行政院國家科學委員會專題研究計畫 期中進度報告
栽培環境對稻米品質影響之蛋白質體研究:溫度調節性蛋白
質之功能分析(1/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2313-B-002-067- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學農藝學系暨研究所 計畫主持人: 盧虎生 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 5 月 31 日
行政院國家科學委員會補助專題研究計畫
期中進度報告
栽培環境對稻米品質影響之蛋白質體研究:
溫度調節性蛋白質之功能分析(1/3)
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 93-2313-B-002-067-
執行期間:2004 年 8 月 1 日至 2005 年 7 月 31 日
計畫主持人:盧虎生 教授
共同主持人:
計畫參與人員: 林韶凱、王湘儀
成果報告類型(依經費核定清單規定繳交):■精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究
計畫、列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公
開查詢
執行單位:國立台灣大學農藝系
中 華 民 國 九十四年 五月 二十日
(一)中文摘要 蛋白質為影響水稻穎果發育及稻米品質形成的主要因子,高溫環境則為主要 影響稻米品質的環境因子。本研究建立水稻穎果發育期間蛋白質表現之雙向電永 圖譜,並且利用逢機質譜分析(LC\MS\MS)的方法鑑定具表現差異的蛋白質。在表 現的蛋白質中,超過 400 個於水稻穎果發育期間表現的蛋白質於雙向電泳膠中被 分離出。其中有對溫度處理會產生表現差異的蛋白質,亦有品種兼具表現差異的 蛋白質,其中利用質譜分析鑑定出具表現差異的蛋白質共有 70 個,病找到其中 54 個已知功能的蛋白質。包括 21 個與碳水化合物生合成與代謝相關的蛋白質, 14 個與蛋白質生合成與累積相關的蛋白質,9 個與逆境反應有關的蛋白質。Waxy (Wx) 蛋 白 質 與 glutelins 為 隨 著 水 稻 穎 果 發 育 期 呈 顯 著 增 加 的 蛋 白 質 。 Allergen-like proteins, PPDK and NADH-SDH, 也在穎果發育期間表現,且扮 演其重要的生理角色與功能。其中, Wx 蛋白質各 isoforms 的表現量與 amylose 的含量成正相關。此外,本研究找出一個基因,具有高度 GC 比例的 DNA 序列, 且可能與心腹白的形成有關。高溫(35/30 ℃)會降低 Wx 蛋白質、allergen-like proteins 以及 elongation factor 1 β 的表現。,但是一群低分子量的 heat shock proteins (sHSP)、glyceraldehyde-3-phosphate dehydrogenase 以及 prolamin 等蛋白質的表現量會增加。sHSP 的含量與心腹白的程度呈正相關。此外,本期 計畫亦利用 RT-PCR 確認相關基因在 RNA 層次上的表現。
關鍵詞:水稻,蛋白質體學,高溫,稻米品質,Waxy 蛋白質,熱休克蛋白。
Abstract
Proteins are essential to rice caryopsis development and quality formation. High
temperature is an important environmental factor which may decrease grain quality. In the present study rice caryopsis proteins were profiled by two-dimensional polyacrylamide gel electrophoresis, and differentially expressed proteins were analyzed by liquid chromatography/tandem mass spectrometry (LC\MS\MS). Expressions of more than 400 polypeptide spots during caryopsis development, in response to temperature treatments or between varieties were monitored. Among them, more than 70 differentially expressed polypeptides were analyzed by LC\MS\MS. We identified 54 proteins with known functions. Of these, 21 were involved with
carbohydrate metabolism, 14 with protein synthesis and sorting, and 9 with stress responses. Waxy (Wx) proteins and glutelins were the most significant spots increased significantly during development. Allergen-like proteins, PPDK and NADH-SDH, also were expressed during development, implying their physiological roles in caryopsis. Expression of large isoforms of Wx proteins was correlated with the amylose content of rice caryopses. One protein with high GC content in its DNA sequence was correlated with the chalky trait of kernels. High temperature (35/30 ) ℃ decreased expression of Wx proteins, allergen-like proteins and elongation factor 1 β, but increased expression of small heat shock proteins (sHSP),
glyceraldehyde-3-phosphate dehydrogenase and prolamin. sHSP was positively correlated with the appearance of chalky kernels. In this study, we used RT-PCR to confirm the expression profiles of those genes.
