行政院國家科學委員會專題研究計畫 期中進度報告
登革病毒感染產生自體抗體對內皮細胞的影響及疫苗研發
的考量(2/3)
計畫類別: 個別型計畫 計畫編號: NSC92-3112-B-006-003- 執行期間: 92 年 05 月 01 日至 93 年 04 月 30 日 執行單位: 國立成功大學微生物免疫學研究所 計畫主持人: 林以行 計畫參與人員: 劉校生,莊偉哲,林秋烽,王欣怡,鄭獻仁,萬書文 報告類型: 完整報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 93 年 3 月 26 日
Specific Aims
The onset of vascular leakage and hemorrhagic diathesis is one of the life-threatening complications that occur in dengue patients, yet the pathogenic mechanisms are not well understood. We have proposed a mechanism of molecular mimicry in which antibodies directed against dengue virus (DV) nonstructural protein 1 (NS1) can cross-react with endothelial cells and induce these cells to undergo apoptosis. Immune responses to infectious agents are the critical defense mechanisms in a host, yet abnormal immunity may also be involved in the pathogenesis of disease. We hypothesize that the host-virus interactions that induce autoimmune responses may have implications in dengue hemorrhagic fever and dengue shock syndrome (DHF/DSS) pathogenesis. Gene expression profiling after anti-DV NS1 stimulation in endothelial cells will be studied which may have implications in dengue disease pathogenesis. Furthermore, the epitopes in NS1 that may elicit autoimmune responses should be identified and avoided for safety concern in vaccine development.
Specific Aims
1. The mechanisms involved in the effects of anti-NS1 on endothelial cells after binding will be examined. Using microarray technique, gene expression profiling after anti-DV NS1 stimulation will be dissected. The expression of proteins will be confirmed by immunohistochemistry. The expression of cytokines, chemokines, adhesion molecules, and the molecules involved in apoptotic signaling will be investigated.
2. Endothelial cell surface molecules recognized by anti-DV NS1 will be screened using cDNA library from endothelial cells. For alternative approaches, endothelial cell membrane extracts will be applied on one-dimensional or two-dimensional gel electrophoresis followed by Western blotting and mass spectrometry. The epitopes shared between NS1 and specific surface molecules will be identified. The recombinant NS1 with modifications to remove epitopes that may elicit the pathogenic effects will be prepared and analyzed for: (a) their immunogenicity, (b) protective efficacy in the animal model, and (c) lack of pathogenic effects both in vitro and in vivo.
3. Based on the similarity of partial peptide sequences existed between mucin 5AC, 5B and dengue NS1 protein, specific surface molecules of endothelial cells for
anti-NS1 antibodies recognition are also suspected to be mucin-like proteins. The anti-mucin antibodies will be used to test their binding and apoptosis-inducing effect on endothelial cells, the results will be compared to the properties of anti-NS1 antibodies.
4. We propose to develop DNA vaccine by construction of modified NS1 genes in a mammalian expression system and testing the protective efficacy and potential pathogenic effects based on the knowledge obtained from Specific Aims 1, 2 and 3.
Progress Summary
Vascular leakage, liver abnormality, and hemorrhagic diathesis are the clinical features associated with DHF/DSS, yet the mechanisms remain unclear. We have proposed a mechanism of molecular mimicry in which antibodies directed against DV NS1 cross-react with endothelial cells and cause damage. The presence of autoantibodies could be detected in both dengue patient sera and NS1-hyperimmunized mouse sera (Lin et al., 2002, 2003; Hung et al., submitted). Endothelial cells underwent apoptosis via production of NO following anti-NS1 induction (Lin et al., 2002, 2004). Meanwhile, immune activation including cytokine and chemokine production and adhesion molecule expression after treatment with anti-NS1 in endothelial cells was demonstrated (Lin et al., manuscript in preparation), which can be correlated with the cytokine profiles in patients with DHF/DSS (Hung et al., 2004). Studies directed toward Specific Aim 1 concerning the mechanisms involved in the effects of anti-NS1 antibodies on endothelial cells include signal transduction, cytokine and chemokine production, adhesion molecule expression, and interaction between mononuclear cells and endothelial cells. Protein phosphorylation and NF-κB activation were observed after anti-NS1 stimulation. The production of IL-6, IL-8 and MCP-1, but not RANTES, showed an increase in endothelial cells. The elevation of ICAM-1 expression and the adhesive ability of peripheral blood mononuclear cells to endothelial cells were also observed, and these effects were inhibited by pretreatment with anti-MCP-1 or anti-ICAM-1 antibodies. This part of studies is being prepared as a manuscript for submission. The establishment of animal models by either passive administration of anti-NS1 antibodies or active immunization of NS1 has also been done and the manuscript is currently prepared. The study of gene expression patterns of anti-NS1-stimulated endothelial cells is ongoing. After stimulation with anti-NS1, anti-JEV or control mouse IgG, the total RNA of treated and untreated endothelial cells are extracted for hybridization analysis.
