行政院國家科學委員會專題研究計畫 成果報告
血管內皮細胞發炎反應的細胞與分子機制--在發炎物刺激
下血管內皮細胞中剪流所扮演之保護角色: 剪流對干擾素
引發 Stat1 訊息傳導及相關基因表現之調控(3/3)
研究成果報告(完整版)
計 畫 類 別 : 整合型 計 畫 編 號 : NSC 95-2320-B-002-052- 執 行 期 間 : 95 年 08 月 01 日至 96 年 10 月 31 日 執 行 單 位 : 國立臺灣大學化學工程學系暨研究所 計 畫 主 持 人 : 謝學真 計畫參與人員: 博士班研究生-兼任助理:陳柏仰 碩士班研究生-兼任助理:張弘勳 其他:蔡宇致 處 理 方 式 : 本計畫可公開查詢中 華 民 國 97 年 01 月 07 日
行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告
□期中進度報告
整合型總計畫名稱:血管內皮細胞發炎反應的細胞與分子機制
本計畫名稱:在發炎物刺激下血管內皮細胞中剪流所扮演之保護角色:
剪流對干擾素引發Stat1訊息傳導及相關基因表現之調控
(Protective role of shear flow in inflammatory agent-stimulated vascular endothelial cells: Regulation of interferon-induced Stat1 signaling and related gene expression by shear flow)計畫類別:□ 個別型計畫 ■ 整合型計畫
計畫編號:
NSC93-2321-B-002-034, NSC94-2321-B-002-016, NSC95-2320-B-002-052執行期間:93 年 8 月 1 日至 96 年 10 月 31 日
計畫主持人:
謝學真
整合型總計畫主持人:王 寧
計畫參與人員:
蔡宇致、謝宗佑、吳畹儀、王安姬、張弘勳等
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列管計
畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:
國立台灣大學 化學工程學系暨研究所
中 華 民 國 96 年 12 月 31 日
國科會專題研究計畫完整報告
整合型總計畫名稱:血管內皮細胞發炎反應的細胞與分子機制
本計畫名稱:在發炎物刺激下血管內皮細胞中剪流所扮演之保護角色:剪流對干
擾素引發Stat1訊息傳導及相關基因表現之調控
(Protective role of shear flow in inflammatory agent-stimulated vascular endothelial cells: Regulation of interferon-induced Stat1 signaling and related gene expression by shear flow)
計畫編號:NSC93-2321-B-002-034, NSC94-2321-B-002-016, NSC95-2320-B-002-052 執行期間:93 年 8 月 1 日至 96 年 10 月 31 日 計畫主持人:謝學真 整合型總計畫主持人:王 寧 計畫參與人員:蔡宇致、謝宗佑、吳畹儀、王安姬、張弘勳等 執行單位:國立台灣大學 化學工程學系暨研究所 一、摘要 動脈粥狀硬化是一種慢性疾病,而發炎 反應在動脈硬化的病程中扮演相當重要的角 色 。 發 炎 過 程 中 包 括 細 胞 素 如 interferon-gamma (IFN-γ) 會刺激內皮細胞, 誘導發炎基因如CXC chemokines 的表現,其 中需要 JAK/STAT1 訊息傳導路徑參與。由 於血管壁的內皮細胞(ECs)持續受到血液流 動所產生的剪力(shear stress,簡寫 SS)刺激, 穩定的層流(laminar flow,簡寫 LF)剪力作用 能保護內皮細胞,抑制動脈粥狀硬化。本研 究欲探討在發炎反應中重要的細胞素IFNγ刺 激下,層流剪力作用是否能扮演抑制發炎反 應的調控角色。以細胞素IFNγ刺激內皮細胞
會活化 JAK1/2 及 STAT1,造成 JAK1/2 及 STAT1 的酪氨酸(Tyr701)、絲氨酸(Ser727) 磷酸化增加。Tyr701 磷酸化在 30 分鐘達到 最高點,而 Ser727 磷酸化在 30 分鐘後才開 始明顯上升。然而,剪力會抑制IFNγ所引發 的JAK1/2 及 Tyr701 磷酸化,隨著剪力施予 的時間增加,抑制的效果也越明顯。而隨著 剪力大小的增加,抑制 IFNγ引發的 Tyr701 磷酸化,其效果也越明顯,但並不會影響 Ser727 磷酸化的程度。再進一步探討剪力抑 制STAT1 磷酸化的機制,發現 protein tyrosine phosphatase (PTP)參與剪力的抑制效果。IFNγ 的刺激,會經由JAK/STAT1 訊息傳導路徑, 引發下游發炎基因的表現。本研究以Taqman real-time PCR 方法測定各發炎基因之 mRNA 的含量,發現以IFNγ刺激內皮細胞,會引發 化 學 趨 化 蛋 白 CXC chemokines (CXCL9, CXCL10, CXCL11)和 CIITA 的 mRNA 大量 表現。再以 siRNA 去除細胞中的 STAT1 蛋 白質,發現 IFNγ所造成的 CXC chemokines 和CIITA 的 mRNA 表現會被顯著地降低,說 明以IFNγ刺激內皮細胞,會經由活化 STAT1 訊息傳導路徑,引發下游 CXC chemokines 和CIITA 的表現。血液流動所產生的剪力能 抑 制 IFNγ 所 引 發 的 上 述 三 種 CXC
chemokines mRNA 表現,也能大量降低 IFNγ
所引發的 CXCL10 蛋白質表現,以及 CIITA mRNA 表現。另外,不同的剪力大小可對 CXC chemokines mRNA 有不同程度的抑 制,對內皮細胞施予低剪力(2 dyn/cm2),IFNγ 所引發的CXC chemokines mRNA 表現會被 部分抑制下來,而高剪力(25 dyn/cm2)則幾乎 完全抑制CXC chemokines mRNA 表現。綜 言之,本研究證實了層流剪力作用可抑制 IFNγ引發的 JAK/STAT1 訊息傳導路徑,進 而抑制 STAT1 下游與發炎反應相關的 CXC chemokines 和 CIITA 的基因表現,因而可望 對於血管壁的內皮細胞產生保護效果,抑制 動脈粥狀硬化。 關 鍵 詞 : 剪 力 , 層 流 , 內 皮 細 胞 ,CXC chemokines, interferon-gamma, STAT1
Abstract
