行政院國家科學委員會專題研究計畫 成果報告
綠竹超氧岐化脢 之抗氧化機制研究
計畫類別: 整合型計畫 計畫編號: NSC93-2321-B-002-029- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學植物科學研究所 計畫主持人: 靳宗洛 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 12 月 25 日
Abstract
The total superoxide dismutase (SOD) isozymes activity in green bamboo were analyzed, at least 7 isoforms of CuZnSOD and 1 MnSOD were identified. Southern blotting analysis suggested that there are at least 4 CuZnSOD and MnSOD genes. When supplying 0.05 to 0.2 mM copper or manganese ion into the bamboo leaf crude protein extract resulted in no activities band-shifting of isoforms. When denaturing and renaturing of the protein extract with 8 M urea followed by dialysis, shown the same result as copper ions treatment. The abundance of green bamboo SOD genes and the complexity of CuZnSOD isoforms suggested that green bamboo has a more complex regulation in its antioxidant system. Which may tribute to the physiologically special properties of green bamboo under oxidative stress?
By alignment of the published MnSOD cDNA sequences in monocots, the degenerate primers were designed according to the conserved regions. Two full-length cDNA sequences coding for green bamboo BoMnSOD were cloned by the RACE and RT-PCR. The deduced amino acid of BoMnSOD consists of 231 amino acids, including the essential domains of MnSOD and a mitochondrial transit peptide (27 a.a.) in the N-terminus. The BoMnSOD share 75-89% identity with MnSODs from other plants. The cDNA coding regions were cloned into the pGEX-6P-1 expression vector, and expressed in the Escherichia coli BL21 (DE3) strain. Four full-length cDNA sequences coding for green bamboo BoCuZnSOD also cloned. One full-length BoCuZnSOD cDNA clone of 793 bp consists of 152 amino acids, without transit peptide was analyzed. This cytosolic
BoCuZnSOD shares 81-95% identity with other plant’s.
Recombinant GST-BoCuZnSOD and GST-BoMnSOD remained of the SOD activity, both of which were stable at alkaline pH and declined to 10% after incubation at 60°C for 20 min. The SOD activity of recombinant BoCuZnSOD and BoMnSOD were stable more than 3 days incubated at room temperature. BoMnSOD and BoCuZnSOD cDNA sequences from green bamboo not only overexpressed in prokaryotes but also remained stable under a broad range of pH, higher temperature, also very stable in the room temperature. These properties are beneficial for applications in commercial, such as in cosmetics for skin protection or defending un-esthetic effects caused by oxygen-containing free radicals.
The promoter of BoCuZnSOD and BoMnSOD were also studied, analysis of the cis-element within the promoter is proceeding to help us understand the physiological regulation of BoCuZnSOD and BoMnSOD.
Introduction
Reactive Oxygen Species (ROS)
The evolution of aerobic metabolic processes such as respiration and photosynthesis unavoidably led to the production of reactive oxygen species (ROS) in mitochondria, chloroplasts, and peroxisomes. ROS are molecules like hydrogen peroxide, ions like hypochlorite ion, radicals like hydroxyl radical, and superoxide anion which is both ion and radical. Hydroxyl radical is the most reactive ROS. A common feature among the different ROS types is their capacity to cause oxidative damage to proteins, DNA, and lipids. Increasing evidence indicates that ROS also function as signaling molecules in plants involved in regulating development and pathogen defense response (Apel and Hirt, 2004).
Generation of ROS
In plants ROS are continuously produced as byproducts of various metabolic pathways localized in different cellular compartments (Foyer and Harbinson, 1994). The production and scavenging of ROS is balanced in plants at normal condition. Some environmental factors (high light, drought, low or high temperature), mechanical stress and the pathogen attack may perturb the equilibrium and cause the rapid increase in ROS. In plant, NADPH-oxidase associated with plasma membrane which was required for ROS production during defense response (Torres et al., 2002). Cell wall-bound peroxidases may produce ROS for physiological responses such as the controlled breakdown of structural polymers, like cell wall in root, hypocotyls, or coleoptiles (Vianello et al., 1991; Ogawa et al., 1997; Frahry et al., 2001). Predominantly, ROS are continuously produced in chloroplasts, mitochondria, and peroxisomes through many metabolism pathways: photosynthesis, photorespiration, oxidative phosphorylation and fatty acid -oxidation.
ROS Detoxification
Plant cell had to evolve sophisticated strategies to keep the concentrations of superoxide, hydrogen peroxide, and transition metals such as Fe and Cu under control were described following:
Nonenzymatic ROS Scavenger: Nonenzymatic antioxidants include the major cellular redox buffers ascorbate and glutathione (GSH), as well as tocopherol, flavonoids, alkaloids, and carotenoids. GSH is oxidized by ROS forming oxidized glutathione (GSSG), ascorbate is oxidized to monodehydroascorbate (MDA) and dehydroascorbate (DHA). Through the ascorbate-glutathione
cycle, GSSG, MDA, and DHA can be reduced reforming GSH and ascorbate. Reduced states of the anti- oxidants are maintained by glutathione reductase (GR), monodehydroascorbate reductase (MDAR), and dehydroascorbate reductase (DHAR), using NADPH as reducing power (Asada and Takahashi, 1987; Tsugane et al., 1999). Little is known about flavonoids and carotenoids in ROS detoxification in plants. However, overexpression of carotene hydroxylase in Arabidopsis leads to increased amounts of xanthophyll in chloroplasts and results in enhanced tolerance towards oxidative stress induced in high light (Davison et al., 2002).
Enzymatic ROS Scavenger
Enzymatic ROS scavenging mechanisms in plants include superoxide dismutase (SOD), ascorbate peroxidase (APX), glutathione peroxidase (GPX), and catalase (CAT). Superoxide dismutases (SODs; EC 1.15.1.1) are ubiquitous metalloenzymes that catalyze the disproportionation of superoxide (O2.-) radicals into hydrogen peroxide and molecular oxygen. APX, GPX and CAT
subsequently detoxify H2O2. They play an important role in protecting cells against the toxic effects
of superoxide radicals produced in different cellular loci (Fridovich, 1986; Halliwell and Gutteridge, 1989). In chloroplasts, the superoxide radical is produced by the univalent reduction of dioxygen during photosynthetic electron transport, particularly under conditions where CO2 is limited and
light intensity is high (Asada and Takahashi, 1987). Because superoxide has been postulated as a reactant in the formation of the highly reactive hydroxyl radical (Asada and Takahashi, 1987), SODs may play a central role in protecting chloroplast proteins and membranes against damage from ROS.
Plant Superoxide Dismutase (SOD)
Three types of SOD have been characterized based on the nature of the metal co-factor present at the catalytic site, i.e. copper/zinc (CuZnSOD), iron (FeSOD), or manganese (MnSOD) SODs. The CuZnSODs are localized in the cytosol, chloroplasts, nucleus, and apoplast; the Mn-SODs in the mitochondria and peroxisomes; and the FeSODs in the chloroplasts (Sandalio et al., 1987; Van Camp et al., 1990; Bowler et al., 1994; Ogawa et al., 1996). Unlike most other organisms, plants contain multiple SOD isozymes. The SOD activity in plants increases differentially in response to various environmental stresses (Bowler et al., 1989; Tsang et al., 1991) and often correlates with the severity of stress (Bowler et al., 1992).
