(-)-Epigallocatechin-3-gallate 藉由抑制基本表現及第六白介素誘導之訊息傳遞及轉錄因子3路徑誘發頭頸部腫瘤細胞凋亡; (-)-Epigallocatechin-3-gallate Induces Apoptosis via the Repression of Constitutive and Interleukin-6 Inducible Signal Transducer and Activators of Transcription 3 Pathway in Head an
77
0
0
全文
(2)
(3) 致謝 首先,誠摯的感謝指導教授林慧怡博士及魏宗德博士,兩位老師 悉心的教導使我得以一窺生技藥學領域的深奧,不時的討論並指點我 正確的方向,使我在碩班的這兩年中獲益匪淺。老師們對學問的嚴謹 更是我輩學習的典範。 本論文的完成另外亦得感謝貴儀中心的戴如淳組長大力協助,以 及貴儀中心魏秀涓小姐的支持。因為有你們的體諒及幫忙,使得本論 文能夠更完整而嚴謹。 兩年裡的日子,實驗室裡共同的生活點滴,學術上的討論、言不 及義的閒扯、趕實驗進度的革命情感,感激眾學長姐、同學、學弟妹 的共同砥礪,你/妳們的陪伴讓兩年的研究生活變得絢麗多彩。 感謝高銘欽老師總是在我對實驗灰心的時候鼓勵我,感謝薇至學 姐、思樺學姐、玲玲學姐、傑文學長、柏村學長們不厭其煩的指出我 研究中的缺失,且總能在我迷惘時為我解惑,也感謝怡婷、秀學、岑 容、瓊之、佳霓同學的幫忙,恭喜我們順利走過這兩年。實驗室的宜 禎、怡君、怡鈴、宜靜學妹、嘉群、尚杰學弟們當然也不能忘記,妳 們的熱心協助我銘感在心。 最後,謹以此文獻給我摯愛的雙親。.
(4) 目錄 目錄………………………………………………………………….. I. 圖目錄……………………………………………………………….. III. 縮寫表……………………………………………………………….. IV. 中文摘要…………………………………………………………….. VI. 英文摘要…………………………………………………………….. VIII. 第一章 序論……………………………………………………….... 1. 1.1 頭頸部癌症(Head and neck squamous cell carcinoma)之 介紹……………………………........................................... 1. 1.2 訊息傳遞及轉錄因子 3(Signal transducer and activator of transcription 3)之介紹…………………………………….. 1. 1.3 Signal transducer and activator of transcription 3(STAT3) 與口腔癌之關係…………………………………………... 3. 1.4 第六白介素(Interleukin-6)誘導之 STAT3 路徑………..... 4. 1.5 第六白介素與頭頸部癌症之關係………………..……….. 4. 1.6 茶的介紹………………………………………………….... 5. 1.7 綠茶與抗癌之簡介……………………………………….... 8. 1.8 表沒食子兒茶素沒食子酸脂(EGCG)之抗癌背景介紹…... 10. 1.9 實驗動機………………………………………………….... 11. 第二章 材料與方法……………………………………………….. 12. 2.1 實驗材料………………………………………………….... 12. 2.2 細胞培養………………………………………………….... 13. 2.2.1 細胞株………………………………………………... 13. 2.2.2 細胞培養與繼代……………………………………... 14. 2.3 細胞存活率分析………………………………………….... 14. I.
(5) 2.4 流式細胞儀分析 ( Flow cytometry )…………………….... 15. 2.5 細胞總蛋白收集與定量………………………………….... 16. 2.6 蛋白質之電泳分析(SDS-PAGE)…………………………. 17. 2.7 西方點墨法 ( Western Blotting ) …………………………. 18. 2.8 免疫螢光染色( Immuocytochemistry )……………………. 19. 2.9 統計分析………………………………………………….... 20. 第三章 結果……………………………………………………….. 21. 3.1 EGCG 抑制頭頸部癌細胞的增生……………………….... 21. 3.2 EGCG 促使頭頸部癌細胞細胞週期停滯於 G1 期並誘導 細胞凋亡……………………………………………………. 22. 3.3 EGCG 促使細胞週期停滯 G1 期、細胞凋亡之相關蛋白 的表現………………………………………………………. 22. 3.4 EGCG 抑制頭頸部癌細胞中 STAT3 蛋白的磷酸化….... 23. 3.5 EGCG 抑制 STAT3 蛋白之下游蛋白的表現……….……. 25. 3.6 EGCG 抑制頭頸部癌細胞中 Interleukin-6 誘導 STAT3 蛋白的磷酸化………………………………………….…. 3.7 EGCG 抑制由 Interleukin-6 所引發的細胞增生………. 25 26. 第四章 討論……………………………………………………….. 28. 第五章 參考文獻………………………………………………….. 35. 附圖………………………………………………………………….. 53. 附錄………………………………………………………………….. 62. II.
(6) 圖目錄 Figure 1. Effect of EGCG on HNSCC cell line……..………………………. 53. Figure 2. Effect of EGCG on cell cycle progression and apoptosis……….. 54 Figure 3. Effect of EGCG on cell cycle proteins and apoptosis related proteins……………………………………………………….…….. 55. Figure 4. Effect of EGCG on phosphorylated STAT3………………..…..… 56 Figure 5. Western blot analyses for STAT3 downstream proteins……...… 59 Figure 6. Interleukin-6 induces STAT3 phosphorylation in human HNSCC cell, and EGCG inhibits it………………..…………. 59. Figure 7. No effect of EGCG on phosphorylated STAT3 in HSC3 cell line. 61. III.
(7) 縮寫表 5GG. Penta-O-galloyl-β-D-glucose. Bcl-2. B cell lymphoma protein 2. BSA. Bovine serum albumin. Caspase. Cysteine dependent aspartate specific protease. DAPI. 4,6-diamidino-2-phenylindole. DMSO. Dimethyl sulfoxide. ECL. Enhanced-chemiluminescence. EDTA. Ethylenediaminetetraacetic acid. EGCG. (-)-Epigallocatechin gallate. ELISA reader. Enzyme-linked immunosorbent assay reader. FBS. Fetal bovine serum. HNSCC. Head and Neck Squamous Cell. IL-6. Interleukin-6. Jak. Janus kinases. LR1. Laminin Receptor 1. MTT. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl. Carcinoma. tetrazolium. bromide OSCC. Oral squamous cell carcinoma. PARP. Poly ADP-ribose polymerase. PBS. Phosphoate-buffered saline. PI. Propidium iodide. P-STAT3(705). PhosphoTyr705-STAT3. P-STAT3(727). PhosphoSer727-STAT3. rpm. Revolutions per minute. SDS. Sodium dodecyl sulfate. SDS-PAGE. Sodium. dodecyl. electrophoresis IV. sulfate-polyacrylamide. gel.
(8) STAT3. Singal Transducer and Activators of Transcription 3 Pathway. TF1. Theaflavin. TF2. Theaflavin-3-gallate. TF3. Theaflavin-3,3’-digallate. Tris. Tris(hydroxymethyl) aminomethane. V.
(9) 中文摘要 表 沒 食 子 兒 茶 素 沒 食 子 酸 脂 ( (-)-epigallocatechin-3-gallate (EGCG))已被發現有許多益處,如:化學防治活性、抗癌活性與 抗氧化活性等,但對於頭頸部癌細胞(HNSCC)的影響還未被清楚 的研究。從多種黃酮類、多酚類的天然化合物中,我們發現 EGCG 明顯的抑制頭頸部癌細胞的生長。 根據研究指出癌細胞中持續活化的 STAT3 和癌細胞的存活、增 生是有相關性的。且在眾多癌症病患中,大於 90%比例的口腔癌病患 中可以發現 STAT3 被過度活化的現象,此情形在其他的癌症病患中 卻非如此。使我們想探究 EGCG 對頭頸部癌細胞株的抑制活性,是 否跟 STAT3 有所關聯? 在我們使用四株頭頸部癌細胞株(SAS、 Cal-27、Ca9-22、HSC3) ,進一步的研究 EGCG 對於頭頸部癌細胞 中 STAT3 訊息傳遞路徑的影響,發現 EGCG 抑制 STAT3 磷酸化的效 果與給藥劑量、給藥時間有著依存性的關係;在細胞層面的觀察裡, STAT3 磷酸化表現量逐漸下降; 在處理 EGCG 後,STAT3 明顯的移 動至細胞核外;STAT3 路徑下游蛋白表現量也下降。 interleukin-6 (IL-6)可藉由磷酸化訊息蛋白 singal transducer and activator 3 (STAT3)而促進腫瘤細胞的存活與生長。頭頸部癌細胞株 會自體分泌 IL-6 刺激自身的 STAT3 活化。我們發現 EGCG 可以消除 VI.
(10) IL-6 在頭頸部癌細胞中誘導 STAT3 活化的能力,更進ㄧ步的抑制 IL-6 所誘導的細胞增生現象。在與 Apigenin、Curcumin、AG490 比較抑制 IL-6 所誘導的細胞增生中,EGCG 也是最有效的抑制者。 總結而論,我們實驗的結果強烈的指出 EGCG 是一個有潛力去 抑制 IL-6 誘導 STAT3 磷酸化的抑制劑,而此機轉可反應出 EGCG 抑 制口腔癌細胞增生的原因。. VII.
(11) 英文摘要 Many. beneficial. properties. (-)-epigallocatechin-3-gallate. (EGCG),. have. been. including. attributed. to. chemopreventive,. anticarcinogenic, and antioxidant active. The antitumor effect of green tea compound EGCG have not been studied clearly in head and neck squamous cell carcinoma cells. Previous study indicated that the constitutive activation of singal-transducer-and-activator-3 (STAT3) were associated with the proliferation of HNSCC cells. Numerous reports suggest that interleukin-6 (IL-6) promotes survival and proliferation of tumor cells through the phosphorylation of STAT3. Constitutive of STAT3 in HNSCC was relative with its proliferation and survival. Thus, compound that can suppress STAT3 activation have potential for the treatment of HNSCC. Therefore, we examined in detail the molecular effect of EGCG on four HNSCC cell lines, SAS, Cal-27, Ca9-22 and HSC3. In the beginning, we found the great inhibition on HNSCC cells of EGCG. Further, we focused the effect of EGCG on STAT3 signal pathway. In this study, the 70% lethal dose (IC70) of EGCG for three cell lines, SAS, Cal-27 and Ca9-22, was 20 μM, and EGCG inhibited STAT3 phosphorylation in a dose- and time- dependent manner. Nuclear translocation of STAT3 was VIII.
(12) also inhibited by EGCG. Besides inhibiting constitutive expression, EGCG also abrogated the IL-6-induced activation of STAT3 and further inhibited IL-6-induced proliferation on HNSCC cell. Compare with Apigenin, Curcumin and AG490, EGCG was more effective inhibitor of IL-6-induced proliferation on HNSCC cells. Overall, our results strongly suggest that EGCG is a potent inhibitor of constitutive and IL-6-induced STAT3 phosphorylation. This mechanism may be at least partially responsible for EGCG’s ability to suppress proliferation of HNSCC cells. Taken together, these finding suggest that this naturally occurring compound may be useful, in the chemoprevention and/or treatment of HNSCC.. IX.
