轉錄抑制因子HBP1經由對氧化還原平衡的調控抑制MDA-MB-231乳癌細胞配體依賴型之上皮生長因子受體的訊息傳遞; Transcriptional repressor HBP1 inhibits ligand-dependent epidermal growth factor receptor signaling through modulation of redox homeostasis in MDA-MB-231 breast cancer cells
97
0
0
全文
(2) 中國醫藥大學碩士班研究生 論文指導教授推薦書. 營養學系碩士班:. 林孟儀. 君所提之論文. 轉錄抑制因子 HBP1 經由對氧化還原平衡的調控來 抑制 MDA-MB-231 乳癌細胞配體依賴型之上皮生長 因子受體的訊息傳遞 (題目). 係由本人指導撰述,同意提付審查。. (簽章). 指導教授. 中 華 民 國. 97. 年. 5. 月. 28. 日.
(3)
(4) 目錄 縮寫對照表………………………………………………………………………………..I 中文摘要…………………………………………………………………………………III 英文摘要…………………………………………………………………………...…….Ⅳ 第一章 前言…………………………………………………………………………….1 第二章 文獻探討…………………………………………………………………….....2 一、乳癌(Breast Cancer)……………………………………...……………………….2 I. 介紹……………………………………………………………………………….2 II. 病理特徵…………………………………………………………………………3 III. 乳癌分期………………………………………………………………………...4 二、上皮生長因子受體(epidermal growth factor receptor;EGFR)………………….6 I. 上皮生長因子受體家族……………………………………….…………………6 II. 上皮生長因子受體之訊息傳遞……………………..…………………………..7 III. EGFR (HER1)之磷酸化位點…….……………………………………………...8 IV. EGFR 與乳癌………...…………………………………………………………..9 三、氧化壓力與訊息傳遞……………………………...…………………………….11 I. NADPH 氧化酶複合體(NADPH oxidase complex)..….…….………………….11 II. 氧化壓力訊號與生長因子……………...…………….…………………..........13 四、HBP1 (HMG (High-mobility group)-Box Protein 1)……………………………..15 I. HBP1 對細胞週期的調控……..………………………………………………...15 II. HBP1 為 p38 MAPK 路徑之作用目標..………………………………………..16 III. HBP1 標的基因….……………………………………………………………..17 五、RNA 干擾術 (RNA inference;RNAi)…………………………………………..21 I. 背景……………………………………………………………………………...21 i.
(5) II. 原理…………………………………………………………………………….21 III. 應用……………………………………………………………………………22 第三章. 材料與方法……………………………………………………………………23. 一、實驗設計………………………………………………………………………….23 二、實驗藥品與儀器………………………………………………………………….24 三、細胞培養………………………………………………………………………….27 四、質體(plasmid)的製備……………………………………………………………..28 I. 使用的質體 (乾燥)………..…………………………………………………...28 II. 轉型(Transformation)…………………………………………………………..28 III. 質體小量製備--使用 QIAGEN Plasmid Mini Kit (QIAGEN)….…………….28 IV. 質體鑑定--以限制酶切割……………………………………………………..29 V. 質體大量製備--使用 QIAGEN Plasmid Maxi Kit (QIAGEN)….…………….30 五、建立 HBP1 overexpresssion 與 knockdown 之細胞株(使用反轉錄病毒感染)..31 I. 轉染至 phoenix 細胞……………….…………………………………………...31 II. 感染目標細胞…….…………………………………………………………….31 III. 以 puromycin 篩選……….…………………………………………………….31 六、抽取 RNA--使用 RNeasy Mini Kit (QIAGEN)………………………………….32 七、RT-PCR--使用 SuperScriptTMⅢ One-Step RT-PCR System with Platinum® Taq DNA Polymerase Kit (invitrogen)……………………………………………….33 八、ROS 測定………………………………………………………………………….35 九、蛋白質萃取………………………………………...……………………………...36 十、蛋白質定量…………………………………………..…………………………...37 十一、Western blot………………………………………..…………………………...38 I. 鑄膠……………………………………………………………………………...38. ii.
(6) II. 跑膠……………………………………………………………………………..39 III. 轉印…………………………………………………………………………….39 IV. 免疫染色………………………………………………………………………40 第四章. 結果……………………………………………………………………………42. 一、HBP1 調控 EGFR 及其下游 Akt 的活性………………………………………..42 I. 細胞內 HBP1 表現上升可抑制 EGFR 及其下游 Akt 活性…………………...42 II. 細胞內 HBP1 表現降低時會增強 EGFR 與其下游 Akt 之活性……………….43 二、HBP1 調控細胞內 EGFR 活化所伴隨的 ROS 生合成………………………....44 三、細胞內 ROS 生成量可調控 EGFR 及其下游的活性……………………………45 四、HBP1 調控 EGFR 及其下游的活性是藉由抑制 NADPH 氧化酶次單位 p47phox 的基因轉錄……………………………………………………………………… 46 第五章. 討論……………………………………………………………………………66. 第六章. 結論……………………………………………………………………………72. 參考資料…………………………………………………………………………………73 附錄. pBabe.puro 與 pSUPER.puro 載體………………………………………………82. iii.
(7) 圖目錄 圖 2-1. DCIS 與 IBC 乳癌示意圖……………………………………………………………………...3. 圖 2-2. 上皮生長因子受體結構圖……………………………...…………………….…6. 圖 2-3. 上皮生長因子受體的活化與訊息傳遞…………………...…………………….8. 圖 2-4 EGFR 磷酸化位點與其下游路徑……………………………………………….9 圖 2-5 NADPH 氧化酶複合體之活化過程……………………………………………12 圖 2-6. 受體酪胺酸激酶與蛋白質酪胺酸水解酶之拮抗作用.……………………….14. 圖 2-7. HBP1 結構圖……………………………………………………………………15. 圖 2-8. HBP1 對細胞週期的調控………………………………………………………16. 圖 2-9. p38 對 HBP1 的調控…………………………………………………………….17. 圖 2-10 Wnt 訊息傳遞與 HBP1 的作用……………………………………………….18 圖 2-11. p47phox 之啟動子…...………………………………………………………….18. 圖 2-12 HBP1 對 NADPH 氧化酶的調控…………………………………………….19 圖 2-13 RNAi 之作用原理…………………………………………………………….22 圖 4-1. pBabe 控制組與 HBP1 過度表現細胞中之 HBP1 mRNA 表現量…………….47. 圖 4-2 HBP1 過度表現對 EGF 刺激不同時間後之 EGFR 活性的影響………………48 圖 4-3 HBP1 過度表現對不同濃度的 EGF 誘發之 EGFR 活性的影響………………49 圖 4-4 HBP1 過度表現對 EGF 刺激不同時間後之 Akt 活性的影響………..………50 圖 4-5 HBP1 過度表現對不同濃度的 EGF 誘發之 Akt 活性的影響………….…….51 圖 4-6. pSR 控制組與 HBP1 knockdown 細胞中之 HBP1 mRNA 表現量…………..52. 圖 4-7. HBP1 表現降低時對 EGF 刺激不同時間後之 EGFR 活性的影響………..…53. 圖 4-8 HBP1 表現降低時對不同濃度的 EGF 刺激之 EGFR 活性的影響……….…54 圖 4-9 HBP1 表現降低時對 EGF 刺激不同時間之 Akt 活性的影響…………….….55 圖 4-10 HBP1 表現降低時對不同濃度的 EGF 刺激之 Akt 活性的影響…………....56 iv.
(8) 圖 4-11. HBP1 表現增加對細胞內 EGF 誘發之 ROS 的生成之影響…………….….57. 圖 4-12 HBP1 表現降低時對細胞內 EGF 誘發之 ROS 生成之影響……….……….58 圖 4-13 以 DPI 降低細胞中 ROS 的生成量對 EGFR 活性的影響……………………59 圖 4-14. 以 DPI 降低細胞中 ROS 的生成量對 Akt 活性的影響……………………..60. 圖 4-15 以 NAC 降低細胞中 ROS 的生成量對 EGFR 活性的影響………………….61 圖 4-16. 以 NAC 降低細胞中 ROS 的生成量對 Akt 活性的影響…………………….62. 圖 4-17 pBabe 控制組與 HBP1 過度表現細胞中之 HBP1 mRNA 表現量…………..63 圖 4-18 模擬 HBP1 調控 p47phox 表現來觀察 EGFR 活性的變化…………………….64 圖 4-19 模擬 HBP1 調控 p47phox 表現來觀察 Akt 活性的變化……………………….65 圖 5-1. p47phox 之啟動子………...……………………………………………………...68. 圖 5-2 DPI 結構式……………………………………………………………………...69 圖 5-3 NAC 結構式…………………………………………………………………….69 圖 5-4 附圖 1-1. 正常的 HBP1 與突變的 HBP1 結構……………………………………………70 pBabe.puro 載體……………………………………………………………...82. 附圖 1-2 pSUPER.puro 載體…………………………………………………………...83. v.
(9) 表目錄 表 2-1. 乳癌分期…………………………………………………………………………5. vi.
(10) 縮寫對照表 縮寫. 全名. AJCC. American Joint Committee of Cancer. AR. amphiregulin. CDK. cyclin dependent kinase. DCIS. ductal carcinoma in situ. DPI. diphenylene iodonium. EGF. epidermal growth factor. EGFR. epidermal growth factor receptor. EREG. epiregulin. ERK. extra-cellular signal regulated kinase. FAD. flavin adenine dinucleotide. GAP. GTPase activating protein. GDI. guanine dissociation inhibitor. HB-EGF. heparin-binding EGF. HBP1. HMG-box protein 1. HMG. high-mobility group. IAP. inhibitor of apoptosis protein. IBC. invasive breast cancer. LCIS. Lobular Carcinoma in Situ. MAPK. mitogen-activated protein kinase. MEK. MAPK/ERK kinase. MPO. myeloperoxidase. NAC. N-acetylcysteine I.
(11) PI3K. phosphoinositide 3- kinase. PDGF. platelet-derived growth factor. PTP. protein tyrosine phosphatase. RB. retinoblastoma. Rho-GEF. Rho guanine exchange factor. RISC. RNA-induced silencing complex. RNAi. RNA inference. ROS. reactive oxygen species. RTK. receptor tyrosine kinases. SH2. Src-homology 2. SOD. superoxide dismutase. SOS. Son of sevenless. STAT. signal transduction and activators of transcription. TGF-α. transforming growth factor-α. VEGF. vascular endothelial growth factor. II.
