行政院國家科學委員會專題研究計畫 成果報告
CXCR4 之表現對基底細胞癌腫瘤增生以及基因調控影響之
研究
計畫類別: 個別型計畫
計畫編號: NSC93-2314-B-039-007-
執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日
執行單位: 中國醫藥大學醫學系
計畫主持人: 吳孟澤
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 94 年 10 月 31 日
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※
※
※
CXCR4 之表現對基底細胞癌腫瘤增生以及基因調控影響之研究
※
※
※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:▇個別型計畫
□整合型計畫
計畫編號:NSC 93-2314-B-039-007
執行期間:
93 年
8 月
1 日至 94 年
7 月
31 日
計畫主持人:吳孟澤
共同主持人:無
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:中國醫藥大學醫學系皮膚學科
中
華
民
國
94
年
10 月
日
一. 中英文摘要
趨化激素及其受體較為人所熟悉的角色為調控白血球之移行, 然而 研究顯示它們在細胞分裂,凋亡, 生存以及血管增生各方面的調控 也扮演了重要的角色, 因此可能也有助於癌細胞的生長與轉移.在 九十二年度的國科會計劃(92-2314-B-039-024)中,我們探討趨化激 素及其受體在基底細胞癌腫瘤生長病程中所可能扮演的角色.我們 使用定量RT-PCR 測定各種趨化激素受體於基底細胞癌之表現, 發 現CXCR4 於培養基底細胞癌細胞中有高度之表現. 進一步檢驗 CXCR4 於其它皮膚正常細胞(角質細胞與黑色素細胞)以及培養皮 膚癌細胞(鱗狀細胞癌以及黑色素細胞癌)之表現, 發現CXCR4 於 這些細胞中並無顯著表現.我們也利用傳統RT-PCR, western blotting 方法進一步驗證CXCR4 於培養基底細胞癌細胞中之表現. 我們也 利 用 Immunohistochemistry 以 及 insitu hybridization 方 法 證 實 了 CXCR4 於基底細胞癌組織中的表現.顯示CXCR4 於基底細胞癌細 胞進犯過程中可能扮演某種角色.為了進一步確定CXCR4 之表現是 否 在 基 底 細 胞 癌 腫 瘤 增 生 過 程 中 扮 演 重 要 之 角 色 , 我 們 利 用 retroviral transduction 技術將CXCR4 cDNA 基因轉入基底細胞癌細 胞, 而得到過度表現CXCR4 之基底細胞癌細胞株(CXCR4-BCC). 於細胞增生實驗中, 培養於低濃度血清中的CXCR4-BCC 細胞與 CXCL12 (CXCR4 之ligand) 反應後, 其細胞數目與對照組相較有明 顯之增加, 若進一步加入anti-CXCR4 mAb 抗體則可中和此反應. 於細胞凋亡實驗中, 我們以紫外線B來引發CXCR4-BCC 之凋亡, 並發現CXCR4-BCC 與CXCL12 反應後其細胞凋亡與對照組相較 有明顯之減少. 若進一步加入拮抗CXCR4 之blocking peptide T22 則 可 抵 消 CXCL12 所 帶 來 之 減 少 凋 亡 之 作 用 . 這 些 發 現 顯 示 CXCR4/CXCL12 之反應可以於體外實驗中增加基底細胞癌細胞細 胞增生以及保護其免於凋亡.以上之成果顯示CXCR4 之表現對於基 底細胞癌腫瘤增生過程可能扮演了重要之角色. 於此我們提出本計 劃以進一步探索此一主題. 我們計劃以以下三個方向來進行. 第 一, 我們將繼續進行系列之體外實驗(如細胞移行實驗, 腫瘤侵犯實 驗, 血管新生實驗)以探討CXCR4 之過度表現是否會對基底細胞癌 之行為表現造成影響. 第二, 我們擬以裸鼠進行異種腫瘤移殖之動 物實驗, 將CXCR4-BCC 注射於老數皮下, 以了解CXCR4之過度表 現是否會增進動物體內之基底細胞癌腫瘤增生. 最後, 由於確定了 CXCR4/CXCL12 之相互反應對於基底細胞癌腫瘤增生會有顯著影 響, 我們將進一步探索其所產生的訊息傳導對下游基因之調控現象. 我 們 擬 以 siRNA 技 術 干 擾 CXCR4 於 基 底 細 胞 癌 之 基 因 表 現 . CXCR4-BCC 以及缺少CXCR4 表現之基底細胞癌與CXCL12 反應 後 將 進 行 cDNA 微 陣 列 分 析 (microarray analyses), 以 了 解 CXCR4/CXCL12 之相互反應對下游基因之調控情形. 我們預期本 研究將能進一部釐清CXCR4 之表現對於基底細胞癌腫瘤增生之生 物意義與重要性.Beside the well-known role to regulate leukocyte trafficking, chemokines and their receptors alsoplay important roles in the regulation of mitosis, apoptosis, survival and angiogenesis, which mayenhance tumor growth or metastasis. In our previous NSC project (92-2314-B-039-024), we investigated the possible involvements of chemokines and chemokine
receptors in thecarcinogenesis of basal cell carcinoma (BCC). By real-time RT-PCR, the expression CXCR4 inBCC cell line was
~64-fold more than in other cancer cell lines and normal controls (cultured humankeratinocytes and fibroblasts). The expression of CXCR4 in BCC cell line was further confirmedusing conventional RT-PCR and western blotting. Immunohistochemistry and in situ
hybridization studies using tissue samples from BCC lesions showed
clear CXCR4 staining as well.To further determine whether CXCR4 expression may play an important role in BCC tumorigenesis,retroviral
transduction was employed to transfer CXCR4 cDNA into BCC cell
line. CXCR4expression by BCC was thus significantly enhanced in CXC4-transduced BCC cells (CXCR4-BCC).We next address whether overexpression of CXCR4 may alter BCC tumor progression in vitro. In
cell proliferation assay, CXCR4-BCC cells were cultured under low
serum concentration (0.5%).CXCL12 (ligand for CXCR4) treatments resulted in significant increase of CXCR4-BCC proliferation compared to PBS treatments, which could be completely neutralized by co-treatments with anti-CXCR4 mAb. To determine whether CXCR4
