嘉南藥理科技大學 生物科技系
碩士論文
Tomatidine 抑制人類肺癌細胞 A549 基質金屬蛋白酶的表現 Tomatidine inhibits the expression of matrix
metalloproteinases in human lung carcinoma A549
指導老師:陳 品 晟 博士 研 究 生:李 佳 真
中華民國一百年七月十四日
嘉南藥理科技大學生物科技系 Department of biotechnology
Chia-Nan University of Pharmacy and Science
碩士論文
Thesis for the Degree of Master
Tomatidine 抑制人類肺癌細胞 A549 基質金屬蛋白酶的表現 Tomatidine inhibits the expression of matrix metalloproteinases in
human lung carcinoma A549
指導老師:陳 品 晟 博士(Dr. Pin-Shern Chen)
研 究 生:李 佳 真(Chia-Chen Li)
中華民國ㄧ百年七月十四日
July 14, 2011
謝誌
我真的很珍惜嘉南藥理科技大學給予這兩年寶貴的學習機會,回首求 學的過程,學習的初始雖然辛苦,除了面臨考試與專題討論報告時備感壓 力之外,生物科技系的老師授業解惑及專業指導下,受益良多,也使學生 這兩年的日子更為充實、精彩。
我親愛的老師—品晟老師,在我碩士班生涯中,給予我很多的幫助,
從題目的擬定建議到研究動機乃至於論文架構、研究方法與觀念上提供諸 多寶貴意見,讓學生從煩亂的思緒中理清正確的方向,使學生獲益匪淺。
這篇論文得以付梓,特別要感謝指導教授陳品晟老師鉅細糜遺的細心 指導,感謝林美惠教授和蔡孟峯副教授對論文的撰寫方向及資料的蒐整方 面提出寶貴的意見及指正,實驗室的祥青、恩澤、阿傑、小力、幸文、雅 珊、佩青大家一起互相切磋,還有共同為生活而努力的新樓醫院檢驗科的 同室晨愉、郁晴、曉菁以豐富的經驗不吝嗇的給予寶貴的意見,時時關切 論文的進度,使我在文思枯竭時,注入活水,銘感五內。
感謝碩士班一起打拼論文的同學們,恭喜你們也拿到了碩士論文學 位。另外,還在持續努力的同學們,你們也要加油,再加把勁將論文完成 喔!尤其,要特別感謝我的好同學,一同為論文努力的日子,相互鼓勵而建 立起的革命情感,不但讓我們成為莫逆之交,更讓我們這輩子都永久懷念。
最後我要謝謝我的家人祥釗、聖紘、崇祐對於我在多重角色的扮演上,
常常無法面面俱到、力不從心的體諒與支持,以感恩的心於現在的職責中,
我還可利用時間增進知識與學習,真的心存感激。
最後,我想將這一份榮耀獻給影響我最深的外婆,衷心感謝她的教誨,
時常告誡「人要不斷的學習」,因為有您,終於在碩士學位這重要的人生階
段,劃上美好的句點。加油! 加油! 加油!共勉之~
李佳真 謹致 中華民國 100 年 7 月
中文摘要
肺癌是男性和女性的癌症死亡原因中最常見的一種,其可分為小細胞 肺癌和非小細胞肺癌。肺癌的形成和侵襲是一個多步驟的過程,其中涉及 癌細胞與鄰近周圍基質 間的多種蛋白 交互作用,包括基質金屬 蛋白酶
(matrix metalloproteinase, MMP)和組織金屬蛋白酶抑製劑(tissue inhibitor of metalloproteinase, TIMPs)。基質金屬蛋白酶是一種鋅依賴性內肽酶家 族,具有切割細胞外基質的能力。MMP 家族中的明膠基質金屬蛋白酶,特 別是 MMP-2 和 MMP-9,會參與一些病理過程,例如腫瘤細胞轉移和血管 新生。許多的侵入性惡性腫瘤,包括肺癌,皆表現大量的 MMP 相關酵素,
例如 MMP-2、MMP-7、MMP-9、TIMP-1 等。Tomatidine 是由番茄萃取得 到的天然固醇類生物鹼。最近的研究發現 tomatidine 可以降低發炎作用。
Tomatidine 對癌細胞轉移方面的研究尚不清楚。本實驗室先前的研究發現,
tomatidine 對人類肺癌細胞 A549 的侵入作用有明顯的抑制。本論文擬以 reverse transcription- PCR (RT-PCR) 探討 tomatidine 對 MMP-2/-9/-7 mRNA 表現的影響。結果發現,tomatidine 在 40 μM 以下的劑量並不會影響 A549 細胞的增生。在此無細胞毒性的劑量下,tomatidine 可抑制 A549 細胞的侵 入作用。Tomatidine 可抑制 MMP-2/-9/-7 mRNA 基因的表現。同時 tomatidine 也抑制組織金屬蛋白酶抑製劑 TIMP-1 的表現。此結果顯示 tomatidine 可能 藉由抑制 MMP-2/-9/-7 基因的表現而抑制人類肺癌細胞 A549 的侵入作用。
關鍵字:基質金屬蛋白酶蛋白酶、組織金屬蛋白酶抑製劑、Tomatidine。
Abstract
Lung cancer is the most common cause of cancer-related death in men and women and can be divided into small cell lung carcinoma and non-small cell lung carcinoma. Progression and invasion of lung cancer is a multi-step process involving multiple interactions between the tumor and the surrounding stroma mediated by many proteins, such as metalloproteinase (MMP) and tissular inhibitors of metalloproteinase (TIMP). MMP comprise a family of zinc-dependent endopeptidases that have the capacity to cleave extracellular matrix. The subgroup of MMPs known as gelatinases, specifically gelatinase A (MMP-2) and gelatinase B (MMP-9), have been implicated in the pathological processes that contribute to tumor progression, such as tumor cell invasion and angiogenesis. Many invasive malignant tumors, such as lung cancers, express high levels of MMPs, including MMP-2, MMP-7, MMP-9, and TIMP-1.
Tomatidine, isolated from tomato, is a naturally occurring steroidal glycoalkaloid. Recent study demonstrated that tomatidine suppressed inflammation. However, the effect of tomatidine on metastasis of caner cell is still unclear. In our previous study, we demonstrated that tomatidine suppressed invasion of human lung cancer cell A549. In the present study, we aimed to elucidate the effect of tomatidine on the expressions of MMP-2, -9 and -7 by
RT-PCR. Results show that treated of tomatidine at concentrations below 40 μM did not affected viability of A549 cells significantly. When treated with non-toxic doses of tomatidine, cell invasion was markedly suppressed.
Tomatidine also reduced the expression of MMP-2, -9 and -7. Meanwhile, tomatidine suppressed the expression of TIMP-1. Taken together, these results suggest that tomatidine suppresses the expression of MMP-2 -9 and -7 and may subsequently contribute to the inhibition of invasion of A549 cells.
Key word : Matrix metalloproteinases (MMPs), tissular inhibitors of metalloproteinase (TIMP), Tomatidine.
