國立交通大學
生化工程研究所
碩士論文
研究白色念珠菌 REP3 突變株的功能性補救
Study of REP3 (Regulator of Efflux Pump)
functional rescue in rep3/rep3 homozygous mutants
of Candida albicans
研究生 : 吳金蓉
指導教授 : 楊昀良博士
研究白色念珠菌 REP3 突變株的功能性補救
Study of REP3 (Regulator of Efflux Pump) functional rescue in
rep3/rep3 homozygous mutants of Candida albicans
研究生 : 吳金蓉 Student : Chin-Jung Wu
指導教授 : 楊昀良 博士 Adviser : Dr. Yun-Liang Yang
國立交通大學
生化工程研究所
碩士論文
A Thesis
Submitted to Institute of Biological Engineering
National Chiao Tung University
in Partial Fulfillment of the Requirements
for the Degree of
Master
in
Biological Engineering
July 2007
Hsinchu, Taiwan, Republic of China
中文摘要
伺機性病原菌—白色念珠菌 (Candida albicans) 是造成人體黴菌感染最常見的病 原。Azole 類藥物常用來治療白色念珠菌的感染。然而,廣泛使用 fluconazole 使得抗藥 性白色念珠菌菌株之案例大增,進而導致臨床上治療失敗的機會增高。因此,瞭解白色 念珠菌抗藥機轉對於克服黴菌感染是很重要的。排藥幫浦CDR1 和 CDR2 的過量表現常 見 於 抗 藥 性 臨 床 菌 株 , 然 而 其 表 現 被 如 何 調 控 卻 所 知 甚 少 。 實 驗 室 先 前 經 由 在Saccharomyces cerevisiae 所 進 行 之 library screening 發 現 在 白 色 念 珠 菌 中 , REP3
(Regulator of Efflux Pump) 能增加 CDR1Ym990348啟動子-lacZ 的 β-galactosidase 活性約
四倍。再者根據藥物敏感性測試的結果顯示 REP3 突變株對 azole 類藥物的感受性有提
高的現象,但REP3 突變補救株對藥物感受性卻沒有恢復。本研究重新建構突變株,但
結果仍同,經Real-time PCR 檢測顯示 REP3 mRNA 確實存在 rep3/rep3::REP3 突變補 救株中且表現量與野生株SC5314 相近。意外的是,在 rep3/rep3 突變株、rep3/rep3::REP3
突變補救株及野生株 SC5314 中 CDR1 的 mRNA 量相近。再者,在藥物誘導下,另一
個距REP3 基因下游處 266 個核酸的 orf19.3926 表現量在 rep3/rep3::REP3 突變補救株
中比 rep3/rep3 突變株及野生株提高了 3.5 倍。即使利用 SAT1 的策略再次重新建構
rep3/rep3::REP3 突變補救株,以減少額外載體 DNA 序列介入 rep3/rep3 突變株中可能
造成的干擾,仍然無法補救rep3/rep3 突變株對藥物敏感的表現型。由於在 rep3/rep3 突
變株中,CDR1 之 mRNA 並未下降,因此,對於 REP3 是否透過其他非調控 CDR1 的
路徑而影響白色念珠菌的藥物抗藥性,以及 rep3 突變株的功能難以補救回復的原因,
Abstract
Candida albicans is an opportunistic fungal pathogen and is the most common cause of
deep mycoses in humans. Azole therapy is commonly used to treat C. albicansinfections. However the widespread use of azoles led to an increased frequency of treatment failure due to azole-resistant C. albicans in clinical setting. Therefore, understanding the molecular mechanisms of drug resistance in C. albicans is important to render the fungal infection. Over expression of drug efflux pumps— CDR1 and CDR2 (Candida Drug Resistance 1 and 2) in azole resistant C. albicans is commonly observed but the regulatory mechanism is poorly understood. Previously in the laboratory, REP3 (Regulator of Efflux Pump) was isolated from Candida genomic library due to its ability to increase the β-galactosidase activity of
CDR1YM990348 promoter-lacZ about four folds in Saccharomyces cerevisiae in the presence of
miconazole. According to the results of the drug susceptibility tests, rep3/rep3 homozygous mutant seems to be more susceptible to miconazole, itraconazole, ketoconazole, fiuconazole and voriconazole than the SC5314 wild-type strain in spite of rep3/rep3::REP3 rescued strains not showing a restoration of drug susceptibility phenotype. In this study, REP3 rescued strains were re-constructed and the outcome remained the same. No REP3 mRNA could be detected in rep3/rep3 mutant and the real-time PCR results showed that the REP3 mRNA could be detected in the rep3/rep3::REP3 rescued strains in similar quantity as that of the SC5314 wild-type strain. Surprisingly, the expressions of CDR1 were similar in the
rep3/rep3 mutants, rep3/rep3::REP3 rescued strains, and SC5314 wild type strain.
Furthermore, the expression level of the orf19.3926, which locates at the down stream 266 bps from REP3, increased about 3.5 folds in the REP3 rescued strain than that of the wild-type strains and rep3/rep3 mutants in the presence of miconazole. Even though the
REP3 rescued strains were reconstructed through SAT1 flipping method to eliminate the
potential interference from the integration of vector DNA fragment in C. albicans, the regenerated rep3/rep3::REP3 rescued strains still cannot restore completely the drug susceptibility phenotype of rep3/rep3 mutant. Since the mRNA level of CDR1 in rep3/rep3 mutant was the same as that in the wild type. REP3 may be involved in drug resistance
through pathways other than CDR1 in C. albicans. There is no clear reason as the why the
rep3/rep3::REP3 rescued strain could not restore the REP3 phenotype. Both will require
誌謝
我常以為,學如逆水行舟,而今輕舟已過萬重山。一路行來,點滴在心,因此下筆 之時,自己便預料這將是一篇冗長的謝詞,雖然這不是一本完美的論文,但這本論文的 完成,要感謝的人真的很多,僅以此文表達我的誠摯謝意。 由衷的感謝楊昀良教授兩年來悉心指導與啟發,培養學生獨立思考與解決問題的能 力,於論文研究及撰寫期間,細心斧正,使學業得以順利完成。論文研究期間,承蒙國 家衛生研究院羅秀容教授之關愛,予以學生實驗上諸多協助與指導,銘感衷心。文稿初 成,復蒙本校彭慧玲教授細心校稿與口試時的指導,使本論文得已益臻完善。 於大學專題期間,感謝中興大學林金和教授懇切的教導,大葉大學游志文與洪淑嫻 教授的循循善誘與細心指導,帶領我進入科學世界,讓我對科學有更深一層的體認,於 此一併敬表謝忱。 國家衛生研究院的一群夥伴們;學姊佳君不論生活或實驗上的幫助,訓練我對邏輯 思考的重要性,伴我走過這沒有掌聲的舞台-沒有妳給我的引導,相信今天的研究之路 不會如此順利;感謝學長彰勳(Scott),不時的給予我幫助和鼓勵;學姊惠菁,謝謝你與 我一同經歷交大與國衛院來往穿梭的旅程;俐君、國鋆、仁忠、誌偉、緯婷、安慧、志 兆謝謝你們的相互砥礪與支持。另一群還在交大的熱血澎湃青年;學弟妹淑萍、淑貞、 旻秀、敏書;小朋友佳叡、阿毛、小倩;你們的笑聲,是這緊湊、忙碌的研究生活中不 可或缺的緩衝液,永遠在我耳邊迴響,請繼續加油喔!一起並肩作戰的同學新彬、育穎, 好開心我們一起劃下碩班句點。已畢業的學長姐們,大小聲怡瑾、好人杏枚、好脾氣建 斈、鑽石嘴砲志豪,謝謝你們讓我以歡笑而非淚水,度過了這研究所的歲月。 嘉佑,認識你讓我知道人生的相遇並非偶然,而是必然,距離對我們倆而言並非阻 礙而是另一分幸運,何其有幸的是能透過遠距離擁有你默默在背後永不歇息的支持。 我最親愛的家人阿公,感謝有你在天上的庇祐;爸、媽,謝謝你們從小到大對我無 怨無悔的栽培與付出,若不是你們的支持,我不會有今日小小的成就。 最後,將此論文獻給所有愛我及我愛的你們...Contents
Abstract (Chinese)...i Abstract (English)...ii Acknowledgement...iv Contents...v Contents (Figures) ...x Contents (Tables)...xiiiChapter 1. Introduction...1
1.1 Clinical significance of Candida infection...1
1.2 Candida albicans is an opportunistic pathogen...2
1.3 Resistance of C. albicans to antifungal agents...3
1.4 Mechanism of resistance to azole antifungal agents ...5
1.5 Candida drug resistance gene: CDR1...7
1.6 Cis- and trans-acting factors regulating of CDR genes...8
1.6.1 Cis-regulatory factors of efflux pumps involved in azole resistance ...8
1.6.2 Trans-regulatory factors of efflux pumps involved in azole resistance ...10