signaling may render apoptosisresistance for BCC cells, UVB
irradiation was used to induce apoptosis in CXCR4-BCC cells. With CXCL12 pre-treatment, there was ~38% decrease of apoptosis in CXCR4-BCC cells. This protective effect could be negated by
that CXCL12/CXCR4 interactions enhance the capacities of BCC cells for proliferation andapoptosis-resistance.The above results
indicate that CXCR4 expression may play a critical role in BCC tumorigenesis.Herein we propose to further investigate this subject
with the following approaches. Firstly, weseek to address whether
CXCR4 expression may also affect other essential aspects of BCCtumorigenesis (migration, invasion and angiogenesis) in vitro. Secondly, whether CXCR4 overexpression may specifically enhance BCC tumor progression in vivo also needs to be addressed. We plan to set up a xenograft tumor transplant model using immuno-compromised nude mice. Thirdly, given that CXCR4/CXCL12 interactions play an important role in BCC tumorigenesis, it would be interesting and important to investigate what are the exact downstream molecular targets of CXCR4/CXCL12 signaling in BCC cells. We propose to use the newly developed RNA interference technique (small interfering RNA, or
siRNA) to knock down the CXCR4 expression in BCC cell lines.
CXCR4-BCC and CXCR4-deficient BCC cells (generated by siRNA expression vector transfection) will then be exposed to CXCL12 and subject to cDNA microarray analyses to obtain the differentially expressed genes in these two cell lines. In this way, we expect to further determine the biological significances of CXCR4 expression, especially for BCC tumorigenesis.
二、緣由與目的
Chemokines and their receptors are well known for their ability to regulate the directional migration of leukocytes, and thus broadly involved in normal developmental processes like organogenesis and host homeostasis like immune surveillance, as well as pathological conditions like infection or inflammation (reviewed by (Rossi and Zlotnik 2000) and (Baggiolini 2001)). Furthermore, emerging evidences also show that chemokines/chemokine receptors may play important roles in several essential aspects of tumorigenesis, including tumor growth, angiogenesis, invasion and metastasis (review by(Vicari and Caux 2002)).
CXC chemokine receptor-4 (CXCR4) is one of those chemokines having been implicated in tumor progression recently. Müller et al (Muller, et al 2001) demonstrated that CXCR4 was highly expressed in breast cancer cells and chemokine CXCL12 (the only cognate ligand for CXCR4, also known as stromal-derived factor-1/SDF-1) had peak levels of expression in organs known as common metastatic sites for breast cancer. Moreover, in vivo neutralization of CXCR4 by mAb significantly impaired metastasis of breast cancers to regional lymph nodes and lung, indicating CXCR4’s involvement in selective metastases of breast cancer to certain organs. In an attempt to further clarify CXCR4’srolein cancermetastasis,Murakamietal(Murakami,
et al 2002) demonstrated thatCXCR4’sexpression in vivo selectively
enhances the metastatic potential of melanoma cells to lung, but not to other organs; and further showed that CXCR4 might act by enhancing tumor adhesion to endothelial cells and tumor growth under stress. Moreover, CXCR4 were also found expressed in other types of tumors and may be involved in tumor progression(Koshiba, et al 2000, Staller,
et al 2003, Taichman, et al 2002).
