行政院國家科學委員會專題研究計畫 期中進度報告
玉葉金花花部器官決定基因之功能與蛋白質交互作用研究
(1/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2311-B-002-024- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學生態學與演化生物學研究所 計畫主持人: 胡哲明 共同主持人: 賴爾 計畫參與人員: 徐馨怡,張世白,黃俊溢 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 6 月 6 日
一、中英文摘要
毛玉葉金花(Mussaenda pubescens)是台灣低海拔中常見的植物之一。玉葉金花
屬植物常可以見到花萼其中的一個裂片尖端膨大成一個外形類似葉子、但顏色為白色 或紅色的構造,稱為葉狀萼片(calycophylls, 或 floral semaphylls)。而玉葉金花名稱 的由來,即源於它的花序中黃色的花瓣和白色的葉狀萼片而來。這些葉狀萼片不管在 細胞形狀或細胞排列上,都與正常的花瓣類似,加上有著顯著的顏色,一般皆認為這 些葉狀萼片在功能上與花瓣無異,我們稱之為類花瓣(petaloid)的構造。我們在這個三 年期的研究計畫中,期望藉由花部基因的選殖與功能性的研究,希望瞭解有關花瓣與 類花瓣調控的各類因子。在第一年的研究中,我們順利的取得了毛玉葉金花中一個 A 群,三個 B 群,以及一個 E 群基因的全長序列,並構建了 Yeast-2-hybrid 所需的各個 clones。另外也完成毛玉葉金花的各部器官的 RNA 萃取,俾後續的 cDNA library 建 置,以及 RNA 表現分析。
關鍵詞:玉葉金花,花部器官決定基因,蛋白質交互作用
Abstract
Mussaenda pubescens is a plant with enlarged and petaloid calyx lobe, named
calycophylls, they are phenotypically similar to true petals in having bright color and papilliate epidermal cells. We proposed the petaloid structure were formed by the ectopic expression of genes that specifying petal organ identity, but our preliminary data suggested that there might be unknown factors involved other than floral ABE genes. In the first year of the three-year project, we have successfully identify the full-length sequences of one A, three B, and one E class genes. We have also successfully constructed clones for yeast-2-hybrids. The RNAs were extracted and ready for cDNA library construction and expression analysis.
Keywords: Mussaenda pubescens, floral organ identity genes, protein-protein interactions
二、前言與研究目的
The majority of flowering plants uses petals as the attractant agent for luring pollinators. However, there are many examples that use non-petal organs to mimic showy petals, and thus called "petaloid structures". In order to elucidate the possible genetic
mechanism for the formation of petaloid structures, we selected Mussaenda pubescens as our working system. M. pubescens has an enlarged and petaloid sepals named "calycophylls" on the peripheral flowers of a inflorescens. The calycophylls have vivid white color and loose mesophylls, and have papilliate epidermis, all typical features of a true petal. Based on the current floral genetic model, petals are determined by combination of three classes of genes, A (e.g. AP1 and AP2), B (e.g. PI and AP3), and E (SEPs). In Arabidopsis studies,
overexpression a combination of AP1, AP3, PI, and SEP showed an interesting phenotype that converts all its leaves into petals (Honma and Goto, 2001). Theses results suggest that, petaloid structures could be regulated by ectopic expression of petal identity genes, as proposed by several authors (Bowman, 1997; Albert et al., 1998; Baum, 1998; Baum and
Whitlock, 1999). The project's rationale is to apply this model to the petaloidy regulation in
Mussaenda pubescens.
In our previous survey, we have successfully identified A, B, and E class genes from
M. pubescens, namely MupAP1 (A class homologue); MupDEF, MupGLO, MupTM6 (all B
class homologues); and MuSEP (E class homologue). The expression patterns based on RT-PCR results suggest that B and E class genes are expressed in the normal sepals as well as calycophylls. The only exception is MupGLO, its transcript is not detected in calycophylls and very weak in normal sepals. The complex expression pattern is intriguing and indicates that the formation of calycophylls might correlate with MupDEF and MupTM6 expression, both these two genes are homologues to AP3 of Arabidopsis. These floral gene products needed to form a multimer complex in order to function properly, and different functions are likely carried out by combinations of different proteins (Theiβen and Saedler, 2001, Theiβen, 2001). The preliminary results prompted us to propose a thorough screening of candidate proteins that interact with MupDEF, MupTM6 and/or MupSEP, in order to identify the unknown factors that are responsible for the petaloid structure.
三、研究方法
(1) Sample collections and RNA isolation
Plant materials of Mussaenda pubescens have been collected from the populations at Shu-Mei-Pin (Taipei Co.) and Kenting (Pingtung Co.). Total RNAs were isolated from different organs of Mussaenda pubescens by Pine Tree method (Chang et al. 1993).
(2) Full-length open reading frame of cDNAs
Since previously identified ABE genes were obtained through 3' RACE (Rapid Amplification of cDNA Ends, Invitrogen, Life technologies, Carlsbad, CA, USA), specific primers were designed at K domain region of previously identified ABE genes for 5' RACE (Rapid Amplification of cDNA Ends, Invitrogen, Life technologies, Carlsbad, CA, USA) reaction in order to obtain 5' region of the genes. The 5' RACE products were then cloned into pGEM®-T vector (Promega, Madison, WI, USA), and determined the sequences. New specific primers at the 5' end of the genes were then designed to amplify full-length cDNAs in conjugated with 3' specific primers for PCR. The products were then cloned again into pGEM®-T vector (Promega, Madison, WI, USA), and determined the sequences.