Keywords: rice; proteomics; high temperature; grain quality; Wx protein; heat shock
protein.
(二)文獻探討與試驗目的 台灣稻米的生產,隨著生活及消費水準的提昇,目前以米質提升為主要目 標。良質米除了需要有優良的品種外,更需要適當的環境條件配合,方能生產高 品質的米。 在環境因素中,榖粒充實期不適當的溫度常導致水稻低產,且更降低了米粒 的品質。研究報告指出,水稻榖粒充實期間的溫度對榖粒產量及米質關係最為密
切(稻津 1979、1990; Tamaki et al. 1989a、b、c)。台灣的一、二期稻作生
育溫度截然不同,稻米品質也有差異,兩期稻作分別處於高溫(南部一期作)與低 溫(中北部二期作)環境。因此,溫度對水稻榖粒充實期的影響,實為台灣重要的 研究課題。以米質而言,米粒中的主要成分如澱粉、蛋白質以及游離脂肪酸等皆 易受到環境影響,其成分的改變將影響米粒外觀及食味品質(Tamaki et al. 1989a、b、c)。以本省的栽培管理而言,水稻生育溫度隨著地點以及時間的不同, 致使一、二期稻作在不同的栽培地榖粒充實期所處的溫度環境差異甚大(盧等 1999)。 水稻開花後,胚乳開始分裂及膨大,此時期澱粉與貯藏性蛋白質會隨發育期 逐漸累積。在水稻開花約第 6 天至第 15 天,為貯藏性物質最快速累積的時期, 此時期也是對環境最敏感的階段。在高溫環境中,會影響穎果充實的速率,且往 往導致心腹白的產生,導致品質的下降。 然而,影響此重要生育階段的分子基 礎仍未知,且環境如何影響水稻穎果發育的調控機制之研究也相當缺乏。 蛋白質是植體主要的功能性分子。水稻榖粒中的蛋白質受溫度環境的影響相 當大,早期水稻榖粒蛋白質的研究多集中於粗蛋白質含量的變化,並指出蛋白質 含量與米質呈負相關。水稻胚乳蛋白質約佔乾重之 20%,依溶解度分類可分為四 群,其中鹼溶性蛋白質 (Glutelins)與醇溶性蛋白質(prolamins)為兩種最主要 的貯藏性蛋白質,也是影響稻米品質的重要因子。而水溶性蛋白質(Albumins) 與鹽溶性蛋白質(globulins)通常扮演胚乳中酵素或調節生理代謝的功能。不同 種類的貯藏性蛋白質分子與稻米食用、營養及消化品質也有不同的相關性 (Jliano 1985; Masumura et al. 1991a、b; Souza et al. 1993; Chrsatil 1992)。 以往的水稻穀粒研究多集中在醇溶性蛋白質與鹼溶性蛋白質,很少有探討水溶性 蛋白質與鹽溶性蛋白質的角色。 蛋白質也是基因表現後的產物,栽培環境因素會影響基因表現。因此,從蛋 白質的表現層面可反應出不同生育環境條件及不同榖粒發育充實期的生理狀 態,若再配合上生理上乾鮮重變化、蛋白質含量以及細胞數目的變化等研究分 析,可建立基因表現、環境因素與穀粒生長發育上的關聯性。 蛋白質體學的發展可以在整體的層面上探討基因表現與穀粒發育間的相關 性,並歸納出其中的調控方式。