Cutaneous basal cell carcinoma (BCC) is the most common human cancer and its incidence is continuously increasing(Gloster and Brodland 1996, Miller and Weinstock 1994). Typical lesions of BCC feature local skin invasion and angiogenesis, whereas the metastasis is rare. Although several factors may account for the pathogenesis of BCC, including ultraviolet light exposure(Gailani, et al 1996) and mutations in Hedgehog genes(Oro, et al 1997, Unden, et al 1997), the molecular mechanisms for BCC tumor progression are still limited.
We hence hypothesized that certain chemokine receptor(s) may be involved in BCC tumor progression. In our previous work (supported by NSC 91-2314-B-037-299 and NSC 92-2314-B-039-024), we have tried to address whether CXCR4 may play a critical role in BCC tumorigenesis and obtained some interesting results as follows:
CXCR4 is expressed by BCC cells. To address whether chemokine
receptors may play a role in the tumorigenesis of BCC, we extensively examined the expression levels of CC and CXC chemokine receptors (CCR1CCR10 and CXCR1CXCR6) in a human BCC cell line (BCC-1/KMC)(Chiang, et al 1994) using real-time quantitative PCR (qPCR). CXCR4 was predominantly expressed by BCC-1/KMC cells compared with other chemokine receptors (~72-fold higher than CCR10, the second highest expressed). We next determined the relative
expression levels of CXCR4 in BCC-1/KMC, other skin cancer cell lines (melanoma and squamous cell carcinoma) and normal cells from human skin (keratinocytes, melanocytes and fibroblasts), and observed similarly highly expressed CXCR4 in BCC-1/KMC in comparison with other kinds of normal and transformed cells from skin (~1000- to 3000-fold higher expression than other cells). The expression of CXCR4 in BCC-1/KMC cells was further confirmed at protein level by immunoblotting.
We next determined whether CXCR4 is expressed in BCC tissues. BCC samples were subject to immunohistochemical studies. Anti-CXCR4 mAb staining was found positive in a subset of BCC samples. In situ hybridization studies using CXCR4 anti-sense oligonucleotide probes further showed positive staining of CXCR4 mRNA in BCC tumor cells.
Functional Transduction of BCC Cells with CXCR4. To determine
whether CXCR4 alone may play important roles in BCC tumorigenesis, we used retroviral vector (pLNCX2, from Clontech, Palo Alto) to transfer CXCR4 gene into BCC-1/KMC cells, which were then subject to magnet-bead selection to enhance the CXCR4 expression as described(Murakami, et al 2002). Magnet-bead positive selection was further used to increase CXCR4 expression, as ~91% CXCR4-BCC showed clear staining with anti-CXCR4 mAb by flow cytometry. Measured by qPCR, CXCR4 expression was significantly enhanced in CXCR4-BCC compared with parental BCC-1/KMC (~37-fold) and pLNCX2-transduced
共 BCC cells (pLNCX2-BCC, serving as vector-control) (~14-fold). Calcium flux assay was further used to confirm that CXCR4 receptors were functional in CXCR4-BCC.
CXCL12/CXCR4 Enhance CXCR4-BCC Proliferation by CXCR4 in vitro. To address whether expression of CXCR4 may enhance BCC
tumor growth, pLNCX2-BCC and CXCR4-BCC cells were cultured with normal serum (10% FCS) or low serum (0.5% FCS) in the presence or absence of CXCL12. CXCR4-BCC and pLNCX2-BCC cells showed similar proliferation rates in normal serum, either with or without CXCL12. By contrast, CXCL12 treatments resulted in significant increase of CXCR4-BCC cell proliferation compared with PBS treatments (24-hr, ~1.2-fold p=0.027; 48-hr, ~2.2-fold, p=0.004). The enhanced growth with the treatments of CXCL12 could be neutralized by anti-CXCR4 (300 ug/ml) mAb, but not by the isotype control mouse IgG2b. CXCL12 did not significantly enhance the proliferation of pLNCX2-BCC cells in low serum (p=0.4, data not shown). Thus, CXCL12 may enhance BCC tumor growth by CXCR4 expression under stressful condition like low serum.