(12) 摘要. 乳癌是婦女常見的癌症之一,其中 30%以上之乳癌有上皮生長因子受體 EGFR (epidermal growth factor receptor)過度表現的現象,這也影響了乳癌的治療與癒後情 形。因此,若能有效阻斷 EGFR 過度表現所造成的失控性細胞增生,將對乳癌的防 治有很大的貢獻。有研究指出,在生長因子的刺激下,NADPH 氧化酶(NADPH oxidase)的活化促使了 ROS (reactive oxygen species)的生成,進而增強 EGFR 及其下 游訊息傳遞因子的活性,增加腫瘤的發生。同時,也有研究發現,轉錄抑制因子 HBP1 (HMG box protein 1)可藉由抑制 p47phox (NADPH 氧化酶其中一個次單位)的基因表現 而減少 ROS 的生成。而且,HBP1 也具有抑制細胞生長,導致細胞週期停滯的作用。 因此,我們假設 HBP1 可經由抑制 p47phox 基因表現來調控 NADPH 氧化酶的活性, 減少 ROS 的生成,進而抑制乳癌細胞中 EGFR 的訊息傳遞。首先,我們利用 RNA 干擾技術將 MDA-MB-231 乳癌細胞中 HBP1 的表現降低。結果顯示,HBP1 表現降 低時,EGF 所刺激的 EGFR 及其下游 Akt 的活性有顯著的增強,伴隨著比控制組較 高的 ROS 生成;反之,當 HBP1 在 MDA-MB-231 細胞過度表現時,確實抑制了 EGF 所誘發的 EGFR 訊息傳遞和 ROS 生合成。另外,給予 DPI (diphenyleneiodonium chloride,NADPH oxidase 抑制劑)與 NAC (N-acetyl cysteine,ROS 清除劑)來降低細 胞內 EGF 刺激所產生的 ROS 濃度後,EGFR 及其下游的活性皆有被抑制的現象。 接著,我們更進一步利用 p47phox siRNA 降低細胞中 p47phox 基因表現後發現,EGFR 及下游 Akt 的活性均有減低的情形。因此,由上述結果可知,HBP1 確實可經由抑 制 p47phox 基因表現來調控 NADPH 氧化酶的活性,減少 ROS 的生成,進而抑制 EGFR 的訊息傳遞。. III.
(13) Abstract. Breast cancer is one of the most frequent and spontaneous malignancies diagnosed in women. Greater than 30% of human breast cancers have EGFR overexpression and are often associated with poor prognosis. Inhibition of EGFR family signaling has become an important treatment option for specific breast cancers. The activation of NADPH oxidase and subsequent ROS generation are required for EGFR signal transduction cascade. Importantly, the transcriptional repressor HBP1 has been reported to decrease ROS production through inhibiting the expression of p47phox gene, one of the NADPH oxidase subunit. Therefore, we hypothesized here that HBP1 may block EGFR signaling through inhibition of NADPH oxidase activity in MDA-MB-231 breast cancer cells. To test the model, HBP1 shRNA was used to knockdown endogenous HBP1 gene, resulting in a higher EGF-stimulated EGFR activation and ROS generation than the control. In contrast, the HBP1 overexpressing cells showed decreased EGFR activation and ROS generation relative to the control. NAC (N-acetylcysteine) or DPI (Diphenyleneiodonium chloride) treatment also reduced EGFR signaling with a concomitant decrease in ROS production. Finally, to mimic HBP1 down-regulation of p47phox function, the p47phox siRNA was used and the result was consistent with the HBP1 effect, showing a down-regulated EGFR signaling. Thus, our study demonstrates that HBP1 can block EGF-induced EGFR signaling through regulating NADPH oxidase function in breast cancer cells. The study may provide a mechanistic framework for understanding the potential chemopreventive function of phytochemicals in breast and other cancers.. IV.
(14) 第一章 前言. 第一章、前言. 乳癌是全世界女性最常見的癌症之一,在歐美國家,平均每 4 個女性癌症患者 中就有一個罹患的是乳癌;而在台灣,隨著生活及飲食習慣日益西化,乳癌的發生 率也日漸升高。近年的臨床治療發現,第一期乳癌治療後的存活率達 80%,而原位 性乳癌治療後的存活率更接近 100%。因此,早期的預防與治療便已成為一個很重 要的課題。. 乳癌發生的原因有很多,其中有 30%以上與上皮生長因子受器(EGFR)的過度表 現有關,而我們希望能夠找出可降低 EGFR 活性表現的途徑,針對這 30%的乳癌患 者來做有效的控制。已有研究證實,EGFR 受到配體刺激而活化後,會促進 NADPH 氧化酶活性,增加體內活性氧化物的生成;而所產生的活性氧化物會經由另外的途 徑去促進 EGFR 的活性表現。. 近年來有文獻指出,HBP1 轉錄因子可以抑制體內 NADPH 氧化酶的活性,減 少活性氧化物的生成;除此之外, HBP1 也可阻斷一些與細胞週期相關的因子來抑 制細胞的增生。因此,我們也希望能夠提出更多關於 HBP1 的研究,藉以建立 HBP1 的地位。. 本研究主要是探討 HBP1 經由影響氧化還原平衡來抑制 EGFR 活性,希望對於 乳癌的防治有進一步的貢獻。而在未來,也可將此機制應用於一些已知具有抗癌功 效之天然的植物性化學物質的研究。. 1.
(15) 第二章 文獻探討. 第二章、文獻探討. 一、乳癌(Breast cancer). I. 介紹 根據民國 96 年行政院衛生署的統計,乳癌在女性癌症死因排名第四位。乳癌好 發年齡在 40-50 歲之間,較歐美國家的好發年齡約提早十歲,但任何成年婦女皆有 罹患此症的危險。. 乳癌是因為乳房乳腺管細胞或是乳小葉細胞產生不正常分裂、繁殖而形成之惡 性腫瘤。乳癌除了局部侵犯乳房組織外,更可能轉移到遠處組織,如:肺、骨骼、 肝臟、腦等器官,而危害生命安全。. 乳房組織主要由乳小葉(負責乳汁分泌)、乳管(乳汁輸送管道,連接於乳小葉及 乳頭間)及脂肪組織、血管、淋巴管所構成;大部份乳癌源自乳管(約佔 80%),小部 份來自乳小葉(約佔 10~15%),極少數乳房惡性腫瘤源自淋巴、血管、脂肪組織。 乳癌病理組織學上可區分為原位性乳癌(Ductal Carcinoma in Situ;DCIS、Lobular Carcinoma in Situ;LCIS)及侵犯性乳癌(Invasive Breast Cancer;IBC)。原位乳癌大多 來自乳管,稱為原位性乳管癌(DCIS),少部份來自乳小葉稱為原位性乳小葉癌 (LCIS);同樣的侵犯性乳癌也大多來自乳管稱為侵犯性乳管癌;少部份來自乳小葉 而稱為侵犯性乳小葉乳癌。. 2.
(16) 第二章 文獻探討. Ductal Carcinoma in Situ (DCIS). Invasive Breast Cancer (IBC) 圖 2-1. DCIS 與 IBC 乳癌示意圖. 資料來源:National Cancer Institute (http://www.cancer.gov). II. 病理特徵 臨床上最常見之侵犯性乳癌,其病理組織學中三分之二屬於非特殊外型的侵犯 性乳管癌,三分之一屬於具有特殊外型的侵犯性乳癌,如黏液癌 (mucinous)、髓質 癌 (medullary)、管狀癌 (tubular)。侵犯性乳癌因具有侵犯乳管-乳小葉周圍間質組 織能力,會經由淋巴、血管產生遠處轉移而威脅生命。理論上原位乳癌沒有能力產 生遠處轉移,但在臨床上卻有極少數原位乳癌會有淋巴腺、遠處器官的轉移。原位 乳管癌在病理學主要分為乳突狀 (papillary),微小乳突 狀(micropapillary),篩狀 (cribriform),實心型 (solid)及粉刺型 (comedo) 等種類。其中以粉刺型較有演變成. 3.
(17) 第二章 文獻探討. 侵犯癌傾向,也較易為多發病灶及產生腋下淋巴腺轉移。在國外新發現的乳癌病例 中,原位乳癌佔 30%以上,台灣原位乳癌病例也逐漸增加中。原位乳癌在分期上屬 於零期乳癌,其預後極佳。唯臨床上國內一般乳癌發現時期,已屬侵犯癌程度為多, 為了早期發現乳癌,乳癌的篩檢工作應廣泛推廣。. III. 乳癌分期 病理分期常採用依照腫瘤大小,淋巴腺轉移程度及是否遠處轉移(即 TNM 分類 系統)分成 0、Ⅰ、Ⅱ、Ⅲ、Ⅳ期;一般依據 AJCC (American Joint Committee of Cancer) 系統: 0 期:原位乳癌,含乳管原位癌及乳小葉原位癌。 I 期:腫瘤小於或等於 2 公分,無淋巴腺轉移。 II 期: II A:腫瘤小於 2 公分,淋巴腺有轉移;或腫瘤介於 2-5 公分,無淋巴腺轉移。 II B:腫瘤 2-5 公分,淋巴腺有轉移;或腫瘤大於 5 公分且淋巴腺無轉移。 III 期: III A: (1)腫瘤大於 5 公分,淋巴腺有轉移。 (2)腫瘤小於 5 公分但腋下淋巴腺彼此連黏,或腋淋巴周圍組織發生連黏。 (3)腫瘤小於 5 公分,發生 4~9 顆淋巴腺轉移。 III B:乳癌腫瘤直接侵犯胸壁或乳房皮膚潰瘍、橘皮變化。 III C:不論腫瘤大小,但發生多於 10 顆以上淋巴腺轉移,或同側腋下及內乳淋巴 結 (internal mammary node) 轉移或鎖骨上、下淋巴腺轉移。 IV 期:任何腫瘤大小,但有發生遠處轉移(如:肺、肝、骨骼、腦部等)。. 4.
(18) 第二章 文獻探討. 表 2-1. 乳癌分期 Tumor (T). Node (N). Metastasis (M). 0. T0. N0. M0. I. T1. N0. M0. II A. T0. N1. M0. T1. N1. M0. T2. N0. M0. T2. N1. M0. T3. N0. M0. T0. N2. M0. T1. N2. M0. T2. N2. M0. T3. N1. M0. T3. N2. M0. T4. N0. M0. T4. N1. M0. T4. N2. M0. III C. any T. N3. M0. IV. any T. any N. M1. II B. III A. III B. 資料來源:breastcancer.org (http://www.breastcancer.org/). 5.