1.7 Previous work in the laboratory ...11
Chapter 2. Materials and Methods...13
2.1 Materialsts ...13
2.1.1 Strains...13
2.1.2 plasmids and primers...13
2.1.3 Chemicals and reagents ...13
2.1.4 Buffers ...15
2.1.5 Medium...16
2.1.6 Enzymes ...17
2.1.7 Equipments ...17
2.1.8 Strains and growth media ...18
2.2 Methods ...19
2.2.1 DNA methods... 19
2.2.2 Preparing electrocompetent cells...19
2.2.3 E. coli. transformation with electroporation...20
2.2.4 Competent cell prepared using Calcium chloride ...21
2.2.5 E. coli. transformation using Calcium chloride...21
2.2.6 Constructions of plasmids and strains ...22
2.2.7 Complementation of the REP3 gene (CJC7 and CJC10)in rep3/rep3 mutant strains (CSC80 and CSC81) ...23
2.2.8 Complementation of the REP3 gene (CJC13~CJC18)in rep3/rep3 his1::hisG/ his1:: hisG::HIS1 mutants (CJC2 and CJC4) by SAT1 method...24
2.2.9 The REP3 rescued strains (CJC13 ~ CJC18) excise the SAT1 flipping ...25
2.2.10 Isolation of genomic DNA ...26
2.2.11 Transformation of Candida albicans by lithium acetate method...27
2.2.12 Transformation of Candida albicans by electroporation method ...28
2.2.13 Southern blot analysis...29
2.2.14 Antifungal susceptibility test with agar dilution assay...31
2.2.15 Antifungal susceptibility test with CLSI M-27A method ...31
2.2.16 Isolation of RNA by the hot acid phenol method...32
2.2.17 Isolation of RNA by the Masterpure Yeast Purification Kit ...34
2.2.18 Quantitative analysis of the mRNA level by real-time polymerase chain reaction (real-time PCR)...35
Chapter 3. Results...38
3.1 Generation of histidine prototrophy of rep3/rep3 mutant ...38
3.2 The antifungal drugs susceptibility of histidine prototrophy rep3/rep3 is identical with h- istidine auxototrophy rep3/rep3 ...38
3.3 The REP3 rescued strains YLO221 and YLO222 cannot restore the drug susceptibility p- henotype of rep3/rep3...39
3.4 The REP3 sequence of the rescue plasmid (pWJB24) is identical with the REP3 sequenc- e of Candida Genome Database (CGD) ...40
3.6 The REP3 was disrupted at the proper chromosomal location and the ARG4 or URA3
marker did not integrated into unpredicted chromosome locations ...41
3.7 The REP3 rescued strains (CJC7 and CJC10) did not show a restored drug susceptibility phenotype of rep3/rep3 ...43
3.7.1 The agar dilution assay...43
3.7.2 Broth microdilution method (according to NCCLS standard)...44
3.8 Comparison of the genes expression level of rep3/rep3 mutants and the rescued strains (CJC7 and CJC10) in C- andida albicans ...45
3.8.1 The expression of REP3 in wild type and rescued strains (CJC7 and CJC10) but not in knockout strains (CJC2 and CJC4) ...45
3.8.2 Mutations in REP3 do not decrease the CDR1 expression ...45
3.8.3 The expression level of orf19.3926 increased about 3.5 folds in the REP3 rescued strains than that of wild-type and rep3/rep3 in the presence of miconazole ...46
3.9 Construction of the REP3 rescue plasmid (SATI flipping method)...46
3.9.1 Construction of A fragment into pSFS2 to create CJB1 ~ CJB5 ...46
3.9.2 Construction of CJB6 ~ CJB10: pGEM-T easy vector with B fragment ...47
3.9.3 Comparison of the REP3 rescue plasmid sequence between pWJB24 and pCJB6 ~ pCJB8 ...47
3.9.4 Construction of the plasmid CJB11and CJB15 with A and B fragments on both sides of SAT1 from pSFS2...48
3.10 Construction and confirmung the rep3/rep3::REP3 (SATI flipping method) rescued str- ains (CJC13 ~ CJC18) ...48
3.11 The REP3 rescued strains (SAT1 cassette method) (CJC19 ~ CJC24) can not show a
restored phenotype of drug susceptibility...49
3.11.1 The agar dilution assay...49
3.11.2 Broth microdilution method (according to NCCLS standard)...50
Chapter 4. Discussion...51
4.1 Genomic organization of the REP3 locus ...51
4.2 The phenotype of rescued strains ...51
4.3 The relationship with drug resistance between trans-regulator factor, REP3 and CDR1 .... ...53
4.4 Other uncharacterized protein factors probably bind to elements within the MDR1 prom- oter and trans-activate the MDR1 gene...55
4.5 REP3 is a homolog of CRZ1, a target of the calcineurin pathway, in Candida albicans.56 4.6 Orf19.3926 has high similarity to uncharacterized Candida albicans Rny1p ...58
Chapter 5. Future work ... 61
References ...63
Tables ...73
Figures ...79
Contents (Figures)
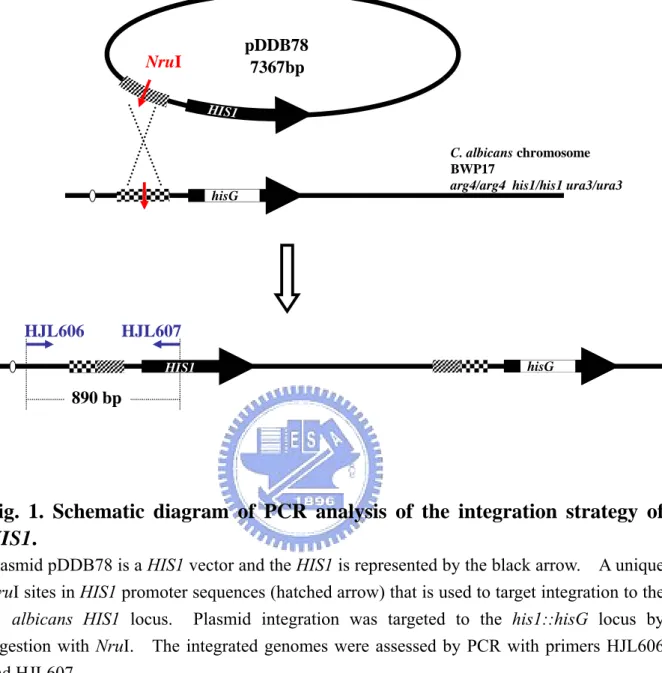
Fig. 1. Schematic diagram of PCR analysis of the integration strategy of HIS1...79 Fig. 2. Assessment of HIS1 integration into rep3/rep3 by PCR...80 Fig. 3. The antifungal drugs susceptibility of histidine prototrophy rep3/rep3 is identical
with histidine auxototrophy rep3/rep3 ...81 Fig. 4. The REP3 rescued strains YLO221 and YLO222 can not restore the drug susceptibil-
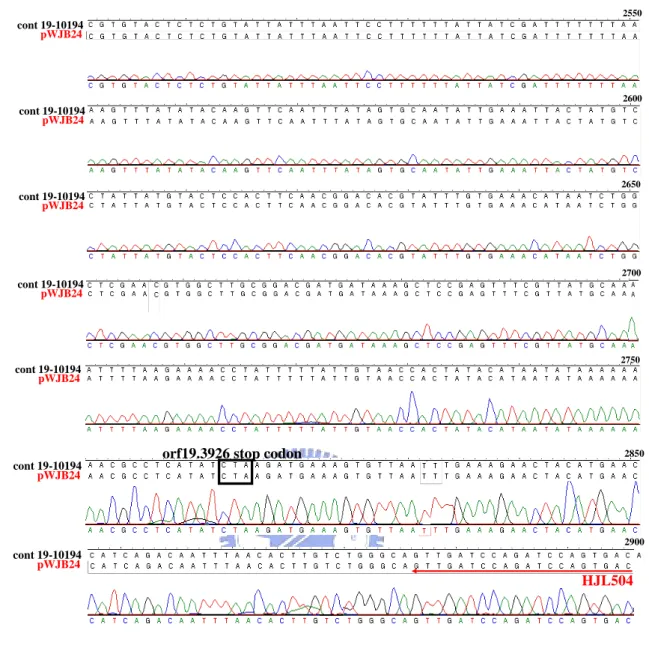
ity phenotype of rep3/rep3 ...82 Fig. 5. The REP3 sequence of the rescue plasmid (pWJB24) is identical to the REP3 sequen- ce of Candida Genome Database (CGD)...88 Fig. 6. Schematic diagram of analysis of integration of REP3 into rescued strains with pWJB
24 by PCR...89 Fig. 7. Assessment of the REP3 gene transformed to rep3/rep3 null mutant strains by PCR
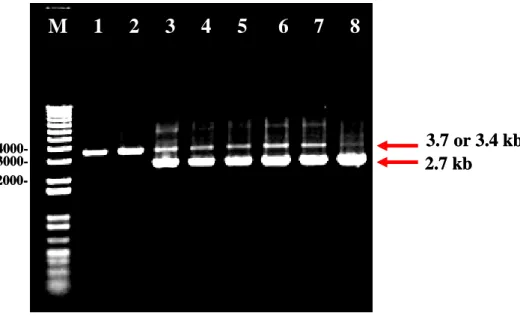
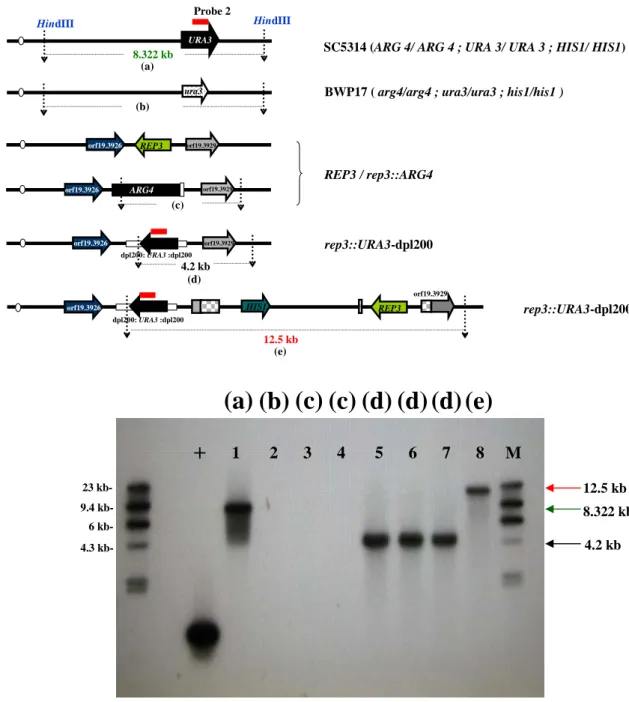
with primers HJL458, HJL505...90 Fig. 8. Southern blot analysis of HindIII-digested genomic DNA of the parent strain BWP17,
mutant and rescued derivatives with ARG4-specific probe 1...91 Fig. 9. Southern blot analysis of HindIII-digested genomic DNA of the parent strain BWP17,
mutant and rescued derivatives with URA3-specific probe 2...92 Fig. 10. Mutations on REP3 increase the susceptibility to antifungal agents but the rescued
strains (CJC7 and CJC10) did not show a restored drug susceptibility phenotype in C.