本試驗以三年為期,以水稻穀粒為材料,利用高 解析度的蛋白質雙向電泳與銀染將穎果蛋白質分離,再以 Q-TOF 或 LC/MS/MS 之 蛋白質定序等方法,研究水稻穎果發育早期蛋白質表現及與溫度環境之關係,探 討關鍵性功能蛋白質所扮演的角色,及其與溫度、穎果發育、甚至於稻米理化性 質間的關係,作為稻米品質改良的學理依據。本計畫亦針對可能影響稻米品質的 基因,以具有不同遺傳背景的水稻品種,配合溫度環境處理,分析其具表現差異 的關鍵蛋白質,以及 RT-PCR 的測定確認相關蛋白質在 mRNA 層面上的表現是否類 似,進而了解相關蛋白質表現與稻米品質形成之關係。 1
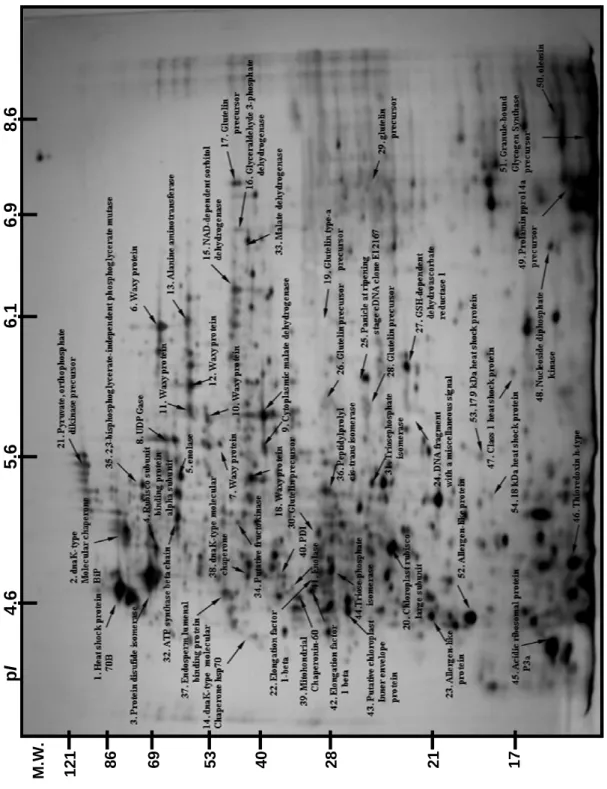
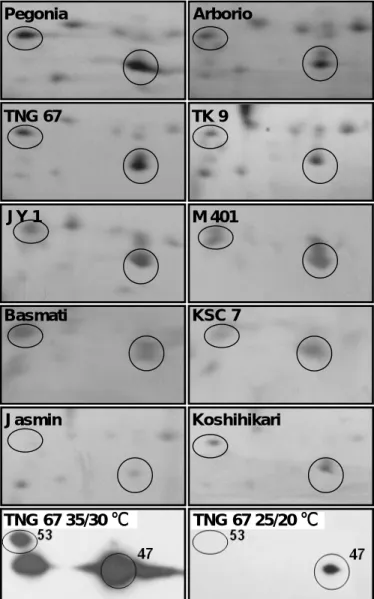
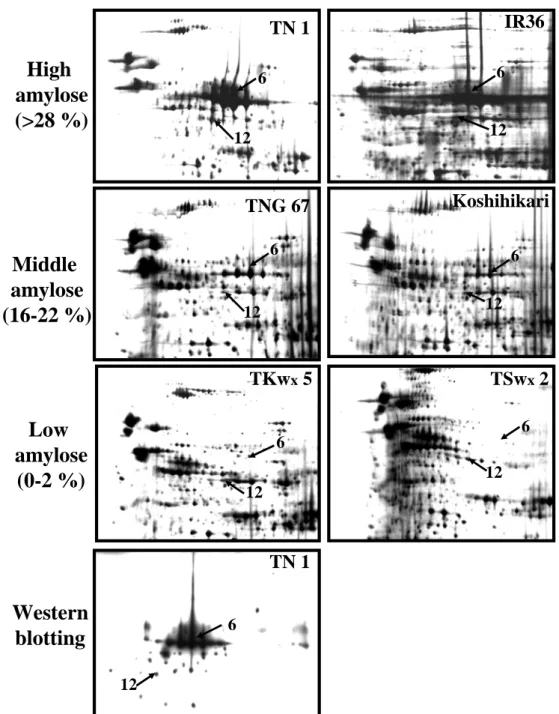
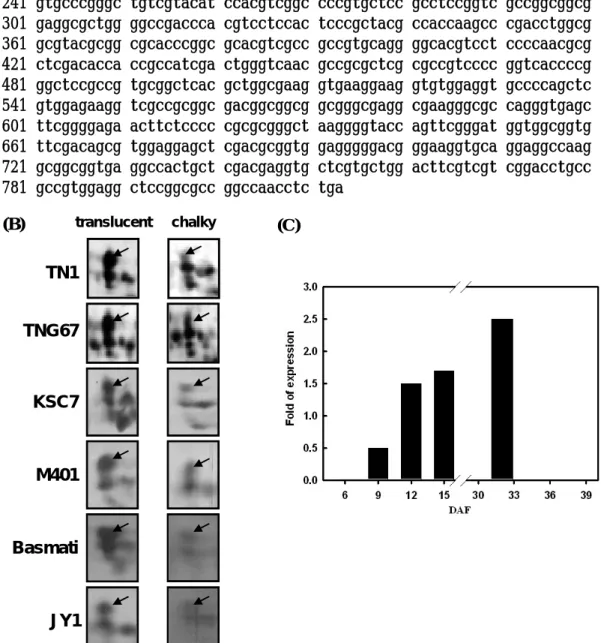
(三)結果與討論 (一)蛋白質鑑定(Protein identification) 在水稻穎果發育第 6 天至第 15 天,經雙向電泳與銀染,可分離超過 300 個 以上的主要蛋白質。並依分子量由大到小,pI由低至高的順序,兩年來經 Q-TOF 或 LC/MS/MS 分析鑑定超過 70 個蛋白質 (Fig. 1.)。在此方法於各參試處理或重 複試驗中呈穩定表現與高度的再現性。本研究再將已鑑定的 70 個蛋白質依功能 區分為五大類,分別為碳水化合物生合成、貯藏性蛋白質生合成、逆境蛋白質、 其他功能蛋白質與未知功能的 EST clones。 (二)熱休克蛋白質與稻米品質之關係 本試驗以在遺傳背景上,具有不同程度心腹白特性的品種為材料,其中 Pegonia、Arborio 與 JY1 等三品種為一般栽培環境下稻米即有明顯心腹白的品 種,TNG67、TK9、M401 與 Basmati 則為在高溫環境易產生心腹白的品種。KSC7、 Jasmin、Koshihikari 則為不易產生心腹白的品種。本試驗以這些品種為材料, 分析其第一群熱休克蛋白(sHSP)表現的情形,發現心腹白程度愈高的品種,其 16.9、17.9、18 這三群的 sHSP 會大量累積。反之,心腹白程度愈低的品種,則 sHSP 的累積量則較少甚是無表現(Fig 2.)。為確認該區域確為 sHSP,本試驗以 TNG67 進行高溫以及正常溫度處理,以 class 1 HSP 之抗體進行 Western blotting 分析的結果,顯示電泳該區域表現的蛋白質確為 sHSP。 本期結果顯示,第一群的熱休克蛋白可能與心腹白的形成有關,本研究室正 進一步的針對熱休克蛋白與心腹白的關聯進行分析,期能找出其可能的機制。 (三)直鏈澱粉合成酵素 直鏈澱粉(amylose)含量可顯著影響稻米品質。一般在水稻中直鏈澱粉合成基 因 Waxy gene 為單一基因。本研究經蛋白質體分析後得到 8-10 個 isoforms。可 分為兩群,其中一群分子量較高的 4 個 isoforms 之蛋白質含量與直鏈澱粉含量 呈正相關(Fig. 3.)。