It is known that inflammatory response plays an important role in atherogenesis. Interferon-gamma (IFNγ) is an inflammation-related cytokine that can stimulate endothelial cells (ECs) to express CXC chemokines via JAK/STAT1 signaling pathway. ECs are constantly exposed to blood flow-induced shear stress (SS). Laminar flow (LF)-associated shear stress protects ECs from endothelial dysfunction and thus is
atheroprotective. This study was conducted to analyze whether ECs under laminar shear stress
were protected from IFNγ-induced inflammatory responses. IFNγ triggered the activation of JAK/STAT1 signaling pathway with the phosphorylation of JAK1/2 and phosphorylation of Tyr701 and Ser727 in STAT1. The Tyr701 phosphorylation approached the highest level at 30 minutes and Ser727 phosphorylation obviously raised after 30 minutes. IFNγ induced Tyr701 and Ser727 phosphorylation in a dose-dependent manner, with their highest levels occurring at the IFNγ concentration of 2.5 ng/ml. When SS was applied to IFNγ-treated ECs, it significantly inhibited the IFNγ-induced Tyr701 phosphorylation of STAT1 in a shear force- and time course-dependent manner whiles Ser727 phosphorylation was unaffected. In the presence of sodium orthovanadate (an inhibitor of protein tyrosine phosphatase), SS was unable to suppress IFN-γ-induced STAT1 Tyr-701 phosphorylation, indicating that protein tyrosine phosphatase (PTP) was involved in the suppression process. Next, we performed Taqman real-time PCR to detect the mRNAs of downstream target genes such as CXC chemokines and CIITA in the JAK/STAT1 signaling pathway. IFN-γ (2.5 ng/ml) stimulation dramatically increased the mRNA levels of CXC chemokines (CXCL9, CXCL10 , and CXCL11) and CIITA. To examine the atheroprotective effects of laminar flow on ECs, IFNγ-treated ECs were subjected to LF for 4 hours. The expression of IFNγ-inducible CXC chemokines was significantly attenuated. Consistently, LF greatly suppressed CXCL10 secretion into a cultured medium. In addition to CXC chemokines, the IFNγ-induced expression of CIITA, a crucial transcriptional
regulator of MHCII expression, was also attenuated by LF. We further transfected ECs with STAT1-targeting siRNA to reduce the amount of STAT1 protein and found that the expressions of the IFNγ-induced CXC chemokines and CIITA were significantly suppressed. This result revealed that CXC chemokines and CIITA are the downstream target genes of STAT1. We also found that LF suppressed IFNγ-induced CXC chemokines expression in a shear stress-dependent manner, with
high shear stress (25 dyn/cm2) being more effective
than low shear stress (2 dyn/cm2) in suppressing
CXC chemokines expression. In summary, LF suppressed IFNγ-induced JAK/STAT1 signaling and downstream inflammation-related target genes including CXC chemokines and CIITA, thereby protecting ECs from dysfunction and suppressing atherogenesis.
