and Methods
Study population and sample collection. Two hundred and eighteen patients
diagnosed with prostate cancer were recruited at the outpatient clinics of general surgery between 2003-2009 at the China Medical University Hospital, Taichung, Taiwan, Republic of China. All patients voluntarily participated, completed a self-administered questionnaire and provided peripheral blood samples. Twice as many of non-prostate cancer healthy volunteers as controls were selected by matching for age, gender and habits after initial random sampling from the Health Examination Cohort of the hospital. The exclusion criteria of the control group included previous malignancy, metastasized cancer from other or unknown origin, and any familial or genetic diseases. Both groups completed a short questionnaire which included habits and they were recorded. Our study was approved by the Institutional Review Board of the China Medical University Hospital and written-informed consent was obtained from all participants.
Genotyping assays. Genomic DNA was prepared from peripheral blood leukocytes
using a QIAamp Blood Mini Kit (Blossom, Taipei, Taiwan) and further processed
according to previous studies (6-14). The PCR cycling conditions were: one cycle at
94
oC for 5 min; 35 cycles of 94
oC for 30 s, 55
oC for 30 s, and 72
oC for 30 s; and a
final extension at 72
oC for 10 min. Pairs of PCR primer sequences and restriction
enzyme for each DNA product are all listed in Table I.
Statistical analyses. Only those individuals with complete SNP data and smoking
status (cases/controls =218/436) were selected for final analysis. To ensure that the
controls used were representative of the general population and to exclude the
possibility of genotyping error, the deviation of the genotype frequencies of MTHFR
SNP in the controls from those expected under the Hardy-Weinberg equilibrium was
assessed using the goodness-of-fit test. Pearson’s Chi-square test or Fisher’s exact test
(when the expected number in any cell was less than five) was used to compare the
distribution of the genotypes between cases and controls. Data were recognized as
significant when the statistical P-value was less than 0.05.
Results
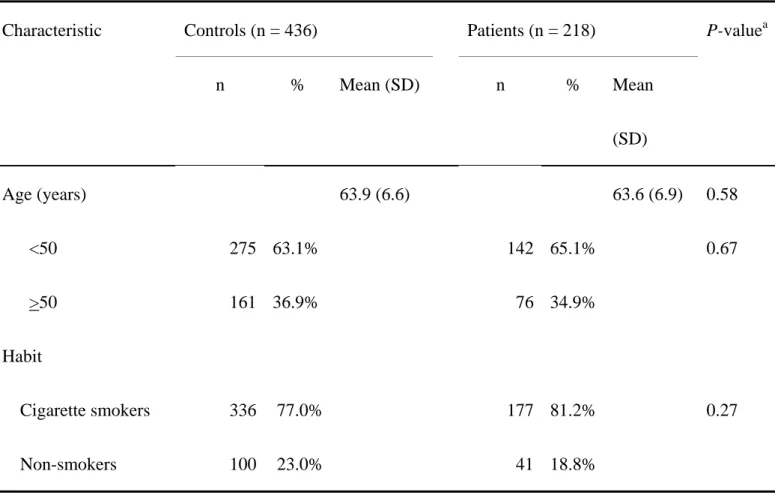
The frequency distributions of selected characteristics of 218 prostate cancer patients and 436 controls are shown in Table II. These characteristics of patients and controls are all well matched. None of the differences between groups were statistically significant (P>0.05) (Table II).
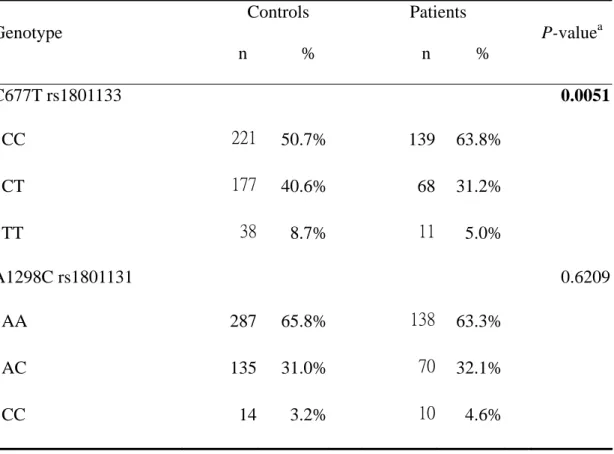
The frequency of the genotypes for the MTHFR C677T and A1298C in controls and prostate cancer patients is shown in Table III. The genotype distribution of the genetic polymorphisms of MTHFR C677T was significantly different between prostate cancer and control groups (P=0.0051), while that for A1298C polymorphisms was not significant (P>0.05) (Table III). The data indicated that only the MTHFR C677T polymorphism was significantly associated with prostate cancer.
