厚朴酚對培養大鼠腦皮質細胞缺氧所引起細胞傷害的保護作用; Protective Effect of Magnolol on Hypoxic Cell Injury in Rat Cultured Cortical Cells
125
0
0
全文
(2) 目錄 縮寫字對照表 中文摘要暨關鍵字 英文摘要暨關鍵字 第壹章 總論 一、 鼠腦細胞缺氧 <一>、細胞之缺氧模式-氰化鉀 <二>、細胞缺氧後神經訊號傳遞物質的變化 二、 厚朴酚 <一>、厚朴酚之來源 <二>、厚朴酚溶液的製備 <三>、厚朴酚之藥物作用 <四>、厚朴酚之藥物動力學 三、 實驗假說 第貳章 厚朴酚對大鼠腦部血管梗塞引發組織壞死面積之影響 一、緒言 二、材料與方法 <一>、厚朴酚影響血管梗塞引發組織壞死面積 1. 實驗動物 2. 動物腦部血管梗塞的方法 3. 厚朴酚之給藥方法 4. 動物腦部壞死面積的測量方法 <二>、厚朴酚與其它藥物對氰化鉀引起細胞損傷影響之比較. 1.
(3) 1. 細胞培養 2. 細胞毒性分析---乳酸脫氫脢測定 <三>、藥物或化學物 <四>、統計分析 三、結果 <一>、厚朴酚改善大鼠因血管性梗塞引起腦組織壞死現象 <二>、細胞培養之細胞形態的變化 <三>、細胞毒性試驗:乳酸脫氫脢釋放量的變化 四、討論 第參章 厚朴酚對氰化鉀引起細胞損傷影響之初探 一、 緒言 二、 材料與方法 <一>、細胞培養 <二>、細胞免疫化學染色法與細胞形態 <三>、螢光染色分析與細胞核變化 <四>、細胞毒性分析------乳酸脫氫脢 (LDH) 之測定 <五>、細胞生存力的分析------粒腺體琥珀酸脫氫脢 (MTT) 之測定 <六>、藥物或化學物 <七>、統計分析 三、 結果 <一>、神經元及神經膠細胞形態的改變 <二>、細胞乳酸脫氫脢 (LDH) 釋放量的影響 <三>、細胞生存力的影響 <四>、厚朴酚對細胞乳酸脫氫脢的影響. 2.
(4) <五>、厚朴酚對細胞核的影響 <六>、厚朴酚對神經元細胞數目的影響 四、討論 第肆章 氰化鉀對細胞 Ins (1,4,5) P3 活性的影響. 一、緒言 二、材料與方法 <一>、細胞培養 <二>、放射線[3H]分析法分析 Ins (1,4,5) P3 <三>、藥物或化學物 <四>、統計分析. 三、結果 <一>、氰化鉀對細胞內之 Ins (1,4,5) P3 活性的影響 <二>、厚朴酚對細胞內之 Ins (1,4,5) P3 活性的影響 第伍章 厚朴酚對氰化鉀引起細胞前列腺素 E2 與一氧化氮反應之影響 一、 緒言 二、 材料與方法 <一>、細胞培養 <二>、細胞釋放前列腺素 E2 之測定 <三>、細胞生成一氧化氮之測定 <四>、藥物或化學物 <五>、統計分析 三、 結果 <一>、 在含糖培養液情況下,乳酸脫氫脢與細胞生成之前列腺素 E2 的 變化 3.
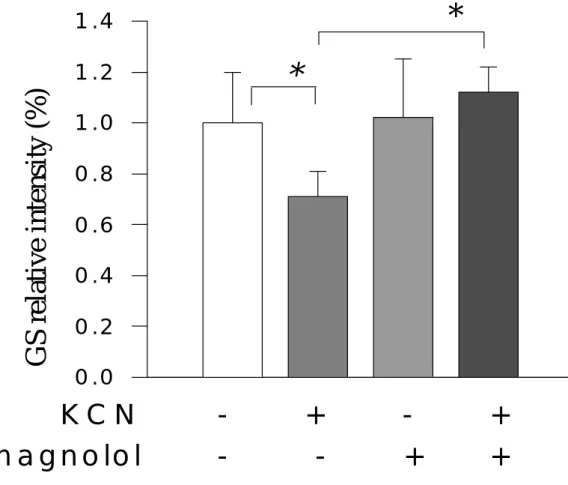
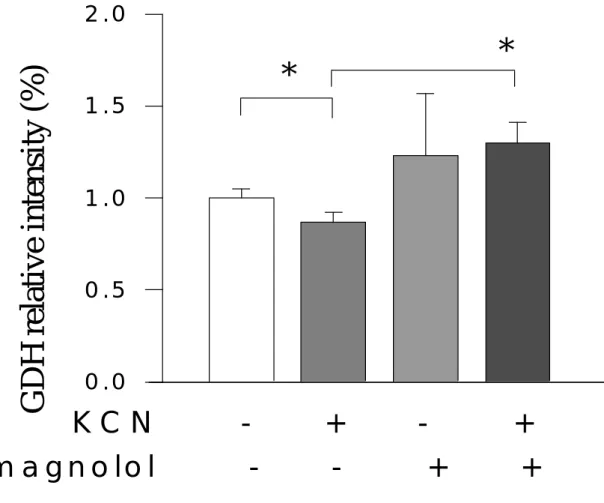
(5) <二>、 在不含糖培養液情況下,細胞生成之乳酸脫氫脢的變化 <三>、 在不含糖培養液情況下,細胞生成之前列腺素 E2 的變化 <四>、 在缺糖情況下,細胞生成之一氧化氮的變化 四、討論 第陸章 厚朴酚對氰化鉀抑制細胞代謝過程之調節作用 一、緒言 二、材料與方法 <一> 細胞培養 <二> 細胞毒性分析------乳酸脫氫脢 (LDH) 之測定 <三> 酵素分析法測定細胞外乳酸 (lactate) 濃度 <四> 酵素分析法測定細胞外麩胺酸 (glutamate) 濃度 <五> 、酵素分析法測定細胞外葡萄糖 (glucose) 濃度 <六> 、北方墨點法 (Western Blot method)測定神經膠細胞酸性纖維蛋白 (GFAP),微管相聯性蛋白(MAP-2),麩胺合成脢(GS),麩胺酸脫氫 脢(GDH),葡萄糖傳送接受體-1(GLUT-1) <七> 、藥物或化學物 <八> 、統計分析 三、結果 <一>、 神經元標誌性蛋白質(MAP-2)的變化 <二>、 神經膠細胞酸性纖維蛋白(GFAP)的變化 <三>、 細胞外乳酸脫氫脢(LDH)之濃度變化 <四>、 細胞外乳酸(lactate)之濃度變化 <五>、 神經膠細胞內麩胺合成脢(GS)的蛋白質強度變化 <六>、 神經膠細胞內麩胺酸脫氫脢(GDH)的蛋白質強度變化. 4.
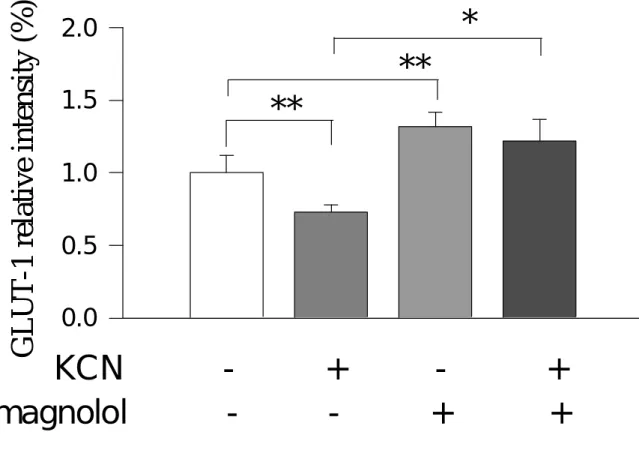
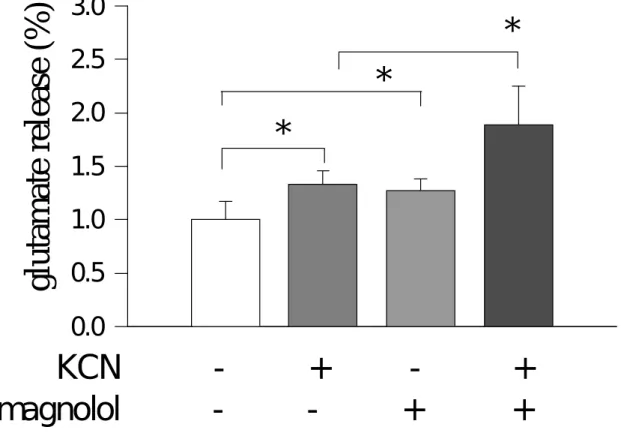
(6) <七>、 葡萄糖傳送接受體-1(GLUT-1)之蛋白質強度變化 <八>、 細胞外麩胺酸(glutamate)之濃度變化 <九>、 細胞外葡萄糖(glucose)之濃度變化 <十>、 麩胺酸對細胞外乳酸脫氫脢(LDH)之濃度變化 四、討論. 5.
(7) 縮寫字對 照表 µg. microgram. 7-NI. 7-nitroindazole. AG. aminoguanidine. CCA. common carotid arteries. DAB. 3,3'-diaminobenzidine tetrahydrochloride. DAPI. 4, 6-diamidino-2-phenylindole. DMEM. Dulbecco’s Modified Eagles Media. DMSO. dimethylsufoxide. Dnas I. Deoxyribonuclease I. EDTA. ethylenediaminetetraacetic acid. G. Gram. GFAP. glial fibrillary acidic protein. GDH. glutamate dehydrogenase. GLUT-1. glucose transporter-1. GS. glutamine synthetase. HBSS. Hanks’ Balanced Salt Solution. Hepes. N-[2-hydroxyethyl] piperazine-N-[2-ethane-sulfonic acid]. Hr. Hour. HS. Horse serum. KCN. potassium cyanide. LDH. lactate dehydrogenase. L-NAME. NG-nitro-L-arginine methl ester dihydrochloride. LPS. Lipopolysaccharide. 6.
(8) LTB4. leukotriene B4. M. Molar. MAP-2. microtubule associated protein-2. MCA. middle cerebral artery. mg. milligram. min. minute. ml. milliliter. mM. millimolar. MTT. 3-(4,5-dimethylthiazol-2,5-diphenyl tetrazolium bromide. ng. nanogram. NO. nitric oxide. PBS. phosphate-buffered saline. PGE2. prostaglandin E2. PS. penicillin/streptomycin. SDS. sodium dodecyl sulphate. TBS. Tris [hydroxymethyl]amino-methane buffered saline. TBST. TBS and Tween 20. TTC. 2, 3, 5-triphenyl tetrazolium chloride. Tween 20. Polyoxyethylenesorbitan monolaurate. 7.