目錄(Contents)
中文摘要 ...I Abstract... III 目錄(Contents)... V 圖目錄 ... VIII 附錄圖表 ... IX 縮寫表 ... X
第一章 緒論 ... 1
1.1 肺癌(Lung cancer)... 1
1.2 肺癌細胞 A549 ... 2
1.3 癌細胞的轉移(Cancer of metastasis) ... 2
1.4 基質金屬蛋白酶(Matrix metalloproteinases) ... 3
1.5 Tomatidine 簡介 ... 6
1.6 實驗目的... 7
第二章 材料與方法 ... 9
2.1 試劑及相關實驗耗材: ... 9
2.2 檢體採集:細胞培養(Cell culture)... 12
2.3 Tomatidine 處理 ... 14
2.4 細胞存活率實驗(MTT assay): ... 15
2.5 細胞侵入性實驗 (Matrigel invasion assay)... 15
2.6 RNA 抽取流程(Total RNA extraction)... 16
2.7 核醣核酸定量(RNA quantitative) ... 18
2.8 反轉錄聚合酶連鎖反應(Reverse transcriptase polymerase chain reaction ,RT-PCR) ... 18
2.9 去氧核醣核酸定量(cDNA quantitative) ... 19
2.10 聚合酶連鎖反應(Polymerase chain reaction,PCR) ... 20
2.11 統計分析 (Stastistical analysis) ... 22
第三章 結果 ... 23
3.1 Tomatidine 對 A549 細胞存活率的影響 ... 23
3.2 Tomatidine 對 A549 細胞侵入能力的影響 ... 23
3.3 Tomatidine 抑制 A549 細胞 MMP-2 基因的表現 ... 23
3.4 Tomatidine 抑制 A549 細胞 MMP-9 基因的表現 ... 24
3.5 Tomatidine 抑制 A549 細胞 MMP-7 基因的表現 ... 24
3.6 Tomatidine 對於 A549 細胞 TIMP-1 基因的表現... 25
3.7 Tomatidine 對於 A549 細胞 TIMP-2 基因的表現... 25
第四章 討論 ... 26
第五章 結論 ... 29
第六章 參考文獻 ... 30
第七章 圖表 ... 39 附錄圖表 ... 52
圖目錄
圖 1. Tomatidine 對 A549 細胞存活率的影響... 39
圖 2. Tomatidine 對 A549 細胞侵入能力的影響... 41
圖 3. Tomatidine 抑制 A549 細胞 MMP-2 的表現 ... 42
圖 4. Tomatidine 抑制 A549 細胞 MMP-9 的表現 ... 44
圖 5. Tomatidine 抑制 A549 細胞 MMP-7 的表現 ... 46
圖 6. Tomatidine 對於 A549 細胞 TIMP-1 的表現 ... 48
圖 7. Tomatidine 對於 A549 細胞 TIMP-2 的表現 ... 50
附錄圖表
附錄 1. MMP 在腫瘤細胞移動中所扮演的角色。... 52
附錄 2.腫瘤轉移的過程。... 53
附錄 3. Tomatidine 的化學結構式。 ... 54
附錄 4.Tomatidine 抗發炎的可能機制。 ... 55
縮寫表
BSA:Bovine serum albumin ddH2O:Double distilled water DEPC:Diethylpyrocarbonate
DMEM:Dulbecco's modified eagle's medium DMSO:Dimethyl sulfoxide
ECM:Extracellular matrix
ERK:Extracellular sinaling-regulated kinase FBS:Fetal bovine serum
JNK/SAPK:c-Jun N-terminal kinase/stress-activated protein kinase kDa:Kilo-dalton
MAPK:Mitogen-activated protein kinase MMPs:Matrix metalloproteinases
β-ME:2-Mercaptoethanol NF-κB:Nuclear factor kappa B OD:Optical density
P/S:Penicillin / Streptomycin PBS:Phosphate buffered saline PCR:Polymerase chain reaction
PI3-K:Phosphoinositide 3-kinase
RT-PCR:Reverse transcriptase polymerase chain reaction TIMPs:Tissue inhibitor of metalloproteinases
第一章 緒論 1.1 肺癌(Lung cancer)
台灣人罹患癌症的比例中,肺癌是最常見且最重要的一種癌症,佔男 性癌症死亡率的第二位(僅次於肝癌)、女性的第一位,是男女合計中癌症死 亡率的第一位(1)。Lee, G. 等學者的研究表明,肺癌病例是由吸煙,空氣污 染,環境風險因素(即風險輻射,石棉,重金屬和多環芳烴碳氫化合物)
和致癌基因所引起(2)。肺部腫瘤細胞可能發展出強勁的侵襲和轉移特性,而
成為惡性腫瘤。在臨床上,患者身上常出現肺癌腫瘤細胞的浸潤和轉移,
目前的治療方法包括手術,放射線治療,化療等對此症狀的治療效果皆不 佳。通常 85%的肺癌患者診斷存活期約 5 年(3)。因此,運用分子生物技術 研究肺癌,設計新的治療藥物,以防止或減緩肺癌細胞侵襲和轉移,乃成 為重要課題。
臨床上將肺癌簡單區分為非小細胞肺癌 ( NSCLC ) 和小細胞肺癌 ( SCLC ) 。非小細胞肺癌和小細胞肺癌為不同形式的肺癌、具有不同的遺 傳原因,治療方法也不同(4)。非小細胞肺癌可分為:(1)鱗狀細胞癌,發生 率約30~50%,病灶發生於肺門附近,吸煙為最主要的發生因素。(2)腺癌,
發生率為25%,病灶發生於肺部周邊,好發於女性、非吸煙者、肺部瘢痕 組織病變者。(3)大細胞癌,發生率約10%,可發生在任何部位。小細胞癌,
發生率為20~25%,好發於肺門附近,與吸菸有關(5)。
1.2 肺癌細胞 A549
A549 肺癌細胞源自 58 歲白人的肺葉細胞癌。在 1972 年由 Giard DJ 等學者建立細胞系,使用電子顯微鏡觀察 A549 細胞,在早期和後期有多
層細胞質包涵體,屬於典型的第二型肺泡上皮細胞(6),就是臨床上所謂的
非小細胞肺癌。現在市售的肺癌細胞株種類有很多,例如: MRC -5 是人 類胚胎肺成纖維細胞,被用來作為正常對照; H226 和 H520 是鱗狀細胞 癌的細胞株;H358 和 A549 是腺癌細胞系(7)。A549 細胞常用於肺癌的研 究,例如 Jing-Hsien Chen 等學者利用小鼠為實驗品,結果發現 Gaseous Nitrogen Oxide (GNO)可促進肺癌細胞 A549 轉移到肺(8)。
1.3 癌細胞的轉移(Cancer of metastasis)
一 般 癌 細 胞 的 轉 移 過 程 包 括 : (1) 細 胞 移 動 ( motility ) 和 侵 入 ( invation )能力的增加。細胞爬行的部分細胞會先向前伸出偽足後,而細 胞表面的受體會與細胞外基質黏合,尾端的細胞表面受體會被蛋白水解酶 水解,使細胞能向前移動 (附錄 1);(2)細胞與細胞間相互作用力被破壞;
(3)細胞與細胞外基質間的結合能力 ( adhesion ) 增加;(4)胞外基質被分 解。惡性腫瘤發生轉移的首要步驟即是侵入細胞外基質。惡性腫瘤轉移時 會產生大量的分解酵素破壞細胞外基質,包括 serine proteinase 、 Matrix
metalloproteinases 、 cathepsins 、 plasminogen activator 及 heparanitase 等(9,10,11)。
1.4 基質金屬蛋白酶(Matrix metalloproteinases)
蛋白水解酶 ( proteases ) 影響範圍廣泛,例如細胞外蛋白質水解酶是腫 瘤細胞穿過組織所須的物質;水解 ECM 蛋白會造成腫瘤的轉移,而內皮 細胞水解 ECM 導致的血管新生,並會促進腫瘤生長。有四種內胜肽酶 ( endopeptidases ) :絲氨酸 ( serine ) 蛋白酶、胱氨酸 ( cystine ) 蛋白酶、
天門冬氨酸 ( aspartyl ) 蛋白酶,基質金屬蛋白酶等,都和腫瘤的發生、轉 移過程有關(12)。
基質金屬蛋白酶 ( Matrix metalloproteinases ,簡稱 MMPs ) 是指一家 族的鋅依賴型蛋白水解酶,其主要的工作就是水解蛋白質(13)。MMPs 可分 解細胞外基質 ( extracellular matrix, ECM ) ,不管在正常生理或病理狀態,
他們都扮演重要的角色。MMP 家族成員的蛋白結構不盡相同,但至少都含 有信號肽、前肽和催化區 3 個結構域,酶催化區和前肽區具有高度保守性。
目前已經發現的 MMP,依蛋白的功能區和可分解的基質種類不同(14,15,16), 可分為六大類:
(1).膠原酶 ( MMP-1, -8, -13 )
(2).明膠酶 ( MMP-2 和 MMP-9 )
(3).Stromelysins 蛋白酶 ( MMP-3,-10, -11 )
(4).基質溶解蛋白酶 ( MMP-7 和 MMP-26 )
(5).膜型—基質金屬蛋白酶蛋白酶 ( MMP-14, -15, -16, -17, -24 和 -25 )
(6).其他蛋白酶 ( MMP-12, -19, -20, -21, -23, -27 和 -28 )
此外,基質金屬蛋白酶還可以調節大量非細胞外基質蛋白的活性,例 如生長因子,細胞因子,趨化因子,細胞受體,絲氨酸蛋白酶抑製劑和蛋
白酶等(17,18)。因此,基質金屬蛋白酶的複雜生物功能具有影響正常生理和病
理的特性,包括能造成形態發生改變、傷口癒合、組織修復、傷口重塑、
以及心肌梗塞動脈粥樣硬化、關節炎、癌症和慢性潰瘍組織(19,20,21)等。
細胞在癌化過後基質金屬蛋白酶的表現量會大幅的提升,惡性腫瘤發 生轉移的部位亦可測得大量的 MMPs 。在相關研究中證實癌細胞藉由 MMPs 來分解細胞外基質及破壞基底膜(22),再藉由血管及淋巴系統將癌細 胞轉移到其他器官(23)(附錄 2)。在 MMP 家族中, MMP-9 和 MMP-2 同 時都具有分解細胞外基質及破壞基底膜的能力,與癌細胞的轉移與侵入有 密切的相關性(24)。 Stetler-Stevenson W.G.等學者研究發現在腫瘤細胞中(明 膠酶 A / MMP-2,基質分解素 1 / MMP-3, matrilysin / MMP-7 ,明膠酶 B / MMP-9, stromelysin-2 / MMP-10 ,和 MT1-MMP / MMP-14),或晚期腫 瘤基因(stromely-sin-3 / MMP-11 , collagenase-3 / MMP-13)皆有表現(25)。
MMPs 許多種腫瘤都有 MMP 表現,例如食道癌有 MMP-1 / -2 / -3 / -9 表 現,胃癌 MMP-1 / -2 / -7 / -9 及 MT1-MMP ,大腸癌有 MMP-1 / -2 / -3 / -9 / -11 表現,胰臟癌有 MMP-1 / -2 / -3 及 TIMP-1 ,乳癌有 MMP-2 / -9 / -11 , 前 列 腺 癌 有 MMP-2 , 肺 癌 有 MMP-2 / -11 、 MT1-MMP 及 TIMP-1,卵巢癌有 MMP-2 / -9 及 MT1-MMP (26)。
基質金屬蛋白過度表現導致細胞外基質瓦解時,身體內會產生組織金屬蛋 白酶抑製劑 ( Tissue inhibitor of metalloproteinases , TIMPs ),其主要目的 是預防 MMPs 的過量活化。 TIMP 是由包括皮膚在內的各種器官的細胞 製造出來的。 TIMP 與 MMP 以 1:1 的比例的方式連結成一個可逆的、
非共價鍵結合的複合物,可抑制 MMPs 的活性。目前所知的 TIMPs 有四 大族群: TIMP1 ~ TIMP4 ,均屬於富含胱胺酸 ( cysteine ) 的蛋白質,透 過雙硫鍵來穩定抑制的作用。 TIMPs 的主要功能是保持組織蛋白水解的平 衡,例如在中樞神經系統,小膠質細胞和星形膠質細胞利用 TIMPs 調解組 織的分解和修復(27)。 TIMP-1 表現於纖維母細胞和角膜上皮細胞,也與肝 損傷後修復的基質重塑過程中有關(28)。 TIMP-1 會受到基礎纖維母細胞生 長因子、血小板生長因子、間白素-1等刺激而表現。 TIMP-2 的功能是在 主導細胞外基質代謝(29)。在腫瘤細胞遷移中基質金屬蛋白酶2(MMP-2)能 夠降解第四型膠原蛋白。 TIMP-2 可抑制 MMP-2 活性。 MMPs 與 TIMPs 兩者之間的不平衡可能與癌細胞的生長有關(30)。 TIMP-3 具有抑制血管新
生的作用。例如眼睛發生黃斑退行性病變或黃斑脈絡膜病變時,會有血管 內皮因子表現而刺激血管新生,此時亦會有 TIMP-3 的表現,抑製血管生 成。 Apte S.S. 等學者的研究發現, TIMP3 會抑制血管內皮因子受體-2
(VEGFR-2)和破壞下游信號而抑制血管新生(31)。 TIMP- 4 表現於心臟,
腎臟,胰腺,大腸,睾丸,腦和脂肪組織(32)。 TIMP-4 是一種非糖基化,
由195個氨基酸組成的長多肽(33),是目前所發現最大的人類基質金屬蛋白酶 抑製劑。 Nicholas A.等學者研究發現 , TIMP-4 除了可去除 MMP-1
(34)。,也與抑制血管新生的作用有關。
1.5 Tomatidine 簡介
蕃茄素 ( tomatine ) 與蕃茄素定 ( Tomatidine ) 屬於類固醇醣基生物 鹼,廣泛地存在於茄科植物的根、莖及葉片中,其中以根部的含量為最多。
Tomatine 與 tomatidine 可在腸內和膽固醇形成一不可吸收的複合物,影響 膽 固 醇 吸 收 及 影 響 細 胞 膜 的 通 透 性 , 它 們 亦 可 作 為 疫 苗 之 佐 劑 ( adjuvant ),而增加其作用。 Tomatidine 是氮類似物類固醇,這是一種固
醇類激素和抗發炎類固醇的前驅物(35)。曾有研究使用各種的類固醇生物鹼
去 處 理 不 同 的 癌 細 胞 , 例 如 α - tomatine 、 dehydrotomatine 、 tomatidenol 、 tomatidine ,來檢測這些類固醇生物鹼的毒性,其結果表示 tomatidine 不具有很強的毒殺性(36)。而這些類固醇生物鹼有許多生物活性,
包括抗真菌和抗菌活性。這些類固醇生物鹼的抗發炎作用是非常有限的,
其結構(附錄3)和醣皮質激素類似(37)。
在茄科類植物,如西紅柿,土豆和茄子,具有固醇類生物鹼基於 C27 膽固醇骨架,如 tomatidine 和 solasodine 。這些化合物基本上氮類似物類 固醇如 diosgenin ,它是一個前體的類固醇荷爾蒙的前趨物和具抗發炎的類
固醇(38)。固醇類生物鹼和醣皮質激素生物活性類似,包含抗黴菌及抗細菌
的作用(39)。 Chiu F.L.等學者研究發現 tomatidine 可以抑制 inducible NOS ( iNOS ) 和 cyclooxygenase-2 ( COX-2 ) 而降低發炎的作用(40)。同時發現在 LPS-刺激老鼠的巨噬細胞經由抑制 NF–κB 與 JNK 路徑有關(41) (附錄4)。
過去文獻中使用 tomatidine 分別處理不同的癌細胞,測試結果中顯示 tomatidine 可以抑制 hedghog pathway的相關基因表現(42)。同時也有文獻表 示 , hedghog pathway 會 影 響 MMP-9 的 表 現(43)。 最 近 的 研 究 發 現 tomatidine 可抑制 Sonic Hedghog ( SHH ) 蛋白所誘發的 Gli 轉錄因子活 化(44),同時也抑制 Gli 轉錄因子調控的下游基因 Ptc 1 的表現(45,46)。 Hh 訊息傳遞路徑除了影響胚胎發育,傷口癒合和修復的過程外,也與腫瘤形 成有關。因此 tomatidine 也可能藉由抑制 Hh 訊息路徑而抑制腫瘤形成。
1.6 實驗目的
本論文擬以人類肺腺癌上皮細胞 ( A549 ) 為體外試驗,首先延續實驗
室先前學長的研究成果,分析 tomatidine 的細胞毒性,並了解 tomatidine 對於癌細胞轉移過程(如細胞爬行/侵入)之影響。細胞爬行/侵入的同時也會 分泌 MMPs 來分解細胞外基質而影響 MMPs 的活性,因此再利用反轉錄 聚合酶連鎖反應 ( RT-PCR ) 分析 tomatidine 對 A549 細胞的 MMPs 相 關基因表現。
第二章 材料與方法 2.1 試劑及相關實驗耗材:
(1).實驗耗材:
15cm 離心管、50cm 離心管,15cm dish,blue tip、yellow tip、
micro tip,1.5ml tubes、0.2ml PCR tubes ( Flat Caps )、0.2ml Q-PCR 專 用 tubes ( PCR Strip with attached Optically Clear Cap Low Profile )、96 孔培養盤。
(2).實驗相關試劑:
(A).DNase I : Deoxyribonuclease I 20000U(251U/ μ L ) ; 廠 牌 是 Invitrogen TM ;Cat no.18047-019 ; Lot no.810266. ;Conc 為 50-375U/μl;保存於-20℃。
(B).Oligo dT :Oligo (dT)12-18primer 0.5μg/μL;廠牌是 Invitrogen
TM ;Lot#767206 ;保存於-20℃。
(C).5 × FS Buffer:5 × First Strand Buffer;廠牌是 Invitrogen TM
;Lot no.709042 ;保存於-20℃。
(D).10mM dNTP Mix:10mM dNTP(2’-deoxynucleoside 5’-triphospate) Mix consists of all four nucleotides(dATP,dCTP,dGTP,dTTP),
Size:1mL。廠牌是 Gene Direχ®;Cat No. DN0010.;保存於-20
℃。
(E).0.1M DTT:500μL;廠牌是 InvitrogenTM;Lot no.707966.