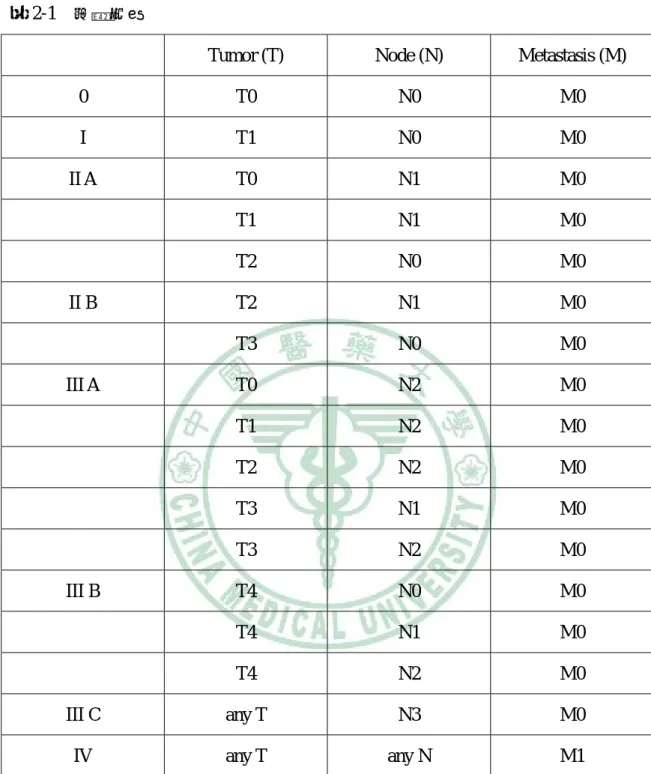
(19) 第二章 文獻探討. 二、上皮生長因子受體 (epidermal growth factor receptor;EGFR). I. 上皮生長因子受體家族 上皮生長因子受體是一種橫跨細胞膜的受體,屬於 ErbB 家族,具有受體酪胺 酸激酶 (receptor tyrosine kinases;RTKs)的活性,為許多訊息傳遞路徑之首,可參與 調控細胞增生、存活、黏附、遷移及分化(1, 2)。. 上皮生長因子受體家族共包含四種不同的受體,分別為 EGFR (HER1)、HER2、 HER3 與 HER4,EGFR (也就是 HER1)是最早被發現的受體。這些受體結構依所在 位置分為三個部分: --細胞外介質結合區 (extracellular ligand-binding domain) --穿透細胞膜區 (transmembrane domain) --細胞內區 (intracellular domain)(1)。. 圖 2-2. 上皮生長因子受體結構圖. 資料來源:(1). 6.
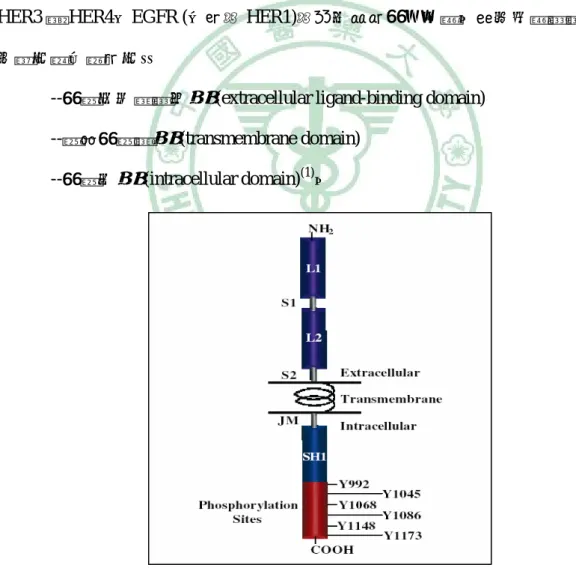
(20) 第二章 文獻探討. 其他家族成員 (HER2~HER4)也有相同的構型,但在 HER2 之細胞外介質結合 區卻無法與配體結合,須與 EGFR 形成異質雙元體 (heterodimer)的形式來活化(3, 4); 而 HER3 則是缺乏酪胺酸激酶活性,也是需靠其他受體的激酶活性來完成訊息的傳 遞(1, 5)。. II. 上皮生長因子受體之訊息傳遞 已知有十種不同的配體可分別和 EGFR、HER3 與 HER4 結合 (因 HER2 細胞 外介質結合區無法與配體結合),較常見的包括有:生長因子 (epidermal growth factor; EGF)、α變形性生長因子 (transforming growth factor-α;TGF-α)、生長 因子雙調蛋白 (amphiregulin;AR)、表皮調節素 (epiregulin;EREG)與肝素結合上 皮生長因子 (heparin-binding EGF;HB-EGF)(3)。當配體與受體結合後,會誘導受體 形成 EGFR-EGFR 的同質雙元體 (homodimer)形式,或是 EGFR-HER2、EGFR-HER3 或 EGFR-HER4 的異直雙元體的形式,並活化受體酪胺酸激酶,使得位於受體尾端 的酪胺酸殘基(tyrosine residues)磷酸化(4)。而這些磷酸化的酪胺酸殘基則會被其他含 有 Src 同源區 2 (Src-homology 2 domains;SH2)的蛋白質結合,像是 Grb2、Shc、PLC γ,進而活化一連串的訊息傳遞,使細胞外的訊號被帶往細胞核中(3, 6)。EGFR 主要 活化的下游訊息傳遞路徑包括:Ras-Raf-MEK-ERK1/2、STAT3、STAT5,這些路徑 主要是調控基因轉錄、細胞週期 G1~S 期與細胞的增生、分化(6, 7);PI3K-Akt-mTOR 路徑則與促進細胞存活、抗細胞凋亡有關(5, 8, 9)。. 7.
(21) 第二章 文獻探討. 圖 2-3. 上皮生長因子受體的活化與訊息傳遞. 資料來源:(9). III. EGFR (HER1)之磷酸化位點 位於 EGFR 的酪胺酸磷酸化位點位於細胞內區與其尾部 C 端 (carboxyl terminus)194 個胺基酸位置(10, 11)。隨著這些位點的不同,所活化的下游路徑也有所 區別。以下為幾個常見之位點: Tyr845:位於細胞內區上之活化環,當此位點磷酸化後可增加 EGFR 受質 Shc 與 PLCγ磷酸化,促進細胞增生與腫瘤形成。 Tyr1068、1086:會與 Src 受質 Grb2,活化 SOS-Ras-Raf-MAPK-ERK 路徑,調控. 8.
(22) 第二章 文獻探討. 細胞增生與分化。 Tyr1173:會與磷酸化的 EGFR 受質 Shc 和磷酸化的 Src 受質 Gab1 結合, 活化 Grb2-SOS-Ras-Raf-MAPK-ERK 路徑與 p85-PKD-Akt 路徑。 Tyr1045:會和磷酸化的 Src 受質 CBL 結合,與泛素化作用 (ubiquitination) 有關。. 圖 2-4 EGFR 磷酸化位點與其下游路徑 資料來源:(12). IV. EGFR 與乳癌 在許多細胞型態中,EGFR 在每個細胞中分佈約為 2×104~2×105,而在許多癌症 細胞中皆可發現 EGFR 的數量大於 106,有過度表現的現象,且也與其預後不佳的. 9.
(23) 第二章 文獻探討. 情形有關(5, 8, 9)。有文獻指出,這些受體的增加會減少癌症患者的存活率、降低對化 療的敏感性;在乳癌的治療上,也大幅降低內分泌療法的效果(5, 9, 13, 14)。. 在 1992 年的一篇文獻回顧中有提到,在 40 篇不同的研究裡的 5232 個乳癌病患 當中,有 45%的患者是 EGFR 陽性(15);在另一篇回顧的 370 名乳癌患者中,也有 47% 為 EGFR 陽性(16)。有大於 20%的乳癌病患,體內的 EGFR 持續不斷的活化表現 (EGFRVIII);另外,在侵入性乳癌的患者中也發現 25%有 HER2 過度表現的情形(17); 最近的臨床研究中也提到,EGFR 與 HER2 磷酸化的增加,對於轉移性乳癌的診斷 與治療都有負面的影響(18, 19)。以上資料在在顯示,EGFR 對乳癌的影響是不容忽視 的。. 10.
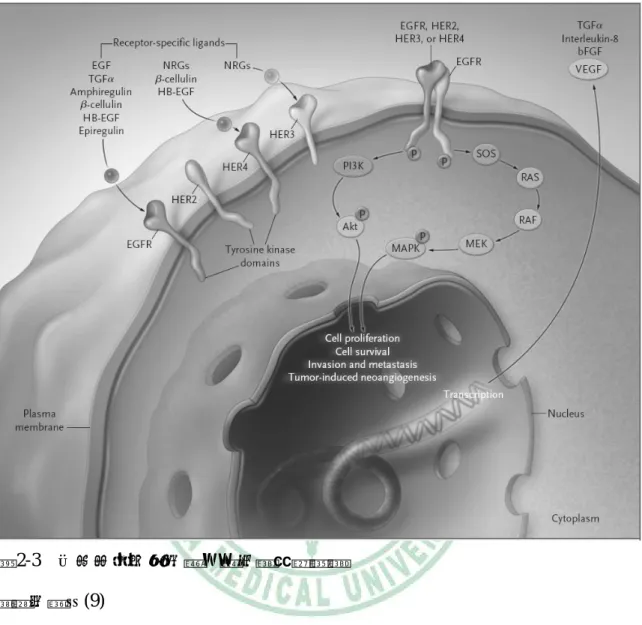
(24) 第二章 文獻探討. 三、氧化壓力與訊息傳遞. I. NADPH 氧化酶複合體(NADPH oxidase complex) NADPH 氧化酶是由多個次單位所組成的複合體,主要是將 NADPH 上的電子 傳遞給 O2,使其轉變成超氧化物 (superoxide)(20-22)。其組成包括位於細胞膜上之具 催化作用的 gp91phox、調節作用的 p22phox,及其他位於細胞質的調節次單位 p47phox、 p67phox 與 p40phox (20, 23, 24);另外,還有一個小分子的 GTP 結合蛋白 Rac(21, 25)。當 NADPH 受到刺激後,這些位於細胞質中之次單位會位移至細胞膜上而活化系統並 產生超氧化物,而產生的超氧化物會再經由超氧化物歧化酶 (superoxide dismutase; SOD) 及觸酶 (catalase)作用而生成其他的活性氧化物 (reactive oxygen species;ROS) (20, 21). 。. gp91phox gp91phox 為 NADPH 氧化酶重要的組成因子,具有可攜帶電子的 FAD (flavin adenine dinucleotide) 與兩個具有還原能力的血基質 (heme group),並提供其他 NADPH 次單位與氧分子結合的位置(20)。gp91phox 與另一個細胞膜上之次單位 p22phox 合稱為細胞色素 b558 (cytochrome b558)(21)。. p47phox 當受到刺激時,p47phox 在絲胺酸殘基 (serine residues) 磷酸化後會造成結構上的 改變,使其兩個 Src 同源區 3 (Src homology 3 domain;SH3) 與 p22phox C 端之富含 脯胺酸 (proline) 區域結合;此外,p47phox 之 phox 同源區 (phox homology domain; PH) 會與細胞膜之磷酸肌醇 (phosphoinositide) 結合,使 p47phox 可貼附於細胞膜 上。而 p47phox 會藉由其尾端富含脯胺酸之區域與 p67phox 之 SH3 作用,促使 p67phox. 11.