(9) 中文摘要 厚朴酚,為自藥用植物厚朴皮部抽出之主要藥用成份,對於動物腦 部缺氧性傷害的保護作用首先用 TTC 染色法評估大腦梗面積來作檢視。 腹腔注射 chloral hydrate (400 mg/kg) 麻醉大鼠,以手術方法結紮兩側總 頸動脈以及右側中腦動脈造成局部腦缺氧模式達九十分鐘。厚朴酚以腹 腔注射方式給藥,劑量分別為 1,10,30 mg/kg,給藥方式為血管結紮前 30 分鐘以及再灌流前 30 分鐘兩次給藥,腦切片(2mm 厚度)以 TTC 染 色,呈現出腦梗塞面積。結果發現以 10 mg/kg 厚朴酚最可有效減少梗塞 面積達控制組之 58%。 厚朴酚對抗缺氧引起細胞傷害的保護作用主要以體外培養大鼠腦 皮質細胞作試驗。將細胞曝露於 0.5mM 氰化鉀溶液中,神經元產生形態 上之變化,包括細胞外觀結構的破壞和數目的喪失,此變化可被厚朴酚 所減弱;而氰化鉀對神經膠細胞則無明顯可觀察之變化。氰化鉀會引起 細胞外乳酸脫氫脢於劑量及時間上依存性的增加以及存活細胞數目的減 少。加入厚朴酚(10-100µM)處理後可減少氰化鉀所引起的細胞外乳酸 脫氫脢,並有劑量依存性作用。細胞核的萎縮現象並未在氰化鉀處理的 細胞上觀察到,此暗示化學性缺氧導致的細胞死亡方式較傾向於壞死 (necrotic)而少表現在凋零死(apoptosic)。本篇論文為首篇利用培養正常大 鼠腦皮質細胞研究厚朴酚對抗缺氧傷害或壞死性死亡的保護作用。 本篇研究試驗一實驗假說:乙醯膽鹼媒介之訊號傳遞系統可被氰化 鉀或厚朴酚所改變。將細胞曝露於不同的藥物:carbachol,KCN,glutamate 及 bradykinin 溶液中為時 5-30 秒,然而只有 carbachol 及 carbachol 合併 KCN 的處理能明顯提高細胞內三磷酸肌醇的活性,且 carbachol 合併 KCN 的處理要較 carbachol 高 14% (於 5 秒時);此由 carbachol 合併 KCN 的處. 8.
(10) 理引起的反應可被厚朴酚加強 21%。總而言之,此結果說明氰化鉀的處 理可加強乙醯膽鹼媒介之細胞內三磷酸肌醇的形成而厚朴酚會加強此作 用。 另外,我們試驗了厚朴酚的神經保護作用是否包括調節氰化鉀所誘 發之發炎因子,前列腺素 E2 或一氧化氮的路徑中。結果發現,在缺乏葡 萄糖的環境下,氰化鉀或氰化鉀合併細菌脂多糖蛋白內毒素均可引起細 胞外乳酸脫氫脢及前列腺素 E2 的增加達 32%,34%及 12%,32%;而這 兩方面的增加均可受到厚朴酚的抑制。此外,雖然氰化鉀或細菌脂多糖 蛋白內毒素單獨處理無法刺激一氧化氮的產生,然而氰化鉀合併細菌脂 多糖蛋白內毒素可引起一氧化氮的產生。此增加可因厚朴酚或 L-NAME 的處理而減少,但細胞外乳酸脫氫脢及前列腺素 E2 的增加卻無法被 L-NAME 的處理而降低。這些發現暗示厚朴酚對抗氰化鉀或氰化鉀合併 細菌脂多糖蛋白內毒素均引起細胞傷害的保護作用包括在抑制前列腺素 E2 的產生,但是一氧化氮的抑制似乎較不重要。 氰化鉀的毒性會引起細胞代謝性的障礙(例如細胞外麩胺酸的堆 積),影響細胞在能量來源上的利用並且造成能量喪失。我們接著研究 是否厚朴酚能夠調節細胞代謝過程以保護細胞對抗氰化鉀之缺氧性傷 害。細胞傷害的程度以細胞外乳酸脫氫脢及乳酸活性作評估。細胞外麩 胺酸的分析利用酵素分析法,細胞內麩胺酸代謝酵素的蛋白質強度,包 括麩胺酸脫氫脢 (glutamate dehydrogenase) 及麩氨合成脢 (glutamine synthetase),以北方墨點法作評估。厚朴酚可減少氰化鉀引起的細胞外乳 酸脫氫脢及乳酸的增加,但加強了麩胺酸在細胞外的蓄積。氰化鉀會造 成抑制麩胺酸代謝酵素(麩胺酸脫氫脢及麩氨合成脢),而此抑制可被 厚朴酚所減弱。此外,氰化鉀會減少神經元標誌性蛋白(MAP-2)以及神經. 9.
(11) 膠細胞標誌性蛋白 (GFAP) 的強度。厚朴酚可減少氰化鉀對 MAP-2 的抑 制,但無法影響氰化鉀對 GFAP 的抑制。總之,厚朴酚保護細胞對抗氰 化鉀之缺氧性傷害,也包括了調節細胞代謝過程並且減弱氰化鉀對麩胺 酸代謝酵素的抑制作用。. 10.
(12) 英文摘要 The protective effects of magnolol , a component of Magnolia officinalis, on hypoxic injury were first examined by measuring infarct size by TTC method. In chloral hydrate (400 mg/kg, intraperitoneally) anesthetized rats, a focal cerebral ischemia was produced by occluding two common carotid arteries (CCA) and right middle cerebral artery (MCA) for 90 min. Intraperitoneal injection of magnolol (1, 10, 30 mg/kg) was gived twice at 30 min prior to the occlusion and at 30 min before reperfusion. Brain slides (2 mm thick) were stained by TTC. The infarct size was attenuated by 10 mg/kg magnolol by 58% (p<0.05) when compared with control. Protective effect of magnolol against hypoxia-induced cell injury in cortical neuron-astrocyte mixed cultures was examined. Exposure of the cells to chemical hypoxia (0.5 mM KCN) produced morphological changes in neurons but not in astrocytes. KCN induced dose- and time-dependently increases in release of LDH and decreases in viable cell number. Treatment of magnolol (10 and 100 µM) significantly reduced the KCN-induced LDH-release in a concentration-dependent manner. The higher concentration (750 µM) was toxic. Nuclear condensation was not observed in KCN-treated cells, suggesting chemical hypoxia-induced cell death via necrosis, rather than via apoptosis. This is the first report demonstrating magnolol protected neurons against chemical hypoxic damage or necrotic cell death in cortical neuron-astrocyte mixed culture. This. study. tested. the. hypothesis. that. acetylcholine. (ACh). receptor-mediated signal transduction systems in mixed cortical cultured cells. 11.
(13) were altered after treatment with KCN or magnolol. Exposure to various stimuli of cabachol, KCN, glutamate and bradykinin for 5-30 secs, while only carbachol and KCN plus carbachol significantly elevated the activity of Ins (1,4,5) P3 by 14% (p<0.05 vs. carbachol) at 5 secs. This response of KCN combined with carbachol was further enhanced in the cells treated with magnolol by 21% (p<0.05 vs carbachol plus KCN) at 5 secs. Taken together, the results demonstrated that treatment with 0.5 mM KCN enhanced ACh receptor-mediated Ins (1,4,5) P3 formation and magnolol potentiated this effect. In the present study, we examined whether the neuroprotective effect of magnolol involve modulating inflammatory mediators, prostaglandin E2 (PGE2) and nitric oxide (NO), induced by KCN (hypoxia). In glucose-absent (hypoglycemia) media, KCN or KCN plus LPS induced increases in lactate dehydrogenase (LDH) activity by 32% and 34%, and PGE2 production by 12% and 32%, respectively. Both LDH and PGE2 increases were suppressed by 100 µ M magnolol. In addition, although KCN or LPS alone did not increase NO generation, KCN plus LPS increased NO generation. This increase was reduced by 100 µ M magnolol or 10 µ M L-NAME, but the LDH increase and PGE2 production were not reduced by L-NAME. These findings suggest that the protective effects of magnolol against brain damage by KCN or KCN plus LPS in hypoglycemic media may involve inhibition of PGE2 production, but inhibition of NO generation may not be important. KCN intoxication may cause cellular metabolic disorders, affect cell utilization in energy source and result in energy depletion. We studied whether magnolol may modulate cellular metabolic processes to protect cell. 12.
(14) against hypoxic injuries in rat cortical neuron-glia mixed cultured cells exposed to 0.5 mM KCN in the glucose-free medium for 60 min. Cell injury was evaluated on release of lactate dehydrogenase (LDH) and activity of lactic acid (lactate). Extracellular glutamate levels were assessed by an enzymatic method. Protein intensities of intracellular metabolic enzymes (GDH and GS) were evaluated by Western blot method. Magnolol attenuated the LDH- and lactate-increases induced by KCN and enhanced the glutamate-accumulation elevated by KCN. KCN resulted in suppressions of metabolic enzyme (protein) intensities, such as glutamine synthetase (GS) and glutamate dehydrogenase (GDH), the suppressions by KCN were markedly attenuated by 100 µM magnolol. In addition, KCN suppressed microtubule associated protein-2 (MAP-2), a neuronal cell marker protein, as well as glial fibrillary acidic protein (GFAP), an astrocyte marker protein. Magnolol attenuated the suppression in MAP-2 intensity, but did not affect the suppression in GFAP intensity. In conclusion, magnolol protects neuronal cell against KCN-injuries, involving modulation of metabolic processes by attenuating KCN-induced suppressions in GS and GDH protein intensities.. 13.
(15) 第壹章 總論 一、 鼠腦細胞缺氧 <一>、體外培養細胞之缺氧模式-氰化鉀 化學性缺氧(chemical hypoxia)模式為應用在探討缺氧(ischemia)引起 神經變性分子機轉之一相當有用的模式(Gibson-GE, 1991)。氰化鉀(KCN) 可引起細胞產生缺氧性傷害,干擾電子傳遞鏈及阻斷呼吸作用,現今已 知對神經元可產生不可逆的傷害。而氰化鉀對呼吸電子傳遞鏈的阻斷效 果與缺氧結果類似,因而氰化鉀中毒常被使用來作為化學性缺氧模式 (Goldberg-MP, 1987; Gibson-GE, 1991)。 <二>、細胞缺氧後神經訊號傳遞物質的變化 氰化鉀中毒會造成鼠腦部位神經傳遞物質(neurotransmitters)在量上 的變化,如胞外麩胺酸(glutamate)、乙醯膽鹼(acetylcholine)、(catecholamine) 釋放量的增加,以及 GABA 的減少(Persson-SA, 1985)。而對於細胞內之 訊號 (signal) 傳遞物質的影響則包括:鈣離子的衡定,自由基(free radical) 的生成。並且可能產生發炎性物質。所以本論文擬以白鼠培養大腦皮質 細胞,包括神經細胞及神經間膠細胞,以 KCN 引發細胞缺氧 ,建立缺 氧或缺葡萄糖對神經細胞及神經間膠細胞的致毒性,以乳酸脫氫脢 14.
(16) (lactate dehydrogenase, LDH)、粒腺体琥珀酸脫氫脢 (MTT) 的活性評估 細胞的損傷;評估中藥萃取物厚朴酚 (magnolol) 是否可減低缺氧性細胞 傷亡;探討細胞的損傷及厚朴酚的作用機轉是否與訊號傳遞物質之釋 放,花生四烯酸或一氧化氮有關;探討並比較神經細胞及神經間膠細胞 的相互關係。 二、 厚朴酚 <一>、厚朴酚之來源 厚朴酚 (magnolol) 之植物來源為木蘭科(Magnoliaceae)植物厚朴 (Magnolia officinalis)之乾燥樹皮的活性成份。實驗中使用之厚朴酚為購 自建吾藥品公司代理日本米山純化合物公司 (Yoneyama company, Osaka, Japan) 於大阪 (Osaka) 生產純化之厚朴酚純化合物 (附圖 1)。 <二>、厚朴酚溶液的製備 體外試驗中所使用之厚朴酚溶液為溶於 DMSO 溶液中,作成 100 mM 之 stock 溶液,貯存於-20℃,實驗時用 DMEM 無 phenol red 培養液 稀釋至終濃度(100 µM)。體內試驗中所使用之厚朴酚溶液為溶於 0.1 N NaOH 溶液中。. 15.
