國 立 交 通 大 學
生 物 科 技 學 院
生 物 科 技 學 系
碩 士 論 文
利用工業煙道廢氣培養微藻對其生物質與
油脂產量之最佳化探討
Optimizations of microalgae cultivated with the flue gas from
steel plant on micaroalgal biomass and lipid productivity
研 究 生:黃 子 庭
指導教授:林 志 生 博士
利用工業煙道廢氣培養微藻對其生物質與油脂產量之最佳化探討
Optimizations of microalgae cultivated with the flue gas from steel plant on
micaroalgal biomass and lipid productivity
研 究 生:黃子庭 Student: Tzu-Ting Huang
指導教授:林志生 博士 Advisor: Chih-Sheng Lin, Ph D
國 立 交 通 大 學
生 物 科 技 學 院
生 物 科 技 學 系
碩 士 論 文
A Thesis
Submitted to Department of Biological Science and Technology College of Biological Science and Technology
National Chiao Tung University In partial fulfillment of the requirements
for the degree of M.S. in
Biological Science and Technology July 2012
Hsinchu, Taiwan, Republic of China
Acknowledgement
眨眼間,兩年就過去了。從剛開始進入實驗室懵懵懂懂的菜鳥,變成一個在內可以 對著自己專業領域侃侃而談的準碩士生,在外可以抬反應器、搬藻桶,甚至連鐵架的扛 得起來的壯漢。相信這一段充滿陽光、汗水以及屎臭味的光陰,讓原本的我更加成熟, 更有自信的面對未來的挑戰。這段求學路途中,最要感謝的就是我的指導老師林志生教 授,在您嚴厲的教導下,總是時時刻刻透露出對我們未來的關心,害怕我們將來出社會 沒有競爭力,所有教誨和叮嚀皆因恨鐵不成鋼,謝謝老師的指導,在這裡獻上最誠摯的 感謝。感謝這次的口委老師曾慶平教授和鍾竺均院長,因為你們而讓這場口試蓬蓽生輝, 給我很多十分受用的建議,謝謝你們的指導。 而過去的日子歷歷在目,首先我要先感謝實驗室的首領:棠青學長和千雅學姊。阿 關學長,謝謝你常常帶著我們上山下海,南征北討,且能用比較正面且客觀的態度去分 析各種事情,不僅在實驗上和做人處事上,都能用各種不同的角度去觀察,真的讓我學 到很多東西,很開心能在碩士班這兩年能遇到你!千雅學姊,你就像領航員一樣帶領我 們,不管是藻類的計畫,還是實驗室的事務,因為你一切才能如此的井然有序。再來我 要感謝我的兩位同學品萱和睦元。品萱,在這段實驗室的日子裡總是有你能和我一起分 享快樂與憂愁,做實驗寫論文的過程中總是會遇到很多挫敗,也是我們一起互相鼓勵, 一起挺過這段難關。睦元,感謝你平時當我的戰友,在我被許多人圍剿的時候,適時地 出現一起殺退敵人,希望我們以後都能武運昌隆。我們家唯一的藻類研究助理,戴樂, 有你的出現,讓我身上需要背負的藻液桶重量減輕很多,一起在炙熱太陽底下揮熱汗撒 熱血的日子,讓我們成為最好的 partner。還有兩位可愛的學妹,PCR 狂人燕秋和化工系 女孩芳沅,實驗室總是少不了你們兩個的笑聲,做實驗很認真的你們,相信以後也都能 順利畢業!另外還有現任專題生佩衡和一華,佩衡,其實你也算是和我同期進來實驗室 的同學,平常也只有你會和我一起耍白癡,少了你,我的研究生活一定會少了些色彩, 我想我以後會很想念和你品萱兩人一起聊天紓解壓力的日子的。也要感謝在我研究路途 中,曾經一起努力,一起歡笑的曜禎學長、佳蓉學姊、聖壹學長和俞任學長,很開心能 認識你們,常常收到你們如即時雨般的關心和祝福,祝福你們的人生能更加順遂。最後 祝福四位活潑的碩一新學妹,采郁、葛麗、意涵、碧珊,相信你們的認真努力,在將來 研究的路上也能順利。而最後要把這本論文獻給我的老爸和老媽,不是你們從小砸大錢花一大堆的心力在 我們兩兄弟上,我也沒機會來這裡讀這個碩士,你們一路上默默的努力和付出我都有看 到,謝謝你們,希望你們將來都能健健康康,快快樂樂的,也希望我未來的成就能讓你 們感到驕傲。 黃子庭 謹誌 國立交通大學 生物科技學系碩士班 中華民國一百零一年七月
利用工業煙道廢氣培養微藻對其生物質與油脂產量之最佳化探討
研 究 生:黃子庭 指導教授:林志生 博士
國 立 交 通 大 學
生 物 科 技 學 院
生 物 科 技 學 系 碩 士 班
摘 要
近年來,大氣中過量累積的二氧化碳成為全球暖化的主因,其中主要多為工廠所排 放含大量二氧化碳之工業廢氣,而微藻養殖則被視為解決此問題的方法之一。微藻可行 光合作用,利用光照做為能量,將二氧化碳和水轉化為生物質,達到二氧化碳減量與生 產生質能料源的目的。本研究應用本實驗室所篩選出之耐溫、高生長速率,且具高二氧 化碳耐受性的微藻株 Chlorella sp. TT-1,通入中鋼煙道廢氣做為碳源,進行廢氣養殖微 藻之試驗,用以產製微藻生物質,並萃取藻油生產生質柴油。本研究更結合反應曲面法 (response surface methodology, RSM) 進行微藻養殖,找出微藻之最適化培養條件,例如 微藻養殖之初始濃度、通入廢氣組成比率及廢氣通氣速率等。本研究利用不同廢氣比例組成的氣體進行微藻養殖,實驗結果顯示以廢氣稀釋為 25 %的混合氣體進行微藻培養時,可達到最佳的生長效率,其生物質產率可達 0.421 g/L/day;再則,以廢氣比率為 25 %所培養的微藻,其油脂累積量最多,可達約 40%的 油脂含量,因此在不同的廢氣組成比例的培養下,微藻的生長速率和產脂效應會有所差 異,不同的廢氣組成比率並不會顯著的影響微藻脂肪酸甲酯 (fatty acid methyl ester, FAME) 的組成。另外不同的廢氣通氣速率對微藻的生物質產率會有顯著性的影響,在 0.3 vvm 的通氣速率下微藻有最大的生物質產率,為 0.286 g/L/day;在油脂含量的部分, 在通氣速率 0.3 vvm 下的油脂累積可達約 40%;通氣速率對 FAME 的累積也有影響,於 0.3 vvm 之高通氣速率下有最多之 FAME,比低通氣速率下之微藻其 FAME 在微藻內的 含量多約 4~5 %。 為探討光照於利用工廠廢氣養殖微藻試驗上之影響,我們將養殖的 Chlorella sp.
TT-1 通入工廠廢氣,並以全日照和半日照兩種不同的光照模式進行微藻養殖。結果顯示 微藻之生物質產率在全日照和半日照下,分別為 0.299 g/L/day 和 0.137 g/L/day,明顯可 得知微藻在以廢氣培養下,全日照會有較高的產率;另外,微藻之油脂含量在全日照和 半日照下分別為 37 %和 15 %,可看出油脂的合成累積與光照時間是具有相關性;在 FAME 的部分,以半日照所培養之 Chlorella sp. TT-1 的飽和脂肪酸(C16:0)比例較多,而 以全日照培養則有較多的不飽和脂肪酸(C18:1 和 C18:2)的累積。 我們根據上述的實驗結果,結合反應曲面法進行微藻養殖最佳化探討,RSM 可做 實驗因子設計並模擬反應曲面,以較少的實驗成本和時間獲得可信賴且有效的資訊,並 可討論因子間的交互作用,進而探討多因子對實驗結果的影響性,並以實驗結果得一模 擬公式,找出最適之操作條件。本研究利用微藻培養初始濃度、廢氣組成比率及廢氣通 氣速率為實驗因子,探討微藻培養最適化之條件。實驗結果顯示,當微藻株在初始濃度 為 0.37 g/L、通氣量為 0.30 vvm 及工業煙道廢氣比率為 75 %時,可得到最佳的生物質產 率 0.486 g/L/day;在初始濃度為 0.35 g/L、通氣量為 0.24 vvm 及工業煙道廢氣比率為 75 %時,可得到最佳的油脂產率 0.216 g/L/day;在初始濃度為 0.37 g/L、通氣量為 0.25 vvm 及工業煙道廢氣比率為 74 %時,可得到最佳的脂肪酸甲酯產率 0.157 g/L/day。此外,在 上述實驗所得出之最佳微藻養殖參數下,我們另外再增加光照強度至 500 μmol/m2/s 和 700 μmol/m2/s,以探討其對生長速率和產脂效應的影響。在 500 μmol/m2/s 下微藻有最 大的生物質產率 0.390 g/L/day,且油脂累積可高達約 49 %,另外在 FAME 的組成部分, 500 μmol/m2/s 可顯著促進 C16:0 脂肪酸的累積。
Optimizations of microalgae cultivated with the flue gas from steel
plant on micaroalgal biomass and lipid productivity
Graduate student: Tzu-Ting Huang Advisor: Chih-Sheng Lin, Ph. D.