(25) 第二章 文獻探討. 位移至膜上(21)。. p67phox p67phox 的功能是將 NADPH 上之電子轉移至 FAD,此過程為超氧化物產生之速 率限制步驟(21)。. Rac Rac 屬於 Rho 家族,有三種同質體,為 NADPH 氧化酶活化所需。NADPH 氧 化酶未活化時,Rac 是以 GDP 的形式存在細胞質中,並藉由 GDI (guanine dissociation inhibitor) 來使其穩定(26-28)。當受到外界刺激後,Rho-GEF (Rho guanine exchange factor)會促進 Rac 上之 GDP 轉換成 GTP 的形式,促使 Rac 與其他細胞質中之次單 位位移至細胞膜,啟動 NADPH 氧化酶而產生超氧化物(29-31)。活化的 Rac 可藉由 GAP (GTPase activating protein) 將 Rac 上的 GTP 水解,使其恢復成不活化狀態(21)。. 圖 2-5 NADPH 氧化酶複合體之活化過程 資料來源:(31). 12.
(26) 第二章 文獻探討. II. 氧化壓力訊號與生長因子 有文獻指出,血小板源生長因子 (platelet-derived growth factor;PDGF)、上皮 生長因子 (epidermal growth factor;EGF) 等配體可刺激細胞在短時間之內產生活性 氧化物(32, 33),而這樣的現象與受體酪胺酸激酶-Ras 的路徑有關。Ras 在細胞增生、 分化等也扮演重要的角色,在許多癌症當中也有發現 Ras 有突變或過度表現的情形 (34)。. 當配體與受體結合並促使磷酸化發生後,酪胺酸激酶活化且伴隨著 SOS (Son of sevenless;Ras-specific guanine exchange factor) 表現(35-37),SOS 會與活化的受體酪 胺酸激酶和 Grb2 形成三元複合體 (ternary complex),促進 Ras 上的 GDP 轉換成 GTP,並位移至細胞膜上,活化 NADPH 氧化酶之 Rac 的表現,進而促使超氧化物 及其他活性氧化物的產生(38)。. 另一方面,配體刺激所產生之活性氧化物在細胞中除了增加氧化壓力外,同時, 適度的氧化壓力也可藉由抑制蛋白質酪胺酸水解酶 (protein tyrosine phosphatase; PTP) 的活性來調節受體酪胺酸激酶的磷酸化,扮演訊息傳遞的角色(24, 28, 29, 35, 39)。 蛋白質酪胺酸水解酶與受體酪胺酸激酶在細胞酪胺酸的磷酸化扮演重要的角色,這 兩個酵素的作用彼此是互相拮抗的,精密的調控細胞中酪胺酸的磷酸化作用,分別 發揮促進信號與抑制信號的作用(20, 40)。. 細胞在氧化狀態下可調節蛋白的半胱胺酸殘基 (cysteine residues),形成穩定的 Cys-SOH 結構(22, 39),將蛋白質酪胺酸水解酶上的半胱胺酸殘基轉變成穩定的氧化態 結構(29, 35),使蛋白質酪胺酸水解酶的活性受到抑制而無法阻斷受體酪胺酸激酶的促. 13.
(27) 第二章 文獻探討. 磷酸化與其下游之訊息傳遞。. 圖 2-6. 受體酪胺酸激酶與蛋白質酪胺酸水解酶之拮抗作用. 資料來源:http://www.bc.sinica.edu.tw/PI_DetailE.asp?Auto=36. 14.
(28) 第二章 文獻探討. 四、HBP1 (HMG (High-mobility group)-Box Protein 1). HBP1 屬於 HMG (high-mobility group) 家族的成員,為一轉錄因子,大小約 513 個胺基酸,其結構包含: --HMG-box DNA 結合區 (HMG-box DNA binding domain) --抑制區 (repression domain) --RB 結合區 (RB binding domain) --p38 結合區 (p38 binding domain). 圖 2-7 HBP1 結構圖 資料來源:(41). I. HBP1 對細胞週期的調控 HBP1 對細胞週期的影響與 RB (retinoblastoma) 對細胞分化與 G1 期的調控有相 似的功能。RB 家族 (RB、p107、p130) 為細胞週期調控的核心之一,它可以抑制 細胞週期的進行,使其停滯於 G1 期。RB 及其他成員 p107 和 p130 可分別與轉錄因 子 E2F 結合來調控細胞週期(42),當 RB 被 cyclin 與 CDK (cyclin dependent kinase)磷 酸化後,會使 RB 與 E2F 分離,E2F 釋放而使細胞週期得以進行。而 HBP1 可和 RB 家族成員中的 RB 與 p130 作用,抑制細胞週期 G1 期的進行,促進細胞分化(42-44)。. 15.
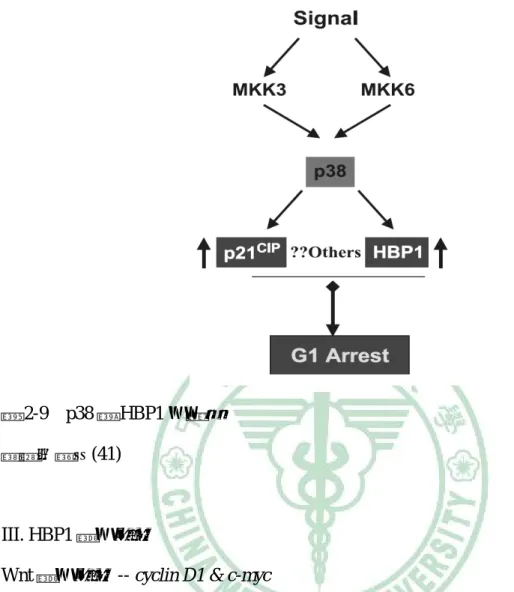
(29) 第二章 文獻探討. 圖 2-8 HBP1 對細胞週期的調控 資料來源:(41). II. HBP1 為 p38 MAPK 路徑之作用目標 p38 MAPK 路徑參與包括細胞凋亡與細胞週期停滯,有抑制腫瘤的作用。有文 獻指出,p38 MAPK 的活性喪失與乳癌及其他癌症的發生有關。最近的研究中有提 到,p38 MAPK 可藉由直接調控 p21 與 HBP1 來達到抑制腫瘤作用與促使細胞週期 停滯於 G1 期(41, 45)。. 16.
(30) 第二章 文獻探討. 圖 2-9. p38 對 HBP1 的調控. 資料來源:(41). III. HBP1 標的基因 Wnt 標的基因 -- cyclin D1 & c-myc Wnt 的訊息傳遞失調與乳癌、直腸癌及其他癌症的發生有關(46, 47)。當 Wnt 訊息 持續表現時,會抑制細胞內β-catenin 裂解,增加其穩定性,使β-catenin 在細胞質 裡堆積,並位移至細胞核中與 Tcf/Lef 轉錄因子形成複合體,促進下游基因像是 cyclin D1、c-myc 等轉錄,促使細胞異常增生而導致癌化(47);而 HBP1 可藉由與 Lef/Tcf 競爭結合至 cyclin D1、c-myc 之啟動子 (promoter) 來降低基因的轉錄作用(48)。. 17.
(31) 第二章 文獻探討. 圖 2-10 Wnt 訊息傳遞與 HBP1 的作用 資料來源:(48). NADPH 氧化酶之次單位 -- p47phox NADPH 氧化酶為細胞內活性氧化物生成的重要酵素複合體,其次單位之一 p47phox 的啟動子在-1243 至-1318 處上含有 HBP1 之 DNA 結合序列,HBP1 可藉由 結合至啟動子上來抑制 p47phox 的轉錄,使 NADPH 氧化酶無法活化而降低超氧化物 的生成(49)。. 圖 2-11. p47phox 之啟動子. 資料來源:(49). 18.
(32) 第二章 文獻探討. 圖 2-12 HBP1 對 NADPH 氧化酶的調控 資料來源:(49). N-myc N-myc 也和細胞週期 G1 的調控有關,且在許多癌症當中都有過度表現的現象 (41)。. histon H10 組織蛋白 histon 為染色質的蛋白質,共有六種,其中 H10 連接組織蛋白可將核 小體核心顆粒與 DNA 的進入位點及 E 位點結合,因而可以將 DNA 緊扣在位。而 HBP1 可活化此基因的表現,因此推測 HBP1 對於染色質的重建可能有一定的影響 (41). 。. 19.
(33) 第二章 文獻探討. myeloperoxidase 骨髓過氧化酶 (myeloperoxidase;MPO) 為骨髓性白血病 (myeloid leukemia) 臨 床診斷重要的指標,而 HBP1 可活化 MPO 基因。在骨髓細胞中,缺乏 HBP1 可能 會導致這個重要的分化指標有異常的表現(41, 50)。. 20.
(34) 第二章 文獻探討. 五、RNA 干擾術 (RNA interference;RNAi). I. 背景 RNAi 最早於 1990 年時被發現,原本是將查爾酮合酶 (chalcone synthase) 的基 因轉殖於矮牽牛花中,希望藉由增加一套基因而使花色更鮮豔。結果卻發現不但多 一套的基因沒有表現,原本存在的那一套基因反而失去功能而出現白花,當時稱此 現象為「共同抑制 (co-suppression)」。直到 1998 年,Fire 與 Mello 在線蟲 (Caenorhabditis elegans) 上也發現這種 RNA 干擾現象,並首度被稱為 RNAi。直到 2001 年初,Thomas Tuschl 率先證明了在哺乳動物細胞中,亦可以使用 RNAi 來抑 制特定基因的表現,因此 RNAi 技術也由植物延伸到了哺乳類動物,對於其機制也 有了更深入的了解。在 2006 年時,RNAi 技術也透過諾貝爾獎受到肯定(51)。. II. 原理 RNAi 是利用細胞內的雙股 RNA 所引發的基因沈默作用。長鏈的雙股 RNA 先 經由 Dicer (一種 RNase) 將其切割成多個小段,每段約 19~25nt,稱為小干擾 RNA (small interfering RNA),也就是所謂的 siRNA。siRNA 會被細胞質中的核糖核酸誘 導沉默複合體 (RNA-induced silencing complex;RISC) 辨識並結合,使雙股的 siRNA 解開,形成 sense 與 antisense 兩個單股的 RNA,並將其中的正義股 (sense) 分解, 保留反義股 (antisense)。而保留的反義股則會引導 RISC,利用其序列專一性找到目 標 mRNA,進而將其降解(51-53)。. 21.
(35) 第二章 文獻探討. 圖 2-12 RNAi 之作用原理 資料來源:(52). III. 應用 RNAi 技術的應用相當廣泛,在生物醫學方面,許多疾病的發生是因為特定基 因表現異常或過度表現,我們可以依所需來關閉非必要或致病基因的功能,以抑制 這些異常或過度活化的基因。理論上說,若能關閉致病基因的表達,則很多疾病將 被治癒。在動物實驗中已證明,可以通過 RNAi 的方法抑制導致血膽固醇升高的基 因;病毒性疾病,眼疾,心血管代謝性疾病等方面的臨床試驗也正在進行中。因此, 利用 RNAi 技術可用來開發出對抗癌症、感染性疾病、心血管與腦血管疾病、神經 退化性疾病、糖尿病…等各種疾病的新療法。. 22.