Keywords: shear stress, laminar flow, endothelial
cells, CXC chemokines, interferon-gamma,
STAT1 二、計畫緣由與目的 目前已有眾多研究指出動脈粥狀硬化與 血流動力學有關,本研究室之前的研究發現 穩定的層流會導致內皮細胞 reactive oxygen species (ROS)的增加,此外,層流也會抑制 monocyte chemotactic protein-1 (MCP-1)基因 的表現而有保護效果[1]。然而在身體血管內 的轉彎分支處往往是粥狀斑塊容易堆積的地 方,而此處的血液流動較為緩慢或伴隨著擾 流的發生。動脈粥狀硬化是一種慢性疾病, 從初期的內皮細胞功能異常發展到形成粥狀 班塊進而導致血栓,發炎反應在整個惡化的 過程扮演相當重要的角色。許多文獻指出 interferon-gamma (IFNγ)與動脈硬化有關, IFNγ會引發內皮細胞發炎相關的基因表現, 如 CXC chemokines 和 major
histocompatibility complex class II (MHCII),
讓 T cell 更容易沾黏在內皮細胞上,並穿過 內皮細胞層引發後續的發炎反應[2,3]。而 CXC chemokines 中 CXCL9 (Mig), CXCL10 (IP-10), CXCL11 (I-TAC)已被發現會在動脈 硬化斑塊處大量表現。另一方面,細胞素也 會引發內皮細胞中的訊息傳導,例如 IFNγ
訊息傳導路徑[4],轉錄因子 Stat1、Stat3 可 調控細胞中之發炎反應、生長及死亡[5,6]。 本研究室之前的研究指出,剪流可抑制內皮 細胞中由血清或 IL-6 誘導的 JAK/STAT3 活 化[7,8]。根據這些發現,本研究欲探討在發 炎反應中重要的細胞素IFNγ刺激下,層流剪 力作用是否能扮演抑制發炎反應的調控角 色。 三、實驗方法 3.1 細胞株與細胞培養 本 實 驗 採 用 的 細 胞 為 bovine aortic endothelial cells(BAEC),以 DMEM (含 10 %FBS、penicillin、streptomycin)培養。從臍 帶 初 級 培 養 取 下 的 human umbilical vein endothelial cells (HUVEC),以 M199 (含 20% FBS、penicillin、streptomycin)培養,並置於 於37℃、5%CO2含飽和濕度的培養箱中。繼 代 培 養 時 , 以 緩 衝 液 洗 過 , 再 用 1X trypsin-EDTA 將細胞取下,離心收集,移去 上清液,加入上述培養基使細胞懸浮分散, 再接種到附有gelatin 的玻片上。 3.2 剪流實驗 將培養細胞的培養基裝入玻璃瓶內層, 並且通入含 5%CO2的空氣,以保持培養基之 pH 值為 7.4。外層以恆溫水流通,保持溫度 為 37℃。開啟轉動式幫浦(roller pump), 輸送培養基至波動調節器(pulse dampener) 以消除流體的脈動,並且阻止氣泡進入流動 室。待調節器內液面達適當高度時,將調節 器平放。此時培養基便經由塑膠管流回瓶 中,而成為一循環狀態之流動系統。整個系 統以恆溫水域槽控溫,保持流動系統溫度為 37℃。將長滿單層細胞的玻璃片裝置在設計 好 之 平 行 板 流 動 室 (parallel plate flow chamber),連接上流動管路,調節幫浦轉速 至所需流量後,在恆溫下開始流動實驗。 3.3 細 胞 內 蛋 白 質 含 量 測 定 ( 西 方 墨 點 Western blot) 經實驗處理後的細胞以 cord buffer 清洗 過之後,吸乾殘留之液體,加入 lysis buffer
(RIPA buffer,with proteinase inhibitor、0.1%
SDS),充分溶解細胞後,將 sample 離心取上 清液。以BCA 蛋白質定量液於 530 nm 之吸 光值來計算總細胞之蛋白質含量。取同量之 蛋白質並調成同體積,加入還原劑,於95℃ 加熱五分鐘,之後以8~10 % polyacrylamide gel 進行電泳約二到三個小時,取出凝膠,將 蛋白質轉移到 nitrocellulose 膜片上。然後以 含5%脫脂奶粉及 0.2% Tween-20 的 TBS 溶 液(TBS-T)浸泡膜片一小時,然後依序再以欲 定量之單株或多株抗體浸泡搖晃二小時,然 後再以 TBS-T 洗三次,每次五分鐘,倒掉 TBS-T 後加入含二次抗體之 TBS-T 浸泡搖晃 一小時,然後以TBS-T 洗三次,每次五分鐘, 最 後 則 以 Western-Star Chemiluminescence Detection System 來 測 量 其 強 度 , 使 用 CDP-Star chemiluminescence substrate 當受 質。
3.4 萃取細胞 RNA 與 Northern blot
經實驗處理後的細胞以 cord buffer 清洗 過之後,吸乾殘留之液體,加入4M guanidine thiocyanate 溶 液 溶 解 細 胞 , 以 phenol-chloroform 有 機 溶 液 萃 取 細 胞 中 之 total RNA , 再 以 酒 精 沈 澱 的 方 式 分 離 出 RNA,並且溶於 DEPC 處理過的 H2O 中。 由細胞中所分離出之RNA (10 µg/lane) 將以 1 % MOPS-formaldehyde agarose gel,在 1 x MOPS 溶液中進行電泳 (100 V) 約 2.5 小 時,以vacuum bloting 方式將 gel 上之 RNA 轉移到nylon membrane 膜片上,並以 UV 光