The first plant SOD gene was cloned from maize (Cannon, White and Scandalios, 1987). The sequences of many SODs have been cloned in intact plants, e.g. tobacco (Bowler, Aliotte, De Loose,
Van Montagu and Inze, 1989), tomato (Perl-Treves, Nacmias, Aviv, Zeelon and Galun, 1988), maize (Cannon and Scandalios, 1989), rice (Kaminaka, Morita, Yokoi, Masumura and Tanaka, 1997; Sakamoto, Ohsuga and Tanaka, 1992), and aspen (Akkapeddi, Shin, Stanek, Karnosky and Podila, 1994). Moreover, nine SOD isoenzymes have been described in maize (Baum and Scandalios, 1981), and seven in Arabidopsis (Kliebenstein, Dietrich, Martin, Last and Dangl, 1998). The subcellular localization of seven SOD genes in Arabidopsis was: cytosol (CSD1), chloroplasts (CSD2, FSD1, FSD2, FSD3), mitochondria (MSD1), and peroxisome (CSD3).
The growth rate of green bamboo shoot was extremely fast, which may lead to the large amount of ROS accumulation. We analyzed the SODs from green bamboo which was the first line of enzymatic defense against ROS and revealed the complexity of its SOD activity. By cloning of green bamboo CuZnSOD and MnSOD and overexpress the recombinant proteins in E. coli, we can characterize the property of the recombinant protein and to raise the antibody against green bamboo SODs for further physiological study. Many commercial skin-care products were contained SOD as antioxidant, so the understanding of green bamboo SODs system may have economic potential.
Materials and Methods
Plant Materials
Green Bamboo (Bambusa oldhamii) leaf and shoot were excised and frozen in liquid nitrogen. Samples were stored at -80°C until RNA, DNA, and protein extraction. (Samples were harvested from Muzha, suburb of Taipei)
Protein Extraction and Quantification
Green bamboo leaf and shoot samples were ground to powder in liquid nitrogen and suspended with 150 mM Tris-HCl buffer (pH 7.2) (tissue:medium ratio 1:3, w/w). The homogenate was centrifuged at 13,000g at 4°C for 10 min to collect the supernatant. Protein quantification was conducted by the method of Bradford (1976) with the Bio-Rad protein assay reagent (Bio-Rad, Hercules, CA). Bovine serum albumin (Sigma) was used as a reference standard.
Electrophoresis and SOD Activity Analysis
Protein was separated on a 10% native PAGE in the Tris-glycine buffer (pH 8.3). A photochemical method modified from (Beauchamp and Fridovich, 1971) was used to visualize SOD activity. The gel was soaked in 0.1% (w/v) nitroblue tetrazolium (NBT) solution for 15 min, rinsed with distilled water and transferred to 100 mM potassium phosphate buffer (pH 7.0) containing 0.028 mM riboflavin and 28 mM TEMED (N,N,N’,N’ -tetramethyl-ethylenediamine) for another 15 min. After being washed with distilled water, gel was illuminated on a light box, with a light intensity of 30 μmol m-2 s-1 for 15 min to initiate the photochemical reaction. The resulting intensity
of SOD activity and the banding area after the SOD activity stain was measured by a computing densitometer LAS-3000 imaging system (FUJIFILM). The SOD activity was verified by KCN and H2O2 as described by Pan et al. (1999). KCN is an inhibitor of CuZnSOD, whereas H2O2 inhibits
both CuZnSOD and FeSOD. The MnSOD activity is not inhibited by either chemical (Table. 5).
Western Blot Analysis
Protein was separated on a 10% native- or SDS-PAGE then transferred to a PVDF membrane (Amersham Biosciences), blocked with 5% nonfat-milk in 1X PBST for 1 h, and then incubated with anti-GST antiserum (1:2,000 dilution) for another hour as described by (Lee et al., 2003). The washed membrane was incubated with 1:2,000 diluted goat anti-rabbit IgG conjugated with alkaline
phosphatase (PerkinElmer, Boston, MA); the signals were detected by colorimetric reaction with BCIP/NBT (PerkinElmer).
DNA Isolation and Southern Blot Analysis
Green bamboo leaf and shoot samples were ground to powder in liquid nitrogen. Genomic DNA was extracted using DNAzol reagent (Invitrogen) according to the manufacturer’s instructions. 5 μg of the genomic DNA was digested with restriction enzymes (ex., EcoRV, BamHI, HindIII, and
XbaI) and electrophoresed on a 0.8% TBE agarose gel and blotted to a nylon membrane
(Immobilon-Ny+, Millipore) then probed with the 32P-radiolabeled BoMnSOD or BoCuZnSOD cDNA fragment. The membrane was washed in a high stringency condition and analyzed by scanning the phosphorscreen on Typhoon 9400 (Amersham Biosciences).
RNA Isolation and cDNA Synthesis
Green bamboo leaf and shoot samples were ground to powder in liquid nitrogen. Total RNA was isolated using REzol (PROtech Technologies) reagent as described by the manufacturer. 1 μg of total RNA was used for cDNA synthesis using BD SMART PCR cDNA synthesis kit (Clontech, Palo Alto, CA) according to the manufacturer’s instructions.
Analysis of MnSOD and CuZnSOD Gene Sequence
Sequence information of genes, proteins, and cDNAs were retrieved by searching public database with the BLAST algorithm (Altschul et al., 1997) at National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov), The Prosthetic groups and Metal Ions in Protein Active Sites Database Version 2.0 (PROMISE, http://metallo.scripps.edu/PROMISE/), Plant Cis-Acting Regulatory Element (Plant CARE, http://intra.psb.ugent.be:8080/PlantCARE/) and the Expert Protein Analysis System proteomics server of the Swiss Institute of Bioinformatics (ExPASy, http://au.expasy.org/). The mitochondrial-targeting signal was predicted with the TargetP version 1.1 (Emanuelsson et al., 2000; http://www.cbs.dtu.dk/servic es/ TargetP).