Department of Biological Science and Technology
College of Biological Science and Technology
National Chiao Tung University
Abstract
In the recent years, global warming becomes more serious problem due to the increasing carbon dioxide (CO2) accumulated in the atmosphere, and the plant steel for industry plays
the important role in emitting flue gas which is CO2-rich. Microalgae are the candidate to
solve the problem by photosynthesis, which use sun light as energy source to convert water and CO2 into biomass, and it can reduce the CO2 emission and produce biomass. We utilized
the isolated thermal- and CO2-tolerant mutant microalga Chlorella sp. TT-1 to reduce CO2 in
flue gas from the steel plant and produce microalgal biomass which can be extracted oil to produce biodiesel. Furthermore, we cultivated the microalgae combined with response surface methodology to get the optimized cultivation conditions under the specific initial density, aeration rate, and flue gas ratio.
To investigate the effect of flue gas ratio, there were different flue gas ratio gases utilized to study the effect of flue gas ratio in microalgal cultivation. The microalga Chlorella sp. TT-1 aerated with 25 % flue gas had more biomass productivity, which was 0.421 g/L/day. Lipid content of Chlorella sp. TT-1 cultures with 25 % flue gas ratio aeration were 40%. In the part of FAME production, there was no difference between the fatty acid methyl ester (FAME) content of Chlorella sp. TT-1 cultivated with different flue gas ratios. In the part about the effect of aeration rate, the biomass productivity of Chlorella sp. TT-1 at 0.3 vvm aeration rate has the maximum value, and it is 0.286 g/L/day. The maximum lipid contents of Chlorella sp. TT-1 was 40 % when it cultivated with 0.3 vvm aeration rates, and the experimental result was obtained that the biomass production and lipid content in microalga cells increased with the increasing aeration rate. The experimental result about FAME content showed it would
slightly increase with high aeration rate comparing to low aeration rate, and the variance was 4~5 % in total FAME content.
In order to investigate the effect of illumination in microalgae culture aerated with flue gas, Chlorella sp. TT-1 was cultivated under full and half illumination. The biomass
productivity of Chlorella sp. TT-1 in full and half illumination were 0.299 g/L/day and 0.137 g/L/day, respectively. The better irradiation time is full illumination for microalgal cultivation, and the light should be devised when the microalgae cultivated with flue gas. The lipid
content of Chlorella sp. TT-1 under full and half illumination were approximately 37 % and 15 %, respectively. Obviously, the lipid in microalgal cells under full illumination was more than it cultivated under half illumination. The saturated fatty acid (C16:0) accounted for the most part in Chlorella sp. TT-1 cultivated under half illumination, while the microalgae had more long and unsaturated fatty acid (C18:1 and C18:2) under full illumination.
Instead of enormous experiments to test, we cultivated microalga Chlorella sp. TT-1 combining response surface methodology which can reduce the experimental times to diminish the cost, and simulate the experimental formula to get the optimum condition for microalgal cultivation. The optimized biomass productivity of Chlorella sp. TT-1 is 0.486 g/L/day when the initial density of 0.37 g/Laerated with 75 % flue gas ratio at 0.30 vvm aeration rate; the optimized lipid productivity of Chlorella sp. TT-1 is 0.216 g/L when the initial density of 0.35 g/Laerated with 75 % flue gas ratio at 0.24 vvm aeration rate; the optimized FAME productivity of Chlorella sp. TT-1 is 0.157 g/Lwhen the initial density of 0.37 g/Laerated with 74 % flue gas ratio at 0.25 vvm aeration rate. In order to investigate whether the enhancement of the illumination is necessary in microalgal culture aerated with flue gas under the optimum condition of microalgal cultivation and the effect of different illumination intensity for microalgal cultivation, the different illuminations were utilized and irradiated for the microalgal cultivation with flue gas. The microalgal cultivation cultivated with 500 μmol/m2
/s had the maximum microalgae biomass productivity 0.390 g/L/day. The illumination of 500 μmol/m2/s could enhance the lipid accumulation, and it could obtain the lipid content of 49%. Furthermore, the culture irradiated under 500 μmol/m2/s had more C16:0 content, and it means higher illumination would induce microalgae to synthesize the saturated carbon compound of shorter chain.
Content
Acknowledgement……… i
Abstract in Chinese………... iii
Abstract in English………..……… v Content………..……… vii List of Figures...………..……….…... ix List of Tables.………..……….………… x I. Literature Review 1.1 General introduction……… 1 1.2 Microalgae………... 2 1.3 Microalgae cultivations...……… 5 1.4 Experimental design……… 12
1.5 Bio-mitigation of CO2, NOx and SOx in flue gas with microalgae.……….... 14
1.6 Biodiesel………... 18
II. Materials and Methods 2.1. Microalgal cultures………. 20
2.2. Culture medium and chemicals……….. 20
2.3. Experimental system of indoor photobioreactor……… 21
2.4. Preparation of the inoculums………... 21
2.5. Experiment design………... 21
2.6. Lipid extraction……….. 23
2.7. Transesterification……….. 23
2.8. Fatty acid profile analysis……….. 23
2.9. Analyses ……… 24
2.9.1 Microalgae cell counting………...………..………... 24
2.9.2. Measurement of growth rate………...……….………..…………... 24
2.9.3 Measurements of pH……….……...………..…… 25
2.9.5 Determinations of CO2(g) ………...……….………... 25
III. Results and Discussion 3.1 Profile of Chlorella sp. TT-1 cultivated with different flue gas ratio….………... 26
3.1.1 Growth profile in Chlorella sp. TT-1 cultures with different flue gas ratios……… 26
3.1.2 Lipid content and production in Chlorella sp. TT-1 cultures with different flue gas ratios…. 28 3.1.3 Lipid composition in Chlorella sp. TT-1 cultures with different flue gas ratios…………... 29
3.2 Profile of Chlorella sp. TT-1 cultivated at different aeration rate………... 29
3.2.1 Growth profile in Chlorella sp. TT-1 cultures with different aeration rates ……… 29
3.2.2 Lipid content and production in Chlorella sp. TT-1 cultures with different aeration rates... 30
3.2.3 Lipid composition in Chlorella sp. TT-1 cultures with different aeration rates……… 31
3.3 Microalgae cultivation under the illumination of full and half illumination aerated with flue gas 31 3.3.1 Growth profile of Chlorella sp. TT-1 under full and half illumination.……... 31
3.3.2 Lipid content and production of Chlorella sp. TT-1 under full and half illumination……... 32
3.3.3 Lipid composition of Chlorella sp. TT-1 under full and half illumination………... 33
3.4 The optimization of Chlorella sp. TT-1 aerated with flue gas in biomass productivity, lipid productivity, and FAME productivity…………... 34
3.4.1 The optimization of Chlorella sp. TT-1 in biomass productivity.……… 35
3.4.2 The optimization of Chlorella sp. TT-1 in lipid productivity………... 37