交聯使RNA 固定在膜片上。將膜片浸泡在預
雜交溶液中(42℃) 16~20 小時,再將膜片與
32P-labeled IP-10 cDNA probe 雜交 (42℃) 24
小時,雜交完之膜片在室溫下以wash buffer (1x SSC,1% SDS) 清洗 2~3 次,每次 30 分 鐘,清洗完的膜片置於-70℃,以 x-ray film 感光2~3 天。 3.5 轉 錄 因 子 -DNA 複 合 體 的 偵 測 (gel shifting) 將 1~2 ng 的特定序列(32P-end-labeled high-affinity sis-inducible element (hSIE) oligonucleotide probe)與 5 µg 的核蛋白質反應 30 分鐘,將反應後的樣品以 5% nondenaturing polyacrylamide gel 進行電泳約三個小時,再
以真空乾燥的方式轉印到3M membrane 上, 以autoradiography 方式,用 X-ray film 偵測 membrane 上之放射線訊號。
3.6 萃取細胞 RNA 與 Taqman real-time PCR assay 經實驗處理後的細胞以 cord buffer 清洗 過之後,吸乾殘留之液體,加入4M guanidine thiocyanate 溶 液 溶 解 細 胞 , 以 phenol-chloroform 有 機 溶 液 萃 取 細 胞 中 之 total RNA , 再 以 酒 精 沈 澱 的 方 式 分 離 出 RNA,並且溶於 DEPC 處理過的 H2O 中。
將100 ng 的 total RNA 加入 Taqman one step RT-PCR master mix solution (Applied Biosystems)中,並加入所欲偵測基因之探針 (委請瑞林公司設計並製作),一管的總反應體 積為20 µl,進行 RT and real-time PCR 之反 應。RT 的反應條件為 48℃30 分鐘,緊接著 95℃10 分鐘開啟 DNA polymerase 的活性, 接著40 個 PCR cycles (95℃15 秒,60℃1 分 鐘),由 ABI 7700 real-time PCR 機器記錄每 一個cycle 的螢光訊號並分析所得的結果。每 一個基因的 mRNA 含量除以 GAPDH 的 mRNA 含量,來標準化每一管的結果。 四、實驗結果與討論 4.1 剪 力 不 影 響 內 皮 細 胞 在 基 礎 狀 態 下 STAT1 的活性 STAT1 是一種可被 IFNγ引發的細胞內重 要的訊息轉錄因子,與細胞的生長,凋亡和 發炎反應有關,STAT1 的 Tyr701 及 Ser727 磷酸化程度與其活性有關,本研究首先要探 討剪力是否會影響 STAT1 的活化狀態。如 Fig. 1 所示,實驗發現對未受細胞素(cytokine) 刺激的內皮細胞施予剪力(25 dyn/cm2)之情 況下,並不會引起STAT1 酪氨酸(Tyr701)的 磷酸化,也不會改變 STAT1 絲氨酸(Ser727) 已有的磷酸化。以2.5 ng/ml IFNγ刺激內皮細 胞30 分鐘後,可以看到 Tyr701 及 Ser727 磷 酸化增加。同時並觀察剪力對ERK1/2 及 Akt 之調控,剪力刺激會造成 ERK1/2 磷酸化在 10 分鐘達到最高點,並在兩小時後回到基礎 狀態下的磷酸化程度,同時並觀察到 Akt 受 到剪力的刺激之後也會產生短時間的磷酸化 增加,ERK1/2 及 Akt 的結果與其他報導所觀 察到的現象一致。 4.2 IFNγ刺激內皮細胞中 STAT1 之活化 當內皮細胞受到細胞素的影響後,會產 生一連串的訊息傳導過程以調控下游的基 因,引發內皮細胞異常並導致發炎反應,IFNγ 是由 T cell 所釋放出來的一種干擾素,會活 化 JAK/STAT1 的訊息傳導路徑,活化的 STAT1 會與其目標 DNA 結合並促進目標基 因的轉錄,最後導致目標基因的表現。如Fig. 2A 所示,本實驗以細胞素 IFNγ (2.5 ng/ml) 刺激內皮細胞,IFNγ會活化 STAT1,造成 STAT1 的 Tyr701、Ser727 磷酸化增加,加入 IFNγ刺激內皮細胞之後,Tyr701 磷酸化在 5 分鐘就開始上升,並在30 分鐘達到最高點, 而 Ser727 磷酸化在 30 分鐘後才開始明顯上 升。如Fig. 2B 所示,以不同濃度的 IFNγ刺 激內皮細胞30 分鐘,Tyr701、Ser727 磷酸化 程度會隨著 IFNγ濃度的增加而上升,在 2.5 ng/ml 高濃度 IFNγ的刺激之下,Tyr701、 Ser727 磷酸化程度達到最高。 4.3 剪力會抑制由 IFNγ所引發的 JAK1/2 及 STAT1 Tyr701 磷酸化 本研究室先前的研究發現穩定的層流會 抑制MCP-1 基因的表現而有保護效果[1],本 研究欲觀察剪力對受到發炎反應因子IFNγ刺 激的內皮細胞是否同樣具有保護作用。如Fig. 3A 所示,對臍帶靜脈內皮細胞(HUVEC)施 予剪力,會抑制 IFNγ所引發的 Tyr701 磷酸 化,在預處理2.5 ng/ml IFNγ 10 分鐘之後, 再繼續施予剪力(25 dyn/cm2) 10 或 20 分鐘, 與靜止狀態下加入IFNγ的細胞相比,有施予 剪力的內皮細胞其 Tyr701 磷酸化會被抑 制,隨著剪力施予的時間增加,抑制的效果 也越明顯。如Fig. 3B 所示,由 IFNγ所引發 的Tyr701 磷酸化,隨著剪力大小的增加,抑 制效果也越明顯,施予5 dyn/cm2大小的剪力 並不影響 Tyr701 磷酸化程度,當施予 12.5 dyn/cm2 剪力,Tyr701 磷酸化會有比較明顯 的抑制效果,增加剪力大小到 25 dyn/cm2 時,抑制效果最明顯,與 IFNγ所引發的 Tyr 701 磷酸化程度相比,約可抑制 50%,另一
