國立高雄大學生物科技研究所
碩士論文
阿黴素對心臟內皮素表現的影響
Ariamycin
Effects
Endothelin-1
Expression
of
Heart
研究生:馮婉萱
撰
指導教授:施能朗
謝誌 這篇論文能夠順利完成首先要由衷感謝我的指導教授施能朗博士。老師兩年 來費心地指導我,並且在實驗和生活上給我最大的支持與協助。老師親自教導我 實驗的技術,並且不厭其煩一再講解實驗的觀念和關鍵技巧讓我獲益良多,實在 是非常感謝老師。同時也要感謝論文口試委員黃永森博士和翁炳孫博士對於論文 內容的詳細審閱,並在論文口試期間對於論文內容所提供的許多指正與建議。 此外,非常感謝台大醫學院醫學系副教授陳錦澤醫師和慈濟大學醫學系助理 教授鄭敬楓醫師提供豐富的研究資源,使得本篇論文能如期完成,尤其感謝陳醫 師對我的關心與照顧。還有感謝鄭志鴻博士、林恆博士和我討論實驗上問題。以 及欣翰、燕玲、韋雄、幸雯、英琪佩池對我的關心和照顧。 也要感謝我的同學及好朋友,惠菁、佑任、淙偉、文傑,謝謝你們伴我一 路 走來,幫我度過許多困難,和你們一起度過的時光真的非常快樂。 最後要感謝父母和家人的支持讓我沒有後顧之憂可以專心於研究中,尤其父 母親是引導我對自然及科學產生興趣的啟蒙老師,謹將此篇論文獻給我親愛的父 母親。
阿黴素對心臟內皮素表現的影響
指導教授:施能朗博士 國立高雄大學生物技術研究所 學生:馮婉萱 國立高雄大學生物技術研究所 摘要 本篇論文研究阿黴素對心臟內皮素調控的影響。阿黴素(多比柔星),是含有醌結 構的四環黴素,為最典型的抗腫瘤藥物之一,在臨床上屬於常規的治療藥物,在人體中 的累積劑量約達 550mg/m2時會導致嚴重的心臟毒性而限制了阿黴素的使用。雖然阿黴 素引起的心肌症之機制屬於多因性,但是自由基的過度產生以及抗氧化物的不足所引起 的氧化壓力增加扮演著重要的角色。此外,最新的研究指出endothelin-1 (ET-1)有當作此 種心肌損傷指標的潛力。在之前的研究指出,接受DOX治療而出現心臟衰竭的肺癌病 人血清中,可以發現ET-1 的量逐漸增加;另外動物實驗的研究發現,以DOX處理的老 鼠,血漿中ET-1 的量會增加。但是也有相反的結果發表。因此我們有興趣的是在DOX 處理下,心肌與非心肌細胞中的ET-1 表現模式與分子機轉。我們實驗室之前的研究發 現ROS會引發心臟纖維母細胞及血管內皮細胞的ET-1 表現,因此我們也想了解ROS在 DOX處理的心肌細胞與非心肌細胞中扮演何種角色。在我們的發現中,心肌細胞ET-1 mRNA和ET-1 的分泌在DOX的刺激下表現量都會增加,且有時間延遲的現象。非心肌 細胞ET-1 基因在DOX的影響下初期表現量會減少,但是ET-1 的分泌有早期且短暫性的 明顯增加。我們在進一步的細胞內ROS測定研究中發現,DOX是經由ROS活化心肌細胞 ET-1 的轉錄。但是在非心肌細胞的結果中,DOX不會增加細胞中的ROS,DOX可能是 經由影響非心肌細胞內的ET-1 儲存及運輸系統,以增加ET-1 的分泌。 關鍵字:阿黴素(Adriamycin,Doxorubicin,DOX),內皮素-1(endothelin-1,ET-1), 心肌細胞(myocyte),非心肌細胞(non-myocyte,non-M)。Ariamycin
Effects
Endothelin-1
Expression
of
Heart
Advisor: Dr. Shih, Neng-Lang Institute of Biotechnology National University of Kaohsiung
Student: Fong, Wen-Hsuan Institute of Biotechnology National University of Kaohsiung
ABSTRACT
The current study investigated the effects of adriamycin on ET-1 regulation in the heart. Adriamycin(Doxorubicin), a quinone-containing anthracycline antibiotics, is one of the most typical antineoplastic drugs and is in routine clinical use as therapeutic agents; the development of severe cardiac toxicity in human limits administration exceeding an accumulated dose of approximately 550mg/m2 . Although the mechanism of adriamycin-induced cardiomyopathy is multifactorial, increased oxidative stress due to overproduction of free radicals and antioxidant-deficit play an important role. In addition, recent studies are reported that endothelin-1 (ET-1) has the potential to be the maker of DOX induced caidac damage. Yamashita et al. (1994) and Yamashita et al. (1995) reported that they found that plasma ET-1 level increased in patients with lung cancer who receipted DOX treatments and have the progression of heart failure. Sayed-Ahmed et al. (2001) also reported that plasma ET-1 level increased in rat treated with DOX but there were reversed results. So we are interesting in the ET-1 expression patterns and molecular mechanisms of myocytes and non-myocytes treated with DOX. In our previous studies, we found that ROS can induce ET-1 expression in fibroblasts and endothelial cells. Then we are also interesting in the role of ROS in myocytes and non-myocytes treated with DOX. In this present study, we found that DOX could induce ET-1 mRNA expression and ET-1 secretion of myocytes and there exited a time lag. In non-myocytes, DOX inhibited ET-1 mRNA expression before 6 hours but ET-1 secretion rapidly increased after stimulated by DOX. Following intracellular ROS assay revealed that DOX induced ROS formation then with the consequence of ET-1 promoter activation in myocytes. In non-myocytes, DOX treated non-myocytes had similar ROS level with control. It’s clear that DOX can stimulate ET-1 expression in myocytes by activating ET-1 promoter through ROS mediated signal pathway. On the other hand, DOX may stimulate ET-1 secretion through the storage and transport system in non-myocytes .
目錄 頁次 壹、緒言……… 1 貳、實驗材料……… 18 參、實驗方法……… 27 肆、結果……… 35 伍、討論……… 40 陸、參考文獻……… 61
圖目錄
頁次
圖 A Doxorubicin………· 11
圖 B DOX 的 One-electron redox cycling……… 11
圖 C DOX, lipid peroxidation, 和 MDA-DNA adducts 的形成……… 12
圖 D Virtual cross-linking 的形成……… 13
圖 E Anthracyclines 的 C-13 secondary alcohol 代謝物……… 14
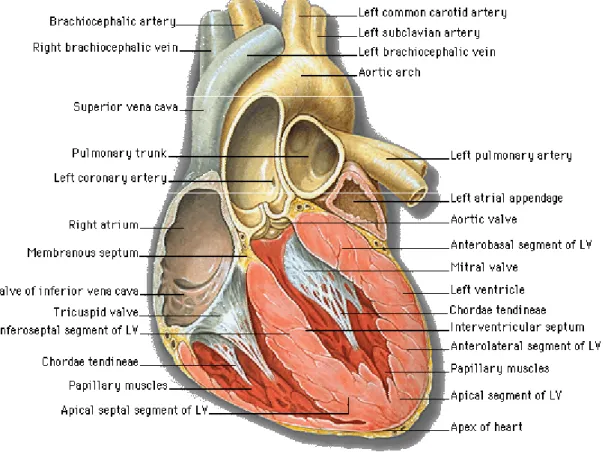
圖 F 心臟解剖圖……… 15
圖 G 心臟細胞間的溝通………· 16
圖 H Endothelin-1 的轉錄後修飾……… 16
圖目錄 頁次 圖 1. DOX 會刺激心肌細胞 ET-1 的分泌……… 47 圖 2. 不同劑量的 DOX 刺激心肌細胞 ET-1 之分泌……… 48 圖 3 DOX 刺激非心肌細胞的 ET-1 分泌迅速增加……… 49 圖 4 DOX 刺激非心肌細胞 ET-1 的分泌……… 50 圖 5 DOX 引發心肌細胞與非心肌細胞中不同時間模式的 ET-1 分泌… 51 圖 6 DOX 刺激心肌細胞 ET-1 基因的表現……… 52 圖 7 DOX 短暫抑制非心肌細胞 ET-1 基因的表現……… 53 圖 8 DOX 不影響 24 小時後非心肌細胞 ET-1 基因的表現……… 54 圖 9 DOX 刺激心肌細胞 ET-1 的表現有延遲的現象……… 55 圖 10 DOX 刺激非心肌細胞的 ET-1 表現受到其他機轉的調控………· 56 圖 11 DOX 處理使心肌細胞中的 ROS 量增加………· 57 圖 12 ROS 參與心肌細胞轉錄的活化……… 58 圖 13 人類冠狀動脈的 ET-1 運輸模式……… 59 圖 14 細胞中的 ROS………· 60
縮寫表 AP-1 BrdU CAT DOX DOXol DMEM ECE ERK ET-1 FBS GAPDH JNK MAPK MDA mRNA PBS ROS rRNA activator protein-1 5-bromo-2- deoxyuridine chloramphenicol acetyl transferase doxorubicin
doxorubicinol
Dulbecco's modified eagle's medium endothelin converting enzyme extracellular-regulated kinase endothelin-1
fetal brovine serum
glyceraldehyde-3-phosphate dehydrogenase c-Jun N-terminal kinase
mitogen-activated protein kinases malondialdehyde