3.4.3 The optimization of Chlorella sp. TT-1 in FAME content………... 39
3.4.4 The predicted value and experimental productivity under optimum microalgae cultivation condition……… 40
3.5 Different light illumination used in the optimum microalgae cultivation………... 41
3.5.1 Growth profile of Chlorella sp. TT-1 cultivated with different illuminations………... 41
3.5.2 Lipid content and production of Chlorella sp. TT-1 cultivated with different illuminations... 42
3.5.3 Lipid composition and production of Chlorella sp. TT-1 cultivated with different illuminations……….. 42
IV. Conclusions ……….………..………... 44
V. References ……….………..……… 47
Figures……….………..………... 59
List of Figures
Figure 1-1 Central composite designs for the optimization of two variables and three variables.……… 59
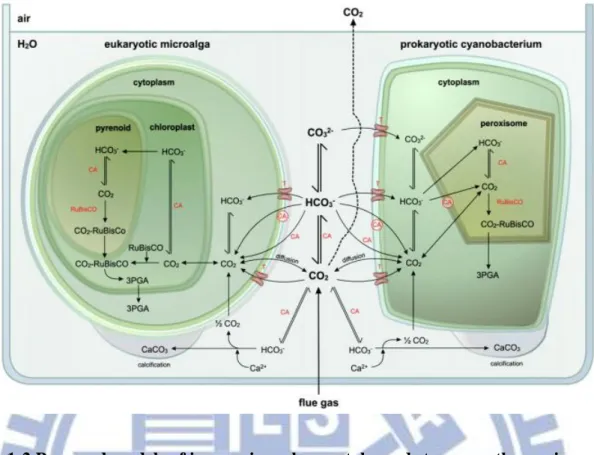
Figure 1-2 Proposed models of inorganic carbon uptake and storage pathways in eukaryotic microalgae and prokaryotic cyanobacteria.……… 60
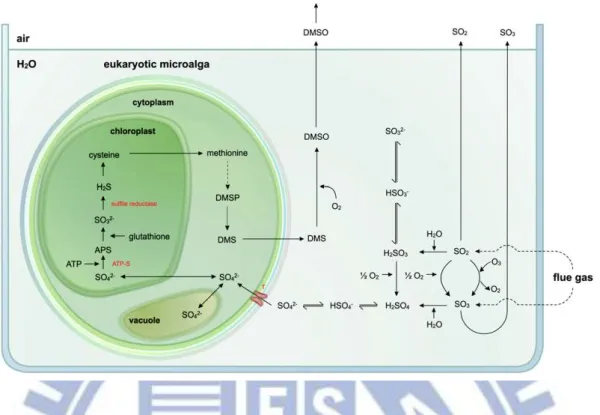
Figure 1-3 Proposed models for nitrogen uptake and storage by eukaryotic microalgae and prokaryotic cyanobacteria………... 61
Figure 1-4 Proposed models for sulphur uptake and storage by both prokaryotic cyanobacteria and eukaryotic microalgae. ………... 62
Figure 1-5 Transesterification of triglycerides ………... 63
Figure 2-1 The gas chromatography (GC) profile of fatty acid methyl ester (FAME) from microalgae... 64
Figure 2-2 Microalgae cultivation system………. 65
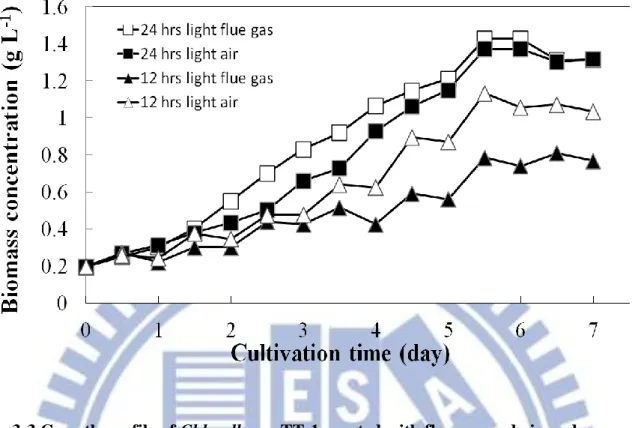
Figure 3-1 Growth profile of Chlorella sp. TT-1 aerated with different ratios of flue gas………... 66
Figure 3-2 Growth profile of Chlorella sp. aerated with different aeration rate of flue gas.………... 67
Figure 3-3 Growth profile of Chlorella sp. aerated with flue gas and air under different illumination time (12 hr and 24 hr)………... 68
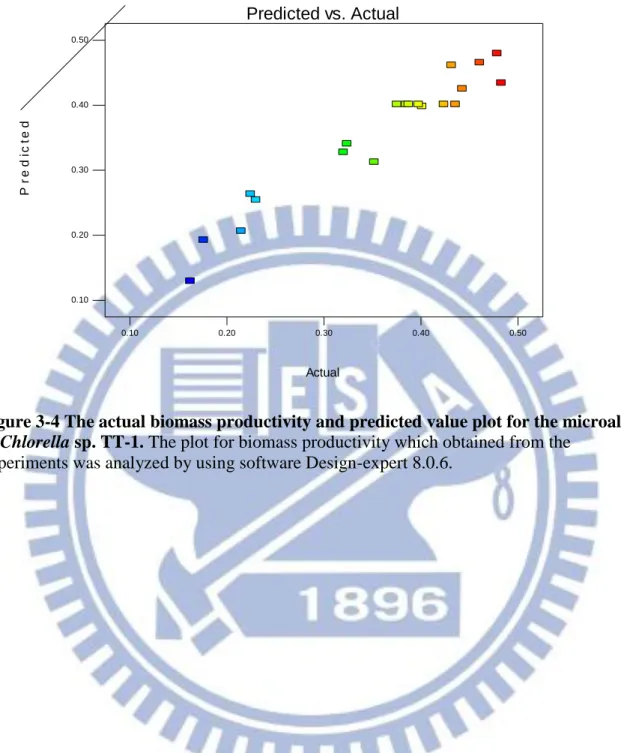
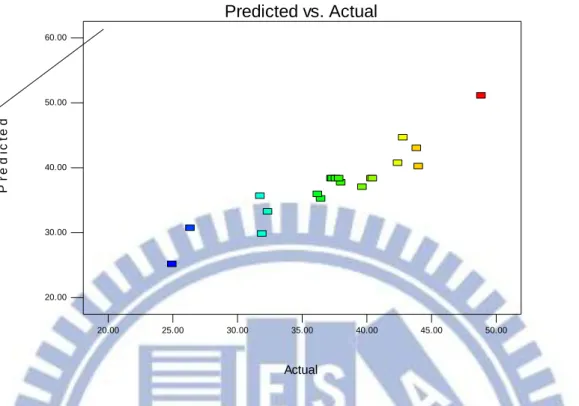
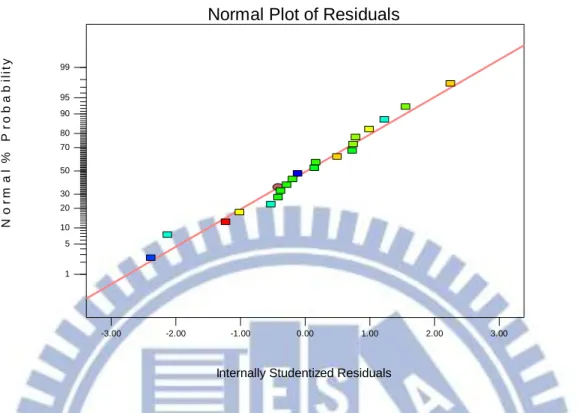
Figure 3-4 The actual biomass productivity and predicted value plot for the microalgae of Chlorella sp. TT-1 69 Figure 3-5The internally studentized residuals and normal % probability plot for the biomass productivity of Chlorella sp. TT-1………... 70
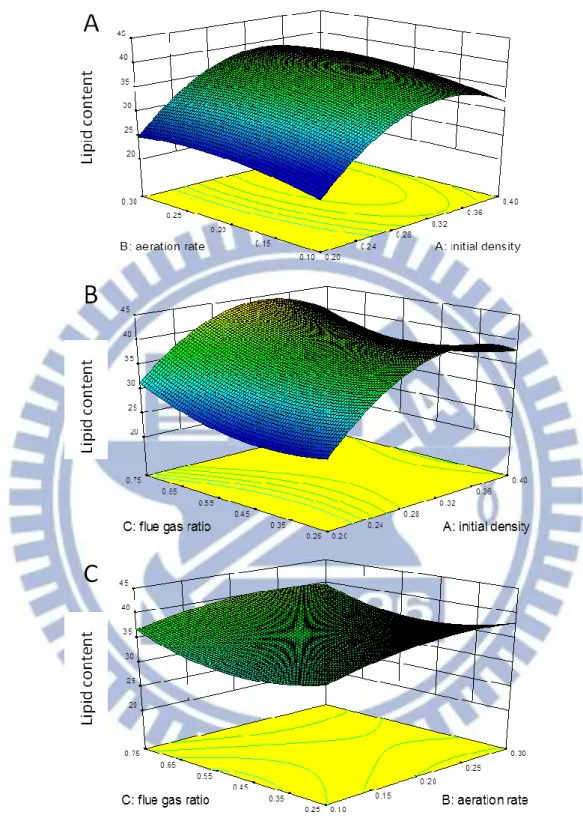
Figure 3-6 The 3D reaction surface plot for the biomass productivity of Chlorella sp. TT-1……….. 71
Figure 3-7 The actual lipid content and predicted value plot for the microalgae of Chlorella sp. TT-1……….. 72
Figure 3-8 The internally studentized residuals and normal % probability plot for the lipid content of Chlorella sp. TT-1.………... 73
Figure 3-9 The 3D reaction surface plot for the lipid content of Chlorella sp. TT-1.………... 74
Figure 3-10 The actual FAME content and predicted value plot for the microalgae of Chlorella sp. TT-1……. 75
Figure 3-11 The internally studentized residuals and normal % probability plot for the FAME content of Chlorella sp. TT-1.………... 76
Figure 3-12 The 3D reaction surface plot for the lipid content of Chlorella sp. TT-1……….. 77
List of Tables
Table 1-1 Present state of microalgae production………...………... 79
Table 1-2 Temperature and flue gas tolerance of various algal species………. 80
Table 1-3 Experimental matrices for central composite design: two variables and three variables ……... 81
Table 1-4 Comparison of microalgae with other biodiesel feedstocks……….. 82
Table 1-5 Comparison of properties of biodiesel, diesel fuel and ASTM standard………... 83
Table 3-1 Biomass and lipid production of Chlorella sp. TT-1 aerated with different ratios of flue gas………. 84
Table 3-2 Lipid composition profile of Chlorella sp. TT-1 aerated with different ratios of flue gas.………... 85
Table 3-3 Biomass and lipid production of Chlorella sp. TT-1 aerated at different aeration rates.…………... 86
Table 3-4 Lipid composition profile of Chlorella sp. TT-1 aerated at different aeration rates.……… 87
Table 3-5 Biomass and lipid production of Chlorella sp. TT-1 cultivated under full and half illumination……. 88
Table 3-6 Lipid composition profile of Chlorella sp. TT-1 aerated at different aeration rates………... 89
Table 3-7 Experimental factors and the values of each level for the optimization of Chlorella sp. TT-1 in biomass productivity, lipid content, and FAME content………... 90
Table 3-8 Experiment layout designed by Design-Expert 8.0.6 and the results expressing the biomass productivity, lipid content, and FAME content of Chlorella sp. TT-1 in different conditions……… 91
Table 3-9 ANOVA table about the biomass productivity of Chlorella sp. TT-1 for the quadratic model……… 92
Table 3-10 Parameter values found for the quadratic model about the biomass productivity of Chlorella sp. TT-1 and their significances………...……….. 93
Table 3-11 ANOVA table about the lipid content of Chlorella sp. TT-1 for the quadratic model…………...… 94
Table 3-12 Parameter values found for the quadratic model about the lipid content of Chlorella sp. TT-1 and their significances………... 95
Table 3-13 ANOVA table about the FAME content of Chlorella sp. TT-1 for the quadratic model………….. 96
Table 3-14 Parameter values found for the quadratic model about the FAME content of Chlorella sp. TT-1 and their significances………... 97
Table 3-15 The predicted value and experimental productivity under optimum microalgae cultivation condition……… 98
Table 3-16 Biomass and lipid production of Chlorella sp. TT-1 cultivated under different illumination……… 99
I. Literature Review
1.1 General introduction
The industry revolution from 18th century brought with the rapid economic development, and it improved our living standard. However, there are enormous by-products like
greenhouse gas accumulated in the atmosphere. These greenhouse gases absorb and emit radiation at specific wavelengths within the spectrum of thermal infrared radiation emitted by the earth’s surface, and this property results in the greenhouse effect. Originally, the
greenhouse gases, which include water vapor, carbon dioxide (CO2), methane (CH4), nitrous
oxide (NOx), ozone (O3) and chlorofluorocarbons, play the important roles in maintaining the
temperature of the earth, and its character makes the environment of the earth habitable for livings [Karl et al., 2003]. However, more and more greenhouse gases especially CO2 are
emitted, and it makes the heat accumulated in the atmosphere. Global warming, which is induced by increasing concentrations of greenhouse gases in the air, is of great concern [Watanabe et al., 1996]. CO2 is the principle greenhouse gas, and the global temperature is
rapidly rising with the increasing concentration of CO2. The global atmospheric concentration
of CO2 has increased from 280 ppm of a pre-industrial value to 390 ppm in 2010 [McGin et
al., 2011]. This causes the drastic climate change, and it makes a lot of organisms endangered. In addition, energy crisis is another serious problem.
There are various strategies on CO2 sequestration which have been carried out, such as
