紫杉醇對人類肝癌細胞株乙醯轉移酵素的基因表現及細胞週期的影響; Paclitaxel Affected N-Acetyltransferase Gene Expression and Cell Cycle on Human Liver Tumor Cells
90
0
0
全文
(2) 誌. 謝. 俗話說:「活到老,學到老」 ,尤其以救人為職志的臨床醫師,更應時時充 實本職學能,跟上時代腳步。離開學校的學生生活已二十年,畢業後進入外 科醫師的養成階段—住院醫師訓練,著重在疾病診斷與手術技術及手術前後 的照護。到了主治醫師階段,才逐漸行有餘力去思考「What?」「Why?」「How?」 等問題。醫學的進步日新月異,為學真如逆水行舟,不進則退。近二十年來 的細胞及分子生物學的蓬勃發展,與臨床醫學更是息息相關,這些學問都不 是昔日的我所能接觸的到。身為一個二十一世紀的外科醫師,在腫瘤學及移 植醫學的領域,更應該有所鑽研。兩年前,在蔡董事長、林院長、洪副院長 及張文正博士的鼓勵下報考醫研所,並且幸運地能在張教務長與鍾景光博士 的諄諄指導下從事研究。兩年來更在陳偉德所長以及所裡各位教授的鞭策, 同學間的互相勉勵下,拓展人生視野,學習研究方法等為學之道。尤其感謝 鍾教授自始至終地教誨與指正,才能有今天的成果。 幕起幕落,舞台上的汗水希望沒有白流。感謝醫院同仁的支持,更感謝 我最愛的家人,吾妻長湘及小孩爵因、爵安、上儀,沒有你們的陪伴、支持、 幫忙與鼓勵,我是無法順利畢業的。 「結束是另一個階段的開始」,我將秉持 師長們的教導,傳授給我的學生,並繼續從事研究,希望應用所學,而服務 於社會人群。. i.
(3) 中文摘要 惡性腫瘤已經連續 20 年位居國人十大死亡原因之首,每年因肝癌死亡的 平均人數約有 5,000~6,000 人,原發性肝癌如未積極治療,病人往往在數個月 或數週內死亡,手術切除目前仍是唯一能治癒原發性肝癌的治療方法。當手 術後腫瘤多發性復發,或因身體狀況不適合手術,抑或是腫瘤發生遠處轉移 時,尋找有效的化學治療藥物來代替手術極為迫切。 紫杉醇(Paclitaxel)在乳癌,卵巢癌和在非小型細胞肺癌的治療效果良好, 在肝癌則未定論。本實驗以 Paclitaxel 對肝癌細胞株(HA 22 T/VGH)的細胞週 期的作用,發現 Paclitaxel 會抑制細胞週期在 G2/M 期,而其 cyclins 的控制與 cyclin-dependent. kinase 的關係研究,發現 Paclitaxel 在高濃度時( > 2.5 mM). 可抑制 cyclin B、D 和 CDK1 的表現,對 cyclin A、E 和 CDK2 則相反地呈過度 表現。另外,在 Paclitaxel 對 NAT 的活性研究發現,Paclitaxel 可以抑制細胞 內胞質液 NAT 的活性,且與藥物濃度、作用時間相關。這些發現似乎可以做 為 Paclitaxel 治療肝癌的立基。. ii.
(4) 目. 錄 頁 數. 誌謝....................................................................................................i 中文摘要 ............................................................................................ii 目錄....................................................................................................iii 表目錄 ................................................................................................iv 圖目錄 ................................................................................................v 符號及縮寫.........................................................................................vii 第一章 前言.......................................................................................1 第二章 材料.......................................................................................13 第三章 方法.......................................................................................18 第四章 結果.......................................................................................26 第五章 討論.......................................................................................30 第六章 結論.......................................................................................35 參考文獻 ............................................................................................37 英文摘要 ............................................................................................79 作者簡歷 ............................................................................................81 授權書 ................................................................................................82. iii.
(5) 表 目 錄 頁 數 表 1 常見的化學致癌物. ........................................................................................52 表 2 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH NAT 活性的影響 ........53. iv.
(6) 圖 目 錄 頁 數 圖 1 三個重要的,且實驗上常用的化學致癌物之構造. .........................................54 圖 2 2-Aminofluorene 之結構式.....................................................................................54 圖 3 2-Aminofluorene (2-AF)的代謝及其代謝物與 DNA 之作用 ...............................55 圖 4 細胞週期及其調控因子之間關係 .........................................................................56 圖 5 不同濃度的 Paclitaxel 在不同作用時間下對 HA 22T/VGH 的影響 ...................57 圖 6 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH DNA 的影響 .....................60 圖 7 不同濃度的 Paclitaxel 在 24 小時作用時間下對 HA 22T/VGH 細胞週期 的影響 .....................................................................................................................61 圖 8 在不同時間不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH 細胞週期 的影響 .....................................................................................................................67 圖 9.1 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH cyclin A 的影響 .............70 圖 9.2 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH cyclin B 的影響 .............70 圖 9.3 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH cyclin D1 的影響 ...........71 圖 9.4 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH cyclin D3 的影響 ...........71 圖 9.5 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH cyclin E 的影響..............72 圖 10.1 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH CDK1 的影響 ..............73 圖 10.2 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH CDK2 的影響 ..............74 圖 11 HPLC 檢測不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH NAT 活性 .....75. v.
(7) 圖 11-1 在 24 小時不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH 胞質液 NAT 活性的影響 .......................................................................................................76 圖 11-2 在不同時間同一濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH 胞質液 NAT 活性的影響 .......................................................................................................77 圖 12 不同濃度的 Paclitaxel 對肝癌細胞株 HA 22T/VGH NAT1 基因表現的影響 ..78. vi.
(8) 符號與縮寫 HA 22/VGH: human hepatocellular carcinoma cell line DMSO:dimethyl sulfoxide DTT: 1,4-dithiothreitol Rnase: ribonuclease A PMSF: phenylmethylsulfonyl fluoride EDTA: ethylenediaminetetraacetic acid Acetyl CoA: acetyl coenzyme A MgCl2 : magnesium chloride KCl: potassium chloride NaCl: sodium chloride BSA: bovine serum albumin PBS: phosphate Buffered Saline NAT: N-acetyltransferase 2-AF: 2-aminofluorene 2-AAF: 2-acetylaminofluorene CDK: cyclin dependent kinase PS: phosphatidylserine PI: propidium iodide. vii.
(9) RTPCR: reverse transcriptase polymerase chain reaction PARP: poly (ADP-ribose) polymerase FACS: flow cytometry HPLC: high performance liquid chromatography PHS: prostaglandin H synthase UGT: UDP-glucuronosyltransferase OAT: O-acetyltransferase NOAT: N-O-acetyltransferase SULT: Sulfotransferase DAC: Deacetyltransferase. viii.
(10) 第一章. 前. 言. 根據衛生署 91 年 6 月所公佈的資料,自民國 71 年起至今,惡性腫瘤連 續 20 年位居國人十大死因之首,其死亡率每十萬人口為 147.68 人,民國九十 年約有三萬三千人死於惡性腫瘤。平均每十五分鐘就奪走一條寶貴的生命。 台灣每年因肝癌死亡的平均人數約有 5,000~6,000 人,在男性惡性腫瘤死亡原 因中佔第一位,在女性癌症中也名列第二位。男性和女性病患比例可高達 8:1; 其好發年齡層在 50 歲到 70 歲間 (1-2)。因此,肝癌的研究在臨床醫學及公共衛 生領域顯得格外重要。原發性肝癌是一種非常惡性的癌症,臨床上如未積極 治療,病人往往在數個月甚或數週內死亡。手術切除目前仍是唯一能治癒原 發性肝癌的治療方法。遺憾的是,當手術後腫瘤多發性復發,或病患因身體 狀況不適合手術,抑或是腫瘤發生遠處轉移時,尋找有效的化學治療藥物來 代替手術就顯得更為迫切。. 第一節【致癌的原因】 惡性腫瘤是一種基因變異的疾病,它的形成是多步驟的,並且可以發現幾 乎都與某些基因的改變有關(3)。基本上惡性腫瘤的發展過程約可分成下列四個 階段: (1)引發階段 (initiation):此階段肇因於基因上的變異 (2)促進階段 (promotion):乃細胞表現型上的改變 (3)惡性轉型階段(malignant coversion) (4) 腫瘤形成、生長及惡化(tumor formation, growth and progression)。構成惡性腫 瘤的癌細胞的特徵是癌細胞失去正常細胞的調控功能、失去細胞分化的能 1.
(11) 力,並且以不正常的比例快速增殖,可能侵犯周圍的正常組織或具轉移性--轉移至其他器官或組織。 一 般 而 言 , 致 癌 物 質 (carcinogen) 可 以 分 為 三 類 : 輻 射 能 (ionized radiation),病毒(virus)及化學物質(chemical compounds)。研究發現很多與癌症 有關的基因能與後天環境中各種因子產生交互作用,例如病毒基因影響肝細 胞基因組的正常功能即一重要促發癌變的機轉(1,4-5)。B 型或 C 型肝炎感染與肝 癌的形成機制有所不同,前者與 DNA 病毒基因組鑲嵌入肝細胞基因組有關, 後者則因細胞反覆發炎、壞死、增生、肝硬化形成等因素所致。B 型肝炎病毒 的感染,在原發性肝細胞癌的致癌原因中所扮演的角色相當重要。根據統計, 台灣目前可能有多達三百萬名 B 型肝炎帶原者,若年輕即感染病毒,則約有 1/3 的人會在 40 歲以後,慢慢轉變成肝硬化或肝癌。 除了 B 型或 C 型肝炎感染等病毒因素外,其他原因所導致的肝硬化(如長 時間或大量酒精攝取致肝硬化),外在環境的化學致癌物如 Aspergillus flavus 黃麴毒素(被污染的玉米、花生以及醃漬物)、香煙(含有四氨基聯苯)、含砷井 水、燒烤物及拜香(產生多環芳香烴)等等皆為引發肝癌的因子;最近國人更藉 由流行病學研究,報告長期嚼食檳榔(含肝癌致癌物 safrole)會罹患肝癌,其致 癌機轉有待探究。一般而言,B 型肝炎病人罹患肝癌比一般人高出 15-390 倍, 若加以暴露於這些化學致癌物,則會比一般人高出 40 至 60 倍罹患肝癌的機 率 (1-3)。 據估計,80%以上人類的癌症與環境中化學物質關係密切 (6)。西元 1775 2.
(12) 年英國的一位外科醫師 Percival Pott(7)首先提到環境中的化學物質與癌症的發 生可能有關。他發現煙囪的清潔工人有較高的比率得到鼻腔癌及陰囊皮膚 癌。在過去數十年來,已發現多種化學致癌物質存在煙囪之煤灰中,且可在 動物實驗中引發惡性腫瘤。有些化學物質可以本身就是致癌物,有些則是經 由細胞中某些特殊酵素的作用而轉變為致癌物。 如表(1)所示,許多化學物質都有致癌作用(2, 8),圖(1)所示即為三種常被廣 泛研究的化學物質的化學結構。人們可因為不同的環境因素:職業、飲食、 生活型態等等,接觸到這些化學物質。例如:油漆工人(苯 benzen),石棉瓦製 造業(石棉 asbestos),發霉的花生 (由黃麴霉菌 Asperqillas flavus 產生黃麴毒素 Aflatoxin B1) 或其它食物,如烤肉所產生的多環芳香胺類致癌物 (polycyclic arylamine)等。 在芳香胺類方面,已有研究顯示經由餵食芳香胺類化合物可能導致動物產 生癌症,但是將芳香胺類化合物直接注射動物的任何器官卻不會產生癌症(9)。 經研究後發現許多化學致癌物均需經過體內酵素代謝活化才會造成活性更強 的化學致癌物,再與細胞內的 DNA 結合形成 DNA 添加物(DNA adduct),最 後造成基因突變引發癌症 (10)。乙醯轉移酵素(N-acetyltransferase;NAT)就是參 與芳香胺類化合物代謝的一個代表酵素 (11)。而在多環芳香胺類方面,日本 Yamagiwa 與 Ichikawa(12)將柏油塗抹於兔子上導致皮膚癌而證實柏油中含有致 癌物質。後來經研究發現柏油中所含的多環芳香胺碳氫化合物 benzo(α)-pyrene 是一種致癌物質(13)。而在無機化合物方面,已有研究指出長 3.