(17) <三>、厚朴酚之藥物作用 厚朴酚係一酚類化合物,為自厚朴皮部分離之重要藥用成份(Fujita et al., 1972)。已經實驗證實之藥理活性如:中樞神經抑制作用(Watanabe et al., 1983),抑制大腦皮質血清素 (serotonin) 之釋放 (Tsai-TH, 1995),肌 肉鬆弛作用(Teng-CM, 1990),抗菌(antibacteria)活性(Clark-AM, 1981),抗 血 小 板 凝 集 (anti-aggregation) 作用(Teng-CM, 1988) ,抑制凝血脢原 (collagen) 誘發之血小板 (platelet) 釋放血清素。在保護作用方面,厚朴 酚能夠保護大鼠心臟粒線體對抗脂質過氧化作用引起的傷害 (Lo-YC, 1994) , 並 對 大 鼠 具 有 抗 因 冠 狀 動 脈 結 紮 (ligated) 引 發 之 心 律 不 整 (arrhythmic) 及 缺 氧 (ischemia) 傷害(Hong-CY, 1996) ,或作為抗氧化劑 (antioxidant) 抑 制 氫 氧 自 由 基 (hydroxyl free radical) 的生成 (Fujita-S, 1994) 。在抗發炎 (anti-inflammation) 作用方面,厚朴酚能夠抑制小鼠因 藥物 A23187 誘發後足蹠水腫 (hindpaw edema) 的程度,因其能減少前列 腺 素 E2 (PGE2) 及 白 三 烯 素 B4 (LTB4) 在水腫液中的形成 (Wang-JP, 1995)。 <四>、厚朴酚之藥物動力學 (Yong-Hua MA, 1988; Masao Hattori, 1986) A. 溶解度:0.1N NaOH 可提高溶解度達 86% 以上. 16.
(18) B. 腸肝循環:口服六小時後血液中濃度開始下降 C. 靜脈注射:線性藥物動力學。約七十五分鐘(5×t1/2)後達穩定狀態 濃度(steady-state concentration) D. 20 mg / kg, p.o⇒0.1 µg / ml in serum E. 0.5 mg / kg, iv →10 min → cortex (均勻分佈 Brain 各區) → 12 µg / g, tissue 二、 實驗假說. 厚朴酚已經實驗證實能使心臟缺氧及再灌流造成的梗塞性傷害面積 減少 (Hong-CY, 1996),並且能對抗脂質過氧化作用對大鼠心臟粒線體造 成的傷害,此效果甚至遠超過生育醇 (tocopherol) (Lo-YC, 1994)。因此, 厚朴酚是否具有保護細胞對抗缺氧性傷害的作用,我們將利用生體外(in vitro)細胞培養的方式,探討厚朴酚對氰化鉀引起細胞損傷的影響。. 17.
(19) 第貳章 厚朴酚對大鼠腦部血管梗塞引發組織壞死面積之影響 一、 緒言 中風引起的腦缺血造成腦部損傷與細胞死亡,主要是由於腦血流不 足與缺氧,而其病理機轉不清楚。腦缺血造成腦部受損與細胞損傷之病 理機轉可能為能源不足,鈣離子恆定系統受到干擾,活化磷脂質 C 或 A2,產生磷脂肌醇,花生四烯酸,大量釋出神經傳導物質。腦缺血及再 灌流過程也會引起發炎,釋放介白質及一氧化氮造成神經細胞的傷害。 神經間膠細胞也可能參與腦缺氧引起發炎反應導至神經細胞傷害的作用 過程。文獻報導中藥萃取物,厚朴酚 (magnolol),是抗發炎藥物,但其 對中樞神經系統是否具抗發炎作用,是否經由抗發炎效應保護神經細胞 有待探討。 二、材料與方法 <一>、厚朴酚影響血管梗塞引發組織壞死面積 1. 實驗動物 本實驗動物採用 Sprague-Dawley 雄性大鼠,體重 300 至 350 克,由台中 榮民總醫院醫學研究部動物室飼育提供。. 18.
(20) 2. 動物腦部血管梗塞的方法 將 雄 性 大 鼠 以 腹 腔 注 射 Chloral hydrate (400mg/ml/kg, intraperitoneally) 麻醉,以保溫板維持體溫 37±1℃,手術前先剃除頸部及 頭部之毛髮,大鼠腹部朝上, 以手術刀劃開頸部皮膚,露出左右頸動脈 (common carotid arteries,以下簡稱 CCA),以手術縫線稍固定之,再將 其頭部固定在小動物用立體定位儀 (Kopf stereotaxic apparatus)上,以手 術刀劃開頭部皮膚,用棉球除去血跡,並撥開皮下組織,使頭骨露出,用 骨剪將一側中腦動脈 (middle cerebral artery,以下簡稱 MCA)上方頭骨剪 開,並將中腦動脈以手術縫線稍固定之.進行血管梗塞狀態試驗時.同時結 紮 (occlusion) 兩側頸動脈及一側中腦動脈,以「雷射都卜勒」 (laser dopplar) 監測中腦動脈內血流量變化情形,以確定血管結紮之完全。血 管結紮 (ligation) 時間為時九十分鐘,時間到鬆開頸動脈及中腦動脈縫 線,此時血液再次通過血管,稱為再灌流(reperfusion)。 3. 厚朴酚之給藥方法 厚朴酚之給藥濃度為 1mg,10 mg, 30 mg/kg/ml (i.p),泡製時溶於 0.1 N NaOH 水溶液中。. 19.
(21) 實驗方法. Ligation time (min). magnolol. ligation. -30. 0. magnolol. 30. 60. reperfusion. 90. . 4. 動物腦部梗塞面積的測量方法 此方法根據林天南教授所指導之「TTC 染色法」。將大鼠腹部麻醉後 剪開腹腔及胸腔肋骨 (附圖 2),裸露出肝臟及心臟,以 19G×3/4 蝴蝶針 插入左心室,並以 PE 管連接灌流幫浦。在右心房開一小孔後立即開啟幫 浦,以 200ml/10min 流速灌流(perfusion) Normal saline,直至大鼠肝臟呈 現灰白色後停止。將大鼠自頸部截斷取腦,挑除腦部血管膜(dura)後 將全腦置入腦切片模型中,製成厚度 1.5 mm 之大腦橫向切片,再將切片 (註 1). 移置事先加熱至 37℃之 TTC. 溶液中,持續保溫加熱五至十分鐘後可. 20.
(22) 見切片呈現紅白兩色,紅色部分為存活組織,白色部分則為死亡組織(附 圖 5)。可將腦部之各部分切片保存於 4﹪paraformaldehyde 中,以利下次 照相並避免切片組織乾縮變形。腦梗塞面積之計算為利用「 Image-Pro plus soft propram」,結果以全腦面積之百分比表示之。. <二>、厚朴酚與其它藥物對氰化鉀引起細胞損傷影響之比較 1. 細胞培養 (1) 細胞來源 實驗中使用的細胞種類屬於正常細胞原始混合型培養(primary mixed culture),細胞取自出生一日齡之內 Sprague-Dawley rat 的大腦皮質部位。 (2) 培養基及化學試劑 (A)細胞培養液: horse serum (HS) 購自 Hyclone (Logan, Utah, USA); Dulbecco's modified Eagle's (DME) medium, neurobasal, B27 supplement. 註1. 購自 GibcoBRL (Grand Island, NY, USA), (B) 試藥及酵素: dimethylsulfoxide(DMSO), CaCl2, EDTA, papain, deoxyribonuclease I (DNas I), Hanks‘ Balanced Salt Solution (HBSS), cysteine, poly-D-lysine, N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid. 21.
(23) (Hepes), boric acid, penicillin, streptomycin 購自Sigma Chemicals Co (St. Louis, MO, USA) (3) 細胞培養方法 註2. 收集取下之大腦皮質部位,浸於 5 ml papain 溶液. 中,並在 37℃水. 浴中浸煮 20 分鐘以分離腦細胞組織,此分解反應當加入 10% horse serum 註. 3. 而終止。接著以 200 離心力的速度離心,細胞沉澱再次懸浮於 DMEM. 包含 10% horse serum 及 penicillin, streptomycin 的溶液中,將細胞懸浮液 移置到預先覆蓋(coating)過 poly-D-lysin. 註. 4. 的細胞培養皿中。細胞接種密. 度因培養皿的種類而不同,96 孔洞的平盤培養皿(96-well plate)為每孔洞 1 ×105 個細胞;4 孔槽的平盤培養皿(4-well chamber slide)為每孔槽 2×105 個細胞,細胞接種後培養於 5% CO2,37℃之恆溫培養箱。接種後 2 小時 更換細胞培養液為 neurobasal,並補充 B27 及 penicilin/streptomycin,0.5 mM glutamine 和 25 µM glutamate. 培養後 4 天,更換不含 glutamine 和 glutamate 的細胞培養液,即 neurobasal 補充 B27 及 penicilin/streptomycin,然後另外 再培養 10 天直至實驗當日。 註 1:B27 為一群化學物質組成的濃縮液,由 Brewer’s B-18 的組成改變而來 (Brewer-GJ, 1989)。. 22.
(24) 註 2:papain 溶液組成份包括:1.5 mM CaCl 2, 0.5 mM EDTA , 0.6 mg/ml papain, 0.063 % DNas I, 5.32 g/l HBSS, 0.2 mg/ml cysteine 註 3:horse serum 應為熱不活化性(heat-inactivated),使用前先在 60℃水浴中浸煮 30 分鐘,以破壞其中之補體反應的作用。 註 4:poly-D-lysin 濃度為 3 µg/ml 溶於 0.15 M boric acid. 2. 細胞毒性分析---乳酸脫氫脢測定 細胞毒性的分析可藉由測定細胞培養液中乳酸脫氫脢的活性來表 現,利用 LDH diagnostic kit 試藥與乳酸脫氫脢形成一紫色化學物質 -formazan,根據比色法算出乳酸脫氫脢之活性濃度。實驗當日,吸掉細 胞培養液,並將細胞曝露於已預先配製好之 0.5 mM KCN 溶液當中,直 到實驗所需時間 30, 60, 120 min,實驗過程均在 37℃之水浴中操作。KCN 需當日新鮮配製,秤量好之 KCN 溶於 DMEM 含 Hepes 而不含 glucose 的 溶液中,以鹽酸溶液調整酸鹼度至 pH 7.4,注意絕不可超過 pH 7.4,否 則易因酸化而使得 KCN 分解而釋出氰氫酸。實驗時間到時,以定量吸管 吸取細胞液,並隨即將細胞液置於冷凍離心機中於 4℃,14,000×離心力 下離心 5 min。吸取 50µl 之細胞上清液並置於 96 孔洞微滴平盤中,加入 50µl LDH diagnostic kit 之 reaction mixture 溶液,於室溫下反應 30 min 而 呈色. 23.
(25) <三>、藥物或化學物 TTC(2,3,5-triphenyl tetrazolium chloride)購自 Sigma Chemical Co(St. Louis,. MO,. ). USA. N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic. ; acid. D-(L)-glucose; (Hepes);. lactate. dehydrogenase (LDH) kit 購自 Boehringer Mannheim (Germany). potassium cyanide (KCN)購自 Merk Chemicals (Darmstadt, Germany)。 註一:TTC 溶液濃度為 2 ﹪(w/v) 溶於 Normal saline solution. <四>、統計分析 結果之表示以平均值加減標準誤差(mean±s.e.m.),體內試驗以動 物隻數為平均,體內試驗以細胞培養之次數為平均。為單因子變異數分 析時,統計上平均值之差異以 Student’s t-test 決定之。為多重因子變異數 比較時,結果之分析以 one-way ANOVA 表示之。P 值小於 0.05 稱為有顯 著統計上之差異。. 24.