The 5’- and 3’-RACE to clone Green bamboo BoMnSOD and BoCuZnSOD genes
BoMnSOD gene-specific degenerate primers were synthesized according to the conserved
regions between cDNA sequences of MnSOD from Oryza sativa (NCBI accession no. L34038 and L34039), Triticum aestivum (NCBI accession no. AF092524, U73172, and U72212) and Zea mays
(NCBI accession no. L19461, L19462, L19463, M33119, and X12540). The gene-specific primers sequences of BoMnSOD-Fw-0, BoMnSOD-Rv-0, and BoMnSOD-Rv-1 were shown in the Table 1 and Figs. 1 and 2. The first time RT-PCR (by BoMnSOD-Fw-0, BoMnSOD-Rv-0 primers) following by a nested PCR (BoMnSOD-Fw-0, BoMnSOD-Rv-1 primers), one 0.4-kb cDNA fragment was amplified, then subcloned into yT&A vector (Yeastern Biotech, Taipei, Taiwan) for sequencing. Another gene-specific primer BoMnSOD-Fw-1 (see Table 1 and Fig. 2) was designed against to the former RT-PCR amplified sequence. Then, the 5’- and 3’-RACE were conducted using the 5’-RACE CDS primer and the 3’-RACE CDS primer-A provided in the commercial kit, combined with the primers: BoMnSOD-Rv-0, BoMnSOD-Rv-1 and BoMnSOD-Fw-0, BoMnSOD-Fw-1, respectively. The two RACE products were confirmed the specificity by a nested PCR, were also cloned into the yT&A vector for sequencing. The sequenced data used to designed primers for RT-PCR to clone the full length BoMnSOD cDNA genes (The primers used here as shown in the next section: Recombinant Proteins Expression). The full length BoCuZnSOD cDNA cloning was adapted according to BoMnSOD cDNA cloning described above. Sequencing results were analyzed with EditSeq and MegAlign programs of Lasergene software (DNASTAR, Inc.).
Recombinant Proteins Expression
The full-length BoMnSOD open reading frame was amplified using synthesized primers, the BoMSD-5’, BoMSD-3’-0, and for nested PCR BoMSD-3’-1 primers (Table 3). Two full-length
BoMnSOD open reading frames about 0.7-kb were cloned and named as BoMnSOD11 and BoMnSOD31. The primer pair set for amplification of the full-length BoCuZnSOD open reading
frame are BoCZSD-5’ and BoCZSD-3’ (Table 3) (Liao et al., 2003). Four full-length BoCuZnSOD open reading frames with about 0.5-kb were cloned and named as BoCuZnSOD3, BoCuZnSOD4,
BoCuZnSOD16, and BoCuZnSOD28. Theses BoCuZnSOD and BoMnSOD ORFs were constructed
in the pGEX-6P-1 vector (Amersham Biosciences) and sequenced. After transformation of the constructed expression vectors into E. coli BL21 (DE3) competent cells, the recombinant proteins were induced, expressed, and purified according to protocol described by the manufacture (Pharmacia Biotech).
Biochemical Characterization of the Recombinant Protein
Each purified recombinant protein was underwent the following treatments, all treatments were repeated three times. Samples after treatments were separated on a 10% native PAGE and following
by SOD activity analysis.
(1) pH stability: recombinant protein was amended with a half volume of the buffer at different pH: 0.2 M Na2HPO4 and 0.1 M citrate buffer (for different pH at 2.2, 3.0, 4.0, 5.0, and 7.0) and 0.2
M glycine-NaOH buffer (for pH at 9.0 and 10.4). Each sample was incubated at 37°C for 1 h.
(2) Thermal stability: recombinant protein was incubated at the different temperature (ex., 25, 40, 50, 60, 80, and 100°C) for 20 min.
(3) Room temperature stability: recombinant protein was incubated at different time interval for 2, 4, 8, 12, 24, 36, 48, 60, and 72 h in the room temperature.
Urea Denaturation and Metal Ion Application of Green Bamboo Leaf Crude Protein Extract Green bamboo leaf crude protein extract (60 μg) was treated with 8 M urea for denaturing and dialyzed for renaturation of the endogenous SODs. Different concentration of copper and manganese ion (0, 0.05, 0.1, 0.15, and 0.2 mM) were applied to green bamboo leaf crude extract and incubated at 37°C for 1h. Treated samples were separated on a 10% native PAGE and following by SOD activity analysis.
TAIL-PCR to Clone the Promoter Region of BoCuZnSOD and BoMnSOD
Specific TAIL-PCR primers were designed from the 1st intron sequence of BoCuZnSOD and
BoMnSOD (Tables 3 and 4). The genomic DNA was used for TAIL-PCR as described (Lieu, 1995;
Results
Sequence Analysis of Green Bamboo MnSOD and CuZnSOD
The nucleotide sequences of the two full-length MnSOD cDNA open reading frame (ORF) clones (696 bp): BoMnSOD11 and BoMnSOD31 are highly conserved (with 89-90% identity) compared with a wheat mitochondrial form MnSOD and the deduced amino acid sequences (231 a.a.) are also highly conserved (with 89% identity) compared with a rice mitochondrial form MnSOD. The MITOPROT and TargetP programs indicated that the BoMnSOD11 and BoMnSOD31 with a mitochondrial targeting signal and also with a Mn/FeSOD signature was identified by the PROSITE SCAN program. Suggesting that the BoMnSOD11 and BoMnSOD31 are putative mitochondrial localized MnSOD (Fig. 3, A and B). Comparison with the PROMISE database version 2.0 predicted that His-27, His-75, Asp-164 and His-168 are residues for manganese ion binding after the mitochondrial targeting sequence (Fig. 3B). The four full-length BoCuZnSOD cDNA ORF clones (459 bp) are BoCuZnSOD3, BoCuZnSOD4, BoCuZnSOD16, and BoCuZnSOD28 highly conserved (with 91% identity) compare with a rice cytosolic form CuZnSOD and the deduced amino acid sequences (152 a.a.) are also highly conserved (with 95% identity) compare with a maize cytosolic CuZnSOD. The BoCuZnSOD amino acid sequences were analyzed by PROSITE SCAN and without any transit peptide was proposed. Suggesting these four clones are to be a cytosolic CuZnSOD (Fig. 4, A and B). Comparison with the PROMISE database version 2.0 predicted that His-45, His-47 and His-119 are for copper ion binding; and His-70, His-79, and His-82 are for zinc ion binding, the His-62 is for both copper and zinc ion binding (Fig. 4B).
Expression and Purification of the Recombinant BoMnSOD and BoCuZnSOD
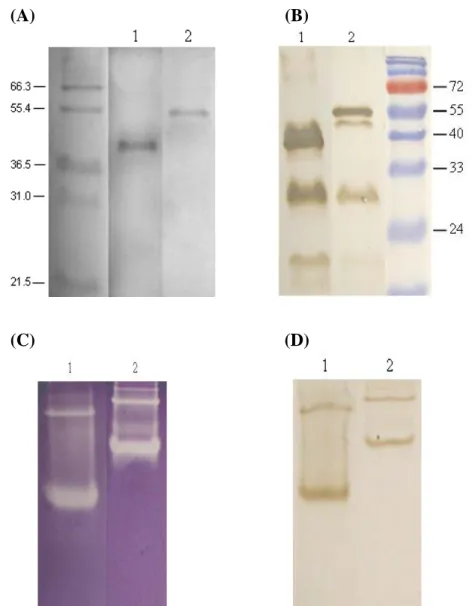
The recombinant BoMnSOD and BoCuZnSOD were overexpressed in E. coli BL21 (DE3) and analyzed on the 10% SDS-PAGE. The recombinant BoMnSOD and BoCuZnSOD were affinity purified to a homogeneity as judged by, SDS-PAGE analysis (Fig. 5A), SOD activity stain (Fig. 5C), and by Western blot analysis on SDS-PAGE (Fig. 5B) or native PAGE (Fig. 5D). The molecular weight of the purified GST-BoCuZnSOD (41 kD) and GST-BoMnSOD (51 kD) on SDS-PAGE agreed with the molecular weight deduced from its amino acid sequence.