albicans ...93
Fig. 11. Mutations on rep3 gene of C. albicans increases the susceptibility to miconazole....94 Fig. 12. Mutations on rep3 gene of C. albicans increases the susceptibility to itraconazole...95 Fig. 13. Mutations on rep3 gene of C. albicans increases the susceptibility to 4-NQO...96 Fig. 14. Mutations in REP3 do not decrease the CDR1 expression in C. albicans...97 Fig. 15. The expression of CPH1 and MDR1 in the rep3/rep3 null mutations (CJC2 and CJC4)
and rescued strains (CJC7 and CJC10) in the present of miconazole...98 Fig. 16. The expression of ERG3 and ERG11 in the rep3/rep3 null mutations (CJC2 and CJC4 ) in the present of miconazole...99
Fig. 17. The orf19.3926 in the REP3 rescued strains showed higher mRNA level after the cells were treated with miconazole ...101 Fig. 18. Schematic diagram of construction REP3 rescue plasmid (pCJB11 and pCJB15) by
SAT1 flipper method to transform rep3/rep3 (CJC2 and CJC4) homozygous mutants
... 103 Fig. 19. All the process of construction REP3 rescue plasmid (pCJB11 and pCJB15) by SAT1
flipper method shown below... 104 Fig. 20. Constructing the plasmid pCJB1 ~ pCJB5and confirming the transformants by kpnI
and XhoI digestion ... 105 Fig. 21. Constructing the plasmid pCJB1 ~ pCJB5 and confirming the transformants by ScaI
and SacII digestion ... 106 Fig. 22. Constructing the plasmid pCJB6 ~ pCJB10and confirming the transformants by enz-
yme digested... 107 Fig. 23. Comparison REP3 sequences of cont19-10194 nucleotide sequences with pCJB11,
pCJB7 and pCJB15 ... 109 Fig. 24. Comparison of the REP3 amino acid sequences among pCJB11, pCJB7 and pCJB15 ... 110 Fig. 25. Schematic diagram of construction strategy of pCJB11 and pCJB15...111 Fig. 26. Constructing the plasmid pCJB11 ~ pCJB16and confirming the transformants by en- zyme digestions...113 Fig. 27. Schematic diagram of integration strategy of REP3 by SAT1 flipping method...115 Fig. 28. Assessing the REP3 integration to rep3 null mutant strains and excision of the SAT1
cassette by PCR... 117 Fig. 29. Mutations on REP3 increase the susceptibility to antifungal agents but the rescued
strains (CJC19 ~ CJC24) did not show a restored drug susceptibility phenotype in C.
albicans ... 118
Fig. 30. Comparing the rep3 homozygous mutants and the REP3 rescued strains (CJC19 ~ CJ C 21) by SAT1 flipper method for the susceptibility to miconazole ... 119
Fig. 31. Comparing the rep3 homozygous mutants and the REP3 rescued strains (CJC19 ~ CJ C 21) by SAT1 flipper method for the susceptibility to itraconazole ... 120
Contents (Tables)
Table 1. The recombinant plasmid...73 Table 2. Candida albicans used in this study ...75 Table 3. Primer used in this study...77
Chapter 1. Introduction
1.1 Clinical significance of Candida infection
Candida albicans is a diploid asexual fungus, and a causal agent of opportunistic oral
and genital infections in humans. There are a number of reasons for an increase in fungal infections. Systemic fungal infections have emerged as important causes of morbidity and mortality in immunocompromised patients (for example, AIDS, cancer chemotherapy, organ or bone marrow transplantation). C. albicans is a common commensal yeast that colonizes the gastrointestinal or genital tract of 15 – 30 % of healthy humans.
C. albicans is the fourth leading cause of bloodstream infections in the United States
(Beck-Sague et al., 1993; Pfaller et al., 1998; Edmond et al., 1999). It can cause either benign and frequent infections such as oral and vaginal candidiasis or more serious infections such as life-threatening invasive infections in immunocompromised hosts. Invasive candidiasis mainly occur in hospitalized patients and C. albicans, which is currently a leading cause of nosocomial infections, is responsible for 60% of cases of candidemia. The annual Medicare coasts for treating candidemia estimated to exceed $1 billion in the USA (Miller et al., 2001). Through a simple calculation based on an incidence rate of 8 out of 100,000 every year (Kao et al., 1999), 40% mortality (Edmond et
al., 1999), and 300 million population size, there are almost 10,000 deaths per year due to Candida infections in the USA.
In Taiwan the prevalence of nosocomial candidemia increased 27-fold from 1981 through 1993 at one medical center at National Taiwan University Hospital in Taipei and C.
hospital-related infections in patients not previously considered at risk (for example, patients on an intensive care unit) have become a cause of major health concern.
1.2 Candida albicans is an opportunistic pathogen
C. albicans is among the gut flora, the many organisms that live in the human mouth
and gastrointestinal tract. Under normal circumstances, C. albicans lives in 80% of the human population with no harmful effects, although overgrowth results in candidiasis. It is the causes of mucosal, cutaneous, and systemic infections, including oropharyngeal candidiasis (OPC), the most frequent opportunistic infection among AIDS patients (Klein et
al ., 1984; Feigal et al., 1991).
Candidiasis is often observed in immunocompromised individuals such as HIV-positive patients. HIV-positive individuals and AIDS patients often develop oropharyngeal candidiasis and, in many countries, the triazole antifungals have been the mainstay of their treatment. The widespread use of prolonged fluconazole therapy in the 1990s led to an increased frequency of treatment failure due to fluconazole-resistant C.
albicans (Ruhnke et al., 1994; Boschman et al., 1998). It also may occur in the blood and
in the genital tract. Candidiasis, also known as "thrush", is a common condition that is usually easily cured in people who are not immunocompromised.
Among the different antifungal agents available, the class of azoles has been used extensively during the past 20 years. C. albicans cells repetitively exposed to these antifungals pressure can lead to phenomenon of azole resistance (Coste et al., 2006). Due to the repeated use of this agent especially in human immunodeficiency virus-positive patients with recurrent oropharyngeal candidiasis, treatment failures were observed to be associated with emergence of azole-resistant C. albicans strains (Coste et al., 2004).
1.3 Resistance of C. albicans to antifungal agents
Antifungal agents exert their activity through a variety of mechanisms, some of which are poorly understood. Drug targets that are distinct between pathogens and hosts are more difficult to identify in fungi than in bacteria, at least in part because fungi and animals are relatively close as both are eukaryotes. The available drugs to therapy fungal infection suffer from many drawbacks in term of side effects, efficacy, spectrum, resistance and cost (Sundriyal et al., 2006). Currently, antifungal drugs that are available for treatment of
Candida infections are those against ergosterol and its biosynthesis, nucleic acid synthesis,
and cell wall synthesis (Ghannoum and Rice, 1999; Yang and Lo, 2001). Most of the antifungal drugs are directed against ergosterol, the major sterol of the fungal plasma membrane, which is analogous to cholesterol in mammalian cells. Ergosterol is the main component of the fungal plasma membrane and plays many important roles within the cell (Joseph-Horne and Hollomon, 1997; Morschhauser, 2003; Parks, 1995). It helps maintain membrane integrity and fluidity, as well as ensuring proper function of several membrane-bound enzymes (Lupetti, 2002). Accordingly, many antifungal drug classes target enzymes in the ergosterol biosynthetic pathway. In Candida, mutations or changes in levels of expression of ERG genes can lead to alterations in drug sensitivity (Jensen-Pergakes et al., 1998; Jia et al., 2002; Sanglard et al.,2003; Young et al., 2004). Available antifungal drugs can be divided into four major classes including (1) polyenes (2) ergosterol biosynthesis inhibitors (3) nucleic acid synthesis inhibitor and (4) cell wall synthesis inhibitor.
1. polyenes
The polyenes, such as amphotericin B, are very potent agents acting by binding to the fungal cell membrane and causing the fungus to leak electrolytes. Amphotericin B is the
gold standard for severe, life-threatening systemic fungal infections.
The polyene compounds are so named because of the alternating conjugated double bonds that constitute a part of their macrolide ring structure. The polyene antibiotics are all products of Streptomyces species. These drugs interact with sterols in cell membranes (ergosterol in fungal cells; cholesterol in human cells) to form channels through the membrane, causing the cells to become leaky. The polyene antifungal agents include nystatin, amphotericin B, and pimaricin.
