玉蘭花、檄樹及咖啡等植物對基質金屬蛋白酶及彈性蛋白酶作用之探討; The effect of Michelia alba, Morinda citrifolia and Coffea arabica on MMPs and elastase
113
0
0
全文
(2)
(3) 目 錄 目錄………………………………………………………………………I 附圖目錄………………………………………………………………IV 附表目錄………………………………………………………………VI 中文摘要………………………………………………………………VII 英文摘要………………………………………………………………IX 第一章 緒言……………………………………………………………1 第二章 總論……………………………………………………………3 第一節 皮 膚 生 理 結 構 … … … … … … … … … … … … … … 3 第二節 膠原蛋白………………………………………………6 第三節 基質金屬蛋白酶(MMPs)…………………………………10 第四節 光老化……………………………………………………13 第五節 玉蘭花、檄樹與咖啡之文獻考察………………………24 第三章 材料與方法………………………………………………30 一、 實驗材料……………………………………………30 二、 實驗方法……………………………………………35 (一) 咖啡、玉蘭花及檄樹之葉甲醇萃取物之製備……35 (二) 明膠分解試驗…………………………………………36 (三) 膠原蛋白酶抑制試驗…………………………………38. I.
(4) (四) 彈力蛋白酶抑制試驗…………………………………39 (五) 細胞培養……………………………………………40 (六) 纖維母細胞之 UVB 照射試驗………………………41 (七) 分離細胞內細胞質之蛋白質…………………………42 (八) 西方墨點法……………………………………………43 (九) 細胞毒性試驗………………………………………45 (十) 皮膚一次刺激性試驗…………………………………46 (十一). 眼球刺激性試驗…………………………………47. (十二). 統計方法………………………………………47. 第四章 結果…………………………………………………………48 一、 植物萃取及水解物製備……………………………48 二、 膠原蛋白酶活性抑制之試驗………………………48 三、 彈力蛋白酶抑制試驗………………………………50 四、 抑制基質金屬蛋白酶(MMP-1, -3, -9)表現…………50 五、 咖啡及玉蘭花萃取物之安全性評估………………54 第五章 討論……………………………………………………………57 第六章 結論與建議……………………………………………………66 第七章參考文獻………………………………………………………67 圖……………………………………………………………………77. II.
(5) 表……………………………………………………………………94 附錄……………………………………………………………………98. III.
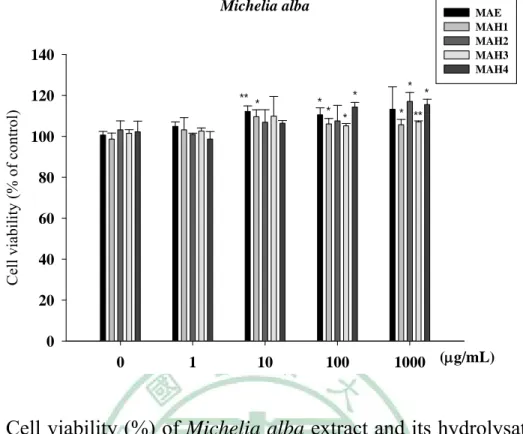
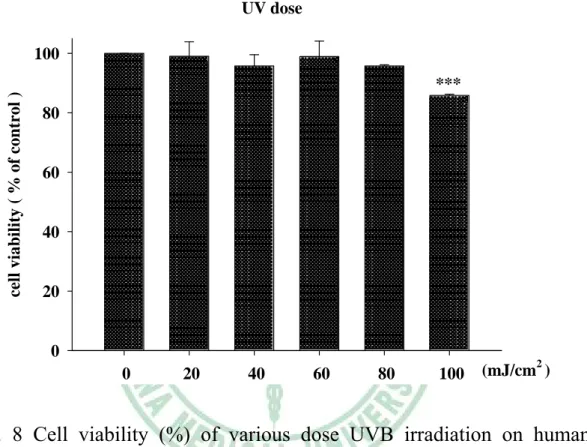
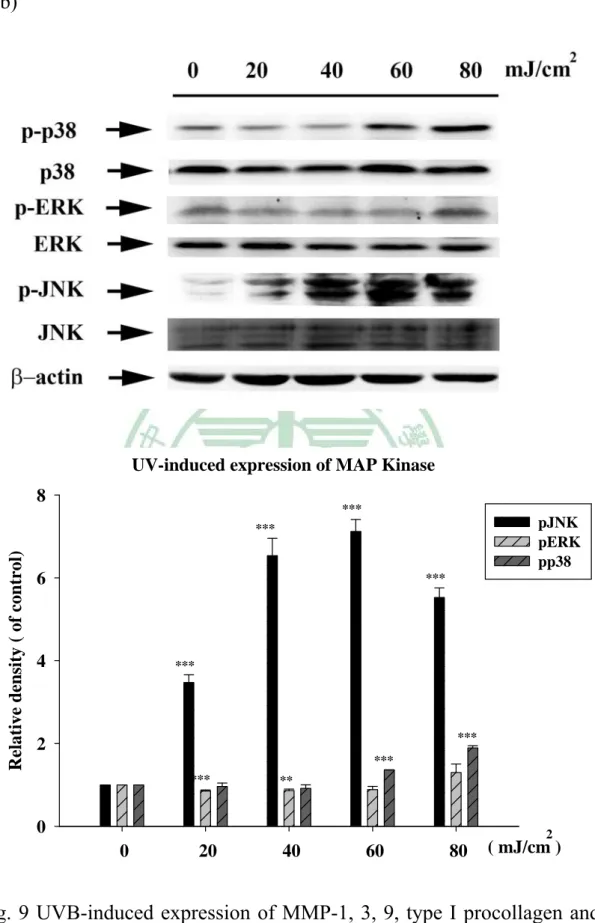
(6) 附 圖 目 錄 Fig.1 The inhibition of Coffea arabica, Michelia alba and Morinda citrifolia extract and their hydrolysates on collagenase activity.... ………………………………………………………79 Fig. 2 The inhibition (%) of Coffea arabica extract and its hydrolysates on the activity of bacterial collagenase..………………………80 Fig. 3 The inhibition (%) of Michelia alba extract and its hydrolysates on the activity of bacterial collagenase.…………………………80 Fig. 4 The inhibition (%) of Michelia alba extract and its hydrolysates on the activity of porcine elastase..………………………………...81 Fig. 5 Cell viability (%) of Coffea arabica extract and its hydrolysates on human foreskin fibroblast. ……………………………………81 Fig. 6 Cell viability (%) of Michelia alba extract and its hydrolysates on human foreskin fibroblast..………………….………………...82 Fig. 7 Cell viability (%) of chlorogenic acid and caffeic acid on human foreskin fibroblast..…………………………………………….82 Fig. 8 Cell viability (%) of various dose UVB irradiation on human foreskin fibroblast.