方面,剪力並不會對Ser727 磷酸化有影響。 而在牛的動脈內皮細胞(BAEC)也可以看到
剪力的保護效果,如Fig. 4 所示,剪力也同
樣會抑制IFNγ所引發的 STAT1 Tyr701 磷酸
化,並且也抑制 IFNγ所引發的 JAK1/2 磷酸 化。由上述實驗可以證明剪力對於兩種來源 不同的內皮細胞,都具有抑制 JAK/STAT1 訊息傳導活化的效果。 4.4 IFNγ透過活化 STAT1,引發內皮細胞 中化學趨化蛋白 CXCL10 (IP-10)的表現 IFN-γ 會 引 發 內 皮 細 胞 中 CXC chemokines、MHCII 基因的表現,這些基因 也都透過 JAK/STAT1 訊息傳導路徑的調 控,其表現出之蛋白則會刺激內皮細胞導致 後 續 的 發 炎 反 應 。 本 研 究 欲 探 討 CXC chemokines 對 內 皮 細 胞 的 影 響 , CXC chemokine 家 族 中 包 括 CXCL9 (Mig), CXCL10 (IP-10), CXCL11 (I-TAC)這三種,已 有報導指出在動脈硬化的斑塊表面,內皮細 胞會表現大量的CXC chemokine 並且會吸引 許多活化的 T cell 沾黏到班塊表面並轉移到 斑塊的組織裡面,並在此處放出許多細胞 素,擴大發炎反應[8]。如 Fig. 5 所示, IFNγ (2.5 ng/ml)會刺激內皮細胞,使 IP-10 mRNA 的表現量隨刺激時間增長而上升,在 刺激 4 小時後達到最高點。由之前所得的結 果 , 證 實 剪 力 會 抑 制 IFNγ 所 引 發 的 JAK/STAT1 訊息傳導路徑,未來我們將探 討,剪力是否同樣會抑制IFNγ所引發的下游 目 標 基 因 群 表 現 , 如 CXC chemokines 、 MHCII,以達到保護內皮細胞免於細胞素刺 激的效果。
4.5 Protein tyrosine phosphatase 參與層流 (LF) 剪 力 作 用 抑 制 IFNγ 所 引 發 STAT1 Tyr701 磷酸化 STAT1 屬於一種可被 IFN-γ引發磷酸化 的細胞內重要的訊息轉錄因子,與細胞的生 長,凋亡和發炎反應有關。之前的結果顯示 層流(LF)剪力作用會抑制 STAT1 Tyr701 的 磷酸化。本研究欲探討LF 所產生的剪力抑制 STAT1 Tyr701 磷酸化的作用機制為何。如 Fig. 6 所示,比較 lane 3 和 lane 5,對 BAECs 施予剪力,剪力會抑制 IFNγ (2.5 ng/ml)所引
發的STAT1 Tyr701 磷酸化。比較 lane 5 和 lane 6,以 100 µM Na3VO4(protein tyrosine
phosphatase 抑制劑)預先處理 30 分鐘後,再 施予LF (25 dyne/cm2)20 分鐘之後,LF 抑制 IFNγ所引發 STAT1 Tyr701 磷酸化的效果就 會消失,表示protein tyrosine phosphatase 參 與LF 對 STAT1 Tyr701 去磷酸化的過程。 4.6 層流(LF)剪力作用會抑制由 IFN-γ所引 發的 STAT1-DNA 結合
IFNγ會促使內皮細胞中 SΤΑΤ1磷酸化, 形成STAT1/1 homodimer,此 homodimer 再
進入細胞核,與核內 DNA promoter 上的特
定序列(hSIE)進行結合,,以啟動下游相關基 因的表現。如Fig. 7 所示,lane 1 表示在基礎 狀況的HUVECs 並沒有產生 DNA binding 的 現象。lane 2 表示以 IFNγ (2.5 ng/ml)刺激細胞 4小時之後,引發 DNA binding,比較 lane 2 和 lane 3,當施予剪力 4 小時於 IFNγ-induced 的
內皮細胞,發現剪力會明顯地抑制 STAT1/1
的DNA binding。在 lane 4 和 lane 5,分別使 用 anti-STAT1 和 anti-STAT3 抗體加入到核
蛋白和DNA 結合反應的溶液中,發現當加入
anti-STAT1 抗體之後,DNA binding 的現象
就消失,據此確認核蛋白中與 DNA (hSIE) binding 的分子就是 STAT1。 4.7 IFNγ經由活化 JAK/STAT1 訊息傳導路 徑,增加 CXC chemokines 和 CIITA 的表現 已有報導指出,IFN-γ會刺激內皮細胞, 引發許多發炎相關基因如 CXC chemokines 和CIITA (MHCII 基因轉錄的重要調控因子) 的表現。本實驗欲觀察層流(LF)對上述基因 表現的影響,以Taqman real-time PCR 的方 式分析細胞中基因的表現量。如Fig. 8 所示, 以 IFNγ (2.5 ng/ml)刺激 HUVECs,在刺激 1,2,4,6 小時後,觀察不同時間點的基因表 現,在基礎狀況未受刺激的狀態下,內皮細 胞 CXC chemokines (CXCL9, CXCL10, CXCL11)和 CIITA 的表現量非常低,當受到 IFNγ刺激 1 小時之後,這些基因開始大量表 現,隨著刺激的時間增長,這些基因的表現 量也會慢慢升高。為確認這些受到IFNγ刺激 而表現的基因是透過 STAT1 的活化而表 現,本研究以siRNA 技術去除內皮細胞中的
STAT1 蛋白質,如 Fig. 9 下半圖所示,lane 3 和lane 4 表示,HUVECs 中的 STAT1 蛋白質