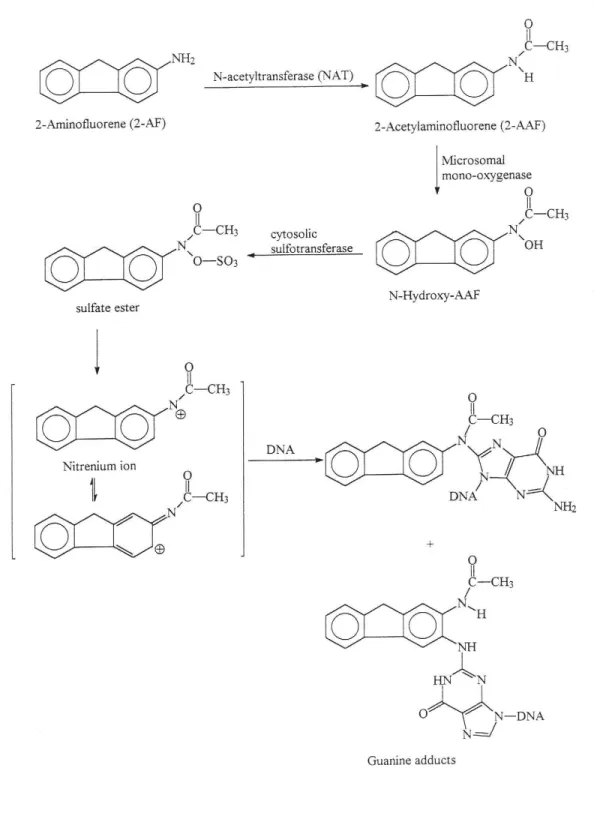
(13) 期攝食含有亞硝機化合物的醃製醬菜食物可能導致胃癌與食道癌(14)。而天然 性致癌物方面,如存在於發霉稻米、花生中的黃麴毒素(aflatoxins)已被證實易 引起動物及人類肝癌(15)。而 UV 紫外線方面,亦有研究指出長期暴露於 UV 紫外線下易得皮膚癌(16)。 因為 NAT 與芳香胺類化合物的致癌作用有相關連性已被證實,如果個體 暴露在含芳香胺類化合物的環境下,提高 NAT 的活性,則會增加致癌的機率。 有學者指出,改變 NAT 的活性與乳癌和膀胱癌的發生有相關聯,故參與致癌 性芳香胺類乙醯化的酵素近來甚受重視。若能發現影響此酵素活性的新藥物 或化合物,抑制 NAT 的活性,那就可以減少芳香胺類化合物的乙醯化,使化 學 致 癌 物 不 會 變 成 活 性 強 的 代 謝 產 物 , 進 而 減 少 終 極 致 癌 物 (ultimate carcinogen)的生成,減少細胞突變致癌的危險性,或許能預防或治療某些癌症。. 第二節【芳香胺類及化學致癌流程】 芳香胺類如 2-Aminofluorene (2-AF)是目前已知最強化學致癌物的其中一 種,存在於自然界和食物中,本身穩定且活性低,其結構式如圖(2): 芳香胺類須被生物轉換才會產生反應代謝物,亦即需先經由生物體內 NAT 代謝。據 Umemura(17)等人的研究發現,累積不同劑量與時間的 2-AAF,可以 對大鼠肝臟細胞產生 DNA 破壞,功能改變及可能發生惡性腫瘤的轉變。因此 當人類暴露於職業、環境或食物中的芳香胺類化合物 2-aminofluorene (2-AF) 後,該 2-AF 需先經 NAT 的乙醯化形成 2-Acetylaminofluorene (2-AAF),接著 4.
(14) 再 經 細 胞 內 Prostaglandin H synthetase 或 Cytochrome P450 代 謝 形 成 hydroxylamine. 及. hydroxamic. acid , 再 由. N-O-acetyltransferase ,. UDP-glucuronosyltransferase, Sulfotransferase 或 O-acetyltransferase 進行進一步 代謝,所形成之代謝化合物會自發性的分解產生 nitrenium ions,並且與 DNA 結合形成 DNA adducts (18-25),形成活性強的致癌物質(ultimate carcinogen),最 終造成目標器官癌化(26)。(如圖 3 所示)。而 2-AF 具有組織專一性,只會與特 定器官結合 (27)。. 第三節【乙醯轉移酵素(N-acetyltransferase,NAT)】 乙醯轉移酵素(N-acetyltransferase, NAT) 的存在因不同的物種、細胞而 異,並且與癌症的發生具有密切關係。NAT 已被證實存在兔子、天竺鼠、大 老鼠、小老鼠等齧齒類、豬及人類的各組織中,也存在於魚類、鳥類、靈長 類、兩棲類(青蛙),甚至白色念珠菌、沙門氏菌、大腸桿菌和胃幽門螺旋桿菌, 以及我們平常食用的蔬菜和水果中都有 (28-38)。 實驗顯示,若直接餵食小狗 2-AF,並不會引起任何惡性病變;但若直接 餵食 2-AAF 則容易引發肝癌,可見小狗血液中沒有 NAT 的存在,後來經研究 證實在小狗身上缺乏 NAT 基因 (39)。由此似乎間接證明 NAT 基因-NAT-AF-AAF 與肝癌之間有某種關係存在。 文獻報告指出,人類控制 NAT 的基因位於第八對染色體 8p21.3-23.1 上, 具有多變異性(polymorphism)(40),包括 NAT1 基因、NAT2 基因及假基因 NATP 5.
(15) Locus (41)。NAT1 及 NAT2 基因是屬於單一、無 intron 的 exon,具 870bp 的 open reading frames,可轉譯出 290 個胺基酸,共用 87%的核甘酸,產生約 55 個相 異的胺基酸,且分別決定出 NAT1 與 NAT2 兩種酵素 (42)。另外有學者在小老 鼠身上發現了第三種 NAT 基因為 NAT3(43-44)。NAT1 及 NAT2 基因的種類,截 至目前已發現 24 種不同的 NAT1 及 26 種不同的 NAT2 (45-46)基因型,且不同的 NAT 基因型在不同民族的發生率明顯不同(47)。NAT1 與 NAT2 兩種酵素都具 有 N-acetylation, O-acetylation 和 N,O-acetylation 作用,並且與某些致癌物的 活化(activation)及去毒化(detoxication)有關 (48)。依 NAT 對受質的催化速率,可 將 NAT 分為快的或慢的 NAT (49-53)。NAT 活性快的個體,暴露到 2-AF 較易得 大腸直腸癌 (54-56) 、NAT 活性慢的個體,暴露到 2-AF 較易得膀胱癌和食道癌 (57-63). 。. 在人體組織中 NAT 的分布並不均勻,NAT2 主要分布在肝臟,NAT1 則大 都分佈在肝臟和肝臟以外的組織中(64)。本實驗所用的 2-AF 是 NAT1 與 NAT2 的共同受質。 NAT 其分子量為 34 kDa,基本作用是將藥物及其他化合物乙醯化(64)。化 合物含有 sulf-hydry1、amino、hydroxy1 基,進入體內後須先經 NAT 的乙醯 化代謝後,利用 Acety1-Co A 上的乙醯基轉移至受質(例如 2-AF)的 NH2 基上 而形成 Acety1 amine (例如 2-AAF (N-acety1-2-aminofluorene))。然後經由體內 其 他 酵 素 , 如 細 胞 色 素 P450(Cytochrome P450) , 硫 基 轉 移 酵 素 (Sulfotransferase) , 和 甲 基 轉 移 酵 素 (Methyltransferase) , 黃 基 加 氧 酵 素 6.
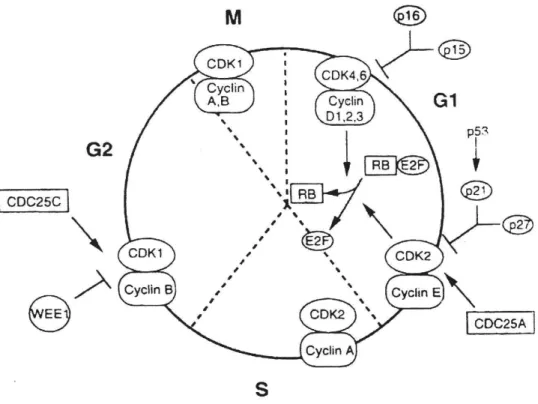
(16) (Oxygenase) 等 的 進 一 步 代 謝 , 才 會 成 為 活 性 的 終 極 致 癌 物 (ultimate carcinogen),然後與靶的器官細胞內的 DNA 的 Guanine 共價結合,形成核酸 衍生物(Nucleic acid adducts),最後造成標的器官細胞 DNA 的突變,而致癌化 (18-25). 。這種乙醯轉移的過程是芳香氨類化合物的必經代謝路徑的先前步驟。. 簡單地說,反應式如下所示: AcCoA + NAT. <=> CoA + Ac-NAT. Ac-NAT + Amine <=> Acetyl-amine +NAT. 因為 NAT 與芳香胺類化合物的致癌作用的相關性已被證實,如果個體暴 露在含芳香胺類化合物的環境下,提高 NAT 的活性,則會增加致癌的機會。 故參與致癌性胺類芳香胺類乙醯化的酵素近來甚受重視。若能發現影響此酵 素活性的新藥物或化合物,抑制 NAT 的活性,那就可以減少芳香胺類化合物 的乙醯化,使化學致癌物不會變成活性強的代謝產物,進而減少終極致癌物 (ultimate carcinogen)的生成,減少細胞突變致癌的危險性,或許能預防或治療 某些癌症。因此本實驗將探討 Paclitaxel 對肝癌細胞株的 NAT 的影響效果以 作為化學治療的基本依據。. 第四節【細胞週期】 細胞週期是指細胞經一連串規律的成長步驟而分裂為兩個子細胞的過 程 (65-68),如圖(4)。G0 期是細胞週期的停止期,若給予適當的刺激即可誘導 7.