CXCL12/CXCR4 Interactions Protect CXCR4-BCC from Apoptosis by UVB Irradiation. To determine whether CXCR4 signaling may provide
apoptosis resistance for BCC cells, UVB irradiation was used to induce apoptosis in pLNCX2-BCC and CXCR4-BCC cells. With CXCL12 pre-treatment, there was ~38% (p=0.04) decrease of apoptosis in CXCR4-BCC cells (using caspase-3 as marker). This protective effect could be negated by co-treatment with CXCR4-blocking T22 peptide but not control peptide ALA. The presence of CXCL12 and T22 did not significantly affect the UVB-induced apoptosis in pLNCX2-BCC cells. Thus, CXCL12/CXCR4 pathway might provide BCC cells with significant resistance to apoptosis.
The above results indicate that CXCR4 may play a critical role in BCC tumorigenesis. Herein we propose to further investigate this subject with the following approaches. Firstly, to consolidate these results as above, we seek to address whether CXCR4 expression may also affect other essential aspects of BCC tumorigenesis (migration, invasion and angiogenesis) in vitro. We plan to perform angiogenesis assays to test CXCR4-BCC and pLNCX2-BCC cells. Secondly, whether CXCR4
overexpression may specifically enhance BCC tumor progression in vivo also needs to be addressed. In this regard, we plan to set up a xenograft tumor transplant model using immuno-compromised nude mice. CXCR4-BCC and pLNCX2-BCC cells will be subcutaneously injected into nude mice. Tumor growth will then be assessed. Thirdly, given that CXCR4/CXCL12 interactions play an important role in BCC tumorigenesis, it would be interesting and important to investigate what are the exact downstream targets of CXCR4/CXCL12 signaling in BCC cells. In other words, we want to know what molecules would be turn on or turn down by the CXCR4/CXCL12 axis. We propose to use the newly developed RNA interference technique (small interfering RNA, or siRNA) to knock down the CXCR4 expression in BCC-1/KMC cells. CXCR4-BCC and CXCR4-deficient BCC cells (generated by siRNA
expression vector transduction) will then be exposed to CXCL12 and subject to cDNA microarray analyses to render the differentially expressed genes in these two cell lines. In this way, we expect to further determine the biological significances of CXCR4 expression, especially for BCC tumorigenesis. More importantly, the discovery of CXCR4 and CXCR4-regulated genes being actively involved herein may lead to establishment of novel therapeutic strategies to fight against cancer in general. The results of the proposed experimental are detailed as follows.
三、結果與討論
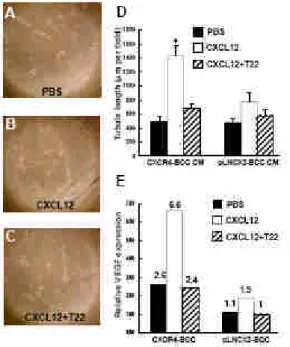
CXCL12/CXCR4 pathway enhances angiogenesis of CXCR4-BCC in vitro To determine whether CXCR4 expression may enhance tumor
angiogenesis, concentrated conditioned media from pLNCX2-BCC or CXCR4-BCC cells were used to stimulate the growth of human endothelial cells in vitro, as described in Materials and methods. With CXCL12 treatments, conditioned media from CXCR4-BCC cells significantly enhanced tubule formation of human microvascular endothelial cells, HMEC-1 (Fig. 1B and D), compared with control (no CXCL12 treatment) conditioned medium incubation (Fig. 1A and D; ~2.19-fold, P<0.05). Co-treatment with T22 peptide led to the reverse of enhanced angiogenesis, as shown in Fig. 1C and D. In contrast, HMEC-1 cells incubated with conditioned media from CXCL12-treated pLNCX2-BCC did not show significantly enhanced tubule formation (Fig. 1D, P=0.086) Thus, CXCR4/CXCL12 interactions in CXCR4-BCC cells may enhance tumor angiogenesis by production of soluble mediators. In order to determine whether CXCR4/CXCL12 signaling axis may promote the expression of certain angiogenic factors, pLNCX2-BCC and CXCR4-BCC cells, with CXCL12 pre-treatments, were subject to qPCR analysis for the expression of vascular endothelial growth factor (VEGF). As shown in Fig. 1E, 24 h after the exposure to CXCL12, VEGF expression was up-regulated in both CXCR4-BCC and pLNCX2-BCC cells, with a less degree in the latter (CXCR4-BCC, ~2.5-fold; pLNCX2-BCC, ~1.7-fold). CXCL12 treatments did not enhance expression of other angiogenic factors (VEGF-B, VEGF-C, VEGF-D and cycloxygenase-2, data not shown). Furthermore, the up-regulation of VEGF could be reversed by the addition of T22 peptide (Fig. 1E), indicating that VEGF expression may be specifically dependent on CXCR4R/CXCL12 signaling axis in BCC cells.