(13) 第一章、序論 1.1 頭頸部癌症(Head and neck squamous cell carcinoma)之介紹. 頭頸部癌症(Head and neck squamous cell carcinoma, HNSCC) 或其他人稱之口腔上皮細胞癌(Oral squamous cell carcinoma, OSCC) 是全球第八常見的癌症 1,2。在台灣,癌症死因的排名中,頭頸部癌排 名第六;台灣每年每壹百萬人中約有兩千人死於口腔癌。根據統計, 國人吃檳榔、抽煙、酗酒的習慣和罹患口腔癌的機率有著高度的關聯 性. 3,4,5. 。但要戒除這些不良的習慣是困難的。也是為什麼即使早期發. 現口腔癌,加以治療,仍有一半以上的口腔癌病患其病程會繼續惡化 6. ,導致 HNSCC 末期的存活率仍然很不理想 3。因此,尋找治療藥物. 是迫切需要的。. 1.2 訊息傳遞及轉錄因子 3 (Signal transducer and activator of transcription 3)之介紹. Signal transducer and activator of transcription 3 (STAT3)是許多 生長因子與細胞激酶(cytokine)的重要訊息傳遞分子 7。Jak/Stat 訊息 傳導路徑是許多 cytokines 和生長因子在接受到訊息後,結合細胞核 1.
(14) 中特定的基因,以傳達出訊息 8。以下介紹 STAT3 在細胞中是如何傳 遞訊息的: 當細胞膜上的接受器皆受到 cytokines 或生長因子的訊息時, Jak/tyrosine kinase 2( JAK family protein kinases) 將啟動磷酸化,接著 磷酸化細胞質中的 STAT3。被磷酸化的 STAT3 會組合成 dimer,再進 入細胞核與特定的 DNA 片段結合,並轉錄這些 DNA 片段,使細胞 9. 製造出所要的蛋白 。所以,平常不活化的 STAT3 是存在於細胞質中 的,當有訊息要傳遞時,被磷酸化的 STAT3 才能進入細胞核中進行 轉錄的動作。這是 STAT3 很特別的機制,也有一些學者致力於此機 制失常的研究 10。 STAT3 有 兩 個 可 磷 酸 化 的 位 置 , 一 個 是 位 於 705 序 列 的 Tyrosine,另一個則是位於 727 序列的 Serine。不同的 cytokines 或生 長因子所傳遞的訊息,使 STAT3 磷酸化於不一樣的位置。目前的研 究發現,刺激 STAT3 磷酸化於 705 序列 Tyrosine 位置的 cytokine 為 Interleukin-611;而刺激 STAT3 磷酸化於 727 序列 Serine 位置的生長 因子為 EGFR12。 細胞中有許許多多錯綜複雜的訊息傳遞路徑,在抑制癌細胞生長 研究上 STAT3 深受矚目,主要歸因於 STAT3 所結合的這些特定的 DNA 片段。目前已證實的 STAT3 結合片段有:bcl-2、bcl-xL、mcl-1、. 2.
(15) cyclin D1、VEGF13-17,這些片段與細胞的存活、週期的運轉有關。從 癌細胞的角度來看,bcl-2、bcl-xL、mcl-1 過度表現可幫助抑制癌細 胞發生凋亡;cyclin D1 的過度表現,可幫助癌細胞逃過細胞週期的檢 查點,使細胞週期繼續進行,使癌細胞增生;VEGF 的表現,則是促 成癌細胞周圍的血管新生,提供癌細胞生長的養分 7。由此可知, STAT3 在癌細胞的增生中扮演著很重要的腳色。. 1.3 Signal transducer and activator of transcription 3(STAT3)與口 腔癌之關係. STAT3 在致癌基因傳遞訊息中是不可或缺的中間者,常常在許 多癌症中可看到 STAT3 不正常的持續活化 7,18-20,如:有 82%的前列 腺癌可發現 STAT3 的持續活化 21;而在乳癌中,則是 70%22;在肺癌 中,則只有半數的癌細胞有此現象 23。但是,在口腔癌中,卻是超過 90%以上的口腔癌病患 8,皆可發現 STAT3 不正常的過度活化。 在 2002 年 Cancer 期刊中報導臨床上罹患口腔癌的病患跟腫瘤內 STAT3 變化的統計調查。從 90 位口腔癌的病患檢體中,有 74 種品 系的口腔癌細胞被發現有不正常活化的 STAT3 表現量。在這些採樣 的病人中,有 74%的病患在癌症病程早期,就已經發現 STAT3 不正. 3.
(16) 常高度活化的情形 24。同一年,在 Cancer research 的期刊中,也有學 者發現,若是在口腔癌細胞中 STAT3 磷酸化的程度越高,則病患的 存活率越低 25。 由這些證據指出,STAT3 在治療頭頸部癌,是一個重要的指標。. 1.4 第六白介素(Interleukin-6)誘導之 STAT3 路徑. Interleukin-6 是一個 cytokine 常與免疫調節發炎反應有關;也在 調節細胞凋亡、增生、血管新生和分化上扮演重要的角色 26,27。先前 的 研 究 證 實 , 在 頭 頸 部 癌 中 分 泌 interleukin-6 主 要 刺 激 STAT3 Tyrsine705 位置的磷酸化 11。. 1.5 第六白介素與頭頸部癌症之關係. 先前的研究報告指出 interleukin-6 會藉由刺激 STAT3 Tyr705 位 置的磷酸化而促進腫瘤細胞的存活與生長. 。並且,也有研究指出. 11,28. HNSCC 細 胞 株 會 自 體 分 泌 (autocrine) 或 旁 分 泌. (paracrine). interleukin-6 來刺激自身的 STAT3 活化 11。當 IL-6 結合在細胞表面的 接受器上時(JAK family protein kinases) ,在細胞質中未活化的 STAT3 會被磷酸化,而活化的 STAT3 會形成 dimer 並進入細胞核。活化的 4.
(17) STAT3 dimer 會和特定的基因片段結合例如:cyclin D1, Bcl-2,並轉 錄這些片段,而使頭頸部癌細胞達成增生、逃脫細胞凋亡…等目的 11。 在臨床上的研究也發現 interleukin-6 與頭頸部癌相關的例子:在 頭頸部癌病患血中可以測量到高濃度的 interleukin-629;在頭頸部癌病 患從口腔轉移到淋巴節的腫瘤組織切片裡,可以觀察到 interleukin-6 被高度的轉錄. 30. ; interleukin-6 會增進頭頸部癌細胞轉移侵犯骨頭. 31. ;頭頸部癌病患 interleukin-6 的血中濃度越高,則其放射線治療阻. 抗性也越高. 32. ;將從頭頸部癌病患所取下的口腔癌細胞,在 primary. culture 後,外加 interleukin-6 可發現促進頭頸部癌細胞生長之現象 33。 由這些例子指出,interleukin-6 對於頭頸部癌病患的治療而言, 是一明顯的生物指標(biomark),對研究頭頸部癌而言有相當的助益。. 1.6 茶的介紹 茶在中國被當作日常飲料和天然的藥草已有幾千年的歷史,而在 世界上也是最常飲用的農產品之ㄧ,在植物分類上屬山茶科 (Theaceae)、山茶屬(Camellia),每年消耗數百萬公噸以上。一 般公認茶的原產地為中國雲南地區,之後才往亞洲其他氣候溫暖的地 方擴散。而對於飲茶的由來,依據中國古老的傳說,茶的效用是由神 農氏傳給平民百姓的【陸羽茶經:「茶之為飲,發乎神農氏」】。在. 5.
(18) 當時,茶主要是被當作藥品來服用,直到唐朝時期才漸漸成為一種日 常的飲料。而西方飲茶的歷史,則是十六世紀航海時代,葡萄牙主掌 中國與歐洲的貿易,並於 1557 年,以廣東的澳門作為基地,開始向 歐洲輸入中國茶葉。後來,因為英王與葡萄牙皇后聯姻的關係,茶葉 展轉由葡萄牙人手中介紹到英國。由於當時中國茶葉在歐洲市場中十 分的昂貴,使得英國人想直接從中國進口茶葉以節省成本,便於 1869 年在福建的廈門建立茶葉的通路。茶在福建的方言中唸做 ”tai”, 也就是英文中 ”tea”的音源,【中國茶文化史】。 中國歷代以茶治病,其應用相當的廣,在漢代【神農本草經】中 有記載著「茶味苦,飲之使人益思、少臥、輕身、明目。」,而明代 李時珍的【本草綱目】中也有記載著,茶主治「痿瘡、中風昏愦、多 睡不醒、傷暑泄痢、大便下血、久年心痛、痔瘡作癢、陰囊生瘡、風 痰癲疾、霍亂煩悶、脈不通、痰喘咳嗽」等。清代食醫王孟英在【隨 息居飲食譜】中記載著:「茶,清心神、醒睡除煩、涼肝膽、滌熱清 痰、肅肺胃、明目解渴。凡暑穢、痧氣腹痛、乾霍亂、痢疾等症,初 起飲之,輒癒。」 。歷代的醫學家,皆從不同角度來闡明茶的治療效 用。唐代醫學家陳藏器在【本草拾遺】中記載道: 「茶,破熱氣、除 瘴氣、利大小便。」 ;元代醫學家王好古在【湯液本草】中記載: 「茶, 清頭目、治中風昏愦、多睡不醒。」 ;明代醫學家吳瑞元的【日用本. 6.
(19) 草】則記載: 「茶,炒煎飲,治熱毒赤白痢。」 ;而明代大醫學家李時 珍著的【本草綱目】有寫到:茶「濃煎,吐風熱痰涎。」,「合醋, 治泄痢,甚效。」;清代醫學家趙學敏在【本草綱目拾遺】中,更詳 細的記載 21 種茶的性味、功能、主治之差異和特點。經我國歷代醫 學家們的不斷實踐,茶在醫療上應用的範圍不斷擴大,不僅僅用於治 療,還用於天花等疾病之預防。另外在古代醫書中所記載的單味茶與 複方茶飲就有一百多個,其中【直指方論】的薑茶飲、【局方】的川 芎調散、【經驗百病良方】的午時茶、【醫學新語】的治痢散…等, 尤稱著於世。隨著醫學科學的不斷精進,近年來茶葉及其製劑在醫療 上的應用價值,也漸漸受到國際學者們的矚目。 市售茶葉種類複雜,以發酵程度不同來區分可分為不發酵、半發 酵、全發酵及後發酵茶。由於製茶過程不同,不同類型茶葉所含成分 也有所差異。一般來說,不發酵茶以綠茶為代表;半發酵茶,其發酵 程度以 10%~50%不等,以包種茶、烏龍茶為代表;全發酵茶,則以 紅茶為代表。所謂的發酵,是指茶葉中 oxidases 對各種物質進行的氧 化反應。在不發酵製程中的”炒菁”、”蒸菁”的步驟,其目的便是 利用高溫來早期破壞 oxidases 的活性,以阻斷發酵反應;相反的, 在全發酵茶的製程中,破壞 oxidases 活性的步驟則是在最後才進行, 使得茶葉能充分的進行發酵。. 7.
(20) 茶的成分很複雜,其主要活性成分有(一)黃烷酮醇類的兒茶素 (catechins) , (二)酚酸類(gallic acid) , (三)花青素類(anthocyanins) , (四)茶黃素(theaflavin),(五)生物鹼類(caffeine)等。而茶 多酚類化合物為茶葉中最廣泛研究的物質,其藥理特性較其他物質研 究的較為清楚,目前主要可分為四個方面:抗菌 34、抗發炎 35、影響 catecholamines 代謝 36,37、抑制脂質過氧化(lipid peroxidation)38。. 1.7 綠茶與抗癌之簡介. 綠茶與紅茶雖然都是以相同的茶葉來源製造,但卻因為製造過程 不同,不但有不同的風味,對健康上的功效也完全不同。而綠茶之所 以能發揮健康功效,主要是因製程中保存了較多完整的、還原態的多 酚類,對人體健康上有更多保護的效果 39。在流行病學上,也早已有 調查證實,飲用綠茶可以降低食道癌的發生率 40。而另一篇關於抽 菸、酗酒和飲茶的研究上更指出,飲用綠茶可以降低食道癌與胃癌的 發生率 41。 綠茶含有許多有生物活性的多酚類,因此被考慮用於在抑制腫瘤 生成和抑制腫瘤生長上 42。近幾年來,綠茶被廣泛的發現在許多不同 的癌症中有化學防治(chemopreventive)與化學治療. 8.