Biological Characterization of the Recombinant BoCuZnSOD and BoMnSOD
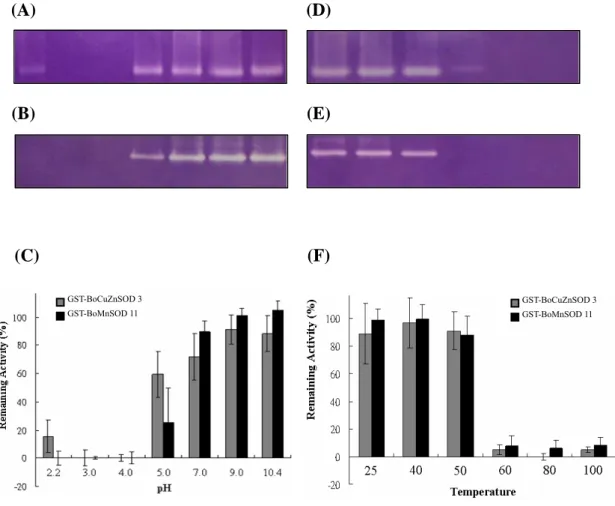
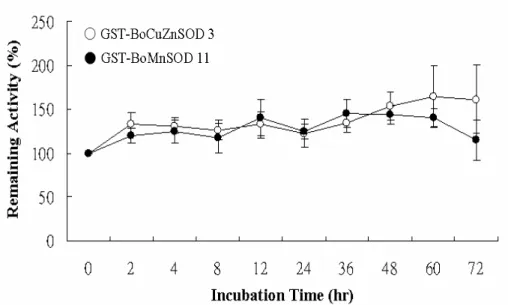
endogenous CuZnSOD isolated from the green bamboo leaves (Fig. 6A, lanes 2, 3; and Fig. 6B, lanes 3 and 5), meanwhile the recombinant BoMnSOD is also insensitive to any used inhibitors, same as the endogenous MnSOD (Fig. 6A, lanes 2, 3; and Fig. 6B, lanes 4, 6). Both the recombinant proteins are stabled at the alkaline pH value. Recombinant BoMnSOD showed better stability at pH 7-10.4, however, BoCuZnSOD is more stable than recombinant BoMnSOD during pH 5 and still remained the activity at pH 2.2 (Fig. 7C). Both recombinant proteins are retained the SOD activity during the 50°C treatment, and will lose the 90% activity when above 60°C (Fig. 7F). The stability of recombinant proteins is stable at room temperature even more than 3 days latter (Fig. 8).
Genomic Organization of BoCuZnSOD and BoMnSOD
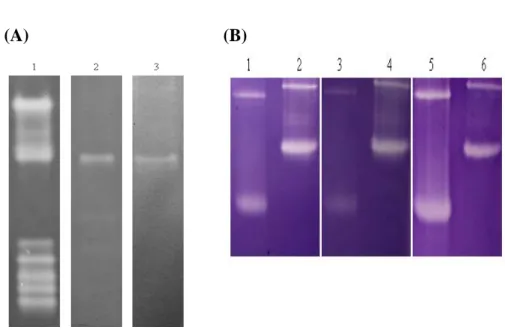
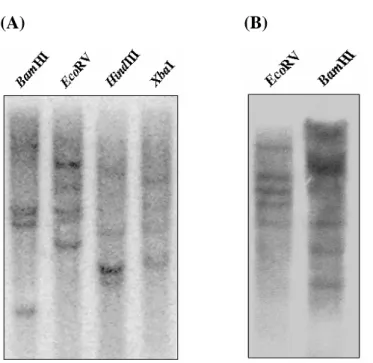
In order to determine the copy number of the BoCuZnSOD and BoMnSOD, a 5 μg genomic DNA of bamboo leaves was digested with BamHI, EcoRI, HindIII and XbaI. Southern blotting result was shown with 4 to 5 signals either probed with BoMnSOD or BoCuZnSOD cDNA fragment (Fig. 9, A and B), which strongly suggested that the green bamboo genome contains a multiple copy of
BoCuZnSOD and BoMnSOD genes.
Characterization of SODs Activities from the Green Bamboo
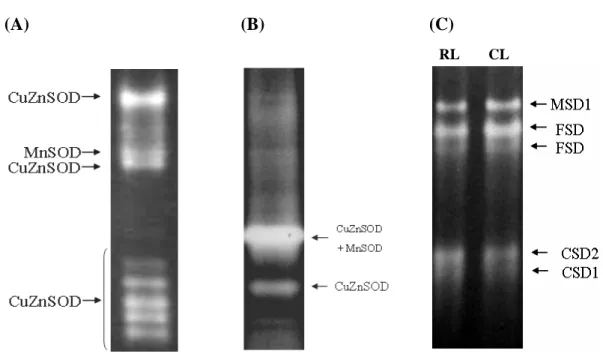
Green bamboo was shown with complexes of SOD activity profiles, especially for the CuZnSOD activities were compared with rice and Arabidopsis (Fig. 10). However, the SOD activity of green bamboo is only composed of the CuZnSOD and MnSOD, which was characterized and identified by inhibitors of KCN and H2O2 treatments (Fig. 6A). No band-shifting of SOD isoforms on the SOD
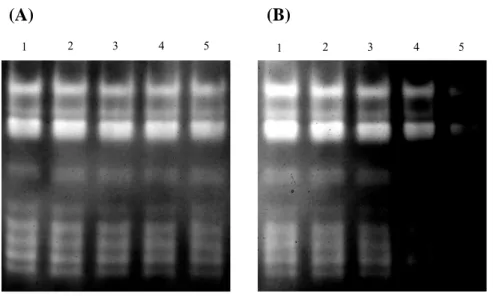
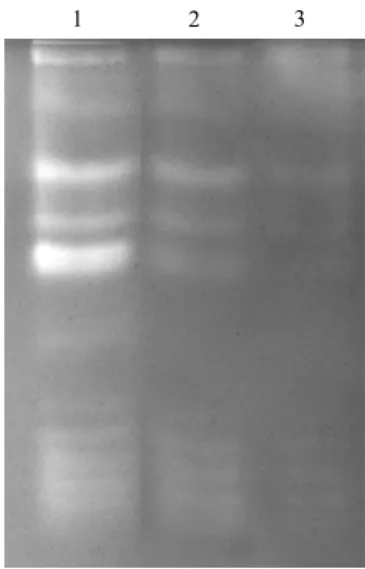
activity stain profile of green bamboo leaf crude protein extract after the application of different concentration of copper or manganese ion (Fig. 11 A, B). Using 8 M urea as denaturant to denature and renature of the endogenous SODs in green bamboo leaf protein extract. After the urea treatment, endogenous SODs still remained the SOD activity (Fig. 12, lane 2) and the activity stain profile is not interfered. Dialysis of treated sample resulted in largely losing SOD activity (Fig. 12, lane 3) but the activity stain profile was still the same.