……………………………………………83 Fig. 9 UVB-induced expression of MMP-1, 3, 9, type I procollagen and MAP kinase. ……………………………………………………85 Fig. 10 Effect of Coffea arabica extract on the UVB-induced expression of MMP-1, 3 and 9 and type I procollagen in human fibroblast.86 Fig. 11 Effect of Coffea arabica extract on the UVB-induced expression of MAP kinases in human fibroblast. …………………………87 Fig. 12 Effect of caffeic acid and chlorogenic acid on the UVB-induced expression of MMP-1, 3, 9 and type I procollagen in human fibroblast. ………………………………………………………88 Fig. 13 Effect of Michelia alba extract on the UVB-induced expression of MMP-1, 3, 9 and type I procollagen in human fibroblast. ……89 Fig. 14 Effect of Michelia alba extract on the UVB-induced expression of MAP kinases in human fibroblast. ……………………………90 Fig. 15 The primary skin irritation test of Coffea arabica extract in the rabbit. …………………………………………………………91 IV.
(7) Fig. 16 The primary skin irritation test of Michelia alba extract in the rabbit. …………………………………………………………92 Fig. 17 The Draize eye test of Coffea arabica extract in the rabbit. …93 Fig. 18 The Draize eye test of Michelia alba extract in the rabbit. ……93. V.
(8) 附 表 目 錄 Table 1 The inhibition rate of Coffea arabica and Michelia alba extract on bacterial collagenase by gelatin digestion assay….94 Table 2 The inhibition rate of Coffea arabica, Michelia alba and Morinda citrifolia extract and their hydrolysates on bacterial collagenase by gelatin digestion assay ……………………………………94 Table 3 The inhibition rate of Coffea arabica extract and its hydrolysates on bacterial collagenase using fluorometric assay…………….94 Table 4 The inhibition rate of Michelia alba extract on bacterial collagenase using fluorometric assay………………….95 Table 5 The inhibition rate of Michelia alba extract and its hydrolysates on porcine elastase……………………………………………..95 Table 6 The scores of primary skin irritation test of Coffea arabica extract at low dose (0.1 g) in the rabbit………………………………..95 Table 7 The scores of primary skin irritation test of Coffea arabica extract at high dose (0.5 g) in the rabbit………………………………96 Table 8 The scores of primary skin irritation test of Michelia alba extract at low dose (0.1 g) in the rabbit.………………………………96 Table 9 The scores of primary skin irritation test of Michelia alba extract at high dose (0.5 g) in the rabbit.………………………………96 Table 10 The scores of Draize eye test of Coffea arabica.……………97 Table 11 The scores of Draize eye test of Michelia alba………………97. VI.