messenger ribonucleic acid phosphate buffer saline reactive oxygen species ribosomal ribonucleic acid
壹、 緒言
阿黴素(adriamycin又稱作doxorubicin,簡寫為DOX ) ,14-hydroxy- daunorubicin,NSC-123127,FI-106,化學式C27H29NO11,屬於anthracycline
antibiotics 一 族 , IUPAC 名 稱 : 10-[(3-amino-2,3,6-trideoxya-L- lyxohexo-
pyranosyl ) oxy ]- 7,8,9,10 - tetrahydro - 6,8,11 -trihydroxy- 8 - (hydroxylacetyl)
- 1methoxy- 5,12-naphthacenedione (詳見圖A),是由放線菌目,鏈黴菌科的
Streptomyces peucetius var. caesius 所產生具抗癌活性的抗生素之一,因為 DOX會呈現橘紅色,因此在臨床使用上俗稱為「小紅莓」。臨床治療時,經
由靜脈注射後,DOX在血漿中濃度很快就下降,之後就分佈到肝、脾、腎、
肺 和 心 臟 。DOX 主 要 在 肝 臟 代 謝 , 而 由 膽 汁 中 排 出 ; 尿 排 泄 量 約 為
4~5%(Bachur et al., 1974; Peters et al., 1981) 。
它的抗癌機轉主要有5種 : 1. 阻礙拓樸異構酶(Topoisomerase)_而造 成DNA損傷:拓樸異構酶II具有解開DNA超螺旋的功能,可以在不改變去 氧核苷酸的結構和序列的情況下,暫時地造成DNA雙股斷裂,以解開雙股 螺旋的纏繞。拓樸異構酶II解開DNA超螺旋的活性,受到細胞週期及轉錄 活化的調節(Binaschi et al.,2001)。拓樸異構酶II是DOX的作用目標,當DNA 被切開並與拓樸異構酶II的tyrosine形成共價結合時,DOX的環狀結構會插 入DNA,形成DOX-DNA-topoisomerase II三元體,DOX沒有插入DNA的部
分扮演穩定三元體的角色,結果使得DNA斷裂(Binaschi et al.,2000,2001)。
這 樣 的DNA 斷 裂 造 成 細 胞 生 長 停 在 G1和G2期 , 以 及 造 成 細 胞 凋 亡
(Apoptosis)(Perego et al., 2001; Zunino et al., 2001) 。反之,腫瘤細胞會對 DOX產生抗藥性的原因之一,即來自於topoisomerase II基因的表現或活性 發生改變(Lage et al., 2000) 。 2. 形成自由基:DOX的C環quinone結構(詳 見圖B)在氧化還原作用中受到NAD(P)H-oxidoreductases (cytochrome P450 reductase、cytochrome b5 reductase、mitochondrial NADH dehydrogenase、 xathine dehydrogenase 和 endothelial nitric oxide synthase ) 所催化,當作反 應的電子接受者(Vasquez-Vivar et al., 1997; Minotti et al., 1999) 。一個自由 電子的加入使得quinones轉變成semiquinone free radicals,接著semiquinone free radicals藉由還原氧原子使本身迅速恢復為quinone結構,因此產生活性
氧族群(ROS),像是superoxide anion(O2‧﹣
)、hydrogen peroxide (H2O2)等;另 一方面semiquinone free radicals也會氧化A環和daunoamine之間的鍵結,產 生deglycosidation、7-deoxyaglycone以及H2O2。這些氧化還原作用增加了 DOX的脂溶性、醣苷鍵嵌入生物膜之能力以及ROS的產生(Gille and Nohl, 1997; Licata et al., 2000) 。DOX的自由電子氧化還原循環也伴隨著細胞內儲 存之iron(III)的釋放,DOX與釋放的iron(III)作用形成DOX-iron(II)化合物,
並且將O2‧﹣
和H2O2轉換成hydroxy radicals(OH‧
)。這些反應所產生的ROS不 只是會氧化細胞內的核酸、蛋白質與脂質,而造成細胞損傷。ROS也具有
訊息傳遞的功能,像是活化neutral sphingomyelinase與ceramide的產生。另
外, ROS還會調節許多與控制細胞週期以及促進或抑制細胞凋亡路徑有關
的kinases 或 transcription factors。 (Laurent and Jaffrezou, 2001; Bezombes et al., 2002; Kim et al., 2002; Martin et al., 2002) 。 3. 造成DNA損傷:最為 人所熟知的DNA損傷指的是DNA的鹼基被ROS氧化, DOX會經由氧化還
原 過 程 產 生 的 自 由 基 , 例 如OH‧
會 氧 化DNA 鹼 基 產 生 8-oxo-dG 和
8-oxo-dA。在接受DOX治療的肺癌病人的peripheral blood mononuclear cells (PBMC)中發現有Thymine glycol (ThyGly), 5-hydroxyhydantoin (5-OH-Hyd), 5-(hydroxymethyl) uracil (5-OH-MeUra), 4,6-diamino-5-formamido-pyrimidine (FapyAde)的產生,會增加造成變異的趨勢(Jaruga and Dizdaroglu, 1996)。 FapyGua 和 ThyGly 會阻礙 DNA replication 或增加 DNA polymerase 的 reading error frequency,造成細胞毒性或變異性的DNA損傷(Doroshow et al., 2001; Marnett et al., 2003)。DOX經由使脂質過氧化產生的malondialdehyde (MDA) , 會 和 DNA 之 deoxyguanosine (dG), deoxyadenosine (dA) 以 及 deoxycytidine (dC)的6環胺基作用,產生鹼性化的產物,例如dG 變成 pyrimidopurinone (M1dG) (詳見圖C) (Marnett et al., 2003)。MDA對細胞而言 是突變劑,其主要引起的突變發生在GC sites上,造成大範圍的插入或缺失 (Niedernhofer et al.,2003)。4. DNA 交叉連結:經由iron產生的自由基反應使
1998,1999; Taatjes and Koch, 2001) 。 DOX 和 HCHO 反 應 產 生 共 軛 物 (conjugate) :DOXFORM(Cutts et al.,2003),DOXFORM的代謝物能夠以共 價鍵結嵌入DNA的minor groove的5’-NGC-3’的dN與dG之間並與dG形成共 價鍵結,另一方面則與相對的另一股DNA的dG之間形成氫鍵,這種結構稱 作virtual cross-linking(詳見圖D) (Taatjes and Koch, 2001)。由於DOX會專一
性 地 引 起DNA 雙 股 在 5’-NGC-3’ 序 列 處 交 叉 連 結 , 5’-NGC-3’ 序 列 是
5'-untranslated regions的明顯特徵 (包括transcription factor-binding sites, promoter, and enhancer regions), DOX 作用在5’-NGC-3’ sites 會影響與 DNA 作用之蛋白質的結合,因此會抑制腫瘤細胞的DNA合成與細胞生 長。DOX抑制DNA的合成,受到迅早期基因的生長抑制訊息調控,這個訊 息調控與p53的功能有關(Kastan et al.,1991)。5.引起細胞凋亡(apoptosis): DOX作用的結果之一,是藉由活化p53和DNA的結合引起細胞凋亡,臨床 與非臨床的許多研究支持這點(Penault-Llorca et al., 2003; Ruiz-Ruiz et al., 2003; Stearns et al., 2003),但也有負面的結果(Inoue et al., 2000;Perego et al., 2001; Bertheau et al., 2002; Gariboldi et al., 2003),這是因為實驗所用腫瘤細 胞的不同,或是檢測p53情況及腫瘤細胞反應的方法有所不同。除了p53外 也有其他的機制參與例如, c-Jun N-terminal kinase (JNK) 的刺激與活化、 c-Jun/AP-1的c-Jun N-terminal kinase (JNK) 的刺激與活化 (Laurent and Jaffrezou, 2001); Akt/protein kinase B survival pathway之serine-threonine Akt
的degradation 和 down-regulation (Martin et al.,2002). DOX在臨床上屬於第一線及第二線的抗癌藥物,用於治療許多固體腫 瘤和血液腫瘤,例如乳腺癌、肉瘤、膀胱癌、甲狀腺癌、胃癌、卵巢癌和 小細胞肺癌、霍奇金氏和非霍奇金氏淋巴瘤,以及急性淋巴球性和急性骨 髓性白血病,是應用廣泛且有效的藥物。 但是DOX會對心臟造成傷害,因為DOX進入心肌細胞後會經由以下方 式產生心臟毒性: 1.產生自由基:自由基的產生如上所述。心臟對自由基 的傷害特別敏感,是因為相較於其他代謝的器官含有較少的去除超氧化物 毒性的物質(superoxide dismutase、glutathion 和 catalase),例如肝臟或腎臟 (Olson et al.,1981; Olson and Mushlin, 1990)。此外DOX已知對cardiolipin(心 肌細胞粒線體膜上的主要成分之一)有高度親和力,造成DOX累積在心肌細
胞中(Goormaghtigh and Ruyss-chaert, 1984) 。2.造成Ca2+
的過度累積:之前
的研究指出,在接受慢性之DOX處理的兔子心肌細胞中發現Ca2+異常增
加 , 且 累 積 在 心 室 的 心 肌 層 以 及 粒線體內 的鈣包 含體中(Olson et al.,
1974)。DOX造成的粒線體內鈣離子累積,需消耗氧化磷酸化產生的ATP, 造成高能磷酸鍵的減少(Ohhara et al., 1981) 。許多研究也指出DOX會造成 不正常的鈣離子運輸 (Boucek et al., 1997; Caroni et al., 1981);詳細的研究
reticulum中鈣離子的釋放 (Boucek et al., 1987; Ondrias et al., 1990; Boucek et al.,1993; Dodd et al., 1993 ) 。最近的研究發現DOX引起sarcoplasmic
reticulum Ca2+-ATPase 2的dowm-regulation,而且是經由H2O2的調節(Arai et