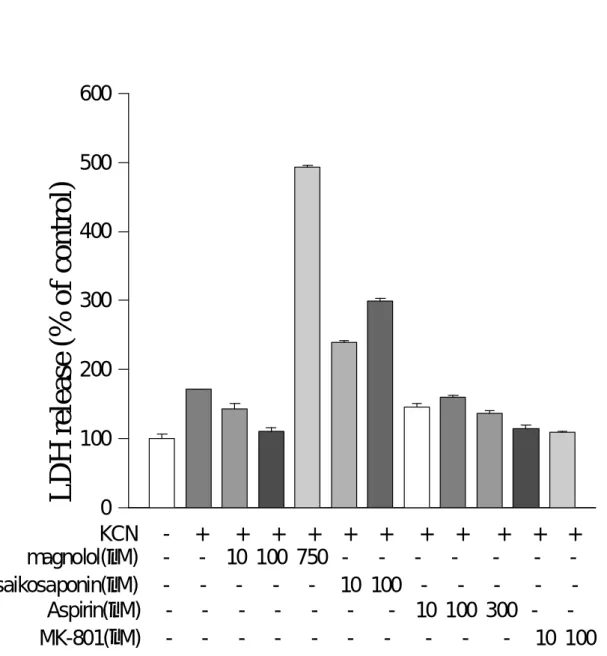
(26) 三、結果 <一>、厚朴酚改善大鼠因血管性梗塞引起腦組織壞死現象 圖 1-1 實驗完成之大鼠腦切片經 TTC 染色後,依切片順序排列 (由大腦 額葉至枕葉),照相結果如下。控制組之梗塞面積自 S1 至 S7 均明顯可見, 而厚朴酚之實驗組則以 10 mg/kg 之梗塞面積自 S3 至 S5,治療效果最好。 圖 1-2 經統計結果顯示,大鼠腦梗塞面積在控制組為 17±2 %,而厚朴 酚之實驗組則以 10 mg/kg 之劑量治療效果為最好 (腦梗塞面積為 7±1 %)。 <二>、細胞培養之細胞形態的變化 圖 2 利用逆相顯微鏡 (reverse-phase microscope) 分別觀察一日、四日 及十四日齡之細胞,顯見其細胞成熟之過程以十四日齡之細胞有較完整 之細胞樹突生成。 <三>、細胞毒性試驗:乳酸脫氫脢釋放量的變化 圖 3 實驗中比較厚朴酚與其它抗炎性藥物,aspirin (NSAID drug), saikosaponin A (柴胡皂甘 A) 及 MK-801(麩胺酸接受體拮抗劑),對氰化 鉀引發細胞缺氧性傷害的影響。. 25.
(27) 26.
(28) 圖 1-1. Fig. 1 Photograph presentation for brain slides of TTC-stained. Rats were treated with CCA and MCA occlusion (control), treatment of magnolol (1, 10 and 30 mg/kg, intraperitoneal) reduced the infarct volume. Magnolol was given 30 min before the start of occlusion and 60 min after the start of occlusion, the occlusion was maintain 90 min.. 27.
(29) 圖 1-2. 25. *. *. n=5. n=3. Infarct Size (%). 20 15 10. n=2. n=10. 5 0. magnolol(mg/kg) 0. 10. 1. 30. Fig.1-2 Effect of pretreatment of magnolol on infarct volume. Pretreatment of magnolol (1, 10 and 30 mg/kg, intraperitoneal) reduced the infarct volume by 5% (n=5), 59% (n=10) and 53% (n=2) of the control (n=3), respectively. Magnolol was given 30 min before the start of occlusion and 60 min after the start of occlusion, the occlusion was maintain 90 min. *P<0.05 vs. Control ( without magnolol), Mean ± S.E.M.. 28.
(30) 圖2. Fig.2. The reverse-phase microscope photograph presenzated the growth stations of mixed cultured cortical cells. Cells were grown in neurobasal containing B27 and PS for one(A), four(B) and fourteen(C) days.. 29.
(31) 圖3. 600. LDH release (% of control). 500 400 300 200 100. 0 KCN magnolol(µM) saikosaponin(µM) Aspirin(µM) MK-801(µM). -. + + + + - 10 100 750 - - - - - - - - - -. + + - 10 100 - -. + + + + - - - 10 100 300 - - 10. + 100. Fig. 3 Effect of magnolol on KCN-stimulated LDH release compared with various anti-inflammatory drugs on rat cultured cortical cells. Treatment of 100 µM of magnolol (mag) or MK-801 (MK) or aspirin (ASP) or saikosaponin A (SAIKO) affected LDH-release induced by 0.5 mM KCN.. 30.
(32) 三、 討論. 由本章研究結果顯示,厚朴酚可以有效預防大鼠腦部血管梗塞所造成 之腦皮質細胞之死亡現象,減少腦梗塞之受損面積。而由乳酸脫氫脢之 實驗結果得知,厚朴酚保護神經元的效果顯然優於其它常用抗腦缺血的 藥物,如 Aspirin, MK-801。 TTC 染色法已被用來評估腦組織受損面積 (Yang-Y, 1998),此方法便利 而且有效。厚朴酚已經實驗證實能使心臟缺氧及再灌流造成的梗塞性傷害 面積減少 (Hong-CY, 1996),本章研究結果更進一步證實厚朴酚對腦及再 灌流造成的梗塞性傷害也極為有效。而在本章體外實驗中也發現厚朴酚可 以有效減少氰化鉀對神經細胞的缺氧性傷害。由於至今尚無研究報告探討 厚朴酚對腦部缺氧性傷害的影響,因此本篇論文將利用體外培養腦神經細 胞的方式,及氰化鉀之缺氧模式來探討厚朴酚是經由何種機制來保護細胞 免於缺氧作用的損傷。. 31.
(33) 第參章 厚朴酚對氰化鉀引起細胞損傷影響之初探 一、 緒言. 缺氧/缺血後的神經變性的機轉可能包括鈣離子恒穩狀態 (homeostasis)的改變,神經傳遞物質的釋放,氧化性壓力的產生以及 發炎反應。化學性缺氧的模式是相當可用來探討神經變性的分子機轉 (Gibson,1991)。氰化鉀(KCN),其作用在干擾電子鏈的傳遞並且阻 斷呼吸(Jones,1984),已知可對神經元產生不可逆的改變。而其對呼吸 電子傳遞的阻斷作用與缺氧狀況產生的作用相似。因此,氰化物中毒已 常被用作化學性缺氧阻斷劑(Chemical hypoxia)(Gibson-GE, 1991) (Goldborg,1984)。 研究發現,大鼠在氰化物中毒的情況下,會引起胞外麩銨酸 (glutamate)及乙醯膽鹼(acetycholine),catecholamine 的增加,但 GABA 的濃度減少(Persson, 1985)。化學性缺氧也會造成海馬迴腦切片和突觸 細胞體釋放麩銨酸(Patel, 1991; Jose, 1988)及 PC12 神經元細胞釋放 catecholamine(Kanthasamy, 1991)。最重要的是,過量的興奮性氨基酸 和活化其接受體都在神經元的傷害及死亡中扮演了極重要的角色。而再 活化氧原子基(Reactive oxygen species, ROS)所引起的脂質過氧化作用. 32.
(34) 也有極大的關聯,所以,自由基(free radical)清除者(scavenger)及抗 氧化劑(antioxidant)也經研究證實具有保護神經元避免肝臟缺血所造成 的損傷(Marubayashi, 1987)。 厚朴酚,一酚類化合物,是中藥植物厚朴皮部的一種活性成份。 在實驗室的研究方面,其能減少大鼠心臟部位經缺血/再灌流 (ischemia/reperfusion)後的梗塞受損面積(infarct size)(Hong-CY, 1996)。其可保護離體鼠心臟粒腺體對抗脂質過氧化作用,且效果比生 育醇(tocopherol)更好。此外,厚朴酚的抗發炎作用能夠緩和 A23187 所引起小鼠後蹠浮腫的程度,經由減少前列腺素 E2(PGE2)及白三烯素 B4(LTB4)的產生(Wang-JP, 1995),和降低小鼠後蹠浮腫液中 eicosanoid 介質的含量(Wang-JP, 1992)。然而,厚朴酚是否具有保護神經元避免 受到缺氧性傷害而死亡,卻從未有人探討過,因此,我們利用體外培養 神經細胞,以氰化鉀(KCN)作為細胞缺氧的實驗模式,專責探討厚朴 酚能否在此情況下保護神經細胞。 二、 材料與方法. <一>、細胞培養 實驗中所使用的細胞材料相同於第貳章之「材料與方法」內「細胞. 33.
(35) 培養」中所述。 <二>、細胞免疫化學染色法與細胞形態 接種於 4 孔槽平盤培養皿並培養 14 天之細胞先吸去培養液,以 PBS 註1. 緩衝溶液洗 2 次,然後加入 4% paraformaldehyde. 註2. 固定 10 min。再次. 以 PBS 緩衝溶液洗 2 次之後加入 3% H2O2 和 10% methanol 溶於 0.1 M PBS 之混合溶液行過氧化作用 15 min。然後將細胞浸於 5% 脫脂奶粉(skim milk)溶於 0.05M TBS. 註3. 的溶液中 30 min 當作 blocking buffer,填補抗原. 沒有吸附到的孔槽空隙,以避免非專一性的吸附。接著以 0.05 M TBST 4. 註. 浸洗細胞,吸乾液體,加入對特殊細胞具特異性之單株抗體(monoclonal. antibody),並在 4℃下培養過夜。神經元細胞(neuron)的形態(morphology) 染 色 使 用 的 抗 體 為 微 管 相 聯 性 蛋 白 質 -2 (microtubule associated protein-2),濃度為 1:400;星狀神經膠細胞(astrocyte)的抗體為神經膠細 胞酸性纖維蛋白質(glial fibrillary acidic protein),濃度為 1:500。之後用 0.05 M TBST 浸洗細胞,吸乾液體,加入二次抗體(secondary antibody). 註5. 於. 室溫下培養 2 小時。再次用 TBST 浸洗後,培育於 avidin-biotinated horse-radish peroxidase 中 30 min,並接著使用 DAB 呈色 15 min 及 hematoxylin 作背景染色。 註 1:PBS 組成為 0.1 M Na 2HPO4, 0.1M NaH2PO4, 0.9 % NaCl,酸鹼度調至 pH=7.4. 34.
(36) 註 2:paraformaldehyde 溶於 0.1 M PB 緩衝溶液中,0.1 M PB 組成為 0.1 M Na 2HPO4, 0.1M NaH2PO4 註 3:0.05 M TBS 組成為 0.05 M Tris [hydroxymethyl]amino-methane 加入 0.9 % NaCl, 酸鹼度調至 pH=7.6. 註 4:0.05 M TBST 組成為 TBS 加入 0.02 % Tween-20. 註 5:實驗中使用之二次抗體為 horse anti-mouse IgG,濃度為 1:25,稀釋於 0.05 M TBS. <三>、螢光染色分析與細胞核變化 接種於 4 孔槽平盤培養皿並培養 14 天之細胞先吸去培養液,以 PBS 1. 緩衝溶液洗 2 次,然後加入 4% paraformaldehyde. 註2. 註. 固定 10 min。再次以. PBS 緩衝溶液洗 2 次之後加入濃度 1:1000 的螢光染色劑 DAPI (稀釋於 100 ﹪methanol),其能與細胞核中的 DNA 鍵結,培育 10 min 作細胞核染色。 於螢光顯微鏡下觀察比較細胞核有無分裂或縮小情況,並計算完整細胞核 的數目有無變化。 1、細胞計數 細胞經免疫化學染色後,經顯微鏡(Olympus)放大 200 倍取景照相, 底片為 35 mm Kodak VX-100 彩色負片。沖洗出 3×5 吋彩色相片,以相 片分別計算出神經元細胞及神經膠細胞之總數,並利用墨汁點去已算過. 35.