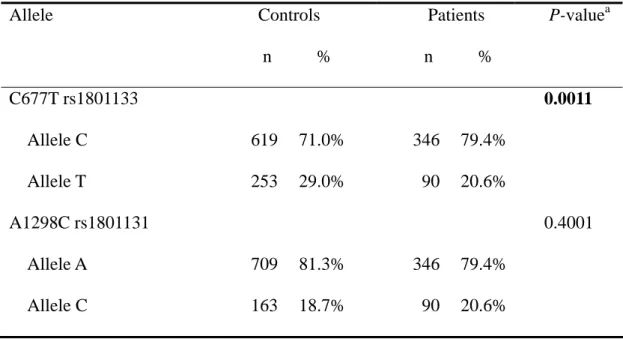
The frequency of the alleles for MTHFR C677T and A1298C in controls and prostate cancer patients is shown in Table IV. The C allele of the MTHFR C677T polymorphism was significantly associated with prostate cancer (P=0.0011). The conclusion deduced from Tables III and IV is that the MTHFR C677T T allele seems to be associated with a lower risk for prostate cancer in Taiwan.
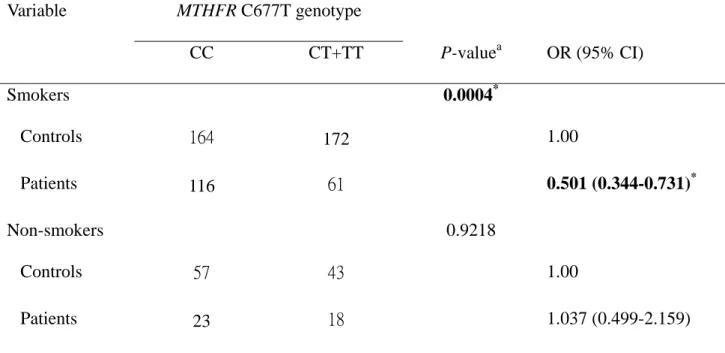
The interaction between MTHFR genotype and individual smoking habits was
further analyzed. The genotype distribution of MTHFR C677T was significantly
different between prostate cancer patients and controls who have smoking habit
(P=0.0004) (Table V), while that for MTHFR A1298C was not significant (P >0.05)
(data not shown). The T allele frequency was significantly lower in cancer patients
who smoked than in controls. The frequency of individuals with MTHFR C677T CT
or TT who smoked were approximately 0.5-fold lower than those with CC and
prostate cancer than those who did not smoke.
Discussion
In order to investigate the role of MTHFR and to find potential biomarkers of prostate cancer, we selected two SNPs of the MTHFR gene and investigated their associations with the susceptibility for prostate cancer in a population of central Taiwan. The C677T MTHFR polymorphism has been related to acute leukemia (15), endometrial carcinoma (16), and colon adenocarcinoma (17, 18). We found that the T variant genotypes of MTHFR C677T were significantly associated with a lower susceptibility for prostate cancer (Tables III and IV). The conclusion was inconsistent with previous findings. Heijmans (19) reported that the incidence of prostate cancer was higher among men with the Val/Val genotype, but others found no such differential distribution between this gene alone and in combinations with other genes (20-22).
On the other hand, Van Guelpen and his colleagues (22) after adjusting for serum levels of folates, vitamin B12 and homocysteine, reported that there was a positive association between the heterozygote C677T and the risk of prostate cancer risk. The wide inconsistency may be caused by differences in ethnicity and population. More importantly, a limited sample size may also cause the variation.
We have further analyzed the association between C677T genotype and prostate cancer risk in patients and controls who have cigarette smoking habits. Interestingly, the interaction between MTHFR C677T and cigarette smoking habit is clear, i.e.
smoking people with the CT or TT genotype have a 2-fold reduction in the odds of the
prostate cancer than those smoking people with the CC genotype (Table V).
To sum up, this is the first study which focuses on the SNPs of MTHFR and their joint effects with smoking habit on prostate cancer risk in Taiwan, and the presence of the C allele of C677T was associated with a higher risk of prostate cancer. The C allele of MTHFR C677T may be a useful marker in prostate oncology for anticancer application, and early cancer detection. In order to further elucidate the importance of the MTHFR C677T in prostate carcinogenesis, larger studies assessing circulating levels, as well as dietary intake of folate, are warranted in the future.
