檜木林經營模式對於地被植物多樣性 與林木天然更新之影響
蘇聲欣
1,2)邱志明
1)彭炳勳
1)摘 要
森林經營對於天然檜木林生態之影響過去在台灣曾引起重大爭議,相關研究亦未有定論,因此仍 需投入更多研究工作進行驗證。本研究在具有長久林業經營歷史而且仍保有廣大天然檜木林的棲蘭山 區,選擇4種不同經營模式下的檜木林分進行地被植群及天然更新樹苗調查,以比較不同林型之植物多 樣性與林下更新狀態。調查結果發現,枯立倒木整理後老齡林具有較高的地被植物多樣性,天然老齡 林則具有較高的樹苗多樣性,而擇伐後天然更新林無論在地被植物或樹苗的多樣性皆為最低。從物種 組成來看,天然老齡林的地被植物及樹苗物種均與其他林型存有明顯的區隔。不同林型的天然更新狀 態也有極明顯的差異,其中以天然老齡林的樹苗量最高,分別在全部樹種以及檜木(均為台灣扁柏)樹 苗上佔有77、87%。此外,91%檜苗是建立在木質殘材之上,顯見與其他樹種具有不同的生長基質偏 好。
本研究顯示不同的經營模式對於檜木林的地被植群、林木天然更新具有長期的影響,人為作業過 之林分雖歷經20~40年的恢復,仍未發展出近似天然老齡林的狀態。我們建議應保護及長期監測天然老 齡林之生態,並可運用枯立倒木整理林內之不同植生階段來增加生物棲地種類,進而提升天然林之整 體生物多樣性。至於皆伐後人工栽植林與擇伐後更新林,應積極進行中、後期撫育工作以促進林木生 長並改善林況。
關鍵詞:台灣扁柏、枯立倒木整理、檜木林、地被植群、天然更新。
蘇聲欣、邱志明、彭炳勳。2018。檜木林經營模式對於地被植物多樣性與林木天然更新之影響。台灣 林業科學33(2):125-40。
1)
林 業 試 驗 所 森 林 經 營 組 ,10066台北市南海路53號 Forest Management Division, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan.
2)
通訊作者 Corresponding author, e-mail:sush@tfri.gov.tw
2017年9月送審 2018年1月通過 Received September 2017, Accepted January 2018.
Research paper
Impacts of Forest Management Regimes on the Ground Vegetation Diversity
and Natural Regeneration of Chamaecyparis Forests
Sheng-Hsin Su,
1,2)Chih-Ming Chiu,
1)Ping-Hsun Peng
1)【 Summary】
The impact of forest management on the ecology of natural Chamaecyparis (cypress) forests previously caused momentous disputes in Taiwan. However, the scientific questions behind this is- sue have never been settled and thus require further investigations. This study was conducted in the Chilanshan area of northeastern Taiwan, where there is a long history of forest management, and extensive natural Chamaecyparis forests are still preserved. We investigated the ground vegetation and naturally regenerating seedlings in stands under 4 different management regimes to compare their plant diversity and tree regeneration. Our results showed that the diversity of ground vegeta- tion was higher in the old-growth forest where snags and logs had previously been harvested, while the diversity of tree seedlings was higher in the natural old-growth forest. The lowest diversities of ground vegetation and seedlings were both observed in the naturally regenerated stand established after selective cutting. The natural old-growth forest was also distinct from the other stand types in terms of the species composition. The natural regeneration of seedlings significantly differed among stand types. The natural old-growth forest contained the greatest amount of both cypress seedlings (all of which were Chamaecyparis obtusa var. formosana; 87%) and overall seedlings (77%). Moreover, 91% of cypress seedlings were established on coarse woody debris, revealing a distinct substrate preference from the other species.
Our study indicated long-term impacts of management regimes on the ground vegetation and natural regeneration in Chamaecyparis forests. After 20~40 yr of recovery, stands that had expe- rienced management practices were still dissimilar to natural old-growth forests. Therefore, we would argue that natural old-growth forests should be protected and ecologically monitored, and that old-growth forests where snags and logs had previously been removed could offer various vegetation stages to create more-heterogeneous habitats and thus enhance the overall biological di- versity. As for plantations and naturally regenerated stands, middle- or later-stage tending practices should promptly be applied to improve tree growth and stand conditions.
Key words: Chamaecyparis obtusa var. formosana, removing snags and logs, Chamaecyparis forest (cypress forest), ground vegetation, natural regeneration.
Su SH, Chiu CM, Peng PH. 2018. Impacts of forest management regimes on the ground vegetation diversity and natural regeneration of Chamaecyparis forests. Taiwan J For Sci 33(2):125-40.
緒 言
檜木林是台灣極為珍貴的自然資源,其重 要性非僅限於檜木本身的林產利用價值,還包 括檜木林生態系所蘊藏的生物多樣性、生態服 務功能、自然襲產意義、森林文化與美學價值 等等(Kuo 1995, Lin et al. 2016)。台灣自日據時 期1912年開始大量砍伐天然檜木,直至1989年 政府宣布全面禁伐天然檜木林,原始檜木林的 開發行為才告中止,進入到森林保育時期(Jen 1995, Horng et al. 2000)。自此之後,林業機關 不再以木材生產做為天然檜木林的唯一經營目 標,並開始發展全新的多目標森林經營規劃。
然 而 在 過 去 二 十 多 年 來 , 有 關 於 天 然 檜 木林是否應該以人為方式介入經營,例如枯立 倒木整理作業、檜木保育更新作業等等(Huang 1995, Lin et al. 2016),仍舊引起極大的民間保 育運動與學術爭論(Li 1988, HH Chen 1999, YF Chen 1999, Council of Agriculture 2000)。這些 爭議凸顯出兩項亟待驗證的科學問題:
1. 人為經營是否會對天然檜木林之生物多樣性 造成負面影響?
2. 檜木天然老齡林是否需要人為經營以促進林 分更新?
隨著國內自然保育觀念的逐漸深化與政策 演進,近年來相關管理單位雖然陸續有為台灣 天然檜木林成立「國家公園」或申請「世界自 然遺產」之提議(Ministry of the Interior 2002, Lin et al. 2016),但是關於前述檜木林保育與經 營的根本議題,則未有更進一步的討論。
在 檜 木 林 的 研 究 進 展 方 面 , 在 此 期 間 陸 續有許多研究人員投入於檜木林調查與監測,
進行諸如植群生態(Chen et al. 2002, Li et al.
