0 1993 Kluwer Academic Publishers. Printed in the Netherlands. 197
Osmotic stress and water stress have opposite effects on putrescine and proline
production in excised rice leaves
Chien Teh Chen & Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China *Author for correspondence
Received 24 December 1992; accepted 20 February 1993
Key words: abscisic acid, jasmonic acid methyl ester, Oryza sativa, osmotic stress, proline, putrescine, water stress
Abstract
The effects of water stress and osmotic stress (sorbitol treatment) on the production of putrescine and proline in excised rice leaves were compared. Osmotic stress and water stress were found to affect differentially the levels of putrescine and proline in excised rice leaves. Putrescine accumulation is induced by osmotic stress, whereas proline accumulation is induced by water stress. The effects of ABA on the levels of proline and putrescine are similar to those of water stress, whereas the effects of jasmonic acid methyl ester (JA-Me) are similar to those of osmotic stress. Water stress results in an increase of endogenous ABA is excised rice leaves. However, neither osmotic stress nor JA-Me has effect on endogenous ABA levels in excised rice leaves. Of particular interest is the finding that proline levels increase when putrescine levels induced
by osmotic stress or JA-Me are reduced by D-arginine and a-methylornithine. L-arginine and L-ornithine
applied exogenously also cause an increase in proline levels. It seems that L-arginine and L-ornithine are preferentially utilized as precursors for putrescine accumulation in excised rice leaves treated with osmotic stress and JA-Me, and for proline accumulation in excised rice leaves exposed to water stress and ABA.
Abbreviations: ABA = abscisic acid; BSA = bovine serum albumin; ELISA = enzyme-linked immunosor-
bent assay; HPLC = high performance chromatography; JA-Me = jasmonic acid methyl ester; PVP =
poly-vinylpyrrolidone
1. Introduction
Recently, considerable interest has been shown in the changes that take place in polyamine levels in the leaves subjected to various kinds of environ- mental stress. Water stress has been shown to increase the level of putrescine in leaf tissues [l 1, 211. Putrescine accumulation also occurs in cereal leaf segments exposed to osmotic stress [8, 9, 10, 221. It has long been known that proline accumu- lates in leaves of many plants in response to water
stress [l]. Proline accumulation has also been
observed in tobacco callus exposed to osmotic stress [6]. In experiments where osmotic stress is imposed in plant tissue by use of sorbitol or manni-
tol, it is difficult to separate purely osmotic effects from effects due to water stress generated during
such treatments. The present investigation was
undertaken to compare the effects of water stress and osmotic stress (sorbitol treatment) on the accu- mulation of putrescine and proline in excised rice leaves.
Proline accumulation can be markedly induced
by ABA in excised leaves of several plant species [5, 171. However, ABA did not affect the levels of putrescine in detached barley leaves [17]. Thus it
was of great interest to understand whether the
levels of putrescine and proline in excised rice leaves treated with ABA are similar to those exposed to water stress or osmotic stress. Jasmo-
nates and ABA show many and remarkable simila- rities - structurally, functionally, and regulatory [ 161. Thus, the effects of jasmonic acid methyl ester (JA-Me) on the accumulation of proline and putrescine were also investigated.
2. Materials and methods
2.1 Plant material and incubation condition
Rice (Oryza saliva cv Taichung Native 1) was cul- tured as previously described [12]. The apical 3 cm segments excised from the third leaves of 12- day-old seedlings were used. A group of 10 seg- ments was floated in a Petri dish containing lOm1 of test solutions. Incubation was carried out at 27 “C in the light (16.7pmol quanta mp2s-‘). Water stress was applied by a previously described method [3]. Briefly, leaf segments were exposed to the vapor above a solution of 0.5 A4 NaCI. Water stress was judged by loss of fresh weight. The fresh weight loss was about 9, 15 and 20% at 4, 8 and 12 h after the experiment, respectively. Osmotic stress was achieved by floating leaf segments on 0.6M sorbitol solution. The fresh weight loss in osmotically stressed leaf segments was about 3, 5 and 10% at 4, 8 and 12 h after the experiment, respectively. For ABA and JA-Me treatments, the optimum concentration (45 ,&Y) to induce proline or putrescine accumulation was used.