Figure 1. CXCR4 expression enhances BCC angiogenesis in vitro. (A-C) HMEC-1 cells were cultured in matrigel-coated wells and then exposed to conditioned media collected from CXCR4-BCC and pLNCX2-BCC cells in various conditions as indicated. Representative pictures showing tubule formation of HMEC-1 cells, from three independent experiments with similar results, 100. HMEC-1 cells in conditioned medium from CXCR4-BCC with addition of PBS only (A), in conditioned medium from CXCR4-BCC
with previous CXCL12 treatment (B), and in conditioned medium from CXCR4-BCC with CXCL12 and T22 treatment (C). (D) Tubule formation of HMEC-1 cells, as shown in (A-C), was quantified after incubation with various conditioned media from CXCR4-BCC and pLNCX2-BCC cells for 8 h; CM: condition media; *, P < 0.05, versus HMEC-1 cells incubated with control (PBS only) conditioned medium. (E) qPCR expression analysis of VEGF by CXCR4-BCC and pLNCX2-BCC cells, in the presence of CXCL12 or not. T22 was added in some experiments as indicated.
CXCR4 transduction increase BCC tumor growth in vivoTo determine
whether CXCR4 expression may enhance tumorigenesis of BCC in vivo, BCC-1/KMC, pLNCX2-BCC, and CXCR4-BCC cells were injected s.c. into nude mice. At the end-point of observation (day 31), the injections of CXCR4-BCC cells resulted in larger tumor burden (volume) in nude mice, compared with pLNXC2-BCC cell injection (~3.6-fold, P< 0.05), as shown in Fig. 2A. Gross examination of inoculated CXCR4-BCC tumors revealed marked angiogenesis (Fig. 2B) and ulceration (Fig. 2C), mimicking typical human BCC tumors. Furthermore, tumor growth of CXCR4-BCC could be suppressed by serial injections of specific CXCR4-blocking peptide, T22 (Fig. 2A). Thorough examination of internal organs (lung, liver and gastrointestinal tract) and draining lymph nodes were performed in tumor-bearing mice, and revealed no gross metastasis. Taken together, these results indicate that CXCR4 expression may enhance tumor growth and angiogenesis of BCC in vivo.
Figure 2. CXCR4 expression enhances BCC tumorigenesis in vivo. Parental BCC-1/KMC, pLNCX2-BCC and CXCR4-BCC cells (with T22 treatments or not) were injected (5105
cells in 100 l PBS) subcutaneously into the flanks of nude mice respectively. (A) BCC tumor burden from day 11 to day 31 (n=5). Growth of CXCR4-BCC tumor (without T22 treatment) was significantly increased (P<0.05) compared with other groups. (B-C) Representative pictures (at day 31) showing CXCR4-BCC tumor progression mimicking human BCC, with angiogenesis in (B) and ulceration in (C); scale bar (B-C): 5 mm.
Microarray analysis of CXCR4 signaling in CXCR4-BCC To further
investigate the effect of CXCR4 signaling for BCC, CXCR4-BCC with CXCL12 treatment was subject to cDNA microarray comparison versus CXCR4-BCC with PBS treatment only. Up- and down-regulated genes are listed herein as follows.
Fig. 3 up-regulated genes
Fig. 4 down-regulated genes
Discussion
The molecular mechanisms for BCC tumor growth are still largely unknown. Recent studies have shown that chemokines and chemokine receptors may play important roles in the tumor growth and metastasis of various cancers. Prompted by this, we have investigated whether certain chemokine receptors may be involved in the tumorigenesis of BCC. Taking advantage of a human BCC cell line, we found high expression of CXCR4 by this cell line at mRNA level, which is comparable to, but less (~2.0- or 2.8-fold) than other CXCR4-expressing cancer cell lines (small cell lung carcinoma and breast cancer). By ISH and IHC assays, CXCR4 expression was also detected in a subset of tumor cells in BCC tissues; quantification of IHC assay further revealed CXCR4 was significantly more expressed by noduloulcerative- and sclerosing-type BCC than superficial-type BCC, indicating that CXCR4 expression may be correlated with more aggressive nature in certain BCC subtypes. Given that CXCL12 is constitutively expressed by dendritic cells and endothelial cells in human skin(Pablos, et al 1999), and there is no detectable expression of CXCL12 (the ligand for CXCR4) by BCC-1/KMC cells, CXCR4/CXCL12 interactions in BCC may take place in a paracrine, rather than an autocrine fashion.