al., 2000 )。3. 阻礙ATPase的功能:DOX具有C-13 carbonyl functional group ,在心臟中會被代謝為C-13 alcohols(詳見圖E) ( Young and Raymond, 1986 )。Doxorubicinol (DOXol)是DOX主要的C-13 alcohol代謝物,會累積在 心臟組織中(Del et al., 1985; Rossini et al., 1986) ,在DOX引起的心臟毒性中 扮演重要的角色 (Danesi et al., 1987; Olson et al., 1988) 。從藥理學的觀點 來看,這可以解釋DOX處理後有延遲心臟毒性的現象,例如在動物活體的 實驗中,DOX注射到老鼠體內1小時後,心臟中的DOX濃度達到最高,然 而DOXol幾乎測不到;經過24小時後,心臟中的DOXol濃度逐漸增加,高 過肝臟中的濃度(Peters et al., 1981) 。DOXol對心臟具有毒性,會影響心臟
的 收 縮 及 舒 張 功 能 , 且 會 阻 礙sarcoplasmic reticulum 、 mitochondria和
sarcolemma 的ATPase活性( Olson et al., 1988 )。 4. 引起細胞凋亡:直到最 近幾年,細胞凋亡才被視為DOX引起之擴大性心肌症以及心臟衰竭的可能 之機轉,許多研究指出DOX會造成Cytochrome c release,之後經由Bax的
up-regulation(Wang et al., 1998a,b)或是Bcl-XL的down-regulation (Kim et
al.,2003; Kitta et al., 2003)形成apoptosome complex (apoptosis activating factor (Apaf-1/cytochrome c/pro-caspase-9)。一些研究結果指出細胞凋亡與
DOX的氧化還原有關,O2‧﹣
和H2O2 會誘發p53(Miyashita and Reed, 1998) 或是直接打開粒線體通透之暫時性孔洞 (Green and Leeuwenburgh, 2002; Clementi et al., 2003)。DOX的親脂性7-deoxyaglycone會累積在粒腺體內膜
上,大大增加呼吸鏈轉移給氧的電子,使得O2‧﹣
和H2O2增加,結果加速使 得粒線體失去功能 (Gille and Nohl, 1997)。DOX也會經由ROS活化p38 mitogen-activated protein kinases (MAPK) , 引 起 細 胞 凋 亡 (Kang et al., 2000)。DOX引起的心肌細胞凋亡可以經由一些參與活化phosphoinositide kinase (PI-3K)-serine/threonine kinase(Akt) 的 survival factors 來防止,像是 insulin growth factor-1 (Wu et al., 2000)、leukemia inhibitory factor( Negoro et al., 2001)。另外ET-1也會減輕DOX引起的心肌細胞凋亡(Araki et al.,2000; Suzuki et al.,2001; Tokudome et al.,2004)。
因為DOX的毒性會累積,當在人體中的劑量累積到550mg/m2以上就會 造成心臟損傷,嚴重者足以致命,所以DOX的使用需要受到小心地監控, 目前臨床上的檢測方法有下列幾種,endomyocardial biopsy:診斷DOX引起 之心肌症的黃金標準,具有高度敏感性與專一性、angiocardiography:對心 肌細胞壞死的偵測具有高度敏感性但卻為低專一性、echocardiograghy:非 侵 入 性 之 可 信 賴 的 檢 測 工 具 , 但 須 有 經 驗 的 操 作 員 來 操 作 、electro-
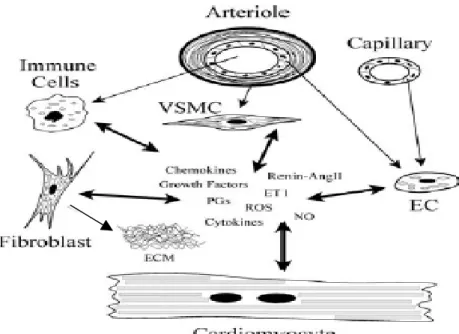
究指出endothelin-1 (ET-1)除了可以減輕DOX引起的心肌細胞凋亡外,還有 當作DOX引起心肌損傷指標的潛力(Yamashita et al.,1994,1995)。因此我們有 興趣的是DOX直接處理心臟細胞是否會引發ET-1的表現,以及其表現的模 式為何。 心臟(詳見圖 F)是人體非常重要的器官之一,藉由心臟的跳動帶動了全 身的循環。心臟是由心壁圍成的腔室,心壁的結構包括心内膜(endo- cardium)、心肌膜(myocardium)與心外膜(epicardium),由心肌細胞、 平滑肌細胞、內皮細胞、纖維母細胞、間皮細胞、束細胞、脂肪細胞以及 細胞外基質等所組成。在心臟的功能中,心肌細胞負責心臟的收縮,而其 他非心肌細胞則維持心臟運作正常,兩者之間藉由細胞激素、生長激素等 等的旁泌以互相溝通協調(詳見圖 G)(Ortega et al.,1997),內皮素是其中之 一,健康時參與調節心臟收縮功能,並在疾病發生時參與致病過程的機轉 (Goldie,1999)。 內皮素-1(endothelin-1,ET-1)是Yanagisawa et.al.(1988)由猪大動脈血管 內皮細胞培養液中分離、純化出來,它是由21個胺基酸所構成,共有三種 異構物(ET-1、ET-2、ET-3),其中ET-1是目前已知最有力的血管收縮劑。人 類ET-1的前驅物為一個含有203個胺基酸的大分子物質,經酵素切斷而形成 含38個胺基酸的big ET-1 ,再經轉換酶 (endothelin converting enzyme, ECE)
轉換成ET-1(詳見圖H),最後分泌至細胞外,此外ET的接受器也有二種: ET-A、ET-B(Ortega et al.,1997)。內皮素主要由血管內皮細胞所分泌,此外 還有上皮細胞、 神經元細胞、平滑肌細胞、腎間質細胞、網狀內皮細胞、 單核球及 多形核白血球細胞、造骨細胞、神經膠質細胞,及副甲狀腺細胞 等。內皮素在健康的情況下其功能是負責維持脈搏與血壓、調控鹽份與水 份的恆定、調控平滑肌的收縮、以及細胞的修復與生長。除血管組織外, 內皮素還廣泛地分佈在腦、肺、腎、心臟、脊髓、胃腸道等非血管組織。 在動物的心臟,包括動脈、靜脈、心房、心室中均含有內皮素,與心臟的 收縮功能有關。內皮素在血液中的半衰期約為1分鐘,其作用卻可以持續1 小時,肺、腎、肝的endopeptidase會清除內皮素,肺為清除血流中內皮素之 主要器官,當內皮素第一次流經肺臟時約有50%的內皮素被清除,約有90% 的內皮素是由肺臟所清除,其餘由腎清除,只有很少的部分由肝清除。心 臟疾病中,內皮素在心室肥大、心臟重塑、與心臟衰竭等的疾病機轉上被 廣泛地研究討論,此外急性心肌梗塞、心因性或敗血性休克、原發性高血 壓、肺動脈高血壓、狹心症,嚴重之動脈硬化症,出現雷諾氏現象之病人, 血漿中都有較高的內皮素值(Ortega et al.,1997; Goldie,1999)。
因為ROS在DOX引發的心臟毒性上扮演著重要的角色,另一方面在我 們實驗室之前的研究發現,心臟纖維細胞受到機械力、angiotensin II的作
用,會引起的轉錄因子AP-1與ET-1的轉錄活化且與ROS有關(Cheng et al., 2003a; Cheng et al.,2003b; Cheng et al.,2001),在內皮細胞中angiotensin II是 經由extracellular signal-regulated kinase pathway活化轉錄因子AP-1與ET-1基 因的表現(詳見圖I)(Hsu et al.,2004),angiotensin II引起的ET-1的轉錄活化會 受到抗氧化物17-beta-oestradiol的抑制(Chao et al.,2005)。在目前的研究中發 現,接受DOX治療而出現心臟毒性的肺癌病人(Yamashita et al.,1994,1995) 與老鼠的血清中(Sayed-Ahmed et al.,2001)可以發現血漿中的ET-1量增加; 但也有相反的結果(Mostafa et al.,2000;Mima et al.,2001 ;Murata et al.,2001; Zsary et al.,2002)。因此我們有興趣的是經由DOX氧化還原循環產生的ROS 是否會引發ET-1的轉錄。 我們經由細胞培養的方法將心臟細胞分為心肌細胞與非心肌細胞,當 DOX分別處理心肌細胞與非心肌細胞時,我們發現心肌細胞的ET-1 mRNA 和ET-1 分泌在DOX的刺激下表現量都會增加,且有時間延遲的現象。此 外,DOX會引發心肌細胞中ROS的明顯增加,且DOX刺激心肌細胞ET-1 的 轉錄活化與ROS(H2O2)有關。非心肌細胞受到DOX的刺激會引發ET-1 之早 期的短暫分泌高峰,而ET-1 mRNA在DOX的影響下初期表現量反而減少, 另外DOX不會引發非心肌細胞中的ROS增加,顯示非心肌細胞的ET-1 表現 可能受到其他機制的調控 。
圖A. Doxorubicin (Adriamycin; 10-[(3-amino-2,3,6-trideoxya-L- lyxohexo- pyranosyl ) oxy]-7,8,9,10-tetrahydro-6,8,11-trihydroxy-8-(hydroxylacetyl)-
1methoxy- 5,12-naphthacenedione) 由一個羥化的 tetracycline quinone 加上
一個醣化的duanosamine sugar側鏈,這個側鏈在 in vivo 和 ex vivo中都很容
易被羥化。( Minotti et al.;,2004)
圖C. DOX, lipid peroxidation, 和 MDA-DNA adducts 的形成。( Minotti et al.,2004 )
圖D. Virtual cross-linking的形成
經由iron產生的自由基反應使得DOX能作用產生formaldehyde (HCHO, FORM),DOX和HCHO反應產生conjugate (DOXFORM), DOXFORM的代謝物能夠以共價鍵結嵌入DNA的minor groove的
5’-NGC-3’的dN與dG之間,並與dG形成共價鍵結,另一方面則與相對 的另一股DNA的dG之間形成氫鍵,這種結構稱作virtual cross-linking。 R=-OH (DOX); Me,methyl residue in daunosamine. ( Minotti et al.,2004 )
圖E. Anthracyclines的C-13 secondary alcohol 代謝物。Anthracyclines受到 酶還原作用,產生 C-13 secondary alcohol 代謝物。 ( Minotti et al.,2004 )
圖F. 心臟解剖圖
圖G. 心臟細胞間的溝通。(Ortega et al.,1997)
圖I. ROS參與Angiotension II對ET-1 表現的調控。Angiotension II經由
ERK1/2 signal pathway活化內皮細胞的AP-1 轉錄因子並促進ET-1 的表
現。而ERK1/2 是經由ROS(H2O2)所活化,ROS的產生受到細胞膜上的
貳、實驗材料:
一、心肌細胞與非心肌細胞的初級細胞培養 器材
MillexGS(0.45µm): Millipore Corp. (Massachusetts ,U.S.A.) 。 眼科鑷子、眼科剪刀:Karl Klappenecker Surgical Instruments
(TN, German) 。
培養瓶(Trypsinizing Flask, Graduated with Screw Cap): Bellco Biotechnology(Vineland, U.S.A.) 。
培養皿,70µm 細胞濾篩(BD Falcon™ Cell Strainers) :BD Biosciences(CA ,U.S.A.)。
Shaker Bath,Model903 : HOTECH Instruments Corp.(Taipei, Taiwan, R.O.C.) 。