physical, chemical, and biological methods. There are some examples of physical method like wet absorption or dry adsorption, and geological storage of CO2, which CO2 is injected into
reservoirs [Granite et al., 2005; Yang et al., 2008; Favre et al., 2009; Ball et al., 2010; Aydin et al., 2010]. However, to use these techniques, purified CO2 is required, and it is expensive to
separate and purify CO2 from the industrial emissions such as flue gas. It is very necessary
and valuable to develop a technique which can directly remove or fix CO2 from the flue gas.
Whereas the examples chemical methods are by washing with alkaline solutions, multiwalled carbon nanotubes, amine coating activated carbon for CO2 capture in order to reduce the
emission of CO2. The biological method using microalgae photosynthesis is considered as an
effective approach for biological CO2 fixation. By the biological approaches, CO2 can be fixed
into microalgae biomass via photosynthesis. Microalgae has approximately 10~50 times higher CO2 fixation rates than terrestrial plants and can thus utilize CO2 from flue gas to
produce biomass.Brennan showed One kilogram of algal dry cell weight utilizes around 1.83 kg of CO2. Annually around 54.9 ~ 67.7 tones of CO2 can be sequestered from raceway ponds
corresponding to annual dry weight biomass production rate of 30~37 tons per hectare [Brennan et al., 2010]. Therefore, reduction of the emission from industries or power plants by the use of microalgae incorporated photobioreactor is a potential method for removing CO2 from waste gas.
In addition, microalgae can use CO2 efficiently and accumulate lipid which is chemically
similar to common vegetable oils. Furthermore, it is high potential source for biodiesel
production. Instead of fossil fuels, the biodiesel produced from microalgae is more renewable, biodegradable, and low pollutant produced. Thus, reducing atmospheric CO2 by microalgae
photosynthesis is considered safe and reliable for nature.
1.2 Microalgae
Microalgae cover all unicellular and simple multi-cellular microorganisms, including both prokaryotic microalgae and eukaryotic microalgae. In a multistep process of
photosynthesis plants and algae (green algae and cyanobacteria) fix CO2 into sugar using light
and water as energy and electron source, respectively. The overall reaction for photosynthesis is given by:
CO2 + H2O + light → (CH2O)n + O2
They can grow almost anywhere, requiring sunlight and some simple nutrients, although the growth rates can be accelerated by the addition of specific nutrients and sufficient
aeration.
Microalgae can either be autotrophic or heterotrophic. The former requires only inorganic compound such as CO2, salts and light energy source for growth while the latter
which is non-photosynthetic requires external source of organic compounds for nutrients as energy source. Sometimes microalgae can be autotrophic or heterotrophic when it exists in different condition, and it is called as mixotrophilic. It performs photosynthesis as the main energy source, though both organic compounds and CO2 are essential. Amphitrophy, subtype
of mixotrophy, means that organisms are able to live either autotrophically or
available. Photoheterotrophycally, also known as photoorganitrophy, photoassimilation, photometabolism, describes the metabolism in which light is required to use organic
compounds as carbon source. The photoheterotrophic and mixotrophic metabolisms are not well distinguished, in particular they can be defined according to a difference of the energy source required to perform growth and specific metabolite production.
Microalgae are present in all existing earth ecosystems, not just aquatic but also terrestrial, representing a big variety of species living in a wide range of environmental conditions. It is estimated that more than 50,000 species exist, but only a limited number, of around 30,000, have been studied and analyzed. While among the 10,000 species which are believed to exist, only a few thousand strains are kept in collections, a few hundred are investigated for chemical content and just a handful are cultivated in industrial quantities [Richmond, 2004; Olaizola, 2003]. This collection attests to the large variety of different microalgae available to be selected for use in a broad diversity of applications, such as value added products for pharmaceutical purposes, food crops for human consumption and energy source.
Many research reports and articles described many advantages of using microalgae for biodiesel production in comparison with other available feed stocks, like microalgae are considered to be a very efficient biological system for harvesting solar energy for the production of organic compounds. Microalgae are non-vascular plants, lacking complex reproductive organs, and many species of algae can be induced to produce particularly high concentrations of chosen, commercially valuable compounds, such as proteins, carbohydrates, lipids and pigments. Microalgae are microorganisms that undergo a simple cell division cycle. The farming of microalgae can be grown using sea or brackish water, and microalgae biomass production systems can easily be adapted to various levels of operational or technological skills.
Production of biodiesel and other bio-products from microalgae can be more
environmentally sustainable, cost-effective and profitable, if combined with processes such as wastewater and flue gas treatments. In fact various studies demonstrated the use of microalgae for production of valuable products combined with environmental applications. In addition, depending on the microalgae species various high-value chemical compounds may be extracted such as pigments, antioxidants, β-carotenes, polysaccharides, triglycerides, fatty acids, vitamins, and biomass, which are largely used as bulk commodities in different
industrial sectors. These materials are utilized in pharmaceuticals, cosmetics, nutraceuticals, functional foods, and biofuels fields, and the application of microalgae in producer country are listed in Table 1-1[Lorenz et al., 2000; Hejazi et al., 2004; Pulz et al., 2004; Ratledge, 2004; Spolaore et al., 2006a; Loubiere et al., 2009]. For examples, some microalgae have been exploited for millennia (Nostoc in China and Arthrospira in Chad and Mexico).
Currently, they have several applications from human and animal nutrition to cosmetics and the production of high-value molecules (fatty acids, pigments, and stable isotope
biochemicals). Recently the utilization of microalgae can serve enormous purposes which are potential and feasible. Some possibilities currently being considered are listed below [Mata et al., 2010].
Microalgae can provide feedstock for several different types of renewable fuels such as biodiesel, methane, hydrogen, ethanol, among others. Algae biodiesel contains no sulfur and performs as well as petroleum diesel, while reducing emissions of particulate matter, CO, hydrocarbons, and SOx. However emissions of NOx may be higher in some engine
types [Delucchi, 2008].