本研究進一步發現不同的 isoform 間可能是藉由磷酸化作 用的調控所形成。目前結果顯示,直鏈澱粉合成酵素(GBSS)在不同品種間及不同 溫度環境表現不同的量,最終可能影響澱粉性質。 (四)WB 基因與稻米品質之關係 利用蛋白質雙向電泳,配合具有不同心腹白特性的品種,以輕微至嚴重心腹 白程度區依序為 TN1、TNG67、KSC7、M401、Basmati、JY1 等,依序分析其蛋白 質表現圖譜,發現一個於本試驗中編號 25 之蛋白質,其累積量與心腹白的嚴重 程度呈負相關(Fig 4B)。其中,TN1 為輕微心腹白的品種,其 No. 25 蛋白表現 量高,反之,JY1 為嚴重心腹白的品種,其 No. 25 蛋白表現量相對偏低。在 NCBI 資料庫比對的結果顯示,此基因的功能目前並不清楚。但是進一步的比對其序 列,發現序列中具有 74% GC 比例,但無明顯的特徵序列(Fig 4A)。此蛋白質於 受粉後九天開始累積,直至榖粒完全成熟,其累積量亦達最大值(Fig 4C)。 此基因的表現亦可能與心腹白的形成有關,目前正著手進行基因轉殖的工 作,期能找出此基因可能的功能及其扮演的角色,本實驗室暫時將其命名為 WB(white belly)基因。 2
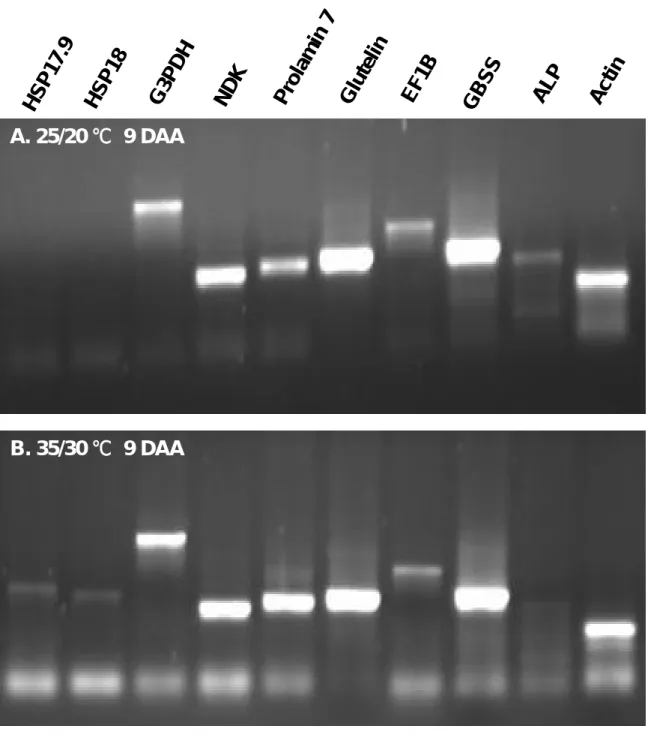
(五)利用 RT-PCR 確認具表現差異的基因與蛋白質
為了確認蛋白質雙向電泳上,各主要具表現差異蛋白質的表現量,與其 mRNA 層次上的表現量是否一致,將主要具有差異的數個基因進行 RT-PCR 分析,如 HSP17.9 and HSP18; prolamin 7: prolamin gene no. 7; G3PDH: G-3-P
dehygrogenase; NDK: nucleoside diphosphate kinase 1; GBSS: granule bound starch synthase; EF1B: elongation factor 1 β; ALP: allergen like protein. 且以 Actin 作為對 照,結果顯示,高溫下此十個基因的表現量與蛋白質的表現量一致,在蛋白質層 面上表現量較高,則在 RT-PCR 之表現也較高(fig 5.)。此結果反應這類基因確 實受溫度影響,進而可能影響稻米理化性質,影響品質之形成過程。 (四) 計畫成果自評 本計畫著重於利用蛋白質體學技術瞭解稻米理化性質的生理成因,進而期待 能達成改善米質的目標。本研究內容與原定計畫目標一致,本試驗為三年期的研 究計畫之第一年期,目前已順利將 proteomics 技術應用於水稻穎果在不同溫度 環境下之蛋白質表現圖譜的差異分析。已完成 70 個具有表現差異的蛋白質分子 之序列分析與比對,並得知各分子在水稻基因組中的位置及其可能的功能。 目前已知在日本型與印度型品種,在各穎果發育期於不同溫度環境下的蛋白 質差異表現,藉由 proteomics 的技術將各具有差異表現的蛋白質一一進行功能 分析,提供水稻基礎的蛋白質圖譜分析研究,判定各蛋白質在水稻穎果發育中所 扮演的角色,以供作將來育種與分子操作上的重要參考指標。 