(21) (chemotherapeutic)效果 43-50。綠茶中多酚類化合物主要是由 catechin compounds 所組成,而在許多先前的研究中發現 catechin compounds 才是酚類化合物中活性的部份 51。目前認為 catechin compounds 在預 防與治療癌症上的功能大致上可分為,清除自由基、抗致癌物質、抑 制發炎反應。 清除自由基:catechin compounds 具有很強的抗氧化能力 52,可 將細胞產生之過多的自由基清除,以防止這些身體來不及代謝的自由 基去攻擊 DNA,而造成細胞突變或癌化。因此,許多學者們認為 catechin compounds 可以有效降低 initiation stage 的發生,而達到預防 癌症的效果 53。 抗致癌物質(Carcinogen):catechin compounds 除了可間接的因降 低自由基的含量而阻斷致癌物質對身體的影響外,某些致癌物質甚至 是直接與 catechin compounds 進行反應 54,如 tea catechins 抑制 nitrosation reaction,便是因為 catechin compounds 與 nitrosating species 作用,大量降低了 N-nitro compounds 的量 55。 抑制發炎反應:許多報告指出 tea catechins 對於抑制 inflammatory56 與 hyperplasia57 具有相當的活性。發炎反應會導致細胞 產生 hyperplasia 的現象,長期處於 hyperplasia 下的細胞,可能會導 致 tumor promotion,這種情形在皮膚癌、血癌中尤其明顯。目前認為,. 9.
(22) tea catechins 抑制發炎反應的作用,是因其明顯降低白介素 mRNA 的 表現量的關係 58。 近幾年來的研究發現,(-)-Epigallocatechin-3-gallate (EGCG)是 catechin compounds 中最主要、最有效的抗氧化活性多酚類成分,所 以有非常多研究綠茶機轉的學者們開始專注於 (-)-Epigallocatechin-3-gallate 的探討。. 1.8 表沒食子兒茶素沒食子酸脂(EGCG)之抗癌背景介紹. (-)-Epigallocatechin-3-gallate (EGCG)是綠茶中主要的活性多酚類 成分,也擁有抑制癌化因子的潛力 59。EGCG 在許許多多不同的癌症 中陸陸續續的被發現其抗癌效果,例如:在前列腺癌中,EGCG 藉由 活化 p53 蛋白並抑制 MDM2 蛋白來誘使癌細胞凋亡;在多發性骨髓 瘤細胞中,EGCG 藉由調控 LR1 蛋白,抑制骨髓細胞的癌化,並使 骨髓瘤細胞週期停滯且凋亡 60。EGCG 在許多不同的癌細胞上產生的 抗癌機制,主要的機轉為:抑制血管新生 61、藉由調控生長因子來抑 制癌細胞增生 62-65、抑制氧化傷害 66、誘導癌細胞凋亡 67,68、使細胞週 期停滯 69。而且,更重要的是,已有研究證實,EGCG 是有選擇性的 抑制癌細胞的生長以及誘導凋亡,但對於正常細胞的生長卻沒有任何. 10.
(23) 影響 70,使得我們對 EGCG 更加好奇並投身於 EGCG 研究。. 1.9 實驗動機. 綠茶的抗癌活性成分 EGCG 在許多的癌細胞上都有發現不同的 功效,而即使流行病學研究已指出綠茶可降低口腔癌的罹患率,但是 EGCG 對於抑制口腔癌細胞的詳細機轉還未被清楚的研究。在得知 STAT3、IL-6 和頭頸部癌的增生、癌化有高度相關的背景下,我們著 手研究 EGCG 是否對於由 IL-6 誘導的 STAT3 路徑有所影響。以確定, 是否 EGCG 也將是個可用於治療頭頸部癌症的新希望。. 11.
(24) 第二章 材料與方法. 2.1 實驗材料. 1. Osthole、Magolol、Geniposide、Emodin 得自於林慧怡老師實驗室, China Medical University (Taichung, Taiwan)。 2. Ritin、TF1、TF2、TF3、5GG 得自於林仁混老師實驗室,National Taiwan University (Taipei, Taiwan)。 3. Apigenin 、 (−)-epigallocatechin-3-gallate (EGCG),購自Sigma公司。 4. Interleukin-6購自Santa Cruz Biotechnology公司。 5. Anti STAT3、PhosphoSer727-STAT3與β-actin antibody,購自Cell Signaling Technology, Inc.公司。 6. PhosphorTyr705-STAT3 antibody購自Santa Cruz Biotechnology公司。. 12.
(25) 2.2 細胞培養. 2.2.1 細胞株. 本研究之實驗細胞SAS、Cal-27、Ca9-22、HSC3等頭頸部癌症細 胞株皆由高銘欽老師實驗室所贈。 SAS: 培 養 於 90% 的 DMEM ( Dulbecco Modified Eagle Medium),包含有10%的FBS(fetal bovine serum)、1%青黴素與鏈 黴 素 的 混 合 抗 生 素 ( 100× stock solution : Penicillin 50U/ml ; Streptomycin 50 μg/ml, Gibco) Cal-27 : 培 養 於 90% 的 DMEM ( Dulbecco Modified Eagle Medium),包含有10%的FBS(fetal bovine serum)、1%青黴素與鏈 黴 素 的 混 合 抗 生 素 ( 100× stock solution : Penicillin 50U/ml ; Streptomycin 50 μg/ml, Gibco) Ca9-22:培養於90%的MEM,包含有10%的FBS(fetal bovine serum)、1%青黴素與鏈黴素的混合抗生素(100× stock solution: Penicillin 50U/ml;Streptomycin 50 μg/ml, Gibco) HSC3:培養於90%的DMEM/F12,包含有10%的FBS(fetal bovine serum)、1%青黴素與鏈黴素的混合抗生素(100× stock solution:. 13.
(26) Penicillin 50U/ml;Streptomycin 50 μg/ml, Gibco). 2.2.2 細胞培養與繼代. 各品系之口腔癌細胞株皆培養於5%CO2,恆溫37℃的細胞培養箱 中,每隔48小時以trypin-EDTA(0.05%trypin 與 2.5mM EDTA)將 細胞打散,以6×105 個細胞分種於新的10公分培養皿中做繼代培養。. 2.3 細胞存活率分析. 本篇論文所採用的分法為MTT試驗(MTT assay) 。原理如下述, MTT(3-(4,5-Dimethylthiazol-2-yl)-2,5-dipheny tetrazolium bromide)進 入細胞內而被粒線體內膜上的琥珀酸去氫酶(Succinate dehydrogenase on Mitochondrial inner membrane)給還原成MTT,不溶於水,呈現紫 色針狀結晶;因為只有在活細胞中其粒線體才有活性,因此只有活細 胞才會生成結晶,在結晶完全形成之後,會結在細胞上造成細胞死 亡,再用DMSO溶出結晶,用595mm的波長在ELISA reader 下讀取吸 光值。 細胞以1.5×104/well種於24孔盤中,24小時後,換成含有指定濃 度指定藥物的培養液(一個孔1ml),培養24小時後,加入MTT試劑 14.
(27) (MTT 2mg/ml 溶於PBS中)反應3小時待結晶形成,將24孔盤中的 培養液與試劑吸乾,加入500μl的DMSO溶解結晶,取200μl溶解液轉 入96孔盤以590mm之波長在ELISA reader 下讀取吸光值。. 2.4 流式細胞儀分析(Flow cytometry). 口腔癌細胞種入6公分培養皿,每盤細胞數為3×105個,培養液為 5ml,24小時後,換成含有20μM EGCG的培養液,放入 37℃、5% CO2 的培養箱培養0、12、24、36、48小時。將各組別的細胞用trypsin-EDTA 打下連同培養液一起收集至15 ml離心管中,離心1500rpm、5分鐘。 倒乾上清液,加入1 ml PBS 將細胞打散,離心1500rpm、5分鐘。倒 去上清液,將細胞完全打散後,加入4℃、70%的冰酒精進行細胞固 定步驟 (以”SHAKE 3”速度震盪,一滴一滴緩慢將酒精滴入),再冰 入-20℃冰箱中隔夜。第二天,將固定好之細胞從冰箱取出,離心 1500rpm、5分鐘,倒乾上清液。加入1 ml PBS 將細胞打散,離心 1500rpm、5分鐘,倒去上清液,將細胞完全打散加入500μl的PI stain 染劑(Sigma),避光反應30分鐘。再將細胞轉至FACS專用管,以流 式細胞儀(Flow cytometry)分析。. 15.
(28) 2.5 細胞總蛋白收集與定量. 細胞以1×106個種植於10公分培養皿中,待16~20小時細胞貼附 後,給予指定劑量的EGCG並培養指定的時間於細胞培養箱中。待處 裡後,將各組別的細胞用trypsin-EDTA打下連同培養液一起收集至15 ml離心管中,離心1500rpm、5分鐘。去除上清液,加入1 ml PBS 將 細胞打散,離心1500rpm、5分鐘。將上清液倒乾,加入1 ml PBS 將 細胞打散並轉到1.5 ml eppendorf,離心12000rpm、1分鐘。用微量吸 管確實吸乾上清液,收集的細胞可存放於-80℃冰箱一個月備用。 將收集的細胞加入適量的lysis buffer(10% glycerol, 1% Triton X-100, 137 mM NaCl, 10 mM NaF, 1 mM EGTA, 5 mM EDTA, 1 mM sodium. pyrophosphate,. 20. mM. Tris-HCl,. pH. 7.9,. 100. mM. β-glycerophosphate, 1 mM sodium orthovanadate, 0.1% SDS, 10 μg/ml aprotinin, 1 mM phenylmethylsulfonyl fluoride, and 10 μg/ml leupeptin) 置於冰上,每10分鐘震盪一次(以”vortex 9”速度震盪)。待震盪三次 後,再以超音波震盪1分鐘,最後離心12000rpm、30分鐘、4℃。以微 量吸管吸取上清液,收集於新的eppendorf中,此上清液即為細胞總蛋 白(total protein) 。當總蛋白質定量後,以每管50 μg的蛋白質分裝儲 存於-20℃備用。 蛋白質定量,以Bio-Rad Protein Assay Dye來測定。其原理簡述 16.