(37) 的細胞以為記號。細胞計數採兩次實驗之結果,每兩個孔槽取十處景並 平均計算之。 <四>、細胞毒性分析------乳酸脫氫脢 (LDH) 之測定 相同於第貳章之「材料與方法」內「細胞毒性分析------乳酸脫氫脢 (LDH) 之測定」中所述。 <五>、細胞生存力的分析------粒腺體琥珀酸脫氫脢 (MTT) 之測定 使用 MTT 來作定量測定細胞增生(cell proliferation) 及活化之程度 (Denizot-F, 1986)。此項分析基礎在於存活細胞中含有粒線體琥珀酸脫氫 脢 (mitochondrial succinate dehydrogenase),可以將黃色 tetrazolium salt (MTT) 轉換成紫色 formazan 結晶,此紫色 formazan 結晶為可溶性並且 形成有顏色之溶液,藉由測定其於 570 nm 波長下之吸光值,對照細胞數 目所得到之標準曲線,可得到實驗結果,參考波長為 630 nm。 <六>、藥物或化學物 D-(L)-glucose;. N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic. acid. (Hepes); D-(L)-glucose; glial fibrillary acidic protein (GFAP); (microtubule associated protein-2) MAP-2; 4,6-diamidino-2-phenylindole; dihydrochloride (DAPI); 3,3'-diamidinobenzidine tetrahydrochloride (DAB) 購 自 Sigma. 36.
(38) Chemicals. Co. (St.. Louis,. MO,. USA).. (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) 及 lactate dehydrogenase (LDH) kit 購自 Boehringer Mannheim (Germany). Potassium cyanide (KCN)購自 Merk Chemicals (Darmstadt, Germany) and ABC kit 購自 Vector (Vector, Burlingame, CA, USA). <七>、統計分析 結果之表示以平均值加減標準誤差(mean±s.e.m.),並以細胞培養 之次數為平均。為單因子變異數分析時,統計上平均值之差異以 Student’s t-test 決定之。為多重因子變異數比較時,結果之分析以 one-way ANOVA 表示之。P 值小於 0.05 稱為有顯著統計上之差異。. 37.
(39) 三、 結果. <一>、神經元及神經膠細胞形態的改變 圖 1 由免疫化學細胞染色法可看出,鼠腦皮質細胞受到氰化鉀的傷害, 神經元 (neuronal cells) 會發生形態上之改變,但神經膠細胞(glial cells)則 無。培養14天之成熟細胞,經細胞計數後結果約有30﹪為神經膠細胞,70 ﹪為神經元細胞。而在無糖的實驗情況下,氰化鉀引起之神經毒性會造 成神經元細胞數目減少及網狀結構形成之損害 (impairment of network formation)。 <二>、細胞乳酸脫氫脢 (LDH) 釋放量的影響 圖 2 對細胞處理氰化鉀並培養於含糖或不含糖之培養液中 30, 60, 120 分 鐘發現,在含糖之培養液中 60, 120 分鐘後,細胞外 LDH 分別較控制組 增加 50﹪及 30﹪;而在不含糖之培養液中 60, 120 分鐘後,細胞外 LDH 分別較控制組增加 63﹪及 20﹪。由結果比較得知,缺氧並且缺糖(含 KCN,不含 glucose)的情況下培養 60, 120 分鐘後,細胞外 LDH 分別較缺 氧不缺糖組(含 KCN,含 glucose)增加 156﹪及 179﹪。此一結果明顯指 出,細胞缺糖的結果會加重化學性缺氧造成之神經細胞毒性。. 38.
(40) <三>、細胞生存力的影響 圖 3 細胞處理方法同結果<二>,存活細胞數目之測定利用 MTT 之分析。 在含糖之培養液中 30, 60, 120 分鐘後,存活細胞數目均較控制組減少約 20﹪;在不含糖之培養液中 30, 60 分鐘後,存活細胞數目分別較控制組 減少 9﹪及 37﹪,而 120 分鐘後,存活細胞數目幾乎無法測得。由結果 比較得知,缺氧並且缺糖(含 KCN,不含 glucose)的情況下培養 30, 60 分 鐘後,存活細胞數分別較缺氧不缺糖組(含 KCN,含 glucose) 減少 17﹪ 及 66﹪。此一結果可以看出,KCN 對細胞生存力的影響與細胞乳酸脫氫 脢的影響相同。 <四>、厚朴酚對細胞乳酸脫氫脢的影響 圖 4 為了研究厚朴酚對化學性缺氧引起細胞傷害的保護作用,對細胞處 理氰化鉀並且併用不同濃度厚朴酚,分析培養液中 LDH 含量。結果發現 厚朴酚對 KCN 引起的 LDH 有濃度依存性的減少作用,厚朴酚在 10 µM 及 100µM 濃度時分別較 KCN 組減少 17﹪及 36﹪;然而,750µM 濃度厚 朴酚不但未減少 LDH,反而引發較 KCN 組更大量之 LDH。由此結果可 得知,100µM 濃度厚朴酚最能有效減少 KCN 造成之神經細胞毒性,而過 量之厚朴酚反而對細胞有害。. 39.
(41) <五>、厚朴酚對細胞核的影響 圖 5 為了研究 KCN 造成的神經細胞死亡方式為凋零死(apoptosis)亦或 壞死(necrosis),我們利用螢光染色劑 DAPI 觀察比較細胞核有無分裂或 縮小情況,並計算完整細胞核的數目有無變化。缺氧(hypoxia:含 KCN, 不含 glucose)及厚朴酚對神經元及神經膠細胞細胞核的影響則同時分別 使用 MAP-2 及 GFAP 作免疫染色法標識。由染色結果可看出,GFAP 標 識之細胞無外形或細胞核上的變化,而 MAP-2 標識細胞之細胞核數目減 少,但沒有細胞核萎縮(condensation)的現象,此一結果可以說明細胞的 死亡應該是經由 necrosic 的過程,而非 apoptotic。厚朴酚對於細胞的保 護作用,則利用計算完整細胞核的數目有無變化,結果發現 KCN 培養 60, 120 分鐘後,細胞核的數目分別較控制組減少 57﹪及 51﹪,此一結果 與經 MAP-2 免疫標識的細胞減少數目相同。另一方面,於培養 60, 120 分鐘後,100µM 厚朴酚則增加細胞核的數目分別較 KCN 組高 50﹪及 121 ﹪。由此可見,厚朴酚保護細胞免於 KCN 的毒性作用與細胞乳酸脫氫脢 的結果一致。 <六>、厚朴酚對神經元細胞數目的影響 圖 6 對細胞處理氰化鉀並培養於不含糖之培養液中 120 分鐘後,神經元. 40.
(42) 細胞使用 MAP-2 作免疫染色法標識,明顯發現神經元細胞數目減少而且 神經結構受損,此氰化鉀引起之神經毒性可被厚朴酚所減弱。. 41.
(43) 圖1. Fig.1. Neuron toxicity in mixed cortical cultures following chemical hypoxia treatment. Fixed 14-day-old MAP-2-positive neurons (A) and GFAP-positive astrocytes (B) were treated with glucose-free vehicle media (C for MAP-2-positive and D for GFAP-positive) or KCN for 2 hr (E for MAP-2-positive and F for GFAP-positive). Scale bar = 50 µ m.. 42.
(44) 圖2. 156% 179% 63%. LDH content ( % of control). 20% **. 400. **. 350 300 250. 50%. 30%. 200 # # #. # #. 150 100 50 0. g lu c o se K C N. T im e (m in ). + -. + +. -. +. + + - +. 30. - - +. 6 0. + + - +. -. +. 12 0. Fig.2. Effect of glucose and KCN on cell viability by extracellular LDH activity. Primary mixed cultures were grown as described in Materials and Methods. On the day of the experiment, the media were removed, and the cultured cells were treated with KCN (0.5 mM) in glucose-free DMEM for the indicated times (30, 60 and 120 min). The release of LDH activity in the culture medium was determined and expressed as a percentage of vehicle (mean ±S.E.M., n=10). Asterisks indicate statistically significant increase in KCN-induced LDH release (**P<0.01 vs. glucose(-) and KCN(-); ##P<0.01, ###P<0.001 vs. glucose(+) and KCN(-)).. 43.
(45) 圖3. 66%. 17% 120. 17%. 83%. 24%. 17%. 110. MTT level (% of control). 100 ##. 90 80. 9%. #. **. 70 60. 37%. 50 40. ***. 30 20 10 0. glucose + + - - + - + K CN Tim e(min). + + - - + -+. 30. 60. ++ - - + - + 12 0. Fig.3. Effect of glucose and KCN on cell viability by MTT reduction. Primary mixed cultures were grown as described in Materials and Methods. On the day of the experiment, the media were removed, and the cultured cells were treated with KCN (0.5 mM) in glucose-free DMEM for the indicated times (30, 60 and 120 min). The amount of MTT formazan produced was expressed as a percentage of vehicle (mean±S.E.M., n=9). Asterisks indicate statistically significant reduction of KCN-induced formazan . [**P<0.01, ***P<0.001 vs. glucose(-) and KCN(-); #P<0.01, ##P<0.001 vs. glucose(+) and KCN(-)].. 44.
(46) 圖4. 500. ***. 36%. LDH content ( % of control). *** 17%. *. 200 ### 150 100 50. g lu c o se K C N M a g (µ M ). -. + -. + 10. + 100. + 750. Fig.4 Effect of magnolol (mag) on KCN-induced release of LDH in mixed cultured cells. Magnolol was dissolved in DMEM without glucose but with 0.1 % DMSO. Data were expressed as a percentage of vehicle (mean ±S.E.M.) from two separate experiments, each performed using five dishes (n=10). Asterisks indicate statistically significant increase in KCN-induced LDH release and significant decrease in LDH release in the presence of magnolol [###P<0.001 vs. glucose(-) and KCN(-), *P<0.05, ***P<0.001 vs. glucose(-) and KCN(+)].. 45.
(47) 圖 5-1. Fig. 5-2 The total numbers of fluorescent DAPI-stained nuclei following KCN or magnolol (Mag) treatment for 2 hr. Cells were seeded onto a 4-well chamber slide at 2×105 cells/well for 14 days, then treated with 0.5 mM KCN, 100 µM magnolol or a combination of the two for 1 and 2 hr. The number of cells counted by DAPI-staining in (A) and (B) for vehicle, (C) and (D) for KCN-treatment, and (E) and (F) for KCN combined with magnolol.. 46.
(48) 圖 5-2. 120. control 0.5 mM KCN 100 µM Mag + KCN. cell nuclei number ( % of control). 100 80 60. #. **. ##. 40 ** 20 0. 60 min. 120 min. Fig. 5-2 The total numbers of fluorescent DAPI-stained nuclei following KCN or magnolol (Mag) treatment for 2 hr. Cells were seeded onto a 4-well chamber slide at 2×105 cells/well for 14 days, then treated with 0.5 mM KCN, 100 M magnolol or a combination of the two for 1 and 2 hr. Data expressed as a percentage of vehicle (mean S.E.M.) from two separate experiments, each performed using five wells (n=10). Asterisks indicate statistically significant difference when compared with control groups [**P<0.001 vs. glucose(-) and KCN(-); #P<0.01,##P<0.001 vs. glucose(-) and KCN(+)].. 47.