Acknowledgements
We thank Wen-Shin Chang, Hao-Ting Lan, Tzu-Ting Weng and Judy Wang for their
technical assistance. This study was supported by research grants from the Terry Fox
Cancer Research Foundation, National Science Council (NSC
98-2320-B-039-010-MY3) and China Medical University and Hospital
(DMR-99-069).
References
1 Gronberg H: Prostate cancer epidemiology. Lancet 361: 859-864, 2003.
2 Hsing AW and Chokkalingam AP: Prostate cancer epidemiology. Front Biosci 11: 1388-1413, 2006.
3 Krajinovic M, Lamothe S, Labuda D, Lemieux-Blanchard E, Theoret Y, Moghrabi A and Sinnett D: Role of MTHFR genetic polymorphisms in the susceptibility to childhood acute lymphoblastic leukemia. Blood 103: 252-257, 2004.
4 Homberger A, Linnebank M, Winter C, Willenbring H, Marquardt T, Harms E and Koch HG: Genomic structure and transcript variants of the human methylenetetrahydrofolate reductase gene. Eur J Hum Genet 8: 725-729, 2000.
5 Jeng YL, Wu MH, Huang HB, Lin WY, You SL, Chu TY, Chen CJ and Sun CA: The methylenetetrahydrofolate reductase 677C—>T polymorphism and lung cancer risk in a Chinese population. Anticancer Res 23: 5149-5152, 2003.
6 Bau DT, Tsai MH, Huang CY, Lee CC, Tseng HC, Lo YL, Tsai Y and Tsai FJ:
Relationship between polymorphisms of nucleotide excision repair genes and oral cancer risk in Taiwan: evidence for modification of smoking habit. Chin J Physiol 50: 294-300, 2007.
7 Chang CH, Chang CL, Tsai CW, Wu HC, Chiu CF, Wang RF, Liu CS, Lin CC and Bau DT: Significant association of an XRCC4 single nucleotide polymorphism with bladder cancer susceptibility in Taiwan. Anticancer Res 29:
1777-1782, 2009.
8 Chiu CF, Tsai MH, Tseng HC, Wang CL, Wang CH, Wu CN, Lin CC and Bau DT: A novel single nucleotide polymorphism in XRCC4 gene is associated with oral cancer susceptibility in Taiwanese patients. Oral Oncol 44: 898-902, 2008.
9 Chiu CF, Wang CH, Wang CL, Lin CC, Hsu NY, Weng JR and Bau DT: A novel single nucleotide polymorphism in XRCC4 gene is associated with gastric cancer susceptibility in Taiwan. Ann Surg Oncol 15: 514-518, 2008.
10 Chiu CF, Wang HC, Wang CH, Wang CL, Lin CC, Shen CY, Chiang SY and Bau DT: A new single nucleotide polymorphism in XRCC4 gene is associated with breast cancer susceptibility in Taiwanese patients. Anticancer Res 28:
267-270, 2008.
11 Hsu CF, Tseng HC, Chiu CF, Liang SY, Tsai CW, Tsai MH and Bau DT:
Association between DNA double strand break gene Ku80 polymorphisms and oral cancer susceptibility. Oral Oncol 45: 789-793, 2009.
12 Hsu NY, Wang HC, Wang CH, Chiu CF, Tseng HC, Liang SY, Tsai CW, Lin
CC and Bau DT: Lung cancer susceptibility and genetic polymorphisms of
Exo1 gene in Taiwan. Anticancer Res 29: 725-730, 2009.
13 Shrubsole MJ, Shu XO, Ruan ZX, Cai Q, Cai H, Niu Q, Gao YT and Zheng W:
MTHFR genotypes and breast cancer survival after surgery and chemotherapy:
a report from the Shanghai Breast Cancer Study. Breast Cancer Res Treat 91:
73-79, 2005.
14 Yang MD, Hsu YM, Kuo YS, Chen HS, Chang CL, Wu CN, Chang CH, Liao YM, Wang HC, Wang MF and Bau DT: Significant association of Ku80 single nucleotide polymorphisms with colorectal cancer susceptibility in Central Taiwan. Anticancer Res 29: 2239-2242, 2009.
15 Pereira TV, Rudnicki M, Pereira AC, Pombo-de-Oliveira MS and Franco RF:
5,10-Methylenetetrahydrofolate reductase polymorphisms and acute lymphoblastic leukemia risk: a meta-analysis. Cancer Epidemiol Biomarkers Prev 15: 1956-1963, 2006.