2015)、動物群聚生態(Yuan et al. 2004, Chen et al. 2015)、生物多樣性清查(Lee 2000, Wang 2000)、林分結構與動態(Chang et al. 2001, Liao et al. 2003a, 2003b)、生態系功能(Chang et al. 2008)等不同面向的研究,逐漸拓展吾人 對於檜木林生物資源與生態的了解。有關於生 物多樣性議題,Yuan et al. (2004)比較棲蘭山 枯立倒木整理後與未整理林地的鳥類相,發現 林地作業易造成林下植被的破壞、改變鳥類食
物來源,因而影響鳥類群聚組成,但鳥類種數 未見差異。Chen et al. (2015)則是調查棲蘭山 3種檜木林型的野生哺乳動物群聚,發現枯立 倒木整理林與原始林差異不大,但是人工林的 動物群聚則明顯不同。有鑑於哺乳動物、鳥類 均具有長距離的移動能力,生活範圍並不受限 於單一林分,相較之下,行固著生活(sessile) 的植物會受到種子或孢子產生數量、散播機制 (dispersal)與生育地篩選(habitat filtering)等生 態過程的高度限制,因此在不同林分經營模式 之下很可能產生與動物群聚不同的反應。而地 被層植物與樹苗做為最接近地面的植生組成,
可 提 供 許 多 動 物 食 物 來 源 與 棲 息 環 境 , 因 此 與林地之整體生物多樣性有密切關聯(Yuan et al. 2004, Teixeira et al. 2017)。Wei and Chen (2012)曾在棲蘭山研究不同人為作業擾動下林 分的植物多樣性,發現受擾動林分中的冠層樹 種組成雖已近似於天然林,但次冠層以下的植 被則與天然林相差甚多。但是該研究並未探討 檜木的天然更新情形,而且取樣調查地點廣泛 分布於棲蘭山各林道區塊,可能會受到不同地 點的背景條件差異(區域性氣候、周遭植被)而 影響分析結果,因此仍有必要進行不同的取樣 設計,並增加林分天然更新情形的調查分析,
以進一步釐清相關問題。
而關於檜木天然更新,向來即為檜木林經 營的重要議題,許多人為作業之目標均是為了 促進林分持續天然更新,以期達成森林永續經 營(Hung 1982, Kuo 1991, Huang 1995, Lin et al. 2016)。在檜木天然更新調查方面,Chang et al. (2001)針對棲蘭山不同時期枯立倒木整理後 的林分進行試驗處理,並調查檜木種子雨、種 子發芽與樹苗建立之情形,Liao et al. (2003a, 2003b)則在鴛鴦湖及鄰近地區的天然檜木林進 行檜木稚樹調查,他們均發現到林地內的倒木 殘材與檜木樹苗(以下簡稱檜苗)之建立有明顯關 聯性。然而,這些研究的地點均設定在天然林 或人為經營林分其中之一,並未擴及不同經營 模式下林分的比較,而且在研究方法(人工試驗 處理,或純粹野外調查)、調查對象(初生幼苗,
或直徑≥ 1 cm之樹苗及稚樹)上皆有所區隔。因
此,仍有必要針對不同經營方式下的林分直接 進行比較性研究。
綜上所述,對於與檜木林經營實務密切相 關的前述科學問題,相關的研究進展顯然仍需 累積更多成果,方可提供資源管理單位更完整 的決策知識。為此,本研究試圖同時納入人為 經營林分與天然林為研究對象,分析不同檜木 林經營模式對於地被植物多樣性與林木更新的 影響。希冀能為台灣天然檜木林保育與經營議 題,提供更多的學研知識基礎。本研究將針對 以下問題進行探討,並且對目前林分經營現況 提出建議:
1. 在不同經營模式下,檜木林的地被植物多樣 性是否具有明顯差異?其物種組成有何不同?
2. 在不同經營模式下,林分的天然更新狀態是 否有明顯差異?檜苗於天然老齡林的建立情形 如何?
材 料 與 方 法
本研究選擇在具有長期林業經營歷史,而 且目前仍保有大面積天然檜木林的棲蘭山地區 進行研究。棲蘭山地區的天然檜木林為臺灣現 存最大面積的原始檜木林,同時具有台灣扁柏 (Chamaecyparis obtusa var. formosana)與紅檜 (C. formosensis)兩種原生檜木的分布,此地的 人工檜木林也是兩種均有栽植(Wang 2000, Lin et al. 2016)。退輔會榮民森林保育事業管理處 過去在此地的檜木林經營,除了早期的伐木收 穫 作 業 之 外 , 亦 著 力 於 檜 木 人 工 林 建 造 及 撫 育、柳杉與檜木混合複層林之營造、檜木天然 更新造林(包括留伐及擇伐天然更新作業、枯立 倒木整理更新作業)等各種保續經營計畫(Huang 1995, Lin et al. 2016)。因此,棲蘭山為臺灣目 前最重要的檜木林天然分布與人為經營區域。
本研究於4種不同經營模式下的檜木林型進 行取樣調查:
1. 皆伐後人工栽植林(plantation after clear cutting, PCC;簡稱皆伐後人工林):位於棲 蘭山100線林道約25.5 km處,屬於太平山事 業 區 第39林班。此林分於1974年進行皆伐
作業,並於隔年進行造林,栽植樹種以紅檜 為主,鄰近地區存有天然檜木林。2005年曾 對造林木進行修枝,但未曾進行過任何伐採 作業。2016年調查林分狀況,檜木平均密度 為1152 ha-1,平均胸高直徑(以下簡稱胸徑) 22.3 cm,平均樹高11.8 m (未發表資料)。
2014年本研究於此區進行調查時,此林分已 歷經30~40年未有大規模人為擾動。
2. 擇伐後天然更新林(naturally regenerated stand after selective cutting, NSC;簡稱擇伐 後更新林):位於棲蘭山170線林道約0.5 km 處,屬於大溪事業區第94林班。此地於1976 年劃設約8公頃之試驗地進行擇伐作業,分 成弱度(20~25%,以胸高斷面積計算)、中 度(30~35%)、強度(40~45%)、對照區等4種 擇伐程度;並於隔年春季進行整地處理以實 施天然下種更新試驗(Hung 1982, 1984, Lo- Cho et al. 1989)。依據12年後之調查,天然 更新檜木稚樹以台灣扁柏為主,佔95% (Lo- Cho et al. 1989)。由於稚樹密度過高,1992 年再進行除伐、修枝等撫育作業試驗,將檜 木平均密度調整為2000、3300 ha-1與未除伐 (11,040 ha-1) 3種處理(Lo-Cho et al. 1999)。
2013年調查林分狀況,3種處理林地的檜木 平均密度分別為1770、3060、5510 ha-1,平 均胸徑10.6、9.3、7.3 cm,平均樹高7.1、
6.7、5.7 m (未發表資料);1976年擇伐後保 留的檜木母樹,估計每公頃約有90株。2013 年本研究於此區進行調查時,此林分已歷經 20年未有人為擾動。
3. 枯立倒木整理後老齡林(old-growth forest after removing snags and logs, RO;簡稱枯 立倒木整理林):位於棲蘭山170線林道約1 km處,亦屬於大溪事業區第94林班。此林分 原為天然老齡林,但曾在1984年進行枯立倒 木整理作業(Huang 1995, Lin et al. 2016)。
我們於2010~2012年設置50×50 m樣區調查 檜木分布,平均密度為692 ha-1,平均胸徑 15.3 cm (最大達100.6 cm),冠層樹高估計約 20~30 m;區內胸徑< 10 cm之小徑木數量豐 富,佔71% (未發表資料)。此區檜木以台灣