(9) 中文摘要 多酚類具有許多生物活性如抗氧化,抑制 MMPs 與彈力蛋白酶活 性,若能抑制彈力蛋白酶或基質金屬蛋白酶之分泌或活性,則可延緩 皺紋生成。本研究針對茜草科植物咖啡(Coffea arabica, CA)、檄樹 (Morinda citrifolia, MC)和木蘭科植物玉蘭花(Michelia alba, MA)的葉 萃取物及其水解物、咖啡成分綠原酸和咖啡酸等探討其抑制 MMP 活 性。上述三種植物以甲醇製備其萃取物,再以鹽酸水解製備水解物。 三種植物之萃取物及其水解物先以膠原蛋白酶與彈力蛋白酶抑制實 驗進行初篩。具活性者再以紫外線 B 照射纖維母細胞誘導 MMPs 產 生之模式,探討對第 I 型膠原蛋白原的生成及 MMPs 之作用,進而再 探討對 MAP Kinases 磷酸化的影響。並針對具活性者進行安全性評 估。 CA、MA 及 MC 之甲醇萃取率分別為 10.4%、16.9%、15.0%。於 明膠分解初篩試驗結果顯示:咖啡萃取物(CAE,相當於原植物 9.6 mg/mL)與玉蘭花萃取物(MAE,相當於原植物 5.9 mg/mL)及兩者的水 解物(CAH, MAH 1000 μg/mL)抑制率皆大於 90%。檄樹萃取物(MCE 相當於原植物 6.7 mg/mL)抑制率 74.3%,水解物抑制率皆低於萃取 物。而咖啡酸與綠原酸均無此抑制效果。 膠原蛋白酶活性抑制試驗結果顯示:CAE 具抑制效果,且具有濃. VII.
(10) 度依存性,序列稀釋萃取物,相當於原植物濃度 0.096~4.8 mg/mL 之 抑制率 84.6~100.3%。CAH 之抑制效果未優於 CAE。MAE 具抑制效 果,且具有濃度依存性,序列稀釋萃取物,相當於原植物濃度 0.059~ 2.96 mg/mL 之抑制率 92.7~100.1%。MAH 之抑制效果未優於 MAE。 彈力蛋白酶活性抑制試驗結果顯示:CAE 及 CAH 皆無抑制效果。而 MAE 與 MAH,抑制效果亦不佳。 對於 UVB 所誘導 MMP-1, -3, -9 之抑制表現試驗結果顯示,CAE 能抑制 MMP-1, -3, -9 之濃度分別為相當於原植物 0.048, 0.96, 0.48 mg/mL。MAE 能抑制 MMP-1, -3 之濃度分別為相當於原植物 0.029 及 0.059 mg/mL。咖啡酸能抑制 MMP-1, -3 的表現之濃度分別為 5 及 50 μM。綠原酸能抑制 MMP-1, -3 表現之濃度分別為 10 及 50 μM。 CAE 能提高第 I 型膠原蛋白原生成的濃度為相當於原植物 0.096 mg/mL。MAE 無提高生成的效果。 CAE 可抑制 JNK、ERK 及 p38 磷酸化之濃度分別為相當於原植 物 0.048, 0.24 及 0.048 mg/mL。MAE 可抑制 JNK 及 ERK 磷酸化之濃 度分別為相當於原植物 0.029 及 0.148 mg/mL。CAE 與 MAE 於高濃 度(相當於原植物 4.8 及 2.9 g/mL)無皮膚刺激性。兩者皆無眼球刺激 性。此結果顯示咖啡及玉蘭花具有開發成為抗光老化化妝品之潛力。. VIII.