CO2恆溫培養箱,Model6300: NAPCO Inc.( Winchester Virginia, U.S.A.) 。
變頻式泛用冷凍離心機,Model5910:KUBOTA Corp.(Tokyo, Japan) 。
研究用顯微鏡(OPTISHOP),倒立式顯微鏡(TMS-F ba):Nikon(Tokyo, Japan) 。
(1)心臟組織之取得 (1-1)實驗動物
初生1-2 天的 Wistar rat,來自台大動物中心。
(1-2) 1XPBS buffer
Phosphate Buffered Saline : Bio Basic Inc.( Ontario,Canada) 。 (1-2-1)Phosphate Buffered Saline 一罐,溶於去離子水,調 pH 到
7.4 後,補去離子水到 1 升,再稀釋 10 倍,滅菌之後使用。
(2)心肌細胞之取得
(2-1) 0.125% trypsin digesting solution
trypsin (1:250): Gibco BRL(Grand Island,NY,U.S.A.) 。 (2-1-1)秤 0.125g trypsin (1:250),溶在 100ml 的 1XPBS buffer 中,
將50c.c.針筒裝上 0.45µm 的針孔過濾器,把溶液過濾到滅
菌過的玻璃瓶中,放在冰上備用。
(2-2)細胞培養液
F10 Nutrient Mixture、Dulbecco’s Modified Eagle Medium
(DMEM)、FBS (fetalbovine serum)、Penicillin-streptomycin:
Insulin, Transferrin, 5-bromo-deoxyuridine(BrdU): Sigma(St. Louis, MO, U.S.A.) 。
(2-2-1) F10
80% F10 Nutrient Mixture、20% FBS、5 µg/ml 的 insulin 和 transferrin、1% 的 penicillin 和 streptomycin。
(2-2-2)DMEM-BrdU
90% DMEM、10% FBS、30.7µg/ml 的 BrdU 、5 µg/ml 的 insulin 和 transferrin、1% 的 penicillin 和 streptomycin 。
(3)非心肌細胞之取得 (3-1) DMEM
90% DMEM、10% FBS、5 µg/ml 的 insulin 和 transferrin、 1% 的 penicillin 和 streptomycin 。
二、以 Doxorubicin 處理細胞 (1) Serum-Free DMEM
DMEM、 5 µg/ml 的 insulin 和 transferrin、 1% 的 penicillin 和 streptomycin 。
Doxorubicin, Dimethyl Sulfoxide(DMSO): Sigma(St. Louis, MO, U.S.A.) 。
(2-1)Doxorubicin 溶在 DMSO 中。
三、ET-1 ELISA
Endothelin-1 Biotrak Assay : Amersham Biosciences (Buckinghamshire, England) 。
四、RT-PCR 器材
Spectrometer U-2000: HITACHI Inc. (Tokyo, Japan) 。
小型電泳裝置(Major science MP-150): Instrument of Biomedical Science (London,England) 。
Firefox Dry Bath 6100: Pantech instruments(Taipei, Taiwan, R.O.C.) 。
GeneAmp PCR system 2700 : AME Bioscience Ltd.(Toroed, Norway) 。
(1) total RNA 之取得
(2)RNA量測定 滅菌水。
(3)total RNA 的 rRNA 比例檢測
Agarose A: Bio Basic Inc. ( Ontario,Canada) 。
EtBr, 10X TAE buffer : Sigma(St. Louis, MO, U.S.A.) 。
(4) Superscript II RT
oligo dT、25mM dNTP、 RNase inhibitor、Superscript II reverse transcriptase、5X 第一次反應緩衝液 : Invitrogen(San Diego, CA, U.S.A.) 。
(5)PCR
2mM dNTP、Taq DNA polymerase buffer、Taq DNA polymerase : Qiagen (Hilden, Germany)。
合成的引子: 誠樺(Taipei, Taiwan, R.O.C.) 。
ET-1 primers 5’端 GCTCCTGCTCCTCCTTGATG 3’端 CTCGCTCTATGTAAGTCATGG GAPDH primers 5’端 TATGATGACATCAAGAAGGTGG
(6) 瓊脂凝膠電泳
Agarose A: Bio Basic Inc. ( Ontario,Canada) 。
EtBr, 10X TAE buffer : Sigma(St. Louis, MO, U.S.A.) 。
五、細胞內ROS測定 器材
SYBRON/Thermolyne Speci-Mix : Quality Lab Excess, Inc. (Michigan, U.S.A.) 。
Fluorescence Spectrophotometer F-4010: HITACHI , Inc. (Tokyo, Japan) 。
(1) 10mM nonpolar 2’,7’-DCFH-DA
nonpolar 2’,7’-DCFH-DA(nonpolar 2’,7’-dichlorofluorescein
diacetate):Molecular Probes, Inc.( Oregon, U.S.A.) 。
(1-1) 1mg nonpolar 2’,7’-DCFH-DA溶在205µl酒精中。
六、以 CAT assay 測 ET-1 promoter 的活性
水浴槽:SPG Media Limited(London,United Kingdom) 。
X 光底片: Kodak (Rochester, NY, U.S.A.) 。
Whatman 3MM filter paper: Whatman (Middlesex,UK)。
(1) 細胞轉染
lipofectamineTM Transfection Reagent: Invetrogen(San Diego, CA,
U.S.A.) 。
(2) 以 catalase,DOX 處理細胞 (2-1) Serum-Free DMEM
DMEM、 5 µg/ml 的 insulin 和 transferrin、 1% 的 penicillin 和 streptomycin 。
(2-2)Doxorubicin
Doxorubicin, Dimethyl Sulfoxide(DMSO): Sigma(St. Louis, MO, U.S.A.) 。
(2-3)catalase from human erythrocytes: SIGMA CHEMICAL CO.(St. Louis, U.S.A.)。
(3) CAT assay
(3-1) 細胞萃取液之製備
(3-1-1)EDTA(0.1M) 1.861gNa2.EDTA.2H2O溶在去離子水,調到pH8.0,最終 體積 50ml。 (3-1-2)Tris.Cl(1M) 24.2gTris.Cl 溶在離子水,調到 pH7.5,最終體積 200ml。 (3-1-3)NaCl(0.6M) 7gNaCl 溶在 200ml 去離子水。 (3-1-4)TEN solution 取 5ml EDTA(0.1M)、20mlTris.Cl(1M)、125mlNaCl(0.6M) 加上 350ml 去離子水,配成 500ml TEM solution。 (3-1-5)0.25M Tris.Cl(pH7.5) 1M Tris.Cl(pH7.5)以去離子水稀釋 4 倍。 (3-2)細胞萃取液反應進行之測定
200µCi[14C]-chloramphenicol: Amersham( Buckinghamshire,
England) 。
Ethyl acetate, Acetyl CoA:Merck (Darmstadt, Germany) 。 (3-2-1)10mM acetyl CoA
(3-3)薄層色層分析法之進行
參、實驗方法:
一、心肌細胞與非心肌細胞的初級細胞培養 (1)心臟組織之取得 將初生 1 到 2 天的小鼠放在舖有鋁箔紙的平台上,以膠帶固定 四肢,用酒精棉球消毒胸腹部,以眼科剪刀沿肋骨中間剪開,以眼科鑷 子輕輕夾取心臟,將心臟剪下放在盛有20c.c. 1XPBS的培養皿中,剪刀 與鑷子用酒精棉清理並以火烤過備用。將心室剪下並從中間剪開,輕輕 撥動以洗去血液,在不同培養皿中清洗至少3 次。最後放在盛有 10c.c. 1XPBS的培養皿中,用眼科剪刀將心臟剪成 1mm3 的組織塊。 (2)心肌細胞之取得 將所有組織吸到震盪三角瓶中,以1XPBS洗一次,然後trypsin digesting buffer (約 10c.c./ heart),以 37℃、50-100rpm 震盪 10 分鐘,之後收集懸 浮液到離心管中,加入1/10 體積的 F10 ,放在冰上。重複數次,待組 織分解完畢,將所收集之懸浮液以 1600rpm、3 分鐘離心,去掉上清液, 用 F10 重新懸浮細胞,以 70µm 細胞濾篩過濾,再用 1600rpm、3 分鐘 離心, 2.5 小時,仍懸浮的細胞則大部分為心肌細胞(>90%)。以 1600rpm、3 分鐘離心收集懸浮的細胞、計數,每 60-mm培養皿種 3x106DMEM-BrdU,待 2 天後可觀察到會跳動的心肌細胞,接著就可以進行 實驗。 (3)非心肌細胞之取得 以0.125% trypsin處理心臟所得的細胞,用 F10 培養基培養 2.5 小時後, 黏附在培養皿上的細胞即為非心肌細胞,以DMEM 培養 10 天(2 代) 的 非心肌細胞,取1.5×106種在100-mm培養皿中,即可進行實驗。 二、以 Doxorubicin 處理細胞 (1)Dox 處理之時間點 細胞以serum-free DMEM 培養 24 小時後,用 1X PBS 清洗一次,換上
含1µM DOX 的 serum-free DMEM 分別培養 1、3、6、12 、24 小時,
之後分別收集培養液與細胞,收集的培養液儲存在-20℃,沉澱的細胞以 350 µl RLT solution 打散後儲存在-70℃,可以保存數個月。
(2) Dox 處理之劑量
細胞以 serum-freeDMEM 培養 24 小時後,以 1X PBS 清洗一次,分別 換上含 0.1、0.01、1、10 µM DOX 的 serum-free DMEM 培養 24 小時, 之後分別收集培養液與細胞收集的培養液儲存在-20℃,沉澱的細胞以
350 µl RLT solution 打散後儲存在-70℃,可以保存數個月。
三、ET-1 ELISA