(3) Construction of clones for yeast-2-hybrid
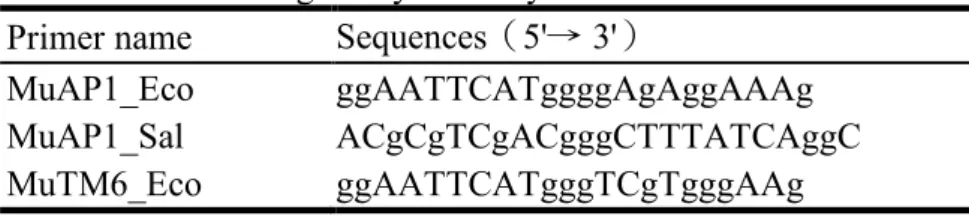
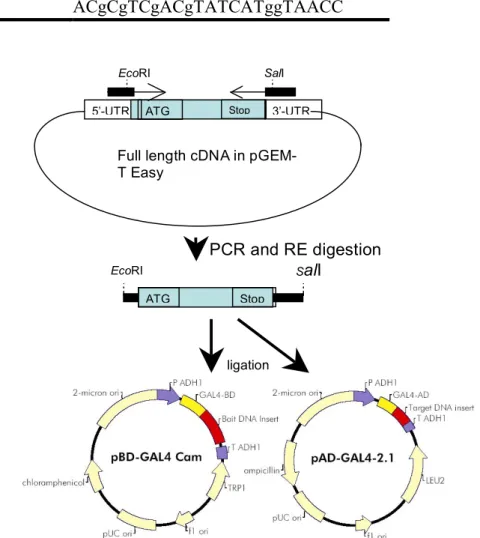
The cDNAs were constructed into the lambda vector using the HybriZAP 2.1® two-hybrid system, into pAD-GAL4 and pBD-GAL4 following the manufactory's suggestions (Stratagene, La Jolla, CA, USA). New primers were designed to include a EcoRI or SalI overhead at their 5' (Table 1 and Fig. 1) for amplifying open reading frame of cDNAs, and then cloned into pAD or pBD vectors and transformed into E. coli. Colony PCR were used to confirm the identity of these clones.
Table 1. Primer design for yeast-2-hybrid constructs Primer name Sequences(5'→ 3')
MuAP1_Eco ggAATTCATggggAgAggAAAg
MuAP1_Sal ACgCgTCgACgggCTTTATCAggC
MuTM6_Sal ACgCgTCgACCATgCAATAggATg MuGLO_Eco ggAATTCATggggAgAggTAAg MuGLO_Sal ACgCgTCgACgTTCTTgggTAgAgC MuDEF_Eco ggAATTCATggCTCgTgggAAg MuDEF_Sal ACgCgTCgACgTAAATAAATTTGCTAC MuSEP_Eco ggAATTCATgggAAgAggTAgg MuSEP_Sal ACgCgTCgACgTATCATggTAACC
Fig. 1. Cloning strategy for yeast-2-hybrid constructs.
四、研究結果與討論
(1) Full length cDNA construction
We have obtained full-length cDNA sequences of all five genes (1 A, 3 B, and 1 C class genes), and have them cloned into pGEM-T vectors. No new homologues were found during 5' RACE or full-length cDNA PCR amplification after examining the sequence alignments.
(2) Clone constructs for yeast-2-hybrid
We have obtained full-length cDNA clones of all five genes in either pAD-GAL4 or
5’-UTR ATG Stop 3’-UTR
Full length cDNA in pGEM-T Easy EcoRI PCR and RE digestion ATG Stop EcoRI SalI SalI ligation
pBD-GAL4 vectors, except for MuDEF (Table 2). The sequence of the MuDEF clone contained a stop codon, and was different from cDNA sequences we had, therefore, needed to be re-amplified. The sequences of all of the clones were examined to prevent any errors at every cloning step. We are designing another sets of clones that only include IKC domains of these genes, i.e. excluding MADS-box domain. This is to eliminate a possible auto-regulation effect of these constructs (Elena Kramer, personal communication).
Table 2. A check-list for the obtained constructs. Checked marks () are correct clones we had. Triangles are not finished cloning, whereas the cross marks mean a wrong sequence was obtained as explained in the text.
Insert Transformation Ligation & Colony PCR extraction Plasmid Sequence
AD-MuAP1 BD-MuAP1 AD-MuDEF Δ Δ Δ Δ BD-MuDEF Δ Δ Δ Δ AD-MuGLO BD-MuGLO AD-MuTM6 BD-MuTM6 AD-MuSEP BD-MuSEP 五、參考文獻
Albert, V. A., M. H. G. Gustafsson, and L. D. Laurenzio. 1998. Ontogenetic systematics, molecular developmental genetics, and the angiosperm petal. Pages 349-374 in Molecular systematics of plants II: DNA sequencing (D. E. Soltis, P. S. Soltis, and J. J. Doyle, eds.). Kluwer Academic Publishers, Boston.
Baum, D. A. 1998. The evolution of plant development. Current Opinion in Plant Biology 1: 79-86.
Baum, D. A., and B. A. Whitlock. 1999. Plant development: Genetic clues to petal evolution.
Current Biology 9: R525-R527.
Bowman, J. L. 1997. Evolutionary conservation of angiosperm flower development at the molecular and genetic levels. Journal of Biosciences 22: 515-527.
Chang, S., J. Puryear, and J. Chirney. 1993. A simple and efficient method for isolating RNA from pine trees. Plant Molecular Biology Reporter 11: 113-116.
Honma, T. and K. Goto. 2001. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 409: 525-529.
Theiβen, G. 2001. Development of floral organ identity: stories from the MADS house.
Current Opinion in Plant Biology 4: 75-85.