2.2 Determination of proline
Proline was extracted and its concentration deter- mined by the method of Bates et al. [2]. Leaf seg- ments were homogenized with 3% sulfosalicylic acid and the homogenate was centrifuged. The
supernatant was treated with acetic acid and acid ninhydrin, boiled for 1 h, and then absorbance at 520nm was determined. Levels of proline are expressed as pmol g-’ initial fresh weight.
2.3 Determination offree putrescine
For putrescine determination, leaf segments were homogenized in 5% perchloric acid. Putrescine levels were determined using HPLC after benzoyla- tion as described previously [4]. Levels of putres- tine are expressed as nmol g-’ initial fresh weight.
2.4 Determination of ABA
For ABA extraction, leaf samples were homoge- nized with extraction solution (80% methanol con- taining IOOmg 1-t butylated hydroxytoluene). Extracts were filtered through Whatman No. 1 fil- ter paper and rinsed twice with extraction solution. Filtrate was reduced to dryness in vacua at 30 “C. Samples were resuspended in 100% methanol. (NH&HP04 (500mM) was then added and the samples were allowed to stand for 10min at 4 “C until ammonium salts formed. Pigments and phe- nolics in ammonium salt solution were removed by passing through a PVP column [14]. The com- bined PVP column-filtrated solutions were adjusted to pH 3.0 with acetic acid. The acidified solution was eluted through a Cl8 cartridge. Methanol (20%, pH 3.2) was passed through the Cl8 cartridge to remove polar compounds. ABA trapped in the Cl8 cartridge was then eluted with 65% ethanol (pH 3.2). ABA solution was reduced to dryness in vacua, resuspended in Tris-buffered saline (50mM Tris HCl, 10mM NaCl, 1 mM MgC12, 15 mMNaNs, pH 7.5) and then subjected to HPLC. Samples were injected into a Waters M-6 UK Universal Liquid Chromatograph. They were eluted through a 250- x4 mm Cl 8 reverse- phase column (5 pm particle size) at a flow rate of 1 ml min-’ with 40% methanol in 0.02% acetic acid. ABA was collected by a fraction collector. The ABA fractions were then dried in vacua. Sam- ples were resuspended in Tris-buffered saline and 100% methanol and stored at -20 “C for ELISA analysis. ABA was quantitated by indirect ELISA according to Walker-Simmons [23] and Norman et al. [15]. ABA-4’-BSA conjugate was prepared according to Weiler [25] and used to determine ABA. ABA levels are expressed as pmol g-t initial fresh weight.
3. Results
Figure 1 shows the changes with time in levels of putrescine and proline in excised rice leaves incu- bated in water or exposed to osmotic stress or water stress in the light. Osmotic stress markedly increases putrescine levels. However, only a slight increase in putrescine levels was observed in excised leaves exposed to water stress. At variance
rrn
z 900 cz
3 600 ii a 2 p 300 0 0 0 H,O l WATER STRESS A OSMOTIC STRESS I I I I 0 H,O l WATER STRESS A OSMOTIC STRESS I 0 4 6 12 TIME, hFig. 1. Changes with time in the levels of putrescine and proline in excised rice leaves treated with water stress and osmotic stress. Excised rice leaves were treated with either water, water stress or 0.6 M sorbitol in the light. The levels of putrescine were quanti- tated at the times indicated. Bars represent SE; 3 replicates.
with the results of putrescine accumulation, water stress markedly increased proline levels, but osmo- tic stress only caused a slight increase in proline levels. The effects of ABA and JA-Me on the accu-
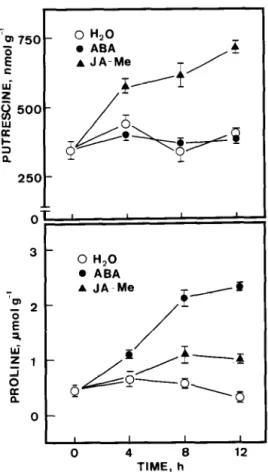
mulation of proline and putrescine are shown in
Figure 2. The increase of putrescine by JA-Me was evident at 4 h after incubation in the light.