受到siRNA STAT1 的影響而被去除掉,但細 胞中 STAT3 和 tubulin 蛋白質的含量並沒有 改變,由此證明siRNA STAT1 可以專一性地 去除細胞中 STAT1 而不影響其它蛋白質含 量 。 如 Fig. 9 上 半 圖 中 所 示 , 經 過 STAT1-silenced 內皮細胞,再以 IFNγ (2.5 ng/ml) 處理 4 小時後,觀察 STAT1 下游基因 的表現,發現CXC chemokines 和 CIITA 的 mRNA 表現都會被明顯地抑制下來,證明內 皮細胞當受到 IFNγ刺激,主要會透過活化
STAT1,來表現 CXC chemokines 和 CIITA。 4.8 層流(LF)剪力作用會抑制由IFNγ所引發 的CXC chemokines及CIITA基因表現 為了探討層流(LF)剪力作用對內皮細胞 所產生保護效果,若對HUVECs 施予剪力 4 小時,如 Fig. 10A 所示,並不會引發 CXC chemokines (CXCL9, CXCL10, CXCL11)基 因表現,若以IFNγ (2.5 ng/ml)刺激 HUVECs 則會大量增加CXC chemokines 的表現,然而 若以 IFNγ刺激 HUVECs 再施予剪力 4 小 時,則層流(LF)剪力作用會顯著抑制由 IFNγ 所引發的 CXC chemokines 基因表現(Fig. 10A)。另外,LF 剪力作用也會顯著抑制由 IFNγ所引發的 CXCL10 蛋白質的釋放量(Fig. 10B),與上述現象一致。 此外,LF 剪力作 用 也 會 顯 著 抑 制 由 IFNγ所引發的 CIITA (MHCII 基因轉錄的重要調控因子)的基因表 現(Fig. 10C)。綜言之, LF 剪力作用會抑制 由IFNγ所引發的 CXC chemokines 及 CIITA 基因表現。 4.9 LF 剪力越大可產生對 IFNγ引發 CXC chemokines 基因表現越顯著的抑制效果 在血管分支處,會產生disturbed flow, 伴隨低剪力或是非單一方向流動,來刺激內 皮細胞,與穩定剪力所產生的效果有所不 同。本實驗欲觀察不同剪力大小,對內皮細 胞所產生保護效果之差異。如Fig. 11 所示,
lane 2 表示以 IFNγ (2.5 ng/ml)刺激 HUVECs 4 小時又10 分鐘,CXC chemokines 會大量表 現。若對內皮細胞施予低剪力(2 dyn/cm2)4 小 時,則 IFN-γ所引發的 CXC chemokines 的 mRNA 表現會被部分抑制下來(約抑制掉 50~60%),而高剪力(25 dyn/cm2) 則有更好的 抑制CXC chemokines mRNA 的效果(約抑制 掉80~90%)。因此,LF 可抑制發炎基因表現, 進而保護內皮細胞,而此效果強弱也與剪力 的大小呈現正相關的趨勢。 五、結論 根據本研究上述各項實驗結果,歸納出 以下重點: 1. 以發炎反應中重要的細胞素 IFNγ刺激內 皮細胞會造成STAT1 Tyr701 及 Ser727 磷
酸化上升,引發STAT1 之活化。 2. 剪力作用可抑制由 IFNγ所引發的 JAK1/2 及STAT1 Tyr701 磷酸化,因而血管中血 液 流 動 所 產 生 的 剪 力 可 以 保 護 內 皮 細 胞,緩和細胞素刺激所引發的後續發炎反 應。 3. IFNγ的刺激會引起內皮細胞表現 CXC chemokines,吸引活化的 T cells 沾黏到內 皮細胞表面,可能導致動脈硬化斑塊區域 的發炎反應。 4. 在 層 流 (LF) 剪 力 作 用 抑 制 IFNγ所引發 STAT1 Tyr701 磷酸化的過程,有 protein tyrosine phosphatase 參與其中。
5. 層流(LF)剪力作用會抑制由 IFNγ所引發 的STAT1/1 與 DNA binding 現象。 6. IFNγ可經由活化 JAK/STAT1 訊息傳導路 徑,增加CXC chemokines 和 CIITA 的表 現。 7. LF 剪力作用會抑制由 IFNγ所引發的 CXC chemokines (CXCL9, CXCL10, CXCL11) 及CIITA 基因表現。 8. LF 剪力作用的抑制效果與剪力的大小呈 現正相關的趨勢。 綜合上述,本研究明確顯示層流剪力作 用可抑制 IFNγ引發的 JAK/STAT1 訊息傳導 路徑,進而抑制STAT1 下游與發炎反應相關 的 CXC chemokines 和 CIITA 的基因表現, 因而可望對於血管壁的內皮細胞產生保護效 果,抑制動脈粥狀硬化。
六、參考文獻
[1] Ni CW, Wang DL, Lien SC, Cheng JJ, Chao YJ, Hsieh HJ., “Activation of PKC-epsilon and ERK1/2 participates in shear-induced endothelial MCP-1 expression that is repressed by nitric oxide”,
J. Cell. Physiol. 195, 428-434 (2003)
[2] Mach F, Sauty A, Iarossi AS, Sukhova GK, Neote K, Libby P, Luster AD, “The interferon- inducible CXC chemokines IP-10, Mig, and I-TAC are differentially expressed by human atheroma-associated cells: implications for lymphocyte recruitment in atherogenesis”, J. Clin.