扁柏為主,佔68%;紅檜多為小徑木,經觀 察其植株配置,可知部份為人工栽植。2013 年本研究於此區進行調查時,此林分已歷經 近30年未有人為擾動。
4. 天然老齡林(natural old-growth forest, NO):
位於棲蘭山170線林道約1 km處,亦屬於大 溪事業區第94林班,為從未施以人為經營的 殘留原始檜木林地。我們於2010~2012年設 置50×50 m樣區調查檜木分布,平均密度 為720 ha-1,平均胸徑22.4 cm (最大達96.2 cm),冠層樹高估計約20~30 m;區內胸徑<
10 cm之小徑木數量亦多,佔39% (未發表資 料)。此區檜木皆為台灣扁柏,比對Chen et al. (2002)與Wei and Chen (2007)之植群分類 成果,此林分應屬於「木荷―台灣扁柏型」
植群。
其 中 ,NSC、RO、NO林分的間隔不到1 km,PCC與此3種林分距離約2 km。由於本研 究中不同林型的地點十分鄰近,因此應可將各 林分之區域性氣候、周遭天然植被組成等背景 條件視為相同。這樣的取樣調查設計有助於本 研究釐清不同經營模式對於林分的影響。
調查工作於2013年9月及2014年8月進行。
每一種檜木林型設置10個2×2 m樣區,位置均 從既有的每木調查樣區選定。其中PCC、NSC 林分的樣區為隨機配置,RO、NO林分則採系 統 配 置 , 同 一 林 分 內 地 被 樣 區 的 最 小 間 距 皆
≥ 10 m。樣區調查時記錄各種微生育地之覆蓋 度 , 共 區 分 成 土 壤 、 苔 蘚 、 木 質 殘 材(包括倒 木)、樹幹基部4型,並進行以下兩項調查:
1. 地被植物組成:調查地面草本層與灌木層植 物的種類與覆蓋度(包含草本、藤本、灌木及 胸徑< 1 cm的喬木種樹苗)。覆蓋度採用百分 率級距來記錄,每達到5%為一級,但未達 5%的少量種則計為1%;若是明顯未達1%的 偶見種,則記為0.1%,例如少量的初生幼苗 或小型草本植物。
2. 天然更新樹苗組成:調查胸徑< 1 cm的喬木 種天然更新樹苗,包括檜苗與其他喬木類闊 葉樹種的樹苗。除了鑑別樹種、計數樹苗數 量之外,也記錄樹苗所處的微生育地類型。
以上共調查到40個樣區的地被植物組成(覆 蓋度)與天然更新樹苗組成(株數)資料,可供分 析不同林型的植物多樣性、物種組成以及林分 的天然更新狀態。在植物多樣性、樹苗數量、
微生育地覆蓋度的比較上,本研究採用Kruskal- Wallis單向等級變異數分析(Kruskal-Wallis´
one-way ANOVA by ranks)檢定林型間差異。
此 法 為 無 母 數 統 計 方 法 , 適 用 於 統 計 分 布 形 式不明或少量之資料(Legendre and Legendre 2012)。各樣區之植物多樣性除了直接計數物 種數(species richness)之外,同時也採用多樣 性指數Hill diversity of order 1 (H1)來評估(Jost 2006)。多樣性指數之計算納入了各物種的數 量比例(地被植物為覆蓋度,樹苗為株數),因 此與物種數相比較可反映出物種組成的均勻度 (evenness)。而選用H1多樣性指數可計算出群落 之「有效種數」(effective number of species,
或稱numbers equivalent),代表該群落等同於多 少種「數量相等之物種」(equivalents)所組成,
因此為一更直覺、適合於比較多樣性的評估方 法(Jost 2006, Ellison 2010)。
前述之統計分析在空間尺度上是以單一樣 區做為比較的單元。然而,物種多樣性會隨著 空 間 尺 度 而 變 化 , 且 具 有 「 非 相 加 性 」(non- additive)的統計性質,故而會呈現出「種數―
面積曲線關係」(species-area relationship)。
因此,本研究亦採用sample-based rarefaction curve分析方法來比較不同取樣面積下的物種數 變化,其中的統計檢定是以平均值±1.96倍標 準差(mean±1.96×SD)做為95%信賴區間,以 進行林型間的比較(Magurran 2004, Kindt and Coe 2005)。
在 物 種 組 成 分 析 方 面 , 本 研 究 採 用 principal coordinate analysis (PCoA)多變量分 析,比較各林型之組成變異情形以及林型間的 差異。本研究使用Jaccard相異性指數,分別先 以地被植物之覆蓋度資料、天然更新樹苗之株 數資料計算樣區間的相異性矩陣,再進行PCoA 分析(McCune and Grace 2002, Legendre and Legendre 2012)。
以 上 統 計 分 析 是 以R 3.4.1統計語言執行
完成(R Core Team 2017),其中的rarefaction curve分析使用BiodiversityR擴充套件(Kindt and Coe 2005),PCoA分析則使用vegan擴充套件 (Oksanen et al. 2017)。
結 果
在40個調查樣區內,總計記錄到102種(50 科)地被植物,以及19種(11科)、346株天然更 新樹苗。依據H1多樣性指數的估算,地被植物 的整體有效種數為17.3種,而天然更新樹苗則
僅有5.8種;與實際調查到的物種數兩相比較,
可知地被植物與樹苗的物種組成情形相當不平 均,導致多樣性指數偏低。
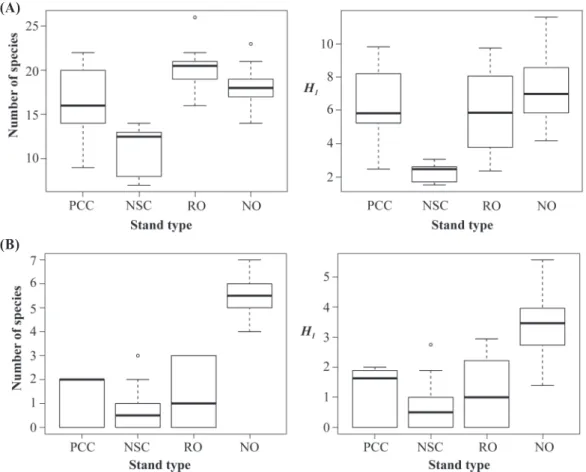
在地被植物多樣性方面,4種檜木林型的地 被物種數在單一樣區的比較下呈現顯著的差異 (p < 0.001),其中以枯立倒木整理林(RO)略高於 天然老齡林(NO)、皆伐後人工林(PCC),而以 擇伐後更新林(NSC)明顯最低(Fig. 1A)。若是改 以H1多樣性指數來比較,不同林分之間同樣存 在顯著差異(p < 0.001),只是改由NO的有效種 數略高於RO、PCC,而NSC仍然為明顯最低者 (A)
Fig. 1. Comparison of the diversity of ground vegetation (A) and natural seedlings (B) among different Chamaecyparis forest stand types. In each panel, the plant diversity of the 4 stand types significantly differs (see text). The ordinates represent 2 diversity measures in 2×2-m plots: left panel, numbers of species; right panel, Hill diversity of order 1 (H1). The abscissas represent the 4 stand types: PCC, plantation after clear cutting; NSC, naturally regenerated stand after selective cutting; RO, old-growth forest after removing snags and logs; NO, natural old-growth forest.