Amphotericin B binds to sterols, preferentially to the primary fungal cell membrane sterol, ergosterol. This binding disrupts osmotic integrity of the fungal membrane, resulting in leakage of intracellular potassium, magnesium, sugars, and metabolites and then cellular death. The mechanism of action is the same for all the preparations and is due to the intrinsic antifungal activity of amphotericin B (Vanden Bossche et al., 1994a).
2. ergosterol biosynthesis inhibitors
Polyenes including azoles are ergosterol biosynthesis inhibitors and specifically inhibit fungal ergosterol biosynthesis. Azoles are antifungal agents that inhibit the cytochrome P450-dependent 14-α-lanosterol demethylase enzyme. Inhibition of this enzyme results in the depletion of ergosterol, a major component needed to maintain the integrity of the fungal cell wall. The azoles are subdivided into two groups: the triazoles (fluconazole, itraconazole and voriconazole) and the imidazoles (clotrimazole, ketoconazole miconazole, econazole, and sulconazole). Fluconazole and voriconazole are the two triazoles that are most widely used to treat C. albicans infections. It inhibits the sterol 14-α-demethylase, which is encoded by ERG11 (Hitchcock, 1991). Inhibition of lanosterol 14-α-demethylase by azoles not only results in ergosterol depletion but also in the accumulation of the methylated sterol, which inhibits cell growth (Kelly et al., 1997). The triazoles replaced earlier drugs like ketoconazole because they have better pharmacokinetics, are safer, and
have better efficacy when treating systemic fungal infections. Imidazoles are chiefly used to treat cutaneous fungal infections and vulvovaginal infections.
3. nucleic acid synthesis inhibitor
5-flucytosine (5-FC) is a synthetic fluorine analog of cytosine. It is deaminated to 5-fluorouracil (5-FU) and then converted to 5-fluorodeoxyuridylic acid monophosphate, a noncompetitive inhibitor of thymidylate synthetase. This interferes with DNA synthesis. It inhibits both DNA and RNA synthesis via intracytoplasmic conversion to 5-fluorouracil. The latter is converted to two active nucleotides: 5-fluorouridine triphosphate, which inhibits RNA processing, and 5-fluorodeoxyuridine monophosphate, which inhibits thymidylate synthetase and hence the formation of the deoxythymidine triphosphate needed for DNA synthesis (White et al., 1998). As with other antimetabolites, the emergence of drug resistance is a problem. Therefore, 5-fluorocytosine is seldom used alone. In combination with amphotericin B it remains the treatment of choice for cryptococcal meningitis and is effective against a number of other mycoses, including some caused by the dematiaceous fungi and perhaps even by C. albicans.
4. cell wall synthesis inhibitor
Although both fungi and humans are eukaryotics in nature, fungal cell wall is significantly unique structure feature. The major component of fungal cell wall is 1, 3-β-glucan. The glucan synthesis inhibitors are, collectively, agents that are presumed to block fungal cell wall synthesis by inhibiting the enzyme 1,3-β-glucan synthase. The lipopeptides papulocandins and echinocandins obtained from fungal kingdom through blocking the 1,3-β-glucan synthase to inhibit fungal cell wall synthesis (Leelaporn et al., 1994; Radding et al., 1998). Inhibition of this enzyme results in depletion of glucan polymers in the fungal cell, resulting in an abnormally weak cell wall unable to withstand
osmotic stress. There are three such agents at present, with all three belonging to the chemical family known as the echinocandins: caspofungin, micafungin, and anidulafungin. Caspofungin (MK-0991; L-743, 872) has antifungal activity against yeasts of the genus
Candida (including isolates resistant to azoles and amphotericin B), several species of
filamentous fungi, including Aspergillus, and certain dimorphic fungi, such as Histoplasma,
Blastomyces and Coccidioides (Letscher-Bru and Herbrecht, 2003). It is fungicidal with
minimal host toxicities and is the only compound in this class approved by US Federal Drug Agency for clinical use for therapy of aspergillosis and disseminated Candida infection (Georgopapadakou, 2001; Tkacz and DiDomenico, 2001; Odds et al., 2003).
1.4 Mechanism of resistance to azole antifungal agents
To date, four resistance mechanisms have been identified in the development of azole resistance in C. albicans: (1) the cellular content of the azole target encoded by the ERG11 gene can be increased; (2) the affinity of Erg11p to azoles can be decreased by mutations in
ERG11; (3) the ergosterol biosynthetic pathway can be altered; and (4) azoles can fail to
accumulate inside the cells. (Sanglard et al., 1998b). In C. albicans, one of the well-documented mechanisms of resistance to azole antifungal agents is the upregulation of multidrug transporter genes (Perea et al., 2001; Sangland et al., 1997; Sangland et al., 1995; White et al., 1997). The upregulation of multidrug transporter genes leads to the enhanced efflux of azoles and therefore results in decreased drug accumulation and reduced inhibition of their target encoded by the ERG11 gene. Among these mechanisms, the last is the most commonly observed in clinical strains developing azole resistance and involves the upregulation of multidrug transporter genes from at least two families, i.e. CDR1 and CDR2 (Candida drug resistance 1 and 2) of the ATP-binding cassette (ABC) transporter family (Prasad et al., 1995; Sanglard et al., 1995; 1997) and CaMDR1 (Candida albicans
MultiDrug Resistance 1) of the major facilitator (MF) superfamily (Sanglard et al., 1995; Franz et al., 1998). The most important mechanism of resistance to azoles is the overexpression of multidrug transporters, encoded by either the major facilitator efflux pump CaMDR1 (MultiDrug Resistance 1) or the ABC transporters CDR1 (Candida Drug Resistance 1) and CDR2. CaMDR1 expression is controlled by at least two regulatory cis-acting promoter regions as reported recently by Harry et al (2005).
1.5 Candida drug resistance gene: CDR1
The Candida drug resistance 1 (CDR1) gene, which encodes an ABC efflux pump, is identified by complementation of the pleiotropic drug resistance 5 (pdr5) mutant, which is hypersensitive to cycloheximide, chloramphenicol, and azole drugs, in Saccharomyces
cerevisiae (S. cerevisiae) (Prasad et al., 1995).
Cdr1 protein (~170kDa) contains two highly hydrophobic transmembrane domains (TMD) and two cytoplasmically localized nucleotide binding domains (NBD). ABC transporter proteins are located in the plasma membrane, or in organelle membranes, of organisms as diverse as Escherichia coli and humans. They are ATP-dependent translocators of a wide variety of small molecules, including many xenobiotics, and typically comprise alternating pairs of cytoplasmic NBDs and membrane embedded TMDs that contain six transmembrane spans (Dean, 2005). All NBD regions contain conserved motifs: Walker A and Walker B also found in other nucleotide-binding proteins and the family defining C-loop or ABC signature motif (LSGGQ). Saccharomyces cerevisiae Pdr5p is the archetype of the fungal pleiotropic drug resistance PDR family of drug transporters. C. albicans Cdr1p and Cdr2p show approximately 70% homology with Pdr5p.
relationships for the primary equences of the Cdr pump proteins of C. albicans. C. albicans cdr1mutant resulted in increasing susceptibilities to azole drugs (Coste et al.,
1996), which is consistent with the observation that overexpression of CDR1 contributes to the drug resistance of clinical isolates of C. albicans (Lopez-Ribot et al., 1999; Yang et al., 2001). However, the molecular mechanism and the gene network regulating the expression of CDR1 and drug resistance are poorly understood.
Understanding the transcriptional control of CDR genes, by both cis- and trans-acting effectors, is therefore important for determining how azole resistance and transport mechanisms are regulated in C. albicans.
1.6 Cis- and trans-acting factors regulating of CDR genes
1.6.1 Cis-regulatory factors of efflux pumps involved in azole resistance
There are two major strategies to identify the important cis-regulatory elements of the efflux pumps involved in drug resistance. (1).Through comparing the promoter sequence of the CDRs and MDR1 of fluconazole-resistant C. albicans strains with the published promoter sequences and that of fluconazole-susceptible strains. The difference among their promoters may cause the mRNA level of efflux pumps increasing in fluconazole-resistant strains. However, like previously mentioned in the ERG11 sequence comparison, such sequence differences may simply reflect allelic variation and by themselves do not prove a causal relationship with resistance. Morschhäuser’s group compared the promoter sequences of both MDR1 alleles of two matched pairs of clinical C.
albicans isolates in which fluconazole resistance correlated with constitutive MDR1
expression and did not reveal promoter mutations that might be responsible for MDR1 activation in the resistant isolates (Wirsching et al., 2000). (2). Using the method of
promoter mutation analysis, for example, deletion or site direct mutation. The expression of the CDR1 gene has been shown to be induced by agents such as miconazole, fluconazole, nystatin, vinblastine or steroid hormones (Krishnamurthy et al., 1998). Using the promoter serial deletion analysis, four activating and four repressing domains were identified along the entire CDR1 promoter. The AP-1 site and the drug-responsive element of the CDR1 promoter have been reported to be the cis-regulatory elements (de Micheli et
al., 2002; Puri et al., 1999). Several elements of CDR genes are important for the
regulation of CDR1 and CDR2. A basal response element (BRE) is located between nt -860 and -810 in the CDR1 promoter, and a drug response element (DRE) is present in the promoters of both CDR1 and CDR2 (de Micheli et al., 2002). Further, the DNase I footprinting revealed four cis-acting regulatory elements (W1:-272/-265, W2:-243/-234, W3:-209/-198 and W4:-167/-160) in the proximal promoter (-289/-34). Their mutations led to modulation of basal promoter activity to varying degrees (W1: 4.5 folds, W2: 0.2 fold, W3: 0.6 fold, and W4: 1.9 folds). A considerable promoter activity enhancement caused by W1 mutation suggests that this region is involved in downregulating basal CDR1 promoter activity by possibly interacting with a repressor protein, and thus this sequence was designated an NRE (negative regulatory element: -272~-265).