Cloning the Promoter Region of BoCuZnSOD and BoMnSOD
A 543 bp of the BoCuZnSOD promoter region and 1678 bp of BoMnSOD promoter region were obtained through TAIL-PCR and PCR. Analysis the sequences of promoter region at PlantCARE, the predicted cis-element are summarized in Tables 8 and 9.
Discussion
Study of the SOD activity in green bamboo is composed of MnSOD and CuZnSOD that was confirmed and identified by treated with different inhibitors (Fig. 6A). The SOD activity stained patterns of green bamboo is more complex than other plants which we studied in this research, especially with very complexes of CuZnSOD activity profiles (Fig. 10). Compared with maize, a most studied monocot plant, which has 9 SOD genes with 5 CuZnSOD genes and 4 MnSOD genes. The SOD-1, -2, -4, -4A, and -5 are CuZnSOD, in which SOD-1 is a chloroplastic and the others belong to the cytosolic CuZnSOD, meanwhile, the SOD-3.1, -3.2, -3.3, and -3.4 are mitochondrial MnSOD. In rice which also with the CuZnSOD (SodC), MnSOD (SodA), and FeSOD activities.
The Southern blot analysis of CuZnSOD and MnSOD in green bamboo showed that there belong to a multiple copy genes family (> 4 copies) (Fig. 9). The activity profile of CuZnSOD in green bamboo with 7 different CuZnSOD activities but only with 1 MnSOD activity band were analyzed. The extracted total protein when treated with excess Cu and Mn ions, as well as by urea treatments for denatured and renatured caused without any SOD activities changed (Fig. 11 and 12), revealed these multiple CuZnSOD activity bands were not caused by incomplete incorporation of the catalytic metal ions, instead, it might be by itself specific genes/protein structures. The CuZnSOD genes with more diversity according to the gene alignment result, however, gene structures and/or protein structures of the MnSOD gene were more similarity than each other. However, why only one MnSOD activity was analyzed, it might because some of the MnSOD genes were not constitutively expressed.
We had cloned 2 mitochondrial BoMnSOD clones and 4 cytosolic BoCuZnSOD clones in green bamboo, for the reason of why with a less BoMnSOD get cloned, was because a highly GC content at the 5’-UTR which leaded to an unsuccessful 5’-RACE in BoMnSOD cloning (Fig. 2). The 3’-RACE clones revealed with 4 different 3’-UTR sequences in agreed with the results as shown in the Southern blot analysis which with 4 different copies of BoMnSOD in green bamboo genome.
We characterized and analyzed of the purified recombinant BoCuZnSOD and BoMnSOD expressed in E. coli. The BoMnSOD11 and BoCuZnSOD3 cDNA clones were conducted. During an overexpression of the recombinant BoMnSOD experiment, the protein was led to the inclusion body and this recombinant BoMnSOD was elaborated purified from the inclusion body without the SOD activity. The same result was also reported on the overexpression and purification of a rice recombinant MnSOD in E. coli (Tzeng, 1997). Because urea was used to release the recombinant
protein from the inclusion body, and followed by dialysis which might cause protein inactivation. In this case, dialysis might result in releasing of the ion cofactors from the active protein and cause the loss of SOD activity. In other published documents also indicated that the formation of the inclusion body mostly depended on the property of induced protein. When we induced of the recombinant BoMnSOD during 30°C for overnight culture, the recombinant BoMnSOD from the soluble fraction was purified and with SOD activity. We suggested that induction of the recombinant protein in a lower incubation temperature would be a good way for induction of the MnSOD among other plant species.
Both recombinant proteins of the BoCuZnSOD and BoMnSOD were stable in the alkaline pH condition, recombinant BoCuZnSOD remaining with a 60% SOD activity at pH 5 which was more stable than the recombinant BoMnSOD (about 20%). Compared with the rice recombinant CuZnSOD and MnSOD in a various pH treatment (Liu, 1997; Tzeng, 1997), the recombinant BoMnSOD was more stable (remaining more 90% SOD activity) than the rice recombinant MnSOD (less than 80% SOD activity) when the pH>7 (Tzeng, 1997). The recombinant BoCuZnSOD was less stable than the rice recombinant CuZnSOD at any treated pH condition. Interestingly, both recombinant CuZnSOD of green bamboo and rice were stable at alkaline pH but still remained the SOD activity specially at pH 2.2 (Liu, 1997).
In the thermal sensitivity assay, the recombinant BoCuZnSOD and BoMnSOD activities were agreed with the results of the rice recombinant CuZnSOD and MnSOD, both were stable under 50°C for 20 min treatment (Liu, 1997; Tzeng, 1997). The recombinant BoCuZnSOD and BoMnSOD activities were stable more than 3 days at room temperature, which were more stable than the rice recombinant CuZnSOD (decreased to 80% after 50h incubation).
In comparison of the biochemical characterization in green bamboo recombinant CuZnSOD and MnSOD with other CuZnSOD and MnSOD in different plant species were summarized in the Table 6 and 7. In summary, BoMnSOD and BoCuZnSOD ORFs from green bamboo not only overexpressed in prokaryotes but also remained stable under a broad range of pH, higher temperature, also very stable at the room temperature. These properties are beneficial for applications in commercial, such as in cosmetics for skin protection or defending un-esthetic effects caused by oxygen-containing free radicals.
References
Akkapeddi, A.S., Shin, D.I., Stanek, M.T., Karnosky, D.F. and Podila, G.K. (1994) cDNA and derived amino acid sequence of the chloroplastic copper/zinc-superoxide dismutase from aspen (Populus tremuloides). Plant Physiol. 106, 1231-1232.
Alscher, R.G., Erturk, N. and Heath, L. (2002) Role of superoxide dismutase (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 372, 1331-1341.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 17, 3389-3402.
Apel, K. and Hirt, H. (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 55, 373-399.
Asada, K. and Takahashi, M. (1987) Production and scavenging of active oxygen in
photosynthesis. In Photoinhibition, pp. 227-287. Kyle, D.J., Osmond, C.B., and Amtzen, C.J., eds. Elsevier Science Publishers, Amsterdam.
Bannister, W.H., Bannister, J.V., Barra, D., Bond, J. and Bossa, F. (1991) Evolutionary aspects of superoxide dismutase: the copper/zinc enzyme. Free Radical Res. Commun. 12-13, 349-361. Baum, J.A. and Scandalios, J.G. (1981) Isolation and characterization of the cytosolic and
mitochondrial superoxide dismutases of maize. Arch. Biochem. Biophys. 206, 249-264. Beauchamp, C.O. and Fridovich, I. (1971) Superoxide dismutase: improved assays and an assay
applicable to acrylamide gels. Anal. Biochem. 44, 276-287.
Bowler, C., Alliotte, T., Van den Bulcke, M., Bauw, G., Vandekerckhove, J., Van Montagu, M. and Inzé, D. (1989) A plant manganese superoxide dismutase is efficiently imported and correctly processed by yeast mitochondria. Proc. Natl. Acad. Sci. U S A. 86, 3237-3241. Bowler, C., van Montagu, M. and Inzé, D. (1992) Superoxide dismutase and stress tolerance.