Removal of CO2 from industrial flue gases by microalgae bio-fixation, reducing the
GHG emissions of a company or process while producing biodiesel [Wang et al., 2008]. Wastewater treatment by removal of NH4+, NO3-, PO43-, making algae to grow using
these water contaminants as nutrients [Wang et al., 2008].
After oil extraction the resulting algae biomass can be processed into ethanol, methane, livestock feed, used as organic fertilizer due to its high N:P ratio, or simply burned for energy cogeneration (electricity and heat) [Wang et al., 2008]
Combined with their ability to grow under harsher conditions, and their reduced needs for nutrients, they can be grown in areas unsuitable for agricultural purposes
independently of the seasonal weather changes, thus not competing for arable land use, and can use wastewaters as the culture medium, not requiring the use of freshwater. Depending on the microalgae species other compounds may also be extracted, with
valuable applications in different industrial sectors, including a large range of fine chemicals and bulk products, such as fats, polyunsaturated fatty acids, oil, natural dyes, sugars, pigments, antioxidants, high-value bioactive compounds, and other fine
chemicals and biomass [Li et al., 2008a; Li et al., 2008b; Raja et al., 2008]. Because of this variety of high-value biological derivatives, with many possible
commercial applications, microalgae can potentially revolutionize a large number of biotechnology areas including biofuels, cosmetics, pharmaceuticals, nutrition and food additives, aquaculture, and pollution prevention [Rosenberg et al., 2008; Raja et al., 2008].
Heterotrophic and mixotrophic cultivation could be a possible avenue of research. The genetic improvement of microalgae strains is also a present challenge. The use of transgenic microalgae for commercial applications has not yet been reported but holds significant promise. Modified strains could overproduce traditional or newly discovered algae
compounds and also serve to express specific genes that cannot be expressed in yeast. This could be of great importance for the production of hydrogen, for example. However, a successful drug discovery is the most promising aspect of microalgae biotechnology because the potential is immense although screening remains limited [Tramper et al., 2003].
1.3 Microalgae cultivation
There are several factors influencing algal growth abiotic factors such as light (quality, quantity), temperature, nutrient concentration, O2, CO2 , pH, salinity, and toxic chemicals;
biotic factors such as pathogens (bacteria, fungi, and viruses) and competition by other algae; operational factors such as shear produced by mixing, dilution rate, depth, harvest frequency, and addition of bicarbonate.
1.3.1 Temperature
Temperature is the most important limiting factor, after light, for culturing algae in both closed and open outdoor systems, and it even regulates cellular, morphological and
physiological responses of microalgae. Higher temperatures generally accelerate the metabolic rates of microalgae, whereas low temperatures lead to inhibition of microalgae growth [Munoz et al., 2006]. The optimal temperature varies among microalgae species. Whereas, Many microalgae can easily tolerate temperatures up to 15 ℃ lower than their optimal, but exceeding the optimum temperature by only 2~4 ℃ may result in the total
culture loss. Furthermore, overheating problems may occur in closed culture systems during some hot days, where the temperature inside the reactor may reach 55 ℃. In the application of removing the CO2 and other oxides in flue gas emitted from power plants, temperature of flue
gas and other sources are around 120 ℃. Feasibility of sequestering CO2 from flue gas
depends on either installing heat exchanger system or using thermophilic species. Several species have been identified which can tolerate high temperature up to 60 ℃ (Table 1-2).
1.3.2 Illumination
Sunlight is the most common source of energy for the CO2 fixation and biomass
production optimum of microalgae, to an extent that is rather species-dependent. The light intensity would be the limiting factor if the irradiation does not achieve the level to carry out photosynthesis, and microalgae productivity becomes proportional to the light conversion efficiency [Richmond et al., 2003]. Light intensity requirements of typical microalgae are relatively low compared with those of higher plants, microalgae activity usually rises, with increasing light intensity, up to 400 μmol/m2/s. However, a strong species-dependence exists that should be taken into account [Munoz et al., 2006; Kumar et al., 2010]; as an example, the saturating light intensity of Chlorella and Scenedesmus sp. is of the order of 200 μmol/m2/s [Hanagata, 1992]. While exposure of cells to long period with high light intensity causes photoinhibition due to damage of repair mechanism of photosystem II leading to inactivation of other systems including the oxygen evolving systems, electron carriers and the associated D1/D2 proteins [Rubio et al., 2003]. Whereas, Light above a saturation point causes light inhibition, which can be counterbalanced by exposing microalgae cells to very short cyclic periods of light and darkness [Pulz, 2001]. The ratio of light to dark (or low-intensity light) periods in a cycle is crucial for microalgae productivity [Munoz et al., 2006]. When the light/dark cycle period approaches the photosynthetic unit turnover time (equal to the dark reaction time, estimated to lie within 1~15 ms), maximum photosynthetic efficiencies can be achieved [Richmond et al., 2003]. Furthermore, periods of low light intensity significantly increase growth, CO2 assimilation and lipid productivity in microalgae for a given
illumination compared with periodic darkness. This type of irradiation design can be achieved via artificial light, such as hybrid lighting systems [Chen et al., 2011].
The spatial distribution of the light intensity inside the reactor is, apart from the geometry, mainly influenced by light attenuation caused by mutual shading of the cells via
adsorption by the pigments or via scattering by the cells. Some mechanistic formulae for calculating light gradients in liquid particle systems have been published, however, for a small and flat volume element an exponential development depending on biomass
concentration can be assumed [Posten, 2009; Cornet et al., 1995]. According to Eq. (1), the light intensity decreases exponentially with distance from a reactor wall as the concentrations of both cell and product increase
(1) Where IL is the light intensity at depth L, Io is the original incident intensity, c is the
turbidity. Hence, the light intensity tends to decrease rapidly due to the light shading effects arising from increases in the concentrations of both cell and product or from formation of biofilm on the surface of reactor vessels [Chen et al., 2008; Chen et al., 2011]. Because of the problems and limitations associated with conventional light sources, various photobioreactor designs with different illumination strategies have been developed to enhance the microalgae production rate and oil/lipid content [Ma et al., 1999]. Fernandes studied the effect of circular and plan geometry in light penetration. For similar microalgae cell concentrations, circular geometry allows a better light penetration, than the plane geometry allowing a higher volume fraction of the reactor to receive sufficient amounts of light however, plan geometry helps in uniform distribution of light [Fernandes et al., 2010].
Different lamps generate distinct spectra, and different microalgae species possess dissimilar absorption optima; therefore, each individual case should be studied before deciding on the set point of this important operational parameter [Kumar et al., 2010; Suh et al., 2003a]. In the recent years, LED is the new potential choice to get the cheap, durable, reliable, and highly efficient light source. LEDs which have narrow light emission spectra between 20 and 30 nm can be matched with photosynthetic needs which can match different microalgae species for different appropriate spectrum. For instance, the adsorption
wavelength of blue LED and red LED are around 450~470 nm and 645~665 nm, respectively [Yeh et al., 2009]. Wang found that the highest specific growth rate and biomass production were obtained by using red LED in the photoautotrophic cultivation of Spirulina platensis [Wang et al., 2007].
1.3.3 Aeration rate
Aeration rate plays the important role in both dissolved inorganic carbon and mixing which influence the microalgae cultivation much. The effects of carbon oxide uptake are different under different aeration rates. In addition, different aeration rates will make the different ratios of CO2 dissolve in medium, and this phenomenon will improve the efficiency
to utilize CO2 as inorganic carbon source for microalgae.
In the perspective of mixing, low mixing rates hamper gaseous mass transfer and might even permit biomass settling. In either case, poor mixing leads to emergence of stagnant zones, where light and nutrients are insufficiently available and anoxic/anaerobic conditions will thus prevail, which results in a decrease of productivity. Culture viability might also be compromised by production and accumulation of toxic compounds in stagnant zones [Becker, 1994]. However, High liquid velocities and high degrees of turbulence in photobioreactors can damage microalgae due to shear stress, and shear damage is sometimes used as an argument against mechanical mixing in microalgae cultures [Barbosa et al., 2003;
Vega-Estrada et al., 2005; Carlsson et al., 2007]. Furthermore, Kaewpintong reported a better growth for an airlift bioreactor than for an unaerated column. This is also explained because aeration lead to a better mixing of the microalgae culture, which prevents sedimentation, maintains homogeneous conditions, and helps for a better contact between cells and nutrients [Kaewpintong, 2004].
In the researches of aeration rate for microalgae cultivation, Ryu demonstrated that the high aeration rate resulted in the high cell concentration of Chlorella sp. AG10002 and the low CO2 utilization efficiency. The optimum aeration rate in consideration of both the
productivity and the CO2 utilization efficiency was 0.20 vvm [Ryu et al., 2009]. Also, Chiu
investigated that significant higher of CO2 fixation rate was also found when the aeration rate
decreases in the result of the increasing of CO2 absorption from bubbling gas. This was
caused by the increase of surface area per unit gas volume of the bubble which would also enhance the CO2 removal efficiency [Chiu et al., 2009b].