(17) G0 期進入 G1 期。G1 期是產生新的子細胞所必須經過的生長發育階段。在這階 段中 RNAs 和蛋白質被合成,而此時期並無 DNA 的複製。S 期時,DNA 進行 複製,使 DNA 的含量增加一倍。G2 則為 DNA 合成後至有絲分裂所必經的階 段,在此時期完成時,細胞即擁有兩套完整的染色體。在 M 期,細胞進行細 胞核及細胞質分裂。由於細胞進入 S 期後一直到完成有絲分裂所需的時間相 當固定,因此 G1 期的長短可代表整個細胞的週期長短。又正常人體細胞 DNA 為雙套,但癌細胞的細胞週期與正常細胞不同,其 DNA 可能成單套或多套。 細胞分裂受相當複雜的機制所調控,若其中步驟發生失調就可能導致細胞 週期紊亂,使細胞失去正常的行為與生長方式而產生癌症 (67-69)。CDK(cyclin dependent kinase)與 cyclins 是推動細胞週期進展的物質,而 CDK 須與 cyclins 結合成複合體才具有功能,進而使細胞激活由 G1 進入 S 期或由 G2 進入 M 期, 它們的作用就是將磷酸根加到蛋白質上。其中 cyclin D 結合 CDK4 及 CDK6 作用在 G1 中期至晚期,cyclin E結合 CDK2 作用在 G1晚期,cyclin A 結合 CDK2 作用在 S 至 G2 期,cyclin A 以及 cyclin B 結合 CDK1 作用在 G2 至 M 期。有 些研究發現人類的癌症可能與 cyclins D1 基因的放大(amplification)和 cyclins D1 蛋白質的過度表現(over expression)有關(70)。本實驗將探討 Paclitaxel 對肝 癌細胞株細胞週期的影響,並進一步檢測與肝癌細胞株細胞內的 cyclins 的調 控關係。. 8.
(18) 第五節 【細胞凋亡(Apoptosis)】 細胞凋亡又稱為計劃性細胞死亡(programmed cell death)。它是極特殊的、 自然的細胞自殺過程,由多種遺傳基因導控細胞按步就班死亡的一種生命現 象。它會發生在各種不同的細胞,有四個重要外在特徵,分別為細胞膜分隔 (shrinkage)、染色體濃縮(condensation)、DNA 切段(fragmentation)以及凋亡小 體(apoptotic body)的產生。它的特性是細胞膜不會破裂,不會將細胞內的殘骸 流出細胞外,但有一特殊現象就是細胞膜內的 phosphatidylserine (PS)會轉位至 細胞膜外。而 Annexin V 是一種 Ca2+ dependent phospholipid-binding protein, 對 PS 具有高度親和力,因此我們使用接有螢光的 Annexin V 偵測細胞凋亡的 存在與否(71)。 當細胞 DNA 受損時會引發特定的 DNA 受損辨識蛋白及癌抑制基因的活 性,進而影響下游分子,導致細胞生長週期暫停在 G1 或 G2 以修復 DNA。若 DNA 受損過多則會傳送訊號破壞粒線體以致活化蛋白酵素 caspase,進而活化 核酸酵素引起染色體分解,造成細胞凋亡。而凋亡小體最後會被鄰近細胞或 巨噬細胞吞噬清除。所以細胞凋亡在動物發育上有許多重要的功用,例如形 體改變、去掉不需要構造、控制細胞數、去除不正常、失去功能或是有害的 細胞以及產生分化細胞等等。. 9.
(19) 第六節 【細胞壞死(necrosis)】 細胞壞死(necrosis)指物理性或外界因子導致細胞傷害時,會透過細胞內粒 線體的感應器執行死亡的現象。就形態上而言,細胞壞死時其形體瞬間瓦解; 而細胞凋亡則是細胞外形相當完整的在程式下循序漸進死亡。 當輻射線、化學致癌物或病毒造成細胞 DNA受損時,細胞核內最初的DNA 修復是在 DNA 斷裂處進行不同蛋白質的 ADP-ribose 聚合作用 (72)。poly (ADP-ribose) polymerase (PARP)是藉著水解 DNA,催化 poly(ADP-ribose)和一 些核蛋白的形成,而後再釋放出 nicotinamide(73)。而在細胞凋亡時 PARP 則會 被 Caspase-3 催化切割成 89 及 24 kDa 的片段 (72)。且已有研究指出 PARP 則會 被 Caspase-3 射線誘導形成的 G1 或 G2 arrest 中扮演相當重要角色(74),因此我 們利用可與 ADP-ribose 聚合物專一性結合的抗 poly (ADP-ribose)的單株抗體 (10H)進行免疫檢測(72)。. 第七節 【癌症的化學藥物治療】 癌症如果能早期發現大都可以手術切除而達到治癒的情況,如果太晚發 現,則必須併用化學藥物治療才能緩解,使末期腫瘤病人的生命得以延長。 例如乳癌、大腸癌的手術後輔助性化學藥物治療,使病患有機會長期存活。 又如急性白血病患者接受化學藥物治療,其平均生存期由過去的 2-3 個月延長 到 5 年以上。然而多數常見的癌症如肝癌、胰臟癌、肺癌等至今仍缺乏有效 的治療藥物,當手術後腫瘤復發或有轉移現象時,大都令臨床醫師束手無策。 10.
(20) 而且,不少抗腫瘤藥物在臨床使用過程中會產生抗藥性。因此,新型抗腫瘤 藥物的研發,隨著科學技術的快速發展,細胞生物學與分子生物學的進步而 不斷出現,例如單株抗體的導向治療、腫瘤細胞的誘導分化及治療,抗腫瘤 侵襲及抗腫瘤轉移藥的研究、免疫治療、基因治療等(75-87)。. 第八節 【關於本實驗癌症治療藥物-紫杉醇(Paclitaxel,Taxol)】 紫杉醇是由太平洋紫杉(Pacific yew 學名為 Taxus brevifolis)之樹皮抽取出 來的(88)。紫杉醇在 1960 年代開始有許多藥物學與藥劑學方面之研究。至於原 植物太平洋紫杉具有醫療作用早在幾世紀以前就已知曉。公元前年 Julius Caesar 就記載在他的書中,有人飲用紫杉茶而自殺;在早期美國原住民就利 用紫杉皮之抽出液來作為抗菌劑(disinfectant),流產劑(abortifacient)以及皮膚 癌之治療(89)。 紫杉醇之化學構造相當複雜(88),Nicolaou(90)研究小組已完成了化學合成的 方法,在自然雜誌發表(Nature 367, 630-634, 1994)。共使用 27 個有機化學反 應,並分離出 19 種中間產物,最後終於合成了 Paclitaxel。 在 1978 年,亞伯愛恩斯坦醫學院(Bronz, N.Y.)的一位教授 Dr. Susan B. Horwitiz 及她的研究生 Peter B. Schiff 發現 Paclitaxel 殺死癌細胞之作用原理與 其他的抗癌藥物不同(91-93)。 在正常細胞中,微管(microtubule)是一種具有彃性的特殊裝置,它為了細 胞的順利分裂(mitosis),可以收緊也可以鬆散,它便是構成了有絲分裂過程中 11.
(21) 的紡綞體(mitotic spindle)。它之主要功能在使染色體(chromosome),在細胞分 裂時適時地分開。Paclitaxel 則是不斷促進微管次體(tubulin)聚合成微管並與之 結合,這微管與 paclitaxel 結合之後變為非常穩定,甚至僵硬,使其僵化而失 去分裂之功能,不讓正常的解離進行,破壞了動態平衡,造成有絲分裂終止 於 G2 與 M 期,染色體就無法分開,細胞就不會分裂而走向死亡(91-94)。G2 與 M 期對於放射線敏感度最高,因此 paclitaxel 也被研究用來做為促放射敏感劑 (radiosensitizer)(95-96)。一般來說,癌細胞的分裂較正常細胞來得快速,paclitaxel 之作用目標是正在分裂的細胞。因此,paclitaxel 會優先攻擊正在分裂之癌細 胞。Paclitaxel 主要在肝臟代謝,肝內代謝與細胞色素 p450 有關(97)。 Paclitaxel 第一個被核准用來當臨床惡性腫瘤化學治療的適應症是經第一 線或其它化學治療失敗的轉移性卵巢癌 (98-100),一年半後又增加了另一個適應 症—經含 anthracycline 之化學療法失敗,或經輔助性化學療法後六個月內再發 之轉移性乳癌(101-103)。Paclitaxel 現在被認為是對乳癌與卵巢癌最有用的藥物, Paclitaxel 用於其它惡性腫瘤的治療目前也正在積極研究中(104-106)。截至目前為 止的研究顯示,在非小細胞及小細胞肺癌,頭頸部鱗狀上皮癌,膀胱癌,生 殖細胞瘤等均報告有相當療效(104-106),但在腸胃道癌如大腸直腸癌,胰臟癌方 面則無明顯治療效果,它在肝癌的治療效果則尚無定論 (107-108)。因此,本實驗 希望藉由 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)的 NAT 基因表現及細胞 週期的影響,來探討未來在臨床病患使用藥物治療的立基。. 12.
(22) 第二章. 材. 料. 一、 實驗儀器 1. 高 效 液 相 層 析 儀 (HPLC) (Beckman, Programmable Solvent Module 126; Diode Array Detector Module 168) 利用不同溶液為移動相,通過靜態相的管柱,藉著在移動相中之 檢品各成分對於靜態相管柱結合時間的不同,進而將其分離之操作。 實驗先由已知濃度的標準品溶液,算出峰線面積,再求得檢體峰線面 積,兩者比較後,進而算出檢體的含量或濃度,再分析其總量。. 測定 NAT 活性的條件: (1) C18 reversed-phase column (spherisorb 4.6x250nm) (2) Solvent system: 20 mM KH2PO4 (PH4.5)/CH3CN (53:47) (3) Wavelength: 280 nm (4) Retention time: AAF: 6.5 min, 2-AF: 9.0 min. 2. 流式細胞計數儀:Flow cytometry (美商必帝 FACScan) 流式細胞計數儀是一具有測定及計算單一細胞的細胞計數儀器,它可偵測 生物顆粒的物理特性,例如細胞大小、形狀及複雜性;我們也可利用細胞的 成分或功能使細胞與螢光染料反應後,再將此細胞懸浮液送入計數儀中分. 13.
(23) 析。而流式細胞計數儀依螢光波長的不同可偵測紅、藍、綠三種不同的螢光。 簡而言之,此計數儀乃是利用液流系統所製造出的穩定液流讓細胞以一定的 速度及精準的軌跡通過預先校正對準好的雷射光束後,再由電腦做定量分析。 而我們利用能嵌入雙鍵 DNA 及 RNA 的鹼基對中與氫鍵結合之 Propidium Iodide (PI)與癌細胞反應,再以流氏細胞儀分析其細胞週期,來決定細胞在週 期中的比例。 以流式細胞技術研究凋亡細胞,基本上是分析細胞形態上及生化上的特 徵。在流式分析術中,前方散射光(FSC)代表細胞大小,側方散射光(SSC)表示 細胞的顆粒性。凋亡細胞應該 FSC 變小,先增加後減少,使用方法必需檢測 活細胞才有意義。活細胞或胞膜完整的細胞可將一些染料排斥在外,例如 Propidium Iodide (PI),由於早、中期凋亡細胞仍保留完整的細胞膜,故對 PI 拒染,可利用此方法分辨凋亡細胞和壞死細胞。. 3. 倒立式相位差顯微鏡:Nikon Ellipse TE 300 4. 離心機:Model 3200 Eppendoff / Brinkman 5. 製冰機:King-Seeley Thermos Co. USA 6. 紫外線燈:Spectvoline model ENF-24℃. 二、 實驗材料 1.購自 Sigma Chemical Co. (St. Louis, MO)的材料 14.
(24) Leupeptin Tris-HC1 Trischloroacetic acid Ethylenediaminetetraacetic acid (EDTA) P-aminobenzolic acid (PABA) N-acetyl-p-aminobenzoic acid (N-ac-PABA) Formic acid (Ammonium salt) = Ammonium formate 1, 4-Dithiothreitol (DTT) Bovine serum albumin (BSA) Phenylmethylsulfonyl fluoride (PMSF) Adenosine 5-triphosphate (ATP) Dimethyl sulfoxide (DMSO) Trypsin Trypan blue Ribonuclease A (RNAse) Acetylcarnitine Carnitine acetyltransferase Tergitol NP-40 2.購自美國 K and K laboratories (Plainview, NY) 公司 2-Aminofluorene (2-AF) 15.