Annu. Rev. Plant Physiol. Plant Mol. Biol. 43, 83-116.
Bowler, C., Van Camp, W., Van Montagu, M. and Inzé, D. (1994) Superoxide dismutase in plants. Crit. Rev. Plant Sci. 13, 199-218.
Bradford, M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254.
Cannon, R.E., White, J.A. and Scandalios, J.G. (1987) Cloning of cDNA for maize superoxide dismutase 2 (SOD2). Proc. Natl. Acad. Sci. U S A. 84, 179-183.
Davison, P.A., Hunter, C.N. and Horton, P. (2002) Overexpression of β-carotene hydroxylase enhances stress tolerance in Arabidopsis. Nature 418, 203-206.
Frahry, G., Schopfer, P. (2001) NADH stimulated, cyanide-resistant superoxide production in maize coleoptiles analyzed with a tetrazolium-based assay. Planta 212:175–83
Fridovich, I. (1986) Biological effects of the superoxide radical. Arch. Biochem. Biophys. 247, 1-11.
Gutteridge, J.M. and Halliwell, B. (1989) Iron toxicity and oxygen radicals. Baillieres Clin Haematol. 2, 195-256.
Kaminaka H, Morita S, Yokoi H, Masumura T, Tanaka K. (1997) Molecular cloning and characterization of a cDNA for plastidic copper/zinc-superoxide dismutase in rice (Oryza sativa L.). Plant Cell Physiol. 38, 65-69.
Kliebenstein, D.J., Monde, R.A. and Last, R.L. (1998) Superoxide dismutase in Arabidopsis: an eclectic enzyme family with disparate regulation and protein localization. Plant Physiol. 118, 637-650.
Liu, Y.G., Mitsukawa, N., Oosumi, T. and Whittier, R.F. (1995) Efficient isolation and mapping of Arabidopsis thaliana T-DNA insert junctions by thermal asymmetric interlaced PCR. Plant J. 8, 457-463.
Liu, Y.G. and Whittier, R.F. (1995) Thermal asymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking. Genomics. 25, 674-681.
Madamanchi, N.R., Donahue, J.L., Cramer, C.L., Alscher, R.G. and Pedersen, K. (1994) Differential response of Cu,Zn superoxide dismutases in two pea cultivars during a short-term exposure to sulfur dioxide. Plant Mol. Biol. 26, 95-103.
Neupert, W. (1997) Protein import into mitochondria. Annu. Rev. Biochem. 66, 863-917.
Ogawa, K., Kanematsu, S. and Asada, K. (1997) Generation of superoxide anion and localization of CuZn-superoxide dismutase in the vascular tissue of spinach hypocotyls: their association with lignification. Plant Cell Physiol. 38:1118–26
Ogawa, K., Kanematsu, S. and Asada, K. (1996) Intra- and extra-cellular localization of cytosolic CuZn superoxide dismutase in spinach leaf and hypocotyls. Plant Cell Physiol. 37, 790-799 Pan, S.M., Chen, M.K., Chung, M.H., Lee, K.W. and Chen, I.C. (2001) Expression and
characterization of monocot rice cytosolic CuZnSOD protein in dicot Arabidopsis. Transgenic Res. 4, 343-351.
Perl-Treves, R., Nacmias, B., Aviv, D., Zeelon, E.P. and Galun, E. (1988) Isolation of two cDNA clones from tomato containing two differential superoxide dismutase sequences. Plant Mol. Biol. 17, 745-760.
Perl-Treves, R. and Galun, E. (1991) The tomato Cu-Zn superoxide dismutase genes are developmentally regulated and respond to light and stress. Plant Mol. Biol. 17, 745-760. Purvis, A.C. (1997) Role of the alternative oxidase in limiting superoxide production by plant
mitochondria. Physiol. Plant. 100, 165-170.
Sakamoto A, Ohsuga H, Tanaka K. (1992) Nucleotide sequences of two cDNA clones encoding different Cu/Zn-superoxide dismutases expressed in developing rice seed (Oryza sativa L.). Plant Mol. Biol. 19, 323-327.
Salin,M.L. and Bridges, S.M. (1980) Localization of superoxide dismutase in chloroplasts from Brassica campestris. Zeitschrift für Pflanzenphtsiologie. 99, 37-47
Sandalios, L.M. and del Rio, L.A. (1987) Localization of superoxide dismutase in glyoxysomes from Citrullus vulgaris: functional implications in cellular metabolism, J. Plant Physiol. 127, 395-409.
Torres, M.A., Dangl, J.L., Jones, J.D.G. (2002) Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 99:517–22
Tsang, E.W., Bowler, C., Herouart, D., Van Camp, W., Villarroel, R., Genetello, C., Van Montagu, M. and Inzé, D. (1991) Differential regulation of superoxide dismutases in plants exposed to environmental stress. Plant Cell. 8, 783-792.
Tsugane, K., Kobayashi, K., Niwa, Y., Ohba, Y., Wada, K. and Kobayashi, H. (1999) A recessive Arabidopsis mutant that grows photoautotrophically under salt stress shows enhanced active oxygen detoxification. Plant Cell. 11, 1195-1206.
Van Camp, W., Bowler, C., Villarroel, R., Tsang, E.W., Van Montagu, M. and Inzé D. (1990) Characterization of iron superoxide dismutase cDNAs from plants obtained by genetic complementation in Escherichia coli. Proc. Natl. Acad. Sci. USA. 87, 9903-9907.
Vianello, A., Macri, F. (1991) Generation of superoxide anion and hydrogen peroxide at the surface of plant cells. J. Bioenerg. Biomembr. 23:409–23
pea cytosolic Copper/zinc superoxide dismutase. Plant Physiol. 96, 1391-1392.
Wu, G., Wilen, R.W., Robertson, A.J. and Gusta, L.V. (1992) Differential response of maize catalases and superoxide dismutase and chloroplastic copper/zinc superoxide dismutase genes in wheat. Plant Physiol. 120, 513-520.
Yost, F.J. Jr. and Fridovich, I. (1973) An iron-containing superoxide dismutase from Escherichia coli. J. Biol. Chem. 248, 4905-4908.
Zhu, D. and Scandalios, J.G. (1993) Maize mitochondrial manganese superoxide dismutases are encoded by a differentially expressed multigene family. Proc. Natl. Aca. Sci. USA. 90,
9310-9314. 曾尹貞。1997。水稻含錳超氧歧化酶在細菌中之表達及性質研究。國立台灣大學植物學研究 所碩士論文。 劉宏基。1997。水稻銅鋅超氧歧化酶的表達、純化與性質研究。國立台灣大學植物學研究所 碩士論文。 陳慶隆。2001。阿拉伯芥表現細胞質型含銅鋅超氧歧化酶之研究。國立台灣大學植物學研究 所碩士論文。 李溫琪。2003。阿拉伯芥銅鋅超氧歧化酶與銅離子鑲嵌蛋白質交互作用之研究。國立台灣大 學植物學研究所碩士論文。 廖民惠。2003。綠竹筍 CuZnSOD 基因之選殖及生化特性之研究。國立台灣大學植物學研究所 碩士論文。
BoMnSOD-Fw-0
BoMnSOD-Rv-1
Fig. 1 Complementary DNA alignment of the mitochondrial superoxide dismutase from monocots.