(B)
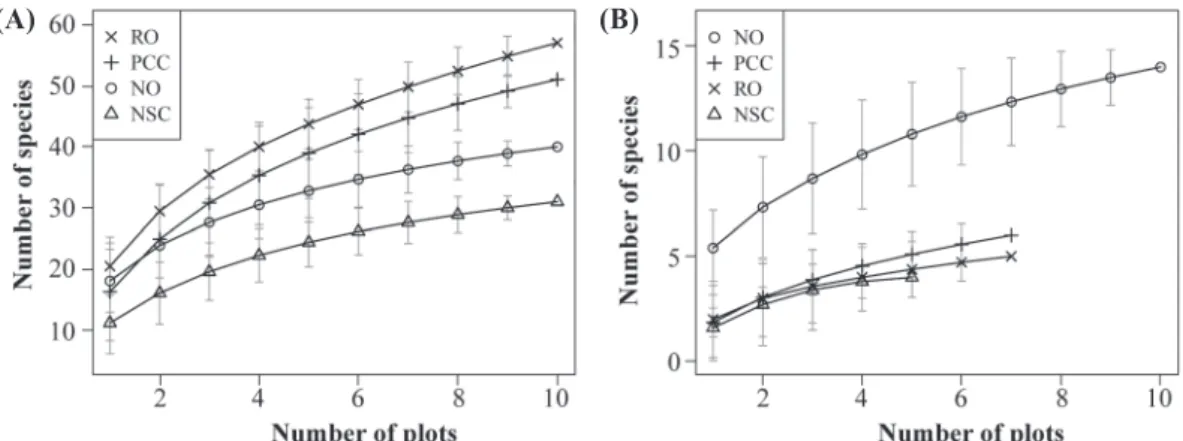
(Fig. 1A)。Rarefaction curve分析結果顯示,在 本研究的取樣範圍內累計物種數最多的林型仍 舊是RO,最低者仍是NSC,但是PCC卻有超越 NO、僅次於RO的情形;而且PCC與RO雖然維 持著4~6種的差距,但其差異均未達到統計顯著 性(Fig. 2A)。另一方面,隨著分析範圍的擴大,
4種檜木林型之間的物種數差異則有逐漸增大的 趨勢。NSC與NO分別在分析範圍大於4個與8個 樣區以後與物種較多的RO、PCC呈現出顯著差 異(p < 0.05),至於NSC與NO之間,則是在大於 6個樣區以後達到顯著差異(p < 0.05; Fig. 2A)。
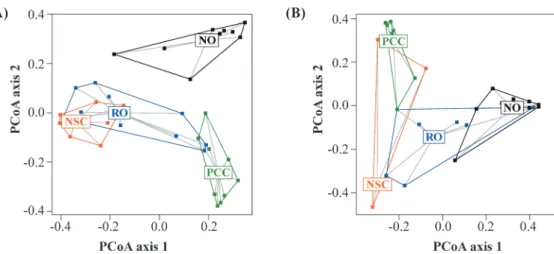
若從地被植物的物種組成來看,PCoA分 析 結 果 顯 示NO的組成與其他林型有明顯的區 隔 ; 而 在 其 他3種林型之間,NSC與PCC也是 有 明 顯 差 異 , 但 居 間 的RO則與兩者皆有重疊 之處(Fig. 3A)。雖然NO、RO、NSC這3型的 最優勢地被植物皆為台灣瘤足蕨(Plagiogyria formosana),但是它在NSC呈現明顯的單一優 勢,總覆蓋度達到75%,與在NO (18%)、RO (40%)的優勢程度不同。此外,NO尚有白花八 角(Illicium anisatum)、台灣杜鵑(Rhododendron formosanum)、深山野牡丹(Barthea barthei)等 優勢種,在其他伴隨的常見種上也與其他兩型
不同(Appendix 1)。值得注意的還有,NO的物 種組成中有部份植物在其他林型均未調查到,
例如台灣杜鵑、蔓竹杞(Myrsine stolonifera)、
台 灣 樹 參(Dendropanax dentiger)、厚皮香 (Ternstroemia gymnanthera)等等。RO的優勢 種 組 成 雖 與NO較為相近,但是在常見種上與 NO明顯不同(Appendix 1);同時也出現許多 喜 好 強 光 、 開 闊 環 境 的 物 種 , 例 如 火 炭 母 草 (Polygonum chinense)、菝契(Smilax china)等 常見種,以及烏皮九芎(Styrax formosana)、戟 葉蓼(Polygonum thunbergii)、芒(Miscanthus sinensis)、數種懸鉤子類(Rubus spp.)等少量種,
這些都是在其他林型罕見的。PCC的優勢種包 括了玉山箭竹(Yushania niitakayamensis)、假 柃木(Eurya crenatifolia)、魚鱗蕨(Acrophorus stipellatus),與其他3型不同(Appendix 1);雖然 台灣瘤足蕨的數量亦多,但總覆蓋度僅約10%。
在林木天然更新方面,總計346株樹苗中 有188株(54%)是檜苗。值得注意的是,棲蘭 山地區雖然同時具有台灣扁柏與紅檜的天然族 群,但是在本研究的樣區範圍內調查到的檜苗 均為台灣扁柏。其他的天然更新樹苗則是以白 花八角(10%)、台灣杜鵑(7%)、紅淡比(Cleyera
Fig. 2. Sample-based rarefaction curves of ground vegetation (A) and natural seedlings (B) among different Chamaecyparis forest stand types. In each panel, the plant diversity of the 4 stand types significantly differs (see text), and the differences increase as the spatial scale increases. The ordinates represent the accumulated numbers of species, and the abscissas represent numbers of 2×2-m plots sampled. The gray vertical segments are 95 confidence intervals composed of the mean±1.96 standard deviations. Abbreviations of stand types are explained in Fig. 1.
(A) (B)
Fig. 3. Two-dimensional ordination plots of a principal coordinate analysis (PCoA) of the ground vegetation (A) and natural seedlings (B). Data points represent 2×2-m plots sampled among different Chamaecyparis forest stand types. The PCoA axes 1 and 2
explained 21 and 13 of the total variation, respectively. The PCoA was performed on the dissimilarity matrix of the Jaccard index computed from coverage data (A) and abundance data (B). Abbreviations of stand types are explained in Fig. 1.
japonica; 6%)較多。就單一樣區內的樹苗物種 數來分析,不同檜木林型之間呈現出顯著差異 (p < 0.001),其中以NO明顯最高,其他林型的 樹種數則都十分低落(Fig. 1B)。若以H1多樣性 指數來比較,仍然維持相同的顯著差異情形(p
< 0.001; Fig. 1B)。Rarefaction curve的分析結 果更清楚地顯示,NO的天然更新樹種數遠高於 其他林型,在分析範圍大於2個樣區以後即達到 統計顯著性,而且差距逐漸加大(p < 0.05; Fig.