(49) 圖6. Fig. 6 Neuron toxicity in cortical mixed cultures following KCN treatment. Primary mixed culture cells were grown in neurobasal containing B27 and PS for 14 days. The seeding density was 2.0×105 cells/well at chamber slide. Fixed 14-days-age cells were examined for MAP-2-positive neurons. Cell morphology after 2 hr. treatment with vehicle (A), KCN (B), and KCN plus magnolol (C) in glucose-free media. Bar = 50 µ m.. 48.
(50) 四、討論 由本章研究結果顯示,KCN 引起鼠腦皮質細胞的缺氧作用會造成細 胞外 LDH 釋放量的增加,並且使神經元以壞死( necrosis)的過程死亡。 而厚朴酚能夠減少 KCN 造成的神經細胞壞死作用。 學界將細胞死亡的方式概括分為凋零死(apoptosis)及壞死(necrosis) (Gerschenson-LE, 1992;Martin-LJ; 1998)。前者在正常成人組織之早期成 長及發育上扮演一重要角色,而其在形態學上之主要特徵為細胞核的斷 裂 (nuclear fragmentation)及細胞破裂 (cellular breakdown);後者則無細 胞核的特徵,但有粒線體腫脹現象細胞 (mitochondrial swelling),及喪失 細胞膜之完整性。從其它學者研究結果顯示,氰化物造成的細胞死亡的 方式,凋零死及壞死兼有之 (Tsujimoto-Y; 1997),由本章研究結果發現 KCN 造成的神經細胞死亡方式為壞死作用之傾向較大。 厚朴酚保護細胞免受缺氧性傷害的機轉並不清楚。例如,它可以作 為抗氧化劑來抑制氫氧自由基(hydroxyl free radical)的產生,藉此作用 來保護 Salmonella typhimurium TA102(Fujita-S, 1994);或是藉由抗炎 作用來減少 cytokine 的釋放(Wang-JP, 1992; 1995)。在周邊組織,厚朴 酚減緩小鼠後蹠之炎性浮腫,藉由降低 A23187 刺激產生之 PGE2 及 LTB4. 49.
(51) 在水腫液的含量(Wang-JP, 1995)。其可保護大鼠心臟粒腺體對抗脂質 過氧化的作用(Lo, 1994)。而對於經冠狀動脈結紮的大鼠有抗心律不整 及抗缺血的作用(Hong-CY, 1996)。然而,厚朴酚是經由何種機制來保 護細胞免於缺氧作用的損傷是接下來我們要探討的重點。. 50.
(52) 第肆章 氰化鉀對細胞內 inositol (1,4,5) trisphosphate (Ins (1,4,5) P3) 活性的影響 一、緒言 經由缺氧或缺血產生的神經變性機轉,包含鈣離子的衡定。而細 胞內游離鈣離子來源為細胞膜上 G 蛋白接受体受到活化,啟動細胞膜之 磷酸二肌醇 PIP2 形成 IP3 後,進而活化細胞內平滑性內質網(smooth ER) 上之 IP3 接受体,使平滑性內質網內貯存之鈣離子釋出至細胞內,而造 成細胞內游離鈣離子增加;大量鈣離子的產生造成細胞酵素活性過度興 奮,導致神經變性(neurodegeneration)。 化學性缺氧的結果已經實驗證實可造成體外培養之神經元細胞內 游離鈣離子的增加 (Gibson-G, 1991)。在體外實驗的結果顯示,厚朴酚能 經由三磷酸肌醇 (IP3) 路徑引發嗜中性白血球胞質內游離鈣離子含量增 加 (Wang-JP, 1998)。而由我們前面研究結果顯示厚朴酚能降低化學性缺 氧對於體外培養之大鼠大腦皮質神元膠質細胞之傷害,因此,本次實驗 接下來要探討化學性缺氧對混合型培養細胞之磷酸肌醇接受體的影響, 以及厚朴酚是否係經由磷酸肌醇-鈣離子路徑活化神經細胞生理活性避 免傷害。. 51.
(53) 二、材料與方法 <一>、細胞培養 實驗中所使用的細胞材料相同於第貳章之「二、材料與方法」中「細 胞培養」之方法。將細胞培養於 3.5 cm 圓形培養皿中,細胞密度為 9.6 × 105 cells/dish。於 5% CO2,37℃之恆溫培養箱中培養至十四天後即可供 實驗之用。 <二>、放射線[3H]分析法分析 IP3 1-1. 細胞膜的製備 用於結合 IP3 的細胞膜材料取自雄性 Sprague-Dawley 成鼠(體重約 200 克)之小腦。將清醒狀況下大鼠以極快之速度斷頭取出小腦部位,並 清除腦上之其它組織後,置入冰冷之三十倍體積之緩衝液 A 註一中以均質 機均質(1500rpm)。將均質液以 13,000 rpm 之速度於 4℃下離心 15 分鐘, 倒去上清液,將沉澱再次懸浮於緩衝液 A 中,再離心,如此重復兩次後, 將最後之沉澱懸浮於十倍體積之緩衝液 A 中,即製成細胞膜懸浮液,分 裝並貯存於-70℃。 1-2. 細胞膜懸浮液之蛋白質含量測定. 52.
(54) 進行放射線[3H]分析法前,試樣均先進行蛋白質含量(protein assay) 測定,以調整分析試樣時有一致含量之蛋白質(3 mg/ml)。根據 「Lowry method」,取 50 µl 試樣溶液置於 plastic curvette 中,加入 2.5 ml Bio-Rad 蛋白質染料試劑(dye reagent),混合後靜置 30 分鐘呈色,於 595 nm 波長 下測其吸光值。蛋白質含量標準曲線由 Bovine Serum Albumin (BSA) 所 配製之標準溶液而得之。 2-1. 實驗方法及細胞內 IP3 之抽取 實驗當天吸去細胞培養液,以 PBS 溶液清洗細胞一次,全部實驗過 程均在 37℃之水浴槽下操作。吸乾細胞清洗液,加入 DMEM 含 glucose 不含 serum 之培養液,於 37℃之水浴槽靜置 4 分鐘,加入不同藥物與細 胞一同培養 (0, 5, 10, 30 sec)。時間到時,立即將細胞培養皿移至冰盤上 放置以終止反應,同時並吸乾培養液,隨即加入 0.5 ml 冰冷之 TCA 溶液 註二. ,細胞培養皿連同冰盤置入 4℃冰箱中靜置 60 min,以將細胞內 IP3. 抽提至 TCA 溶液中。將 TCA 抽提液吸起置入塑膠試管內,而留在細胞 培養皿上之細胞則以蒸餾水浸洗兩次後留作蛋白質含量分析用(步驟 2-2)。TCA 抽提液必須以水飽合之 diethyl ether. 註三. 浸洗過,以將 TCA 萃. 取至 ether 溶液內,留下 IP3 於水層當中。加入兩倍體積之 diethyl ether 至試管內,振盪混合十秒後靜置使溶液分層,吸取上層 ether 溶液,再加. 53.
(55) 入 diethyl ether 至試管內重覆浸洗四次,殘留之 ether 溶液可以氮氣在室 溫下吹乾。吸取剩留於試管內之水溶液,置於 eppendorf tube 中,加入十 分之一體積之 Tris-base 溶液(500 mM, pH=8.4)中和之,混合均勻後置入 -70℃中貯存。 2-2. 細胞之蛋白質含量測定 進行放射線[3H]分析前,細胞均先行進行蛋白質含量(protein assay) 測定,以調整分析結果有一致含量之蛋白質。將以 TCA 溶液抽提過 IP3 之細胞培養皿加入 0.5 ml 之 1N NaOH 溶液於 45℃下靜置 30 min,則附著 於細胞培養皿上之細胞會脫離培養皿而懸浮於 NaOH 溶液中,吸取所有 細胞溶液分別置入 eppendorff tube 中,於 60℃水浴槽內加熱 30 min,之 後待試樣冷卻則進行蛋白質含量(protein assay)測定。根據「Bio-Rad method」,取 50 µl 試樣溶液置於 plastic curvette 中,加入 2.5 ml Bio-Rad 蛋白質染料試劑 (dye reagent),混合後靜置 30 分鐘呈色,於 595 nm 波 長下測其吸光值。蛋白質含量標準曲線由 Bovine Serum Albumin (BSA) 所配製之標準溶液 (0-0.6 mg/ml)而得之。 3. 放射線[3H]分析法分析 IP3 分析過程均於 4℃之冰盤上操作。準備 eppendorff tube,依序加入下. 54.
(56) 列溶液:50 µl Buffer B, 100 µl 細胞試樣或標準 IP3. 註五. 溶液,30 µl 之[3H]. 標誌 IP3 (11 nM),95µl 之小腦細胞膜懸浮液 (60-100 µg protein) 而成反 應混合液。反應混合液於 4℃冰浴下反應 15 min,之後離心 (4℃, 14,000×g, 5 min),吸去上清液,沉澱再次懸浮於 0.5 ml 之 1 N NaOH 溶液中,於 60 ℃下消化 30 min,加入 0.1 ml 之 0.5 N HCl 溶液中和之;最後加入 5 ml 之訊號放大液 (scintillation fluid, amplified agent),即可偵測試樣之放射 線強度。(Tri-carb 1500, Liguid Scintillation Analyser) 註一:緩衝液 A 之組成份為:50 mM Tris-HCl,1 mM EDTA, 1 mM 2-mercaptoethanol; 新鮮配製,酸鹼值調至 7.7。. 註二: TCA 溶液濃度為 1 M,溶於滅菌蒸餾水中,並置於 4℃下避光貯存。. 註三: ether 必須先以水飽合過,以避免處理過程 ether 會吸收細胞水份。. 註四:Buffer B 組成份為:50 mM Tris-HCl, 1 mM EDTA, 1 mM 2-mercaptoethanol; 新鮮配製,酸鹼值調至 8.4。. 註五:標準 IP3 溶液以 Buffer B 溶液配製,配製濃度為 0-1100 picoMolar。. <三>、藥物或化學物 Carbachol, glutamate, potassium chloride (KCl), trichloroacetic acid (TCA), Tris-base 購 自 Sigma Chemicals Co. (St. Louis, MO, USA).. 55.
(57) Potassium cyanide (KCN) 購自 Merk Chemicals (Darmstadt, Germany)。 Ins(1,4,5)P3 購自 Calbiochem (San Diego, CA, USA);[3H]Ins(1,4,5)IP3 購自 England Nuclear (Boston, MA, USA) <四>、統計分析 結果之表示以平均值加減標準誤差(mean±s.e.m.),並以細胞培養 之次數為平均。為單因子變異數分析時,統計上平均值之差異以 Student’s t-test 決定之。為多重因子變異數比較時,結果之分析以 one-way ANOVA 表示之。P 值小於 0.05 稱為有顯著統計上之差異。. 56.
(58) 三、結果 <一>、氰化鉀對細胞內之 IP3 活性的影響 圖 1 為了了解混合型培養細胞內之 IP3 活性對藥物刺激細胞膜上接受體 的反應,實驗中使用幾種藥物以活化細胞膜,包括乙醯膽鹼接受體活化 劑(carbachol)、興奮性胺基酸(glutamate)、鉀離子(KCl)以及 bradykinin, 並比較這些細胞膜活化劑與氰化鉀對 IP3 接受體的反應。IP3 接受體之活 性以細胞合成之 IP3 活性 (IP3 activity) 表示之。對細胞處理 100 µM carbachol 或 0.5 mM KCN 或 100 µM glutamate 或 50 mM KCl 或 1 µM bradykinin,培養 5, 10, 30 秒後分析細胞內 IP3 活性。實驗結果發現 KCN 或 glutamate 或 KCl 或 bradykinin 對 IP3 接受體均無明顯刺激反應,但 carbachol 培養 5 秒時即可達最大提高細胞 IP3 活性至 0 秒時之 85%,5 秒 後 IP3 活性則漸降;而 KCN 併用 carbachol 培養 5 秒時更可較單獨使用 carbachol 時增加細胞 IP3 活性至 14% (p<0.05 vs. carbachol)。由結果可看 出,細胞內 IP3 活性可由刺激細胞膜上乙醯膽鹼接受體所激活,而當細 胞受到缺氧性 (KCN) 傷害時,受到乙醯膽鹼接受體所激活的 IP3 活性會 更高。 <二>、厚朴酚對細胞內之 IP3 活性的影響. 57.