(11) The effect of Coffea arabica, Morinda citrifolia and Michelia alba on MMPs and elastase. Abstract Polyphenols are a group of compounds possessing a variety of biological activities including anti-oxidant, inhibition of MMP-1 and elastase. The inhibition of elastase and MMPs in the skin would slow down the process of aging. In this study, Coffea arabica (CA), Morinda citrifolia (MC) and Michelia alba (MA) extracts and their hydrolysates, and active constituents of CA, chlororgenic acid and caffeic acid, had been selected to screen for MMPs inhibition. The plants were extracted with methanol and the extracts were hydrolyzed with different concentration of hydrochloric acid. The chlororgenic acid, caffeic acid, the various concentration of extracts and its hydrolysates were subject the inhibition tests to MMP-1 and elastase. The fibroblasts were used for the collagen synthesis promotion, MMPs inhibition and MAP Kinases phosphorylation inhibition tests on plants extracts. The active ones were process to safety assessment. The methanolic extraction yield of CA, MA and MC were 10.4%, 16.9% and 15%, respectively. In gelatin digestion test, the inhibition effects of CAE (equal to 9.6 mg dried leaves/mL), MAE (equal to 9 mg dried leaves/mL) and their hydrolysates (1000 μg/mL CAH and MAH) were more than 90%, in MCE (equal to 6.7 mg dried leaves/mL) was 74.3%. However, cholorogenic acid and caffeic acid show no effect on gelatin digestion. The results indicated that CAE would inhibit collagenase activity IX.
(12) and present dose dependent effects. The inhibition results were in range of 84.6%~100.3% at the concentrations of 0.096~4.8 mg/mL (dried leaves), respectively. The collagenase inhibition of CAE was superior to CAH. MAE would inhibit collagenase activity and present dose dependent effects. The inhibition results were in range of 92.7%~100.1% at the concentrations of 0.059~2.96 mg/mL (dried leaves), respectively. The effects of MAE on collagenase inhibition were better than MAH, moreover, CAE, CAH, MAE and MAH had no significantly inhibition results on elastase activity. CAE would inhibit MMP-1, -3 and -9 at concentration of 0.048, 096 and 0.48 mg/mL (dried leaves) and MAE would inhibit MMP-1 and -3 at 0.029 and 0.059 mg/mL (dried leaves). Caffeic acid would suppress MMP-1 and -3 expressions at 5 and 50 μM, moreover, that of chlorogenic acid were 10 and 50 μM. In addition, CAE would elevate type I collagen synthesis at 0.096 mg/mL (dried leaves), but not MAE. In MAP Kinases inhibition, the results indicated that CAE would inhibit the phosphorylation of JNK, ERK and p38 at the concentrations of 0.048, 0.24 and 0.048 mg/mL (dried leaves) and MAE would inhibit JNK and ERK phosphorylation at 0.029 and 0.148 mg/mL. In safety assessment, CAE and MAE did not present toxicity at higher concentrations (equal to 4.8 and 2.9 mg dried leaves/mL) on the skin and eye irritation test. The results show that Coffea arabica and Michelia alba could become the promising anti-photoaging material of cosmetics through this investigation.. X.
(13) . 第一章. 緒言. 皮膚是保護身體的第一道防線,外在環境因素如紫外線,對於皮 膚老化有促進的作用。隨著年紀的增長,皮膚皺紋、鬆弛及膚色黯沉 等老化的徵象一一出現,越來越多人嘗試逆轉歲月留在皮膚上的痕跡。 老化原因分為內因性與外因性,內因性是指身體自然的老化過 程,而外因性老化則是因為外在環境因素所造成的老化。對於皮膚而 言,造成損害最嚴重、加速皮膚老化的最主要因素就是紫外線暴露。 紫外線所造成的老化現象稱之為光老化,光老化會造成真皮層之基質 金屬蛋白酶量及其活性增加。基質金屬蛋白酶會分解細胞外基質,例 如膠原蛋白,皮膚少了膠原蛋白的支撐,外觀會變得鬆弛、彈性降低, 而且角質層過度增生,皮膚變得黯沉(Oba and Edwards, 2006; Pillai et al., 2005; Uitto, 2008)。 因為美容醫學與化妝品工業的發展,人們對於皮膚的生理構造、 恆定性與老化的過程機轉有了更深入的了解。現有市售的抗皺產品有 效成分多為維他命 A 酸及其衍生物(Stefanaki et al., 2005)、果酸、維他 命 C、維他命 E 及其衍生物或具抗氧化作用的化合物(Burke, 2007; Colven and Pinnell, 1996; Glaser, 2004; Luo et al., 2007; Torras, 1996)。A 酸與果酸有刺激性,維他命 C 則不穩定,因此希望能在天然物中尋求. 1 .