在三年期的計畫中,目前第一年的分析,已陸續發現許多以往未知的與穎果 發育及貯藏性物質累積的的代謝機制,如直鏈澱粉合成酵素的調控修飾作用。此 外,亦分析數個與高溫逆境相關的蛋白質,特別是與心腹白形成可能有關的基 因。其中包括 WB 基因、sHSP 基因。本年度計畫亦利用 RT-PCR 來確認這些具有 表現差異的蛋白質,其 mRNA 層次上是否也有類似的差異表現,且已獲得明確結 果,期盼能進一步應用這些結果於育種之分子標誌以及品質的改良工作上。 五、參考文獻 1.稻津 脩。1990。米品質作物學。良質味品種特性栽培。日作紀 59 : 611-615。 標準計測方法﹒1988﹒日本食糧廳﹒東京﹐日本﹒ 2.盧虎生。1999。溫度對水稻穎果充實發育及稻米品質的影響。環境與稻作生產 p.106-118。台灣省農業試驗所中華農業氣象學會。
3. Chien P. J., H. S. Lur and C. Chu. 1997. Effects of culture period on rice storage proteins. Chinese Agron. J. 7:333-341.
4. Chrastil, J. 1992. Correlations between thephysicochemical and functional properties of rice. J. Agric. Food Chem.40:1683-1686.
5. Huang, Y. C., H. S. Lur and C. Chu. 1995. Varity identification and quality characterization of milled rice grains using reverse-phase high performance liquid chromatography I. Technique development, prolamins, glutelins andvariety characterization. J. Agron. Assoc. China 171:1-13.
6. Juliano, B.O. 1985. Biochemical properties of rice. In B.O. Juliano, ed, Rice : Chemistry and Technology. American Assoc. of Cereal Chemists. USA,
pp.175-197.
7. Liang, Y. L., and H. S. Lur. 2001. Conjugated and Free Levels in Normal and Aborting Maize Kernels. Crop Science (in revision).