(59) 圖 2 為了研究 magnolol 對細胞內之 IP3 活性的影響,實驗中對細胞處理 100 µM carbachol 或 0.5 mM KCN 或 100 µM magnolol,培養 5, 10, 30 秒後 分析細胞內 IP3 活性。實驗結果發現 magnolol 可持續性地提高 carbachol 活化的細胞 IP3 活性至 21%, 30%, 26% (相對於 carbachol 併用 KCN),而 IP3 活性仍於 5 秒時達到最高點。由結果可看出,magnolol 可藉由激活細 胞膜上乙醯膽鹼接受體而提高細胞內 IP3 活性。. 58.
(60) 1,4,5-IP3 activity (% of basal). 圖1. carbachol 100 µM KCN0.5 mM carbachol + KCN glutamate 100 µM KCl 50 mM Bradykinin 1 µM. 250. * 200. 150. 100. 0. 5. 10 Time (sec). 30. Fig. 1 Activities of IP3 receptor stimulated by various stimuli including KCN, carbachol, glutamate, KCl and bradykinin as indicated by ativities of IP3, in the glucose-free DMEM media. Plots indicate IP3 values (mean ± SEM., n=8) in DMEM media at 0, 5, 10, 30 sec. Asterisks indicate statistically significant (*P<0.05 by Student's t-test following one-way ANOVA).. 59.
(61) 1,4,5-IP3 activity (% of basal). 圖2. carbachol 100 µM KCN0.5 mM carbachol + KCN magnolol 100 µM mag + car + KCN. 300. # 250. * 200. 150. 100. 0. 5. 10 Time (sec). 30. Fig. 2 Effect of magnolol (mag) on KCN plus carbachol (car) induced IP3 ativities in the glucose-free DMEM media. Plots indicate IP3 values (mean ± SEM, n=8) in DMEM media at 0, 5, 10, 30 sec. Asterisks indicate statistically significant (*P<0.05 vs. carbachol, #P<0.05 vs. KCN plus carbachol by Student's t-test following one-way ANOVA).. 60.
(62) 四、討論 化學性缺氧 (KCN) 的結果造成體外培養大鼠腦皮質細胞內 Ins (1,4,5) P3 的增加,此增加為經由活化 ACh 接受體所致,而細胞在經過厚 朴酚的處理後,細胞內 Ins (1,4,5) P3 的量會持續性地增加較 KCN 更多。 由本章研究結果來看,KCN 所激活之 Ins (1,4,5) P3 與 ACh 接受體 有直接之關係,而厚朴酚對 KCN 作用的影響則是提高 ACh 接受體的作 用,值得思考的是,厚朴酚是增加突觸前 ACh 的合成亦或釋放?或是增 加突觸後 ACh 接受體的活性?可提供研究厚朴酚藥理作用的一個新方 向。唯從以前的研究文獻來看,與厚朴酚植物來源相同且化學結構相似 之另一成份 honokiol,經實驗證實可經由鉀離子刺激引起大鼠海馬迴 (hippocampus) 釋放 ACh ,但厚朴酚卻無此一作用(Tsai-TH, 1995),我們 推測在本論文的實驗環境下,厚朴酚或許會表現出刺激突觸前釋放 Ach 的作用。 化學性缺氧的結果已經實驗證實可造成體外培養之神經元細胞內 游離鈣離子的增加 (Gibson-G, 1991),有學者研究指出厚朴酚能經由 Ins (1,4,5) P3 路徑引發嗜中性白血球胞質內游離鈣離子含量增加 (Wang-JP, 1998)。在本篇論文中雖無法直接測出厚朴酚對細胞質內游離鈣離子含量. 61.
(63) 的影響,但由本研究結果來看,厚朴酚增加細胞內 Ins (1,4,5) P3 的活性, 我們推測厚朴酚應有類似對嗜中性白血球細胞的作用,會增加腦皮質細 胞質內游離鈣離子的含量。但此一作用結果應與 KCN 同樣增加胞質內游 離鈣離子的含量卻造成細胞變性之影響不同,我們推測厚朴酚增加胞質 內游離鈣離子,對維持細胞正常生理功能有正面之貢獻,藉此以保護免 於缺氧性功能之喪失,但此一方面之影響則有待實驗進一步確認。. 62.
(64) 第伍章 厚朴酚對氰化鉀引起細胞前列腺素 E2 與一氧化 氮反應之影響 一、緒言 經由缺氧或缺血產生的神經變性機轉,大致包含下列情形一鈣離子的 衡定、神經傳遞物質的釋放、缺氧性壓力以及發炎反應。化學性缺氧模 式對於探討神經變性的分子機轉是相當便利且有效的(Gibson et al., 1991)氰化鉀會干擾電子鏈的傳遞而阻斷呼吸作用(Jones et al., 1984) 已知對神經元會造成不可逆的改變,而此種改變與缺氧之結果極為相 似。因此,施以氰化物常被使用為模擬化學性缺氧的模式。為了研究細 胞因缺氧而導致傷害或死亡,以氰化鉀引發細胞類似缺氧之功能性傷害 是一個便利的方法。(Gibson, 1991; Golberg, 1989; Lee, 1998) 在花生四烯酸之各種代謝產物中,PGE2 在產生腦部或神經元傷害的 發炎性反應過程中扮演重要環節,在化學性缺氧( Yang, 1994)、缺血性 缺氧(Yoshida, 1980; Abe, 1988; Ellis, 1981)及細菌性脂多醣體(LPS) 引發的炎性反應中亦同。除 PGE2 外,一氧化氮(NO)在炎性反應的過 程中亦扮演指標性角色(Boneh, 1997; Molina, 1995)。 在體內實驗的結果顯示,厚朴酚能減少因缺血後再灌流(reperfusion). 63.
(65) 引起的大鼠心臟部位梗塞壞死面積(Hong-CY, 1996)。在體外實驗中, 厚朴酚能保護大鼠心臟細胞,提升細胞內粒腺體對抗脂質過氧化的作用 (Lo-YC, 1994)。此外,皮下注射厚朴酚能降低前列腺素 E2(PGE2)及 白三烯素(LTB4)含量而減少因 A23187 所引起之大鼠後蹠浮腫程度 (Wang-JP, 1995)。而本次研究結果顯示厚朴酚能降低化學性缺氧對於 體外培養之大鼠大腦皮質神元膠質細胞之傷害,因此本次實驗接下來要 探討厚朴酚是否係經由對抗缺氧引發之發炎介質 PGE2 及 NO 的釋放而 保護神經細胞避免傷害。. 二、材料與方法 <一>、細胞培養 實驗中所使用的細胞材料相同於第貳章之「材料與方法」內「細胞 培養」中所述。 <二>、細胞釋放前列腺素 E2 之測定 前列腺素 E2 之測定乃是使用商業製劑免疫分析系統 (immuno assay kit)(R&D system)。將培養至 14 日齡,平貼於培養皿之細胞自培養箱 取出,置放於水浴恒溫槽內(37℃)以 DMEM 含 20mM Hepes 之培養液 潤洗兩次後,分別加入 0.5mM KCN, 100ng/ml LPS 或 100µM magnolol, 64.
(66) 實驗用藥之配製溶液均使用 DMEM 含 20mM Hepes 之培養液。細胞加入 藥物後於 37℃水浴槽分別放置 30,60,120 分鐘。培養時間到時,吸取 培養皿內之培養液至 ependorff tube 內,以冷凍離心機於 4℃,14,000×g 的速度下離心 5 分鐘。取出細胞上清液,保存於-70℃冷凍櫃中,直至分 析實驗開始。作 PGE2 之分析時,取出細胞上清液 100µl 置於 kit 內預備 好之微量滴盤之孔槽內,PGE2 標準曲線與實驗同步進行。反應最後呈色 之吸光值,以光譜計(spectrophotometer)(MRX, Dynateek Laboratories) 在 405nm 下測其吸光值,參考波長(reference wavelength)為 630nm。 實驗結果數值由 PGE2 標準曲線換算得知。結果以 pg/ml 表示之。 <三>、細胞生成一氧化氮之測定 一氧化氮之測定乃是使用硝酸鹽/亞硝酸鹽製劑分析系統(Cayman’s Nitrate/Nitrite assay Kit)(Cayman Chemical Company, Ann Arbor, MI USA.)(Nitrite 靈敏度大於 1.0 µM)當神經元或神經膠細胞受到發炎刺 激或細胞恒穩狀態受到破壞時,會產生極少量之一氧化氮,因此極難即 時偵測。而經過細胞內的反應,一氧化氮的最終產物為硝酸鹽及亞硝酸 鹽。但轉變成硝酸鹽或亞硝酸鹽之比例是無法預測的,因兩者彼此可互 相轉化。因此,測定一氧化氮總產量的方法必須是一併計算硝酸鹽及亞 硝酸鹽的總量。首先,使用硝酸還原脢(Nitrate reductase)將 Nitrate 還. 65.
(67) 原成 Nitrite,其次,加入呈色試劑(Griess Reagent)使 Nitrite 與之形成 一深紫色沉澱(azo compound)。由呈色反應在 540nm 之吸光值可精確 測定 Nitrite 之含量。 分析實驗開始時,取 80µl 細胞上清液(實驗方法同(三)PGE2 之測定) 置於 96 孔洞之微滴平盤中,接著加入 10 µl 之硝酸還原脢及 10 µl 之輔脢(co-factor) ,混合後於室溫下反應二小時。之後加入 Griess Reagent 後隨即呈色。在 540nm 下測吸光值。而標準曲線由已知亞硝酸鹽濃度測 其吸光值而得到,濃度結果以 micromolar(µM)表示之。 <四>、藥物或化學物 DMSO, CaCl2, EDTA, papain, deoxyribonuclease I (DNas I), Hanks Balanced. Salt. Solution. (HBSS),. cysteine,. N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic Lipopolysaccharide. (LPS),. D-(+)-glucose,. poly-D-lysine,. acid boric. acid,. (Hepes), penicillin,. streptomycin, NG-nitro-L-arginine methl ester dihydrochloride (L-NAME) 購自Sigma Chemicals Co (St. Louis, MO, USA)。horse serum 購自 Hyclone (Logan, Utah, USA) 。 Dulbecco's modified Eagle's (DME) medium, neurobasal, B27 supplement 購自GibcoBRL (Grand Island, NY, USA) 。 lactate dehydrogenase (LDH) assay kit 購 自. 66. Boehringer Mannheim.
(68) (Germany) 。potassium cyanide (KCN) 購自 Merk Chemicals (Darmstadt, Germany) 。 <五>、統計分析 結果之表示以平均值加減標準誤差(mean±s.e.m.),並以細胞培養 之次數為平均。為單因子變異數分析時,統計上平均值之差異以 Student’s t-test 決定之。為多重因子變異數比較時,結果之分析以 one-way ANOVA 表示之。P 值小於 0.05 稱為有顯著統計上之差異。. 67.