(36) 第三章 材料與方法. 第三章、材料與方法 一、實驗設計. MDA-MB-231. Aim 1. 細胞中 HBP1 之表現對 EGF 誘 導之 ROS 的生成及 EGFR 與下 游 Akt 活性的影響. HBP1 過度表現:. HBP1 表現降低:. EGFR 與 Akt 活性. EGFR 與 Akt 活性. ROS 生成. ROS 生成. Aim 2. 降低細胞中 ROS 生成是否可抑制 EGF 刺激之 EGFR 與下游活性. 以 DPI 與 NAC 處理細胞: EGFR、Akt 活性. Aim 3. 模擬細胞內 HBP1 調控 NADPH 氧 化酶的情形來觀察 EGFR 與 Akt 活 性變化. 降低細胞內 p47phox 表現: EGFR、Akt 活性. 23.
(37) 第三章 材料與方法. 二、實驗藥品與儀器 藥品: 名稱. 廠牌. 貯存溫度. Acrylamide. Genepure. 4℃. Agarose. AMRESCO®. 室溫. Ampicillin. Bio Basic Inc.. 4℃. Ammonium Persulfate. J. T. Baker. 室溫. β-mercaptoethanol. J. T. Baker. 室溫. Bovin Serum Album. USBiological. 4℃. Bromophenol Blue. Bio Basic Inc.. 室溫. Deoxycholate. Sigma. 室溫. DMSO. Sigma. 室溫. Dulbecco’s Modified Eagle Medium. Gibco. 4℃. ECL. PerkinElmer. 4℃. EcoR I. BioLabs. -20℃. EDTA. J. T. Baker. 室溫. Ethanol. 東華化工. 室溫. Etidium Bromide. Genepure. 4℃. Fetal Bovine Serum. Gibco, USA. -20℃. A. B. D. E. F. 24.
(38) 第三章 材料與方法. FuGENE® 6. Roche. 4℃. Glycine. USBiological. 室溫. Hind III. BioLabs. -20℃. Isopropanol. Sigma. 室溫. L-Glutamine. Gibco. -20℃. Lucigenin. Sigma. 4℃. Luria Bertani medium. Himedia. 室溫. Luria Bertani Agar. Himedia. 室溫. NaCl. AMRESCO®. 室溫. 10x NEBuffer EcoR I. BioLabs. -20℃. OPTI-MEM®I. Gibco. 4℃. 10x PBS. UniRegion Bio-Tech. 室溫. Penicillin/Streptomycin. Gibco. -20℃. Phosphatase Inhibitors Cocktail. Sigma. -20℃. Polybrene. Sigma. -20℃. Protease Inhibitors Cocktail. Sigma. -20℃. Protein Assay Dye Reagent Contrate. BioRad. 4℃. G. H. I. L. N. O. P. 25.
(39) 第三章 材料與方法. Puromycin. Sigma. -20℃. SDS. Merck. 室溫. Sodium Bicarbonate. Sigma. 室溫. 50x TAE. UniRegion Bio-Tech. 室溫. TEMED. J. T. Baker. 室溫. Tris-base. USBiological. 室溫. Triton X-100. J. T. Baker. 室溫. Trypsin. Gibco. -20℃. Tween 20. Showa. 室溫. S. T. 儀器: 名稱. 廠牌. 倒立式顯微鏡. Olympus. 離心機. Sigma. 微量離心機. DENVILLE. iCycler. BioRad. MultilmageTM Light Cabinet. Alpha Innotech Corporation. Mulpid®-2 plus electrophoresis. ADVANCE. CLC-110 chemiluminescence detector. TOHOKU. CLC-10 CL counter. TOHOKU. PowerPacTM Basic Power Supply. BioRad. Mini-PROTEAN®3 Cell. BioRad. Mini Trans-Blot® Electrophoretic Transfer Cell. BioRad. 26.
(40) 第三章 材料與方法. 三、細胞培養. 人類乳癌細胞株 MDA-MB-231 受贈自楊新玲教授實驗室,培養於含有 10% Fetal Bovine Serum、2% L-Glutamine 與 1% Penicillin/Streptomycin 之 Dulbecco’s Modified Eagle Medium 中,置於 37℃、5% CO2 之培養箱內培養,每隔 2~3 天更換一次培養 液,當細胞長至八分滿時即進行繼代培養。. 繼代培養 將培養皿中之培養液吸除後,加入 10 ml 滅過菌之 PBS 清洗後,小心地吸除。 加入 1 ml trypsin 溶液後,放入 37℃培養箱中處理 5 分鐘,於倒立顯微鏡下觀看細 胞是否全數脫落。確定細胞完全脫落後,加入 10 ml 培養液中和 trypsin 作用,並將 細胞液取出放入 15 ml 離心管中,以 1000 rpm 速度離心 5 分鐘後,吸除上清液,加 入適量的培養液將細胞打散後,分至裝有培養液之培養皿中,置於 37℃、5% CO2 之培養箱內培養。. 細胞冷凍 將培養皿中之培養液吸除後,加入 10 ml 滅過菌之 PBS 清洗後,小心地吸除。 加入 1 ml trypsin 溶液後,放入 37℃培養箱中處理 5 分鐘,於倒立顯微鏡下觀看細 胞是否全數脫落。確定細胞完全脫落後,加入 10 ml 培養液中和 trypsin 作用,並將 細胞液取出放入 15 ml 離心管中,以 1000 rpm 速度離心 5 分鐘後,吸除上清液,每 管加入 3ml 含有 7% DMSO 的培養液,分裝 1 ml 至冷凍保存管中,放入 4℃冰箱裡 10 分鐘後,移至-20℃放置 30 分鐘後,放到-80℃中 24 小時,最後移至液態氮筒中 保存。. 27.
(41) 第三章 材料與方法. 四、質體 (plasmid) 的製備. I. 使用的質體 (乾燥) Name. Relevant characteristics. pBabe-puro. empty vector control. pBabe-puro-AML-HBP1-HA. overexpressing HBP1. pSR-puro. shRNA empty vector control. pSR-puro-213-shHBP1. “213” shRNA against HBP1. pSR-puro-022-shHBP1. “022” shRNA against HBP1 質體來源:Dr. Yee Lab.. II. 轉型 (Transformation) 將 DH5α competent cells (ECOS 101;Yeastern Biotech) 自-80℃取出後回溫, 取 30 μl 至 eppendorf 中,加入 3 μl 質體 DNA 後置於冰上 10 分鐘後,將含有質 體 DNA 及 competent cells 之 eppendorf 放入 42℃水浴 45 秒,立即放回冰上 2 分鐘。 接著,加入 300 μl 之 Luria Bertani medium,放入 37℃培養箱,以 100 rpm 速度震 盪培養 1 小時,取 200 μl 混合液均勻塗抹在 Luria Bertani Agar 上,置於 37℃培養 箱 24 小時。. 挑選菌落:以滅菌過的牙籤輕輕刮起培養皿上單一菌落,連同牙籤一起放入含 有 50 μg/ml 之 Ampicillin 之 Luria Bertani medium 的培養管中,再度放入 37℃培養 箱,以 200 rpm 速度震盪培養 24 小時。. III. 質體小量製備--使用 QIAGEN® Plasmid Mini Kit (QIAGEN) 將上述培養 24 小時之菌體離心 15 分鐘 (6000g,4℃) 後,將上清液倒掉,加 28.
(42) 第三章 材料與方法. 入 300 μl Buffer P1 後震盪均勻後,加入 300 μl Buffer P2,上下搖勻 4~6 次後, 置於室溫下 5 分鐘。接著再加入 300 μl Buffer P3,混勻後放置冰上 5 分鐘後,離 心 10 分鐘 (最大速度,4℃),保留上清液。QIAGEN-tip 20 先以 1 ml Buffer QBT 通 過 column 之後,再加入前述之上清液,待其全部通過 column 後,以 2 ml Buffer QC 清洗 2 次,接著再以 0.8 ml Buffer QF 將 DNA elute 至 eppendorf 中,加入 0.56 ml isopropanol 後離心 30 分鐘 (13000 rpm,4℃)。之後,小心地將上清液抽掉,以 1 ml 70%酒精清洗,離心 10 分鐘 (10000 rpm,4℃),將上清液抽掉,剩餘的 DNA pellet 使其乾燥 5~10 分鐘後,以 50 μl 二次水將其溶解,保存於-20℃。. IV. 質體鑑定--以限制酶切割 EcoR I digestion: Component. Volume. Plasmid DNA. 15 μl. 10x NEBuffer EcoR I. 5 μl 0.5 μl. EcoR I (BioLabs). to 50 μl. ddH2O EcoR I + Hind III digestion: Component. Volume. Plasmid DNA. 15 μl. 10x NEBuffer EcoR I. 5 μl. EcoR I. 0.5 μl. Hind III. 0.5 μl. ddH2O. to 50 μl. 29.
(43) 第三章 材料與方法. 以 37℃水浴 24 小時後,每管加入 6 倍的 DNA loading dye 混合均勻,再以含有 0.26% 之 EtBr 之 1% Agarose 跑膠,並以 MultilmageTM Light Cabinet 分析影像並儲存。. V. 質體大量製備--使用 QIAGEN Plasmid Maxi Kit (QIAGEN) 將上述培養 overnight 之菌體離心 15 分鐘 (6000 g,4℃) 後,將上清液倒掉, 加入 10 ml Buffer P1 後震盪均勻後,加入 10 ml Buffer P2,上下搖勻 4~6 次後,置 於室溫下 5 分鐘。接著再加入 10 ml Buffer P3,混勻後放置冰上 20 分鐘後,離心 30 分鐘 (最大速度,4℃),保留上清液。QIAGEN-tip 500 先以 1 ml Buffer QBT 通過 column 之後,再加入前述之上清液,待其全部通過 column 後,以 2 ml Buffer QC 清洗 2 次,接著再以 0.8 ml Buffer QF 將 DNA elute 至 eppendorf 中,加入 0.56 ml 之 isopropanol 後離心 30 分鐘 (13000 rpm,4℃)。之後,小心地將上清液抽掉,以 1 ml 70%酒精清洗,離心 10 分鐘 (10000 rpm,4℃),將上清液抽掉,剩餘的 DNA pellet 使其乾燥 5~10 分鐘後,以 50 μl 二次水將其溶解,保存於-20℃。. 30.