1.3.4 Culture pH
Most microalgae species are favored by neutral pH, and have a narrow optimal range of pH. The pH of the culture medium is influenced by dissolving CO2 concentration and even
and with higher SOx concentrations even down to pH = 2.6 have been reported [Maeda et al.,
1995]. Some species are tolerant to higher pH (e.g. Spirulina platensis at pH = 9 [Hu et al., 1998b]) or lower pH (e.g. Chlorococcum littorale at pH = 4 [Kumar et al., 2010]). There is a complex relationship between CO2 concentration and pH in microalgae bioreactor systems,
owing to the underlying chemical equilibrium among such chemical species as CO2, H2CO3,
HCO3- and CO32-, and each microalgae strain is appropriate to assimilate different inorganic
compound.
1.3.5 Culture density
Both productivity and light utilization efficiency value are the functions of the cell density. It is crucial to select the optimum cell concentration for the efficient CO2
sequestration. Below the optimum cell concentration, not all the light energy is captured by the cells while at above the optimum cell concentration, a larger proportion of the cell are in the dark due to self-shading which leads the productivity declining due to mainly respiration [Zhang et al., 2001; Kumar et al., 2010]. Light intensity decreases deeper within the culture medium, especially in high-density cultures; hence, the issue of optical depth, which measures the proportion of radiation absorbed or scattered along a path through a partially transparent medium, should be considered in microalgae bioreactor design. The relationship between biomass productivity and cell concentration has been reported in flat inclined modular and vertical flat plate photobioreactor by Hu and Zhang, respectively [Hu et al., 1996; Zhang et al., 2001]. However, highly dense culture also makes cells more tolerant to high percentage of CO2 concentration [Chiu et al., 2008].
1.3.6 CO2 concentration
In addition to light and water, carbon dioxide (CO2) is critical for photosynthesis in
higher plants and microalgae [Li et al., 2008a; Tredici, 2010; Chisti, 2007]. Atmospheric CO2
levels (0.0387 %) are not sufficient to support the high microalgae growth rates and productivities needed for large-scale biofuel production. In principle, flue gas from
combustion process typically contains above 15 % CO2, and this value indicates that flue gas
will supply enough carbon sources for large-scale microalgae production. Whereas, algae cells can tolerate CO2 only up to a certain level after which it becomes detrimental for the
growth of the cells because of the two reasons. Firstly environmental stress induced by the higher CO2 concentration which causes biological reduction in the capacity of algal cells for
CO2 sequestration [Sobczuk et al., 2000; Kumar et al., 2010]. Secondly at higher CO2
concentration, the culture pH decreases due to the formation of high amount of bicarbonate buffer (which is described in the part of 1-3-4 pH). The biomass productivity increases with increase in CO2 % in the gas mixture up to certain percentage beyond which productivity
decreases. CO2 sequestration experiment operated by Chiu at a flow rate of 0.25 vvm reports
that 2 % of CO2 is optimum for the growth of Chlorella while at 10 % specific growth rate
becomes insignificant [Chiu et al., 2008]. However, the experiment conducted by Maeda for the sequestration of CO2 from flue gas emitted by coal fired thermal power plant confirms
that Chlorella sp. T-1 can tolerate up to 100 % CO2 concentration but the maximum growth
rate was obtained when using 10 % CO2 with no significant decreasing growth rate up to
50 % CO2 concentration [Maeda et al., 1995]. They also concluded that pre-adaptation of
cells with lower percentage of CO2 concentration leads the tolerability of cells in higher
percentage of CO2 [Kumar et al., 2011].
1.3.7 O2 accumulation
Photosynthesis is a reversible set of reactions, and excessive dissolved oxygen, DO ( >35 mg/L), can inhibit the metabolic processes [Carvalho et al., 2006]. The water splitting activity of photosystem II is responsible for the oxygen evolution during photosynthesis. Trapped oxygen in the liquid culture causes toxic effects like photo-bleaching and reduces the
photosynthetic efficiency. Furthermore, microalgae are negatively charged on their surface, so they can strongly adsorb polyvalent cations; this ion exchange capacity is the basis of the microalgae potential to remove heavy metals from wastewaters [Munoz et al., 2006]. However, heavy metals are potent inhibitors of microalgae photosynthesis because they can replace or block the prosthetic metal atoms in the active site of relevant enzymes, or otherwise induce morphological changes in the microalgae cells that lead to physiological
incompatibility [Kumar et al., 2010].
An efficient degassing system is required in order to remove formed O2. Accumulation
of O2 is a serious problem in reactors with poor gas exchange like horizontal tubular reactors,
especially when continuous run tubing increases [Miron et al., 1999]. The problem of accumulation of O2 increases when a helical tubular reactor is scaled-up by increasing the
light harvesting unit. Hence it is necessary to have a separate degassing unit in which the distance between the entrance and exit is such that even smallest bubbles can disengage. To prevent inhibition and damage, the maximum tolerable dissolved oxygen level should not
generally exceed about 400 % of air saturation value [Chisti, 2007]. It is not of major concern in reactors which have an open gas transfer area as in stirred tank and vertical reactors
[Kumar et al., 2011].
1.3.8 Nutrient
Except for carbon source from organic (sugar or glucose) and inorganic carbon source (CO2), vitamins, salts and other nutrients like nitrogen and phosphorous are vital for algal
growth [Williams, 2002; Suh et al., 2003b]. Growth medium must provide the inorganic elements that constitute the algal cell. Nitrogen is the most important element that is required for microalgae nutrition, as a constituent of both nucleic acids and proteins, and nitrogen is directly associated with the primary metabolism of microalgae [Becker, 1994]. Fast-growing microalgae species prefer ammonium rather than nitrate as a primary nitrogen source [Green et al., 1996]; intermittent nitrate feeding, however, will enhance microalgae growth if a medium that lacks nitrate is used [Jin et al., 2006]. Under partial nitrogen deprivation,
microalgae grow at lower rates (as expected), but produce significantly more lipids, which are reserve compounds synthesized under stress conditions, even at the expense of lower
productivities [Lardon et al., 2009; Kumar et al., 2010].
Phosphorus is the third most important nutrient for microalgae growth, and should be supplied to significant excess as phosphates because not all phosphorus compounds are bioavailable (e.g. those combined with metalions) [Kumar et al., 2009]. In the case of marine microalgae, seawater supplemented with commercial nitrate and phosphate fertilizers is commonly used for production of microalgae [Green et al., 1996]. Nevertheless, trace species, such as metals (Mg, Ca, Mn, Zn, Cu and Mb) and vitamins, are typically added for effective cultivation [Becker, 1994].
Minimal nutritional requirements can be estimated using the approximate molecular formula of the microalgae biomass, which is CO0.48H1.83N0.11P0.01. This formula is based on
data presented by Grobbelaar [Chisti, 2007]. Nutrients such as phosphorus must be supplied in significant excess because the phosphates added complex with metal ions, therefore, not all the added P is bioavailable. Sea water supplemented with commercial nitrate and phosphate fertilizers and a few other micronutrients is commonly used for growing marine microalgae [Grima et al., 1999; Kumar et al., 2010].
1.4 Response surface methodology
The Response Surface Methodology (RSM) is a collection of mathematical and statistical techniques useful for the modeling and analysis of problems in which a response of interest is influenced by several variables and the objective is to optimize this response [Montgomery, 2006]. It is important in designing, formulating, developing, and analyzing new scientific studying and products. The RSM is important in designing, formulating, developing, and analyzing new scientific studying and products. It is also efficient in the improvement of existing studies and products. There are many applications of RSM in industrial, biological, clinical, social, food, physical and engineering sciences.
According to Hill and Hunter, RSM method was introduced by G.E.P. Box and K.B. Wilson in 1951 [Bruns et al., 2010]. Box and Wilson suggested to use a first-degree
polynomial model to approximate the response variable. They acknowledged that this model is only an approximation, not accurate, but such a model is easy to estimate and apply, even when little is known about the process. Moreover, Mead and Pike stated origin of RSM starts 1930s with use of Response Curves.
The orthogonal design was motivated by Box and Wilson (1951) in the case of the first-order model. For the second-order models, many subject-matter scientists and engineers have a working knowledge of the central composite designs (CCDs) and three-level designs by Box and Behnken (1960). Also, the same research states that another important
contribution came from Hartley (1959), who made an effort to create a more economical or small composite design. There are many papers in the literatures about the response surface models. In contrast, 3-level fractional design has limited works. Thus, 3-level fractional design is an open research subject. Fractional factorial experiment design for factor at 3-levels is a helpful resource conducting this kind of design. Many three-level fractional factorial designs and more importantly their alias tables can be found in their study.