Rice, 1 and 2 (L34038, L34039); wheat, 1 to 3 (AF092524, U73172, and U72212); corn, 1 to 5 (L19461, L19462, L19463, M33119, and X12540). The conserved regions were boxed and used to design the degenerate gene-specific primers, BoMnSOD-Fw-0, BoMnSOD-Rv-0, and BoMnSOD-Rv-1, were underlined for amplification of a partial BoMnSOD cDNA.
Fig. 2 The full-length BoMnSOD cDNAs composed by 5’- and 3’-RACE.
The full-length BoMnSOD cDNAs were composed according to the sequences of 5’-RACE and 3’-RACE. The primers used for RACE or PCR were indicated by long arrows. The BoMnSOD-Fw-0 and BoMnSOD-Fw-1 primers used for 3’-RACE. The BoMnSOD-Rv-0 and BoMnSOD-Rv-1 primers used for 5’-RACE. The primers, BoMSD-5’, MSD-3’-1, and BoMSD-3’-0 were used for amplification of BoMnSOD cDNAs.
BoMnSOD-Fw-1 BoMSD-5’ BoMSD-3’-1 BoMSD-3’-0 BoMnSOD-Fw-0 BoMnSOD-Rv-1 BoMnSOD-Rv-0
(A)
(B)
Fig. 3 The nucleotide and amino acid sequences alignment of BoMnSOD.
(A) Two full-length open reading frames and (B) deduced amino acid sequences of the two BoMnSOD. The mitochondrial targeting sequence was boxed by black line and Mn/FeSOD signature was boxed by the blue line. The residues for manganese ion binding were boxed by red line in His-27, His-75, Asp-164, and His-168 after the mitochondrial targeting sequence.
27 75
(A)
(B)
45 47119
Fig. 4 The nucleotide and amino acid sequences alignment of BoCuZnSOD.
(A) Four full-length open reading frames and (B) deduced amino acid sequences of the four BoCuZnSOD, the CuZnSOD signature 1 was boxed by black line, the CuZnSOD signature 2 was boxed by blue line. The residues for metal binding were boxed by green line as seen in His-45, His-47, and His-119 for copper ion binding; in His-70, His-79, and His-82 were for zinc ion binding; the His-62 was for both copper and zinc ion binding.
Fig. 5 Purification and Characterization of the recombinant BoCuZnSOD 3 and BoMnSOD 11.
(A) 1 μg recombinant protein were separated on a 10% SDS-PAGE and stained with Coomassie brilliant blue. (B) Western blot analysis. 2.5 μg recombinant protein was separated on 10% SDS-PAGE then probed with anti-GST antibodies. (C) 4 μg recombinant protein was separated on 10% native PAGE and followed by SOD activity stain. (D) Western blot analysis. 2.5 μg recombinant protein was separated on 10% native PAGE, probed with anti-GST antibodies. Lane 1, GST-BoCuZnSOD3; lane 2, GST-BoMnSOD11.
(A) (B)
Fig. 6 Inhibition of the SOD activity of green bamboo endogenous SODs and the GST recombinant BoCuZnSOD and BoMnSOD.
(A) 60 μg crude extract of bamboo leaves was separated on 10% native PAGE and followed by the SOD activity assay. Lane 1, without inhibitor as control; lane 2, with 8 mM KCN inhibitor; lane 3, with 8 mM H2O2 inhibitor. (B) 4 μg recombinant protein
was separated on 10% native PAGE and followed by SOD activity stain. Lanes 1 to 2, no inhibition as the control; lanes 3 to 4, with 8 mM KCN inhibitor; lanes 5 to 6, 8 mM H2O2 inhibitor. Lanes 1, 3, and 5 are GST-BoCuZnSOD3, lanes 2, 4, and 6: are
GST-BoMnSOD11.
Fig. 7 The effect of pH and temperature on the recombinant BoCuZnSOD and BoMnSOD.
4 μg of GST-CuZnSOD3 and GST-MnSOD11 were treated with various pH buffer (A and B) at 37°C for 1 h or were heated at various temperatures for 20 min (D and E), samples were separated on 10% native PAGE and followed by SOD activity stain. The GST-CuZnSOD3 and GST-MnSOD11 activity after treatments were quantitated by a computing densitometer to calculate the remaining SOD activity (C and F).
GST-BoCuZnSOD 3 GST-BoMnSOD 11 (A) (D) (B) (C) (E) (F) GST-BoCuZnSOD 3 GST-BoMnSOD 11
Fig. 8 The stability of recombinant BoCuZnSOD3 and BoMnSOD11 at room temperature in different time interval.
Fig. 9 Southern blot analysis of green bamboo genomic DNA extracted from the leaves.
Hybridization with a partial (A) BoMnSOD and (B) BoCuZnSOD cDNA fragments as probes.
Fig. 10 The SODs activity profiles of the green bamboo, rice, and Arabidopsis. The crude extract of (A) the green bamboo leaves, (B) the rice leaves, and (C) the
Arabidopsis rosette (RL) and cauline leaves (CL). The SODs activities were indicated
in the right.
(A) (B) (C)
Fig. 11 Different metal ions applied in green bamboo leaf crude protein extract. 60 μg of green bamboo leaf protein extract was used for each treatment: (A) Lane 1-5: 0, 0.05, 0.1, 0.15, 0.2 mM copper ion supplied. (B) Lane 1-5: 0, 0.05, 0.1, 0.15, 0.2 mM manganese ion supplied.
(A) (B)
Fig. 12 Urea treatment of green bamboo leaf crude protein extract.
40 μg green bamboo leaf crude extract was separated on 10% native PAGE and followed by SOD activity stain. Lane 1, without treatment as control; lane 2, treated with 8 M urea; lane 3, treated with 8 M urea and dialyzed against the urea.