;保存於-20℃。
(F).RNase inhibitor:Rnase OUTTM Recombinant Ribonuclease Inhibitor 5000U (40U/μL) ;廠牌:Invitrogen TM;Lot no .832047.;保存於 -20℃。
(G).RTase (M-MLV RT):M-MLV Reverse Transcriptase 40000U (200 U/μL) ;廠牌是 Invitrogen TM;Lot no. 825017.;保存於-20℃。
(H).10X DNase Buffer:RQ1 DNase 10X Reaction Buffer 1mL
;廠牌是 Promega;Lot no. M198A 21542609;保存於-20℃。
(I).2.0 × Master Mix Red:Taq DNA pol 2.0 × Master Mix Red (MgCl2 1.5mM);廠牌是 Ampliqon II;Lot no.5200300-10H17;保 存於-20℃。
(J)10.RT-PCR primer:
(a).MMP-2 primer:
Forward:5’- CAAGGAGTACAACAGCTGCACTGATA -3’
Reverse:5’- GGTGCAGCTCTCATATTTGTTGC -3’
(b).MMP-7 primer:
Forward:5’- GTATGGGGAACTGCTGACATCATG -3’
Reverse:5’- CTGAATGCCTTTAATATCATCCTG -3’
(c).MMP-9 primer:
Forward:5’- TGGGCAAGGGCGTCGTGGTTC -3’
Reverse:5’- TGGTGCAGGCGGAGTAGGATT -3’
(d).TIMP-1 primer:
Forward:5’-TCAACCAGACCACCTTATAC -3’
Reverse:5’- ATTCCTCACAGCCAACAG -3’
(e).TIMP-2 primer:
Forward:5’-GTAGTGATCAGGGCCAAAG -3’
Reverse:5’- TTCTCTGTGACCCAGTCCAT -3’
(f).β-actin primer:
Forward:5’- TGTTACCAACTGGGACGACA -3’
Reverse:5’- GGGGTGTTGAAGGTCTCAAA -3’
(K).DEPC-H2O:Diethyl pyrocarbonate D5758 - ≥97% (NMR) ;廠牌 是 Sigma-Aldrich®;CAS Number: 1609-47-8; 保存於:2-8℃
(L).10X TBE Buffer:Ultra PureTM10X TBE Buffer 1000mL;廠牌是 Invitrogen TM;Lot no. 376964.;保存於:-15-30℃
(M).100bp DNA Ladder RTU 50μg/500μl;廠牌 Gene Direχ®
;Cat No.DM001-R500;Lot no. 606027.; 保存於-20℃
(N).β-ME:2-Mercaptoethanol,for electrophoresis,﹥﹦98﹪
廠牌是 SIGMA®;Cat No.M7154-100ML Bach#025101271。
(O).PBS 配製:
PBS 配製 500 ml 1L NaHPO42H2O 1.442g 2.884g KCl 0.2g 0.4g NaCl 8g 16g KH2PO4 0.2g 0.4g D.D H2O 500 ml 1L
(P).Dulbecco’s Modified Eagle’s Medium-high glucose (DMEM):High Medium(1L) (2×500ml 過濾)
High Glucose
粉末ㄧ瓶 粉末二瓶
NaHCO3 3.7g 7.4g
D.D H2O 1L 2L 使用前再加入過濾器,過濾分裝
FBS
10% 10%
P/S
1% 1%
(Q).Total RNA extraction kit 使用試劑批號 Cat No.YRB50。
2.2 檢體採集:細胞培養(Cell culture)
(1).解凍細胞
先將冷凍小管取出,於 37℃快速回溫,冷凍小管中的細胞液移入 離心管中離心 5 分鐘 1500 rpm,移除上清液。再加入 1ml 培養基,細胞
再次懸浮後於 10 cm dish 中加入 6-8 ml 的培養基,打散細胞均勻後,置入 37℃、5%CO2 細胞培養箱中靜置 24 小時後更換細胞培養基。
(2).細胞繼代培養方法
A549 培養於內含 90% Dulbecco’s Modified Eagle’s Medium-high glucose (DMEM)、10% FBS、1﹪P/S,待細胞長 8-9 分滿時,移除舊培 養基,以滅菌 PBS 2 ml 清洗 2~3 次後,加入 1ml 的 Trypsin ,並輕輕 搖晃培養皿使 Trypsin 均勻,再放到培養箱 3~5 分鐘後 (Typsin - EDTA 作用將細胞切下),再加入培養基 1ml 混勻,收集細胞液以 25℃、轉速 1500-2000 rpm 離心五分鐘。(沖洗最後一次要將 PBS 液體吸取乾淨,
避免 PBS 抑制掉 Trypsin ,而無法作用),移除上清液再加入適量培 養基打散細胞均勻後,再添加約 6-8ml 培養基並平均分配到 10 cm 培養 皿內,培養於 37℃、5% CO2的恆溫培養箱中,每天更換一次培養基。
(3).冷凍細胞保存,其目的是冷凍保存生長過多的細胞,以備日後繼代 之用途(冷凍保存細胞濃度為 1~5 x 106 cell/ml)
待細胞長至八分滿時,移除舊培養基,以滅菌 PBS 清洗 2~3 次後,
加入 1ml 的 Trypsin-EDTA,並輕輕搖晃 dish 使 Trypsin-EDTA 均勻,
再放到培養箱 3~5 分鐘後 (Typsin-EDTA 作用將細胞切下),再加入培 養基 1 ml 混合均勻,收集細胞液以 25℃、轉速 1500-2000 rpm 離心五 分鐘。移除上清液再加入適量 Freezing medium (冰浴, DMSO: FBS
=1:9) 打散細胞均勻後,平均分配於冷凍管內,並置於冰箱保存。
(4).細胞數計算
移除舊培養基,以滅菌 PBS 2ml 清洗 2~3 次,加入 1ml 的 Trypsin - EDTA ,並輕輕搖晃培養皿使 Trypsin - EDTA 均勻,再放到培養箱 3~5 分鐘後 ( Typsin - EDTA 作用將細胞切下),再加入培養基 1 ml 混 合均勻,收集細胞液,移入 15 ml 離心管中,取 10μl Trypan blue 和 10μl 細胞液到細胞計數器中,均勻抽吸後各取 10μl 至上方與下方細 胞計數器中,用顯微鏡觀察並計算細胞總數。
計算方法:(平均後之每方格細胞數×稀釋倍數)×104/ml
2.3 Tomatidine 處理
(1) Tomatidine 配製
Tomatidine 50mM 將之稀釋成 5mM(即稀釋 10 倍,稀釋液是使用 培養細胞之培養基),分別將之調配成 10μM、20μM、30μM、40μ M 等濃度於無菌的離心管中。
(2) Tomatidine 處理步驟
將 A549 細胞(5×105 cells)培養於 10 cm dish,隔日待細胞貼盤,細 胞約長至 4~5 分滿。加入不同濃度的 tomatidine (10、20、30、40μM) 及 與 tomatidine 最高濃度相同的 DMSO 當作溶媒作控制組,同時以未加
任何試劑作空白試驗,置於 37℃、5% CO2恆溫培養箱中培養 24 小時。
2.4 細胞存活率實驗(MTT assay) :
將 A549 (3x103 cell/well)培養於 96 孔培養盤,培養 24 hr 待細胞 貼附後移除培養液,加入不同濃度的 tomatidine (10、20、30、40 μM)
及未加任何試劑作空白試驗(Control),置於 37℃、5% CO2恆溫培養箱中 反應 24 小時;移除舊培養液,以滅菌 PBS 清洗 2 次後,加入 100 μl/well 內含 MTT (0.5 mg/ml)的培養液,隨即置於 37℃、5% CO2恆溫培養箱 中作用 2 小時;移除內含 MTT 的培養液,以未滅菌 PBS 清洗 1 次,再加 入 100 μl/well DMSO 作用 3 分鐘後,在 570 nm 波長下測定 OD 值,計算 出相對的細胞存活率。以未加藥物處理的細胞吸光值作為對照組,再以實 驗組的細胞吸光值除以對照組,計算出相對的細胞存活率百分比。
2.5 細胞侵入性實驗 (Matrigel invasion assay)
(1) 配製雙面含 0.1% matrigel-coated polycarbonate membrane:
將膜攤平於 10 cm dish 內(無菌操作),而兩面必須無氣泡、完全 浸潤於 0.1% growth factor reduce matrigel solution (BD biosciences),並 置於 37℃、5% CO2恆溫培養箱中作用至少一小時,隨後取出膜晾乾至 少一小時。
(2) 細胞處置:
將 A549 (5x105 cells/well) 培養於 10 cm dish 內,待細胞長至 7~8 分滿後,加入以血清培養基所配置之不同濃度的 tomatidine (10、
20、30、40 μM)及與 tomatidine 未加任何試劑作空白試驗(Control), 置於 37℃、5% CO2恆溫培養箱中反應 24 小時。
(3) Boyden chamber
A.下層:加入 30 μl/well 含血清的培養基,隨即覆蓋上含 0.1%
matrigel-coated polycarbonate membrane,霧面朝下,蓋上橡膠軟片後將 螺絲鎖緊。
B.上層:移除舊培養基,以滅菌 PBS 清洗兩次後,隨即以 Typsin-EDTA 作用一分鐘將細胞切下,再加入含血清培養基 1 ml 混 勻,收集細胞液以 4℃、1000 rpm 離心五分鐘;移除上清液再加入適 量無血清的培養基均勻打散細胞後計數,稀釋成 4 x 105 cells/ml,隨即 以無血清的培養基以 1:1 比例再稀釋混勻,50 μl/well( 2 x 105 cells/ml),隨後置於 37℃、5% CO2恆溫培養箱中作用六小時。
2.6 RNA 抽取流程(Total RNA extraction)
使 用 RBC Bioscience 廠 牌 total RNA extraction kit ﹝ Mini ﹞ Cat No.YRB50;Lot No.PG-924-10165;保存於室溫 15-25℃。
(1). Cell Harvesting
將藥物處理過後的細胞去除上清液,加入 PBS 2 ml 清洗兩次。
加入 500μl RB buffer(含 β-ME),並刮取細胞,冰在- 80℃ or -20℃冰箱 結晶 10~20 min(產生冰晶)讓細胞破更完全。(1ml RB buffer 須添加 10 μl β-ME)。
(2). Lysis
放置 filter Column set. 加入樣品到 Column 離心 2 分鐘,轉速 13000 rpm。將離心後的上清液移至新的 1.5 ml 離心管,filter Column set 丟棄。
(3). RNA Binding
先放置 RB Column set,將樣本與 400μl 70% 酒精混合後,上下 旋轉三次(先前 500μl RB buffer+400μl 70% 酒精=900μl),再取 500 μl 加入 RB Column,離心 2 分鐘轉速 13000 rpm,將剩餘的樣本取出 再一過次 Column,離心 2 分鐘轉速 13000 rpm,將 RB Column 取下放 置新的 1.5 ml tube,下管與廢液丟棄。
(4). Recommended Step:DNA residue degradation
20μl DNase I 和 10μl 10X DNase buffer 70μl 和 DEPC-H2O 加入 RB Column,靜置室溫 20 分鐘。(先將 10μl 10X DNase buffer +70μl DEPC-H2O 混好後再加入 20μl DNase I)
(5).Wash
加入 400 μl R-W1Buffer,離心 1 分鐘轉速 13000 rpm 將下層液 體拋棄,加入 600μl R-Wash Buffer(已加酒精),離心 1 分鐘轉速 13000
rpm 將下層液體拋棄,離心 3 分鐘轉速 13000 rpm 將剩餘酒精液體離乾 淨。
(6).RNA Elution
將 RB Columin 放至於新的 1.5 Tube,加入 50μl RNase free water (DEPC-H2O),離心 3 分鐘轉速 13000 rpm,離心 1 分鐘去 elute purified RNA。
2.7 核醣核酸定量(RNA quantitative)
(1).使用 CB-4500 微量分光光度計。
(2).使用 HITACHI U-1900 Ratio Beam Spectrophotometer 測定。
將吸光值換算成濃度公式:
,DNA inμg/μl。
也可換算 1 OD RNA=40ng/μl;1 OD DNA=50ng/μl(當然也要回 乘稀釋倍數)。
2.8 反轉錄聚合酶連鎖反應(Reverse transcriptase polymerase chain reaction ,RT-PCR)
(1).原理
反轉錄酶以 RNA 為模板合成 DNA 即 cDNA,接下來再利用 PCR 的原理大量增殖這段 cDNA。
(2).RNA 反轉錄過程:
(A).取 RNA sample 後,加入 Oligo-dT 0.5μl,以 DEPC-H2O 補體積 至 10.4μl,以 PCR machine 執行以下時間溫度。
(B).依序加入 4μl 5X First-Strand Buffer、2μl 10 mM dNTP Mix、2 μl O.1M DTT、1μl RNase inhibitor(40μM/μl)、0.6μl Reverse transcriptase 混合後以 PCR machine 執行以下時間溫度。
2.9 去氧核醣核酸定量(cDNA quantitative)
參閱 2.7.(1).與.2.7.(2).說明。
2.10 聚合酶連鎖反應(Polymerase chain reaction,PCR)
(1).擴增目標片段:
依序加入 cDNA sample、0.5μl Forward primer、0.5μl Reverse primer、2μl dNTPs、3.5μl 2.0 × Master Mix Red(MgCl2 1.5mM)混 合後以 DEPC-H2O 補體積至 20μl。
(2).Primer 黏合溫度:
使 DNA 的雙股有一半處於分離或變性狀態下的溫度稱熔點 (Tm),DNA 的 GC 含量對 Tm 有顯著性的影響,一般百力計算 Tm 值的公式:Tm=64.9+41×(NC+NG-16.4)÷(NA+NC+NT+NG),
黏合溫度視 primer 不同而調整黏合溫度,通常 primer 與模板 DNA 黏合的溫度約 Tm 值減 5℃,如果除了預期產物外,有一些非預期的 產物出現,則可逐漸提高黏合溫度約 1-2℃。
以下為本實驗的黏合溫度條件:
(A)MMP-2:primer 黏合溫度為 55℃。
(B)MMP-7:primer 黏合溫度為 57℃。
(C)MMP-9:primer 黏合溫度為 59℃。
(D)TIMP-1:primer 黏合溫度為 56℃。
(E)TIMP-2:primer 黏合溫度為 61℃。
(F)β-Actin:primer 黏合溫度為 56℃。
(3).DNA 凝膠電泳(Gel electrophoresis) (A)原理
利用凝膠電泳分析 DNA 片段大小,DNA 是一種酸,在中性條 件下帶有負電荷,因此會往正極泳動。
(B)操作步驟
將 10×TBE Buffer 稀釋成 0.5×TBE Buffer,取 50ml 或 80ml 溶液,
用微量天秤粉末的 agarose 0.75g 或 1.20g,將膠配製成 1.5% agarose gel,agarose 在高溫下熔解,插上梳子,在膠體冷凝後,拿掉梳子留 下的槽溝,將 DNA 注入井孔中,使電流通過製備的水平凝膠體,以 100V 運行 45 分鐘後,進行染色。
(4).Ethidium Bromide(EtBr)染色,拍照記錄結果。
(A)原理
利用細胞化學染色可觀察細胞內化學分子的位置,DNA、RNA 為酸性,可用鹼性染料。DNA 和染劑結合在紫外光照射下可發出粉
紅色產物。
(B)操作步驟
將產物 1.5% agarose gel 放入 Ethidium Bromide(EtBr)染色 20 分 鐘,退染 20 分鐘,將產物 1.5% agarose gel 撈起,照膠(Digimage system
&UV Transilluminator),拍照記錄結果。
2.11 統計分析 (Stastistical analysis)
數據為平均值±標準差 (Mean ± SD) 來表示,以 one-way ANOVA 來 評估統計上的差異,而 p<0.05 表示具有顯著差異。
第三章 結果 3.1 Tomatidine 對 A549 細胞存活率的影響
為了測試 tomatidine 對於人類肺腺癌上皮細胞的影響,首先利用 MTT assay 測試對細胞存活率的影響。不同濃度的 tomatidine (10、20、30、40、
50 μM)及與 tomatidine 最高濃度相同的溶媒作控制組,同時以未加任何 試劑做空白試驗,分別處理 24 小時。結果發現,在 50 μM 的 tomatidine 處理 24 小時,大約抑制了 15%的細胞存活率(圖 1)。此結果發現 tomatidine 雖然可以降低 A549 細胞的存活率,但並沒有很強的毒性。