(29) 如下,Bio-Rad Protein Assay Dye中酸性的Coomassie® Brilliant Blue G-250會與蛋白質結合,而蛋白質濃度不同會使溶液顏色呈現不同程 度的變化,通常蛋白濃度介於0~20 μg/ml時,溶液顏色將會由棕紅色 轉灰黑色,最後變成深藍色。取稀釋的Bio-Rad Protein Assay Dye(1:4 Milli Q 水)1 ml 分別加入0、2、4、6、8、10 μl的BSA(Bovine serum albumin 2mg/ml),混合均勻後,取200 μl之混合液至96孔盤,以595 mm 之波長在ELISA reader 下讀取吸光值,並以此六點對應之吸光值建立 檢量線(回歸系數須大於0.995才可採用)。取待定量之蛋白質液2μl混 合於1ml 稀釋的Bio-Rad Protein Assay Dye,取200 μl之混合液至96孔 盤,以595 mm之波長在ELISA reader 下讀取吸光值,並以建立好的 檢量線換算蛋白質濃度。. 2.6 蛋白質之電泳分析(SDS-PAGE). 實驗所使用的電泳裝置為Bio-Rad 蛋白質電泳槽,待鑄膠裝置組 合後,先加入分離膠體溶液(separating gel),再加入酒精將上層膠 壓平,待凝固後倒掉酒精並擦拭乾後加入集膠溶液(stacking gel), 插上齒梳,待膠體凝固後置於電泳槽中,注入電泳緩衝液(running buffer)。取50mg蛋白質樣本加入其1/5體積的 6× sample loading dye, 17.
(30) 混勻後在99 ℃加熱5分鐘後,迅速置於冰上再注入樣本槽。先以80V 進行電泳,待藍色染劑跑到分離膠體時,將電壓調整為110V繼續電 泳,直到藍色染劑跑到膠體底部,關閉電源將膠體取出。. 2.7 西方墨點法 (Western blotting). 實驗所使用的電泳裝置為 Bio-Rad 濕式電泳轉漬槽,取出跑好 的 SDS-PAGE 後,切除集膠體部份,膠片浸在轉漬緩衝液(Transfer buffer)中。取轉印紙(硝化纖維紙 nitrocellulose paper)切成膠片大 小,用甲醇浸潤,先鋪上三張濕潤的 3M 濾紙,後鋪上濕潤的轉印 紙,疊上已濕潤的膠片,再鋪上三張濕潤的 3M 濾紙。(注意!過 程需趕走氣泡),轉印紙那面位向正極,膠片那面位向負極,以 70 mv 轉印 3 小時。之後,將轉印紙放入 1x TBST 含 5% 脫脂牛奶, 室溫搖盪 1 小時,以填塞沒有蛋白質轉印上去的空間。倒掉進行 Blocking 的牛奶,加入第一次抗體(1:1000 稀釋),置於室溫搖 擺 1 至 2 小時或放置 4℃隔夜(超過 12 小時)。倒去第一次抗體或 回收重覆使用,以 1 x TBST 清洗 5 分鐘三次。再加入第二次抗體(1: 2000 稀釋),置於室溫搖擺 1 至 2 小時。倒去二次抗體,轉印紙以 18.
(31) 1 x TBST 清洗 5 分鐘三次。最後將轉印紙浸泡於 ECL 試劑(Enhanced chemiluminescence)反應 1 分鐘,再將轉印紙以透明投影片夾住,黏 貼固定於壓片夾(cassette)中,在暗房操作以 X-ray film (Kodak)感光 得蛋白質訊號。. 2.8 免疫螢光染色( Immuocytochemistry ). 先將蓋玻片浸於 99%酒精中,過火,小心置入 6 孔盤中(1 well 一片),加入培養液 3ml/well。再種入細胞,每 well 種 1×105 個細胞。 待 24 小時細胞貼附後,換成含有 20μM EGCG 的培養液,培養不同 的時間後(0、15、30、60、120 分鐘),將培養液吸乾。以 PBS 洗滌 三次,用 4% paraformaldehyde/PBS 固定細胞 10~15 分鐘,吸去 4% paraformaldehyde/PBS 並以 PBS 洗三次。至於冰上,加入 0.2% TritonX-100 / PBS 20 分鐘,再用 PBS 洗三次,再加入 5% BSA/PBS、 室溫、1 小時。移去 5% BSA/PBS 後,加入第一次抗體(1:100,5% BSA/PBS 稀釋),置於 4℃放置隔夜。第二天,回收第一次抗體,用 PBS 洗 3 次。再加入第二次抗體(1:200,1×PBS 稀釋),避光置於 4℃ 放置隔夜。隔日,移去第二次抗體,用 PBS 洗三次。加入 1 ml DAPI(1:50000,1×PBS 稀釋)避光置於室溫反應 15 分鐘,用 PBS 洗 19.
(32) 三次。滴取一滴封片液(Mounting medium)於載玻片中心,小心夾取 蓋玻片,將細胞貼附面朝下蓋於載玻片上。使用共軛焦雷射顯微鏡 (confocal laser scanning microscope)觀察並擷取影像。. 2.9 統計分析. 實驗結果以平均值標準差(mean ± SD)表示,使用 ANOVA(單因 子變異分析) 來決定實驗組與對照組之差異。 *表示 p<0.05;** 表示 p<0.01;***表示 p<0.001,當 p 值小於 0.05 以下時,則認 為具統計上意義。. 20.
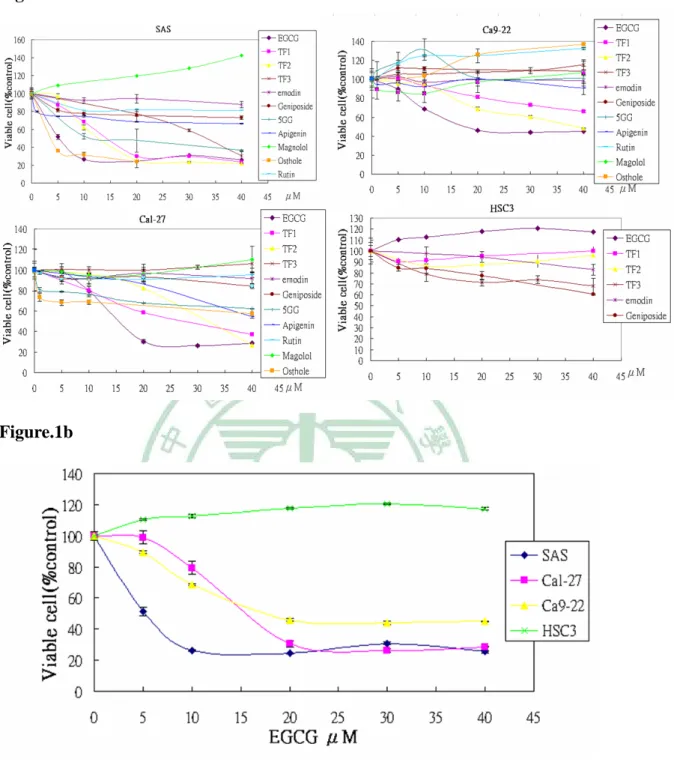
(33) 第三章 結果. 3.1 EGCG 抑制頭頸部癌細胞的增生. 首先,我們將四株頭頸部癌細胞株(SAS、Cal-27、Ca9-22、HSC3) 分別以 0、5、10、20、40 μM 的濃度處理各種黃酮類(Apigenin、Rutin)、 多酚類(EGCG、TF1、TF2、TF3、Magolol、5GG)和其他類別(emodin、 Geniposide、Osthole)的抗癌天然化合物 24 小時,再使用 MTT assay 方 法來測定細胞存活率。在 Figure.1a 中,我們發現 EGCG 在三株頭頸 部癌細胞株(SAS、Cal-27、Ca9-22)中皆為最有效抑制癌細胞生長的 化合物。因此,進一步的將四株頭頸部癌症細胞株在 EGCG 處理下 的存活曲線整理於 Figure.1b 中,SAS 的 IC50 為 5 μM;Cal-27 的 IC50 為 15 μM;Ca9-22 的 IC50 為 20 μM,而 HSC3 細胞株在 EGCG 的處 理下無抑制細胞生長的現象,此特例我們將在之後的段落中研究與討 論。為觀察 EGCG 處理後短時間內細胞中蛋白的變化,我們取 20 μM 此共同引起抑制生長的有效劑量,作為之後觀察細胞中蛋白變化的處 理劑量。. 21.
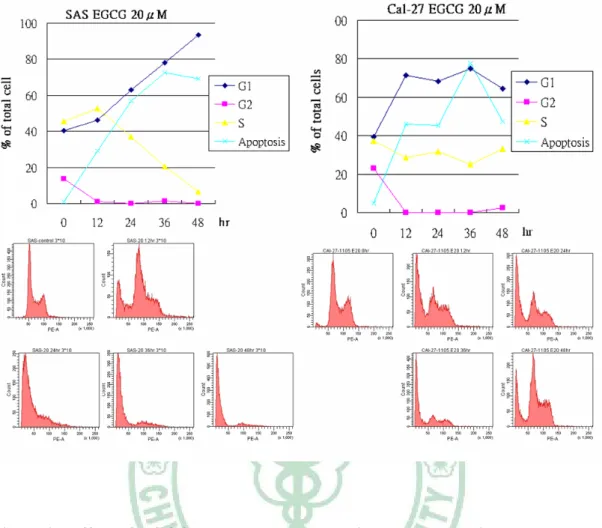
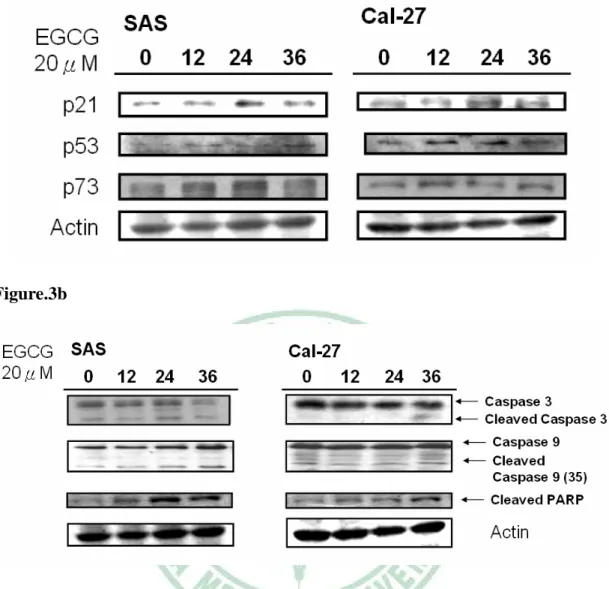
(34) 3.2 EGCG 促使頭頸部癌細胞細胞週期停滯於 G1 期並誘導細胞凋亡. 在前面的實驗中,找到了最有效抑制頭頸部癌細胞生長的天然物 EGCG,而許多研究 EGCG 的報告中,皆指出 EGCG 在許多不同品系 的癌症細胞中能引起許多的抗癌效果,如:細胞週期停滯於 G1 期 23,24,25. 、誘導癌細胞的凋亡 71-74 …等。為了驗證是否這些 EGCG 的抗. 癌效果一樣也發生在頭頸部癌細胞中,我們將 SAS、Cal-27 細胞分別 以 20 μM EGCG 處理 0、12、24、36、48 小時,使用流式細胞儀去分 析其細胞週期的變化。在 Figure.2 中,不論是在 SAS 或是 Cal-27 細 胞中,在處理 12 小時 EGCG 之後,都可以發現停滯於 G1 期細胞的 比例明顯的升高,也可以看到凋亡的癌細胞比例隨著處理時間的變長 也逐漸變多。. 3.3 EGCG 促使細胞週期停滯 G1 期、細胞凋亡之相關蛋白的表現. 經細胞層面的方法證實,也進一步的使用西方點墨法來檢查是否 在蛋白的分子層面上也是如此。將 SAS、Cal-27 細胞分別以 20 μM EGCG 處理 0、12、24、36、48 小時,再收集各組之細胞總蛋白,使 用西方點墨法觀察其蛋白變化。在 Figure.3a 中,發現與停滯於 G1 期 之相關蛋白(p21、p53、p73)的表現量,隨著處理時間的變長而逐漸 22.