Invest. 104, 1041-1050 (1999)
[3] Pober JS, Collins T, Gimbrone MA Jr, Cotran RS, Gitlin JD, Fiers W, Clayberger C, Krensky AM, Burakoff SJ, Reiss CS. Lymphocytes recognize human vascular endothelial and dermal fibroblast Ia antigens induced by recombinant immune interferon. Nature 305, 726-729 (1983) [4] Aaronson DS, Horvath CM, “A road map
for those who know JAK-STAT”, Science 296, 1653-1655 (2002)
[5] Calo V, Migliavacca M, Bazan V, Macaluso M, Buscemi M, Gebbia N, Russo A, “STAT proteins: from normal control of cellular events to tumorigenesis”, J. Cell.
Physiol. 197, 157-168 (2003)
[6] Masuda M, Suzui M, Yasumatu R, Nakashima T, Kuratomi Y, Azuma K, Tomita K, Komiyama S, Weinstein IB, “Constitutive activation of signal transducers and activators of transcription 3 correlates with cyclin D1 overexpression and may provide a novel prognostic marker in head and neck squamous cell carcinoma”,
Cancer Res. 62, 3351-3355 (2002)
[7] Ni CW, Hsieh HJ, Chao YJ, Wang DL, “Shear flow attenuates serum-induced STAT3 activation in endothelial cells”, J.
Biol. Chem., 278, 19702-19708 (2003)
[8] Ni CW, Hsieh HJ, Chao YJ, Wang DL, “Interleukin-6 induced JAK2/STAT3 signaling pathway in endothelial cells is suppressed by hemodynamic flow”, Am. J.
Physiol. Cell. Physiol., 287, C771-C780
(2004)
七、圖表
Fig. 1. Shear stress (SS) does not affect the basal STAT1 Tyr 701 and Ser727 phosphorylation in HUVECs. HUVECs were in static condition or exposed
to SS (25 dyn/cm2) for 10, 30, 60, 120 minutes. ECs
were stimulated by IFNγ (2.5 ng/ml) for 30 minutes as positive controls. Arrows indicate STAT1α and STAT1β.
Fig. 2. IFNγ stimulates STAT1 tyrosine and serine
phosphorylation. A, IFNγ induced Tyr 701 and Ser727
phosphorylation of STAT1 in a time-dependent manner. HUVECs were treated with IFNγ (2.5 ng/ml) for 5, 10, 30, 50, 120 minutes. B, IFNγ induced tyrosine and serine phosphorylation of STAT1 in a concentration-dependent manner. ECs were treated with IFNγ at the indicated concentration for 30 minutes.
A
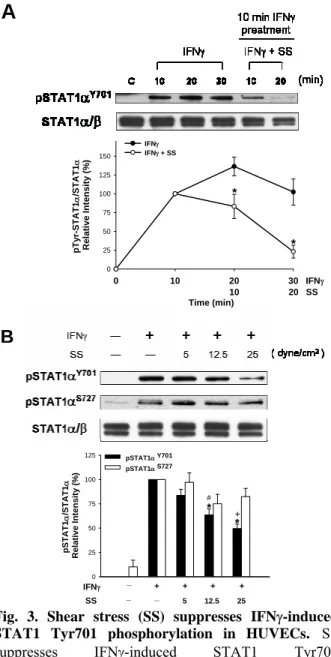
Fig. 3. Shear stress (SS) suppresses IFNγ-induced
STAT1 Tyr701 phosphorylation in HUVECs. SS
suppresses IFNγ-induced STAT1 Tyr701 phosphorylation in a time- and force-dependent manner. A, HUVECs pretreated with IFNγ (2.5 ng/ml) for 10 min were either maintained static or exposed to following SS (25 dyn/cm2) in medium containing IFNγ for an
additional 10 or 20 minutes. After stimulation, cell lysates were subjected to western blot analysis and the Tyr701 phosphorylation was examed. The intensity of Tyr701 phosphorylation was normalized to the level of STAT1α and is presented as mean ± SE of three independent experiments. * P < 0.05 vs. respective IFNγ-treated ECs under static conditions. B, HUVECs pretreated with 2.5 ng/ml IFNγ for 10 min were maintained static or exposed to SS in medium containing IFNγ for 20 minutes with increasing shear stress as indicated, and the Tyr701 and Ser727 phosphorylation of STAT1 was examined. STAT1 phosphorylation was normalized to the level of STAT1 and is presented as mean ± SE of three independent experiments. * P < 0.05 vs. respective IFNγ-treated ECs under static conditions. #P < 0.05 vs. ECs under SS of 5 dyn/cm2. +P < 0.05 vs.
ECs under SS of 12.5 dyn/cm2.
Fig. 4. Shear stress (SS) suppresses IFNγ-induced
JAK/STAT1 activation in BAECs. SS suppresses
IFNγ-induced JAK/STAT1 tyrosine phosphorylation in a time- dependent manner. BAECs pretreated with IFNγ (0.5 ng/ml) for 30 min were either maintained static or exposed to following SS (25 dyn/cm2) in medium
containing IFNγ for an additional 10 or 20 minutes.
Fig. 5. IFNγ induces CXCL10 (IP-10) expression in
HUVECs. Time course study of IFNγ-induced IP-10
mRNA levels. HUVECs were in static condition or treated with IFNγ (2.5 ng/ml) for 1, 2, 3, 4, or 6 hours.