Using the NRE sequence as the affinity matrix, a ~55-kDa nuclear protein specifically interacted with the NRE was purified (Gaur et al., 2004). In addition, another BRE (located between -243 and -234) and a negative regulatory element (NRE) located within the -289 region have been reported in CDR1 (Puri et al.1999; Gaur et al. 2004). The BRE regulates basal expression of CDR1 (de Micheli et al., 2002), while the DRE sequence (5,-CGGAA/TATCGGATA-3,)
is crucial for the upregulation of these genes in azole-resistant strains as well as for the transient upregulation of both genes in the presence of different drugs such as oestradiol, progesterone, or fluphenazine in azole-susceptible strains. Finally, in the same gene, Karnani et al. (2004) identified SRE1 and SRE2 (steroid response elements) between -696
and -521.
1.6.2 Trans-regulatory factors of efflux pumps involved in azole resistance
Trans-acting factors regulating CDR1 and CDR2 were reported recently.
1. This factor, CaNDT80, is a homolog to a meiosis specific transcription factor in S.
cerevisiae (Chu et al., 1998). The overexpression of CaNDT80 decreased the
susceptibility of S. cerevisiae to both fluconazole and ketoconazole. According to the results of agar dilution assays, strains with mutations in either CDR1 or CaNDT80 were more susceptible to fluconazole (25μg/ml) and voriconazole (1μg/ml). According to the real-time PCR data, homozygous null mutations in CaNDT80 abolished the induction of CDR1 expression in the presence of miconazole (100μg/ml). Deletion of
CaNDT80 in C. albicans conferred hypersensitivity to azoles and decreased the
inducible expression of CDR1. This result is consistent with the idea that CaNdt80 is an activator of CDR1 (Chen et al., 2004).
2. Tac1 (transcriptional activator of CDR), a transcription factor belonging to the family of zinc-finger proteins with a Zn2Cys6motif (Coste et al. 2004). Tac1p binds to the DRE, which contains two CGG triplets typical of the DNA-binding sites of Zn2Cys6 transcription factors. Tac1p is responsible for transient upregulation of both CDR genes in azole-susceptible strains in the presence of inducers. Interestingly, TAC1 is located close to (within~14 kb) the mating-type-like (MTL) locus. TAC1, a Candida
albicans transcription factor situated near the mating-type locus on chromosome 5, is
necessary for the upregulation of the ABC-transporter genes CDR1 and CDR2, which mediate azole resistance (Coste et al., 2006). Previous studies reported a strong correlation between homozygosity at the mating-type locus and azole resistance in a number of clinical isolates (Rustad et al. 2002).
cerevisiae Cka2p. CKA2 and its homologue CKA1 specify catalytic subunits of
protein kinase CK2. Although cka1 mutations have little effect on fluconazole resistance,CKA1 overexpression suppresses the fluconazole resistance of a cka2 mutant.
The protein phosphatase calcineurin is required for azole tolerance, and it is found that the calcineurin inhibitor cyclosporin reverses fluconazole resistance of cka2 mutants. The CDR1 and CDR2 transcript was expressed at 1.9 fold higher levels in the cka2/cka2 strain than in the CKA2/CKA2 strain. Therefore, the cka2 mutation affects both CDR1 and CDR2 expression. Because a loss of Cka2p causes elevated expression of CDR1,
CDR2, ERG11, MDR1 genes. It maybe because Cka2p functions as a negative
regulator of CDR1 and CDR2 expression (Bruno and Mitchell, 2005).
The regulation of CDR1 involves both the cis element on the promoter region and other genes acting in trans. The study here focuses on trans-acting regulatory factors of
CDR1.
1.7 Previous work in the laboratory
Although trans-acting factors regulating CDR1 were reported recently, the molecular mechanism and the gene network regulating the expression of CDR1 and drug resistance are poorly understood. Other new trans-factors are still yet to be discovered. In this study, as in previous studies, S. cerevisiae was a successfully used model to study C. albicans, despite the differences between these two organisms. The CDR1 promoter for the library screening is from the Ym990348 strain. The activity of the CDR1 promoter from Ym990348 strain is relatively low compared to that of SC5314.
In addition, the screening was performed in the presence of miconazole (100μg/ml).
Under the screening system, four candidate open reading frames (ORFs) have been isolated. These five ORFs were
named REP2, REP3, REP4, REP5, and REP6.
The candidate genes, REP3, has the C2H2 type zinc finger domain, which is one of the
major nucleic acid-binding structures, indicating their potential roles as transcription regulators. It was then analyzed for this functions by mutagenesis using the homologous recombination technique to understand more about the regulatory mechanism of drug resistance in C. albicans (Chin-Sheng Chi, 2004, Master thesis, Institute of Biological Science and Technology, National Chiao Tung University).
1.8 The purpose of the study
According to the results of the Etest and Agar dilution, rep3/rep3 homozygous mutant seems to be more susceptible to azoles (Chin-Sheng Chi, 2004, Master thesis, Institute of Biological Science and Technology, National Chiao Tung University ; Wan-Jen Wu, 2005, Master thesis, Institute of Biological Science and Technology, National Chiao Tung University). However, this result has to be further confirmed by phenotype complementation. In addition, in their study, not all the strains have the same genetic background. They differ in whether HIS allele is present or not.
Hence, this study focuses on construction and characterization of rep3/rep3::REP3 rescued strains. The homozygous knockout strains were rescued with single copy of wild-type REP3 gene. If the change of phenotype is due to the disruption of REP3, the wild type phenotype may be restored when the wild type REP3 gene is knock-in to the original location. Next, the rep3/rep3::REP3 rescued strains were tested with other antifungal drugs to examine the susceptibility to drugs.
Chapter 2. Materials and Methods
2.1 Materials
2.1.1 Strains
1. Escherichia coli (DH5α) Bacteria
2. Candida strains :Table 1
2.1.2 plasmids and primers :
plasmids, and primers used in this study are listed in Table 2 and Table 3, respectively.
2.1.3 Chemicals and reagents
◎ Difco laboratories
Bacto agar (Cat.No.214040) LB agar (Cat.No.24520)
yeast nitrogen base w/o amino acid (Cat.No.291940) LB broth (Cat.No.244620)
YPD broth (Cat.No.242820) ◎ Invitrogen
Agarose (Cat.No.15510-027)
◎ BDH
20XSSC (Cat.No.443527N) ◎ Bio-Rad
50×TAE(Cat.No.161-0773) ◎ JRH BIOSCIENCES
Fetal bovine serum (Cat.No.12003-500M) ◎ Sigma Chemical Co.
Dithiothreitol (DTT) (Cat.No.D9779)
Disodium ethylenediamine-tetraacetate (EDTA) (Cat.No.E-5134) Isoamyl alcohol (Cat.No.81k1250)
Miconazole (Cat.No.M-3512)
Glassbeads (425~600 μm) (Cat.No.G9268-500G) Lithium acetate (CH3COOLi) (Cat.No.L-6883) PolyethyleneGlycol3350 (PEG3350)
L-leucine
Urdine (Cat.No.U-3003)
Histidine (Cat.No.H-8125) histidine (Sigma H-9511) arginine (Sigma A-5131)
uridine (Sigma U-3003)
◎ Merck KGaA, 64271 Darmsdct, Germany
Ammonium acetate (CH3COONH4) (Cat.No.1.01116.0500) Chloroform (Cat.No.1.0244511000)
Dodecyl sulfate sodium sat (SDS) (Cat.No.113760.0100) Dimethyl sulfoxide (DMSO) (Cat.No.S26740)
Ethanol (Cat.No.K33534874) Glucose (Cat.No.K33069537)
Isopropanol (Cat.No.K32632434)
Potassium chloride (KCl) (Cat.No.K24252236) Sodium acetate (Cat.No.1.06268.0250)
Sodium carbonate (Na2CO3) (Cat.No.A375692) Sodium hydroxide (NaOH) (Cat.No.B886298)
Tris (hydroxymethyl) aminomethane hydrogen chloride (Tris-HCl) (Cat.No.8382T006) Sodium chloride (NaCl) (Cat.No.K29779304) Maleic acid (Cat.No.S27857)
Tween 20 (Cat.No.P-1379) Triton X-100 (Cat.No.K23841503) ◎ USB
Glycerol (Cat.No.US16374)
phenol : chloroform : Isoamyl Alcohol (Cat.No.US75831)
2.1.4 Buffers
◎ TE buffer
10 mM Tris-HCl (pH 8.0), 1 mM EDTA (pH 8.0) ◎ 10X TAE buffer
0.4 M Tris, 0.4 M glacial acetic acid, 10 mM EDTA ◎ 50% PEG3350
200 g polyethyleneglycol3350 added ddH2O to 400 ml.