We next address the functional significance of CXCR4 expression by BCC. Initially, wild-type BCC-1/KMC cells were treated with CXCL12 or not (as control) and then subject to in vitro calcium flux, proliferation and migration assays. Preliminary results (calcium flux assay in Fig. 2C; proliferation and migration assays, data not shown) revealed no significant difference either with or without CXCR4/CXCL12 interactions in wild-type BCC line, which were consistent with the previous report by Zhou et al. (Zhou, et al 2002) demonstrating that CXCL12 treatments did not support the growth, migration and invasion of wild-type, highly CXCR4-expressing glioma tumor cell line in vitro. It is conceivable that CXCR4 expression limited to a subset of BCC cells might not be enough to impose significant impacts on whole tumor cell growth in vitro, or there might exist some endogenous inhibitory factors for tumor cells; however, it is still hard to determine whether it remains the same case in a much more complicated tumor-stroma milieu in vivo. Given that such an approach (using wild-type BCC line only) would be
insufficient to delineate the importance of CXCR4 expression for BCC tumor progression in vivo, mainly due to the difficulty to set up an appropriate control, herein we chose to further amplify CXCR4 expression in wild-type BCC line and then observe whether there are phenotype alterations in tumor cells with increased level of CXCR4 expression, in vitro and in vivo. Using retroviral gene transduction, we found that CXCR4 may enhance BCC tumorigenesis by several mechanisms. Firstly, CXCR4 expression enhances BCC cell proliferation under low serum. This finding is in accordance with several reports demonstrating that CXCR4 is essential for the growth of multiple types of normal and tumor cells.(Barbero, et al 2003, Broxmeyer, et al 2003, Murakami, et al 2002, Zhou, et al 2002) Furthermore, we demonstrate that CXCR4 expression may protect tumor cells from UVB-induced apoptosis, manifested by decreased caspase-3 activity. This result is in line with other reports,(Broxmeyer, et al 2003, Zhou, et al 2002) showing that CXCR4/CXCL12 axis provided anti-apoptotic effects for tumor cells under growth factor withdrawal.(Zhou, et al 2002)
Recent in vitro studies have indicated that CXCR4/CXCL12 interactions may play an important role in tumor angiogenesis.(Koshiba,
et al 2000) As CXCR4 and CXCL12 are constitutively expressed by
endothelial cells,(Pablos, et al 1999) CXCL12 ligation not only induces chemotaxis for CXCR4-bearing endothelial cells, but also induces VEGF release from these cells in vitro.(Mirshahi, et al 2000) Beside serving as an angiogenic factor, VEGF in turn can promote both CXCR4 and CXCL12 expressions by endothelial cells,(Bachelder, et al 2002, Salcedo,
et al 1999, Salvucci, et al 2002) thus forming an amplification loop for
angiogenesis. While these studies clearly showed the possible influence of CXCR4/CXCL12 interactions on endothelial cells, they did not address, under a complex tumor/stroma interacting scenario, whether tumor cells may also play an active role in angiogenesis by chemokine expression. Our study provides in vitro evidences indicating that CXCR4 expression by tumor cells may play a direct role in tumor-associated neovascularization. In support of this, we further found that CXCR4/CXCL12 signaling enhance the expression of VEGF by tumor cells, implying that tumor cells may be actively involved in angiogenesis by VEGF secretion. Moreover, by injections of CXCR4-BCC cells, xenograft tumor transplants manifested visible neovascularization in nude mice. Thus, in the complex tumor-stroma milieu, tumor cells and endothelial cells may interact with each other to amplify angiogenesis by the expression and signaling of CXCR4.
The question remains how some tumor cells re-engineer themselves to express CXCR4 in favor of tumor progression. It has been well known that inherent inactivation mutation of tumor-suppressor PTCH gene in Sonic hedgehog signaling pathway play an important role in the pathogenesis of subsets of BCC,(Unden, et al 1997) a tumor also subject to CXCR4 regulation, as we have shown herein. Given that both CXCR4/CXCL12 and Sonic hedgehog pathways are critical players, not only in normal embryogenesis and development,(Horuk 2001, Taipale and Beachy 2001) but also BCC tumorigenesis,(Oro, et al 1997, Unden,
et al 1997) it would be reasonable to envisage that Sonic hedgehog and
CXCR4 signaling pathways may be correlated, in some way, upon BCC tumorigenesis.