2B)。至於其他3種林型之間的差異則不明顯,
更由於有部份樣區完全未調查到任何樹苗(例如 NSC有半數樣區沒有樹苗),因此這些林型的種 類面積曲線也較短(Fig. 2B)。以PCoA分析樹苗 的物種組成,亦可發現NO與PCC、NSC有明顯 區 隔 , 但 是RO介於兩者之間,與兩群各有重 疊之處(Fig. 3B)。NO的樹苗組成(共267株)以 台灣扁柏為單一優勢種,佔61%,其他常見種 則有白花八角、台灣杜鵑、紅淡比、太平山冬 青(Ilex sugerokii var. brevipedunculata)等,與 其他林型明顯不同(Appendix 1)。RO、PCC、
NSC由於樹苗量過少(分別僅調查到42、23、
14株),因此難以界定出優勢與常見樹種,不過 RO的樹苗組成與NO較為相近,同樣以台灣扁
柏為數量最多者(24株),故可界定為該林型之常 見種(Appendix 1)。
除了在多樣性、物種組成上的差距之外,
不同檜木林型的林木天然更新狀態也有明顯差 異。不同林型之間的樹苗分布數量差異極為懸 殊(p < 0.001),其中以NO的樹苗密度最高,平 均66,750 ha-1(數量佔全部的77%),其餘3種林 型的天然更新情形則都不佳,平均6583 ha-1(佔 23%) (Fig. 4A)。檜苗(台灣扁柏)的天然更新情 形在不同林型間的差異更為顯著(p < 0.001),同 樣是以NO的檜苗密度最高,平均41,000 ha-1(總 數164株,佔全部檜苗的87%),RO次之,平均 6000 ha-1(總數24株,佔13%),其餘兩種林分則 完全未調查到檜苗(Fig. 4B)。若是將天然更新 樹苗最多的NO排除在分析之外,則其他3種林 型在樹苗密度上並未有顯著差異(p = 0.37);但 是在檜苗密度上,由於PCC、NSC未調查到檜 苗,因此仍與RO之間存有顯著差異(p < 0.01)。
至於樹苗生長的微生育地類型,約有95%
的樹苗是建立在木質殘材或土壤之上,苔蘚與 樹幹基部則少見到樹苗發生。值得注意的是,
檜 苗 與 其 他 樹 種 的 苗 木 呈 現 出 不 同 的 偏 好 , 91%的檜苗是建立在木質殘材,而其他樹苗則
(A) (B)
只有48%生長在木質殘材,另外47%則是建立 在土壤介質上(Fig. 5)。就土壤、木質殘材微生 育 地 在4種林型的覆蓋度來分析,土壤的平均 覆蓋度為60%,林型間差異顯著(p < 0.05),其 中以NSC最高(73%),其他3型則差距不大(介 於51~60%)。木質殘材的平均覆蓋度為33%,
在NO及RO的覆蓋度較高(43%、35%),PCC及 NSC較低(30%、22%),但因為各林型之樣區變 異程度相當大(標準差介於13~22%),林型間並 未達到顯著差異(p = 0.09)。
討 論
綜 合 以 上 分 析 結 果 可 知 , 在 不 同 的 經 營 模式下,棲蘭山檜木林的林下地被植物、天然 更新樹苗已發展出明顯不同的物種多樣性與組 成 。 受 到 人 為 作 業(枯立倒木整理、擇伐、皆 伐)擾動過的林分,即使經過長期的自然恢復,
如今在同一地區下的不同林型之間仍然呈現明 顯差異。在地被植物方面,枯立倒木整理後的 老齡林(RO)在物種數上高於其他林型,皆伐後 人工林(PCC)則與之差距不大,而擇伐後更新 林(NSC)則明顯最低。天然老齡林(NO)與其他 林型相比,雖然物種組成上較為均勻(因此多樣 性 指 數 較 高),但是隨著分析的空間尺度加大
(rarefaction curve分析結果),累計物種數不但 比枯立倒木整理林低,差距也逐漸加大,甚至 也低於皆伐後人工林。不過值得注意的是,天 然老齡林的地被植物多樣性雖然較低,它的物 種組成卻顯然與其他林型皆有區別,包含多種 其他林型未調查到的演替中、後期植物。這顯 示在經過人為作業擾動過的檜木林分內,地被 植群雖然可能具有較高的多樣性,但是在物種 組成上卻尚未能發展成接近原始天然林的組成 (Nagaike et al. 2005)。這可能是因為各林分仍 處於恢復階段中,或是因為生長資源受限(例如 林下光度、空間)而影響了植生的恢復程度,以 下將分別進行討論。
以枯立倒木整理林來看,此林型除了存留 部份天然老齡林植物種類之外,也由於倒木的 移除、地表植被的清理而重新釋放出林下生長 空間(Huang 1995, Lin et al. 2016),引入了演替 前期的物種,因此具有最高的地被植物多樣性 (Wei and Chen 2012)。Wei and Chen (2012)同 樣是研究棲蘭山不同人為擾動強度下林分的植 物多樣性,發現冠層、次冠層、灌木層植物均 以天然林的多樣性最高,但在地被層則以整理 過枯立倒木的檜木保育更新地(位於130林道)最 高。他們的地被層分析結果與本研究在枯立倒 木整理林(位於170林道)所觀察到的較高多樣性 Fig. 4. Comparison of seedling densities of all naturally regenerated tree species (A) and Chamaecyparis spp. (B) among different Chamaecyparis forest stand types. In each panel, seedling densities of the 4 stand types significantly differ (see text). The ordinates represent numbers of seedlings per hectare. Note that no cypress seedlings were found in the PCC and NSC stands, and that all cypress seedlings investigated in this study were Chamaecyparis obtusa var. formosana. Abbreviations of stand types are explained in Fig. 1.
(A) (B)
相當類似。至於地被層以上的樹種組成,他們 認為受到人為擾動的林分僅在冠層樹種組成上 趨近於天然林,次冠層以下的植被則仍處於恢 復階段,與天然林相差甚多。綜合以上研究可 知,原始檜木林在受到人為作業擾動後的植被 恢復過程十分緩慢,即使歷經近30年的自然過 程(如本研究的枯立倒木整理林),仍然與天然老 齡林的物種組成有明顯差距。
關於其他兩種人工林分的情形,皆伐後人 工林的優勢種組成與其他林型差異最大,但是 其中與枯立倒木整理林的差異較小,兩者的多 樣性也十分相近。在Wei and Chen (2012)的研 究成果中,兩處人工林的地被植物多樣性呈現 出一高一低的分歧情形,但由於該研究並未提 供詳細的人工林背景資訊(林齡、林分結構等),
因此本研究無法直接與之比較為何不同人工林 呈現出歧異情形。基於這類人工林是在皆伐原 有林分後以人工栽植造林樹種所育成,其林下 植被均是從清理後的裸露林地上再重新自然演 替,因此我們推論影響林下植被恢復的主要因 子應包括物種來源(例如原地之種苗庫、周圍森 林植物之遷入)、各項作業造成的林地環境條件 (例如林下生長空間、光度、伐採後殘材是否移 除、是否進行整地)以及演替時間。就本研究所
比較的4種林型來看,所有林分皆位在十分鄰近 的地區(距離< 2 km),且周圍均有天然檜木林 分布,因此在地被植物的遷入來源上應該不致 受限。而本研究調查之人工林分,其植株密度 適中,林地亦未堆置過多殘材,所以林下生長 環境並無明顯障礙。再加上該林分已有30~40年 未有大規模人為擾動,是3種人為作業過林分中 演替時間最為長久者,因此在植物多樣性與物 種組成上較相近於多樣性最高的枯立倒木整理 林。不過,此林分中的最優勢種玉山箭竹目前 的覆蓋度雖然不高(平均19%),未來仍需注意是 否有擴張的趨向。
至 於 地 被 植 物 多 樣 性 最 低 的 擇 伐 後 更 新 林 , 此 林 分 以 台 灣 扁 柏 的 天 然 更 新 小 徑 木 為 主要組成,過去在實施天然下種更新作業後,
林地即培育出十分稠密的檜木稚樹(Lo-Cho et al. 1989),即使再施以除伐作業(Lo-Cho et al.
1999),目前植株仍然過於密集,為所有林型中 密度最高者。此外,除了部份因倒木而形成的 孔隙之外,林下受光情形也明顯不佳。因此,
此林分的地被植物組成相當單純,以台灣瘤足 蕨佔絕對優勢,其餘種類則稀疏分布。即使在 此 林 分 內 的 不 同 檜 木 密 度 處 理 區 塊 , 也 無 明 顯差異(PCoA結果亦顯示為變異範圍最小的林 Fig. 5. Comparison of proportions of seedlings in different microhabitat types for cypress seedlings (Chamaecyparis obtusa var. formosana) and seedlings of other tree species. Cypress seedlings showed a distinct microhabitat preference from other seedlings.