(44) 第三章 材料與方法. 五、建立 HBP1 overexpresssion 與 knockdown 之細胞株(使用反轉錄病毒感染). I. 轉染至 phoenix 細胞 將 phoenix 細胞培養至 10 cm 培養皿中,待長滿後以 1:10 分盤並使其貼附後 備用。將 18 μl 的 FuGENE® 6 與 serum free medium 反應 5 分鐘後,接著加入 6 μ g 質體 DNA 反應 15 分鐘,將混合液慢慢滴入 phoenix 細胞中,放置 24 小時後更換 7 ml 一般的培養液,繼續培養 24 小時。. II. 感染目標細胞 將 7 ml 的培養液取出後,以 0.45 μm filter 過濾後,加入 8 μg/ml polybrene, 將培養液倒入欲感染之細胞中培養 5 小時。之後再以相同的步驟取得培養液並培養 3 小時後,換成一般的培養液培養。. III. 以 puromycin 篩選 待細胞長滿之後,使用含有 0.5 μg/ml puromycin 之培養液,以 1:10、1:200、 1:2000 與 1:20000 的稀釋倍數進行分盤。分盤後前 3 天須每天更換培養液,之後 則是按照一般細胞培養方式每 2~3 天更換一次培養液。. 篩選約 1 個多星期之後,若細胞長出一個個肉眼可見之 clone,即可進行挑選。 利用沾有 trypsin 的 cloning disc 覆蓋在單一個 clone 上約 1 分鐘,再將 cloning disc 放至含有培養液的 6 孔盤中培養,待細胞長滿後即可移至 10 cm dish 繼代培養,並 抽取 RNA 以做鑑定。. 31.
(45) 第三章 材料與方法. (六)、抽取 RNA--使用 RNeasy® Mini Kit (QIAGEN). 細胞先用 trypsin 刷下後,加入 10 ml PBS 離心 5 分鐘 (1000 rpm),將上清液抽 掉,加入 700 μl RLT buffer 與 7 μl β-mercaptoethanol 後混勻,使其通過 20 G 的 針頭至少 5 次,並加入 700 μl 70% 酒精後混勻。取 700 μl 至 RNeasy spin column (放在 2 ml collection tube 上) 後離心 15 秒 (10000 rpm),將下層的液體倒掉後,再 將剩下的 700 μl 至 RNeasy spin column 後離心 15 秒 (10000 rpm),將下層的液體 倒掉。加入 700 μl RW1,離心 15 秒 (10000 rpm) 後將下層的液體倒掉。加入 500 μl RPE,離心 15 秒 (10000 rpm),將下層液體倒掉後,再加入一次 RPE,離心 2 分鐘 (10000 rpm)。將 column 放到空的 2 ml collection tube 裡,離心 1 分鐘 (13000 rpm),接著換到 1.5 ml collection tube,加入 30 μl RNase-free water,離心 1 分鐘 (10000 rpm),再加入一次 30 μl RNase-free water,離心 1 分鐘 (10000 rpm) 後,即 可將 RNA 放到-20℃冰箱中保存。. 32.
(46) 第三章 材料與方法. 七、RT-PCR--使用 SuperScriptTMⅢ One-Step RT-PCR System with Platinum® Taq DNA Polymerase Kit (invitrogen) Reaction set up: Component. Volume 12.5 μl. 2x Reaction Mix Template RNA (0.5 μg). X μl. Sense primer (10 μM). 0.5 μl. Anti-sense primer (10 μM). 0.5 μl. SuperScriptTM III RT/Platinum® Taq Mix. 0.5 μl to 25 μl. DEPC-water Reaction protocol: Temperature. Time. Cycle. 30 min. 1. 2 min. 1. cDNA synthesis 55℃ Denaturation 94℃ PCR amplification 94℃. 15 s. 55℃. 40 s. 68℃. 45 s. 40. Final extension 68℃. 5 min. 4℃. ∞. 33. 1.
(47) 第三章 材料與方法. 將 RNA、primer 與試劑混合後,放入 PCR 專用之小管中,放入 iCycler,設定上述 之反應條件即可。反應完成後,每管加入 6 倍之 DNA loading dye,再以含有 0.26% 之 EtBr 之 1% Agarose 跑膠,並以 MultilmageTM Light Cabinet 分析影像並儲存。. 34.
(48) 第三章 材料與方法. 八、ROS 測定. 利用化學冷光分析儀來測定。先將 1×106 細胞種至培養皿中,經過 24 小時細胞 貼附之後,依實驗所需將細胞處理之後,加入 trypsin 反應 5 分鐘刷下並離心 3 分鐘, 將上清液抽掉,以 10 ml 之 PBS 清洗。接著加入 100 μl 含有 0.5 mM lucigenin 之 PBS 後,立即放入儀器的偵測槽,置入 photon detector 的密閉腔室中,啟動 chemiluminesence counter 後開始計數。最後數值以 relative light units (RLU)表示。. 35.
(49) 第三章 材料與方法. 九、蛋白質萃取. RIPA buffer: Component. Concentration. NaCl. 150mM. Tris buffer. 10mM. EDTA. 5mM. SDS. 0.1%. Triton X-100. 1%. Deoxycholate. 1%. 細胞以 PBS 清洗過後,加入 100μl RIPA buffer (含 protease inhibitors cocktail 與 phosphatase inhibitors cocktail),以細胞刮勺將細胞刮下,放至 eppendorf,置於冰上 30 分鐘。之後將此 cell lysate 以 15300 rpm 轉速離心 15 分鐘後,取其上清液,放到 -80℃冰箱中保存備用。. 36.
(50) 第三章 材料與方法. 十、蛋白質定量. Standard: Concentration. BSA. 二次水. 0mg/ml. 0μl. 20μl. 0.1mg/ml. 2μl. 18μl. 0.2mg/ml. 4μl. 16μl. 0.4mg/ml. 8μl. 12μl. 0.8mg/ml. 16μl. 4μl. 1mg/ml. 20μl. 0μl. 標準品 BSA 以上述濃度作標準曲線,先將 10μl 之 standard 與 sample 注入 96 孔盤後,加入稀釋之 dye reagent,以 595 波長測其吸光值後,將吸光值代入標準曲 線後,推算蛋白濃度。. 37.
(51) 第三章 材料與方法. 十一、Western blot. I. 鑄膠 4x Lower Tris: Component. Concentration. Tris-base (USBiological). 1.5M. SDS (MERCK). 0.4%. ddH2O. to 200ml. pH. 8.8. 4x Upper Tris: Component. Concentration. Tris-base (USBiological). 0.5M. SDS (MERCK). 0.4%. ddH2O. to 200ml. pH. 6.8. APS: Component. Concentration. Ammonium persulfate (J.T.Baker). 10%. ddH2O. to 10ml. 38.
(52) 第三章 材料與方法. 下膠: 將 30% Acrylamide、Lower Tris、APS、TEMED 與二次水以適當比例混勻之後, 倒入玻璃片凹槽中,再以 100%酒精覆蓋。. 上膠: 待下膠凝固後,將 30% Acrylamide、Upper Tris、APS、TEMED 與二次水以適 當比例混和後,倒在凝固的下膠上,並輕輕插入齒梳(comb)。. II. 跑膠 10x Running buffer: Component. Concentration. Tris-base. 0.25M. glycine. 1.9M. SDS. 1%. ddH2O. to 1000ml. pH. 8.6. 待上膠凝固後,輕輕地拔出齒梳,將膠片移至電泳槽中,倒入適量的 1x running buffer。將製備好的蛋白樣品 loading 到膠片的 well 中,並 loading 適量的 marker。 將電泳槽蓋子上 (紅對紅,黑對黑),以 90 伏特電壓跑膠。. III. 轉印 10x Transfer buffer: 39.
(53) 第三章 材料與方法. Component. Concentration. Tris-base. 0.25M. glycine. 1.9M. ddH2O. to 1000ml. pH. 8.3. 將轉印夾打開,依序放上海棉墊、濾紙、取下的膠片與浸濕的 PVDF 膜後,確 定其緊密貼合後,將轉印夾慢慢地合起來,放到轉印槽中,倒入 1x transfer buffer, 將蓋子蓋上,以 400 mA 電流轉 1 小時。. IV. 免疫染色 10x Tris-base buffer saline: Component. Concentration 0.2M. Tris-base. 80g. NaCl ddH2O. to 1000ml. pH. 7.6. TBS/T: Component. Concentration. 10x TBS. 100ml. Tween 20. 0.1%. ddH2O. 900ml. 40.
(54) 第三章 材料與方法. 將轉印完成的 PVDF 膜以 5%脫脂牛奶搖晃約 1 小時進行 blocking,以 TBS/T 清洗 5 分鐘三次後,加入適當稀釋倍數之一級抗體反應 overnight。將一級抗體洗掉 後,加入稀釋之二級抗體反應 1 小時,清洗 5 分鐘 3 次,以感光試劑 ECL 進行呈色 反應。. 41.
(55) 第四章 結果. 第四章、結果. 一、HBP1 調控 EGFR 及其下游 Akt 的活性. I. 細胞內 HBP1 表現上升可抑制 EGFR 及其下游 Akt 活性 實驗所使用之 MDA-MB-231 乳癌細胞是具有 EGFR 過度表現的特性,因此我 們利用它來當作試驗的細胞株。為了觀察 HBP1 在細胞內的表現對 EGFR 活性的影 響,我們將 HBP1 過度表現的質體利用反轉錄病毒感染的方式帶進細胞當中,建立 HBP1 過度表現之穩定細胞株,使此細胞株之 HBP1 的表現多於一般正常的細胞, 並以轉染空的 pBabe 載體之細胞來當作控制組。圖 4-1 為利用 RT-PCR 來觀察 pBabe 控制組與 HBP1 過度表現細胞之 HBP1 mRNA 表現。在 HPB1 過度表現細胞中之 HBP1 mRNA 表現確實顯著的多於控制組的細胞。接著,我們分別給予控制組與 HBP1 過度表現的細胞 10ng/ml 之 EGF 於 0 分鐘、5 分鐘、15 分鐘、30 分鐘、1 小 時與 2 小時或是 0ng/ml、5ng/ml、10ng/ml、30ng/ml、50ng/ml 與 70ng/ml 之 EGF 刺激 20 分鐘後,以 Western blot 觀察 EGFR、pEGFR、Akt 與 pAkt 的表現。圖 4-2 為時間變化的結果,由圖中可看出無論在那個時間點之下,HBP1 過度表現細胞之 pEGFR 表現皆低於 pBabe 控制組的細胞;而在圖 4-3 中可看出無論在那個濃度下, HBP1 過度表現細胞之 pEGFR 表現也都低於 pBabe 控制組的細胞,顯示細胞中 HBP1 過度表現確實可抑制 EGFR 的活性。在下游 Akt 活性方面也有和 EGFR 相對應的結 果,由圖 4-4 與 4-5 可知。綜合以上結果顯示,細胞內 HBP1 過度表現確實可抑制 EGFR 與其下游 Akt 的活性。. 42.