3.2 Tomatidine 對 A549 細胞侵入能力的影響
此實驗利用 Boyden chamber assay 測試 tomatidine 對 A549 細胞侵入能 力的影響,結果發現在濃度 20μM 的 tomatidine 抑制細胞穿越 0.1 % growth factor reduce matrigel-coated polycarbonate membrane 的程度達 20
%,而濃度 40μM 的 tomatidine 的抑制效果更為明顯,抑制侵入程度達 40 %,且藥物抑制效果亦呈現出劑量依賴性的關係,因此顯示出 tomatidine 能抑制 A549 細胞的侵入能力(圖 2)。
3.3 Tomatidine 抑制 A549 細胞 MMP-2 基因的表現
MMP-2,MMP-9 與 MMP-7,其蛋白可以降解細胞外基質,促使癌細 胞得以通過基底膜,藉由進入血管系統或淋巴系統進而達到轉移的目的。
本研究以 reverse transcription-PCR 分析 tomatidine 對 A549 細胞之 MMPs 基 因表現的影響。以不同濃度的 tomatidine (10、20、30、40μM)處理 A549 細胞 24 小時,萃取細胞 RNA,每組的 RNA 定量並調整至相同濃度,依引 子調整黏合溫度與 PCR 反應的循環數,循環數由 35 個循環數開始遞減,測 試最佳的實驗條件。結果發現操作 31~32 次循環數即可明顯觀察到 A549 細 胞的 MMP-2,MMP-9 與 MMP-7 基因表現。在 MMP-2 基因表現的實驗,
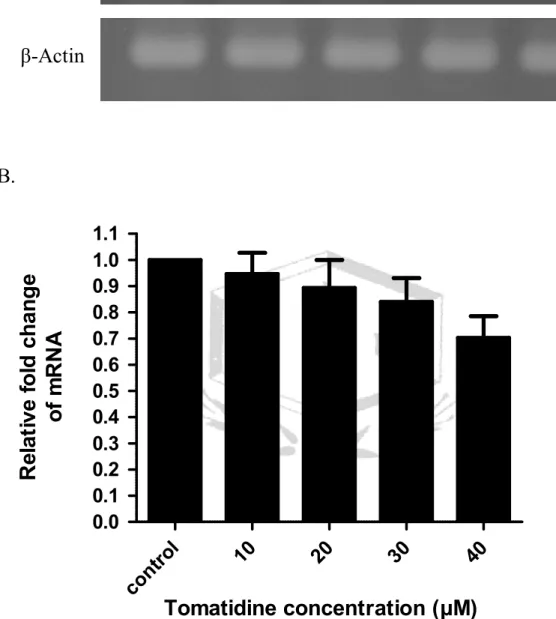
以 55℃引子黏合溫度,進行 PCR 反應 31~32 次循環數,並以 β-actin 的基 因表現當內控制組,進行三重複試驗。結果發現 tomatidine 的處理劑量增 加至 40μM 時,可抑制 MMP-2 基因的表現達 30﹪(圖 3)。
3.4 Tomatidine 抑制 A549 細胞 MMP-9 基因的表現
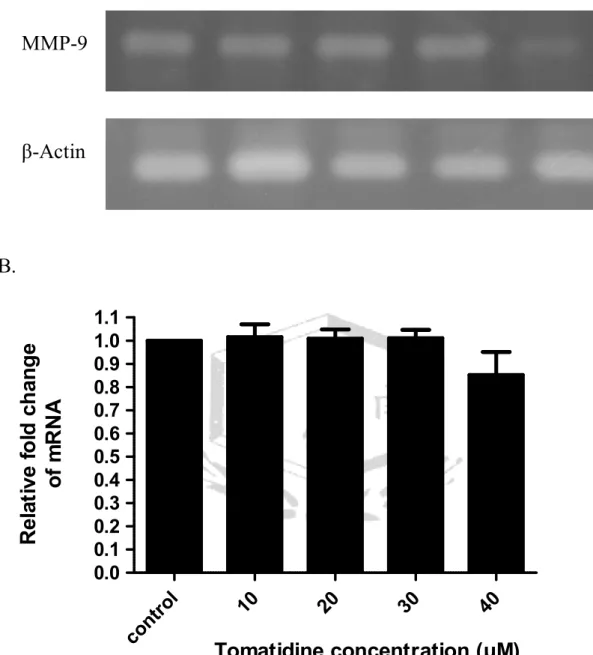
在 MMP-9 基因表現的實驗,以 59℃引子黏合溫度,進行 PCR 反應 32~33 次循環數。結果發現 tomatidine 的處理劑量增加至 40μM 時,可抑制 MMP-9 基因的表現達 15﹪(圖 4)。
3.5 Tomatidine 抑制 A549 細胞 MMP-7 基因的表現
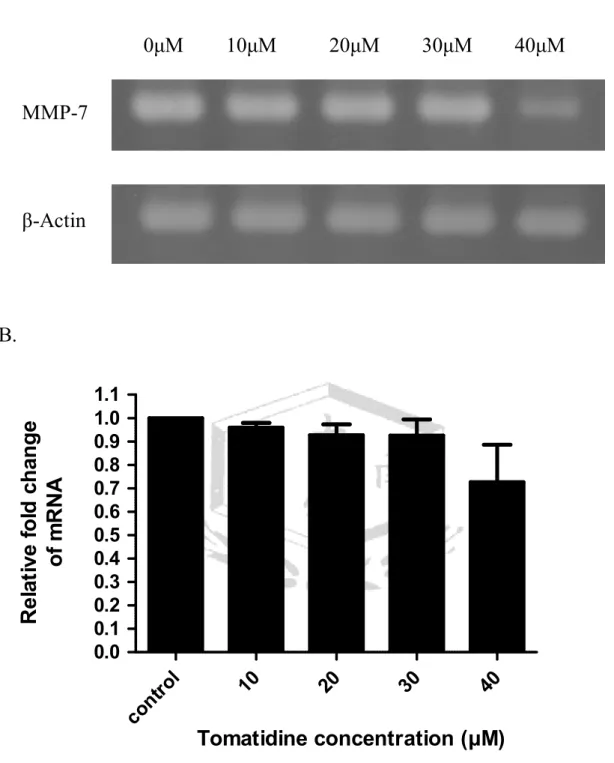
在 MMP-7 基因表現的實驗,以 57℃引子黏合溫度,進行 PCR 反應 32~33 次循環數。結果發現 tomatidine 的處理劑量增加至 40μM 時,可抑制 MMP-7 基因的表現達 27﹪(圖 5)。
3.6 Tomatidine 對於 A549 細胞 TIMP-1 基因的表現
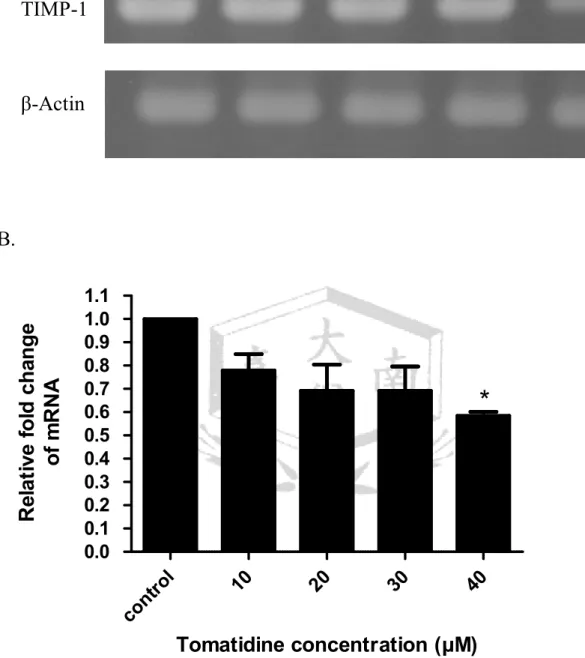
TIMP 能降低基質金屬蛋白酶的活性,而減少細胞外基質的分解。若要 維持細胞外基質的完整性,MMP 和 TIMP 之間的作用必須達到平衡。在 TIMP-1 基因表現的實驗,以 56℃引子黏合溫度,進行 PCR 反應 32~33 次 循環數。結果發現 tomatidine 的處理劑量增加至 40μM 時,抑制 TIMP-1 基因的表現達 42﹪(圖 6)。
3.7 Tomatidine 對於 A549 細胞 TIMP-2 基因的表現
在 TIMP-2 基因表現的實驗,以 61℃引子黏合溫度,進行 PCR 反應 32~33 次循環數。結果發現 tomatidine 的處理劑量增加至 40μM 時,抑制 TIMP-2 基因的表現無劑量效應(圖 7)。
第四章 討論
在腫瘤發生或癌症轉移時,會分泌某些蛋白水解酵素去破壞並分解細 胞周圍組織與細胞外基質,進而促使癌細胞可以轉移至其他的器官或部位
(22,23)
。在過去的文獻表示,癌細胞發生轉移時會釋放大量的基質金屬蛋白酶 來降解細胞外基質。細胞外基質,乃是一種複雜的細胞間凝膠,由構成某 一組織之細胞所分泌與製造。在上皮組織中,上皮細胞所處位置基質或間 質細胞,可產生各類的細胞外基質,在細胞遷移、細胞增殖、分化及癌症 佔重要角色(19,20)。
細胞在癌化過後基質金屬蛋白酶的表現量會大幅的提升,惡性腫瘤發 生轉移的部位亦可測得大量的 MMPs 。在相關研究中證實癌細胞藉由 MMPs 來分解細胞外基質及破壞基底膜,再藉由血管及淋巴系統將癌細胞 轉移到其他器官(47)。在 MMP 家族中, MMP -1 、 MMP - 3 、 MMP - 7 、 MMP - 9 、 MMP- 10 、 MMP - 12 、 MMP - 13 ,分解細胞外基質具有 破壞基底膜的能力(30)。MMP-9 常表現於惡性細胞及發炎的細胞(包含分解組 織巨噬細胞與嗜酸性白血球)。而 MMP-2 增加的免疫反應酵素常見於乳 癌、大腸癌、胃癌的腺癌細胞中。基質金屬蛋白抑制劑能降低基質金屬蛋 白過度表現導致細胞外基質瓦解,其中 TIMP - 1 會影響 MMP - 9 ,而 TIMP - 2 會影響 MMP - 2 (48)。
本研究證明,tomatdine 具有抑制細胞侵入的作用。首先,為了先測試
tomatidine 對人類肺腺癌上皮細胞是否具有毒殺性與抑制生長的效果,利用 MTT assay 進行初步分析。在不同濃度的 tomatidine (10、20、30、40μM),
分別處理 24 小時後,發現 tomatidine 雖然具有抑制生長的效果 ,但並不 具有很強的毒殺性。接著進一步了解 tomatidine 抑制人類肺腺癌上皮細胞 侵入的影響,利用 Boyden chamber assay 進行分析,發現 tomatidine 對人 類肺癌細胞 A549 有明顯的抑制侵入作用。
接著以 reverse transcription - PCR 分析基因表 現。 以不同 濃度的 tomatidine (10、20、30、40μM) 處理 A549 細胞 24 小時後,萃取細胞 RNA ,每組的 RNA 定量後並調整至相同濃度,依引子調整黏合溫度與 PCR 反應的循環數,循環數由 35 個循環數開始遞減,測試最佳的實驗條 件。以 RT - PCR 觀察 MMP - 2、 MMP - 7、 MMP - 9 、 TIMP - 1 、 TIMP - 2 mRNA 的表現量。結果發現 tomatidine 對於 MMP - 2 / - 7 / - 9 的 mRNA 表現量有抑制效果。結果發現進行 31~32 次循環數即可明顯觀察到 A549 細胞的 MMP - 2 , MMP - 9 與 MMP - 7 基因表現。在 MMP - 2 基 因表現的實驗,以 55℃引子黏合溫度,進行 PCR 反應 31~32 次循環數,
並以 β - actin 的基因表現當內控制組,進行三重複試驗。在 MMP - 9 基 因表現的實驗,以 59℃引子黏合溫度,進行 PCR 反應 32~33 次循環數。
在 MMP - 7 基因表現的實驗,以 57℃引子黏合溫度,進行 PCR 反應 32~33 次循環數。結果發現 tomatidine 在 40μM 時,可抑制 MMP - 2 基因表現
達 30﹪、 MMP - 9 基因表現達 15﹪與 MMP - 7 基因表現達 27﹪。 TIMP 能降低基質金屬蛋白酶的活性,而減少細胞外基質的分解。若要維持細胞 外基質的完整性, MMP 和 TIMP 之間的作用必須達到平衡。在 TIMP - 1 基因表現的實驗,以 56℃引子黏合溫度,進行 PCR 反應 32~33 次循環數。
在 TIMP - 2 基因表現的實驗,以 61℃引子黏合溫度,進行 PCR 反應 32~33 次循環數。結果發現 tomatidine 的處理劑量增加至 40μM 時,可抑制 TIMP - 1 基因的表現達 42﹪、但對 TIMP - 2 基因的表現無影響。
TIMP - 1 的表現雖然被認為與抑制癌細胞轉移有關,但是在臨床研究 卻發現,許多癌細胞有 TIMP - 1 有大量表現(49,50,51)。 TIMP - 1 也被發現 可能活化肝癌細胞的轉移(52)。本研究發現 tomatidine 可抑制 TIMP - 1 的 表現,但如何進一步抑制肝癌細胞 A549 的侵入及轉移,仍有待更深入的研 究。
第五章 結論
Tomatdine 能有效的抑制人類肺腺癌細胞入侵的能力。 Tomatdine 可 抑制 MMP -2 / -7 / -9 表現量,進而減少細胞外基質的分解。此外,
tomatdine 會抑制 TIMP-1 表現。本論文的研究發現 tomatdine 可抑制 MMP -2 / -7 / -9 的表現,進而抑制 A549 細胞的入侵作用。