(69) 三、結果 <一>、在含糖培養液情況下,乳酸脫氫脢與細胞生成之前列腺素 E2 的變 化 圖 1 為了研究是否化學性缺氧的傷害會造成細胞膜完整性的改變並且影 響前列腺素的代謝產物(炎性反應),我們分析了處理後細胞培養液中乳 酸脫氫脢 (LDH) 的活性及前列腺素 E2 (PGE2) 的變化,並比較含糖培養 液能否影響其作用。對細胞處理 KCN 或 magnolol 培養 60 分鐘發現,經 KCN 處理之細胞 LDH 及 PGE2 會分別較控制組增加 25% (p<0.01) 及 41 ﹪(p<0.05),而經 magnolol 併用 KCN 處理之細胞 LDH 及 PGE2 會分別較 KCN 組減少 27% (p<0.05) 及 42﹪(p<0.001)。此結果指出即使在含糖培養 液下 KCN 仍然會造成細胞缺氧性的傷害,包括 LDH 及 PGE2 的合成增 加,magnolol 能有效保護細胞減少此類物質之產生。 <二>、在不含糖培養液情況下,細胞生成之乳酸脫氫脢的變化 圖2 為了比較細胞在缺糖狀況下的反應,在不含糖培養液情況下我們分析 細胞生成之LDH,並且使用炎性反應之刺激物LPS,以了解炎性刺激是否 會加重KCN引起的細胞損傷。對細胞處理0.5 mM KCN或100ng/ml LPS或 100µ M magnolol或10 µ M L-NAME培養30, 60, 120分鐘發現,經藥物處理30. 68.
(70) 分鐘時並無明顯變化。而在60, 120分鐘時,KCN處理之細胞LDH會分別較 控制組增加32% (p<0.01) 及8﹪(p<0.01);而經LPS單獨處理之細胞LDH反 而會分別較控制組減少19% (p<0.01)及3﹪(p>0.05);若LPS併用KCN,則 細胞LDH增加之程度與單獨使用KCN時相近;經magnolol併用氰化鉀處理 之 細 胞 LDH 會 分 別 較 KCN 組 減 少 35% (p<0.01) 及 16 ﹪ (p<0.001) ;若 magnolol 併用 LPS 及 KCN ,細胞 LDH會分別較 LPS 併用 KCN 組減少 30% (p<0.001) 及 11﹪(p<0.01)。為了了解NO的產生是否包括在LPS併用KCN引 起的細胞損傷當中,實驗中在變化最明顯的60分鐘時加入NO合成抑制劑 L-NAME,結果發現L-NAME併用LPS及KCN並不會減少LPS併用KCN所產 生之LDH。 <三>、在不含糖培養液情況下,細胞生成之前列腺素 E2 的變化 圖3 對細胞處理0.5 mM KCN或100ng/ml LPS或100µ M magnolol或10 µ M L-NAME培養30, 60, 120分鐘發現,經藥物處理30分鐘時並無明顯變化。而 在60, 120分鐘時,KCN處理之細胞PGE2會分別較控制組增加13% (p<0.05) 及4﹪(p>0.05);而經LPS單獨處理之細胞PGE2並無明顯變化反而會分別較 控制組減少 19% (p<0.01) 及 3﹪ (p>0.05) ;若LPS併用KCN,則LPS會加重 KCN引起之 PGE2 的程度 (21% 對 12% 及 19% 對6%,分別比較於單獨 使用 KCN及 LPS) ;經 magnolol併用氰化鉀處理之細胞PGE2會分別較KCN. 69.
(71) 組減少 26% (p<0.001) 及 57﹪ (p<0.01) ;若 magnolol併用LPS及KCN,細胞 PGE2會分別較LPS併用KCN組減少40% (p<0.001)及58﹪(p<0.01)。為了了 解 NO 的產生是否包括在 LPS 併用KCN引起的細胞炎性反應當中,實驗中 在變化最明顯的60分鐘時加入NO合成抑制劑L-NAME,結果發現L-NAME 併用LPS及KCN並不會減少LPS併用KCN所產生之PGE2。由圖1、圖2及圖 3的結果比較來看,缺糖培養液(hypoglycemia) 確實會加重缺氧(hypoxia) 引 起的炎性反應(inflammation),而由LDH及PGE2的平行反應來看,PGE2的 產生應該與細胞裂解(cell lysis)有關。由圖2及圖3的結果來看,控制組之 LDH及PGE2 釋放量隨時間而增加,可見在缺糖培養液的情況下,細胞也 有炎性反應。而我們也注意到,在 120分鐘的PGE2釋放量,於控制組、KCN 組及 KCN併用 LPS 組之間並無顯著差異;此一結果指出,在120分鐘的時 後,PGE2的產生主要在影響缺糖(hypoglycemia) 的效果。因此,magnolol 在 120 分鐘時顯著地阻斷 KCN或 KCN併用 LPS 引起的 PGE2 產生,也暗示了 magnolol對缺糖引起的PGE2產生也有阻斷的效果。 <四>、在缺糖情況下,細胞生成之一氧化氮的變化 圖 4 為了研究是否一氧化氮路徑包括在 KCN 或媒介 LPS 刺激引起的發 炎反應並且與 magnolol的保護作用有否關聯,我們測量了細胞外液中 NO 的總產量(結果以終產物 nitrite 的含表示之)。對細胞處理 0.5 mM KCN 或. 70.
(72) 100ng/ml LPS 或 100µM magnolol 或 10 µM L-NAME 培養 30, 60, 120 分鐘 發現,單獨處理 KCN 或 LPS 直至 120 分鐘都沒有刺激 NO 的產生,然而, 若合併 KCN 及 LPS 同時處理 30, 60, 120 分鐘可引起 NO 的增加分別為 99%, 70% and 87% (與 KCN 處理相比),以及 62%, 63%, 77% (與 LPS 處 理相比)。這些結果指出 KCN 以及 LPS 之間的交互作用可能在加強 NO 的生成上扮演了某些角色。此外,KCN 媒介 LPS 刺激引起 NO 的增加可 被 magnolol 或 L-NAME 減弱。值得注意的是,雖然 L-NAME 可抑制 NO 的增加,但對 LDH 以及 PGE2 的釋放卻無法影響。. 71.
(73) 圖1. 2 5. *. LDH release ( mU/ml). **. A. 2 0 1 5 1 0 5 0. *. 8 0 0. *. PGE2 concentration ( pg/ml). 7 0 0. B. ***. 6 0 0 5 0 0 4 0 0 3 0 0 2 0 0 1 0 0 0. b a sa l. co n. K C N. m a g + K C N. Fig. 1 Effect of magnolol (mag) on KCN-induced cell cytotoxicity and arachidonate cascades as indicated by LDH and PGE2 releases, respectively, in the DMEM media which contained of glucose (normal glycemia). Bars indicate LDH and PGE2 values (mean ± S.E.M., n=8) in DMEM media at 0 time (basal); after 60 min without treatment (con); with KCN; and magnolol plus KCN. Asterisks indicate statistically significant (*P<0.05, **P<0.01, ***P<0.001 by Student's t-test following one-way ANOVA).. 72.
(74) 圖2. LDH release (mU/ml). 50. control KCN 0.5 mM LPS 100 ng/ml LPS + KCN. 40. *** ** ** ***. ** ** *** **. 30 20. mag 100 µM+KCN mag + LPS + KCN L-NAME +LPS + KCN. ***. 10 0. 30. 120 60 Treated time (min). Fig. 2. Effects of magnolol treatment on LDH release induced by LPS + KCN in glucose-free culture media. Bars indicate mean ± S.E.M. (n=8). *P<0.05, **P<0.01, ***P<0.001 by Student's t-test following one-way ANOVA. 73.
(75) 圖 3. m ag 100 µ M + K C N m ag + L PS + K C N L -N A M E + L P S + K C N. c o n tro l K C N 0 .5 m M L P S 1 0 0 n g /m l LPS + KCN. ** * * ** * * **. 6 00. **. 5 00. **. **. *. 4 00 3 00. 2. PGE release (pg/ml). 7 00. 2 00 1 00 0. 30. 60 120 T r e a t e d ti m e ( m in ). Fig. 3 Effects of magnolol treatment on PGE2 release induced by LPS + KCN in glucose-free culture media. Bars indicate mean ± S.E.M. (n=8). *P<0.05, **P<0.01, ***P<0.001 by Student's t-test following one-way ANOVA. 74.
(76) 圖 4. 6. c o n tro l KC N LPS LPS + KC N m ag + LPS + KC N L -N A M E + L P S + K C N. **. 5. ***. Nitrite content (µM). *** 4. # #. ## 3. ##. ###. ### 2. 1. 0. 30. 60. 120. T r e a te d t im e ( m in ). Fig. 4. Effects of magnolol treatment and L-NAME treatment on KCN+LPS induced nitric oxide generation in glucose-free culture media. Nitric oxide generation was indicated by extracellular nitrite (NO 2-) content. Bars indicate mean ± S.E.M. (n=8). Asterisks indicate statistically significant. **P<0.01, ***P<0.001 vs. KCN; #P<0.05, ##P<0.001, ###P<0.001 vs. LPS+KCN by Student's t-test following one-way ANOVA.. 四、討論 75.
(77) 本部份為討論化學性缺氧引發之炎性反應是否可因厚朴酚而減輕。依 照先前的研究結果(Lee-MM, 1998)以及其他研究者(Millis,1996)的報 告均證實化學性缺氧直接造成神經元的傷害。由本章實驗結果顯示,在 化學性缺氧期間,細胞持續釋放出 LDH 及 PGE2,且 PGE2 的釋放和 LDH 有相當之關聯。在已知的化學性缺氧造成的神經元傷害反應中,包括炎 性 物 質 的 釋 放 ( Lerond, 1994 ; Arya, 1996 ) 、 自 由 基 離 子 的 產 生 (Akira,1994;Maiese, 1993)。而 PGE2 已被公認與腦部炎性反應的產生 有極大的關聯(Engelhardt, 1996)。以混合培養的鼠腦皮質細胞為實驗 的結果顯示,KCN 合併 LPS 使用的結果,較單獨使用 KCN 時產生更大 量的 PGE2;但兩者在於產生 LDH 的效應上則不分軒輊。因此,認定 PGE2 在化學性缺氧期間的炎性反應中是造成細胞死亡的重要因素是合理的推 論。另一方面,實驗結果顯示,厚朴酚能減少因 KCN 而引發之 PGE2 及 LDH 的釋放。因此,厚朴酚能經由抑制炎性物質的產生而保護細胞、降 低缺氧對於細胞的損傷。 此外,在單獨使用 LPS 時可減少細胞於缺醣狀態下釋放的 LDH,但 單獨使用 LPS 或 KCN 時則均增加培養液中 PGE2 的含量。此部份結果似 乎暗示 LPS 能夠保護細胞、降低缺氧對於細胞的損傷,雖然 LPS 合併 KCN 的處理並不會減少細胞 LDH 的釋放。這部份的結果似乎與許多研究. 76.
數據



+7


Outline
相關文件
• 由於細胞代謝、紫外線的電離輻射、不良的生 活習慣 ( 煙、酒、肥胖 ) 、各種感染物所引起 的中性粒細胞、巨噬細胞的激活, 會催化分子 氧發生單價還原產生的 "
Before along with the evolution of time, the development of science and technology, the progress of cell phone, the cell phone that we used, from the beginning has the basic
Now given the volume fraction for the interface cell C i , we seek a reconstruction that mimics the sub-grid structure of the jump between 0 and 1 in the volume fraction
• 是細胞不正常增生,且這些增生的細胞可
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
− The decision of the preferred treatment modality (either closed or open reduction) depends on the time elapsed since injury, the degree and type of dislocation, other
Animal or vegetable fats and oils and their fractiors, boiled, oxidised, dehydrated, sulphurised, blown, polymerised by heat in vacuum or in inert gas or otherwise chemically
2.滴加蒸餾水或亞甲藍液製成水埋標本 3.觀察 石細胞 (厚壁細胞)及