The important development of optimal design theory in the field of experimental design emerged following Word World II. Elfving (1952, 1955, 1959), Chernoff (1953), Kiefer (1958, 1959, 1960, 1962), and Kiefer and Wolfowitz were some of the various authors who published their work on optimality. One of the important facts is whether the system contains a maximum or a minimum or a saddle point, which has a wide interest in industry. Therefore, RSM is being increasingly used in the industry. Also, in recent years more emphasis has been
placed by the chemical and processing field for finding regions where there is an
improvement in response instead of finding the optimum response. In result, application and development of RSM will continue to be used in many areas in the future [Stephen M, 1974]
There is an example to explain the principle of response surface methodology. The growth of a plant is affected by a certain amount of water x1 and sunshine x2, and the plant
can grow under any combination of treatment x1 and x2. Therefore, water and sunshine can
vary continuously. When treatments are from a continuous range of values, then a Response Surface Methodology is useful for developing, improving, and optimizing the response variable. In this case, the plant growth y is the response variable, and it is a function of water and sunshine. It can be expressed as
y = f ( x1 , x2 ) + e
The variables x1 and x2 are independent variables where the response y depends on
them. The dependent variable y is a function of x1, x2, and the experimental error term,
denoted as e. The error term e represents any measurement error on the response, as well as other type of variations not counted in f. It is a statistical error that is assumed to distribute normally with zero mean and variance s2. In most RSM problems, the true response function f is unknown. In order to develop a proper approximation for f, the experimenter usually starts with a low-order polynomial in some small region. If the response can be defined by a linear function of independent variables, then the approximating function is a first-order model. A first-order model with two independent variables can be expressed as
y = β0 + β1 x1 + β2 x2 + ε
If there is a curvature in the response surface, then a higher degree polynomial should be used. The approximating function with two variables is called a second-order model:
y = β0 + β1 x1 + β2 x2 + β11 x112 +β22 x222 +β12 x1x2 + ε
In general all RSM problems use either one or the mixture of the both of these models. In each model, the levels of each factor are independent of the levels of other factors. In order to get the most efficient result in the approximation of polynomials the proper experimental design must be used to collect data. Once the data are collected, the Method of Least Square is used to estimate the parameters in the polynomials. The response surface analysis is performed by using the fitted surface. The response surface designs are types of designs for
fitting response surface [Bradley, 2007]. Therefore, the objective of studying RSM can be accomplish by
(1) Understanding the topography of the response surface (local maximum, local minimum, ridge lines), and
(2) Finding the region where the optimal response occurs. The goal is to move rapidly and efficiently along a path to get to a maximum or a minimum response so that the response is optimized.
1.4.1 Central composite design
The central composite design was presented by Box and Wilson [Box et al., 1951]. This design consists of the following parts: (1) a full factorial or fractional factorial design; (2) an additional design, often a star design in which experimental points are at a distance from its center; and (3) a central point. Figure 1-1 illustrates the full central composite design for optimization of two and three variables. Full uniformly routable central composite designs present the following characteristics:
(1) Require an experiment number according to N = k2 + 2k + cp, where k is the factor
number and cp is the replicate number of the central point;
(2) α-values depend on the number of variables and can be calculated by α= 2(k − p)/4. For two, three, and four variables, they are, respectively, 1.41, 1.68, and 2.00;
(3) All factors are studied in five levels ( −α, − 1, 0, +1, +α).
Figure 1-1 shows representations of central composite designs for two- and
three-variable optimization, respectively. Table 1-3 presents the coded values of the
experimental matrices for the application of these designs. Many applications of the central composite design in the optimization of analytical procedures can be found in the literature [Bezerra et al., 2008].
1.5 Bio-mitigation of CO
2, NO
xand SO
xin flue gas with microalgae
Microalgae can typically be used to capture CO2 from three different sources:
soluble carbonate [Wang et al., 2008]. Capture of atmospheric CO2 is probably the most basic
method to sink carbon, and relies on the mass transfer from the air to the microalgae in their aquatic growth environments during photosynthesis [Wang et al., 2008]. However, the potential yield from the atmosphere is limited by low CO2 concentration in air (360 ppm)
which makes it economically infeasible [stepan et al., 2002]. In contrast, Flue gases from power plants are responsible for more than 7 % of the total world CO2 emissions from energy
use [Kadam, 1997]. Also, industrial exhaust gases contains up to 15 % CO2 [Kadam, 2001;
Maeda, 1995], even the higher CO2 concentration of up to 20 % [Bilanovic et al., 2009],
providing a CO2-rich source for microalgae cultivation and a potentially more efficient route
for CO2 bio-fixation, and adaptability of this process for both photobioreactor and raceway
pond systems for microalgae production. Therefore, to use a flue gas emission from an industrial process unit as a source of CO2 for the microalgae growth is introduced to have a
great potential to diminish CO2 and to provide a very promising alternative to current GHG
emissions mitigation strategies [Mata et al., 2010]. However, only a small number of algae are tolerant to the high levels of SOx and NOx that are present in flue gases. The gases also need
to be cooled prior to injection into the growth medium [Brennan et al., 2010]. A number of microalgae species are able to assimilate CO2 from soluble carbonates such as Na2CO3 and
NaHCO3 [Wang et al., 2008]. Due to the high salt content and resulting high pH of the
medium, it is easier to control invasive species since only a very small number of algae can growth in the extreme conditions [Wang et al., 2008].
The selection of suitable microalgae strains for CO2 bio-mitigation has significant effect
on efficacy and cost competitiveness of the bio-mitigation process. The desirable attributes for high CO2 fixation include as following: high growth and CO2 utilization rates; high
tolerance of trace constituents of flue gases such as SOx and NOx; possibility for valuable
by-products and co-products, like biodiesel and biomass for solid fuels; ease of harvesting associated with spontaneous settling or bio-flocculation characteristics; high water
temperature tolerance to minimize cost of cooling exhaust flue gases [Brennan et al., 2010]. In these requisitions, the tolerant to NOx and SOx is the most important character the
application with flue gas, and the mechanisms of metabolizing CO2, NOx and SOx with
microalgae are mentioned as following.
1.5.1 The reduction of CO2 in flue gas with microalgae
between 36 and 65 %. CO2 can freely diffuse through the plasma membrane (Figure 1-2),
suggesting that a CO2 gradient is not built up along the plasmalemma. Rubisco (ribulose
1,5-bisphosphate carboxylase/oxygenase) is the first enzyme in the Calvin cycle that
assimilates CO2 by converting it into 3-phosphoglycerate (3-PGA). Besides this carboxylase
activity, rubisco displays oxygenase activity, using oxygen to form 3-PGA and
phosphoglycolate. The latter is subsequently oxidatively metabolised via photorespiration, leading to a loss in CO2 fixation [Van Den Hende 0et al., 2012]. Since microalgae have a low
CO2-binding capacity of rubisco and the oxygenase activity of rubisco depends on the CO2:O2
ratio [Graham et al., 2000], an elevated intracellular CO2 concentration is a prerequisite.
Therefore, some microalgae possess carbon concentrating mechanisms (CCMs) that elevate the CO2 level at the site of rubisco up to 1000-fold over that of the surrounding medium
[Price et al., 2008]. Microalgae CCMs involve: (1) cell membrane inorganic carbon transporters/pumps (e.g., HCO3−/Na + symports or ATP-driven uniports), (2) carbonic
anhydrases (CA), (3) specialized cellular structures, and (4) calcification [Van Den Hende S et al., 2012].
A number of research findings have investigated the potential of microalgae for biological carbon capture under various conditions. C. vulgaris grown on wastewater discharge from a steel plant successfully sequestered 0.624 g/L/day CO2 [Yun et al., 1997].
Doucha recorded 10~50 % reduction in CO2 concentration in flue gases using with Chlorella
sp., with the efficacy decreasing with increasing rate of flue gas injection into microalgae culture [Doucha et al., 2005]. Chlorella Strains from hot springs, which also showed to be tolerant to high temperatures up to 42 ℃, for CO2 fixation from industrial flue gases
containing up to 40 % CO2 [Sakai et al., 1995]. Chang and Yang found that certain species of
Chlorella could grow in an atmosphere containing CO2 up to 40 % [Chang et al., 2003]. In
addition to the Chlorella species used in the CO mitigation, de Morais and Costa using
Spirulina sp. obtained a maximum daily CO2 biofixation of 53.29 % for 6 % CO2 and 45.61
% for 12 % CO2 in the injected flue gas, with the highest mean fixation rate being 37.9 % for
6 % CO2. With S. obliquus, de Morais and Costa achieved biofixation rates of 28.08 % and
13.56 % for 6 % and 12 % CO2, respectively [de Morais et al., 2007a]. De Morais and Costa
found the microalgae species S. obliquus and C. kessleri to be capable of growing in media containing up to 18 % CO2 [de Morais et al., 2007b]. When comparing B. braunii, C. vulgaris
and Scenedesmus sp. under flue gas conditions, Yoo found Scenedesmus sp. to be the most suitable for CO2 mitigation due to high rates of biomass production (0.218 g/L/day) [Yoo et
al., 2010]. B. braunii and Scenedesmus sp. were found to grow better using flue gas as
compared to air enhanced with CO2. This is in line with an earlier study by Brown who found
that microalgae can tolerate flue gas very well [Brown, 1996]. The high cost of process technology and lack of price competitiveness of biodiesel extraction from microalgae versus petroleum diesel are key obstacles to commercial exploitation [Chisti, 2008]. Bio-mitigation of CO2 emissions provides a complementary function that may be exploited to reduce cost and
to enable sustained utilization of microalgae as a biofuel resource.