◎ 10X PCR buffer
100 mM Tris-HCl, 500 mM KCl, 15 mM MgCl2 (pH 8.3)
◎ Breaking buffer
◎ Maleic acid buffer
0.1 M maleic acid, 0.15 M NaCl, 0.3% Tween 20 (pH 7.5) ◎ Blocking soiution
1%(W/V) blocking reagent dissolved in maleic acid buffer ◎ Detection buffer 0.1 M Tris-HCl, 0.1M NaCl (pH 9.5) ◎ Denaturation Solution 0.5 M NaCl, 1.5M NaCl ◎ Neutralization Solution 1.5 M NaCl, 0.5 M Tris-HCl (pH 7.5) 2.1.5 Medium ◎ LB (Luria-Bertni) broth
1% tryptone, 0.5% yeast extract, 1% NaCl ◎ LB / ampicillin broth
1% tryptone, 0.5% yeast extract, 1% NaCl, 50 g/ml ampicillin ◎ LB / ampicillin agar
1% tryptone, 0.5% yeast extract, 1% NaCl, 1.5% agar, 50 g/ml ampicillin ◎ YPD/ Uridine broth
2% Bacto-peptone, 1% yeast extract, 2% dextrose, 80 mg/l Uridine ◎ YPD/Uridine agar
2% Bacto-peptone, 1% yeast extract, 2% dextrose, 2% agar, 80 mg/l Uridine ◎ SD (Synthetic Dextrose) / Histidine broth
0.67% Bacto-yeast nitrogen base w/o amino acid, 2% dextrose, 20 mg/l Histidine-HCl
◎ SD agar
0.67% Bacto-yeast nitrogen base w/o amino acid, 2% dextrose, 2% agar, 20 mg/l Histidine-HCl
◎ SD/ Histidine broth
0.67% Bacto-yeast nitrogen base w/o amino acid, 2% dextrose, 20 mg/l Histidine-HCl
◎ SD/ Histidine agar
0.67% Bacto-yeast nitrogen base w/o amino acid, 2% dextrose, 2% agar, 20 mg/l Histidine-HCl
2.1.6 Enzymes
◎ NEB
AflII (R0520S), BamHI (R0136S), EcoRI (R0101S), EcoRV (R0195S), HindIII
(R0104S), NruI (R0192S), KpnI (R0142S), NotI (R0189S), SacII (R0157S), SacI (R0156S), XhoI (R0146S)
CIP (M0290S)
◎ Amersham Biosciences
rTaq DNA polymerase (catalog no. 27-0799-06)
2.1.7 Equipments
Peltier Thermal Cycler PTC-200 (MJ Research) Orbital shaking incubator OSI500 (TKS) AlaImager 2000 (Molecular devices) Vortex-2 genie (Scientific Industry)
Dry bath incubator (Violet Bioscience Inc.)
Brushless microcentrifuge Denville 260D (Scientific Industry) Power pac 300 (Bio-Rad)
Elektroporator Gene pulser II (Bio-Rad) Spectra max plus (Molecular devices) Quick spin (Violet Bioscience Inc.)
Thelco laboratory incubator (Precision Scientific Inc.) High speed refrigerated centrifuges J2-MC (Beckman) High speed refrigerated centrifuges (Heraeus)
Centrifuge (SORVALL RT7)
Mettler AT261 DeltaRange (METTLER TOLEDO) Mettler GG4002-S (METTLER TOLEDO)
TurboBlotter system (Schleicsher & Schuell) Digitale camera COOLPIX 990 (Nikon) Spectrophotometer
pH meter (HANHA instruments) Hot plates / Stirrers (CORNING) Water bath (CHERNG HUEI Co.) UV crosslinker (Stratagene)
2.1.8 Strains and growth media
Escherichia coli DH5α was used as the host strain for all plasmid preparations, and
was grown in Luria-Bertani (LB) medium. All constructed bacteria were grown in Luria-Bertani (LB) medium with or without antibiotics.
al., 1984) and its derivative BWP17 (ura3∆::imm434/ura3∆::imm434 his1::hisG /his1::hisG
arg4::hisG/arg4::hisG) (Wilson et al., 1999). Yeast Peptone Dextrose, (pH6.7) was
adjusting by 5 N NaOH, (YPD, 1% yeast extract, 2% peptone, and 2% dextrose and Synthetic Dextrose (SD, 0.67% yeast nitrogen base without amino acid and 2% dextrose) supplemented with necessary auxotrophic requirements [20 mg of each histidine, and arginine or 80 mg uridine per liter] were prepared as described in Sherman et al (Coste et al., 2004; Sherman, 2002). The pH of SD (pH5.4) was adjusted by 5 N NaOH.
2.2 Methods
2.2.1 DNA methods
All DNA manipulations were performed by standard procedures (Joseph Sambrook and David W.Russell, 1989). Kits from Qiagen, Inc. were used to isolate plasmids (catalog no. 12143, 12125, 27104, 20021) and to purify DNA fragments (catalog no. 28106, 28706) according to the manufacturer’s protocols. Enzymes for digestion, ligation, and dephosphorylation of DNA were from Gibco BRL, BioLabs and Roche Molecular Biochemicals. Taq DNA polymerase (AmershamBiosciences, catalog no. 27-0799-06) and Vent DNA polymerase (BioLabs, catalog no. M0254S) for PCR were purchased from Amersham Pharmacia Biotech and BioLabs, respectively.
2.2.2 Preparing electrocompetent cells
Escherichia coli DH5α was precultured in a tube containing 5 ml of LB broth at 37ºC
overnight. 5 ml overnight culture of E.coli. DH5α was inoculated into 1 liter LB. Cells were grown at 37ºC with shaking to an OD600 of 0.4 ~ 0.6 (about 3 ~ 4 h). The flask along
with the cells were chilled on ice for 15 ~ 30 min. It is essential to keep the cells cold from this point onwards. The cells were transfered to 4 chilled 500 ml centrifuge bottles on ice and spun at 5 K rpm (Beckman J2-MC) for 5 min at 4℃. The supernatant was decanted gently and pellets were resuspended in 200 ml of chilled water per centrifuge bottle (800 ml in total). Then the cells were spun at 5 K rpm (Beckman J2-MC) for 5 min at 4℃. The supernatant was decanted gently. The cells were resuspend in 50 ml of chilled water per centrifuge bottle (200 ml in total). Then the cells were spun at 5 K rpm (Beckman J2-MC) for 5 min at 4℃. The supernatant was decanted gently. The cells were resuspend in 10 ml of chilled 10% of glycerol per centrifuge bottle. Then the cells were transferred cells into 50 ml centrifuge tube and spun at 5 K rpm (Beckman J2-MC) for 5 min at 4℃. The supernatant was decanted gently. The cells were resuspend in 2 ml of chilled 10% of glycerol. 80 μl of cells were used per transformation. The cells can store at –70ºC freezer.
2.2.3 E. coli. transformation with electroporation
Frozen cells were thawed on ice. 80 μl of cells were mixed with 2 μl of transforming DNA in a chilled 1.5 ml microcentrifuge tube. The Bio-Rad Gene Pulse was setted at 25 μF and 2.45 kV. Pulse controller was set to 200 Ω.The cell/DNA mixture were transferred to a chilled 0.2 cm electroporation cuvette and the suspension were shacked to the bottom of the cuvette. The moisture on the outside of the cuvette was wiped off with a tissue and the cuvette was put in the slide, which was pushed into the chamber until the cuvette was seated between the contacts in the base of the chamber. Two red buttons on the machine were pushed on the left at the same time (pulse once and heard the beep). The cuvette was removed from the chamber and 1 ml of SOC broth was added immediately before the resuspension of the cells. The cells were transferred to a 1.5 ml centrifuge tube and
incubated at 37℃ for 1 hr with shaking. Cells were spun for 5 min at 3 K rpm (SORVALL RT7) and the supernatant was poured off (remained medium approximately 100 μl) before the cells were plated on LB contained ampicillin (100 μg/ml) plates after suspension.
2.2.4 Competent cell prepared using Calcium chloride
A single colony from a plate freshly grown was picked and inoculated in 5 ml LB broth for 16 ~ 20 hr. The cells transferred into 100ml LB (1:100) broth and incubated the culture for about 3 hr at 37℃ with 180 rpm till the cells reached OD600 = 0.4 ~ 0.6. The cells
transferred to 50 ml sterile tubes. The culture store on ice for 10 min. The cells were spun 4 K rpm (SORVALL RT7) for 10 min at 4℃. Then the media were decanted and the tubes was inverted for 1 min to allow the last traces of media to drain away. Each pellet was resuspended in 10 ml of ice-cold 0.1 M CaCl2 and stored on ice for 15 min. The cells
were spun at 4 K rpm (SORVALL RT7) for 10 min at 4℃. The media was decanted. And pellets were resuspended in 2 ml of ice-cold 0.1 M CaCl2 for each 50 ml of original
culture. The cells were stored as 200 μl aliquots at -70 ℃.
2.2.5 E. coli. transformation using Calcium chloride prepared competent cells
One 200 μl aliquots of competent cells was taken from -70℃ and thawed on ice. Then the DNA (no more than 50 ng in a volume of 10 μl or less) was added into each tube. The tube was mixed and swirling gently. The cells with DNA were stored on ice for 30 min. The tubes (containing mixture of cells and DNA) were left at 42℃ for 2 min. The tubes were transferred rapidly to ice for 2min. Then 800 μl of SOC broth was added into each tube. The cultures were incubated for 90 min at 37℃ to allow the bacteria to recover and express the antibiotic resistance marker encoded by the plasmid. The cultures were
spun 5 K rpm (SORVALL RT7) for 5 min at room temperature. The supernatant was removed about 800 μl. The cells were resuspended and used up to 200 μl of transformed competent cells per 90-mm plate. Appropriate antibiotics were added to the plate; if blue/white selection is used, plates were coated with X-Gal. Then the plates were left at room temperature, until the liquid has been absorbed. The plates were inverted and incubated at 37℃. The transformants should appear in 12 ~ 16 h.
2.2.6 Constructions of plasmids and strains
The plasmids and primers used in this study are listed in Table II and Table III.