However, ABA treatment did not result in an
increase of putrescine levels. Proline levels, on the other hand, increased significantly in excised rice leaves treated with ABA, but increased slightly in those treated with JA-Me. Clearly, the effects of ABA on the levels of proline and putrescine are similar to those of water stress, whereas the effects of JA-Me are similar to those of osmotic stress.
The levels of endogenous ABA rise markedly in response to water stress in several plant species
o H,O l ABA / 4 A JA-Me 0 H,O l ABA A JA-Me -.-* I I I I 0 4 8 12 TIME, h
Fig. 2. Changes with time in the levels of putrescine and proline in excised rice leaves treated with ABA and jasmonic acid methyl ester (JA-Me). Excised rice leaves were treated with either water, 45 PRIM ABA or 45 PM JA-Me in the light. The levels of putres- tine were quantitated at the times indicated. Bars represent SE; 3 replicates.
[24]. Since exogenously applied ABA mimics the effects of water stress on the levels of proline and putrescine, it is expected that water stress may result in an increase in ABA levels in excised rice leaves. As indicated in Figure 3, the levels of ABA, indeed, increased by water stress at 8 h after
the start of the incubation. Neither JA-Me nor
osmotic stress had any effect on the levels of ABA in excised rice leaves.
To characterize further the effects of osmotic
stress and JA-Me on the accumulation of putres-
tine, inhibitors of the biosynthesis of putrescine,
such as D-arginine and a-methylornithine, were
applied to sorbitol- and JA-Me-treated excised
rice leaves. The results are shown in Table 1. D-
arginine but not a-methylornithine significantly
TREATMENTS: W WATER STRESS OSMOTIC STRESS JA-Me 0 10 20 30 40 ABA, Pmd 9-’ Fig. 3. Effects of water stress, osmotic stress and jasmonic acid methyl ester (JA-Me) on the levels of ABA in excised rice leaves. Excised rice leaves were treated with either water, 0.6 M sorbitol, 45 j&f JA-Me or water stress in the light for 8 h. Bars represent SE; 3 replicates.
leaves in the absence of sorbitol and JA-Me. How-
ever, both D-arginine and cu-methylornithine
decreased the levels of putrescine induced by osmo- tic stress or JA-Me, indicating that osmotic stress and JA-Me may affect the activities of both argi-
nine decarboxylase and ornithine decarboxylase,
enzymes responsible for the biosynthesis of putres- tine [7]. Since D-arginine is more pronounced than
o-methylornithine in reducing the accumulation of
putrescine induced by osmotic stress or JA-Me,
arginine decarboxylase seems to be the major
enzyme responsible for the accumulation of putres- tine induced by osmotic stress or JA-Me.
Proline is known to be originated from L-glutamic acid, L-arginine and L-ornithine [ 191. When rice leaf segments were treated with L-arginine and L-or- nithine, proline level was significantly increased (Figure 4). It seems that proline and putrescine may share L-arginine and L-ornithine as precursors in excised rice leaves (Figure 5). Table 1 shows that
both D-arginine and a-methylornithine resulted in
an increase of proline levels in osmotically stressed and JA-Me-treated excised rice leaves. These results
suggest that D-arginine and o-methylornithine
result in more L-arginine and L-ornithine, respec- tively, available for proline accumulation.
4. Discussion
Sorbitol is a commonly used water stress agent.
Table 1. Effects of D-arginine and a-methylornithine in the ab- sence or presence of jasmonic acid methyl ester (JA-Me) or sor- bitol on the levels of putrescine and proline in excised rice leaves
Treatment Putrescine Proline
(nmol g-‘) Cm01 6) Water 380 f 10 0.25 f 0.02 D-Argine 284 zt 36 0.25 f 0.01 a-Methylornithine 408 f 27 0.25 It 0.01 JA-Me 690 it 10 0.40 f 0.01 JA-Me + D-Arginine 520 f 6 0.58 + 0.03 JA-Me + o-Methylornithine 576 f 4 0.65 310.01 Sorbital 930 f 58 0.45 f 0.01 Sorbital + D-Arginine 562 + 26 0.60 5 0.01 Sorbital + o-Methylornithine 752 zt 43 0.72 zt 0.02 Excised rice leaves were incubated in the light for 8 h in D-argi- nine (5 mM) or a-methylornithine (5 mM) in the absence or pre- sence of JA-Me (45 @) or sorbitol (0.6M) (Mean f SE, 3 replicates).