8. Masumura, T., N. Mitsukawa, K. Tanaka, and S. Fujii. 1991. Rice storage proteins : Genetic analysis of accumulation process.In Biotechnology in Agriculture and Forestry (ed by Y. P. S.Bajaj) Vol 14,pp.495-507.
9. Myers P. N., T. L. Setter, J. T. Madison and J. F. Thompson. 1990. Abscisic acid Inhibition of endosperm cell division in cultures maize kernels. Plant Physiol. 94:1330-1336.
10. Souza, S.R., E.M.L.M. Stark, and M.S. Fernandes. 1993. Effects of
supplemental-nitrogen on the quality of rice proteins. J. Plant Nutr. 16:1739-1751. 11. Tamaki, M., M. Ebata, T. Tashiro, and M. Ishikawa. 1989. Physocoecological
studieson quality formation of rice kernel. Ⅱ.Changes in quality of rice kernel during graindevelopment. Jpn. J. Crop Sci. 58:659-663.
12. Tung C. W., C. Y. Wang, H. S. Lur and C. Chu. 1997. Grain storage proteins in Taiwan rice cultivars:quantifying and RP-HPLC pattern analysis of glutelines and prolamines. Chinese Agron. J. 7:115-125.
p I 4 .6 5. 6 6 .1 6. 9 8 .6 12 1 86 69 53 40 28 21 17 M.W .
Figure 1. A 2D-PAGE protein expression profile of rice caryopses 6 days after anthesis. Caryopses of TN 1 cultivar were used for the analysis. Arrows and numbers indicate the corresponding annotations for the spots.
Pegonia Arborio TNG 67 TK 9 Jasmin Koshihikari TNG 67 35/30 ℃ TNG 67 25/20 ℃ JY 1 M 401 Basmati KSC 7
Figure 2. sHSP expression in rice cultivars with and without the chalky trait.