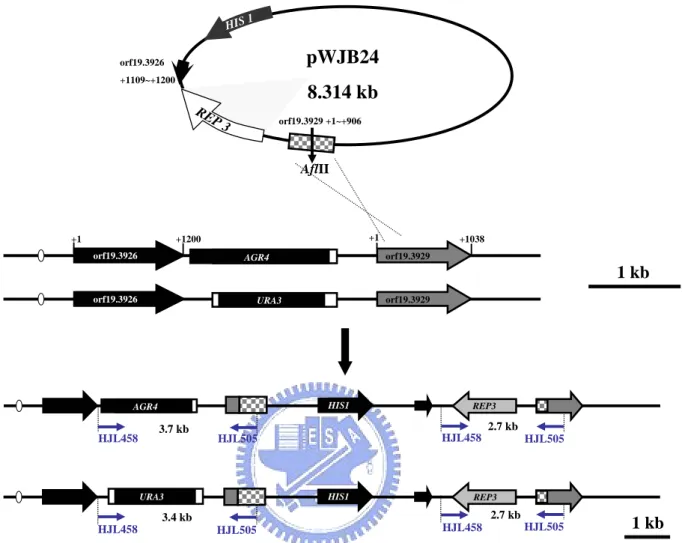
(1). pWJB24: (constructed by Wan-Jan Wu, 2005, Master thesis, Institute of Biological Science and Technology, National Chiao Tung University) pGEM-REP3-
CaHIS1—The C. albicans SC5314 genomic DNA was used as a template for PCR
amplification using the primers HJL504 and HJL505, both containing a BamHI restriction siteat the 5' end. The amplified 2.849 kb fragment started upstreamto the predicted 906 bps of the orf19.3929 start codon and 91 bps downstream to the predicted orf19.3926 stop codon (TAA). After BamHI digestion, the 2.849 kb
fragment was introduced in a sense orientationinto a binary vector (pGEM-CaHIS1) to create pWJB24. To rescue REP3 function in the rep3/rep3 his1::hisG/his1::hisG mutant strains (CSC80 and CSC81), pWJB24 digested with AflII at 310 bps upstream of the translation initiation site of the orf19.3929 was transformed into CSC80 and CSC81 to generate CJC7 ~ CJC9 and CJC10 ~ CJC12, respectively..
(2). pCJB1: A region-pSFS2-CaSAT1—A 201 bps fragment amplified by primers HJL930 (kpnI) and HJL931 (XhoI) from rep3::ARG4/rep3::URA3-dpl200 C. albicans genomic DNA. The PCR amplified program was 94℃ for 3 min followed by 30 cycles of 94 ℃ for 1 min, 55℃ for 1 min and 72℃ for 1 min 20 sec. After a final extension at
72℃ for 1 min then stop reaction at 4℃. The amplified fragment contains the sequences of dpl200:URA3:dpl200 from -558 ~ -721 and the sequence of REP3 from +130 ~ +218 (Fig.18-B). After kpnI and XhoI digestion, the 201 bps fragment was ligated to a binary vector (pSFS2-CaSAT) to create pCJB1 ~ pCJB5.
(3). pCJB6 ~ pCJB10: pGEM-T- B fragment (REP3)—The C. albicans SC5314 genomic DNA was used as a template for PCR amplification usingthe primers HJL928 and HJL929, both containing NotI and SacII restriction sitesat the 5' end. The amplified fragment started +1188 of orf19.3926 to +58 of orf19.3929 (Fig.18-B). The PCR product was amplified by template of SC5314 genomic DNA, 1 μl of a 50 μΜ of each primer, 5 μl of 10 X PCR buffer (Amersham Biosciences), 0.6 μl of 25 mM deoxynucleside triphosphates (Amersham Biosciences), 2 μl of 25 mM MgCl2, 39.2 μl
of water and 0.2 μl (~ 0.9 U) of DNA polymerase mixture (Taq/Vent:9/1). The PCR amplified program was 94℃ for 3 min followed by 30 cycles of 94℃ for 1 min, 55 ℃ for 1 min and 72℃ for 3 min 20 sec. After a final extension at 72℃ for 5 min then stop reaction at 4℃. The PCR product was cloned into pGEM-T to grate pCJB6 ~ pCJB10.
(4). pCJB11 and pCJB15: A fragment-pSFS2-CaSAT1-B fragment (REP3)—The DNA fragment containing full length REP3 purified from pCJB6 and pCJB8 was ligated with the pCJB1 plasmid (A region-pSFS2-CaSAT1) at NotI site to produce pCJB11 and pCJB15.
2.2.7 Complementation of the REP3 gene in rep3/rep3 mutant strains (CSC80 and CSC81)
To verify that the mutant phenotypes were indeed caused by deletion of the target gene, a functional REP3 copy was reinserted into the CSC80 and CSC81 (rep3/rep3 his1::
hisG/his1:: hisG).
Figure 6 showed the genome map of reintegrating the REP3 gene into REP3 knockout strains (CSC80 and CSC81). pWJB24 was digested by AflII which restricted at the +310 of orf19.3929. Then it was transformed into rep3/rep3 strains and the reaction mixture was transformed on SD medium.
The rep3/rep3 rescued strains (CJC7 ~ CJC12) were assessed by PCR with primers HJL458 and HJL505 (Fig.7).
2.2.8 Complementation of the REP3 gene in rep3/rep3 his1::hisG/his1::hisG::HIS1 mutants (CJC2 and CJC4) by SAT1 method (Morschha et al.,2004)
The kpnI-SacII fragments from pCJB11 (pCJB15) containing A and B fragment. B fragment is the complete open reading frame as well as 509 bps of upstream and 279 bps of downstream flanking sequences of REP3 alleles. A fragment includes from -558 ~ -721 of dpl200:URA3:dpl200 to the sequence +130 ~ +218 of REP3. The B fragment from pCJB6 ~ pCJB8 was constructed into pCJB1 in respectively to generate plasmids pCJB11 ~ pCJB13 and pCJB14 ~ pCJB16.
Before transformation, the REP3 rescue plasmids, pCJB11 and pCJB15, (pSFS2 plasmids with both A and B fragments) were restricted by kpnI-SacII in respectively to generate the 6 kb fragment (Fig.18-C) were purified using QIAprep gel extraction Kit (QIAGEN). Generally, 2 ~ 5 μg of gel purified product was suspended in 10 μl of sterile distilled water and was used for one transformation. C. albicans strains were transformed by electroporation as described previously (Kohler et al., 1997), with slight modifications. The kpnI-SacII fragment from pCJB11 and pCJB15 was transformed into strain rep3/rep3
his1::hisG/his1::hisG::HIS1 (CJC2) in respectively to generate CJC13 ~ CJC15. pCJB11
his1::hisG/his1::hisG::HIS1 (CJC4) was integrated the kpnI-SacII fragment from pCJB11
and pCJB15 in respectively to generate CJC16 ~ CJC18. pCJB11 is generated CJC16; pCJB15 is generated CJC17 and CJC18. For the selection of nourseothricin-resistant (NouR) transformants, 200 μg/ml of nourseothricin was added to YPD agar plates. Cells were then spread on YPD plates containing 200 μg/ml of nourseothricin and grown for 2 days at 30 ℃ . The integration of the SAT1 cassette into the rep3/rep3
his1::hisG/his1::hisG::HIS1 genome is selected in the presence of nourseothricin.
First, using the HJL855 and HJL856 primer set, the REP3 integration of rep3/rep3 chromosome will give a 923 bps DNA fragment, when the chromosome carrying rep3::
dpl200-URA3-dpl200-SAT1-REP3. Then were given these strains celled CJC13 ~ CJC15,
of which the parent is CJC2. And the CJC14 ~ CJC15 were obtained by the integration of pCJB15 and the CJC13 was of pCJB11. The strains of CJC16 ~ CJC18 (parent is CJC4); CJC17 ~ CJC18 were obtained by integration of pCJB15 and the CJC16 was of pCJB11.
2.2.9 The REP3 rescued strains (CJC13 ~ CJC18) excise the SAT1 flipping (Fig 28)
In this work, FLP-mediated marker used a new dominant selection marker, caSAT1, conferring resistance to nourseothricin. The REP3 rescued strains (CJC13 ~ CJC18) containing the SAT1 cassette integrated into the rep3/rep3 his1::hisG/his1::hisG::HIS1 genome are selected in the presence of nourseothricin. Originally, I intended to use the
MAL2 promoter to induce caFLP expression by growing the transformants in medium
containing maltose instead of glucose (Backen et al., 2000; Sanchez-Martinez and Perez-Martin, 2002).
FLP-mediated excision of the SAT1 flipper cassette is achieved by simply growing the transformants for a few days in medium with 2% maltose without selective pressure, and nourseothricin-sensitive (NouS) derivatives can easily be identified. After about seven
days, screening for NouS derivatives in which the SAT1 flipper cassette were excised by FLP-mediated recombination. To pick up the single colony from (CJC13 ~ CJC18) after treated with 2% maltose can not grow on YPD plates containing 200 μg/ml nourseothricin to generate CJC19 ~ CJC24.
The CJC19 ~ CJC24 strains were generated and analyzed by PCR with HJL458-HJL133, HJL963-HJL964 and HJL855-HJL856 primers. To confirm the results of excision the
SAT1 flipper in the REP3 rescued strains (CJC19 ~ CJC24) by PCR with primers
HJL963-HJL964. If the SAT1 flipper was excised, the results would be generated a 935 bps fragment (Fig.28-C). Primers of HJL458-HJL133 is positive control the rep3::ARG4
/rep3::URA3 allele would be generated a 1.2 kb fragment (Fig. 28-B). The reintegration of REP3 in CJC19 ~ CJC24 was confirmed by PCR with HJL855 and HJL856 that generated a
923 bps fragment (Fig.28-D). As result shown in Fig.28, I have been got the REP3 rescued strains without SAT1 cassette (CJC19 ~ CJC24).