(25) 2-acetylaminofluorene (2-AAF) 3.購自美國 P-L Biochemicals Inc. (Milwaukee, WI)公司者:Acety1 Co-A 4.購自美國 Gibco laboratories (Grand Island, NY) 公司者 RPMI 1640 cultrue medium Fetal bovine serum Hanks balanced salts Antibiotics (Kanamycin, Penicillin, Streptomycin) Glutamine 5.購自瑞士 Fluka 公司者 Propidium Iodide (PI) 6.購自西德 Merck 公司者 Acetonitrile Acetic acid Ethanol Acetone Triethylamine Iodoacetamide Ethyl acetate 7.購自西德 FERAX (GMBH, Berlin)公司者 Ethylene glycol monoethylether 16.
(26) Trisodium citrate 2-hydrate 8.Working Lysing Buffer Solution (冰浴操作) Working solution [PH7.5, 4℃, 20mM Tris-HCL, 1mM Dithioth-reitol (DTT), 1mM Ethylene diaminete traacetate acid (EDTA)] 20mL, 50mM phenyl methyl sulfonyl fluoride (PMSF) 20μL 及 10μM leupetin 20μ L,後兩者以維持酵素活性。 9.Acetyl-CoA recycling mixture 50mM Tris-HCL (PH 7.5), 0.2mM EDTA, 2mM DTT, 15mM acetylcarnitine, 2U/mL carnitine acetyl transferase, 2-AF(特定濃度)。. 17.
(27) 第三章. 方. 法. 本研究將實驗分成兩大部分,第一部分是檢測 Paclitaxel 對人類肝癌細胞 株(HA 22 T/VGH)生長的影響,包括檢測此藥物對人類肝癌細胞株的細胞毒性. (cytotoxicity)、細胞週期及 cyclins 的影響。第二部分包括 1.在整個活細胞方 面,我們檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) NAT 活性,基因序 列、對(HA 22 T/VGH)細胞內 NAT 蛋白表現的影響。2.在細胞質液方面:我們 檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)細胞質液中 NAT 活性的影 響。而本實驗所用藥物 Paclitaxel 以 100% DMSO 當溶劑,製備了 5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM 五種同濃度。. 一、. 檢測 Paclitaxel 對 人類肝癌細胞株生長方面的影響. 1.檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)細胞 毒性(cytotoxicity). 的影響 準備三個 6 well 培養皿,將人類肝癌細胞株(HA 22 T/VGH)細胞植 入培養,每一 well 培養 1×106 細胞,並加入 2 ml 培養基。第一個 well 加入 20μl DMSO 作為控制組,第二至第六個 well 分別加入配置好之 五種不同濃度 Paclitaxel (5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM) 20μ l,再置於細胞培養箱中,保持 37℃, 5% CO2 的條件下,分別反應 6, 12,24,48 小時後,以 PBS 清洗後,再加入 Trypsin 少許以打下細胞, 經由離心機離心除去培養基。將細胞收集在 15 ml 離心管並離心 1500 18.
(28) rpm,5 分鐘,4℃後,倒掉上清液並輕拍管壁再加入 5ml PBS,使細 胞懸浮在 PBS 中,再離心 1500 rpm,5 分鐘,4℃後,倒掉上清液留 下 100-200μl PBS 並輕拍管壁,使細胞懸浮在 PBS 中,加入 500 ml 4 μg / ml 之 Propidium iodine (PI),再以流式細胞儀(Flow cytometry)分 析存活細胞數。. 1. 檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)細胞週期的影響 取 1×106 細胞放入培養皿中,再移入 37℃ 5% CO2 培養箱,加入 不同濃度藥物,經不同時間培養後收集離心,加入冰浴的 70%酒精後, 放置隔夜,以 PBS 清洗後,各加入染色溶液( 4 μg/mL 的 PI + 0.1 mg/ml RNase + 1% Triton-X 100 ),1 ml 並置於室溫下,再放入流式細 胞計數儀(Flow cytometry;FACS)分析 G0/G1、S、G2 & M 期 DNA 含 量。. 2. 檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)細胞內 cyclins 的影 響 取 1×106 細胞放入培養皿中,再移入 37℃ 5% CO2 培養箱,加入不 同濃度 Paclitaxel,經 24 小時培養後收集離心,加入 1% 福馬林 100 µL 並置於冰上 5 分鐘,加入 99%甲醇(Methanol) 100 µL 並置於冰上 30 分 鐘,以含 0.1% BSA 的 PBS 清洗 2 次後,再加入 0.1% X- 100 及 0.1% 19.
(29) sodium citrate 100 µL 並置於冰上 45 分鐘,再以含 0.1% BSA 的 PBS 清 洗後加入對抗 Cyclin 的單株抗體 50 µL 並於室溫下反應 2.5 小時,再 以含有 0.1% BSA 的 PBS 清洗後加入接有螢光的二次抗體 50 µL 並於 室溫下避光反應 30 分鐘,最後再以 0.1% BSA 的 PBS 清洗後放入 FACS 分析。而抗體是以含有 0.1% BAS 及 0.1% sodium azide 的 PBS 稀釋。. 二、. 以螢光顯微鏡檢測 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH)細. 胞內 poly (ADP-ribose) polymerase 的影響 當 細 胞 DNA 受 損 時 , 細 胞 會 產 生 poly (ADP-ribose) polymerase(PARP)來作為修補之用。準備二個 6 well 培養皿,將人類肝 癌細胞株(HA 22 T/VGH)細胞植入培養,每一 well 培養 1×106 細胞,並 加入 2 ml 培養基。其中一個培養皿的第一個 well 加入 20μl DMSO 作 為控制組,第二至第六個 well 分別加入配置好之五種不同濃度 Paclitaxel (5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM) 20μl,再置於細胞培 養箱中,保持 37℃,5% CO2 的條件下,反應 24 小時,另一培養皿的 第一個 well 作為控制組,其他則加入 0.8 mM Paclitaxel 分別反應 6,12, 24 小時後,以 PBS 清洗後,再加入 Trypsin 少許以打下細胞,經由離 心機離心除去培養基。將細胞收集在 15 ml 離心管並離心 1500 rpm,5 分鐘,4℃後,倒掉上清液並輕拍管壁再加入 5ml PBS,使細胞懸浮在 20.
(30) PBS 中,再離心 1500 rpm,5 分鐘,4℃後,倒掉上清液留下 100-200 μl PBS 並輕拍管壁,使細胞懸浮在 PBS 中,再將細胞移入 96 well 培 養皿中,離心 1500 rpm,4 分鐘,4℃後,倒掉上清液再加入 100μl 之 3.7% formaldehyde 反應 15 分鐘,以 PBS 洗細胞, 離心 1500 rpm, 4 分鐘,4℃後,倒掉上清液再加入 100μl 之 0.2% NP-40 (以 PBS 稀釋) 反應 15 分鐘,以 PBS 洗細胞, 離心 1500 rpm,4 分鐘,4℃,倒掉上 清液再加入含 5%脫脂牛奶之 PBS 並反應 1 小時,以 PBS 洗細胞, 離 心 1500 rpm, 4 分鐘, 4℃後,倒掉上清液再加入含 1% 抗 Poly ADP-ribose 的單株抗體(monoclonal antibody to poly ADP-ribose)之 5% 脫脂牛奶 PBS 溶液,並在 4℃下反應隔夜,以 PBS 洗細胞, 離心 1500 rpm, 4 分 鐘 , 4℃,倒掉上清液再加入 50μ l 之 1% 二次抗體 (FITC-conjugated goat anti-mouse IgG antibody),避光反應 30 分鐘後, 以 PBS 洗細胞並以螢光顯微鏡觀察並照相。. 三、. 檢測 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH)的 NAT 活性的. 影響 1. 檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)的 NAT 將 2-AF 乙醯 化成 2-AAF 的影響。 將培養的人類肝癌細胞株(HA 22 T/VGH),取 5×105 cells / mL 放入 24 well plate (含 1 mL RPMI 1640 培養基,1% glutamine,10%胎牛血清) 21.
(31) 中,再加入 arylamin 受質(2-AF)及分別有無加入不同濃度的 Paclitaxel 試劑(5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM),一起在 37℃,95%空氣, 5%二氧化碳,經過不同的時間(分別為 6,12,24,48 小時)反應終結, 將細胞及培養基移取離心。上清液立即以 ethylacetate / methanol (95:5) 萃取,此溶液冷凍揮發乾燥後,再溶於 50μL methanol 中混合均勻, 取 20μL 打入高效液相層析儀,再經由 HPLC 分析乙醯化受質(2-AAF) 和未乙醯化受質(2-AF)的量。. 2. 檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)細胞內 NAT 基因的影 響 (1) 反轉錄酵素(Reverse Transcriptase)反應 取 5×106 細胞置於培養皿中,再移入 37℃ 5% CO2 培養箱,加 入不同濃度藥物後,經 24 小時培養後將細胞收集離心,以 RNA kit 抽出細胞全部 RNA 並於 260 nm 下測 OD 值(假設 OD 值=1 時, RNA=40 ng/μL),再每管各取 1.5μg 的 RNase 並加入 RNA free water 使體積達 11.5μL,再各加入 1μL (0.5μg/μL) oligo (dT)且 置於 70℃反應 10 分鐘後快速置於冰上且 spin down,再各加入 7.5 μl 的反應溶液(2μL 100 mM DDT + 1μL dNTP + 0.5μL RT+ 4μ L 5X buffer)且靜置 42℃下 1 小時。 (2) 聚合酵素連鎖反應(Polymerase Chain Reaction) 22.
(32) 接著取 1 μL cDNA 加入 24 μL 反應溶液(19.05 μL dd H2O + 2.5 μL 10X buffer + 0.75μL MgCl2 + 0.5μL dNTP + 0.5μL Primer 1 + 0.5μL Primer 2 + 0.2μL Taq DNA polymerase),經混合 及 spin down 後放入 94℃ 3 分,再進行 35 個循環期:94℃ 45 秒, 55℃ 30 秒,72℃ 1 分 30 秒,結束後放置於 72℃ 10 分鐘後再置 於 4℃,加入 loading DNA dye 5μL 且混合及 spin down 後,跑 1.5% agarose 電泳膠,以 ethedium bromide 染色後照相。所使用的 Primer 如(表 2)。 表 2. Primers used for RT-PCR primer. Sequence: (5’ –3’). Size (bp). Act-b1. 5’-GCTCGTCGTCGACAACGGCTC-3’. 21. Act-b2. 5’-CAAACATGATCTGGGTCATCTTCTC-3’. 25. B-MDIEA-NAT1. 5’- CACCCGGATCCCGGGATCATGGACATTGAAGC-3’. 31. VPKHGD-X-NAT1. 5’-GGTCCTCGAGTCAATCACCATGTTTGGGCAC-3’. 31. FP1-NAT2. 5’-CTAGTTCCTGGTTGCTGGCC-3’. 20. RP1-NAT2. 5’-TAACGTGAGGGTAGAGAGGA-3’. 20. 23.