除了空白對照組外,取standard solution 100µl、assay buffer 100µl、 樣本100µl,皆2次重複,加入已吸附有ET-1抗體的96孔盤,放在3-5℃ 到隔天。倒掉所有液體,以400µl wash buffer洗4次,除了空白對照組 外,所有well加入100µl ET-1 antibody conjugate (含 Horseradish Peroxidase標記的Fab' fragment) , 37℃培養30分鐘。倒掉所有液體,以 400µl wash buffer洗4次,確定所有溶液都移除之後,加入100µl TMB substrate,室溫 (15–30℃) 搖晃培養40分鐘。最後加入100 µl 1.0 M H2SO4終止反應,在30分鐘內以 ELISA reader 讀取450nm吸光值。
四、RT-PCR
(1) total RNA 之分離
將-70℃儲存的細胞解凍,震盪打散細胞,將樣本通過直徑 0.9mm 的針頭 5 次,加入等體積的 70% 酒精混合均勻,將所有的樣本加到 RNeasy Mini Column set中,以 10,000 rpm 離心15秒,倒掉管內液體。 加700µl RW1到 RNeasy Mini Column set 中,以 10,000 rpm 離心15 秒,倒掉管內液體。將RNeasy Mini Column 移到新的 2ml 離心管,加
入500µl RPE,以 10,000 rpm 離心15秒,倒掉管內液體。加500µl RPE 到RNeasy Mini Column set中,以 10,000 rpm 離心2分鐘,倒掉管內液 體。最後將RNeasy Mini Column移到新的 1.5ml 離心管,加入30µl RNase-free water,以 10,000 rpm 離心1分鐘,得到total RNA,儲存在 -70℃。
(2) RNA量測定
取 2µl total RNA,加 68µl RNase-free water,以光電比度計測 A260/A280( nm) 吸光值,RNA濃度以下列公式換算求得:
RNA濃度 (µg/ml) = OD260 As值 × 40 × 稀釋倍數
(3) total RNA 的 rRNA 比例檢測
取 5µl total RNA,進行 1%瓊脂凝膠電泳,檢測 18S rRNA、28S rRNA 比例。
(4) Superscript II RT
取2.5µg 的 total RNA, 加入 oligo dT 0.5µl、25mM dNTP 0.5µl、
滅菌之去離子水6.5µl,65℃反應 5 分鐘,接著放到冰上逾 10 分鐘。接
5X 第一次反應緩衝液 2.5µl,37℃反應 1 小時,合成 cDNA。
(5) PCR
取反轉錄作用成的cDNA 2µl,加入 ET-1 引子各 0.5µl 或 GAPDH 引
子各0.5µl、2mM dNTP 5µl、Taq DNA polymerase buffer 5µl、Taq DNA
polymerase 0.25µl、滅菌去離子水 36.25µl。反應 94℃、30 秒,55℃、30 秒,72℃、1 分鐘,25 個循環。 (6)瓊脂凝膠電泳 取10µl RT-PCR 產物,進行 1%瓊脂凝膠電泳,電泳結束,以紫外光影 像分析儀檢測mRNA 的表現。 五、細胞內ROS測定 將以doxorubicin處理的細胞及控制組的細胞用1XPBS洗2次,接著以 trypsin 處理2分鐘,收集下來以200rpm、5分鐘離心,重新以1XPBS打散 細胞,計算細胞數,之後每管加入5µl 的10 mM nonpolar 2’,7’-DCFH-DA (nonpolar 2’,7’- dichlorofluorescein diacetate),避光下,在37℃、震盪反應30 分鐘,之後以1200xg、5分鐘離心,沉澱的細胞重新以1XPBS懸浮,測 525/475nm螢光光譜。
六、以 CAT assay 測 ET-1 promoter 的活性 (1)細胞轉染
(1-1)轉染-1.7kCAT plasmid
5µg -1.7kCAT plasmid 和 10µl lipofectamine 分別加入不含血清及
抗生素的DMEM 到 800µl,彼此混合作用 45 分鐘後,加同樣的培養
液5 毫升,處理細胞 16 小時後,將 DMEM 盡量吸乾淨,換上
10%DMEM 終止反應。 (1-2)轉染 pBLCAT2 plasmid
5µgpBLCAT2 plasmid 和 10µl lipofectamine 分別加入不含血清及
抗生素的DMEM 到 800µl,彼此混合作用 45 分鐘後,加同樣的培養
液5 毫升,處理細胞 16 小時後,將 DMEM 盡量吸乾淨,換上
10%DMEM 終止反應。 (1-3)轉染 pBLCAT3 plasmid
5µg pBLCAT3 plasmid 和 10µl lipofectamine 分別加入不含血清及 抗生素的 DMEM 到 800µl,彼此混合作用 45 分鐘後,加同樣的培養 液 5 毫升,處理細胞 16 小時後,將 DMEM 盡量吸乾淨,換上 10% DMEM 終止反應。
轉染的細胞培養48 小時後,以 catalase(350U/ml)處理心肌細胞 3 小 時,接著以DOX 處理細胞 6 小時,接著收集細胞。 (3) CAT assay (3-1)細胞萃取液之製備 細胞以 PBS 洗 2 次後,加 1ml TEN 溶液,放冰上 5 分鐘,將細 胞刮下以4℃、14,000rpm、1 分鐘離心。之後以 0.25M Tris.Cl(pH7.5) 打散細胞,接著以超音波的方式打破細胞,4℃、14,000rpm、10 分 鐘離心取上清液,儘速凍在-70℃,之後移到-20℃儲存。 (3-2) 細胞萃取液反應進行之測定 取樣本加1MTris.Cl(pH7.5) 到 120µl ,繼續加 35µl H2O 、4µl
200µCi[14C]-chloramphenicol,20µl 10mM acetyl CoA, 37℃水浴 1 小
時後,每管再加入 10µl 10mM acetyl CoA繼續 37℃水浴 2 小時,最後
加入300µl ethyl acetate終止反應,震盪 1 分鐘,14,000rpm、3 分鐘離
心取上層液,之後在快速真空乾燥機中乾燥。 (3-3) Perform thin-layer chromatography
準備TLC tank ,加入190 ml chloroform, 10 ml methanol,以及What- man 3MM filter paper,以凡士林封住空隙,靜置2小時備用。將樣本
點相距1.5cm並吹乾,在有200ml 之 19:1 chloroform /methanol的TLC tank中伸展1小時,乾燥後壓片。
肆、結果:
一、Doxorubicin 刺激心肌細胞 ET-1 的分泌 從 初 生 1-2 天 的 Wistar rat 分 離 培 養 出 來 的 心 肌 細 胞 , 經 由 DMEM-BrdU 培養 2 天,讓心肌細胞可以回復,2 天後心肌細胞會開始 跳動,每分鐘約可跳動125 次。換上 sreum-free DMEM 培養 24 小時 後,以 DOX 1µM 處理細胞 1、3、6、12、24 小時。收集培養液,用Endothelin-1 Biotrak Assay 測 ET-1 的量,結果發現 DOX 會引起心肌細
胞的 ET-1 分泌,在 12 小時 ET-1 的分泌量明顯比對照組多,而 24 小
時 ET-1 的分泌量雖然下降,但是仍然比對照組多。我們也觀察到以
DOX 處理 24 小時後,雖然會造成心肌細胞損傷,但是心肌細胞仍會 跳動。12 小時 DOX 處理的心肌細胞之 ET-1 分泌量是 14.7 fmol/ml,
對照組沒有測到 ET-1 分泌。24 小時 DOX 處理的心肌細胞之 ET-1 分
泌量是8.7fmol/ml,對照組是 6.9 fmol/ml。雖然對照組沒有加入 DOX,
但是在無血清的環境下,隨時間增加心肌細胞的ET-1 分泌量也會逐漸
增加 (詳見圖 1)。若是以 0.01、0.1、1、10µM 的 DOX 處理細胞 24 小
時之後,測量 ET-1 的分泌,發現 1µM、10µM 的 DOX 明顯引起心肌
細胞的ET-1 分泌,但是 0.1µM 的 DOX 會抑制 ET-1 的分泌。0.1µM、
應的現象(詳見圖 2)。 二、Doxorubicin 會刺激非心肌細胞 ET-1 的分泌 從初生 1-2 天的Wistar rat分離培養出來的非心肌細胞,可以繼代 培養,最多可以繼代3 到 4 次,非心肌細胞會增殖,但隨著天數增加, 非心肌細胞的生長能力會逐漸下降,變的容易死亡,對trypsin的耐受 性降低,約 17 天後細胞的型態也會有所改變,約 20 天後細胞便會死 亡,因此我們選擇繼代 2 次的非心肌細胞進行實驗。取繼代 2 次的非 心肌細胞,每培養皿種 1.5x106細胞,以DMEM培養 1 天,接著換上
sreum-free DMEM 培養 24 小時後,以 DOX 1µM處理細胞 1、3、6、 12、24 小時,收集培養液,用Endothelin-1 Biotrak Assay測ET-1 的量,
結果發現處理1 小時的非心肌細胞之ET-1 分泌量是 11.2fmol/ml,對照
組是 3.9 fmol/ml。處理 3 小時的非心肌細胞之ET-1 分泌量是 17.5
fmol/ml,對照組是 3 fmol/ml。處理 6 小時的非心肌細胞之ET-1 分泌量
是 30 fmol/ml,對照組是 11.4 fmol/ml。處理 24 小時的非心肌細胞之
ET-1 分泌量是 13.7 fmol/ml,對照組是 24.5 fmol/ml (詳見圖 3),受到 DOX刺激的非心肌細胞 6 小時內ET-1 分泌量增加 3 到 6 倍,但是 24 小時ET-1 的分泌量卻明顯降低。若是以 0.01、0.1、1、10µM的DOX處
理的非心肌細胞之ET-1 分泌量是 15.5 和 25.3 fmol/ml,對照組是 19 fmol/ml,沒有dose-dependent的現象,以DOX處理非心肌細胞 24 小時 後,1µM DOX會抑制非心肌細胞ET-1 的分泌,但是 10µM DOX則會增 加非心肌細胞ET-1 的分泌(詳見圖 4)。 三、Doxorubicin 會刺激心肌細胞 ET-1 基因的表現 從初生 1-2 天的Wistar rat分離培養出來的心肌細胞,經由DMEM- BrdU培養 2 天後,換上sreum-free DMEM 24 小時後,以DOX 1µM處
理細胞1、3、6、12、24 小時,收集細胞後,加入RLT solution(這樣細
胞可保存在-70℃一個月),接著打散細胞,將細胞通過直徑 0.9mm 的
針頭5 次以打破細胞,接著以RNeasy Product Mini Kit取得total RNA。
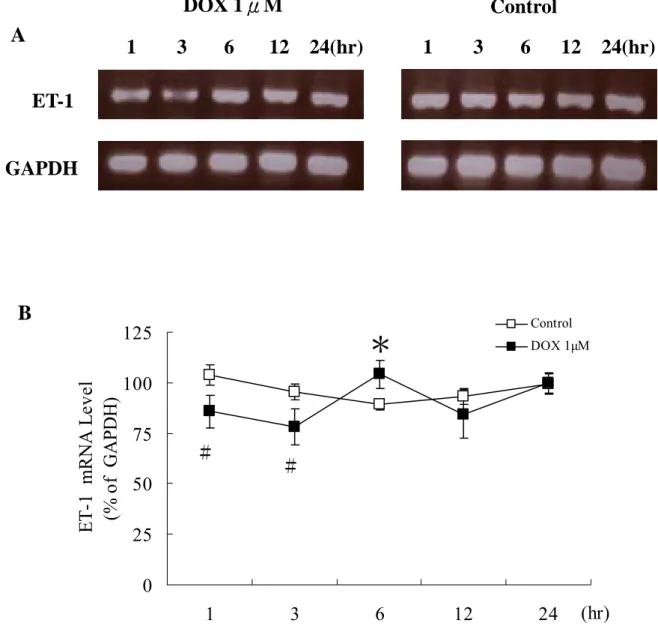
得到的RNA測吸光值,A260/A280比值≧2。將這些total RNA做RT-PCR, 結果發現DOX處理的心肌細胞之ET-1 在 1 小時就有表現,而在 3 小時 則有高量的表現,之後就顯著減少,但在 24 小時的處理,心肌細胞死 亡 很 多(細胞數≦5 x 106 ),所以無法得到足量的total RNA進行 RT-PCR。對照組的心肌細胞在無血清的環境下,隨時間增加ET-1 的表 現也會逐漸增加(詳見圖 6)。我們還發現DOX刺激心肌細胞ET-1 基因 的表現到ET-1 分泌中間有一段時間延遲(詳見圖 9 )。
四、Doxorubicin 會短暫抑制非心肌細胞 ET-1 基因的表現
從初生 1-2 天的Wistar rat分離培養出來的非心肌細胞,經由DMEM
培養2 代後,每培養皿種 1.5x106細胞,換上sreum-free DMEM 24 小時