16 Esteller M, Garcia A, Martinez-Palones JM, Xercavins J and Reventos J:
Germ line polymorphisms in cytochrome-P450 1A1 (C4887 CYP1A1) and methylenetetrahydrofolate reductase (MTHFR) genes and endometrial cancer susceptibility. Carcinogenesis 18: 2307-2311, 1997.
17 Hubner RA and Houlston RS: MTHFR C677T and colorectal cancer risk: A meta-analysis of 25 populations. Int J Cancer 120: 1027-1035, 2007.
18 Ma J, Stampfer MJ, Giovannucci E, Artigas C, Hunter DJ, Fuchs C, Willett WC, Selhub J, Hennekens CH and Rozen R: Methylenetetrahydrofolate reductase polymorphism, dietary interactions, and risk of colorectal cancer.
Cancer Res 57: 1098-1102, 1997.
19 Heijmans BT, Boer JM, Suchiman HE, Cornelisse CJ, Westendorp RG, Kromhout D, Feskens EJ and Slagboom PE: A common variant of the methylenetetrahydrofolate reductase gene (1p36) is associated with an increased risk of cancer. Cancer Res 63: 1249-1253, 2003.
20 Kimura F, Franke KH, Steinhoff C, Golka K, Roemer HC, Anastasiadis AG and Schulz WA: Methyl group metabolism gene polymorphisms and susceptibility to prostatic carcinoma. Prostate 45: 225-231, 2000.
21 Cicek MS, Nock NL, Li L, Conti DV, Casey G and Witte JS: Relationship between methylenetetrahydrofolate reductase C677T and A1298C genotypes and haplotypes and prostate cancer risk and aggressiveness. Cancer Epidemiol Biomarkers Prev 13: 1331-1336, 2004.
22 Van Guelpen BR, Wiren SM, Bergh AR, Hallmans G, Stattin PE and Hultdin J:
Polymorphisms of methylenetetrahydrofolate reductase and the risk of prostate
cancer: a nested case—control study. Eur J Cancer Prev 15: 46-50, 2006.
Table I. The primer sequences, and polymerase chain reaction and restriction
fragment length polymorphism (PCR-RFLP) conditions for analysis of MTHFR gene polymorphisms.
Polymorphism
(rs number)
Primer sequences (5’ to 3’) Restriction
enzyme
SNP
sequence
DNA fragment
size (bp)
C677T (rs1801133)
F: TGA AGG AGA AGG TGT CTG CGG GA R: AGG ACG GTG CGG TGA GAG TG
Hinf I C
T
198 bp 175 + 23 bp A1298C
(rs1801131)
F: GGGAGGAGCTGACCAGTGCAG R: GGGGTCAGGCCAGGGGCAG
Fnu4H I C
A
138 bp 119 + 19 bp
*F and R indicate forward and reverse primers, respectively.
Table II. Characteristics of prostate cancer patients and controls.
Characteristic Controls (n = 436) Patients (n = 218) P-value
an % Mean (SD) n % Mean
(SD)
Age (years) 63.9 (6.6) 63.6 (6.9) 0.58
<50 275 63.1% 142 65.1% 0.67
>50 161 36.9% 76 34.9%
Habit
Cigarette smokers 336 77.0% 177 81.2% 0.27
Non-smokers 100 23.0% 41 18.8%
a
Based on Chi-square test.
Table III. Distribution of MTHFR genotypes among prostate cancer patient and control groups.
Genotype
Controls n %
Patients n %
P-value
aC677T rs1801133 0.0051
CC 221 50.7% 139 63.8%
CT 177 40.6% 68 31.2%
TT 38 8.7% 11 5.0%
A1298C rs1801131 0.6209
AA 287 65.8% 138 63.3%
AC 135 31.0% 70 32.1%
CC 14 3.2% 10 4.6%
a
Based on Chi-square test.
Table IV. Distribution of MTHFR alleles among the prostate cancer patient and control
groups.
Allele Controls n %
Patients n %
P-value
aC677T rs1801133 0.0011
Allele C 619 71.0% 346 79.4%
Allele T 253 29.0% 90 20.6%
A1298C rs1801131 0.4001
Allele A 709 81.3% 346 79.4%
Allele C 163 18.7% 90 20.6%
a
Based on Chi-square test.
Table V. MTHFR C677T genotype and prostate cancer after stratification by smoking
habit.
Variable MTHFR C677T genotype
CC CT+TT P-value
aOR (95% CI)
Smokers 0.0004
*Controls 164 172 1.00
Patients 116 61 0.501 (0.344-0.731)
*Non-smokers 0.9218
Controls 57 43 1.00
Patients 23 18 1.037 (0.499-2.159)
a