(33) 3. 檢測 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)細胞內 NAT 蛋白表現 的影響 取 1×106 細胞放入培養皿中,再移入 37℃ 5% CO2 培養箱,加入 不同濃度 Paclitaxel,經 24 小時培養後收集離心,以福馬林及甲醇固 定細胞,再以 Triton X-100 及 sodium citrate 增加細胞膜滲透性,再加 入本實驗室自製的對抗人類 NAT 多株抗體(polyclonal anti-human NAT antibody),再加入接有螢光的二次抗體,最後放入 FACS 分析。. 4. 檢測人類肝癌細胞株(HA 22 T/VGH)在加入不同濃度 Paclitaxel 後暴露 於細胞外的 NAT 活性 取 1×105 細胞置於培養皿中,再移入 37℃ 5% CO2 培養箱。反應混 合液組成總共體積 90μL:包括取 50μL 的(HA 22 T/VGH) 胞質液, 加入 20μL 的 Acetyl-coenzyme A recycling mixture [(50 mM Tris-HCL, PH7.5),0.2 mM EDTA,2 mM DTT,15 mM acetyl carnitine,2 U/mL carnitine acetyltransferase],以一定量的 AF 或 PABA 當受質,最後加了 20μL 的 Acetyl-Co A 及 10μL 不同濃度的 Paclitaxel (5µM, 50µM, 500µM, 2.5mM, 5mM)。控制組不加 Acetyl-Co A,而加入蒸餾水。培育 於 37℃,10 分鐘,再加入 100μL acetonitrile 終止 2-AF 或 PABA 反應。 實驗組及控制組均作 3 次,再經由 HPLC 分析,得出 Paclitaxel 對人類 肝癌細胞胞質液中不同濃度的 Paclitaxel 對 NAT 活性測定。加入不同濃 24.
(34) 度 Paclitaxel 後,經 24 小時培養,各取 60μL 上清液並各加入 20μL RCM 及 Acetyl CoA,混合後置於 37℃恆溫水槽反應 10 分鐘,再加入 50μL Acetonitrile,混合並離心後吸取 20μL 上清液打入 HPLC 分析。 為維持 NAT 活性,實驗必需在冰上操作。. 25.
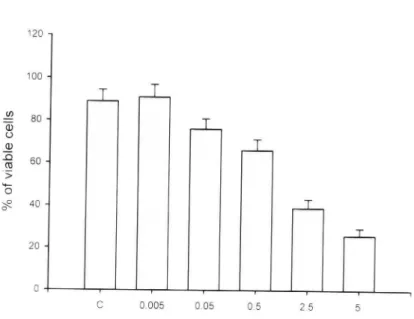
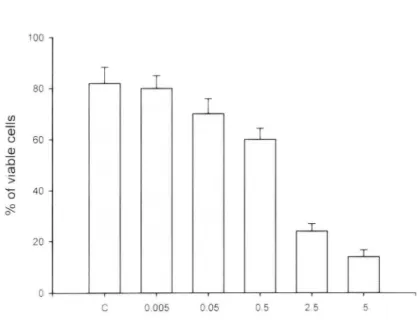
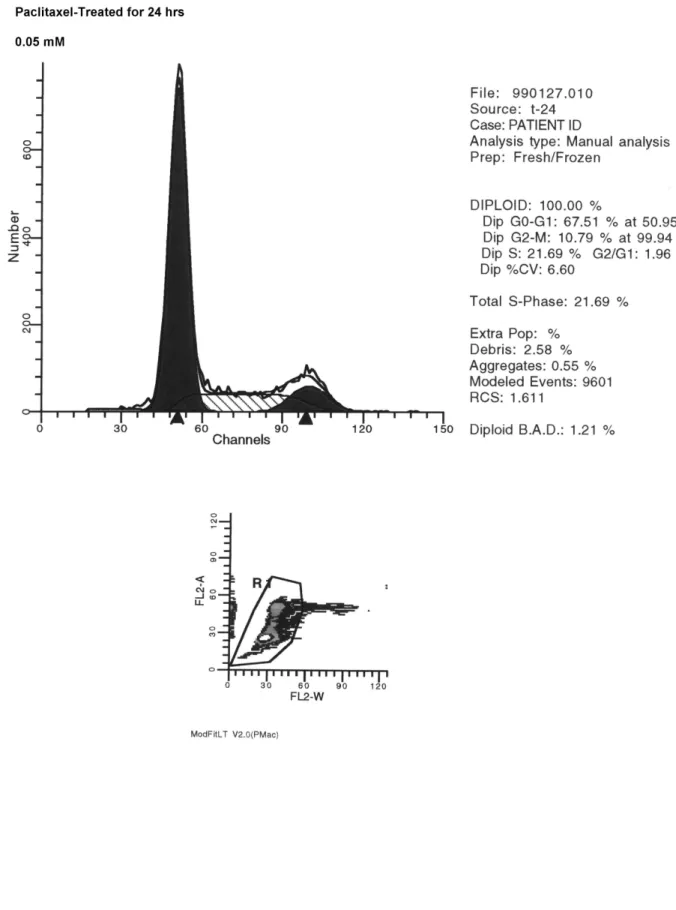
(35) 第四章. 結. 果. 一、 不同濃度的 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH)細胞毒性. (cytotoxicity)的影響 從圖(5.1-5.5)的實驗結果發現,例如在 24 小時的培養時間,所加 Paclitaxel 隨著濃度的上升細胞存活率由 80.00%, 66.66%, 60.00%, 46.66%,36.66%,23.33% 到 10.00%。發現 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH) 的毒殺效應,在相同的培養時間,濃度愈高時,毒殺效 應愈佳。並且在同樣的濃度時,培養時間愈久,細胞存活率就愈低。例 如同樣是加 5 mM/mL 的 Paclitaxel,在 6,12,24,48 小時不同的培養 時間,(HA 22 T/VGH)細胞其存活率依次為 80.76%,60.71%,46.66%, 32.35%。所以 Paclitaxel 對人類肝癌細胞株具有毒殺作用,並且和濃度及 培養時間有正相關,濃度愈高,作用時間愈久,毒殺作用就愈明顯。. 二、 不同濃度的 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH) DNA 的破 壞效應 在細胞培養中,分別有無加入不同濃度的 Paclitaxel 試劑(5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM),結果發現 DNA 有裂解的現象,並且有固定 base-pair DNA 出現(APOPTOSIS 的特徵,如圖 6 )。所以 Paclitaxel 造成人 類肝癌細胞株(HA 22 T/VGH)細胞死亡,是經由計畫性的細胞死亡。. 26.
(36) 三、 不同濃度的 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH) 細胞週期 的影響 在細胞培養中,分別有無加入不同濃度的 Paclitaxel 試劑,利用流式細 胞計數儀檢測 G0/G1,S,G2/M phase 等細胞週期 DNA 量的變化,並利用 電腦分析以測知 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 細胞週期的 影響。所加 Paclitaxel 濃度依序為 control,5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM,其 total S-phase 依序為 8.82%,21.65%,21.69%,33.57%,35.28%, 32.40%,同樣變化也存在 G0/G1 和 G2/M phase。在 G2/M 期則為 7.65%, 8.92%,10.79%,14.97%,16.06% 及 21.44%,由以上結果發現在 24 小時 Paclitaxel 可使(HA 22 T/VGH)細胞週期停止在 G2/M 期及 S 期,隨著藥物 濃度增加,停止在 G2/M 細胞漸增。而且隨著作用增長,(HA 22 T/VGH) 細胞週期停止在 G2/M 期更加明顯。(如圖 7-8)。. 四、 檢測 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH) 細胞週期素的影 響 由細胞週期結果已經發現 Paclitaxel 可使(HA 22 T/VGH) 細胞的細胞 週期停止於 G2/M 期,而在 24 小時 Paclitaxel 作用,(HA 22 T/VGH)細胞 週期發現 S 期及 G2/M 期都增加。在偵測(HA 22 T/VGH)細胞內 cyclins 及 CDKs 的結果,發現調控 G2/M 期的 cyclin B 及 CDK1 隨著藥物濃度增加很 明顯受到抑制,而對於調節細胞週期,由 S 期進入 G2/M 期的 cyclin A 和 27.
(37) CDK2 表現則是受到很明顯的誘導(over expression),對於控制 G1 前期的 cyclins D1 則是受到抑制,控制 G1 後的 cyclin E 則是受到誘導而表現增加 (over expression)。由以上 Paclitaxel 對(HA 22 T/VGH)細胞內 cyclins 及 CDK2 的影響,結果發現是與細胞週期結果相符合的,因此 Paclitaxel 確實 是造成(HA 22 T/VGH)細胞的細胞週期停止於 G2/M 期。(如圖 9-10). 五、 不同濃度的 Paclitaxel 對人類肝癌細胞株 (HA 22 T/VGH) 中 NAT 的 活性效應 1. 在活細胞內 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH)細胞中 N-acetyltransferase (NAT)酵素活性的影響的實驗,在加入不同濃度的 Paclitaxel,依序為 5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM,經作用 24 小時,用 HPLC 檢 測,發現 NAT 活性分別為 4.60%,10.51%,16.42%,33.05%,45.74%, 67.18%。顯示濃度愈高,NAT 活性被誘導愈明顯,(如圖 11)。. 2. 在細胞質內 將培養的人類肝癌細胞株(HA 22 T/VGH)細胞胞質液,利用 2-AF 和 PABA 作受質來決定 Acetyl-Co A 依賴的乙醯轉移酵素乙醯化 2-AF 和 PABA 的量。結果顯示 Paclitaxel 濃度愈高,則人類肝癌細胞株(HA 22 T/VGH) 胞質液中 NAT 活性抑制愈強。譬如在加入不同濃度的 28.
(38) Paclitaxel,依序為 5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM,經作用 24 小 時,用 HPLC 檢測,發現 NAT 活性分別為 7.80%,13.77%,20.40%, 33.67%,51.08%,67.67%,90.05%。. 六、 利用 RT-PCR (Reverse Transcriptase Polymerase Chain Reaction) 方法來檢測 Paclitaxel 是否有影響人類肝癌細胞株(HA 22 T/VGH) 細胞 NAT1 基因(NAT mRNA)的表現 將 5×106 cells,分別有無加入各種不同濃度(5 µM, 50 µM, 500 µM, 2.5 mM, 5 mM)的 Paclitaxel,然後培養 24 小時,經由離心而收集細胞,再利 用 Qiagen RNeasy Mini Kit (Qiagen, inc, Valencia, CA, USA),收集 total RNA,然後用 RT-PCR 的方法,最後跑電泳、照像。觀察發現隨著 Paclitaxel 濃度增加,而(HA 22 T/VGH) 細胞的 NAT 1 基因受到很明顯抑制,(如圖 12)。. 29.
(39) 第五章. 討 論. Paclitaxel (Taxol)是由太平洋杉樹皮或其針葉上提煉出來的一種抗癌藥 (88). ,本實驗藉由探討 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 的影響,. 希望能多了解一些有關 Paclitaxel 抗癌的可能機轉,此實驗結果顯示,Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 有毒殺作用(如圖 5.1-5.5) 。從不同濃度的 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 的細胞毒性效應實驗發現,濃度 愈高,細胞的毒殺效果就愈強。研究顯示,Paclitaxel 與一般 Microtubulin 抑 制劑不同的是,它並不像 vinca alkaloid,podophyllotoxin,colchicine 一樣抑制 tubulin 的形成,反而是促使它形成而使其穩定的無法再分開,以致於細胞將 被迫停留在 G2M 期而無法通過分裂程序(91-93)。根據 Lin HL 等人報告(105),在 胃癌的研究中發現,Paclitaxel 引發的細胞計劃性死亡(apoptosis)與劑量有關, 且可能由 p53 訊息傳導(Signaling. pathway)所誘發。在低劑量時,其所. 引發的計劃性死亡並無 G2/M 期停滯的現象。本實驗發現在(HA 22 T/VGH)肝 癌細胞中,當 Paclitaxel 濃度小於 0.5mM 時,細胞生長並無明顯受到改變。而 在大於 0.5mM 時逐漸顯現其抑制細胞生長的效果,且隨作用時間增長,而更 趨明顯。(HA 22 T/VGH 細胞停滯在 G2/M 期濃度最高)。本研究以人類肝癌 細胞株(HA 22 T/VGH)細胞經不同濃度的 Paclitaxel 處理 24 小時後,再經由流 式細胞計數儀(FACS)分析結果顯示,Paclitaxel 會造成人類肝癌細胞株(HA 22 T/VGH)細胞週期的變化,使其停止在 G2/M 期。其次由人類肝癌細胞株(HA 22 T/VGH) 細胞經不同濃度 Paclitaxel 以螢光顯微鏡檢測 Paclitaxel 對人類肝 30.