後,以DOX 1µM處理細胞 1、3、6、12、24 小時,收集細胞後,加入 RLT solution(這樣細胞可保存在-70℃一個月),接著打散細胞,將細胞 通過直徑 0.9mm 的針頭 5 次以打破細胞,接著以RNeasy Product Mini Kit取得total RNA。得到的RNA測吸光值,A260/A280比值≧2。將這些total RNA做RT-PCR,結果發現DOX處理的非心肌細胞之ET-1 在 1~3 小時 間的表現受抑制,之後就恢復與對照組相同。對照組的非心肌細胞之 ET-1 則在平時就有穩定表現(詳見圖 7)。在處理DOX 24、27、30、36、 48 小時的非心肌細胞中,ET-1 的轉錄和對照組並無差異(詳見圖 8)。 五、受 DOX 刺激使得心肌細胞中 ROS 的量增加 DOX處理會使心肌細胞中的ROS量增加。從初生1-2天的Wistar rat 分離培養出來的心肌細胞,經由DMEM-BrdU培養2天後,心肌細胞開始
跳動,換上sreum-free DMEM 培養24小時後,以DOX 1µM處理細胞,1、 3、6、12、24小時,收集細胞。離心去掉上清液後,以1X PBS重新懸浮 細胞,加入nonpolar 2’,7’-DCFH-DA (nonpolar 2’,7’- dichloro- fluorescein diacetate),避光下,在37℃震盪反應30分鐘,離心去掉上清液,沉澱的
細胞重新以1XPBS懸浮,計算細胞數目以及測525/475nm螢光光譜,DOX 處理心肌細胞 1, 3, 6, 12, 24小時後,與對照組相比,ROS增加132%、 152%、135%、137%、121%(詳見圖11)。但是非心肌細胞以DOX處理1, 3, 6, 12, 24小時後,與對照組相比,ROS並無差異。
六、ROS 參與心肌細胞轉錄的活化
心肌細胞轉染-1.7kCAT plasmid、pBLCAT2 plasmid 及 pBLCAT3 Plasmid,轉染的細胞培養48小時後,以catalase 350U/ml先處理3小時,
再以DOX處理6小時,接著收集細胞。 取樣本加入[14C]-chloramphenicol
和 acetyl CoA作用,之後加入ethyl acetate終止反應,最後進行TLC。 經由X光底片壓片後的結果發現,DOX會活化心肌細胞的ET-1 promotor,且這個作用會受到catalase的抑制,顯示這個活化是經由 ROS相關的訊息傳遞途徑所調控。
伍、討論:
低劑量與高劑量的 DOX 可能經由不同途徑影響心肌與非心肌細胞。在 我們的實驗中,DOX 引起之心肌與非心肌細胞的 ET-1 分泌沒有 dose- dependent 的現象,有趣的是以 DOX 處理心肌細胞 24 小時後,0.1µM DOX
會抑制心肌細胞ET-1 的分泌,但是 1µM 和 10µM DOX 則會增加心肌細胞
ET-1 的分泌。而以 DOX 處理非心肌細胞 24 小時後,1µM DOX 會抑制非
心肌細胞ET-1 的分泌,但是 10µM DOX 則會增加非心肌細胞 ET-1 的分泌。
在之前發表的報告中,DOX 處理的老鼠血漿中 ET-1 值會上升,但是沒有 dose- dependent 的現象。實驗中老鼠從腹腔每次注射 5 mg/kg DOX,隔天
注射一次,使五組老鼠 DOX 的累積劑量分別達到 5 , 10, 15, 20 和 25
mg/kg,以及一組單一次注射高劑量 20 mg/kg。在最後一次注射的 24 小時
後,測血漿中ET-1 量。DOX 累積 10, 15, 20 mg/kg 的組別和只注射生理食
鹽水的組別相比,增加85%, 76% 和 97% ,而且在單一次注射 20 mg/kg 的
組別,發現血漿中 ET-1 量特別高,增加 130% (Sayed-Ahmed MM et.al.,
2001)。除此之外,在細胞凋亡相關研究中,受到高劑量與低劑量的 DOX
處理的細胞表現有所差別,例如以1µM DOX 處理 p388 leukemic cells 會引
起細胞凋亡但在10µM DOX 處理下卻沒有(Ling et al.,1993);Gewirtz 整理
機制殺死腫瘤細胞(Gewirtz,1999)。這些結果似乎顯示 DOX 在低劑量與高 劑量時,採用不同的機制影響細胞。
ET-1 雖然在血漿中的 half-life 只有幾分鐘,但是作用時間卻可以持續 1
小 時 , 所 以 非 心 肌 細 胞 受 到 DOX 刺 激 後 ET-1 的 分 泌 量 迅 速 增 加
(paracrine),ET-1 有保護心肌細胞,減輕 DOX 引起的心肌細胞凋亡的作用,
加上心肌細胞本身受DOX 影響也會自我分泌 ET-1 (autocrine)。在 Takahiko,
S. 和 Takashi, M. 的研究中證明將心肌細胞前處裡 ET-1 24 小時之後,可
以保護心肌細胞,減輕 DOX 引起的心肌細胞凋亡,作用可持續到 24 小時
(Takahiko, S. and Takashi, M. ,2001)。在我們的實驗中,1µM DOX 處理下,
非心肌細胞分泌的高峰出現在 6 小時,而心肌細胞出現在 12 小時(詳見圖
5 ),若綜合兩種細胞的表現模式,ET-1 的分泌增加可以持續到 12 小時, 在 in vivo 上是否能得到相似的表現模式,需要更進一步的研究證明。
非心肌細胞的ET-1 分泌受到其他機制的調控。觀察非心肌細胞以 DOX
處理的結果,我們發現DOX 在 3 小時內會抑制 ET-1 的轉錄,但是在 ET-1
的分泌量上我們卻觀察到,ET-1 的分泌在 6 小時內明顯地增加(詳見圖 1),
我們推論是因為非心肌細胞平時就會穩定地表現 ET-1,即使在無血清的環
ET-1 的轉錄,但卻經由 Weibel-Palade bodies degranulation 的機轉增加了 ET-1 的分泌(詳見圖 13)。在人類內皮細胞中的蛋白質由內質網到細胞膜的 運輸經由二個完全不同的分泌途徑:一個是持續性的途徑,一個是受調節
的途徑。持續性的途徑受mRNA 的表現所調節(Yanagisawa et al.,1988) ;
受調節的途徑則和細胞中的儲存胞器Weibel- Palade bodies 的調節有關。之
前的研究報告,當機械張力作用在培養的牛動脈內皮細胞上,20 分鐘內,
在培養液中就能測到ET-1(Macarthur et al.,1994)。而在老鼠的內皮細胞中有
Weibel-Palade bodies 的存在(Doi et al., 1996; Ozaka et al.,1997)。因此在我們
的 實 驗 中 , 非 心 肌 細 胞 很 有 可 能 是 受 到 DOX 刺 激 後 , 很 快 的 經 由
Weibel-Palade bodies degranulation 分泌出 ET-1 ,使 ET-1 的分泌在 6 小時
內明顯地增加。但是在 6 小時後,因為 ET-1 mRNA 表現的早期抑制、加上
儲存在Weibel-Palade bodies 中的 ET-1 被分泌出去,還有 ET-1 本身的 half-life
只有幾分鐘,因此使得6 小時後培養液中的 ET-1 迅速下降。
DOX對心肌細胞和非心肌細胞的影響有很明顯的不同。在之前的研究 中已經確定拓樸異構酶II是DOX的作用目標,當DNA被切開並與拓樸異構
酶II 的 tyrosine 形 成 共 價 結 合 時 , DOX 的 環 狀 結 構 會 插 入 DNA , 形 成
DOX-DNA-topoisomerase II三元體,DOX沒有插入DNA的部分扮演穩定三 元體的角色,結果使得DNA斷裂(Binaschi et al.,2000,2001)。這樣的DNA斷
裂 造 成 細 胞 生 長 停 在G1和G2期 以 及 造 成 計 畫 性 的 細 胞 死 亡(Perego et al.,2001; Zunino et al., 2001)。反之,腫瘤細胞會對DOX產生抗藥性的原因 之一,就是因為topoisomerase II基因的表現或活性發生改變(Lage et al., 2000)。至於為何DOX對於非心肌細胞的ET-1 RNA的表現有抑制的現象但 是對心肌細胞卻沒有抑制的原因,可能就是因為DOX的會嵌入DNA、抑制 拓樸異構酶II的活性,因此DOX對於會分裂的細胞影響特別大,而心肌細 胞是不會再分裂的細胞,因此心肌細胞的ET-1 mRNA的表現不會受到DOX 的抑制。 此外,我們發現心肌細胞ET-1的轉錄與ET-1的分泌有一時間上的差異 (詳見圖9)。我們推論是因為ET-1轉譯成蛋白質後,還需要經過細胞內酶的 剪切,由prepro-ET變成big ET,還要經ECE(endothelin converting enzyme) 剪切成為ET-1,最後才分泌出去,因此ET-1的轉錄與ET-1的分泌才會有時 間上的差距。從之前研究牛生殖系統ET-1表現的real-time結果發現,ET-1 mRNA的表現高峰出現在12~24小時,但是ET-1 protein分泌的高峰則是出現 在24~48小時(Schams et al.,2003)。此外另一個可能性是由於DOX代謝產物
對 心 肌 細 胞 的 影 響 大 於DOX 的 緣 故 , DOX 在 心 臟 中 會 被 代 謝 為 C-13
alcohols ( Young and Raymond, 1986 ),Doxorubicinol (DOXol)是DOX主要的 C-13 alcohol代謝物,會累積在心臟組織中(Del et al., 1985; Rossini et al.,
1986) ,在DOX引起的心臟毒性中扮演重要的角色(Danesi et al., 1987; Olson et al., 1988) 。從藥理學的觀點來看,這可以解釋DOX處理有延遲之心臟毒 性的現象,從實驗中發現DOX注射到老鼠體內1小時後,心臟中的DOX濃 度達到最高,然而DOXol幾乎測不到;經過24小時後,心臟中的DOXol濃 度逐漸增加,高過肝臟中的濃度(Peters et al., 1981) 。DOXol能比ROS促進 更多的鐵離子之釋放,鐵離子與ROS的作用產生更多自由基,在牛動脈內 皮細胞中及H9c2心肌細胞中會造成apoptosis (Minotti et al., 2001; Kotamraju et al.,2002)。而DOX造成的心臟衰竭與ET-1的表現有正相關性,因此心肌細 胞ET-1的延遲分泌有可能和DOX的延遲心臟毒性有關。 ROS的增加會促進心肌細胞的ET-1的表現,在之前的許多研究已經證 實ROS在心臟血管疾病中扮演訊息傳遞的角色,以活化轉錄因子及下游基 因的表現。ROS的產生是經由細胞內酶的作用,而catalase有去除細胞內H2O2 功能(詳見圖14,A)。細胞外的刺激會增加細胞內的ROS,抗生素,例如DOX 的quinone結構受到NAD(P)H-oxidoreductases (cytochrome P450 reductase、 cytochrome b5 reductase、mitochondrial NADH dehydrogenase、xathine dehydrogenase 和 endothelial nitric oxide synthase ) 所催化,當作反應的電 子接受者,使得quinones轉變成semiquinone free radicals,接著semiquinone free radicals藉由還原氧原子使本身迅速恢復為quinone結構,因此產生活性
氧族群(ROS),像是superoxide anion(O2‧﹣
)、hydrogen peroxide (H2O2)等 (Vasquez-Vivar et al., 1997; Minotti et al., 1999)。另一方面semi- quinone free radicals也會自我氧化,產生deglycosidation和7-deoxyaglycone以及H2O2。此 外,DOX的親脂性7-deoxyaglycone會累積在粒腺體內膜上,大大增加呼吸
鏈轉移給氧的電子,使得O2‧﹣