Grains of the cultivars Pegonia, Arborio, TNG 67, M401 and JY 1 have the chalky trait. TK 9, Jasmin, basmati, KSC 7 and Koshihikari are premium quality cultivars that do not exhibit this trait. Twenty mature grains of each cultivar were randomly sampled,
dekusked, milled and subjected to 2D-PAGE analysis. To confirm sHSP expression, heat-treated samples of TNG 67 were used for immunoanalysis (bottom left and right boxes). Circles and numbers indicate the corresponding sHSP spots.
TSwx2 TKwx5 TN 1 TNG 67 Koshihikari IR36
High
amylose
(>28 %)
Middle
amylose
(16-22 %)
Low
amylose
(0-2 %)
Western
blotting
12 6 TN 1 12 6 12 6 12 6 12 6 12 6 12 6Figure 3. Expression of Wx proteins in cultivars with different amylose contents. TN 1 and IR 36 are indica type cultivars that have high amylose content. TNG 67 and Koshihikari are japonica type cultivars whose amylose content is medium. TKwx 5 and TSwx 2 are japonica and indica type cultivars, respectively, that have low amylose content (wx type variety). The gel of the TN 1 sample was subjected to immunodetection to confirm the expression of Wx protein isoforms. Arrows and numbers indicate the spots corresponding to the Wx protein isoforms.
translucent chalky
(A)
1 atgatttgcc tccgagccgc cttccccttg acctcctcct tctcctcctc ccctctccgc 61 cgcctcgccc tcaagccctc ctcctcccgc gccgccgccg ccgccgccat gtcgtccgcc 121 cccgccaccg ccatcgccgc gccggtcgag cacatcgtgc tcatcaaggt ccgccccgag 181 gcggcggcgt cgggcgccgc cgcggcgatg gtgtcctcgc tgcaggcgct gtccagcgtg 241 gtgcccgggc tgtcgtacat ccacgtcggc cccgtgctcc gcctccggtc gccggcggcg 301 gaggcgctgg ggccgaccca cgtcctccac tcccgctacg ccaccaagcc cgacctggcg 361 gcgtacgcgg cgcacccggc gcacgtcgcc gccgtgcagg ggcacgtcct ccccaacgcg 421 ctcgacacca ccgccatcga ctgggtcaac gccgcgctcg cgccgtcccc ggtcaccccg 481 ggctccgccg tgcggctcac gctggcgaag gtgaaggaag gtgtggaggt gccccagctc 541 gtggagaagg tcgccgcggc gacggcggcg gcgggcgagg cgaagggcgc cagggtgagc 601 ttcggggaga acttctcccc cgcgcgggct aaggggtacc agttcgggat ggtggcggtg 661 ttcgacagcg tggaggagct cgacgcggtg gagggggacg ggaaggtgca ggaggccaag 721 gcggcggtga ggccactgct cgacgaggtg ctcgtgctgg acttcgtcgt cggacctgcc 781 gccgtggagg ctccggcgcc ggccaacctc tga
(B) (C) TN1 TNG67 KSC7 M401 Basmati JY1
Figure 4. Characteristics of protein no. 25 gene. (A) DNA sequence. (B) The 2D expression patterns of chalky and translucent kernels of TN1, TNG67, KSC 7, M401, Basmati and JY 1. (C) Change of abundance during development.
A. 25/20 ℃ 9 DAA
H
S
P
1
7
.9
H
S
P
1
8
G
3
P
D
H
N
D
K
P
ro
la
m
in
7
G
lu
te
li
n
E
F
1
B
G
B
S
S
A
L
P
A
c
ti
n
B. 35/30 ℃ 9 DAA
Figure 5. Expression of putative temperature responsive protein genes was analyzed by reverse transcription PCR. HSP17.9 and HSP18: heat shock protein with Mr of 17.9 and 18 respectively; prolamin 7: prolamin gene no. 7; G3PDH: G-3-P dehygrogenase; NDK: nucleoside diphosphate kinase 1; GBSS: granule bound starch synthase; EF1B: elongation factor 1 β; ALP: allergen like protein. Actin was used to check the RNA and amplification conditions of the RT-PCR reaction internally.