The human microarray of 9,600 spots is used for screening. The RNA samples have been sent to Microarray Core Facility for Genomic Medicine in National Taiwan University Hospital, and the analyzed microarray data are being expected.
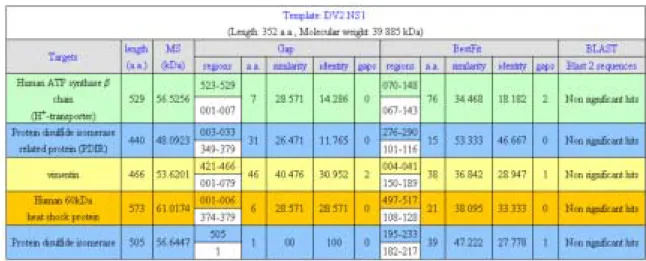
For studies toward Specific Aim 2, some endothelial cell surface molecules recognized by anti-DV NS1 have been identified by two-dimensional gel electrophoresis followed by Western blotting and mass spectrometry. Membrane extracts from endothelial cell line, HMEC-1, were applied on two-dimensional gel electrophoresis followed by Western blotting. By comparing the results of anti-NS1, anti-JEV, and control mouse IgG hybridized membranes (Fig. 1A) with silver-stained gels, fifteen major protein spots were selected and collected (Fig. 1B). The molecular weights of these proteins are around 50 to 60 kDa with isoelectric points ranged from pH 4.5 to 6. These proteins were recollected from silver-stained gels by trypsin digestion and then analyzed by Malti-Tof mass spectrometry and peptide sequencing. After alignment to database of NCBI and SwissPort, these proteins are identified as beta chain of H+-transporter/ATP synthase (spots 1 and 2), protein disulfide isomerase related protein (PDIR) (spots 3 and 4), vimentin intermediate filament (spots 5 and 6), heat shock protein 60 (spots 7 to 12), and protein disulfide isomerase (PDI) (spots 13 to 15), respectively (Table 1). Further peptide sequence alignment of these proteins and dengue NS1 protein by using Bestfit and Gap of GCG Seqweb and Blast2 of NCBI show that none of significant sequential similarity is shared between DV NS1 and these endothelial cell membrane proteins (Table 2). One of the possibilities is that the cross-reaction of anti-DV NS1 antibodies to endothelial cell membrane proteins is caused by epitopes with 3D structural mimicry, but more detail bioinformatic alignment is needed for confirmation.
Studies for Specific Aim 3 have demonstrated that a potential epitope of mucin-like domain shows molecular mimicry with DV NS1. Based on the results obtained from protein database (Fig. 2), mucin 5AC (C-terminal, a.a. 812-831) and mucin 5B (C-terminal, a.a. 902-921) show homology with the C-terminal of DV NS1 (a.a. 312-331) and are expressed on the surface of endothelial cells (HMEC-1) and epithelial cells (A549) (Fig. 3). Using ELISA technique, anti-mucin 5AC/5B cross-reactive with DV NS1 and C-terminal peptides (C20, a.a. 312-331) was shown (Fig. 4). The endothelial binding activity of mouse anti-NS1 antibodies was inhibited by preabsorption with C20 but not N15 (a.a. 1-15) peptides (Fig. 5). Furthermore, the competitive binding assay showed that the binding activities of anti-DV NS1 Abs and anti-mucin 5AC/5B Abs to HMEC-1 and A549 cells were partially inhibited by the preabsorption with anti-mucin 5AC/5B and anti-DV NS1 Abs, respectively (Fig. 6). Therefore, mucin-like epitopes can be recognized by anti-DV NS1. The potential pathological significance requires further studies.
Specific Aim 4 will be conducted in next year project based on the results obtained from Specific Aims 1-3.