和H2O2增加(Gille and Nohl, 1997)。這些ROS 會參與MAPKs pathway的調控以及NF-κB的活化等,MAPKs pathway會活化 基因的AP-1 sites,例如ET-1的promoter上的AP-1 site ,進而活化下游基因 的轉錄;而NF-κB的活化與細胞凋亡有關(詳見圖23,B) (Jun-ichi, A. and Bradford C. B.,1998)。在我們的實驗中發現,DOX會刺激心肌細胞表現 ET-1,並且是經由ROS相關的訊息傳遞路徑活化ET-1的轉錄,因為 這個現象受到catalase的抑制,而catalase具有清除H2O2的功能。 雖然在許多病人的臨床報告和老鼠的活體實驗的結果都指出,DOX 會 引起血漿中 ET-1 量的持續上升,且和心臟衰竭有正相關性,因此 ET-1 具
有當作DOX 引起心臟損傷的指標的潛力,但是要實際使用 ET-1 當作 DOX
引起心臟損傷的指標,需要在機制上有更多的了解,以及在實驗上有更多
數據支持。在我們的實驗中發現 DOX 的確會刺激心臟細胞的 ET-1 分泌增
加,是經由刺激非心肌細胞 ET-1 的分泌以及刺激心肌細胞 ET-1 的表現及
負責較晚期的 ET-1 分泌(12~24 小時)。並且 DOX 是經由引起 ROS 有關的
機制而調控心肌細胞ET-1 表現量的增加,但是在非心肌細胞中是經由其他
*
-10 0 10 20 30 40 1 3 6 12 24 (hr) ET-1 secretion (fmol/ml/1x10 6 cells) DOX 1µM Control □ Control ■ DOX 1μM圖1. DOX 會刺激心肌細胞 ET-1 的分泌。從初生 1-2 天的 Wistar rat 分離培
養出來的心肌細胞,經由 DMEM- BrdU 培養 2 天後,心肌細胞開始跳動,
換上sreum-free DMEM 培養 24 小時後,以 DOX 1µM 處理細胞 1、3、6、
12、24 小時,之後收集培養液,用 Endothelin-1 Biotrak Assay 測 ET-1 的量。 12 小時 DOX 處理的心肌細胞之 ET-1 分泌量是 14.7 fmol/ml,對照組沒有
測到ET-1 分泌。24 小時 DOX 處理的心肌細胞之 ET-1 分泌量是 8.7fmol/ml,
對照組是6.9 fmol/ml。(mean±S.T.D.E.V.;*P<0.05 v.s.沒有處理 DOX 的心肌
(μM)
0
10
20
30
40
C
0.01
0.1
1
10
E
T
-1
s
ecr
et
io
n
(f
m
ol
/m
l/
1x10
6cel
ls
)
# Dxr Control □ Control ■ DOX*
*
圖2.不同劑量的 DOX 刺激心肌細胞 ET-1 之分泌。從初生 1-2 天的 Wistar rat
分離培養出來的心肌細胞,經由DMEM-BrdU 培養 2 天後,心肌細胞開始
跳動,換上 sreum-free DMEM 培養 24 小時後,以 0.01、0.1、1、10µM 的
DOX 處理細胞 24 小時之後,收集培養液,用 Endothelin-1 Biotrak Assay 測 ET-1 的量。發現 1µM、10µM 的 DOX 明顯引起心肌細胞的 ET-1 分泌,但
是0.1µM 的 DOX 會抑制 ET-1 的分泌。0.1µM DOX、1µM DOX 和 10µM DOX
處理的心肌細胞之ET-1 分泌量分別是 5.24 fmol/ml、12.3 fmol/ml 和 11
fmol/ml,對照組是 8.3 fmol/ml,沒有 dose-dependent 的現象。(mean± S.T.D.E.V.;*P<0.05 v.s.沒有處理 DOX 的心肌細胞; # P<0.05 v.s.沒有處理 DOX 的心肌細胞; n=5)。
# 0 10 20 30 40 1 3 6 12 24 (hr) ET-1 secretion (fmol/ml/1. 5x10 6 cells)
*
*
*
DOX 1µM Control □ Control ■ DOX 1μM圖3. DOX刺激非心肌細胞的ET-1 分泌迅速增加。從初生 1-2 天的Wistar rat
分離培養出來的非心肌細胞,經由DMEM培養 2 代後,每培養皿種 1.5x106
細胞,以DMEM培養 1 天,接著換上sreum-free DMEM 24 小時後,以 DOX 1µM處理細胞 1、3、6、12、24 小時,收集培養液,用Endothelin-1 Biotrak Assay測ET-1 的量。1 小時DOX處理的非心肌細胞之ET-1 分泌量是 11.2 fmol/ml,對照組是 3.9 fmol/ml。3 小時DOX處理的非心肌細胞之ET-1 分泌
量是17.5 fmol/ml,對照組是 3 fmol/ml。6 小時DOX處理的非心肌細胞之ET-1
分泌量是30 fmol/ml,對照組是 11.4 fmol/ml。(mean± S.T.D.E.V. ;*P<0.05 v.s.
沒有處理DOX的非心肌細胞; # P<0.05 v.s.沒有處理DOX的非心肌細胞; n=6)。
圖4. DOX刺激非心肌細胞ET-1 的分泌。從初生 1-2 天的Wistar rat分離培養
出來的非心肌細胞,經由DMEM培養 2 代後,每培養皿種 1.5x106細胞,以
DMEM培養 1 天,接著換上sreum-free DMEM 24 小時後,以 0.01、0.1、1、 10µM的DOX處理細胞 24 小時,收集培養液,用Endothelin-1 Biotrak Assay 測ET-1 的量。1µM 和 10µM DOX處理的非心肌細胞之ET-1 分泌量是 15.5 和 25.3 fmol/ml,對照組是 19 fmol/ml,沒有dose-dependent的現象。 (mean±S.T.D.E.V.;*P<0.05 v.s.沒有處理DOX的非心肌細胞; # P<0.05 v.s.沒 有處理DOX的非心肌細胞;n=6)。 (μM)
*
0
10
20
30
40
C
0.01
0.1
1
10
1 s
et
(
fm
ol/m
l/1
.5
x1
0
6c
ells
)
DxrControl*
□ Control ■ DOXio
n
#ecr
E
T
--10 0 10 20 30 1 3 6 12 24 (hr)
ET-1 secretion (nmol/ml/10
cell)
*
*
M,Dxr 1µM M,Control non-M,Dxr 1µM non-M,Control 6 圖5. DOX 引發心肌細胞與非心肌細胞中不同時間模式的 ET-1 分泌。從初生1-2 天的 Wistar rat 分離培養出來的心肌細胞與非心肌細胞,以 DOX 1µM
處理細胞,1、3、6、12、24 小時之後,收集培養液,用 Endothelin-1 Biotrak
Assay 測 ET-1 的量,得到每細胞分泌 ET-1 量。DOX 處理的非心肌細胞在 6 小時 ET-1 分泌量最多。心肌細胞在 12 小時 ET-1 分泌量最多, 綜觀結果 ET-1 的分泌量以非心肌細胞較多。(mean+S.T.D.E.V.;*P<0.05 v.s.沒有處理 DOX 的心肌細胞; n=5;*P<0.05 v.s.沒有處理 DOX 的非心肌細胞; n=6)。
A DOX 1μM
Control
1 3 6 12(hr)
1 3 6 12 24(hr)
ET-1
GAPDH
#*
*
0 25 50 75 100 1 3 6 12 24 (hr) ET-1 m R N A Le ve (% o f GAP D H) Control DOX 1µM B圖6. DOX刺激心肌細胞ET-1 基因的表現。從初生 1-2 天的Wistar rat分離培
養出來的心肌細胞,經由DMEM-BrdU培養 2 天後,心肌細胞開始跳動,換 上sreum-free DMEM 24 小時後,以DOX 1µM處理細胞 1、3、6、12、24 小 時,收集細胞。(A)RT-PCR結果,DOX處理的心肌細胞之ET-1 在 1 小時就 有表現,而在 3 小時則有高量的表現,24 小時心肌細胞因為死亡很多(細胞 數≦5 x 106 ),所以無法得到足量的total RNA。對照組的心肌細胞之ET-1 表現隨時間增加,表現也會逐漸增加。(B) ET-1 mRNA表現量的數值。(mean± S.T.D.E.V.; *P<0.05 v.s.沒有處理DOX的心肌細胞; #P<0.05 v.s.沒有處理 DOX的心肌細胞; n=5)。
DOX 1μM Control A 1 3
6 12 24(hr) 1 3 6 12 24(hr) ET-1 GAPDH B
*
# # 0 25 50 75 100 125 1 3 6 12 24 (hr) E T -1 m R N A L eve l (% o f GAP DH) Control DOX 1µM圖7. DOX短暫抑制非心肌細胞ET-1 基因的表現。從初生 1-2 天的Wistar rat
分離培養出來的非心肌細胞,經由DMEM培養 2 代後,每培養皿種 1.5x106
細胞,換上sreum-free DMEM 24 小時後,以DOX 1µM處理細胞 1、3、6、 12、24 小時,收集細胞。(A)RT-PCR結果,DOX處理的非心肌細胞之ET-1
在 1~3 小時間的表現受抑制,而在 6 小時的表現則有些增加。對照組的非
心肌細胞之ET-1 平時就有穩定表現。(B) ET-1 mRNA表現量的數值。 (mean±S.T.D.E.V.;*P<0.05 v.s.沒有處理DOX的非心肌細胞;#P<0.05 v.s.沒 有處理DOX的非心肌細胞; n=6)。
DOX 1μM Control A 24 27 30 36 48(hr) 24 27 30 36 48(hr) ET-1 GAPDH 0 25 50 75 100 24 27 30 36 48 (hr) ET-1 m R N A Le ve (% o f GAP DH) Control DOX 1µM B 圖8. DOX不影響 24 小時後非心肌細胞ET-1 基因的表現。從初生 1-2 天的 Wistar rat分離培養出來的非心肌細胞,經由DMEM培養 2 代後,每培養皿
種1.5x106細胞,換上sreum-free DMEM 24 小時後,以DOX 1µM處理細胞,
24、27、30、36、48 小時,收集細胞。(A)做RT-PCR,DOX處理的非心肌 細胞之ET-1 在 24~48 小時期間表現穩定不變。對照組亦同。(B) ET-1 mRNA 數值,DOX處理的非心肌細胞之ET-1 在 24~48 小時間表現無明顯差異。 (mean±S.T.D.E.V.; n=6)。
A
*
*
# 0 25 50 75 100 1 3 6 12 24 (hr) ET-1 m R N A Le ve (% o f GAP D H) Control DOX 1µM B -10 0 10 20 30 40 1 3 6 12 24 (hr) E T -1 s ecr et io n ( fm ol /m l/ 1x10 6 cel ls ) □ Control ■ DOX 1μM*
圖9. DOX 刺激心肌細胞 ET-1 的表現有延遲的現象。從初生 1-2 天的 Wistar
rat 分離培養出來的心肌細胞,經由 DMEM-BrdU 培養 2 天後,心肌細胞開
始跳動,換上sreum-free DMEM 24 小時後,以 DOX 1µM 處理細胞,1、3、
6、12、24 小時,收集細胞與培養液。(A)RT-PCR 的結果顯示心肌細胞的 ET-1 表現在 3 小時達到最高量。(B)Endothelin-1 Biotrak Assay 的結果顯示
心肌細胞的ET-1 分泌在 12 小時達到最高量。表示 ET-1 基因的表現到分泌