(56) 第四章 結果. II. 細胞內 HBP1 表現降低時會增強 EGFR 與其下游 Akt 之活性 我們在 HBP1 過度表現的細胞中已觀察到具有抑制 EGFR 活性的作用,相對的, 我們則是將細胞中 HBP1 knockdown 之後來觀察 EGFR 活性是否有顯著的改變。我 們利用 RNA 干擾術將細胞中之 HBP1 knockdown 之後,以 RT-PCR 來觀察 mRNA 的表現,以空的 pSR 載體轉染之細胞當作控制組。圖 4-6 為 pSR 控制組與 HBP1 knokcdown 細胞之 HBP1 mRNA 表現。在 HPB1 knockdown 細胞中之 HBP1 mRNA 表現確實顯著的低於控制組的細胞。接著,我們分別給予控制組與 HBP1 過度表現 的細胞 10ng/ml 之 EGF 於 0 分鐘、5 分鐘、15 分鐘、30 分鐘、1 小時與 2 小時或是 0ng/ml、5ng/ml、10ng/ml、30ng/ml、50ng/ml 與 70ng/ml 之 EGF 刺激 20 分鐘後, 以 Western blot 觀察 EGFR、pEGFR、Akt 與 pAkt 的表現。圖 4-7 為以 EGF 刺激不 同時間後之 EGFR 的活性,結果顯示無論在那個時間點之下,HBP1 knockdown 細 胞之 pEGFR 表現皆高於 pSR 控制組的細胞。而在濃度的改變下也有相同的結果, 無論在那個濃度下,HBP1 knockdown 的細胞 pEGFR 也都顯著的高於控制組,見圖 4-8。圖 4-9 與 4-10 則是 EGFR 下游 Akt 活性的結果,也與 EGFR 結果一致,在 HBP1 knockdown 的細胞中也顯著的提升。綜合以上結果得知,當細胞內 HBP1 表現降低 時,EGFR 的活性與控制組相比有顯著的增加。. 43.
(57) 第四章 結果. 二、HBP1 調控細胞內 EGFR 活化所伴隨的 ROS 之生合成. 因為 HBP1 可藉由調控細胞內 NADPH 氧化酶次單位的活性,進而影響 superoxide 的生成。superoxide 的偵測是利用 lucigenin 試劑,lucigenin 會與 superoxide 發出冷光,我們再以冷光偵測儀來做分析,結果使用 relative light units (RLU)來呈 現。首先,我們將 HBP1 過度表現的質體轉染至細胞內,提升細胞內的 HBP1,並 與控制組相比 superoxide 的生成是否有所變化。圖 4-11 為給予 EGF 誘導後 HBP1 過度表現的細胞內之 superoxide 變化,隨著 EGF 給予的時間增加,與控制組相比有 較低的 superoxide 生成量;相反地,我們以 RNAi 降低細胞內 HBP1,觀察 EGF 所 刺激之活性氧化物的生成有何影響。圖 4-12 HBP1 knockdown 細胞的結果,隨著 EGF 給予的時間增加,超氧化物的生成量顯著的多於控制組。由此結果可知,增加細胞 內 HBP1 的表現可抑制 superoxide 的生成;反之,在 HBP1 表現減少時,EGF 刺激 所伴隨的 superoxide 生成則會增加。. 44.
(58) 第四章 結果. 三、細胞內 ROS 生成量可調控 EGFR 及其下游的活性. 由以上結果得知,HBP1 對 EGFR 的活性與 EGF 所刺激之 ROS 的生成有抑制 的效果,因此我們想要觀察細胞內的 ROS 的濃度是否會影響 EGFR 與其下游的活 性。我們使用已知的 NADPH 氧化酶抑制劑 DPI 與自由基清除劑 NAC 來處理細胞, 減少細胞內 ROS 的濃度,並以 Western blot 觀察 EGFR、pEGFR、Akt 與 pAkt 的蛋 白表現。在使用 DPI 後可觀察到 EGFR 與下游 Akt 的活性有顯著的被抑制,圖 4-13 與 4-14 可見 pEGFR 與 pAkt 表現有顯著的下降;而以 NAC 處理後的細胞也有相同 的結果,EGFR 與 Akt 的活性有顯著的被抑制,見圖 4-15 與 4-16。由此結果可知, 降低細胞內 ROS 濃度增加時可抑制 EGFR 與下游 Akt 之活性表現。. 45.
(59) 第四章 結果. 四、HBP1 調控 EGFR 及其下游的活性是藉由抑制 NADPH 氧化酶次單位 p47phox 的基因轉錄. 由於 NADPH 氧化酶次單位 p47phox 為已知的 HBP1 調控因子,若是 p47phox 的改 變可影響 EGFR 與 Akt 的活性,便可間接證實 HBP1 對 EGFR 的調控。因此我們利 用 p47phox siRNA 來模擬 HBP1 抑制 p47phox 基因轉錄的情形。我們將 p47phox 之 siRNA 轉染至細胞中,並以 GFP 為 transfection marker,觀察 p47phox mRNA 的表現。圖 4-17 為 RT-PCR 的結果,可看出 p47phox siRNA 的細胞中,mRNA 的表現量顯著的低於控 制組。接著,在 p47 phox knockdown 與控制組細胞中分別以 EGF 刺激之後,以 Western blot 來觀察 EGFR、pEGFR、Akt 與 pAkt 的蛋白表現。圖 4-18 顯示,在 EGF 的刺 激下,p47phox siRNA 的細胞中,pEGFR 的表現顯著低於控制組,顯示 EGFR 的活性 有被抑制的情形,圖 4-19 之 Akt 活性亦有相對應的結果。由此可知,當 p47phox 受到 抑制之後的對 EGFR 的活性確實有顯著的影響,也間接證實了可抑制 p47phox 轉錄的 HBP1 確實經由對氧化還原平衡的調控來抑制 EGFR 的活性。. 46.
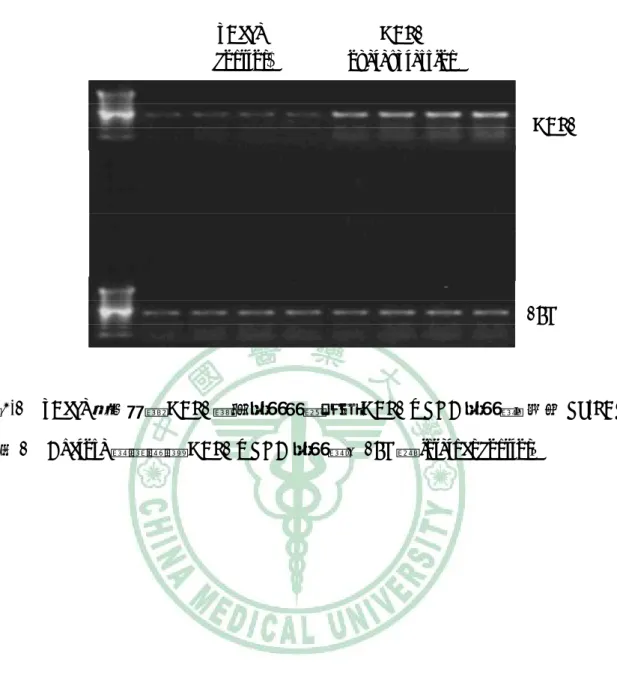
(60) 第四章 結果. pBabe (control). HBP1 overexpression HBP1. 18S. 圖 4-1. pBabe 控制組與 HBP1 過度表現細胞中之 HBP1 mRNA 表現量。使用 RT-PCR. 並以 1% Agarose 跑膠觀察 HBP1 mRNA 表現量,18S 為 internal control。. 47.
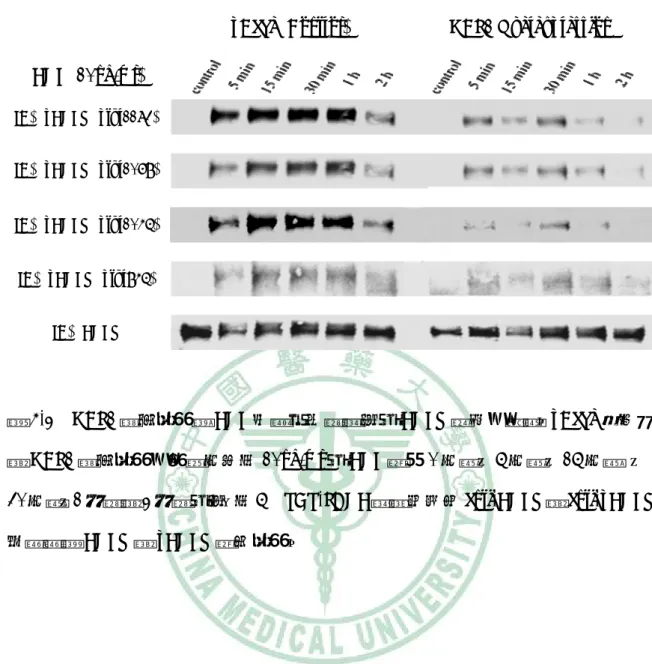
(61) 第四章 結果. pBabe (Control). HBP1 Overexpression. EGF (10ng/ml) IB:pEGFR (ptyr1173) IB:pEGFR (ptyr1068). IB:pEGFR (ptyr1045) IB:pEGFR (ptyr845) IB:EGFR. 圖 4-2 HBP1 過度表現對 EGF 刺激不同時間後之 EGFR 活性的影響。pBabe 控制組 與 HBP1 過度表現的細胞分別以 10ng/ml 之 EGF 處理 0 分鐘、5 分鐘、15 分鐘 、 30 分鐘、1 小時與 2 小時之後,以 5% SDS-PAGE 跑膠並使用 anti-EGFR 與 anti-pEGFR 抗體觀察 EGFR 與 pEGFR 蛋白表現。. 48.
(62) 第四章 結果. pBabe (Control). HBP1 Overexpression. EGF (20min) IB:pEGFR (ptyr1173) IB:pEGFR (ptyr1068). IB:pEGFR (ptyr1045) IB:pEGFR (ptyr845) IB:EGFR. 圖 4-3 HBP1 過度表現對不同濃度的 EGF 誘發之 EGFR 活性的影響。pBabe 控制組 與 HBP1 過度表現的細胞分別以 0ng/ml、5ng/ml、10ng/ml、30ng/ml、50ng/ml 與 70ng/ml 濃度的 EGF 處理 20 分鐘之後,再以 5% SDS-PAGE 跑膠並使用 anti-EGFR 與 anti-pEGFR 抗體觀察 EGFR 與 pEGFR 蛋白表現。. 49.
(63) 第四章 結果. pBabe (Control). HBP1 Overexpression. EGF (10ng/ml) IB:pAkt (pser473) IB:Akt. 圖 4-4 HBP1 過度表現對 EGF 刺激不同時間後之 Akt 活性的影響。pBabe 控制組與 HBP1 過度表現的細胞分別以 10ng/ml 之 EGF 處理 0 分鐘、5 分鐘、15 分鐘 、30 分鐘、1 小時與 2 小時之後,以 10% SDS-PAGE 跑膠並使用 anti-Akt 與 anti-pAkt 抗 體觀察 Akt 與 pAkt 蛋白表現。. 50.