(35) 的上升。與細胞凋亡相關的 Caspase 3、Caspase 9、PARP 等蛋白,當 細胞凋亡時這些蛋白會出現裂解態(Cleaved form),而隨著處理 EGCG 時間的變長也可觀察到這些蛋白裂解態的上升。. 3.4 EGCG 抑制頭頸部癌細胞中 STAT3 蛋白的磷酸化. 先前的研究中指出,癌細胞中持續活化的 STAT3 和癌細胞的存 活、增生是有相關性的 18。且在眾多品系的癌症病患中,大於 90%比 例的口腔癌病患中可以發現 STAT3 過度活化的現象 8,此情形在其他 的癌症病患中卻非如此 75。使我們想探究,EGCG 對頭頸部癌細胞株 的抑制活性,是否跟 STAT3 有所關聯? 因此,我們用西方點墨法來 觀察 EGCG 對於頭頸部癌細胞中 STAT3 磷酸化的影響。將 SAS、 Cal-27 細胞分別處理 0、5、10、20、40 μM EGCG 1 小時,在 Figure.4a 中可以看到隨著 EGCG 處理劑量的提高,不論是在 ser727 位置 (P-STAT3(727))或是 try705 位置(P-STAT3(705))磷酸化的 STAT3 其表 現量越來越少。在 Figure.4b 中,SAS、Cal-27 細胞分別處理 0、15、 30、60、120、240 分鐘 20 μM 的 EGCG,隨著處理時間的延長,STAT3 的磷酸化表現量也逐漸的下降。在 Figure.4c 中,我們再利用免疫螢 光染色法以共軛焦雷射顯微鏡(confocal laser scanning microscope)來. 23.
(36) 觀察細胞層面的變化。Figure.4c 中,我們將 SAS、Cal-27 細胞用 20 μM EGCG 分別處理 0、15、30、60、120 分鐘,並使用免疫螢光染色法 來觀察 P-STAT3(705)的表現量(綠色螢光) (藍色螢光則是細胞核的 位置)。隨著 EGCG 的處理時間的增長,P-STAT3(705)的表現量也逐 漸下降,在 SAS 細胞中,當處理至 30 分鐘時 P-STAT3(705)的表現量 幾乎接近到觀察不到的量;而在 Cal-27 細胞中,則是在處理 1 小時 的時間點,P-STAT3(705)的表現量才幾乎接近到觀察不到的地步。 STAT3 平常在不活化(無磷酸化)的狀態時,它是存在於細胞質 中的,有磷酸化的 STAT3 才能進入細胞核中。當有訊息傳遞出現時, STAT3 才會被磷酸化並兩兩組合成 dimer,再進入細胞核進行轉錄的 工作 9。由於在大多的癌細胞中可發現 STAT3 是持續活化的狀態,而 上述實驗我們發現 EGCG 可以抑制 STAT3 的磷酸化。為了更近一步 探討 EGCG 抑制 STAT3 磷酸化之後,是否也抑制 STAT3 轉入細胞核 中。在 Figure.4d 中,我們將 SAS、Cal-27 細胞用 20 μM EGCG 分別 處理 0、15、30、60、120 分鐘,再用免疫螢光染色法觀察細胞中 STAT3(綠色螢光)的位置,圖中的藍色螢光則是細胞核的位置。隨著 EGCG 的處理時間的增長,STAT3 漸漸往細胞核外移動。在 SAS 細 胞中,在僅僅處理 15 分鐘 20 μM EGCG 即可明顯的觀察到 STAT3 幾 乎全部都移動至細胞核外;在 Cal-27 細胞中,則是處理 2 小時 20 μM. 24.
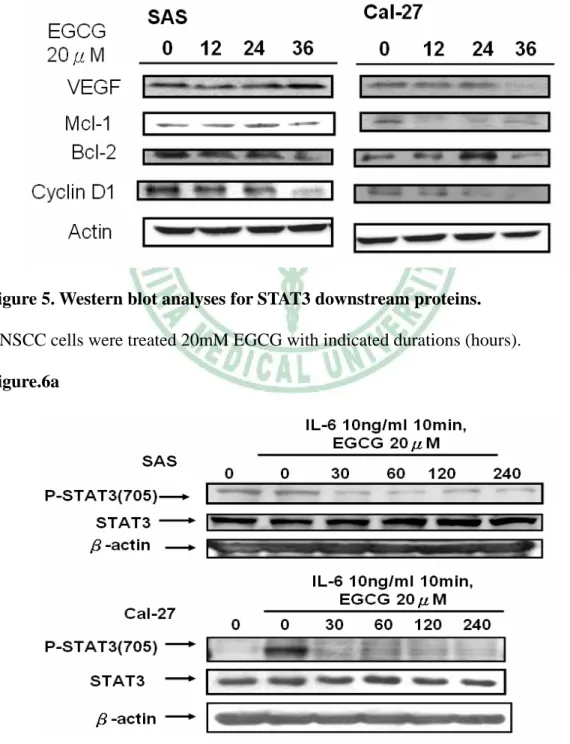
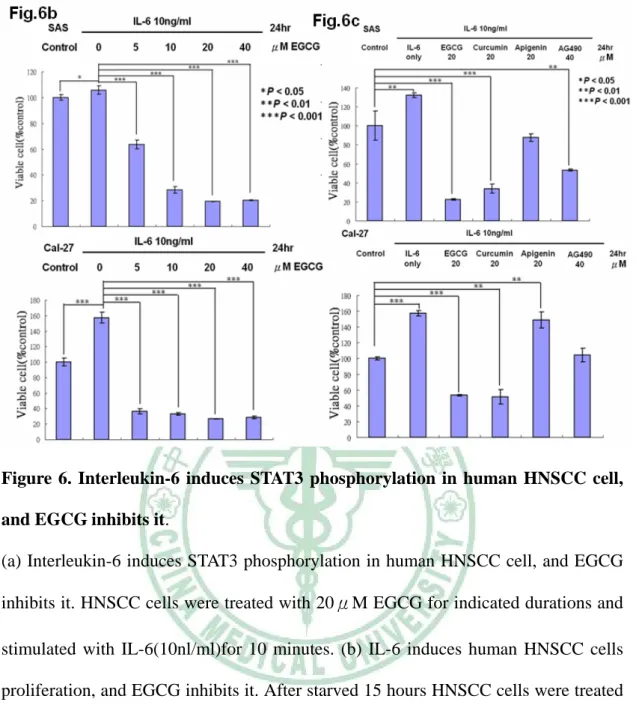
(37) EGCG 後可觀察到 STAT3 明顯的移動至細胞核外。. 3.5 EGCG 抑制 STAT3 蛋白之下游蛋白的表現. 先前許多的研究已找出許多 STAT3 轉錄的目標基因所產生出之 下游蛋白,例如: Bcl-2, VEGF, Mcl-1, and cyclin D113-17,而這些由 STAT3 所轉錄出來的下游蛋白質,大多與癌細胞的存活、增生有關 9。 為了了解在處理 EGCG 之後對於 STAT3 所轉錄之基因其下游蛋白表 現量之影響,SAS、Cal-27 細胞們分別處理 0、12、24、36 小時的 20 μM EGCG,再收集各組細胞的總蛋白,使用西方點墨法觀察各組 STAT3 下游蛋白之變化。在 Figure.5 中可以觀察到,隨著 EGCG 處理 時間的增加,Bcl-2, VEGF, Mcl-1, and cyclin D1 這些 STAT3 轉錄的下 游蛋白表現量也逐漸下降。根據以上的結果得知,EGCG 在抑制 STAT3 磷酸化之後,也阻斷了 STAT3 下游的蛋白表現。. 3.6 EGCG 抑制頭頸部癌中 Interleukin-6 誘導 STAT3 蛋白的磷酸化. 先前的研究報告指出 interleukin-6 (IL-6)會藉由刺激 STAT3 Tyr705 位置的磷酸化而促進腫瘤細胞的存活與生長 28。並且,也有研 究指出頭頸部癌細胞株會自體分泌(autocrine) 或旁分泌 (paracrine) 25.
(38) IL-6 來刺激自身的 STAT3 活化 11。因此,為了了解 EGCG 對於 IL-6 在頭頸部癌細胞中誘導 STAT3 活化之現象的影響,我們使用西方點 墨法來觀察 STAT3 磷酸化的情形。SAS、Cal-27 細胞們在分別處理 0、 30、60、120、240 分鐘 20 μM EGCG 後,移除含有 EGCG 的培養液, 換成含有 10 ng/ml Interleukin-6 的 DPBS 刺激 10 分鐘,再收集各組 細胞的蛋白,以西方點墨法來觀察 P-STAT3(705)蛋白量的變化。在 Figure.6a 中可觀察到,在沒有處理 EGCG 的組別裡,雖然可以明顯 看見 IL-6 增強 P-STAT3(705)的表現量。但是在有處理 EGCG 的組別 中,我們仍然可以發現 P-STAT3(705)的表現量還是被抑制下來的。根 據以上的結果得知,EGCG 除了可抑制 STAT3 磷酸化之外,EGCG 也可以消除 IL-6 在口腔癌細胞中誘導 STAT3 活化的能力。. 3.7 EGCG 抑制由 Interleukin-6 所引發的細胞增生. 先前的研究報告指出 interleukin-6 (IL-6)會藉由刺激 STAT3 Tyr705 位置的磷酸化而促進腫瘤細胞的存活與生長 28。所以,我們也 將 EGCG 對於 IL-6 在頭頸部癌細胞中誘導的細胞增生現象進行試 驗。SAS、Cal-27 細胞待貼盤後,先以無 FBS 的培養液培養 15 個小 時(消除 FBS 中其他會刺激細胞增生的因子的干擾) ,再分別處理 0、. 26.
(39) 5 、 10 、 20 、 40 μM EGCG , 各 組 同 時 加 入 10 ng/ml 濃 度 的 Interleukin-6 ,在處理 24 小時後,使用 MTT assay 分析各組的細胞 存活率。在 Figure.6b 中可觀察到,在沒處理 EGCG 的組別裡,可以 明顯看見 IL-6 有刺激細胞增生的效果;而在有處理 EGCG 的組別中, EGCG 仍然可以抑制頭頸部癌細胞的生長。 AG490 是公認抑制 STAT3 磷酸化的化合物 76;先前的研究指出 Curcumin 也會抑制 interleukin-6 所誘導之 STAT3 磷酸化. 77. ;而. Apigenin 也是目前熱門的有抗癌活性的多酚類成分。所以,更近一步 的,我們將 EGCG 與 Apigenin、AG490、Curcumin 一起比較抑制 IL-6 所誘導的細胞增生現象之效果。SAS、Cal-27 細胞待貼盤後,先以無 FBS 的培養液培養 15 個小時,再分別處理 20 μM EGCG、20 μM Curcumin、40 μM Apigenin、40 μM AG490,各組並同時加入 10 ng/ml 濃度的 Interleukin-6 ,在處理 24 小時後,以 MTT assay 分析各組細 胞的存活率。Figure.6c,在與各組比較之下,可以發現 EGCG 抑制 IL-6 所誘導的細胞增生之效果是最有效的。. 27.