The results of the present investigation show that osmotic stress and water stress affect differentially the levels of putrescine and proline in excised rice
leaves. Putrescine accumulation is induced by
osmotic stress, whereas proline accumulation is
induced by water stress.
It has been reported that ABA accumulation is
not required for proline accumulation in wilted
barley leaves [20]. The present investigation
demonstrated that water stress and ABA treatment behaved similarly in terms of accumulation of pro- line and putrescine in rice leaves. It seems that pro- line accumulation induced by water stress is most
likely mediated through ABA in excised rice
leaves. This suggestion is further supported by the
observation that water stress resulted in an
increase of endogenous ABA in excised rice leaves.
Contrary to water stress, sorbitol and JA-Me
treatments do not result in an increase in endogen- ous ABA levels in excised rice leaves. However, Larsson et al. [13] detected an increase in ABA levels in wheat exposed to osmotic stress by treat- ment with up to 20% polyethylene glycol. It is known that polyethylene glycol does not enter the cell wall space [18] and sorbitol does [S]. Whether different effects of sorbitol and polyethylene glycol on ABA levels are due to the different properties of two osmotica or different plant materials used is deserving of critical experimentation.
The present investigation shows that osmotic
stress mimics the effects of JA-Me on the levels of putrescine and proline in excised rice leaves. At
6 - 5 . ‘0, 4 = : 3. ^ 2, 2 i 1 0 p” 0 0.6 Trn 0.5 E 0.4 =k - 0.3 !i i 0.2 0 E 0.1 0 I ’ ’ ’ ’ ’ ’ 0 10 20 30 40 50 mM
Fig. 4. Effects of L-arginine and L-ornithine on the levels of pro- line in excised rice leaves. Excised rice leaves were treated with various concentrations of L-arginine or L-ornithine in the light for 8 h. Bars represents SE; 4 replicates.
present, we have technical problems in assaying endogenous jasmonates and are unable to provide direct evidence that osmotic stress would result in an increase in jasmonates in excised rice leaves.
However, Parthier et al. [16] demonstrated that
endogenous jasmonates increased in barley leaf segments exposed to 1 M sorbitol. If sorbitol treat- ment behaves similarly in excised rice leaves, then the effects of sorbitol on the levels of putrescine are more likely mediated by changes of endogen- ous jasmonates. Overall, our results suggest that the increase in proline and putrescine during stress are mediated by different mechanisms.
Of particular interest of the present investigation is the finding that proline levels increase when putrescine levels induced by osmotic stress and JA-
Me are reduced by D-arginine and o-methylor-
nithine. We also observed that L-arginine and L-
Glutamate Ornithine v Arginine
y -Glut am!!leh y d\ Agmi ine 1
Putrescine
1
Pyrroline-5-carboxylate Sperimdine
1
Pro1 ine Spermine
1
Fig. 5. Biosynthetic pathways of L-proline and polyamines in plant systems.ornithine applied exogenously resulted in an in-
crease of proline, suggesting that proline and
putrescine may share L-arginine and L-ornithine as precursors in rice leaves. It seems that L-arginine and L-ornithine are preferentially utilized as pre-
cursors for putrescine accumulation in excised
rice leaves exposed to osmotic stress and JA-Me, and for proline accumulation in excised rice leaves exposed to water stress and ABA. However, this
conclusion will not be firmly established until
putrescine and proline accumulation during stress is followed upon labeling with L-[14C] arginine or L-[i4C] ornithine.
Acknowledgements
This work was supported by a grant from the
National Science Council, Republic of China (NSC
82-0409-B002-030). The assistance of Dr.
Huu-Sheng Lur with ABA assay is gratefully
acknowledged.