In summary, we have determined that CXCR4 is expressed by a subset of human BCC cells, particularly those variants with more aggressive nature; Increased CXCR4 expression enhances proliferation, apoptosis-resistance, migration and angiogenesis of BCC cells in vitro, and more importantly, BCC tumorigenesis in vivo. Specific blockade of CXCR4 leads to tumor regression of BCC in vivo and may serve as a potential therapeutic strategy for certain type of BCC with more aggressive nature.
四. 計畫成果自評
We have faithfully executed this granted project and our
paper supported by this grant has been accepted by British
Journal of Dermatology (SCI rank: 3/38 at Dermatology in
2005). Please note the acceptance letter attached herein.
Acceptance letter from Brithish Journal of Dermatology BJD-2005-00437.R1
Dear Dr. Wu,
Thank you for submitting this paper to the British Journal of Dermatology. We are delighted to accept it for publication as Original Article (Dermatopathology) but there may still be some minor changes to make to your manuscript. Should this be the case our technical team will liaise directly with you. After the final amendments are made we will send your paper to the publishers and you will then receive page proofs.
It is now a policy of the Journal and the Publisher that we have hard copies of all of the figures in the manuscript. The reason for this is that when electronic figures are printed they can turn out quite different (with regard to the colour and quality) than they appear on screen. A hard copy of the figures will be a true representation of the quality and can be used for comparison. The first named author of the manuscript will be required to assign copyright in their paper to the British Association of Dermatologists. Copyright assignment is a condition of
publication and papers will not be passed to the publisher for production unless copyright has been assigned. A copyright form
can be obtained from the following link:
http://bjd.manuscriptcentral.com/index.html?mode=instruction Please post/fax the Copyright Form to the address below.(Government employees in both the US and the UK need tocomplete the Author Warranty sections, although copyright in such cases does not need to be assigned.)
Proofs will e-mailed as Acrobat PDF (portable document format) files. The e-mail server must be able to accept attachments up to 4MB in size. Acrobat Reader will be required in order to read this file. This software can be downloaded (free) from the following website:
http://www.adobe.com/products/acrobat/readstep2.html
This will enable the file to be opened, read on screen, and printed out in order for any corrections to be added. Further instructions will be sent with the proof. Proofs will be posted if no e-mail address is available; in your absence, please arrange for a colleague to access your e-mail to retrieve the proofs.
Yours sincerely, Dr. David Eedy Editor
British Journal of Dermatology British Journal of Dermatology BAD House 4 Fitzroy Square London W1T 5HQ Fax: +44 (0)20 7387 0240
五、參考文獻
Bachelder, R.E., Wendt, M.A. & Mercurio, A.M. (2002) Vascular endothelial growth factor promotes breast carcinoma invasion in an autocrine manner by regulating the chemokine receptor CXCR4. Cancer Res, 62, 7203-7206. Baggiolini, M. (2001) Chemokines in pathology and medicine. J
Intern Med, 250, 91-104.
Barbero, S., Bonavia, R., Bajetto, A., Porcile, C., Pirani, P., Ravetti, J.L., Zona, G.L., Spaziante, R., Florio, T. & Schettini, G. (2003) Stromal cell-derived factor 1alpha stimulates human glioblastoma cell growth through the activation of both extracellular signal-regulated kinases 1/2 and Akt.
Cancer Res, 63, 1969-1974.
S., Hangoc, G., Shaheen, M., Li, X. & Clapp, D.W. (2003) Stromal cell-derived factor-1/CXCL12 directly enhances survival/antiapoptosis of myeloid progenitor cells through CXCR4 and G(alpha)i proteins and enhances engraftment of competitive, repopulating stem cells. J Leukoc Biol, 73, 630-638.
Chiang, L.C., Chiang, W., Yu, H.S., Sheu, H.M. & Chen, H.Y. (1994) Establishment and characterization of a continuous human basal cell carcinoma cell line from facial skin (I) cytological behavior of early passages. Gaoxiong Yi Xue
Ke Xue Za Zhi, 10, 170-176.
Gailani, M.R., Leffell, D.J., Ziegler, A., Gross, E.G., Brash, D.E. & Bale, A.E. (1996) Relationship between sunlight exposure and a key genetic alteration in basal cell carcinoma. J
Natl Cancer Inst, 88, 349-354.