(40) 第四章 討論 許多研究已證實天然植物中的多酚類化合物有抑制癌細胞生長 或產生細胞毒性的作用。我們從植物中萃取出具有抗癌活性的天然化 合物來比較其抑制頭頸部癌症細胞的活性。以下簡單的介紹這些天然 物的背景: TF1 、 TF2 、 TF3 、 5GG : TF1 是 Theaflavin 的 簡 稱 、 TF2 是 Theaflavin-3-gallate的簡稱、TF3是Theaflavin-3,3’-digallate的簡稱,這 三個化合物們是從紅茶中萃取出的多酚類化合物;5GG則是有五個 Galloy機團的化合物,其化學名為Penta-O-galloyl-β-D-glucose。其中, TF3與5GG已被證實可藉由抑制前列腺素活化前列腺素受器的功能, 以達到抑制前列腺癌的效果78。 Emodin(大黃素)是由中藥植物裡的大黃萃取出的天然化合物。 在中醫的使用上,主要用於瀉下、治療實熱便秘、食積停滯、腹痛、 急性闌尾炎、急性傳染性肝炎、血瘀經閉、牙痛、吐血、急性結膜炎、 水腫、淋濁等。外用可治療燒燙傷、化膿性皮膚病、癰腫瘡瘍等病症, 【內經、傷寒論、金匱要略】 。現今,有許多研究發現 Emodin 在前列 腺癌 79、胃癌 80 上有誘導細胞凋亡的活性。 Apigenin(芹菜素)常見於春黃菊(Camomile) 、柑橘類的植物中, 而在一些常見的蔬菜水果中也可發現 Apigenin 的存在,如:檸檬、 28.
(41) 蘋果、西洋芹(parsley)、羅勒(basil)、龍蒿(tarragon)、胡荽葉 (cilantro)。許多研究發現 Apigenin 具有多種藥理活性,例如:抗腫 瘤、抗炎症、降血壓、抗動脈硬化和血栓症、抗焦慮、抗菌、抗病毒 以及抗氧化作用等方面 81。 Rutin(芸香苷)可於蕃茄、槐花、蕎麥、柑橘類植物中發現, 有助於維他命 C 的適度吸收和利用,並協助維他命 C 維護膠原蛋白 的健康而有抗老化的功能,也有人稱 Rutin 為維生素 P。而最常見用 途是強化微血管與治療過敏方面,根據研究的顯示,它可減緩肥大細 胞釋放組織胺的速度,而組織胺正是引起一般過敏反應(如鼻塞)的化 學物質。因 Rutin 具抗炎的特性,也顯示抗病毒、抗細菌及抗癌的作 用。目前,在抗癌活性的研究上,已發現 Rutin 藉由活化 macrophages 來抑制 NO 的產生,進而抑制 TNF-α 的產生 84,85。 Magnolol(厚朴酚)常見於木蘭科(Magnolia)植物 中。大多 Magnolol 的研究都是對於其延緩皮膚老化的機制上,且此成分已添加 於保養品中並販售上市。Magnolol 是很好的抗氧化劑,可以保護神經 細胞免於自由基的傷害 84。也可以抑制腫瘤壞死因子,降低發炎反應 85. 。而近年來,也有新的報導發現 Magnolol 在血癌、膀胱癌細胞的抗. 癌效果 86,87。 Osthole ( 蛇 床 子 素 ), 萃 取 於 繖 形 科 (Umbelliferae) 蛇 床. 29.
(42) (Cnidium monnieri (L.) Cuss)的果實中。在中醫的使用上,用於治 療陽痿、子宮虛冷、寒濕帶下、濕痺腰痛、外治外陰濕疹、婦人陰癢 白帶、滴蟲性陰道炎,皮膚騷癢等抗菌用途,【神農本草經、本草求 真、重修政和經史證類備用本草】 。近幾年,有研究指出 Osthole 可抑 制脂肪肝之生成 88,並也發現 Osthole 有抗癌的活性 89。 Geniposide(槴子苷)萃取於茜草科的梔子(Gardenia spp.) 。目 前研究發現具有促進神經細胞分化的功能. 90. ,也有報導指出. Geniposide 能誘導神經膠質瘤細胞凋亡 91。 從 Figure.1a,發現 TF1、TF2、TF3、5GG、Emodin、Apigenin、 Rutin、Magnolol、Osthole、Geniposide、EGCG 在頭頸部癌細胞株 SAS、 Cal-27、Ca9-22 存活分析中, EGCG 對三株頭頸部癌細胞皆具最有 效的抑制作用,也讓我們發現 EGCG 廣效的明顯抑制頭頸部癌細胞 株增生的能力。進一步探討 EGCG 和抑制頭頸部癌細胞增生的關係, 而 將 四 株 頭 頸 部 癌 細 胞 株 在 EGCG 處 理 下 的 存 活 曲 線 整 理 於 Figure.1b 中。 在 Figure.1 的實驗中,找到了最有效抑制頭頸部癌細胞生長的天 然物 EGCG,其他研究 EGCG 的報告中,皆指出 EGCG 在許多不同 品系的癌症細胞中能引起不同的抗癌機制,如:細胞週期停滯於 G1 期 69,92,93、誘導癌細胞的凋亡 67,71-73 等。在 Figure.2 中 Flow cytometry. 30.
(43) 細胞週期分析;與 Figure.3a、Figure.3b 中,使用西方點墨法檢查停滯 G1 期之相關蛋白(p21、p53、p73)、細胞凋亡相關的 Caspase 3、Caspase 9、PARP 的表現量,證實 EGCG 抑制頭頸部癌細胞生長,是藉由誘 導細胞停滯 G1 期之後再誘發細胞凋亡。 在許多期刊中報導頭頸部癌與 STAT3 之關係,我們想測試 EGCG 是否藉由影響 STAT3 訊息傳遞路徑,來抑制頭頸部癌細胞的 生長。 由 Figure.4a、b 發現 EGCG 抑制 STAT3 磷酸化的效果與給藥劑 量、給藥時間有著依存關係。藉由共軛焦顯微鏡也證實 STAT3 磷酸 化表現量也逐漸下降(Figure.4c),而處理 EGCG 後,STAT3 也明顯 的移動至細胞核外(Figure.4d) 。此外,由西方點墨法實驗發現 STAT3 路徑下游蛋白表現量也下降(Figure.5)。從 Figure.4 到 Figure.5 的結 果,說明 EGCG 確實抑制了 STAT3 訊息的傳遞,其主要作用點,是 在抑制細胞質中 STAT3 的磷酸化。 綜合由 Figure.1 到 Figure.5 所展現的結果,我們推論 EGCG 在頭 頸部癌細胞中,是藉由抑制 STAT3 的磷酸化,導致細胞中 STAT3 路 徑無法作用,而使得頭頸部癌細胞凋亡。 在 Figure.1b 中發現 EGCG 沒有抑制頭頸部癌細胞 HSC3 的生 長,進一步用西方點墨法探討其原因,發現經 EGCG 處裡後,HSC3. 31.
(44) 細胞內 STAT3 磷酸化蛋白的表現量,發現 EGCG 的處理並不會展現 抑制 HSC3 細胞中 STAT3 磷酸化(Figure.7)。此結果証實 EGCG 抑 制頭頸部癌細胞增生的效果,的確是因為抑制了 STAT3 路徑的關係 而達成的。 最近的研究報告指出 interleukin-6 會藉由刺激 STAT3 Tyr705 位 置的磷酸化而促進腫瘤細胞的存活與生長 28。另有研究指出頭頸部癌 細胞株會自體分泌(autocrine) 或旁分泌 (paracrine) IL-6 來刺激自身 的 STAT3 活化. 11. 。為了進ㄧ步測試 EGCG 抑制 STAT3 路徑與. interleukin-6 的存在是否相關聯(Figure.6a) ,以 MTT assay(Figure.6b) 測試 EGCG 抑制細胞增生的效果。結果證實,在 EGCG 處理下,可 以抑制 interleukin-6 促進頭頸部癌細胞的增生。 此外,研究報告中指出 AG490 是抑制 STAT3 磷酸化的化合物 76; Curcumin 是也有抑制 interleukin-6 所誘導 STAT3 磷酸化之活性 77; Apigenin 是目前抗癌活性的多酚類成分,在許多研究中常與 EGCG 一起比較抗癌活性。進一步,我們將 EGCG 與 Apigenin、AG490、 Curcumin 一起用於比較抑制 interleukin-6 所誘導的細胞增生現象。觀 察到 EGCG 在 interleukin-6 所誘導的細胞增生,是最有抑制效果的化 合物(Figure.6c)。 Figure.1 與 Figure.6c 的結果指出,EGCG 除了在抑制頭頸部癌細. 32.
(45) 胞增生是最好的化合物之外,對於 interleukin-6 在頭頸部癌細胞上誘 導的細胞增生,也是最好的抑制劑。 臨床上,超過 90%以上的口腔癌病患的細胞中,皆可發現 STAT3 不正常的過度活化 8。Cancer research 的期刊中報導, 若是在口腔癌 細胞中 STAT3 磷酸化的程度越高,則病患的存活率越低 25。在體外 試 驗 中 發 現 , 頭 頸 部 癌 細 胞 株 會 自 體 分 泌 (autocrine) 或 旁 分 泌 (paracrine) IL-6 來刺激自身的 STAT3 活化 11。;從頭頸部癌病患所 取下的口腔癌細胞,在 primary culture 後,外加 interleukin-6 可發現 促進頭頸部癌細胞生長之現象 33;且臨床上,也有許多例子證實頭頸 部癌病患 interleukin-6 的血中濃度的高低與其治療的難易度 32、癌細 胞轉移的機率 30,31 也有直接的關係;再加上本研究中,我們對於 EGCG 可以抑制 interleukin-6 在頭頸部癌細胞上的作用之發現,間接的說明 EGCG 在臨床上用於治療頭頸部癌病患,可能有好的預期與潛力。 已有研究指出了 EGCG 在癌細胞上的接受器(67-kDa laminin receptor ) 94 ; 在 多 發 性 骨 髓 瘤 細 胞 中 , EGCG 也 是 因 為 laminin receptor,而有選擇性的抑制癌細胞的生長以及誘導凋亡,卻對正常 細胞的生長卻沒有任何影響 60。而在頭頸部癌細胞的研究裡,也有人 發現 laminin 的表現量與頭頸部癌細胞的惡性度成正比 95。這些證據 意味著, EGCG 對於治療頭頸部癌症,或許也如同多發性骨髓瘤細. 33.
(46) 胞,是有特異性(有分別性)的抑制癌細胞的生長。 而未來研究計畫的部份:在 EGCG 抑制 STAT3 訊息傳遞上,可 追尋路徑上游 Jak/Tyk2 與 EGCG 的關係,以更確定 EGCG 抑制 STAT3 訊息傳遞的作用點;除了有 HSC3 細胞株的反證外,也可以用轉殖 (transfect)的技術,抹除頭頸部癌細胞 STAT3 的表現,更進一步的探 討 EGCG 和 STAT3 的作用關係;對於,處理 EGCG 無效的 HSC3 細 胞株,也許是因為 HSC3 與實驗中其他的頭頸部癌細胞株比起來,是 比較末期、惡性度比較高的細胞株,其細胞中許多機制也早已突變、 無功能 96,97。所以無法單單只用 EGCG 一個化合物來抑制,或許在治 療這種惡性度比較高的頭頸部癌細胞上,可往複合藥物療法的方向考 慮 98。. 34.