1.5.2 The reduction of NOx in flue gas with microalgae
After carbon, nitrogen is the most important nutrient for algal production. Whereas the Redfield C:N ratio of microalgae biomass is 6.66 (molar), this ratio varies in practice among species, with reported values ranging between 3 and 17 [Geider et al., 2002]. In general,
microalgae can take up nitrogen in several forms: NH4+, NO3−, NO2−, NO and N2 (Figure 1-3).
Both nitrate and nitrite reduction is tightly coupled to energy supply from the photosynthetic electron transfer or external organic carbon [Graham et al., 2000]. Brown found that some of the NO dissolved and was available as an N-source for the microalgae [Brown, 1996]. Nagase concluded that little NO was oxidized in the medium before its uptake by algal cells [Nagase et al., 2001]. Since NO is a small nonpolar molecule, they suggested that NO can diffuse through cell membranes. Moreover, they concluded that NO was preferentially used as a source for microalgae growth rather than nitrate. These researchers based these conclusions on the nitrogen balance calculations. The exact mechanisms through which microalgae use NOx are still to be proven with more accurate techniques such as nitrogen isotopes [Van Den
Hende S et al., 2012].
1.5.3 The reduction of SOx in flue gas with microalgae
Sulphur, an essential component of the amino acids cysteine and methionine, and S-containing thylakoid lipids, is indispensable for microalgae growth [Graham et al., 2000]. As far as it is known at this stage, microalgae acquire sulphur by taking up sulphate into the cytoplasm, for example, by means of high-affinity sulphate transporter systems [Giordano et al., 2005a, b] (Figure 1-4). Sulphate is transported into the plastids or, if present in excess, stored in vacuoles. This stabile sulphate ion is reduced via activation by ATP to
5-adenylsulphate (APS), catalyzed by ATP sulphurylase (ATP-S). APS is then reduced by APS reductase to sulphite (SO32−). The produced sulphite is further reduced by sulphite
reductase to sulphide (S2−), which is immediately incorporated into cysteine. On the other hand, S-compounds can also be released by microalgae [Giordano et al., 2005a, b]. For example, Emiliana sp. and Phaecystis sp. generate dimethyl sulfio proprionate (DMSP) for use in osmoregulation or as a cryoprotectant in cold regions [Graham et al., 2000]. This can then be converted into volatile dimethyl sulphide (DMS), which is released from the cells and oxidised to dimethyl sulphoxide (DMSO) to form sulphate aerosols. These aerosols promote an increase in cloud cover and an increase in the earth's albedo (fraction of incident radiation reflected back into space). DMS can be 29 % of the total sulphate assimilation of microalgae and can also generate SO2 via reactions with NO3 and OH [Giordano et al., 2005a, b].
1.6 Biodiesel
Biodiesel is a derivative of oil crops and biomass which can be used directly in
conventional diesel engines [Clark, et al., 2008]. It is a mixture of monoalkyl esters of long chain fatty acids (FAME) derived from a renewable lipid feedstock which are composed by 90~98 % of triglycerides and small amounts of mono and diglycerides, free fatty acids (1~5 %), and residual amounts of phospholipids, phosphatides, carotenes, tocopherols, sulphur compounds, and traces of water, such as algae oil [Demirbas, 2009; Bozbas et al., 2008]. After the extraction processes, the resulting product algae oil can be converted into biodiesel through a process called transesterification. Transesterification is a multiple step chemical reaction between triglycerides and alcohol in the presence of a catalyst, including three reversible steps in series, where triglycerides are converted to diglycerides, then diglycerides are converted to monoglycerides, and monoglycerides are then converted to esters (biodiesel) and glycerol (by-product) [Sharma, et al., 2009]. The overall
transesterification reaction is described in Figure 1-5 where the radicals R1, R2, R3 represent long chain hydrocarbons, known as fatty acids [Mata, 2010]. The composition of fatty acids significantly depends on the different microalgae species, as they can have a significant effect on the characteristics of biodiesel produced. These are composed of saturated and unsaturated fatty acids with 12–22 carbon atoms, some of them of v3 and v6 families. Thomas analyzed the fatty acid compositions of seven fresh water microalgae species showing that all of them synthesized C14:0, C16:0, C18:1, C18:2, and C18:3 fatty acids. This author reported that the relative intensity of other individual fatty acids chains is species specific, e.g. C16:4 and C18:4 in Ankistrodesmus sp., C18:4 and C22:6 in Isochrysis sp., C16:2, C16:3 and C20:5 in
Nannochloris sp., C16:2,C16:3, and C20:5 in Nitzschia sp. [Thomas et al., 1984].
Although the microalgae oil yield is strain-dependent it is generally much greater than other vegetable oil crops, as shown in Table 1-4 that compares the biodiesel production efficiencies and land use of microalgae and other vegetable oil crops, including the amount of oil content in a dry weight basis and the oil yield per hectare, per year [Hili, 1984; Peterson, 1998; Rathbauer et al., 2002; Zappi, 2003; Callaway, 2004; Vollmann, 2007; Reijnders, 2008; Kheira, 2009].
The conversion of triglycerides or oil to biodiesel can achieve 98 % [Noureddini, 1998] or greater than 98 % [Anderson, 2003] as an alternative fuel for Diesel engines. The results of the biodiesel product should be quite similar to those of conventional. Diesel in its main characteristics or compatible with conventional petroleum diesel, and it can also be blended in any portion with petroleum diesel. The standard specification for biodiesel fuel for B100 is ASTM D6751 [Sarmidi A, 2009]. The level of blending is designated Bxx, where xx indicates the amount of biodiesel in the blend. The physical and fuel properties of biodiesel from
microalgae oil, in general, were comparable to those of diesel fuel. The biodiesel from microalgae oil showed a much lower cold filter plugging point of 11℃in comparison with that of diesel fuel, as shown in Table 1-5 [Xu et al., 2006]. Basically, Algal biodiesel has several advantages over petroleum diesel in that: it is derived from biomass and therefore is renewable, biodegradable, and quasi-carbon neutral under sustainable production; it is non-toxic and contains reduced levels of particulates, carbon monoxide, soot, hydrocarbons and SOx [Brennan, 2010]. Another major advantage of algal biodiesel is in reduced CO2
II. Materials and Methods
2.1 Microalgae strain
The microalgae Chlorella sp. TT-1 was originally obtained from the collection of Taiwan Fisheries Research Institute (Tung-Kang, Ping-Tung, Taiwan) and isolated in our laboratory by chemical mutagenesis.
The species of Chlorella sp. isolated in Taiwan was unidentified. However, the partial sequence of 18S rRNA (599 bp) of the Chlorella sp. has been amplified and sequenced for species identification in this study. This result of sequence alignment was performed by NCBI nucleotide blast [Wu et al., 2001].
2.2 Culture medium and chemicals
Chlorella sp. TT-1 was cultured in artificial sea water enriched with f/2 medium and an
illumination of 300 mol/m2/s by white fluorescent light at 26 ± 1℃. Artificial sea water has following composition (per liter): including 29.23 g NaCl (Showa, Tokyo, Japan), 1.105 g KCl (Showa), 11.0862 g MgSO4 . 7H2O (Amresco, Solon, OH, USA), 1.21 g Tris-base (Merck,
Darmstadt, Germany), 1.83 g CaCl2 . 2H2O (Amresco), 0.25 g NaHCO3 (Amresco). f/2
medium has following composition (per liter): 75 mg NaNO3 (Showa), 5 mg NaH2PO4 . H2O
(Sigma, Saint Louis, MO, USA), 1 mL of trace metal solution, and 1 mL of vitamin solution [Guillard, 1975]. Trace elemental solution (per liter) includes 4.36 g Na2 . EDTA (Amresco),
3.16 g FeCl3 . 6H2O (Sigma), 180 mg MnCl2 . 4H2O (Sigma), 10 mg CoCl2 . 6H2O (Sigma), 10
mg CuSO4 . 5H2O (Sigma), 23 mg ZnSO4 . 7H2O (Showa), 6 mg Na2MoO4 (Sigma). Vitamin
solution (per liter) includes 100 mg vitamin B1 (Sigma), 0.5 mg vitamin B12 (Sigma) and 0.5
mg biotin (Sigma).
The microalgae were selected for the studies of CO2 challenge and the high biomass
concentration which were cultured in modified f/2 medium in artificial sea water at 26 ± 1 ℃ with an illumination of 300 mol/m2/s by white fluorescent light. Modified f/2 medium has following composition (per liter): including 225 mg NaNO3 (Showa), 15 mg NaH2PO4 . H2O