第六章 參考文獻
1. 中華民國行政院衛生屬署.http://www.doh.gov.tw
2. Lee GY, Jang JS, Lee SY, et al. XPC polymorphisms and lung cancer risk.
Int J Cancer. 2005;115(5):807-813.
3. Wang Y, Yang H, Liu H, Huang J, Song X. Effect of staurosporine on the mobility and invasiveness of lung adenocarcinoma A549 cells: an in vitro study. BMC Cancer. 2009;9:174.
4. Pore MM, Hiltermann TJ, Kruyt FA. Targeting apoptosis pathways in lung cancer. Cancer Lett. 2010;10:1016-1025.
5. Shivapurkar N, Reddy J, Chaudhary PM, Gazdar AF. Apoptosis and lung cancer: a review. J Cell Biochem. 2003;88(5):885-898.
6. Lieber M, Smith B, Szakal A, Nelson-Rees W, Todaro G. A continuous tumor-cell line from a human lung carcinoma with properties of type II alveolar epithelial cells. Int J Cancer. 1976;17(1):62-70.
7. Chang HC, Cho CY, Hung WC. Downregulation of RECK by promoter methylation correlates with lymph node metastasis in non-small cell lung cancer. Cancer Sci. 2007;98(2):169-173.
8. Chen JH, Lin HH, Chiang TA, et al. Gaseous nitrogen oxide promotes human lung cancer cell line A549 migration, invasion, and metastasis via
iNOS-mediated MMP-2 production. Toxicol Sci. 2008;106(2):364-375.
9. Ambiru, S. Miyazaki, M. Ito, H. Type IV collagenase activities in human colorectal cancers and its role in cancer invasion and metastasis. Nippon Shokakibyo Gakkai Zasshi. 1993;90(7):155-1561.
10. Thaloor, D. Singh, A. K. Sidhu, G. S. Prasad, P. V. Kleinman, H. K.
Maheshwari, R. K. Inhibition of angiogenic differentiation of human umbilical vein endothelial cells by curcumin. Cell Growth Differ.1998;9(4):305-312.
11. Murphy G, Dochery AJ. The matrix metalloproteinases and their inhibitors. Am J Respir Cell Mol Biol. 1992;7(2):120-125.
12. Curran S, Murray GI. Matrix metalloproteinases in tumour invasion andmet astasis. J Pathol. 1999;189(3):300-308.
13. McCawley LJ, Matrisian LM. Matrix metalloproteinases: they’re not just for matrix anymore! Curr Opin Cell Biol. 2001;13:534–540.
14. PASCHOS KA, CANOVAS D, BIRD NC, Enzymatic function of multiple origins regulates the progression of colorectal cancer and the development of metastases, Hippokratia, 2009, 13(1):23–31.
15. SUN XF, ZHANG H, Clinicopathological significance of stromal variables: angiogenesis, lymphangiogenesis, inflammatory infiltration,
MMP and PINCH in colorectal carcinomas, Mol Cancer, 2006, 5:43.
16. NOVAC L, NICULESCU M, PLESEA IE, TUDORACHE ST, Matrix metaloproteinazele în sarcina normală şi în avortul spontan recurent, InfoMedica, 2007, 3(141):31–33.
17. Butler GS, Overall CM. Updated biological roles for matrix metalloproteinases and new "intracellular" substrates revealed by degradomics. Biochemistry. 2009;48(46):10830-10845.
18. Strongin AY. Mislocalization and unconventional functions of cellular MMPs in cancer. Cancer Metastasis Rev. 2006;25(1):87-98.
19. Nagase H, Visse R, Murphy G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc Res. 2006;69(3):562-573.
20. Cawston TE, Wilson AJ. Understanding the role of tissue degrading enzymes and their inhibitors in development and disease. Best Pract Res Clin Rheumatol. 2006;20(5):983-1002.
21. Hadler-Olsen E, Fadnes B, Sylte I, Uhlin-Hansen L, Winberg JO.
Regulation of matrix metalloproteinase activity in health and disease. FEBS J. 2011;278(1):28-45.
22. Schropfer, A. Kammerer, U. Kapp, M. Dietl, J. Feix, S. Anacker, J.
Expression pattern of matrix metalloproteinases in human
gynecological cancer cell lines. BMC Cancer. 2010;10:553.
23. Coussens LM, Fingleton B, Matrisian LM. Matrix metalloproteinase inhibitors and cancer: trials and tribulations. Science.
2002;295(5564):2387-2392.
24. Chetty C, Lakka SS, Bhoopathi P, Kunigal S, Geiss R, Rao JS. Tissue inhibitor of metalloproteinase 3 suppresses tumor angiogenesis in matrix metalloproteinase 2-down-regulated lung cancer. Cancer Res.
2008;68(12):4736-4745
25. W. G. Stetler-Stevenson. Dynamics of matrix turnover during pathologic remodeling of the extracellular matrix. Am. J. Pathol. 1996 ; 148, 1345-1350.
26. Curran S, Murray GI. Matrix metalloproteinases in tumour invasion and metastasis. J Pathol. 1999;189(3):300-308.
27. Welser-Alves JV, Crocker SJ, Milner R. A dual role for microglia in promoting tissue inhibitor of metalloproteinase (TIMP) expression in glial cells in response to neuroinflammatory stimuli. J Neuroinflammation.
2011;8:61.
28. Wang H, Lafdil F, Wang L, Yin S, Feng D, Gao B. Tissue inhibitor of me ta llo p ro te inas e 1 ( TIM P -1 ) de fi c ie nc y e xace rba tes car bo n
tetrachloride-induced liver injury and fibrosis in mice: involvement of hepatocyte STAT3 in TIMP-1 production. Cell Biosci. 2011;1(1):14.
29. Bahudhanapati H, Zhang Y, Sidhu SS, Brew K. Phage display of tissue inhibitor of metalloproteinases-2 (TIMP-2): Identification of selective inhibitors of collagenase-1 (MMP-1). J Biol Chem.
2011;286(36):31762-31770.
30. Mroczko B, Lukaszewicz-Zajac M, Gryko M, Kedra B, Szmitkowski M.
Clinical significance of serum levels of matrix metalloproteinase 2 (MMP-2) and its tissue inhibitor (TIMP-2) in gastric cancer. Folia Histochem Cytobiol. 2011;49(1):125-131.
31. Norton, S. A. Useful plants of dermatology. III. Corticosteroids, Strophanthus, and Dioscorea. J Am Acad Dermatol. 1998;38(2-1):256-9.
32. Apte SS, Mattei M-G, Olsen BR. Cloning of the cDNA Encoding Human Tissue Inhibitor of Metalloproteinases-3 (TIMP-3) and Mapping of the TIMP3 Gene to Chromosome 22. Genomics. 1994;19(1):86-90.
33. Huang W, Li WQ, Dehnade F, Zafarullah M. Tissue inhibitor of metalloproteinases-4 (TIMP-4) gene expression is increased in human osteoarthritic femoral head cartilage. J Cell Biochem. 2002;85(2):295-303.
34. Tissue inhibitor of metalloproteinases-4. The road less traveled. Molecular Cancer. 2008;7:85-96.
35. Pullen NA, Anand M, Cooper PS, Fillmore HL. Matrix metalloproteinase-1 expression enhances tumorigenicity as well as tumor-related angiogenesis and is inversely associated with TIMP-4 expression in a model of glioblastoma. J Neurooncol. 2011;10:11060-11071.
36. Friedman, M. Levin, C. E. Lee, S. U. Tomatine-containing green tomato extracts inhibit growth of human breast, colon, liver, and stomach cancer cells. J Agric Food Chem. 2009;57(13):5727-33.
37. Chiu FL, Lin JK. Tomatidine inhibits iNOS and COX-2 through suppression of NF-kappaB and JNK pathways in LPS-stimulated mouse macrophages. FEBS Lett. 2008;582(16):2407-2412.
38. Norton SA. Useful plants of dermatology. III. Corticosteroids, Strophanthus, and Dioscorea. J Am Acad Dermatol. 1998;38(2):256-259.