(47) 第五章 參考文獻 1. Murray, C.J.L. and Salomon J.A. (2002). Summary measures of population health: concepts, ethics, measurement and applications. WHO, Geneva. 2. Murray C.J.L. and Lopez A.D. (1996). The Global Burden of Disease. Cambridge: Harvard University Press. 3. Song, J.I., and Grandis, J.R. (2000). STAT signaling in head and neck cancer. Oncogene 19, 2489-2495. 4. Dhooge, I.J., De Vos, M., and Van Cauwenberge, P.B. (1998). Multiple primary malignant tumors in patients with head and neck cancer: results of a prospective study and future perspectives. Laryngoscope 108, 250-256. 5. Yen, A.M., Chen, S.C., and Chen, T.H. (2007). Dose-response relationships of oral habits associated with the risk of oral pre-malignant lesions among men who chew betel quid. Oral Oncol 43, 634-638. 6. Vokes, E.E., Weichselbaum, R.R., Lippman, S.M., and Hong, W.K. (1993). Head and neck cancer. N Engl J Med 328, 184-194. 7. Yu, H., and Jove, R. (2004). The STATs of cancer--new molecular targets come of age. Nat Rev Cancer 4, 97-105.. 35.
(48) 8. Nagpal, J.K., Mishra, R., and Das, B.R. (2002). Activation of Stat-3 as one of the early events in tobacco chewing-mediated oral carcinogenesis. Cancer 94, 2393-2400. 9. Germain, D., and Frank, D.A. (2007). Targeting the cytoplasmic and nuclear functions of signal transducers and activators of transcription 3 for cancer therapy. Clin Cancer Res 13, 5665-5669. 10. Liu, L., McBride, K.M., and Reich, N.C. (2005). STAT3 nuclear import is independent of tyrosine phosphorylation and mediated by importin-alpha3. Proc Natl Acad Sci U S A 102, 8150-8155. 11. Sriuranpong, V., Park, J.I., Amornphimoltham, P., Patel, V., Nelkin, B.D.,. and. Gutkind,. J.S.. (2003).. Epidermal. growth. factor. receptor-independent constitutive activation of STAT3 in head and neck squamous cell carcinoma is mediated by the autocrine/paracrine stimulation of the interleukin 6/gp130 cytokine system. Cancer Res 63, 2948-2956. 12. Levy, D.E., and Darnell, J.E., Jr. (2002). Stats: transcriptional control and biological impact. Nat Rev Mol Cell Biol 3, 651-662. 13. Puthier, D., Bataille, R., and Amiot, M. (1999). IL-6 up-regulates mcl-1 in human myeloma cells through JAK / STAT rather than ras / MAP kinase pathway. Eur J Immunol 29, 3945-3950.. 36.
(49) 14. Karni, R., Jove, R., and Levitzki, A. (1999). Inhibition of pp60c-Src reduces Bcl-XL expression and reverses the transformed phenotype of cells overexpressing EGF and HER-2 receptors. Oncogene 18, 4654-4662. 15. Ivanov, V.N., Bhoumik, A., Krasilnikov, M., Raz, R., Owen-Schaub, L.B., Levy, D., Horvath, C.M., and Ronai, Z. (2001). Cooperation between STAT3 and c-jun suppresses Fas transcription. Mol Cell 7, 517-528. 16. Fukada, T., Hibi, M., Yamanaka, Y., Takahashi-Tezuka, M., Fujitani, Y., Yamaguchi, T., Nakajima, K., and Hirano, T. (1996). Two signals are necessary for cell proliferation induced by a cytokine receptor gp130: involvement of STAT3 in anti-apoptosis. Immunity 5, 449-460. 17. Sinibaldi, D., Wharton, W., Turkson, J., Bowman, T., Pledger, W.J., and Jove, R. (2000). Induction of p21WAF1/CIP1 and cyclin D1 expression by the Src oncoprotein in mouse fibroblasts: role of activated STAT3 signaling. Oncogene 19, 5419-5427. 18. Bromberg, J.F., Wrzeszczynska, M.H., Devgan, G., Zhao, Y., Pestell, R.G., Albanese, C., and Darnell, J.E., Jr. (1999). Stat3 as an oncogene. Cell 98, 295-303. 19. Brown, M.D. (1999). Green tea (Camellia sinensis) extract and its possible role in the prevention of cancer. Altern Med Rev 4, 360-370. 37.
(50) 20. Darnell, J.E. (2005). Validating Stat3 in cancer therapy. Nat Med 11, 595-596. 21. Mora, L.B., Buettner, R., Seigne, J., Diaz, J., Ahmad, N., Garcia, R., Bowman, T., Falcone, R., Fairclough, R., Cantor, A., et al. (2002). Constitutive activation of Stat3 in human prostate tumors and cell lines: direct inhibition of Stat3 signaling induces apoptosis of prostate cancer cells. Cancer Res 62, 6659-6666. 22. Dolled-Filhart, M., Camp, R.L., Kowalski, D.P., Smith, B.L., and Rimm, D.L. (2003). Tissue microarray analysis of signal transducers and activators of transcription 3 (Stat3) and phospho-Stat3 (Tyr705) in node-negative breast cancer shows nuclear localization is associated with a better prognosis. Clin Cancer Res 9, 594-600. 23. Song, L., Turkson, J., Karras, J.G., Jove, R., and Haura, E.B. (2003). Activation of Stat3 by receptor tyrosine kinases and cytokines regulates survival in human non-small cell carcinoma cells. Oncogene 22, 4150-4165. 24. Nagpal, J.K., Mishra, R., and Das, B.R. (2002). Activation of Stat-3 as one of the early events in tobacco chewing-mediated oral carcinogenesis. Cancer 94, 2393-2400.. 38.
(51) 25. Masuda, M., Suzui, M., Yasumatu, R., Nakashima, T., Kuratomi, Y., Azuma, K., Tomita, K., Komiyama, S., and Weinstein, I.B. (2002). Constitutive activation of signal transducers and activators of transcription 3 correlates with cyclin D1 overexpression and may provide a novel prognostic marker in head and neck squamous cell carcinoma. Cancer Res 62, 3351-3355. 26. Cavarretta, I.T., Neuwirt, H., Untergasser, G., Moser, P.L., Zaki, M.H., Steiner, H., Rumpold, H., Fuchs, D., Hobisch, A., Nemeth, J.A., et al. (2007). The antiapoptotic effect of IL-6 autocrine loop in a cellular model of advanced prostate cancer is mediated by Mcl-1. Oncogene 26, 2822-2832. 27. Akira, S., Taga, T., and Kishimoto, T. (1993). Interleukin-6 in biology and medicine. Adv Immunol 54, 1-78. 28. Hong, S.H., Ondrey, F.G., Avis, I.M., Chen, Z., Loukinova, E., Cavanaugh, P.F., Jr., Van Waes, C., and Mulshine, J.L. (2000). Cyclooxygenase regulates human oropharyngeal carcinomas via the proinflammatory cytokine IL-6: a general role for inflammation? FASEB J 14, 1499-1507.. 39.
(52) 29. St John, M.A., Li, Y., Zhou, X., Denny, P., Ho, C.M., Montemagno, C., Shi, W., Qi, F., Wu, B., Sinha, U., et al. (2004). Interleukin 6 and interleukin 8 as potential biomarkers for oral cavity and oropharyngeal squamous cell carcinoma. Arch Otolaryngol Head Neck Surg 130, 929-935. 30. Nagata, M., Fujita, H., Ida, H., Hoshina, H., Inoue, T., Seki, Y., Ohnishi, M., Ohyama, T., Shingaki, S., Kaji, M., et al. (2003). Identification of potential biomarkers of lymph node metastasis in oral squamous cell carcinoma by cDNA microarray analysis. Int J Cancer 106, 683-689. 31. Okamoto, M., Hiura, K., Ohe, G., Ohba, Y., Terai, K., Oshikawa, T., Furuichi, S., Nishikawa, H., Moriyama, K., Yoshida, H., et al. (2000). Mechanism for bone invasion of oral cancer cells mediated by interleukin-6 in vitro and in vivo. Cancer 89, 1966-1975. 32. De Schutter, H., Landuyt, W., Verbeken, E., Goethals, L., Hermans, R., and Nuyts, S. (2005). The prognostic value of the hypoxia markers CA IX and GLUT 1 and the cytokines VEGF and IL 6 in head and neck squamous cell carcinoma treated by radiotherapy +/chemotherapy. BMC Cancer 5, 42.. 40.
(53) 33. Douglas, W.G., Tracy, E., Tan, D., Yu, J., Hicks, W.L., Jr., Rigual, N.R., Loree, T.R., Wang, Y., and Baumann, H. (2004). Development of head and neck squamous cell carcinoma is associated with altered cytokine responsiveness. Mol Cancer Res 2, 585-593. 34. Takahashi, O., Cai, Z., Toda, M., Hara, Y., and Shimamura, T. (1995). [Appearance of antibacterial activity of oxacillin against methicillin resistant Staphylococcus aureus (MRSA) in the presence of catechin]. Kansenshogaku Zasshi 69, 1126-1134. 35. Bu-Abbas, A., Nunez, X., Clifford, M.N., Walker, R., and Ioannides, C. (1996). A comparison of the antimutagenic potential of green, black and decaffeinated teas: contribution of flavanols to the antimutagenic effect. Mutagenesis 11, 597-603. 36. Graham, T.E., and Spriet, L.L. (1995). Metabolic, catecholamine, and exercise performance responses to various doses of caffeine. J Appl Physiol 78, 867-874. 37. Shixian, Q., VanCrey, B., Shi, J., Kakuda, Y., and Jiang, Y. (2006). Green. tea. extract. thermogenesis-induced. weight. loss. by. epigallocatechin gallate inhibition of catechol-O-methyltransferase. J Med Food 9, 451-458. 38. Polette, A., Lemaitre, D., Lagarde, M., and Vericel, E. (1996). N-3 fatty acid-induced lipid peroxidation in human platelets is prevented by catechins. Thromb Haemost 75, 945-949. 41.
(54) 39. Brown, M.D. (1999). Green tea (Camellia sinensis) extract and its possible role in the prevention of cancer. Altern Med Rev 4, 360-370. 40. Gao, Y.T., McLaughlin, J.K., Blot, W.J., Ji, B.T., Dai, Q., and Fraumeni, J.F., Jr. (1994). Reduced risk of esophageal cancer associated with green tea consumption. J Natl Cancer Inst 86, 855-858. 41. Gao, C.M., Takezaki, T., Wu, J.Z., Li, Z.Y., Liu, Y.T., Li, S.P., Ding, J.H., Su, P., Hu, X., Xu, T.L., et al. (2002). Glutathione-S-transferases M1 (GSTM1) and GSTT1 genotype, smoking, consumption of alcohol and tea and risk of esophageal and stomach cancers: a case-control study of a high-incidence area in Jiangsu Province, China. Cancer Lett 188, 95-102. 42. Way, T.D., Lee, H.H., Kao, M.C., and Lin, J.K. (2004). Black tea polyphenol theaflavins inhibit aromatase activity and attenuate tamoxifen resistance in HER2/neu-transfected human breast cancer cells through tyrosine kinase suppression. Eur J Cancer 40, 2165-2174. 43. Ahmad, N., Feyes, D.K., Nieminen, A.L., Agarwal, R., and Mukhtar, H. (1997). Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. J Natl Cancer Inst 89, 1881-1886.. 42.