Gloster, H.M., Jr. & Brodland, D.G. (1996) The epidemiology of skin cancer. Dermatol Surg, 22, 217-226.
Horuk, R. (2001) Chemokine receptors. Cytokine Growth Factor Rev, 12, 313-335.
Koshiba, T., Hosotani, R., Miyamoto, Y., Ida, J., Tsuji, S., Nakajima, S., Kawaguchi, M., Kobayashi, H., Doi, R., Hori, T., Fujii, N. & Imamura, M. (2000) Expression of stromal cell-derived factor 1 and CXCR4 ligand receptor system in pancreatic cancer: a possible role for tumor
progression. Clin Cancer Res, 6, 3530-3535.
Miller, D.L. & Weinstock, M.A. (1994) Nonmelanoma skin cancer in the United States: incidence. J Am Acad Dermatol, 30, 774-778.
Mirshahi, F., Pourtau, J., Li, H., Muraine, M., Trochon, V., Legrand, E., Vannier, J., Soria, J., Vasse, M. & Soria, C. (2000) SDF-1 activity on microvascular endothelial cells: consequences on angiogenesis in in vitro and in vivo models. Thromb Res, 99, 587-594.
Muller, A., Homey, B., Soto, H., Ge, N., Catron, D., Buchanan, M.E., McClanahan, T., Murphy, E., Yuan, W., Wagner, S.N., Barrera, J.L., Mohar, A., Verastegui, E. & Zlotnik, A. (2001) Involvement of chemokine receptors in breast cancer metastasis. Nature, 410, 50-56.
Murakami, T., Maki, W., Cardones, A.R., Fang, H., Tun Kyi, A., Nestle, F.O. & Hwang, S.T. (2002) Expression of CXC chemokine receptor-4 enhances the pulmonary metastatic potential of murine B16 melanoma cells. Cancer Res, 62, 7328-7334.
Oro, A.E., Higgins, K.M., Hu, Z., Bonifas, J.M., Epstein, E.H., Jr. & Scott, M.P. (1997) Basal cell carcinomas in mice overexpressing sonic hedgehog. Science, 276, 817-821. Pablos, J.L., Amara, A., Bouloc, A., Santiago, B., Caruz, A., Galindo,
M., Delaunay, T., Virelizier, J.L. & Arenzana-Seisdedos, F. (1999) Stromal-cell derived factor is expressed by dendritic cells and endothelium in human skin. Am J
Pathol, 155, 1577-1586.
Rossi, D. & Zlotnik, A. (2000) The biology of chemokines and their receptors. Annu Rev Immunol, 18, 217-242.
Salcedo, R., Wasserman, K., Young, H.A., Grimm, M.C., Howard, O.M., Anver, M.R., Kleinman, H.K., Murphy, W.J. & Oppenheim, J.J. (1999) Vascular endothelial growth factor and basic fibroblast growth factor induce expression of CXCR4 on human endothelial cells: In vivo neovascularization induced by stromal-derived
factor-1alpha. Am J Pathol, 154, 1125-1135.
Salvucci, O., Yao, L., Villalba, S., Sajewicz, A., Pittaluga, S. & Tosato, G. (2002) Regulation of endothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1. Blood, 99, 2703-2711.
Staller, P., Sulitkova, J., Lisztwan, J., Moch, H., Oakeley, E.J. & Krek, W. (2003) Chemokine receptor CXCR4
downregulated by von Hippel-Lindau tumour suppressor pVHL. Nature, 425, 307-311.
Taichman, R.S., Cooper, C., Keller, E.T., Pienta, K.J., Taichman, N.S. & McCauley, L.K. (2002) Use of the stromal cell-derived factor-1/CXCR4 pathway in prostate cancer metastasis to bone. Cancer Res, 62, 1832-1837.
Taipale, J. & Beachy, P.A. (2001) The Hedgehog and Wnt signalling pathways in cancer. Nature, 411, 349-354.
Unden, A.B., Zaphiropoulos, P.G., Bruce, K., Toftgard, R. & Stahle-Backdahl, M. (1997) Human patched (PTCH) mRNA is overexpressed consistently in tumor cells of both familial and sporadic basal cell carcinoma. Cancer Res, 57, 2336-2340.
Vicari, A.P. & Caux, C. (2002) Chemokines in cancer. Cytokine
Growth Factor Rev, 13, 143-154.
Zhou, Y., Larsen, P.H., Hao, C. & Yong, V.W. (2002) CXCR4 is a major chemokine receptor on glioma cells and mediates their survival. J Biol Chem, 277, 49481-49487.