Table 1 Synthesized gene-specific primers for 5’- and 3’- RACE
Primers Sequence
BoMnSOD-Fw-0 5’-ACC ACC ARA AGM ACC ACG CCA CCT-3’ BoMnSOD-Fw-1 5’-CAG AGC GCC ATC AAG TTC AAC GGC-3’ BoMnSOD-Rv-0 5’-TCC AGT TCA YCA CCT TCC AGA TGT-3’ BoMnSOD-Rv-1 5’-GCC TVA CRT TCT TGT ACT GCA GGT-3’ (Wobble code: R = A/G, Y = C/T, M = A/C, V = A/C/G)
Table 2 Primers for the amplification of full-length MnSOD and CuZnSOD cDNA open reading frames
Primers Sequence RE
site BoMSD-5’ 5’-TCT GGA TCC ATG GCG CTC CGC ACT TCG-3’ BamHI
BoMSD -3’-0 5’-GCA TAC GAA GAT CCA GAC AGT-3’
BoMSD -3’-1 5’-AGA GAA TTC TCA TGC AGC CAC ATT TTC GTA-3’ EcoRI
BoCZSD-5’ 5’-TCT GGA TCC ATG GTG AAG GCT GTT GCT GTG-3’ BamHI
BoCZSD -3’ 5’-AGA GAA TTC CTA ACC CTG GAG CCC AAT GAT CC-3’ EcoRI
Table 3 Primers used for TAIL-PCR Primers sequence MnSOD-TP-Rv-0: 5'-TACGGGAGGTCAGGGAGCGTGAAC-3' MnSOD-TP-Rv-1: 5'-CGAGCGCCAGGGCTAGGGTTT-3' MnSOD-TP-Rv-2: 5'-TAGGGTTTTCCTCGAGGCCGAAGT-3' CuZnSOD-TP-Rv-0: 5'-GAGACTCGCAGCATGCAGTGGAA-3' CuZnSOD-TP-Rv-1: 5'-GGGTGAAAAAGATGGTGCCCTTGA-3' CuZnSOD-TP-Rv-2: 5'-TAAGCACAGCAACAGCCTTCACCA-3'
Table 4 Primers used for amplification of promoter regions of BoCuZnSOD and BoMnSOD
Primers Sequence RE site
CZSD-PC-Fw: 5'-TCTCCCGGGTTTCGATTCGACTCCGTTTA-3' SmaI
CZSD-PC-Rv: 5'-TCTAAGCTTTGTCTATGTGATCTGCAATC-3' HindIII
MSD-PC-Fw: 5'-TCTCCCGGGCGAGGGGCTAT-3' SmaI
MSD-PC-Rv: 5'-TCTAAGCTTGGCTGCACGTGAGAACGT-3' HindIII
Table 5 Inhibition of SODs analysis
Inhibitors SODs
KCN H2O2
CuZnSOD sensitive sensitive
FeSOD insensitive sensitive
Table 6 Comparison of biochemical characterization of GST-BoCuZnSOD and other CuZnSODs from different species.
CuZnSODs from different species
Native form pI Heat (> 70°C) pH effect H2O2 inhibition
GST-BoCuZnSOD Rice GST-CuZnSOD Rice CuZnSOD (cytosolic) Rice CuZnSOD (chloroplastic) Sweet potato CuZnSOD
Watermelon CuZnSOD (peroxisomal) Cassava CuZnSOD (cytosolic) - Dimmer Dimmer Dimmer
Monomer and dimmer - - 6.22 (prediction) 5.2-5.6 5.0 5.3 - 4.0 5.22 (prediction)
Unstable (less than 10% at 60°C, 20 min) Unstable Stable Stable Dimer dissociated to monomer Remained 50% at 70°C, 15 min - Unstable at acidic pH Unstable at acidic pH - - Dimer dissociated to monomer at acidic pH - - Less sensitive Less sensitive Sensitive Sensitive Sensitive Sensitive (>2.5 mM) -
Table 7 Comparison of biochemical characterization of GST-BoMnSOD and other MnSODs from different species.
MnSODs from different species
Native form pI Heat (> 70°C) pH effect H2O2 inhibition
GST-BoMnSOD Rice GST-MnSOD Norway spruce MnSOD Pine MnSOD (mitochondrial) Carnation MnSOD Pea MnSOD Yeast MnSOD - Dimmer Tetramer Tetramer - - Tetramer 6.77 (prediction) 4.64 5.5 6.5 4.85 - -
Unstable (less than 10% at 60°C, 20 min)
Unstable (less than 30% at 60°C, 20 min) - - - Unstable - Stable at pH > 7 More stable at alkaline
pH
Specific activity was strongly pH dependent - - - Unstable at pH > 7.8 Insensitive Insensitive - - - - -
Table 8 Cis-elements of BoCuZnSOD promoter region (by PlantCARE prediction). The promoter region was 543 bp long upstream of the start codon.
Box Function TATA-box AT1-motif GATA-motif chs-CMA1a I-box AE-box G-Box WUN-motif EIRE P-box HSE CAAT-box Skn-1_motif RY-element Box_I Box_II GC-repeat TATCCAT_C-motif TC-rich_repeats GA-motif ATC-motif ATCT-motif TCA-element CE1 E2Fb
core promoter element around -30 of transcription start part of a light responsive module
part of a light responsive element part of a light responsive element part of a light responsive element part of a module for light response
cis-acting regulatory element involved in light responsiveness wound-responsive element
elicitor-responsive element gibberellin-responsive element
cis-acting element involved in heat stress responsiveness common cis-acting element in promoter and enhancer regions cis-acting regulatory element required for endosperm expression cis-acting regulatory element involved in seed-specific regulation
above elements were conserve in BoMnSOD promoter region part of a light responsive element
part of a conserved DNA module involved in light responsiveness part of a conserved DNA module involved in light responsiveness cis-acting element involved in salicylic acid responsiveness
2
-Table 9 Cis-elements of BoMnSOD promoter region (by PlantCARE prediction). The promoter region was 1678 bp long upstream of the start codon.
Box Function TATA-box AT1-motif GATA-motif chs-CMA1a I-box AE-box G-Box WUN-motif EIRE P-box HSE CAAT-box Skn-1_motif RY-element Box I Box II GC-repeat TATCCAT C-motif TC-rich repeats ACE CATT-motif CG-motif GAG-motif GT1-motif L-box LAMP-element MRE Sp1 TCCC-motif TGG-motif rbcS-CMA7a ABRE AuxRR-core
core promoter element around -30 of transcription start part of a light responsive module
part of a light responsive element part of a light responsive element part of a light responsive element part of a module for light response
cis-acting regulatory element involved in light responsiveness wound-responsive element
elicitor-responsive element gibberellin-responsive element
cis-acting element involved in heat stress responsiveness common cis-acting element in promoter and enhancer regions cis-acting regulatory element required for endosperm expression cis-acting regulatory element involved in seed-specific regulation
above elements were conserve in BoCuZnSOD promoter region cis-acting element involved in light responsiveness
part of a light responsive element
cis-acting regulatory element involved in light responsiveness part of a light responsive element
light responsive element
part of a light responsive element part of a light responsive element
MYB binding site involved in light responsiveness light responsive element
part of a light responsive element part of a light responsive element part of a light responsive element
cis-acting element involved in the abscisic acid responsiveness cis-acting regulatory element involved in auxin responsiveness
ERE CGTCA-motif TGACG-motif TATC-box as1 GCN4 motif GC-motif O2-site MBS Prolamin_box A-box Box-W1
3-AF1 binding site ATGCAAAT motif C-repeat DRE Plant AP-2-like E2Fa
ethylene-responsive element
cis-acting regulatory element involved in the MeJA-responsiveness cis-acting regulatory element involved in the MeJA-responsiveness cis-acting element involved in gibberellin-responsiveness
cis-acting regulatory element involved in the root-specific expression cis-regulatory element involved in endosperm expression
enhancer-like element involved in anoxic specific inducibility
cis-acting regulatory element involved in zein metabolism regulation MYB Binding Site
cis-acting regulatory element associated with GCN4 sequence conserved in alpha-amylase promoters fungal elicitor responsive element


![HPSH [ 氧化數平衡反應式係數 ]](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)