(40) 癌細胞株(HA 22 T/VGH)細胞內 poly (ADP-ribose) polymerase 的影響,發 現,DNA 有出現裂解現象,所以發現 Paclitaxel 造成人類肝癌細胞株(HA 22 T/VGH) 細胞的死亡不是經由細胞壞死,而是經由計劃性的細胞死亡。 在動物的細胞週期中主要的兩個控制點(check point),是在 G1 期和 G2 期 將結束時。從 G1 進入 S 期時,細胞需要激發酵素(kinase)參與作用,如:G1/S 期激發酵素的激活以及 M 期激發酵素的抑制。在哺乳動物中這些酵素的產生 是由 CDK 基因來控制。在動物的基因體中至少有 10 種的 CDK 基因。正常細 胞週期中,G1/S 的過渡期間(transition)是由 CDK2 與 cyclin E 複合體參與作用, 而 S 期的進行由 CDK2 與 cyclin A 參與,在 G0 進入 G1 時則由 CDK2,CDK4 和 CDK5 與 cyclin D 結合形成激發酵素(111)。研究指出 cyclins D1 基因的放大 (amplification)和 cyclins D1 蛋白質的過度表現(over expression)的現象在肝癌 細胞中存在 (112-113)。根據 Joo 等人報告(114),cyclins D1 的過度表現可能是肝癌 病變的早期現象,可能與肝細胞分化有關,但和肝癌腫瘤細胞的分化無關。 而根據 Jung 等人的研究(115),在人類肝癌腫瘤組織中,發現在 cyclins D1 的不 表現時,cyclins E 的 reciprocal deregulation 可能造成 G1 期的進行而導致肝癌。 Paclitaxel 對肝癌細胞 cyclin 和 CDK 的作用仍未定論(112-116),本實驗發現當藥 物濃度大於 2.5mM 時,可以抑制 cyclin B, D 和 CDK1 的濃度,對 cyclin A、 E 和 CDK2 相反地增加。對 cyclin D3 則無明顯作用。本實驗在不同濃度的 Paclitaxel 作用下,cyclin B, D 受到明顯抑制,cyclin E 相對地增加。對 cyclin A 和 cyclin D3 作用則不明顯。另以 CDK 之研究來看,CDK2 比例隨藥物濃度 31.
(41) 增加而上升,CDK1 則明顯下降 (如圖 9-10) 。此外,研究指出(116) cyclin-cdk 複合物受兩套抑制機制調控,一是具專一性抑制 cyclin D-cdk4/6 的 P16INK4A 、 P15INK4B 、 P18INK4C 、 P19INK4D , 二 是 可 抑 制 許 多 種 cyclin-cdk 複 合 物 的 P21WAF/cip1、P27Wip1、P59Wip2。這些更深入的調控機制的異常是否與人類肝癌 的產生有關,仍待進一步的研究。 芳香胺類( Aromatic amine ) 的致癌作用早被報導且已證實,發現若以之餵 食實驗動物會導致癌症的發生,但若直接打到體內的標的器官,並沒有造成 標的器官的癌化。後來才了解,芳香胺類的活化作用,經由 NAT 及 cytochrome P450 酵素的共同作用,最後再與 DNA 結合,形成 DNA 的添加物,最後造成 標的器官的癌化(9-13)。芳香胺類代謝主要著重在肝臟。另有報導如果增加 NAT 的活性,則當個體暴露到芳香胺類致癌物,會增加芳香胺類突變效應的易感 性。有學者指出,改變 NAT 酵素的活性,當個體暴露到相關的芳香胺類致癌 物與引發肝癌和膀胱癌有相關連。本實驗探討不同濃度 Paclitaxel 對人類肝癌 細胞株(HA 22 T/VGH) 細胞中 NAT 的活性及人類肝癌細胞株(HA 22 T/VGH) 細胞胞質液中 NAT 的活性的影響,發現加不同濃度的 Paclitaxel 經 6,12,18, 24 小時作用,可增加(HA 22 T/VGH) NAT 的活性,且濃度愈高,作用時間愈 久,NAT 活性增加就愈明顯。另外人類肝癌細胞株((HA 22 T/VGH)細胞胞質 液的 NAT 活性測試中,也可見到濃度愈高,NAT 活性降低的就愈明顯。 有文獻報導肝癌和 NAT1 基因表現有關,所以我們進一步探討 Paclitaxel 能否影響人類肝癌細胞株(HA 22 T/VGH) 細胞中 NAT 1 基因的表現,因此利用 32.
(42) RT- PCR 的技術進一步探討不同濃度 Paclitaxel 是否會影響人類肝癌細胞株 (HA 22 T/VGH) 細胞 NAT1 基因的表現。在 RT-PCR 的實驗中,經由最後的照 相結果觀察,發現不同濃度的 Paclitaxel 加在人類肝癌細胞株(HA 22 T/VGH) 細胞,(HA 22 T/VGH) 細胞的 NAT 1 基因的表現受到抑制。因為 Paclitaxel 可 以降低人類肝癌細胞株(HA 22 T/VGH) 細胞胞質液的 NAT 活性,並且改變人 類肝癌細胞株(HA 22 T/VGH) 細胞 NAT1 基因的表現,所以 Paclitaxel 可以降 低人類肝癌細胞株(HA 22 T/VGH) 細胞 NAT 活性並非只是透過細胞毒性來降 低 NAT 的活性,而是確實能降低人類肝癌細胞株(HA 22 T/VGH) 的 NAT 活 性。 本實驗藉由探討 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 的影響,希 望能多了解一些有關 Paclitaxel 抗癌的可能機轉,此實驗結果顯示,Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 有毒殺作用,不是透過細胞壞死而是計劃 性的細胞死亡,而且 Paclitaxel 也會使(HA 22 T/VGH)細胞的細胞週期停止在 G2/M 期。Paclitaxel 會降低人類肝癌細胞株((HA 22 T/VGH)細胞胞質液中的 NAT 活性,但這是否跟 Paclitaxel 抗癌效應有關,仍待進一步的研究證實。 由本實驗顯示 Paclitaxel 可以造成人類肝癌細胞(HA 22 T/VGH)的死亡,且 隨濃度增加,效果越趨明顯,這是否意味臨床上可以使用 Paclitaxel 治療肝細 胞癌病患?此外亦需考慮藥物對人體的副作用或其它不良藥物反應(似乎需進 一步做臨床研究來證實)。 故 Paclitaxel 之癌細胞毒殺作用與藥物濃度和接觸時間有關。而在臨床上 33.
(43) 的研究發現,若肝癌細胞侵犯到門靜脈較肝靜脈枝等血管或廣泛性擴散時, 都會影響肝癌的切除效果。如果能在手術前,給予化學治療 (Neoadjuvant chemotherapy),使腫瘤體積縮小,以利切除或在手術後進行輔助性化學治療 (adjuvant chemotherapy)或許能提升病患的存活率。Paclitaxel 因而即可能成為 此類效用的藥物之一。. 34.
(44) 第六章. 結. 論. 本實驗探討 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 的 NAT 基因及 細胞週期的影響。本實驗探討不同濃度 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 細胞中 NAT 的活性及人類肝癌細胞株(HA 22 T/VGH) 細胞胞質液中 NAT 的活性的影響,發現加不同濃度的 Paclitaxel 經 6,12,18,24 小時培養, 可增加 NAT 的活性,且濃度愈高,培養時間愈久,NAT 活性增加就愈明顯。 在 RT-PCR 的實驗中,經由最後的照相結果觀察,發現不同濃度的 Paclitaxel 加在人類肝癌細胞株(HA 22 T/VGH) 細胞,(HA 22 T/VGH)細胞的 NAT 1 基因 的表現被抑制。可是 Paclitaxel 又會降低人類肝癌細胞株(HA 22 T/VGH) 細胞 胞質液的 NAT 活性,而且 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 細胞 NAT 基因的影響,主要是抑制人類肝癌細胞株(HA 22 T/VGH) 細胞 NAT 1 基 因的表現(NAT mRNA)。檢測不同濃度的 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 的細胞毒性作用,結果發現濃度越高,細胞的毒殺效果就愈強。其次 由人類肝癌細胞株(HA 22 T/VGH) 細胞經不同濃度 Paclitaxel 處理 24 小時 後,再萃取細胞 DNA 去跑電泳,結果發現 DNA 有出現裂解現象,所以發現 Paclitaxel 造成人類肝癌細胞株(HA 22 T/VGH) 細胞的死亡方式,不是經由細 胞壞死,而是經由計劃性的細胞死亡。人類肝癌細胞株(HA 22 T/VGH) 細胞 經不同濃度的 Paclitaxel 處理 24 小時後,再經由流式細胞計數儀(FACS)分析 結果顯示,Paclitaxel 會造成人類肝癌細胞株(HA 22 T/VGH) 細胞週期的變 化,發現 Paclitaxel 會使人類肝癌細胞株(HA 22 T/VGH) 的細胞週期停滯於 35.
(45) G2/M 期,利用 FAS 分析也間接證明了 Paclitaxel 對人類肝癌細胞株(HA 22 T/VGH) 細胞死亡方式不是經由細胞壞死,而是經由計劃性的細胞死亡。 此結果顯示,Paclitaxel 會增加人類肝癌細胞株(HA 22 T/VGH) 細胞 NAT 活性及抑制人類肝癌細胞株(HA 22 T/VGH) 細胞 NAT1 基因的表現。但是 Paclitaxel 的抗腫瘤作用,是否與 Paclitaxel 能降低人類肝癌細胞株 (HA 22 T/VGH) 細胞 NAT 1 基因表現有關呢?這待進一步的研究。 在整個細胞中,Paclitaxel 可以促進 NAT 對 2-AF 的乙醯化作用,而此促 進作用與劑量大小有關(dose-dependent)。歸納來說 Paclitaxel 可以且會導致細 胞的計劃性死亡影響人類肝癌細胞株(HA 22 T/VGH)的細胞週期,使細胞週期 停留在 G2/M 期。經由細胞 cyclins 的分析發現,G2 期的停留與 CDK1 –Cyclin B 被 Paclitaxel 抑制有關且與藥物濃度相關聯。. 36.