39. Steel CC, Drysdale RB. Electrolyte leakage from plant and fungal tissue and disruption of liposome membranes by a-tomatine. Phytochemistry.
1988;27:1025–1030.
40. Keukens EA, de Vrije T, van den Boom C, et al. Molecular basis of
glycoalkaloid induced membrane disruption. Biochim Biophys Acta. Dec 13
1995;1240(2):216-228.
41. Shih Y-W, Shieh J-M, Wu P-F, Lee Y-C, Chen Y-Z, Chiang T-A.
[alpha]-Tomatine inactivates PI3K/Akt and ERK signaling pathways in human lung adenocarcinoma A549 cells: Effect on metastasis. Food and Chemical Toxicology. 2009;47(8):1985-1995.
42. Chatel, G. Ganeff, C. Boussif, et al. Hedgehog signaling pathway is inactive in colorectal cancer cell lines. Int J Cancer. 2007;121(12):2622-2627.
43. Nagai, S. Nakamura, M. Yanai, K, et al. Gli1 contributes to the invasiveness of pancreatic cancer through matrix metalloproteinase-9 activation. Cancer Sci. 2008;99(7):1377-1384.
44. Ruiz i Altaba A, Sanchez P, Dahmane N. Gli and hedgehog in cancer:
tumours, embryos and stem cells. Nat Rev Cancer. 2002;2(5):361-372.
45. Incardona JP, Gaffield W, Lange Y, et al. Cyclopamine inhibition of Sonic hedgehog signal transduction is not mediated through effects on cholesterol transport. Dev Biol. 2000;224(2):440-452.
46. Lipinski RJ, Dengler E, Kiehn M, Peterson RE, Bushman W. Identification and characterization of several dietary alkaloids as weak inhibitors of hedgehog signaling. Toxicol Sci. 2007;100(2):456-463.
47. Lai KC, Hsu SC, Yang JS, et al. 2-(3-Methoxyphenyl)-6,
7-methylenedioxoquinolin-4-one, a novel synthetic compound, inhibited migration and invasion in TSGH8301 human bladder cancer cells. Hum Exp Toxicol. 2011;30(8):1045-1052.
48. Reichenberger F, Eickelberg O, Wyser C, Perruchoud AP, Roth M, Tamm M. Distinct endobronchial expression of matrix-metalloproteinases (MMP) and their endogenous inhibitors in lung cancer. Swiss Med Wkly.
2001;131(19-20):273-279.
49. Kemik O, Kemik AS, Sumer A, et al. Levels of matrix metalloproteinase-1 and tissue inhibitors of metalloproteinase-1 in gastric cancer. World J Gastroenterol. 2011;17(16):2109-2112.
50. Safranek J, Pesta M, Holubec L, et al. Expression of MMP-7, MMP-9, TIMP-1 and TIMP-2 mRNA in lung tissue of patients with non-small cell lung cancer (NSCLC) and benign pulmonary disease. Anticancer Res.
2009;29(7):2513-2517.
51. Schrotzlmair F, Kopitz C, Halbgewachs B, et al. Tissue inhibitor of metalloproteinases-1-induced scattered liver metastasis is mediated by host-derived urokinase-type plasminogen activator. J Cell Mol Med.
2010;14(12):2760-2770.
52. Schelter F, Halbgewachs B, Baumler P, et al. Tissue inhibitor of
metalloproteinases-1-induced scattered liver metastasis is mediated by hypoxia-inducible factor-1alpha. Clin Exp Metastasis. 2011;28(2):91-99.
第七章 圖表
MTT assay
Cont rol
DMSO 10 20 30 40
0 10 20 30 40 50 60 70 80 90 100 110
Tomatidine concentration (μM)
Cell viability (%)
圖 1. Tomatidine 對 A549 細胞存活率的影響
將 A549 細胞(3×103 cells/well)培養至 96 孔盤中,以不同濃度的 Tomatidine(10、20、30、40 μM)及未加任何試劑作空白試驗(Control)
做比較,培養 24 小時後以 MTT assay 測定細胞存活率。數值皆以平均值
(Mean)±標準差(SD)表示,三重覆。
A.
20μM 30μM
40μM
Control 10μM
B.
control
10 20 30 40
0 10 20 30 40 50 60 70 80 90 100 110
*
Tomatidine concentration (μM)
Cell invasion (%)
圖 2. Tomatidine 對 A549 細胞侵入能力的影響
(A)A549 細胞以不同劑量(Control、10、20、30、40 μM)的 tomatidine 處 理 24 小時,並以 Boyden chamber assay 測量 tomatidine 對 A549 細胞侵入能 力的影響。將細胞混入無血清培養基培養於上層 chamber,下層 chamber 為 10 %血清培養基,並將 polycarbonate filter (8 μM 孔徑大小) 以 0.1 % growth factor reduce Matrigel 處理。培養六小時後, polycarbonate filters 以 甲醇固定細胞後,以 Liu’s A、Liu’s B reagent 進行染色,並以顯微鏡拍照 記數侵入 polycarbonate filters 下層的 A549 細胞。數值皆以平均值 (Mean) ± 標準差 (SD) 表示,三重複;*代表 p<0.05。
A.
B.
control 10 20 30 40
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 1.1
Tomatidine concentration (μM) Relative fold change of mRNA
圖 3. Tomatidine 抑制 A549 細胞 MMP-2 的表現
(A). Tomatidine 調控 A549 細胞的 MMP-2 mRNA 表現。以不同濃度的 tomatidine (10、20、30、40μM) 處理 A549 細胞 24 小時,萃取細胞 RNA,
以 RT-PCR 分析 A549 細胞 MMP-2 之 mRNA 表現量,三重覆。β-actin 為 MMP-2
β-Actin
0μM 10μM 20μM 30μM 40μM
內控制組。(B).將圖 A. MMP-2 與β-actin 的表現,使用 AlphaImager 2200 軟體將結果量化並繪圖。
A.
B.
control
10 20 30 40
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 1.1
Tomatidine concentration (μM) Relative fold change of mRNA
圖 4. Tomatidine 抑制 A549 細胞 MMP-9 的表現
(A). Tomatidine 調控 A549 細胞的 MMP-9 mRNA 表現。以不同濃度的 tomatidine (10、20、30、40μM) 處理 A549 細胞 24 小時,萃取細胞 RNA,
以 RT-PCR 分析 A549 細胞 MMP-9 之 mRNA 表現量,三重覆。β-actin 為 0μM 10μM 20μM 30μM 40μM
MMP-9
β-Actin
內控制組。(B).將
圖 A.
MMP-9 與β-actin 的表現,使用 AlphaImager 2200 軟體將結果量化並繪圖。A.
B.
control
10 20 30 40
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 1.1
Tomatidine concentration (μM) Relative fold change of mRNA
圖 5. Tomatidine 抑制 A549 細胞 MMP-7 的表現
(A). Tomatidine 調控 A549 細胞的 MMP-7 mRNA 表現。以不同濃度的 tomatidine (10、20、30、40μM) 處理 A549 細胞 24 小時,萃取細胞 RNA,
以 RT-PCR 分析 A549 細胞 MMP-7 之 mRNA 表現量,三重覆。β-actin 為 MMP-7
β-Actin
0μM 10μM 20μM 30μM 40μM
內控制組。(B). 將圖 A. MMP-7 與β-actin 的表現,使用 AlphaImager 2200 軟體將結果量化並繪圖。
A.
B.
control
10 20 30 40
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 1.1
*
Tomatidine concentration (μM) Relative fold change of mRNA
圖 6. Tomatidine 對於 A549 細胞 TIMP-1 的表現
(A). Tomatidine 調控 A549 細胞的 TIMP-1 mRNA 表現。以不同濃度的 tomatidine (10、20、30、40μM) 處理 A549 細胞 24 小時,萃取細胞 RNA,
以 RT-PCR 分析 A549 細胞 TIMP-1 之 mRNA 表現量,三重覆。β-actin 為 TIMP-1
0μM 10μM 20μM 30μM 40μM
β-Actin
內控制組。(B). 將圖 A. TIMP-1 與β-actin 的表現,使用 AlphaImager 2200 軟體將結果量化並繪圖。
A.
B.
cont
rol 10 20 30 40
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 1.1
Tomatidine concentration (μM) Relative fold change of mRNA
圖 7. Tomatidine 對於 A549 細胞 TIMP-2 的表現
(A). Tomatidine 調控 A549 細胞的 TIMP-2 mRNA 表現。以不同濃度的 tomatidine (10、20、30、40μM) 處理 A549 細胞 24 小時,萃取細胞 RNA,
以 RT-PCR 分析 A549 細胞 TIMP-2 之 mRNA 表現量,三重覆。β-actin 為 內控制組。(B). 將圖 A. TIMP-2 與β-actin 的表現,使用 AlphaImager 2200
0μM 10μM 20μM 30μM 40μM TIMP-2
β-Actin
軟體將結果量化並繪圖。
附錄圖表
附錄 1. MMP 在腫瘤細胞移動中所扮演的角色。
附錄 2.腫瘤轉移的過程。
附錄 3. Tomatidine 的化學結構式。
附錄 4.Tomatidine 抗發炎的可能機制。