Microhabitat type
Proportion of seedlings ()
型)。綜上所述,此林分可能是受到林下空間過 於狹密與光度上的共同限制,導致形成最為貧 乏的地被植物組成及多樣性,若未再施以人為 作業改善,未來將難以朝向天然林植生狀態的 方向發展。
在天然更新樹苗方面,則是以天然老齡林 的物種數遠高於其他林型,而擇伐後更新林的 物種數同樣是各林型之中最低者。很明顯地,
不同林型間樹苗多樣性的差距來自於天然更新 樹苗在數量上的懸殊差異,其中尤以天然老齡 林的樹苗量遠高於其他曾經歷人為經營作業的 林 型 。 至 於 不 同 林 型 的 天 然 更 新 樹 種 組 成 , 與 地 被 植 群 的 情 形 相 似 , 天 然 老 齡 林 的 樹 苗 組成與擇伐後更新林、皆伐後人工林存有明顯 區別,但是與枯立倒木整理林則較為相近,同 樣具有天然更新檜苗。以上結果反映出即使是 在相同的氣候、周遭植被等背景條件下,不同 的檜木林經營模式確實會明顯改變地被植群與 樹苗的多樣性及物種組成,也影響了林下天然 更新樹苗的多寡(Nagaike et al. 2005, Liu et al.
2015)。由於本研究調查到的天然更新樹苗有半 數以上是檜苗(台灣扁柏),其他樹種皆佔少數,
而檜木天然更新歷來皆是檜木林經營的重要議 題,因此以下將聚焦在不同林型的檜苗天然更 新情形進行討論。
不同經營模式下的檜木林呈現出極為懸殊 的檜苗更新情形,在本研究的調查範圍內,檜 苗僅出現在兩種老齡林型,而且絕大多數是建 立在原始、未經人為擾動過的天然老齡林分。
若從樹苗生長的微生育地來看,九成以上的檜 苗是建立在木質殘材與倒木之上。早期日本學 者松浦作治郎曾在倒木及根株上進行檜木人工 播種實驗,發現台灣扁柏在倒木上的發芽率高 於在根株上(Kuo 1995)。而在日本木曾地區檜木 林的研究也指出,建立在倒木上的檜苗死亡率 較低(Yamamoto 1993)。Liu (1975)針對全台天然 檜木林主要分布地點進行調查,即發現台灣扁 柏在許多天然林分內的更新情形良好,而且樹 苗、稚樹大多建立於倒木之上或者周遭的土壤 擾動區塊。Chang et al. (2001)曾針對棲蘭山不 同時期枯立倒木整理林分調查檜木種子雨、幼
苗數量與存活動態,發現留存在地表的枯立倒 木與殘材有利於檜苗的存活。Liao et al. (2003a, 2003b)在棲蘭山鴛鴦湖區域的天然檜木林研究也 發現倒木是台灣扁柏苗木更新的重要基質,並 建議檜木林之經營宜於林地內留置倒木,可達 成較有效的檜木天然更新作業。本研究在不同 經營模式下的檜木林分進行天然更新檜苗數量 的比較,同樣發現檜苗主要建立在木質殘材及 倒木上,與其他闊葉樹種不同,顯見檜木對於 林下更新的基質選擇確實有明顯偏好。
本研究調查的4種林型之間,檜苗數量的 多 寡 次 序 與 地 表 木 質 殘 材 覆 蓋 度 的 高 低 次 序 大 致 相 同 , 然 而 其 中2種人工林分完全未調查 到 檜 苗 , 而 這 些 林 分 的 地 表 也 存 有 相 當 程 度 的木質殘材,與老齡林的差異亦未達到統計顯 著性。可見雖然木質殘材是檜苗明顯偏好的微 生育地資源,但是單從在林地上的覆蓋程度並 無法完全解釋天然更新檜苗的數量差距。殘材 的大小、腐朽程度乃至於養份含量等基質特性 很可能對於種子萌發、檜苗建立也有重要影響 (Chang et al. 2001, Liao et al. 2003b)。除此之 外,過去有關檜木更新作業的研究均指出,若 林地空間的整理(如伐採收穫、枯立倒木整理或 整地作業等)能配合上檜木結實的豐年期,可促 進天然更新作業的成功(Lo-Cho et al. 1989, Kuo 1995, Chang et al. 2001);本研究的擇伐天然更 新林即是一成功案例。綜合以上研究,我們推 論檜木天然更新的關鍵因子應包括充足的種子 來源、林地生長空間的釋放、適合的微生育地 三項。其中前兩項的發生時間必須一致,否則 不易達到有效的天然下種更新。而木質殘材則 是提供鬱閉林分內檜木下種更新最合適的微生 育地,除了形成較佳的微環境(如光度較高),
也有利檜苗免於其他植物的競爭壓抑(Liao et al.
2003a, 2003b)。但是木質殘材的質與量必須同 時兼具,才能發揮助益(Chang et al. 2001)。
3種人為作業過的林分目前檜苗數量大不如 天然老齡林分,除了林地上留存木質殘材的質量 差異可能造成影響之外,缺乏充足種子來源與生 長空間兩項條件的搭配也可能是重要原因。以擇 伐後更新林來看,此林分最初雖是以天然下種
更新作業成功育成(Lo-Cho et al. 1989),然而之 後發展成植株密集的林分狀態,即使再經過除伐 作業,目前林況仍是過密。擇伐後更新的幼齡檜 木植株(< 40年生)能否結實下種雖屬未知,但林 分內保留的母樹仍可持續下種,直接提供大量的 種子來源。但即使在這樣的下種條件下,樹冠完 全鬱閉、林分過度擁擠的林況會大幅減少種子順 利散播至林下的數量,林下生長空間與光度的不 足也會進一步限制種子萌發與幼苗建立,形成此 林分檜木更新的雙重障礙。相較之下,皆伐後人 工林並未保留檜木母樹,必須仰賴周圍天然林的 檜木種子以風力飄散至此,能夠抵達的種子量已 然受到限制,再加上鬱閉樹冠的攔阻,導致此林 分天然下種的數量必然不如於其他3種林型。因 此,此林分即使林下生長空間與光度環境較優於 擇伐後更新林(從檜木植株較疏,以及天然更新 樹苗在數量、種數皆較高可知),仍舊難以促成 檜苗更新。
至 於 仍 然 保 有 完 整 天 然 老 齡 檜 木 分 布 的 枯立倒木整理林,過去實施整理作業的目標即 在於釋放出林下生長空間以促進檜木天然更新 (Huang 1995, Lin et al. 2016)。若是從林分目前 的檜木徑級結構與天然老齡林相比,兩者皆具 有充足數量的小徑木(請參見「材料與方法」之 林型描述),然而因為有部份檜木為人工栽植,
因此本研究並無法對過去整理作業的更新成效 進行嚴謹評估。但是從本研究針對目前林分內 檜木幼苗(胸徑< 1 cm者)的調查結果來看,此 林型雖有檜苗分布,但數量明顯少於天然老齡 林。我們推論是因為整理作業後釋放出的生長 空間多年來已被各種天然更新樹木與地被植物 重新佔據,而且可以提供避免與其他植物競爭 的木質殘材資源又已不復如天然林狀態,因此 目前林況條件較不利於以天然下種方式建立檜 木幼苗。而天然老齡林,雖然林下保持原始植 生的狀態,並未釋出生長空間(除了少數倒木形 成的孔隙),但由於地表累積許多大型且質地亦 多樣化的天然生木質殘材,與人為作業後餘留 枝梢殘材的情況大不相同,再加上林地長期持 續著大量天然下種過程(檜木老齡林下種密度可 高達2000~3000 seeds m-2 yr-1; Chang et al. 2001,
Lin et al. 2016),因此能夠存有較多量的檜苗。
綜 上 所 述 , 目 前 不 同 檜 木 林 型 在 地 被 植 物與樹苗的多樣性及組成、林木天然更新情形 均具有明顯差異。在地被植物方面,人為作業 過的林型可釋放出林下生長空間、引入不同演 替階段的物種,因此在植物多樣性上有機會高 於天然老齡林。但若未能適當控制林分密度,
導致林下空間過於狹密,則反而對多樣性有明 顯的負面影響。而在林木天然更新方面,無論 是樹種多樣性或樹苗量,人為作業過的林型都 明顯不如天然老齡林,檜木的樹苗更新情形亦 然。此外,若從物種組成來看,無論是地被植 物或樹苗,人為作業過的林型即使經過20~40年 的自然恢復,仍然未能發展成近似天然老齡林 的狀態。因此,我們認為天然檜木老齡林具有 獨特的生態特性,目前檜木天然更新的狀態亦 未見困難,所以一方面應保護現狀、避免受到 人為作業影響,另一方面也應實施長期生態監 測,關注當前氣候變遷趨勢是否會對天然檜木 林生態產生負面效應。枯立倒木整理過的老齡 林在本研究中呈現出最高的地被植物多樣性,