(46) References 1. 陳定信、賴明陽、陳健弘:肝細胞癌—台灣三十年的經驗。行政院衛生署八十年度委託 研究計劃報告。 2. Zinner M.J,Schwartz S.I,Ellis H.(1997)Maingot’s Abdominal Operations.10th ed.1561-1602. 3. 端本梁等著:生物化學. 藝軒圖書. 2000 年 第 2 版. 945-971 頁。. 4. Chakraborty pr,Ruiz-Oparo N,et al.(1980) Identification of hepatitis B virus DNA and expression of viral RNA in an HbsAg-producing human hepatocellular carcinoma cell line.Nature;286:531. 5. Prince AM.(1981)Hepatitis B virus and hepatocellular carcinoma:moloecular biology providesfurther evidence for an eiologic association.Hepatology;1:73. 6. Beasley RP.(1982) Hepatitis B virus as the etiologic agent in hepatocellular carcinoma-epidemiologic considerations. Hepatology 1982;2:21S. 7. Chambers R. W. (1985) Chemical carcinogensis: A Biochemical Overview. Clinical Biochemistry. 18:158-168. 8. Cerald Karp(1999) Cell and molecular biology. John Willey and Sond, Inc. 2nd ed. 704. 9. Tannock I. F. and Hill R. P. (1992) The Basic Science of Oncology. Chemical carcinogenesis, 102-118. 10. Selkrik J. K. and Maccleod M. C. (1982) Chemical carcinogensis: Nature's metablic mistake. Bioscience. 32:601-605. 11. Adamson R., Gustafsson J. A., Ito N., Nagao M., Sugimura T., Waka abayashi K. and. 37.
(47) Yamazoe Y. (1995) Heterocyclic amines in cooked foods. Proc. 23' Tnt. Priness Takamatsu Cancer Reserch Fund. Princeton Scientific, Priceton, NJ. 1-360. 12. Yamagiwa K. and Ichikwa K. (1915) Experimentelle Studie uber pathogens der epithelial qeschwulste. Mitlelungen Med. Facultat Kaiseri, Uniiu. Tokyo. 15-295. 13. Kennaway E. C. (1995) The identification of a carcinogenic compound in coal- tar: Br. Med. J. 2:749-752. 14. Acad S. (1977) Preliminary Surverys Cancer Institute, Department of Epidemiology, Chinese Academy of Medical Sciences on the epidemiology factor of esophageal cancer in China. Cancer Treat. CWro/2:85. 15. Peers F. G., and Linsell C. A. (1977) Dietary aflatoxin and human primary cancer. Annales de la Nutrition et de I'Alimentation. 31:1005-1017. 16. de Gruiji F. R. (1999) Skin cancer and solar UV radiation. European Journal of Cancer Part A. 35(14): 2003-2009. 17. Umemura T. Tokumo K, Sirma H et al; (1993) Cancer lett. 15;73(1):1-10. 18. Hein D. W., Doll M. A., Rustan T. D. and Ferguson R. J. (1995) Metabolic activation of yv-hydroxyarylamines and A-hydroxyarylamides by 16 recombinant human NAT2 allozymes: effects of 7 specific NAT2 nucleic acid substitutions. Cancer Res. 55: 3531-3536. 19. Hein D. W., Flammang T. J., Kirlin W. G., Trinidad A. and Ogolla F. (1987) Acetylator genotype dependent metabolic activation of carcinogenic A-hydroxy-arylamines by S-acetyl coenzyme A-dependent enzymes of inbred hamster tissue cytosols: relationship to arylamine. 38.
(48) TV-acetyltransferase. Carcinogenesis (Lond.). 8: 1767-1774. 20. Ogolla F., Ferguson R. J., Kirlin W. G., Trinidad A., Andrews A. F., Mpezo M. and Hein D. W. (1990) Acetylator genotype-dependent expression of arylamine A-acetyltransferase and M-hydroxy-arylamine 0-acetyltransferase in Syrian inbred hamster intestine and colon: identity with the hepatic acetylation polymorphism. Drug Metab. Dispos. 18:680-685. 21. Turesky R. J., Lang N. P., Butler M. A., Teitel C. H. and Kadlubar F. F. (1991) Metabolic activation of carcinogenic heterocyclic aromatic amines by human liver and colon. Carcinogenesis (Lond.). 12:1839-1845. 22. Minchin R. F., Reeves R T., Teitel C. H., McManus M. E., Mojarrabi B., Ilett K. F. and Kadlubar F. F. (1992) N-and 0-acetylation of aromatic and heterocyclic amine carcinogens by human monomorphic and polymorphic acetyltransferases expressed in COS-1 cells. Biochem. Biophys. Res. Commun. 185:839-844. 23. Hein D. W., Doll M. A., Rustan T. D., Gray K., Feng Y., Ferguson R. J. and Grant D. M. (1993) Metabolic activation and deactivation of arylamine carcinogens by recombinant human NAT1 and polymorphic NAT2 acetyltransferases. Carcinogenesis (Lond.). 14: 1633-1638. 24. Hein D. W., Rustan T. D., Ferguson R. J., Doll M. A. and Gray K. (1994) Metabolic activation of aromatic and heterocyclic A- hydroxyarylamines by wild-type and mutant recombinant human NAT1 and NAT2 acetyltransferases. Arch. Toxicol.,68: 129-133. 25. Denis M. G., Nicola C. H, Susan A. J., Geoffrey H. G., Hillary J. C., Andrea G., Violeta L. Y.,. 39.
(49) Ranbri G. (1997) Human acetyltransferase polymophisms. Mutation Research. 376:61-70. 26. Lower G. M. (1982) Concepts in causality. Chemically induced human urinary bladder cancer. Cancer 49:1056-1066. 27. Curtis C. Harris. (1985), Future Directions in the Use of DNA Adducts As Internal Dosimeters for Monitoring Human Exposure to Environmental Mutagens and Carcinogens. Environmental Health Perspectives. 62: 185-191. 28. Levy G. N., Chung J. G. and Weber W. W. (1993) Distribution of 2-aminofluorene p-aminobenzoic N-acetyltransferase activity in tissue of C57 rapid and Bb, A-Nats slow acetylator conqenic mice. Drug metabolism and disposition. 1:105 7-1063. 29. Heares D. J. and Weber W. W. (1993) Mutiple N-acetyltransferase and drug metabolism, tissue distribution, characterization and significance of mammalian N-acetyltransferase. Biochem. 32:353-357. 30. Hein D. W., Rustan J. D., Furman E. J. and Martin W. J. (1991) Extrahepatic expression of the N-acetylation poly- morphism toward arylamine carcinogens in tumor target organ of an inbred rat model. J. Pharmacol. Exp. Ther. 258: 232-236. 31. Short C.R., Flory W., Hsieh L. C., Aranas T., Ou S. P. and Weissenger J. (1988) Comparision of hepatic drug metabolizing enzyme activities in several agricultural species. Comp Biochem Physiol. 91C: 419-424 32. Ho C. C., Lin T. H., Chung J. G., Levy G. N. and Weber W. W. (1996) Kinetics of acetyl CoA: Arylamine N-acetyltransferase from rapid and slow acetylation frog tissue. Drug metabolism. 40.
(50) and disposition. 20:137-143. 33. Chung J. G., Kuo H. M., Ho C. C., Levy G. N. and Weber W. W. (1996) Kinetics and characterization of arylamine N-acetyltransferase from Anisakis simplex. Cancer Letters. 106:1-8. 34. Fang S. H., Chung J. G., Levy G. N. and Weber W. W. (1997) Candida albican arylamine N-acetyltransferase and characterization. Toxical left. 35. Watanabe M., Sofuni T and Nohmi T., Involvement of Cys69 residue in the catalytic mechanism of N-hydroxyarylmine 0-acetyltransferase of Salmonella typhimurium. JBiol Chem. 267:8429-8436. 36. Chang F. C. (1996) 2-aminofluorene metabolism by human digest system bacteria. Master Thesis. 37. Chung, J. G.Wang, H. H., Lo, H. H., Chang, S. S., Wu, L. T., Lee, J. H. and Hund C.F. (1998). Evidence for arymine N-acetyltransferase activity in the bacteria Helicobacter pylori. Toxicol Lett. 38. Chung, J. G., Lee, J. H., Ho, C. C., Lai, J. M., Hung, C. F. and Huan, S. C. (1997) A survey of arylamine N-acetyl- transferase in common fruits and vegetables. Joural of Food Biochemistry. 20, 481-490. 39. Trepanier L. A., Ray K, Winand N. J., Spielberg S. P. and Cribb A. E. (1997) Cytosolic Arylamine N-acetyltransferase (NAT) Deficiency in the Dog and Other Canids Due to an Absence of NAT Genes. Biochemical Pharmacology. 54:73-90.. 41.
(51) 40. Brockton N, Little J, Sharp L, Cotton, SC. (2001) N-acetyltransferase polymorphisms and colorectal cancer: a HuGE review. Am J Epidemiol 151(9): 846-61. 41. Hickman D., Risch A., Buckle V., Spun- N., Jeremiah S. J., McCarthy A. and Sim E. (1994) Chromosomal localization of human genes for arylamine/V-acetyltransferase. Biochem. 297:441-445. 42. Blum M., Grant D. M., McBride W., Heim M. and Meyer U.A. (1990) Human arylamine TV-acetyltransferase genes: isolation, chromosomal localization, and functional expression. DNA Cell Biol. 9:193-203. 43. Kelly S. L. and Sim E. (1994) Arylamine A-acetyltransferase in Balbc mice: identification of a novel mouse isoenzyme by cloning and expression in vitro. Biochem. J. 302: 347-353. 44. Fretland A. J., Doll M. A., Gray K., Feng Y. and Hein D. W. (1997) Cloning, sequencing, and functional expression ofNATI, NAT2, and NAT3 derived from C3H/HeJ (rapid) and A/HeJ (slow) acetylator inbred mouse: functional characterization of the activation and deactivation of aromatic amine carcinogens. Toxicol. Appl. Pharmacol. 142: 360-366. 45. Rothman N., Caporaso N. E., Wacholder S., Garcia-Closas M., Lubin J. H., Marcus P., Hoover R. N., Fraumeni J. F. and Jr. (1999) Evaluation of interactions between environmental exposures and common genetic polymorphisms: a population-based epidemiologic perspective. Proc. Am. Assoc. Cancer Res. 40: 762-763. 46. Hein D. W. (2000) N-acetyltransferase genetics and the ir role in predisposition to aromatic and heterocyclic amine- induced carcinogenesis. Toxicology Letters. 12-113:349-356.. 42.
(52) 47. Cascorbi I., Brockmoller J., Mrozikiewicz P. M., Muller A. and Roots I. (1999) Arylamine. V-acetyltransferase activity in man. Drug Metab. Rev. 31:489-502. 48. Fretland AJ, Leff MA, Doll MA, Hein DW. (2001) Functional characterization of human N-acetyltransferase 2 (NAT2) single nucleotide polymorphisms. Pharmacogenetics. 11(3): 207-15. 49. Grant, D.M., Blum, M., Beer, M., and Meyer, U.A. (1991) Monomorphic and polymorphic human arylamine N-acetyltransferase: acomparison of liver isozymes and expressed product of two cloned genes. Mol. Pharmacol. 39:184-191. 50. Smith, G., Stanley, L.A, Strong, R.C. and Wolf, C.R. (1995) Metabolic polymorphisms and cancer susceptibility. Cancer Survey. 25:27-35. 51. Grant, D. M., Blum, M. and Meyer, U.A. Nucleotide sequence of an intronless gene for a human arylamine N-acetyltransferase related to polymorphic drug acetylation. Nucleic Acids res. 17:3978-3986. 52. Coroneos, E., Sim, E. (1993) Arylamine N-acetyltransferase activity in human cultured cell lines. Biochem. J. 294:481-486. 53. Agundez, J. A., Olivera, M., Roman, J. M. and Benitez J. (1995) Genetic analysis of arylamine N-acetyltransferase polymorphism in breast cancer patients. Oncology, 52:7-11. 54. Lang N. P., Chu D. Z., Hunter C. F., Kendall D. C., Flammang T. J. and Kadlubar F. F. (1986) Role of aromatic amine acetyltransferase in human colorectal cancer. Arch. Surg. 121: 1259-1261.. 43.