中間有一段時間差。(mean±S.T.D.E.V.;*P<0.05 v.s.沒有處理 DOX 的心肌細
A # 0 10 20 30 40 1 3 6 12 24 (hr) E T -1 s ec re tio n (f m ol /m l/1.5x10 6 c ells )
*
*
# # □ Control ■ DOX 1μM 0 25 50 75 100 125 1 3 6 12 24 (hr) E T -1 m R N A L ev el (% o f G A PD H) Control DOX 1µM B 圖10. DOX刺激非心肌細胞的ET-1 表現受到其他機轉的調控。從初生 1-2 天的Wistar rat分離培養出來的非心肌細胞,經由DMEM培養 2 代後,每培養皿種1.5x106細胞,換上sreum-free DMEM 24 小時後,以DOX 1µM處理
細胞1、3、6、12、24 小時,收集細胞與培養液。(A)RT-PCR的結果顯示
非心肌細胞之ET-1 在 1~3 小時間的表現受抑制,而在 6 小時的表現則有些
增加。(B) Endothelin-1 Biotrak Assay的結果顯示非心肌細胞之ET-1 分泌在 6
小時達到最高量,之後卻逐漸減少。表示非心肌細胞的ET-1 表現受到其他 機轉的調控。(mean±S.T.D.E.V.;*P<0.05 v.s.沒有處理DOX的非心肌細胞; #P<0.05 v.s.沒有處理DOX的非心肌細胞;n=6)。
0 50 100 150 200 C 1 3 6 12 24 (hr) R el at iv e f lu or escen ce pe rcen tig e (% of c ont ro l, 1x10 6 cel ls )
*
□ Control ■ DOX 1μM*
*
*
圖11. DOX處理使心肌細胞中的ROS量增加。從初生1-2天的Wistar rat 分離培養出來的心肌細胞,經由DMEM-BrdU培養2天後,心肌細胞開始跳 動,換上sreum-free DMEM 24小時後,以DOX 1µM處理細胞,1、3、6、 12、24小時,收集細胞。離心去掉上清液後以1X PBS重新懸浮細胞,加入 nonpolar 2’,7’-DCFH-DA (nonpolar 2’,7’- dichloro- fluorescein diacetate),避 光下,在37℃、震盪反應30分鐘,離心去掉上清液,沉澱的細胞重新以1XPBS 懸浮,測525/475nm螢光光譜。DOX處理心肌細胞 1, 3, 6, 12, 24小時後與對 照組相比ROS增加132%、152%、135%、137%、121%。 (mean±S.T.D.E.V.; *P<0.05 v.s.沒有處理DOX的心肌細胞;n=3)。-1.7kCAT
DOX 1μM
-1.7kCAT -1.7kCAT CAT2 CAT3 + catalase -1.7kCAT + catalase 圖 12. ROS 參 與 心 肌 細 胞 轉 錄 的 活 化 。 心 肌 細 胞 分 別 轉 染 -1.7kCAT
plasmid、pBLCAT2 plasmid及pBLCAT3 plasmid,轉染的細胞培養 48 小時 後,以catalase(350U/ml)先處理 3 小時,再以DOX處理 6 小時,接著收集細
胞。 取同蛋白質量的樣本,加入 [14C]-chloramphenicol和 acetyl CoA作用,
之後加入ethyl acetate終止反應。最後進行TLC。經由X光底片壓片後的結果 發現,DOX會活化心肌細胞的ET-1 promotor,且這個作用會受到catalase的 抑制。
圖13. 人類冠狀動脈的ET-1運輸模式。在這個模式中提出2個不同途徑,使 得ET-1被運輸到細胞表面。ET-1與其他作用於血管的物質儲存在Weibel- Palade bodies中,經由適當的刺激後被釋放到細胞表面。ET-1 也儲存在分 泌的囊泡中,會持續性的釋放,經由cyclic AMP有關的持續性訊息途徑作 用。(Russell et al., 1999)
A
B
圖14.細胞中的ROS (A)細胞內ROS的產生。(B)因外來刺激形成的ROS,在 細胞中參與訊息傳遞路徑,以活化下游基因的表現。(Jun-ichi, A. and Bradford C. B.,1998)
伍、參考文獻:
Arai, M., Yoguchi, A., Takizawa, T., Yokoyama, T., Kanda, T., Kurabayashi, M. and Nagai, R. (2000) Mechanism of doxorubicin-induced inhibition of sarcoplasmic reticulum Ca2q-ATPase gene transcription.Circ. Res. 86, 8-14. Araki, M., Hasegawa, K., Iwai-Kanai, E., Fujita, M., Sawamura, T., Kakita, T., Wada, H., Morimoto, T. and Sasayama, S.(2000) Endothelin-1 as a protective factor against beta-adrenergic agonist-induced apoptosis in cardiac myocytes. J Am Coll Cardiol. 36,1411-8.
Bertheau, P., Plassa, F., Espie, M., Turpin, E., de Roquancourt, A., Marty, M., Lerebours, F., Beuzard, Y., Janin, A. and de The, H. (2002) Effect of mutated TP53 on response of advanced breast cancers to high-dose chemotherapy. Lancet 360,852-854.
Bezombes, C., de Thonel, A., Apostolou, A., Louat, T., Jaffrezou, J.P., Laurent, G. and Quillet-Mary, A. (2002) Overexpression of protein kinase Czeta confers protection against antileukemic drugs by inhibiting the redox- dependent sphingomyelinase activation. Mol Pharmacol 62,1446-1455.
Binaschi, M., Bigioni, M., Cipollone, A., Rossi, C., Goso, C., Maggi, C.A., Capranico, G. and Animati, F. (2001) Anthracyclines: selected new developments. Curr Med Chem1,113-130.
Binaschi, M., Capranico, G., Dal Bo, L., and Zunino, F. (1997) Relationship between lethal effects and topoisomerase II mediated double-strand DNA breaks produced by anthracyclines with different sequence specificity. Mol Pharmacol 51,1053-1059.
II-poisoning anthracyclines.Cancer Res 60,3770-3776.
Boucek Jr., R.J., Buck, S.H., Scott, F., Oquist, N.L., Fleischer, S.and Olson, R.D. (1993) Anthracycline-induced tension in permeabilized cardiac fibers: evidence for the activation of the calcium release channel of sarcoplasmic reticulum. J. Mol. Cell. Cardiol. 25, 249-259.
Boucek Jr., R.J., Dodd, D.A., Atkinson, J.B., Oquist, N. and Olson, R.D.( 1997) Contractile failure in chronic doxorubicin-induced cardiomyopathy. J. Mol. Cell. Cardiol. 29, 2631-2640.
Boucek Jr., R.J., Olson, R.D., Brenner, D.E., Ogunbunmi, E.M., Inui, M. and Fleischer, S. (1987) The major metabolite of doxorubicin is a potent inhibitor of membrane-associated ion pumps. A correlative study of cardiac muscle with isolated membrane fractions. J. Biol. Chem. 262,15851-15856.
Caroni, P., Villani, F. and Carafoli, E. (1981) The cardiotoxic antibiotic doxorubicin inhibits the NaqrCa2q exchange of dog heart sarcolemmal vesicles. FEBS Lett. 130, 184-186.
Chao, H.H., Chen, J.J., Chen, C.H., Lin, H., Cheng, C.F., Lian, W.S., Chen, Y.L., Juan, S.H., Liu, J.C., Liou, J.Y., Chan, P. and Cheng, T.H. (2005) Inhibition of angiotensin II induced endothelin-1 gene expression by 17-beta-oestradiol in rat cardiac fibroblasts. Heart. 91,664-9.
Cheng, C.M., Hong, H.J., Liu, J.C., Shih, N.L., Juan, S.H., Loh, S.H., Chan, P., Chen, J.J. and Cheng, T.H. (2003a) Crucial role of extracellular signal- regulated kinase pathway in reactive oxygen species-mediated endothelin-1 gene expression induced by endothelin-1 in rat cardiac fibroblasts. Mol Pharmacol. 63,1002-11.
Cheng, T.H., Cheng, P.Y., Shih, N.L., Chen, I.B., Wang, D.L. and Chen, J.J. (2003b) Involvement of reactive oxygen species in angiotensin II-induced