Fig. 6. Protein tyrosine phosphatase is involved in tyrosine dephosphorylation of STAT1. BAECs were
pretreated with Na3VO4 (100 µM), an inhibitor to protein
tyrosine phosphatase, for 30 min followed by 0.5 ng/ml IFNγ for 30 min and then maintained static or exposed to LF (shear stress: 25 dyn/cm2) for 20 min. The phosphorylation of STAT1 was analyzed by Western blot using antibody specific to either pTyr-STAT1 or STAT1. Results are representative of 3 independent experiments with similar results.
0 10 20 30 0 25 50 75 100 125 150 IFNγ IFNγ + SS pT yr-STA T1 α /STA T1 α Re lat ive Inte n s it y ( % ) 10 Time (min) 20 IFNγ SS * *
A
0 25 50 75 100 125 pSTAT1α Y701 pSTAT1α S727 * * # pS TAT1 α /S TAT1 α Rel a ti ve I n te n s it y ( % ) IFNγ SS + + + + 5 12.5 25 + _ _ _ BFig. 7. Laminar flow (LF) suppresses IFN-γ-induced DNA binding of STAT1. Nuclear extracts were
prepared and analyzed by gel shifting using a 32P-labeled
high-affinity sis-inducible element (hSIE) oligonucleotide probe. HUVECs were maintained under static conditions (lane 1) or treated with 2.5 ng/ml IFNγ for 4 hours and 10 minutes (lane 2). Followed by IFNγ pretreatment, ECs were exposed to laminar flow (LF) with shear stress of 25 dyn/cm2 for 4 hours in the
presence of IFNγ (lane 3). The specificity of the retarded complex (STAT1/STAT1 homodimer) was confirmed by preincubating the nuclear extracts with an antibody to STAT1 or STAT3. Results are representative of 3 independent experiments with similar results.
Fig. 8. IFN-γ stimulates CXC chemokines (CXCL9,
CXCL10, CXCL11) and CIITA expressions. ECs were
treated with IFNγ (2.5 ng/ml) at different intervals as indicated. Total RNA was extracted, purified and were subjected to Taqman real-time PCR analysis. All these gene expressions (CXCL9, CXCL10, CXCL11, and CIITA) were normalized to the level of GAPDH. Results are representative of 2 independent experiments with similar results.
Fig. 9. IFN-γ induces expression of CXC chemokines
(CXCL9, CXCL10, CXCL11) via STAT1 activation.
HUVECs were transfected with either STAT1-specific or nonspecific siRNA. mRNA expression of CXC chemokines and CIITA was analyzed using Taqman real-time PCR analysis. In a parallel experiment, ECs transfected with siRNA were analyzed by western blotting to detect the expression of STAT1 in ECs transfected with STAT1-specific or nonspecific siRNA. Results are presented as the mean ± S.E. from three independent experiments. *P < 0.05 and #P < 0.001 vs. respective gene mRNA expression in IFNγ-treated HUVECs transfected with nonspecific siRNA.
hours 0 1 2 3 4 5 6 100 101 102 103 104 105 106 107 CXCL9 CXCL10 CXCL11 CIITA ST AT 1-med iat ed g e n e / G AP DH Re la tiv e ra tio
Fig. 10. Laminar flow (LF) suppresses IFNγ-inducible CXC chemokines (CXCL9, CXCL10, CXCL11) and CIITA. (A) ECs were maintained under static conditions
(lane 1) or exposed to LF (25 dyne/cm2) for 4 hours and 10 min (lane 2); ECs were treated with IFNγ for 4 hours and 10 min (lane 3) or exposed to IFNγ for 10 min followed by LF for 4 hours in a medium containing IFNγ (lane 4). CXCL9, CXCL10 and CXCL11 mRNA levels were determined by real-time quantitative RT-PCR. Chemokine expression was normalized to GAPDH. (B) LF suppresses IFNγ-induced CXCL10 protein production. ECs were treated as in lanes 1, 3 and 4 of A. Conditioned medium was collected for ELISA analysis. CXCL10 protein is expressed as pg/mg of total protein. (C) LF attenuated IFNγ-induced CIITA expression in ECs pretreated with IFNγ. The expression of CIITA was analyzed using RT-PCR. Results are presented as the mean±S.E. from three independent experiments. *P < 0.05 vs. IFNγ-treated ECs under static condition.
Fig. 11. Laminar flow (LF) suppresses IFNγ-induced
CXC chemokines (CXCL9, CXCL10, CXCL11) in a shear stress-dependent manner. ECs were maintained
under static conditions (lane 1), treated with IFNγ for 4 hours and 10 minutes, or pretreated with IFNγ (2.5 ng/ml) for 10 minutes and then subjected to LF in medium containing IFN-γ for 4 hours with shear stress of 2 or 25 dyn/cm2. CXCL9, CXCL10 and CXCL11 mRNA levels
were determined using Taqman real-time PCR analysis. Chemokine expression was normalized to GAPDH and is presented as the mean ± S.E. of three independent experiments. *P < 0.05 vs. respective chemokine expression of IFNγ-treated ECs under static conditions. #P < 0.05 vs. respective chemokine expression of IFNγ-treated ECs under LF (2 dyn/cm2).
0 25 50 75 100 125 CXCL9 CXCL10 CXCL11 * CXC Ch emo k ine /GA P DH Relative Int e ns it y (% ) IFNγ LF + + + 2 25 _ _ _ * * # # # * * * (dyne/cm2)