Publications and manuscripts:
1. Lin, C. F., H. Y. Lei, A. L. Shiau, H. S. Liu, T. M. Yeh, S. H. Chen, C. C. Liu, S. C. Chiu, and Y. S. Lin. 2002. Endothelial cell apoptosis induced by antibodies against dengue virus nonstructural protein 1 via production of nitric oxide. J. Immunol. 169:657-664.
2. Lin, C. F., H. Y. Lei, A. L. Shiau, C. C. Liu, H. S. Liu, T. M. Yeh, S. H. Chen, and Y. S. Lin. 2003. Antibodies from dengue patient sera cross-react with endothelial cells and induce damage. J. Med. Virol. 69:82-90.
3. Lin, Y. S., C. F. Lin, H. Y. Lei, H. S. Liu, T. M. Yeh, S. H. Chen, and C. C. Liu. 2004. Antibody-mediated endothelial cell damage via nitric oxide. Curr. Pharm. Design 10:213-221.
4. Hung, N. T., H. Y. Lei, N. T. Lan, Y. S. Lin, K. J. Huang, L. B. Lien, C. F. Lin, T. M. Yeh, D. Q. Ha, V. T. Q. Huong, L. C. Chen, J. H. Huang, L. T. My, C. C. Liu, and S. B. Halstead. 2004. Dengue hemorrhagic fever in infants: a study on clinical and cytokine profiles. J. Infect. Dis. 189:221-232.
5. Hung, N. T., N. T. Lan, Y. S. Lin, C. F. Lin, L. B. Lien, K. J. Huang, T. M. Yeh, D. Q. Ha, V. T. Q. Huong, L. C. Chen, L. T. My, J. H. Huang, C. C. Liu, and H. Y. Lei. Anti-platelet and anti-endothelial cell autoantibodies in Vietnamese infants and children with dengue hemorrhagic fever. (submitted)
6. Lin, C. F., S. C. Chiu, Y. L. Hsiao, H. Y. Lei, A. L. Shiau, H. S. Liu, T. M. Yeh, S. H. Chen, C. C., Liu, and Y. S., Lin. Expression of cytokine, chemokine, and adhesion molecule during endothelial cell activation induced by antibodies against dengue virus nonstructural protein 1. (submitted)
7. Lin, C. F., C. C. Cheng, S. C. Chiu, Y. L. Hsiao, H. Y. Lei, A. L. Shiau, H. S. Liu, T. M. Yeh, S. H. Chen, C. C. Liu, and Y. S. Lin. Pathologic effects of antibodies against dengue virus nonstructural protein 1 in mice. (manuscript in preparation)
(A) (B)
Figure 1. Hybridization of endothelial membrane extracts of HMEC-1 with anti-DV NS1, anti-JEV NS1 and control mouse IgG (A). Fifteen major protein spots specifically hybridized with anti-DV NS1 are selected from 2D gels (B).
Figure 2. Alignment of the homologous amino acid sequences between DV NS1 and human mucin 5AC/5B by Pretty of GCG Seqweb.
Figure 3. The expression of mucin 5AC and mucin 5B on HMEC-1 and A549 cells is determined by flow cytometry (A) and confocal microscopy (B).
Figure 4. The cross-reactivity of anti-mucin 5AC/5B with DV NS1 (A) and C-terminal peptide (C20, a.a. 312-331) (B) is analyzed by ELISA. *: p<0.05, **: p<0.01, ***: p<0.001
Figure 5. Endothelail binding activity of mouse anti-NS1 Abs can be inhibited by preabsorption with NS1 C20 peptides. The percentages of HMEC-1 cells that reacted with non- or preabsorption Abs are
shown as mean ± SD.
Figure 6. Competitive binding of anti-DV NS1 with anti-mucin 5AC/5B on the surface of HMEC-1 and A549 as detected by flow cytometric analysis.
Table 1. Fifteen major proteins hybridized with anti-DV NS1were identified by Malti-Tof MS
Table 2. Alignment results between DV NS1 and proteins specifically hybridized with anti-NS1
BestFit - Makes an optimal alignment of the best segment of similarity between two sequences. Optimal alignments are found by inserting gaps to maximize the number of matches using the local homology algorithm of Smith and Waterman.
Gap - Uses the algorithm of Needleman and Wunsch to find the alignment of two complete sequences. It maximizes the number matches and minimizes the number of gaps.
BLAST 2 SEQUENCES - This tool produces the alignment of two given sequences using BLAST engine for local alignment.