(55) 44. Borska, S., Gebarowska, E., Wysocka, T., Drag-Zalesinska, M., and Zabel, M. (2003). Induction of apoptosis by EGCG in selected tumour cell lines in vitro. Folia Histochem Cytobiol 41, 229-232. 45. Chen, Z.P., Schell, J.B., Ho, C.T., and Chen, K.Y. (1998). Green tea epigallocatechin gallate shows a pronounced growth inhibitory effect on cancerous cells but not on their normal counterparts. Cancer Lett 129, 173-179. 46. Lambert, J.D., and Yang, C.S. (2003). Mechanisms of cancer prevention by tea constituents. J Nutr 133, 3262S-3267S. 47. Morse, M.A., Kresty, L.A., Steele, V.E., Kelloff, G.J., Boone, C.W., Balentine, D.A., Harbowy, M.E., and Stoner, G.D. (1997). Effects of theaflavins on N-nitrosomethylbenzylamine-induced esophageal tumorigenesis. Nutr Cancer 29, 7-12. 48. Otsuka, T., Ogo, T., Eto, T., Asano, Y., Suganuma, M., and Niho, Y. (1998). Growth inhibition of leukemic cells by (-)-epigallocatechin gallate, the main constituent of green tea. Life Sci 63, 1397-1403. 49. Stoner, G.D., and Mukhtar, H. (1995). Polyphenols as cancer chemopreventive agents. J Cell Biochem Suppl 22, 169-180. 50. Yang, C.S., and Wang, Z.Y. (1993). Tea and cancer. J Natl Cancer Inst 85, 1038-1049. 51. Hara, Y. (1994). Antioxidative action of tea polyphenols: Part 1. Am Biotechnol Lab 12, 48. 43.
(56) 52. Chen, H., and Tappel, A.L. (1995). Protection of vitamin E, selenium, trolox C, ascorbic acid palmitate, acetylcysteine, coenzyme Q0, coenzyme Q10, beta-carotene, canthaxanthin, and (+)-catechin against oxidative damage to rat blood and tissues in vivo. Free Radic Biol Med 18, 949-953. 53. Subirade, I., Fernandez, Y., Periquet, A., and Mitjavila, S. (1995). Catechin protection of 3T3 Swiss fibroblasts in culture under oxidative stress. Biol Trace Elem Res 47, 313-319. 54. Kuroda, Y. (1996). Bio-antimutagenic activity of green tea catechins in cultured Chinese hamster V79 cells. Mutat Res 361, 179-186. 55. Wu, Y.N., Wang, H.Z., Li, J.S., and Han, C. (1993). The inhibitory effect of Chinese tea and its polyphenols on in vitro and in vivo N-nitrosation. Biomed Environ Sci 6, 237-258. 56. Ohmori, Y., Ito, M., Kishi, M., Mizutani, H., Katada, T., and Konishi, H. (1995). Antiallergic constituents from oolong tea stem. Biol Pharm Bull 18, 683-686. 57. Valcic, S., Timmermann, B.N., Alberts, D.S., Wachter, G.A., Krutzsch, M., Wymer, J., and Guillen, J.M. (1996). Inhibitory effect of six green tea catechins and caffeine on the growth of four selected human tumor cell lines. Anticancer Drugs 7, 461-468.. 44.
(57) 58. Tokuda, H., Takai, S., Hanai, Y., Matsushima-Nishiwaki, R., Yamauchi, J., Harada, A., Hosoi, T., Ohta, T., and Kozawa, O. (2008). (-)-Epigallocatechin Gallate Inhibits Basic Fibroblast Growth Factor-stimulated Interleukin-6 Synthesis in Osteoblasts. Horm Metab Res. 59. Lin, J.K., and Liang, Y.C. (2000). Cancer chemoprevention by tea polyphenols. Proc Natl Sci Counc Repub China B 24, 1-13. 60. Shammas, M.A., Neri, P., Koley, H., Batchu, R.B., Bertheau, R.C., Munshi, V., Prabhala, R., Fulciniti, M., Tai, Y.T., Treon, S.P., et al. (2006).. Specific. killing. of. multiple. myeloma. cells. by. (-)-epigallocatechin-3-gallate extracted from green tea: biologic activity and therapeutic implications. Blood 108, 2804-2810. 61. Cao, Y., and Cao, R. (1999). Angiogenesis inhibited by drinking tea. Nature 398, 381. 62. Sachinidis, A., Seul, C., Seewald, S., Ahn, H., Ko, Y., and Vetter, H. (2000). Green tea compounds inhibit tyrosine phosphorylation of PDGF beta-receptor and transformation of A172 human glioblastoma. FEBS Lett 471, 51-55. 63. Liang, Y.C., Lin-shiau, S.Y., Chen, C.F., and Lin, J.K. (1997). Suppression of extracellular signals and cell proliferation through EGF receptor binding by (-)-epigallocatechin gallate in human A431 epidermoid carcinoma cells. J Cell Biochem 67, 55-65. 45.
(58) 64. Liang, Y.C., Chen, Y.C., Lin, Y.L., Lin-Shiau, S.Y., Ho, C.T., and Lin, J.K. (1999). Suppression of extracellular signals and cell proliferation by the black tea polyphenol, theaflavin-3,3'-digallate. Carcinogenesis 20, 733-736. 65. Lin, J.K., Liang, Y.C., and Lin-Shiau, S.Y. (1999). Cancer chemoprevention by tea polyphenols through mitotic signal transduction blockade. Biochem Pharmacol 58, 911-915. 66. Hara, Y. (1994). Antioxidative action of tea polyphenols: Part 1. Am Biotechnol Lab 12, 48. 67. Yang, G.Y., Liao, J., Kim, K., Yurkow, E.J., and Yang, C.S. (1998). Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 19, 611-616. 68. Gupta, S., Ahmad, N., Nieminen, A.L., and Mukhtar, H. (2000). Growth inhibition, cell-cycle dysregulation, and induction of apoptosis by green tea constituent (-)-epigallocatechin-3-gallate in androgen-sensitive. and. androgen-insensitive. human. prostate. carcinoma cells. Toxicol Appl Pharmacol 164, 82-90. 69. Khafif, A., Schantz, S.P., al-Rawi, M., Edelstein, D., and Sacks, P.G. (1998). Green tea regulates cell cycle progression in oral leukoplakia. Head Neck 20, 528-534.. 46.
(59) 70. Chow, H.H., Cai, Y., Hakim, I.A., Crowell, J.A., Shahi, F., Brooks, C.A.,. Dorr,. R.T.,. Hara,. Y.,. and. Alberts,. D.S.. (2003).. Pharmacokinetics and safety of green tea polyphenols after multiple-dose. administration. of. epigallocatechin. gallate. and. polyphenon E in healthy individuals. Clin Cancer Res 9, 3312-3319. 71. Li, H.C., Yashiki, S., Sonoda, J., Lou, H., Ghosh, S.K., Byrnes, J.J., Lema, C., Fujiyoshi, T., Karasuyama, M., and Sonoda, S. (2000). Green tea polyphenols induce apoptosis in vitro in peripheral blood T lymphocytes of adult T-cell leukemia patients. Jpn J Cancer Res 91, 34-40. 72. Ahmad, N., Feyes, D.K., Nieminen, A.L., Agarwal, R., and Mukhtar, H. (1997). Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. J Natl Cancer Inst 89, 1881-1886. 73. Paschka, A.G., Butler, R., and Young, C.Y. (1998). Induction of apoptosis in prostate cancer cell lines by the green tea component, (-)-epigallocatechin-3-gallate. Cancer Lett 130, 1-7. 74. Islam, S., Islam, N., Kermode, T., Johnstone, B., Mukhtar, H., Moskowitz, R.W., Goldberg, and Haqqi, T.M. (2000). Involvement of caspase-3 in epigallocatechin-3-gallate-mediated apoptosis of human chondrosarcoma cells. Biochem Biophys Res Commun 270, 793-797.. 47.
(60) 75. Jing, N., Zhu, Q., Yuan, P., Li, Y., Mao, L., and Tweardy, D.J. (2006). Targeting signal transducer and activator of transcription 3 with G-quartet oligonucleotides: a potential novel therapy for head and neck cancer. Mol Cancer Ther 5, 279-286. 76. Meydan, N., Grunberger, T., Dadi, H., Shahar, M., Arpaia, E., Lapidot, Z., Leeder, J.S., Freedman, M., Cohen, A., Gazit, A., et al. (1996). Inhibition of acute lymphoblastic leukaemia by a Jak-2 inhibitor. Nature 379, 645-648. 77. Chakravarti, N., Myers, J.N., and Aggarwal, B.B. (2006). Targeting constitutive and interleukin-6-inducible signal transducers and activators of transcription 3 pathway in head and neck squamous cell carcinoma cells by curcumin (diferuloylmethane). Int J Cancer 119, 1268-1275. 78. Lee, H.H., Ho, C.T., and Lin, J.K. (2004). Theaflavin-3,3'-digallate and penta-O-galloyl-beta-D-glucose inhibit rat liver microsomal 5alpha-reductase activity and the expression of androgen receptor in LNCaP prostate cancer cells. Carcinogenesis 25, 1109-1118. 79. Yu, C.X., Zhang, X.Q., Kang, L.D., Zhang, P.J., Chen, W.W., Liu, W.W., Liu, Q.W., and Zhang, J.Y. (2008). Emodin induces apoptosis in human prostate cancer cell LNCaP. Asian J Androl 10, 625-634.. 48.
(61) 80. Heo, S.K., Yun, H.J., Park, W.H., and Park, S.D. (2008). Emodin inhibits TNF-alpha-induced human aortic smooth-muscle cell proliferation via caspase- and mitochondrial-dependent apoptosis. J Cell Biochem. 81. Patel, D., Shukla, S., and Gupta, S. (2007). Apigenin and cancer chemoprevention: progress, potential and promise (review). Int J Oncol 30, 233-245. 82. Guruvayoorappan, C., and Kuttan, G. (2007). Rutin inhibits nitric oxide. and. tumor. necrosis. factor-alpha. production. in. lipopolysaccharide and concanavalin-a stimulated macrophages. Drug Metabol Drug Interact 22, 263-278. 83. Parkar, S.G., Stevenson, D.E., and Skinner, M.A. (2008). The potential influence of fruit polyphenols on colonic microflora and human gut health. Int J Food Microbiol 124, 295-298. 84. Matsui, N., Nakashima, H., Ushio, Y., Tada, T., Shirono, A., Fukuyama, Y., Nakade, K., Zhai, H., Yasui, Y., Fukuishi, N., et al. (2005). Neurotrophic effect of magnolol in the hippocampal CA1 region of senescence-accelerated mice (SAMP1). Biol Pharm Bull 28, 1762-1765.. 49.
數據

+3
相關文件
A Phase 2/3 Multicenter, Open-label, 3-arm, 2-stage Randomized Study of ASP2215 (Gilteritinib), Combination of ASP2215 Plus Azacitidine and Azacitidine Alone in the Treatment
In Sections 3 and 6 (Theorems 3.1 and 6.1), we prove the following non-vanishing results without assuming the condition (3) in Conjecture 1.1, and the proof presented for the
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
Table 5 - Domestic exports by section and division of SITC-Rev.3, and country/territory of destination Table 6 - Re-exports by section and division of SITC-Rev.3,
Table 5 - Domestic exports by section and division of SITC-Rev.3, and country/territory of destination Table 6 - Re-exports by section and division of SITC-Rev.3,
Table 5 - Domestic exports by section and division of SITC-Rev.3, and country/territory of destination Table 6 - Re-exports by section and division of SITC-Rev.3,
Table 5 - Domestic exports by section and division of SITC-Rev.3, and country/territory of destination Table 6 - Re-exports by section and division of SITC-Rev.3,
Consistent with the negative price of systematic volatility risk found by the option pricing studies, we see lower average raw returns, CAPM alphas, and FF-3 alphas with higher