顯見人為作業的擾動可以在天然林下層營造出 不同演替階段的植生。若能適當運用於與天然 林區塊組成鑲嵌式地景,並輔以監測檜木天然 更新情形,應可將此種正面效應從植物群落擴 及其他生物群聚(例如提供野生動物更多樣化的 食 物 來 源 與 棲 地),提升整體森林生物多樣性 (Yuan et al. 2004, Chen et al. 2015, Teixeira et al. 2017)。皆伐後人工栽植林之建造目的在於培 育優良、均質的木材生產用檜木,因此後續之 經營應依據主林木生長情形進行中、後期撫育 作業,適時調整林分密度與結構。惟亦應留意 玉山箭竹於地被層之生長情形,避免其全面性 擴展、損及地被植物多樣性並造成未來森林作 業上的阻礙。至於擇伐後天然更新林,目前林 木過於密集,林下植生環境明顯不佳,因此無 論是地被植物或樹苗,在多樣性、組成與更新 數量上都呈現最為貧乏的情形。再者,林分過 密的狀態也不利於檜木生長(植株明顯小於林齡 相近的皆伐後人工林)與新生檜苗的再建立。因 此,此林分亟待實施撫育作業方可改善林況。
結 論
本 研 究 顯 示 不 同 的 森 林 經 營 模 式 對 於 棲 蘭山地區檜木林的地被植物與樹苗多樣性及組 成,以及林木天然更新狀態皆有明顯影響,在 同一地區下的不同林型間呈現出顯著差異。而 人為作業過的林分即使歷經20~40年的恢復,
仍然未能發展出近似於天然老齡林的狀態。基 於天然老齡林的獨特生態特性,我們建議應予 保護並長期監測其動態變化。若能適當運用現 有枯立倒木整理後老齡林所營造出的不同植生 階段,則可在天然檜木林區域內增加生物棲地 種類,進而提升整體生物多樣性。至於人為經 營作業介入較多的皆伐後人工栽植林與擇伐後 天然更新林,應積極進行中、後期撫育工作以 促進主林木之生長,並可避免林分之植物多樣 性降低與箭竹覆蓋範圍之擴張。此外,本研究 的取樣對象與調查面積仍不夠廣泛,我們建議 應進行更多檜木林的比較性研究與長期生態監 測,例如調查更多不同經營模式、地區或林齡 序列(chronosequence)下的林分。如此將有助吾 人獲致更為堅實的研究成果,為台灣的檜木林 (無論是天然或人工)提出同時符合林學與生態學 原則的合理經營方式。
謝 誌
本研究承蒙退輔會榮民森林保育事業管理 處及棲蘭山工作區於調查工作期間提供各項行 政協助,特別感謝棲蘭山工作區陳駿銘主任與 同仁協助處理現場調查事故,本研究方可順利 執行。筆者並感謝兩位審查者提供許多寶貴意 見,使本文之質量大幅改善。本文為農業委員 會林業科技計畫(100農科-8.4.2-森-G2、102農 科-13.2.1-森-G6、103農科-13.2.5-森-G1)研究 成果之一。
引 用 文 獻
Chang NH, Hsui YR, Horng FW, Yu HM, Ma FC. 2001. Natural seeding and seedling
occurrence in the Chamaecyparis forest at Chilan Mt. area. Taiwan J For Sci 16(4):321-6.
Chang SC, Tseng KH, Hsia YJ, Wang CP, Wu JT. 2008. Soil respiration in a subtropi- cal montane cloud forest in Taiwan. Agric For Meteorol 148(5):788-98.
Chen HH, editor. 1999. Proceedings of the symposium on snags and natural resource conservation. Taipei, Taiwan: Department of Forestry, National Taiwan University. 197 p. [in Chinese].
Chen TY, Hsu HY, Wu SL. 2002. Study on the Chamaecyparis forest of 170 Roads of Chilan Shan. J Ilan Inst Technol 9:259-75. [in Chinese with English summary].
Chen YF. 1999. Collected works on the nation- al campaign for saving the Chi-lan Chamaecy- paris forest, vol. 1. Kaohsiung, Taiwan: Ai-Chi Books Publisher. 206 p. [in Chinese].
Chen YM, He WL, Wong RH, Kuo CN.
2015. Mammal assemblages of different for- est types in the Cilan cypress forest. Taiwan J For Sci 30(1):75-88. [in Chinese with English summary].
Council of Agriculture. 2000. Reports on evaluating the forest conservation practice of removing snags and logs in Chi-lan-shan. Tai- pei, Taiwan: Council of Agriculture, Executive Yuan. p 53. [in Chinese].
Ellison AM. 2010. Partitioning diversity. Ecol- ogy 91(7):1962-3.
Horng FW, Ma FC, Yu HM, Hsui YR, Chang NH. 2000. An estimation of original Chamaecyparis forest area In Taiwan and its implication for conservation. Q J Chin For 33(1):143-53. [in Chinese with English sum- mary].
Huang CH. 1995. The distribution and man- agement of Chamaecyparis forests under the administration of the Forest Development Administration. In: Kuo PC, editor. The most valuable five conifer timber species in Taiwan.
Taipei, Taiwan: The Chinese Forestry Associa- tion. p 181-98. [in Chinese].
Hung LP. 1982. Method of natural regenera- tion of natural cypress on upper basin of Shin- Men Reservoir. J Agric Assoc China (New Series) 117:49-54. [in Chinese with English summary].
Hung LP. 1984. The effect of improvement by selective cutting methods for the natural forest of cypress on high mountain area in Taiwan. Q J Chin For 17(4):47-56. [in Chinese with Eng- lish summary].
Jen IA. 1995. Expectation and historical re- view of cypress (Chamaecyparis spp.) timber production in Taiwan. Bull Taiwan For Res Inst (New Series) 10(2):227-34. [in Chinese with English summary].