References
1. Aspinall D and Paleg LG (1981) Proline accumulation: phy- siological aspects. In: Paleg LG and Aspinall D (eds) The Physiology and Biochemistry of Drought Resistance in Plants, pp 280-295. Academic press: Sydney
2. Bates LS, Waldren RP, Teare ID (1973) Rapid determina- tion of free proline for water stress studies. Plant Soil 39:
205-207
3. Chen CT and Kao CH (1990) Comparative study of the me- tabolism of I-aminocyclopropane-I-carboxylic acid and se- nescence of water-stressed and ABA-treated excised rice leaves. Plant Cell Physiol 31: 463-468
4. Chen CT and Kao CH (1991) Senescence of rice leaves XXX. Levels of endogenous polyamines and dark-induced senescence of rice leaves. Plant Cell Physiol 32: 935-941 5. Chou IT, Chen CT, Kao CH (1991) Characteristics of the
induction proline by abscisic acid and isobutyric acid in de- tached rice leaves. Plant Cell Physiol 32: 269-272
6. Ebenhardt HJ and Wegmann K (1989) Effects of abscisic acid and proline on adaptation of tobacco callus cultures to salinity and osmotic shock. Physiol Plant 76: 283-288 7. Evans PT and Malmberg RL (1989) Do polyamines have
roles in plant development? Annu Rev Plant Physiol Plant Mol Biol40: 235-269
8. Flares HE and Galston AW (1982) Polyamines and plant stress: activation of putrescine biosynthesis by osmotic shock. Science 217: 1259-1261
9. Flores HE and Galston AW (1984) Osmotic stress-induced polyamine accumulation in cereal leaves I. Physiological parameters of the response. Plant Physiol 75: 102-109 10. Flores HE and Galston AW (1984) Osmotic stress-induced
polyamine accumulation in cereal leaves II. Relation to ami- no acid pools. Plant Physiol 75: 110-l 13
11. Kandpal RP and Rao NA (1985) Changes in the levels of polyamines in ragi seedlings during water stress. Biochem Int 11: 365-370
12. Kao CH (1980) Senescence of rice leaves IV. Influence of benzyladenine on chlorophyll degradation. Plant Cell Phy- siol 21: 1255-1262
13. Larsson M, Larsson CM, Whitford PN, Clarkson DT (1989) Influence of osmotic stress on nitrate reductase activ- ity in wheat (Trificum aesfivum L.) and the role of abscisic acid. J Exp Bot 40: 1265-1271
14. Mousdale DMA and Knee M (1979) Poly-N-vinylpyrroli- done column chromatography of plant hormones with methanol as eluent. J Chromatogr 177: 398-400
15. Norman SM, Poling SM, Maier VP (1988) An indirect en- zyme-linked immunosorbent assay for (+)-abscisic acid in Citrus, R&us, and Xanthium leaves. J Agric Food Chem 36: 225-231
16. Parthier B (1991) Jasmonates, new regulators of plant growth and development: many facts and few hypotheses on their actions. Bot Acta 104: 446-454
17. Pesci P and Reggiani R (1992) The process of abscisic acid- induced proline accumulation and the levels of polyamines and quatenary ammonium compounds in hydrated barley leaves. Physiol Plant 84: 1344139
18. Rubinstein B (1982) Regulation of H+ excretion. I. Effects of osmotic shock. Plant Physiol69: 939-944
19. Stewart CR and Boggess SF (1977) The effect of wilting on the conversion of arginine, ornithine, and glutamate to pro- line in bean leaves. Plant Sci Lett 8: 147-153
20. Stewart CR and Voetberg G (1987) Abscisic acid accumula- tion is not required for prohne accumulation in wilted leaves. Plant Physio183: 747-749
21. Turner LB and Stewart GR (1986) The effect of water stress upon polyamine levels in barley (Ho&urn vulgare L.) leaves. J Exp Bot 37: 170-177
22. Turner L and Stewart GR (1988) Factors affecting polyamine accumulation in barley (Hordeum vulgare L.) leaf sections during osmotic stress. J Exp Bot 39: 1 l-316
23. Walker-Simmons M (1987) ABA levels and sensitivity in de- veloping embryos of sprouting resistant and susceptible cul- tivars. Plant Physiol 84: 61-66
24. Walton DC (1980) Biochemistry and physiology of abscisic acid. Annu Rev Plant Physiol 31: 453-489
25. Weiler EW (1980) Radioimmunoassays for the differential and direct analysis of free and conjugated abscisic acid in plant extracts. Planta 148: 262-272