2.2.10 Isolation of genomic DNA
An isolated single colony was picked and inoculated at 6 ml YPD or YPD + uridine (80 mg/l) broth in 15 ml centrifuge tube. The cells were shaken at about 180 rpm and incubated at 30℃ for 24 ~ 48 h. Cells were spun at 3 K rpm (SORVALL RT7) for 10 minutes, the supernatant was poured off and the cell pellets were washed with 5 ml ddH2O.
Cells were spun at 3,000 rpm (SORVALL RT7) for 10 min, the supernatant was poured off and the cell pellets were resuspended with 500 μl DNA breaking buffer by vortexing and then acid-washed glass beads (~ 400 μl) and 500 μl of phenol/choroform/isoamyl alcohol (25:24:1) were added. The mixtures were vortexed at highest speed for 10 min at room temperature. 500 μl TE was added and the mixtures were vortexes for a few seconds.
Cells were spun at 3 K rpm (SORVALL RT7) for 15 min at room temperature (RT) and aqueous layer were transferred to a new 15 ml centrifuge tube.
Then about 850 μl of (equal volume) phenol/choroform/isoamyl alcohol (25:24:1) were added and the mixtures were vortexed for 60 sec. Phenol extract steps were repeated 3 times then aqueous layer was transferred to 2 new 1.5 ml eppendorf tube (~ 400 μl /tube). 1 ml (2.5 X volume) of cold 100% ETOH was added and the solution was mixed by inversion. The solution were spun at 13 K rpm (Brushless microcentrifuge Denville 260D, Scientific Industry) for 10 min at 4℃ , the supernatant was discarded carefully. The DNA pellet was dried in the air about 10 ~ 15 min. The pellets were resuspended in 0.4 ml of TE buffer with 3 μl of 10 mg/ml RNase A. They were mixed and incubated for ~ 15 min at 37℃. Then, 40 μl of 3 M ammonium acetate (pH 5.2) and 1 ml of cold 100% ETOH were added and mixed by inversion. They were spun at 13 K rpm (Brushless microcentrifuge Denville 260D, Scientific Industry) for 10 min at 4℃ and the supernatant was discarded carefully followed by adding 1ml of cold 70% ETOH to wash DNA pellet. They were spun at 13 K rpm (Brushless microcentrifuge Denville 260D, Scientific Industry) for 5 min at 4℃. The supernatant was removed carefully. The DNA pellet was dried in the air about 15 min and then resuspended in 100 ~ 200 μl of ddH2O.
2.2.11 Transformation of Candida albicans by lithium acetate method
The protocol for yeast transformation was modified from the previous report (Gietz et
al., 1995). C. albicans cells were grown overnight in YPD/Uri medium on a shaker at 30
℃ to stationary phase. The culture was diluted into fresh YPD/Uri medium in 1/100 ratio (OD600 of about 0.1) and was incubated at 30℃ with shaking (about 150 rpm) for an
additional 6 h (OD600 of about 1.0 ~ 0.7). Cells were centrifuged at 3000 rpm in
(pH 7.5), and lithium acetate/TE buffer [100 mM lithium acetate (Sigma L6883), 10 mM Tris-HCl, 1 mM EDTA], sequentially. The washed cell pellet from 10 ml of original culture was resuspended in 0.1 ml lithium acetate/TE buffer for one transformation reaction. Transformation mixtures contained 5 10 µg DNA, 10 µl of 10 mg/ml of boiled Salmon Sperm DNA (sperm DNA was boiled in a PCR machine for 2 min and then it was kept on ice right away), and 100 µl of C. albicans cells. The transformation mixture was incubated at room temperature for 30 min. To this mixture, 700 µl of PEG/lithium acetate/TE buffer [40% polyethyleneglycol 3350PEG (Sigma P4338), 100 mM lithium acetate, 10 mM Tris-HCl, 1 mM EDTA ] was added and briefly vortexed. Subsequently, the transformation mixture was incubated for overnight at 30°C, followed by a heat shock of 60 min at 42°C. Cells were pelleted, washed once with TE buffer (pH 7.5) and plated onto selective media and incubated at 30°C for at least 2 ~ 3 days.
2.2.12 Transformation of Candida albicans by electroporation method
C. albicans strains were transformed by electroporation as described previously
(Kohler et al., 1997), with slight modifications. Cells from a YPD preculture were diluted 10-4 in 50 ml fresh YPD medium and grown overnight at 30℃ to an optical density at 600
nm (OD600) of 1.6 ~ 2.2. The cells were collected by centrifugation and resuspended in 8
ml of water. After addition of 1 ml of 10 ×TE (100 mM Tris–HCl, 10 mM EDTA, pH 7.5) and 1 ml of 1 M lithium acetate (Sigma-Aldrich Chemie, Steinheim, Germany), pH 7.5, the suspension was incubated in a rotary shaker at 150 rpm for 60 min at 30℃. A 250 μl volume of 1 M dithiothreitol was then added, and the cells were incubated for a further 30 min at 30℃ with shaking. After addition of 40 ml of water the cells were centrifuged, washed sequentially in 25 ml of ice-cold water and 5 ml of ice-cold 1 M sorbitol, resuspended in 50 μl of 1M sorbitol, and kept on ice.
The inserts from plasmids were purified by agarose gel electrophoresis and elution with the QIAprep gel extraction kit (Cat. NO 28706). Five microliters (approximately 1 μg) of the linear DNA fragments was mixed with 40 μl of electrocompetent cells, and electroporation was carried out in a 0.2 cm electroporation cuvette, 1.8 kV.
After electroporation, the cells were washed in 1 ml of 1 M sorbitol, resuspended in 1 ml YPD medium, and incubated for 4 h with shaking at 30℃. The cells were spread on YPD plates containing 100 μg/ml of 4 h nourseothricin and grown at 30℃. Resistant colonies were picked after 1 day of growth and inoculated in YPD liquid medium containing 100 μg/ml of nourseothricin for DNA isolation. In parallel, the transformants were streaked on YPD plates with 100 μg/ml of nourseothricin for further use.
2.2.13 Southern blot analysis
Southern blot analysis was carried out with the DIG DNA Labeling and Detection Kit (Roche, catalog no. 1 093 657) according to the manufacturer’s recommendations. Briefly, probe1 is the 634 bps of PCR fragment of ARG4 from plasmid pRSARG4△Spe1 (LOB16) which was amplified by primer HJL311 and HJL312. And probe2 is the 613 bps of PCR fragment of URA3 from plasmid pGEM-URA3 (LOB15) which was amplified by primer HJL632 and HJL633. The probes are DIG labeled by randompriming (PCR DIG Probe Synthesis Kit. Cat. No. 11 636 090 910), using a 1:3 ratio of DIG-dUTP:dTTP according to the length of the probe 1 (634 bps)、probe 2 (613 bps). The PCR amplified program was 95℃ for 2 min followed by 10 cycles of 95℃ for 30 sec, 58℃ for 30 sec and 72℃ for 40 sec followed by 20 cycles of 95℃ for 30 sec, 58℃ for 30 sec and 72℃ for 1 min in a Peltier Thermal Cycler-200 (MJ Research, Watertown, MA). After a final extension at 72 ℃ for 4 min then the reaction was stopped at 4℃.
and CSC22)、rep3::ARG4/rep3::URA3 (CSC80 and CSC81) and rep3/ rep3::REP3 (CJC7 and CJC10) were digested with HindIII and size-fractionated by 0.8% agarose gel electrophoresis on Sub-Cell GT system (BioRad, catalog no. 170-4404). The 634 bps the
ARG4 fragment amplified by primers HJL311 and HJL312 was used as a positive control.
Another positive control URA3 was amplified by HJL631 and HJL633.
The DNAs were transferred by capillarity (TurboBlotter Rapid Downward Transfer system, Schleicher & Schuell, Item no. 10416328) onto Nytran SuPerCharge membrane (Schleicher & Schuell, Item no. 10416296). On UV crosslinker (Stratagene, catalog no. 400075), DNA was immobilized on membrane by pressing two times of autocrosslink mode (120mJ/ pulse).
Appropriate volume of DIG Easy Hyb (approx.20 ml/100 cm2) was pre-heated to hybridization temperature 42℃. The blot was incubated for 30 min with gentle agitation. The DIG-labeled DNA probe (20 ng/ml for DNA-probe) was boiled for 10 min for denaturation and was rapidly cooled on ice. It was then added to pre-heated DIG Easy Hyb (at least 3.5 ml/100 cm2 membrane) and mixed well. Prehybridization solution was then discarded and immediately, the probe/DIG Easy Hyb mixture was added to membrane. The hybridization mixture was incublated with gentle agitation at 42℃ overnight.
The membrane was washed as recommended by the manufacturer under high-stringency conditions with twice of 2 X wash solution (2 X SSC, 0.1% SDS) at room temperature for 5 min, and twice of 0.5 X wash solution (0.5 X SSC, 0.1% SDS) at 65℃ for 15 min. Afterhybridization and stringency washes, the membrane was rinsed briefly 5 min in Washing buffer. Then it was incubated for 30 min in 100 ml Blocking solution. And then it was incubated for 30 min in 20 ml Antibody solution. Next, it was washed 2*15 min in 100 ml Wahing buffer. After equilibrated for 2 ~ 5 min in 20 ml Dectection buffer, the membrane was placed with DNA side facing up on a development folder (or hybridization bag) and 2 ml diluted CSPD solution was applied. The membrance was