Jost L. 2006. Entropy and diversity. Oikos 113(2):363-75.
Kindt R, Coe R. 2005. Tree diversity analysis:
a manual and software for common statisti- cal methods for ecological and biodiversity studies. Nairobi: World Agroforestry Centre (ICRAF). 196 p.
Kuo PC. 1991. Dying trees of the over-mature Chamaecyparis natural stands with regard to its reproduction potentials. Q J Chin For 24(3):35-44. [in Chinese with English sum- mary].
Kuo PC (ed.). 1995. The five most valuable conifer timber species in Taiwan. Taipei, Tai- wan: The Chinese Forestry Association. 510 p.
[in Chinese].
Lee LL. 2000. Survey on wildlife resources in the Chilan cypress forest. Hualien, Taiwan:
Taroko National Park Headquarters, Construc- tion and Planning Agency, Ministry of the In- terior. 64 p. [in Chinese].
Legendre P, Legendre LF. 2012. Numerical ecology. 3rd ed. Amsterdam, The Netherlands:
Elsevier. 1006 p.
Li CF, Zelený D, Chytrý M, Chen MY, Chen
TY, Chiou CR, et al. 2015. Chamaecyparis montane cloud forest in Taiwan: ecology and vegetation classification. Ecol Res 30(5):771-91.
Li K. 1988. The weeping forest. Taipei, Tai- wan: The Commercial Press. 465 p. [in Chi- nese].
Liao CC, Chou CH, Wu JT. 2003a. Popula- tion structure and substrates of Taiwan yellow false cypress (Chamaecyparis obtusa var.
formosana) in Yuanyang Lake Nature Reserve and nearby Szumakuszu, Taiwan. Taiwania 48(1):6-21.
Liao CC, Chou CH, Wu JT. 2003b. Regen- eration patterns of yellow cypress on down logs in mixed coniferous-broadleaf forest of Yuanyang Lake Nature Preserve, Taiwan. Bot Bull Acad Sin 44(3):229-38.
Lin ST, Chen TY, Wei JT. 2016. Chamaecy- paris forests in Chilan Mountain: a potential World Heritage Site. Ilan, Taiwan: Forest Con- servation and Management Administration, Veterans Affairs Council. 287 p. [in Chinese].
Liu TY, Lin KC, Vadeboncoeur MA, Chen MZ, Huang MY, Lin TC. 2015. Understorey plant community and light availability in coni- fer plantations and natural hardwood forests in Taiwan. Appl Veget Sci 18(4):591-602.
Liu VT. 1975. Ecological study on Chamaecy- paris forests in Taiwan. J Agric Assoc China (New Series) 92:143-78.
Lo-Cho CN, Chiu CM, Chen YC. 1999.
Effects of cleaning and pruning on natural- regenerated cypress stands. Taiwan J For Sci 14(3):315-21. [in Chinese with English sum- mary].
Lo-Cho CN, Chung HH, Chiu CM, Chou CF, Lo SS. 1989. Natural regeneration of cypress old growth by selective cutting and intensive site preparation. Bull Taiwan For Res Inst (New Series) 4(4):197-217. [in Chinese with English summary].
Magurran AE. 2004. Measuring biological
diversity. Malden, MA: Blackwell Science.
256 p.
McCune B, Grace JB. 2002. Analysis of eco- logical communities. Gleneden Beach, OR:
MjM Software Design. 300 p.
Ministry of the Interior. 2002. Reports on the delimitation of the Maqaw Cypress Forest National Park. Taipei, Taiwan: Ministry of the Interior, Executive Yuan. 46 p. [in Chinese].
Nagaike T, Kamitani T, Nakashizuka T.
2005. Effects of different forest management systems on plant species diversity in a Fagus crenata forested landscape of central Japan.
Can J For Res 35(12):2832-40.
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, et al. 2017.
vegan: Community ecology package. R pack- age, vers. 2.4-3. Available at https://CRAN.R- project.org/package=vegan.
R Core Team. 2017. R: a language and environ- ment for statistical computing, vers. 3.4.1. Vien- na, Austria: R Foundation for Statistical Comput- ing. Available at http://www.r-project.org.
Teixeira D, Carrilho M, Mexia T, Köbel M, Santos MJ, Santos-Reis M, et al. 2017.
Management of Eucalyptus plantations influ-
ences small mammal density: evidence from Southern Europe. For Ecol Manage 385(Suppl C):25-34.
Wang JC. 2000. Survey on plant resources in the Chilan cypress forest. Hualien, Taiwan:
Taroko National Park Headquarters, Construc- tion and Planning Agency, Ministry of the In- terior. 114 p. [in Chinese].
Wei JT, Chen TY. 2007. Study on vegetation classification in Chi-Lan Shan. J Exp For Natl Taiwan Univ 21(2):133-45. [in Chinese with English summary].
Wei JT, Chen TY. 2012. Different distur- bance intensity situation of plant diversity of Chamaecyparis forest in Chilanshan, northeast of Taiwan. Ilan Univ J Bioresources 8(2):1-11.
[in Chinese with English summary].
Yamamoto S-I. 1993. Seedling establishment of Chamaecyparis obtusa in different micro- environments in the Akasawa Forest Reserve, central Japan. J Jpn For Soc 75(6):519-27.
Yuan HW, Ding TS, Tsai JS. 2004. Effects of removing Chamaecyparis sp. snags and logs on bird communities in Chilan Mt. Q J Chin For 37(1):29-36. [in Chinese with English summary].
Appendix 1. Synopsis of the species composition of ground vegetation and natural seedlings among different Chamaecyparis forest stand types. Dominant and common species are defined as species with ≥ 10 dominance, and of < 10 but ≥ 3 dominance, respectively (counted by the total coverage in ground vegetation and by the total amount of seedlings that naturally regenerated). Note that the number of natural seedlings was so small in the PCC and NSC stands (only 23 and 14 seedlings were found in 10 plots, respectively) that no dominant or common species was designated (here denoted as “N/A”). Likewise, there was no dominant species but 1 common species (Chamaecyparis obtusa var. formosana; 24 seedlings in total) designated in the RO stand.
Stand type Ground vegetation Natural seedlings
Dominant species Common species Dominant species Common species
Plantation after clear cutting (PCC)
Yushania niitakayamensis (玉山箭竹) Eurya crenatifolia (假柃木) Acrophorus stipellatus (魚鱗蕨) Plagiogyria formosana (台灣瘤足蕨)
Ctenitis kawakamii (川上氏肋毛蕨) Dryopteris sparsa (長葉鱗毛蕨) P. euphlebia (華中瘤足蕨) Diplazium kawakamii (川上氏雙蓋蕨) Monachosorum henryi (稀子蕨) I. anisatum (白花八角) B. barthei (深山野牡丹)
N/A N/A
Naturally regenerated stand after selective cutting (NSC)
P. formosana (台灣瘤足蕨) A. stipellatus (魚鱗蕨) E. crenatifolia (假柃木)
N/A N/A
Old-growth forest after removing snags and logs (RO)
P. formosana (台灣瘤足蕨) Illicium anisatum (白花八角)
Polygonum chinense (火炭母草) Smilax china (
拔契)Callicarpa randaiensis (巒大紫珠) C. kawakamii (川上氏肋毛蕨) E. crenatifolia (假柃木)
N/A
C. obtusa
var. formosana (台灣扁柏)Natural old-growth forest (NO)