(53) 55. Ilett K. F., David B. M., Castlecion W. M. and Kwa R. (1987) Acetylator phenotype in colorectal carcinoma. Cancer Res. 47:1466-1469. 56. Gil J. P. and Lechner M. C. (1998) Increased frequency of wild-type arylamine-. V-acetyltransferase allele NAT2*4 homozygotes in Portuguese patients with colorectal cancer. Carcinogenesis (Lond.). 19:37-41. 57. Cui, X; Guo, R; Xu, Z; Wang, B; Li, C. (2000) Relationship between metabolic phenotype of N-acetylation and bladder cancer. Chin Med J. 113(4): 303-5. 58. Cartwright R.A., Rogers H. J., Ahmed R. A., Higgens E. and Kahn M. A. (1982) Role ofN-acetyltransferase. phenotype. in. bladder. carcinogenesis.. A. pharmacogenetic. epidemiological approach to bladder cancer. Lancet. 2:842-846. 59. Risch A., Wallace D. M., Bathers S.and Sim E. (1995) Slow N-acetylation genotype is a susceptibility factor in occupational and smoking related bladder cancer. Hum. Mol. Genet. 4: 231-236. 60. Brockmoller J., Cascorbi I., Kerb R. and Roots I. (1996) Combined analysis of inherited polymorphisms in arylamine A-acetyltransferase 2, glutathione 5-transferases Ml and Tl, microsomal epoxide hydrolase, and cytochrome P450 enzymes as modulators of bladder cancer risk. Cancer Res. 56: 3915-3925. 61. 0kkels H., Sigsgaard T., Wolf A. and Autrup H. (1997) Arylamine TV-acetyltransferase 1 (NAT1) and 2 (NAT2) polymorphisms in susceptibility to bladder cancer: the influence of smoking. Cancer Epidemiol. Biomark. Prev. 6: 225-231.. 44.
(54) 62. Filiadis I. F., Georgiou I., Alamanos Y., Kranas V., Giannakopoulos X. and Lolis D. (1999) Genotypes of V-acetyltransferase-2 and risk of bladder cancer: a case-control study. J. Urol. 161: 1672-1675. 63. Shibuta J, Eto T, Kataoka A, Inoue H, Heo H, Suzuki T, Barnard GF, Mori M. (2001) Genetic polymorphism of N-acetyltransferase 2 in patients with esophageal cancer. Am J Gastroenterol. 96(12): 3419-24. 64. Pacific i G. M., Bencini C. and Rane A. (1986) Acetyltransferase in humans: development and tissue distribution. Pharmacology. 32: 283-291. 65. Hartwell L.et al(1974).Genetic control of the cell division cycle in yeast.Science 183:46-51. 66. Murray A.W and Kirschner M.W(1989).Dominoes and clock:the union of two views of the cell cycle.Science 246,614-621 67. Hunter T and Pines J.(1994).cyclins and cancer II:cyclin D and CDK inhibitors come of age.Cell;79:573-582 68. Lee C. C. R. and Fukushima S. (1998) Alterations in cyclin Dl, P53, and cell cycle related elements: Implications for distinct genetic pathways of urinary bladder carcinogensis. Urologic Oncology, 4:58-72. 69. Leake R.(1998) The cell cycle and regulation of cancer cell growth-Division of Biochemistry and Molecular Biology. 252-261. 70. Joo M, Kany YK, Kim MR .et al. (2001) Cyclin D1 overexpression in hepatocellular carcinoma. Liver. 21(2):89-95.. 45.
(55) 71. Adiba I., Mohamed A. R. (1998) Use of intracellular pH and Annexin-V flow cytometric assays to monitor apoptosis and its suppression by bcl-2 over-expression in hybridoma cell culture. Journal of Immunological Methods, 221: 43-57. 72. Smulson, M.E.; SimbuIan-Rosenthal, C.M.; Boulares, A.H.; Yakovlev, A.; Stoica, B.; lyer, S.; et. al. (2000) Roles of poly(ADP-ribosyl) ation and PARP in apoptosis, DNA repair, genomic stability and functions of p53 and E2F-1, Advances in Enzyme Regulation. 40: 183-215. 73. Affar, El Bachir; Duriez, Patrick J.; Shah, Rashmi G.; Winstall, Eric; Germain, Marc; Boucher, Claudia; et. al., (1999) Immunological determination and size haracterization of poly (ADP-ribose) synthesized in vitro and in vivo. Biochimica et Biophysica Acta (BBA)/General Subjects. 1428: 137-146. 74. Masutani, M; Nozaki, T; Wakabayashi, K; Sugimura, T, (1995) Role of poly (ADP-ribose) polymerase in cell-cycle checkpoint mechanisms following y- irradiation. Biochimie. 77: 462-465. 75. Park CH, Suh JH, et al. (1986) Evaluation of intrahepatic I-131 ethiodol on a patient with hepatocellular carcinoma: therapeutic fesibility study. Clin Nucl Med 11:514. 76. Kobayashi H, Hidaka H, et al. (1986) Treatment of hepatocellular carcinoma by transarterial injection of anticancer agents in iodized oil suspension or of radioactive iodized oil supension. Acta Radiol Diag. 27:139. 77. Ohnishi S, Saibara T, et al. (1989) Adoptive immunotherapy with lymphokine-activated killer. 46.
(56) cells plus recombinant interleukin-2 in patients with unresectable hepatocellular carcinoma. Hepatology. 10:349. 78. Ichida T, Higuchi K, et al. (1989) Treatment of hepatocellular carcinoma utilizing lymphokine-activated killer cells and interleukin-2. Cancer Chemother Pharmacol 23:545. 79. Okuno N, Takagi H, et al. (1986) Treatment for unresectable hepatoma via selective hepatic arterial infusion of lymphokine-activated killer cells generated from autologous spleen. Cancer 58:1001. 80. Farinati F, salvagnini M, et al. (1990) Unresectable hepatocellular carcinoma: a prospective controlled trial with tamoxifen. J Hepatol. 11:297. 81. Ohnishi S, Murakami T, et al. (1986) Androgen and estrogen receptors in hepatocellular carcinoma and in the surrounding noncancerous liver tissue. Hepatology. 6:440. 82. Nagasue N, Ito A, et al. (1986) Estrogen receptors in hepatocellular carcinoma. Cancer. 57:87. 83. Nagasue N, Ito A, et al. (1985) Androgen receptors in hepatocellular carcinoma and surrounding parenchyma. Gastroenteroloty. 89:643. 84. Tang ZY, Liu KD, et al. (1990) Radioimmunotherapy in the multimodality treatment of hepatocellular carcinoma with reference to second- look resection. Cancer. 65:211. 85. Order SE, Stillwagon GB, et al. (1985) Iodine 131 antiferritin: a new treatment modality in hepatoma: a Radiation Therapy Oncology Group study. J Clin Oncol. 3:1573. 86. Sachs E, Di Biscleglie AM, et al. (1985) Treatment of hepatocellular carcinoma with. 47.
(57) recombinant leucocyte interferon. Br J Cancer. 52:111. 87. Takayama T, Makuuchi M, et al. (1991) Distribution and therapeutic effect of intraarterially transferred tumor- infiltrating lymphocytes in hepatic malignancies. Cancer. 68:2391. 88. Wani MC, Taylor HL, Wall ME, et al. (1971) Plant antitumor agents. VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J Am Chem Soc. 93:2325-2327. 89. Nicolaou KC et al. (1996) Taxoids: New weapons against cancer. Scientific American 274: 84. 90. Nicolaou KC et al. (1994) Total synthesis of taxol. Nature. 367: 630. 91. Schiff PB, Horwitz SB (1980) Taxol stabilizes microtubules in mouse fibroblast cells. Proc Nate Acad Sci. 77:1561-1565. 92. Rowinsky EK, Cazenave LA, Donehower RC (1990) Taxol: a novel investigational antimicrotubule agent. J Natl Cancer Inst. 82:1247-59. 93. Torres K, Horwitz SB (1998) Mechanisms of Taxol- induced cell death are concentration dependent. Cancer Res. 58:3620-6. 94. Withers HR, Mason KA, Thames HD (1974) Response of mouse intestine to neutrons and gamma-rays in relation to dose fraction and dividing cycle. Cancer. 34:39-43. 95. Tishler RB, Geard CR, Hall EJ.et al. (1992) Taxol sensitizes human astrocytoma cells to radiation. Cancer Res.. 52:3495-97.. 96. Rowinsky EK et al. (1995) Paclitaxel (Taxol). New Engl J Mad. 322:1044.. 48.
(58) 97. McGuire WP et al. (1996) Cyclophosphamide and cisplastin compared with paclitaxel and cisplastin in patients with stage III and stage IV ovarian cancer. New Engl J Med. 334: 1. 98. Kohn EC, Sarosy G, Bicher A, et al. (1994) Dose- intense taxol: high response rate in patients with platinum-resistant recurrent ovarian cancer, J Natl Cancer Inst. 86:18-24. 99. Trimble EL, Adams JD, Vena D, et al. (1993) Paclitaxel for platinum-refractory ovarian cancer: results from the first 1000 patients registered to Natinal Cancer Institute treatment referral center 9103. J clin Oncol. 11:2405-10. 100. Gianni L et al. (1995) Paclitaxel by 3 hours infusion in combination with bolus doxorubicin in women with untreated metastatic breast cancer. J Clin Oncol. 13:2688. 101. Reichman BS, Seidman AD, Crown JP, et al. (1993) Paclitaxel and recombinant human granulocyte colony-stimulating factor asinitial chemotherapy for metastatic breast cancer. J Clin Oncol. 11:1943-51. 102. Holmes FA, Walters RS, Theriault Rl, Forman AD, Newton LK, Raber MN, et al. (1991) Phase II trial of Taxol, an active drug in the treatment of metastatic breast cancer, J Natl Cancer Inst. 83:1797-805. 103. Murphy WK, Fossella FV, Winn RJ, et al. (1993) Phase II study of Taxol, with untreated advanced non-small-cell lung cancer. J Natl Cancer Inst. 85:384-8. 104. Chang AY, Kim K, Glick J, Anderson T, Karp D, Jophson D. (1993) Phase II study of Taxol, merbarone, and piroxantrone in stage IV non-small-cell lung cancer: the Eastern Cooperative Oncology Group results. J Natl Cancer Inst. 85:388-94.. 49.
數據


+7



相關文件
當時,許多科學家認為“活”的酵素(如酵母細胞)與“沒 有生命”的胃消化液是不同的東西(生機論),但德國生 理學家Kühne則提出後者是所謂的“酵素”
細胞外液(extracellular fluid;ECF) 約佔
GLP :Good Laboratory Practice (藥物非臨床試驗優良操作規範) GTP :Good Tissue Practice(人體細胞組織優良操作規範) GCP:Good Clinical Practice (臨床試驗優良作業準則)
SF11310 張基晟 比較 Ipilimumab 加上 Etoposide/Platinum 與 Etoposide/Platinum
眼睛是心靈之窗,心靈是眼神之源。在眼球後方 感光靈敏的角膜含有 1.37 億個細胞,將收到的信 息傳送至腦部。這些感光細胞,在任何時間均可 同時處理
Your problem may be modest, but if it challenges your curiosity and brings into play your inventive faculties, and if you solve it by your own means, you may experience the tension
• 是細胞不正常增生,且這些增生的細胞可
基因編輯技術以人工核酸酶辨識特定 DNA 位置,並於此處切斷雙股 DNA。DNA 斷