(64) 第四章 結果. pBabe (Control). HBP1 Overexpression. EGF (20min) IB:pAkt (pser473). IB:Akt. 圖 4-5 HBP1 過度表現對不同濃度的 EGF 誘發之 Akt 活性的影響。pBabe 控制組與 HBP1 過度表現的細胞分別以 0ng/ml、5ng/ml、10ng/ml、30ng/ml、50ng/ml 與 70ng/ml 濃度的 EGF 處理 20 分鐘之後,再以 10% SDS-PAGE 跑膠並使用 anti-Akt 與 anti-pAkt 抗體觀察 Akt 與 pAkt 蛋白表現。. 51.
(65) 第四章 結果. pSR-HBP1i. pSR. HBP1. 18S. 圖 4-6. pSR 控制組與 HBP1 knockdown 細胞中之 HBP1 mRNA 表現量。使用 RT-PCR. 並以 1% Agarose 跑膠觀察 HBP1 mRNA 表現量,18S 為 internal control。. 52.
(66) 第四章 結果. pSR (Control). HBP1 shRNA (HBP1 Knockdown). EGF (10ng/ml) IB:pEGFR (ptyr1173) IB:pEGFR (ptyr1068) IB:pEGFR (ptyr1045). IB:pEGFR (ptyr845). IB:EGFR. 圖 4-7 HBP1 表現降低時對 EGF 刺激不同時間後之 EGFR 活性的影響。pSR 控制 組與 HBP1 knockdown 的細胞分別以 10ng/ml 之 EGF 處理 0 分鐘、5 分鐘、15 分鐘 、 30 分鐘、1 小時與 2 小時之後,以 5% SDS-PAGE 跑膠並使用 anti-EGFR 與 anti-pEGFR 抗體觀察 EGFR 與 pEGFR 蛋白表現。. 53.
(67) 第四章 結果. pSR (Control). HBP1 shRNA (HBP1 Knockdown). EGF (20min) IB:pEGFR (ptyr1173). IB:pEGFR (ptyr1068) IB:pEGFR (ptyr1045) IB:pEGFR (ptyr845) IB:EGFR. 圖 4-8 HBP1 表現降低時對不同濃度的 EGF 刺激之 EGFR 活性的影響。pSR 控制 組與 HBP1 knockdown 的細胞分別以 0ng/ml、5ng/ml、10ng/ml、30ng/ml、50ng/ml 與 70ng/ml 濃度的 EGF 處理 20 分鐘之後,再以 5% SDS-PAGE 跑膠並使用 anti-EGFR 與 anti-pEGFR 抗體觀察 EGFR 與 pEGFR 蛋白表現。. 54.
(68) 第四章 結果. pSR (Control). HBP1 shRNA (HBP1 Knockdown). EGF (10ng/ml) IB:pAkt (pser473) IB:Akt. 圖 4-9 HBP1 表現降低時對 EGF 刺激不同時間之 Akt 活性的影響。pSR 控制組與 HBP1 knockdown 的細胞分別以 0ng/ml、5ng/ml、10ng/ml、30ng/ml、50ng/ml 與 70ng/ml 的 EGF 處理 20 分鐘後,再以 10% SDS-PAGE 跑膠並使用 anti-Akt 與 anti-pAkt 抗體 觀察 Akt 與 pAkt 蛋白表現。. 55.
(69) 第四章 結果. pSR (Control). HBP1 shRNA (HBP1 Knockdown). EGF (20min) IB:pAkt (pser473) IB:Akt. 圖 4-10 HBP1 表現降低時對不同濃度的 EGF 刺激之 Akt 活性的影響。pSR 控制組 與 HBP1 knockdown 的細胞分別以 0ng/ml、5ng/ml、10ng/ml、30ng/ml、50ng/ml 與 70ng/ml 的 EGF 處理 20 分鐘後,再以 10% SDS-PAGE 跑膠並使用 anti-Akt 與 anti-pAkt 抗體觀察 Akt 與 pAkt 蛋白表現。. 56.
(70) 第四章 結果. EGF (10ng/ml). 圖 4-11. HBP1 表現增加對細胞內 EGF 誘發之 ROS 的生成之影響。細胞分別以. 10ng/ml 的 EGF 處理 0 分鐘、5 分鐘 、15 分鐘與 30 分鐘後,以發光試劑 lucigenin 並使用化學冷光偵測儀分析 superoxide 的生成量。. 57.
(71) 第四章 結果. EGF (10ng/ml). 圖 4-12 HBP1 表現降低時對細胞內 EGF 誘發之 ROS 生成之影響。細胞分別以 10ng/ml 的 EGF 處理 0 分鐘、5 分鐘 、15 分鐘與 30 分鐘後,以發光試劑 lucigenin 並使用化學冷光偵測儀分析 superoxide 的生成量。. 58.
(72) 第四章 結果. DPI (μM) / 18h. -. -. 5. 10. 30. 50. EGF (30ng/ml)/15min. -. +. +. +. +. +. IB:pEGFR (ptyr1173). IB:pEGFR (ptyr1068) IB:pEGFR (ptyr1045). IB:pEGFR (ptyr845) IB:EGFR. 圖 4-13. 以 DPI 降低細胞中 ROS 的生成量對 EGFR 活性的影響。細胞先以 0μM、. 5μM、10μM、30μM 與 50μM 的 DPI 處理 18 小時後,再以 30ng/ml 的 EGF 刺激 15 分. 鐘後,利用 5% SDS-PAGE 跑膠並以 anti-EGFR 與 anti-pEGFR 抗體觀察 EGFR 與 pEGFR 蛋白的表現。. 59.
(73) 第四章 結果. DPI (μM) / 18h. -. -. 5. 10. 30. 50. EGF (30ng/ml) / 15min. -. +. +. +. +. +. IB:pAkt (pser 473) IB:Akt. 圖 4-14. 以 DPI 降低細胞中 ROS 的生成量對 Akt 活性的影響。細胞先以 0μM、5μM、. 10μM、30μM 與 50μM 的 DPI 處理 18 小時後,再以 30ng/ml 的 EGF 刺激 15 分鐘後,. 利用 10% SDS-PAGE 跑膠並以 anti-Akt 與 anti-pAkt 抗體觀察 Akt 與 pAkt 蛋白的表 現。 .. 60.
(74) 第四章 結果. NAC (15mM) / min. -. -. 5. 10. 20. 30. EGF (40ng/ml) / 15min. -. +. +. +. +. +. IB:pEGFR (ptyr 1086). IB:EGFR. 圖 4-15. 以 NAC 降低細胞中 ROS 的生成量對 EGFR 活性的影響。細胞先以 15mM. 的 NAC 處理 0 分鐘、5 分鐘、10 分鐘、20 分鐘與 30 分鐘後,再以 40ng/ml 的 EGF 刺激 15 分鐘,利用 5% SDS-PAGE 跑膠並以 anti-EGFR 與 anti-pEGFR 抗體觀察 EGFR 與 pEGFR 蛋白的表現。. 61.
(75) 第四章 結果. NAC (min) / 15mM. -. -. 5. 10. 20. 30. EGF (40ng/ml) / 15min. -. +. +. +. +. +. pAkt (pser 473). Akt. 圖 4-16. 以 NAC 降低細胞中 ROS 的生成量對 Akt 活性的影響。細胞先以 15mM 的. NAC 處理 0 分鐘、5 分鐘、10 分鐘、20 分鐘與 30 分鐘後,再以 40ng/ml 的 EGF 刺 激 15 分鐘,利用 10% SDS-PAGE 跑膠並以 anti-Akt 與 anti-pAkt 抗體觀察 Akt 與 pAkt 蛋白的表現。. 62.
(76) 第四章 結果. p47 phox 18S. 圖 4-17. pBabe 控制組與 HBP1 過度表現細胞中之 HBP1 mRNA 表現量。使用. RT-PCR 並以 1% Agarose 跑膠觀察 HBP1 mRNA 表現量,18S 為 internal control。. 63.
(77) 第四章 結果. Control EGF (ng/ml) / 20min. 0. 5. 10. p47phox-siRNA. GFP-siRNA 50. 0. 5. 10. 50. 0. 5. 10. 50. IB:pEGFR (ptyr1173). IB:pEGFR (ptyr1068). IB:pEGFR (ptyr1045) IB:pEGFR (ptyr845). IB:EGFR. 圖 4-18 模擬 HBP1 調控 p47phox 表現來觀察 EGFR 活性的變化。細胞分別以 0ng/ml、 5ng/ml、10ng/ml 與 50ng/ml 之 EGF 處理 20 分鐘後,利用 5% SDS-PAGE 跑膠並以 anti-EGFR 與 anti-pEGFR 抗體觀察 EGFR 與 pEGFR 蛋白的表現。. 64.
(78) 第四章 結果. Control EGF (ng/ml) / 20min. 0. 5. 10. p47phox-siRNA. GFP-siRNA 50. 0. 5. 10. 50. 0. 5. 10. 50. IB:pAkt (pser 473) IB:Akt. 圖 4-19 模擬 HBP1 調控 p47phox 表現來觀察 Akt 活性的變化。細胞分別以 0ng/ml、 5ng/ml、10ng/ml 與 50ng/ml 之 EGF 處理 20 分鐘後,利用 10% SDS-PAGE 跑膠並以 anti-Akt 與 anti-pAkt 抗體觀察 Akt 與 pAkt 蛋白的表現。. 65.
數據

+7


Outline
相關文件
the lymphocyte function-associated antigen 1, or LFA-1, was so named because antibodies recognizing this structure interfere with lymphocyte cell adhesion events and
此為一項針對轉移性或復發性乳癌患者比較 NK105 與 Paclitaxel 的多國第 III 期臨床詴驗研究。此次修 正主要原因為根據 2016 年 1 月 23
10 SF12259A 黃文豊 一項隨機分配、開放性、多中心臨床 試驗,對於先前未接受過治療的 CD20 表面抗原陽性之瀰漫性大型 B 細胞淋 巴瘤或 CD20 表面抗原陽性之濾泡性
IRB 編號 SC19232B#9 【CIRB 副審】 計畫主持人 楊宗穎 計畫名稱.
SF11310 張基晟 比較 Ipilimumab 加上 Etoposide/Platinum 與 Etoposide/Platinum
Rapiacta 因不經肝代謝,故透過 CYP 機轉與其他藥物發生 交互作用之可能性應該很低,就目前所知的排除途徑以及從 體外試驗可推知 Rapiacta 並不會誘導或抑制 CYP 450。 1)
第一期至第三期 實體癌,經標準 治療無效;癌症 別:上皮性卵巢 癌、肺癌、肝 癌、乳癌、大腸 直腸癌、胰臟癌. 長聖國際生
眼睛是心靈之窗,心靈是眼神之源。在眼球後方 感光靈敏的角膜含有 1.37 億個細胞,將收到的信 息傳送至腦部。這些感光細胞,在任何時間均可 同時處理