(14) . 經濟,有效、安定性更佳、安全的成分。有許多研究顯示,多酚類成 分如綠茶及其主成分 EGCG (Bae et al., 2008; Katiyar, 2003; Yusuf et al., 2007)、石榴萃取物(Syed et al., 2006; Zaid et al., 2007)、白藜蘆醇(Baxter, 2008)等皆具有抗老化的效果。 茜草科植物如檄樹(Morinda citrifolia)葉含 flavonol glycosides(Sang et al., 2001a),阿拉比卡咖啡(Coffea arabica)含 chlororgenic acid、caffeic acid 及 catechin 等多酚類成分(Ramirez-Coronel et al., 2004; Zhao et al., 2008),因而具抗老化的潛力。另外木蘭科植物和厚朴亦有研究指出具 有抗光老化活性(Tanaka et al., 2007),屬於此科植物之玉蘭花亦值得研 究探討。 曾有文獻報導苷元之生理活性較配醣體佳(Kim et al., 2004a)而苷元 較配醣體更具脂溶性易吸收,因此本研究針對咖啡、檄樹、玉蘭花等 植物的葉子萃取物及其水解物對基質金屬蛋白酶之影響進行探討。先 以細菌膠原蛋白酶進行明膠分解試驗篩選,確定具抑制作用後,再進 一步探討對於纖維母細胞之基質金屬蛋白酶及膠原蛋白生成影響及調 控機制,開發具對抗光老化作用之成分。. 2 .
(15) . 第二章 總論 第一節 皮膚生理結構 皮膚包覆身體與外在環境相界,可以抵禦外來各種刺激、傷害, 達到保護人體的效果。其面積約 1.6 平方公尺,是身體很重要的器官。 皮膚的厚度隨著年齡、性別及部位而有所變化,由外而內可以分成表 皮(epidermis)、真皮(dermis)和皮下組織(subcutis)三層(如下圖)。茲將 皮膚各層之構造及生理功能列述於下。. 圖一 Scheme of different layers of the skin. (Scharffetter-Kochanek et al., 2000). 3 .
(16) . (一)表皮層 表皮位於身體最表淺處,由細胞相互緊密堆積而成。於身體各部 位的厚薄度不一,以手掌及腳掌最厚,眼皮最薄,且個體間不盡相同。 表皮層可由內而外再細分為四層:基底層(basal layer)、有棘層(spinous layer)、顆粒層(granular layer)、角質層(stratum corneum);手掌及腳掌 在顆粒層與角質層間還有一層透明層(lucid layer),表皮細胞大多是角 質細胞(keratinocytes),角質細胞堆積最後會形成角質層。基底層細胞 與表皮新陳代謝有關,基底層的細胞會持續地分裂,並且慢慢地向表 皮上層移動,而漸漸形成棘狀細胞,形成有棘層。有棘層之細胞間有 狹小間隙隔開,淋巴液可於其間隙間流動,促使營養物質擴散。有棘 層是表皮中最厚的,其成熟後,形成顆粒層,顆粒層細胞呈扁平狀, 平鋪排列二至三層,其細胞內有 keratohyalin 顆粒存在。表皮最外層 為角質層,角質層細胞有別於其他細胞,細胞核及分散於細胞質中之 胞器皆消失,多數只有角質素(keratin) (光井武夫, 2004; 張新湘, 1977)。 (二)真皮層 表皮層與真皮層間靠著基底膜(basement membrane)緊緊地連接 著,基底膜的厚度只有 0.1 μm。基底膜可使表皮細胞正常增生,新陳 代謝。並調控真皮及表皮細胞所產生之生理活性物質與機能。真皮平 4 .
(17) . 均厚度為 1~ 4 mm,約佔皮膚的 90 %。此層細胞較表皮少,大多是 細胞外基質,並以纖維為主,包括膠原蛋白(collagen)及彈力纖維 (elastic fibers)。膠原蛋白以 I 與 III 為主,主要功能為維持組織形狀, 而彈力纖維則分佈於膠原蛋白中,使皮膚具有彈性。真皮層大致可分 為兩層:乳頭狀真皮層(papillary dermis),位於真皮層與表皮交界面, 較薄不如表皮平坦,具有凹凸狀似乳頭狀的結構,嵌入表皮中,與表 面緊密銜接。此層的纖維較細,且稀疏、多成垂直走向;網狀真皮層 (reticular dermis),位於乳頭層下方,其纖維較密且粗,呈水平走向。 有許多毛細血管及神經的末梢存在乳頭層中。真皮中尚有多醣類存 在,以葡萄胺聚醣(glucosaminoglycans)為主,主成分為醣醛酸 (hyaluronic acid)與硫酸皮膚素(dermatan sulfate)。這些成分會與蛋白質 結合形成蛋白聚醣(proteoglycan),可保持大量的水分,呈膠體狀,存 在於纖維間。而膠體內之水分可使血管中的養分、代謝產物及荷爾蒙 擴散到組織之細胞內,並使組織柔軟富有彈性,因此細胞外基質可提 高皮膚之柔軟性與張力(光井武夫, 2004; 張新湘, 1977)。 (三)皮下組織 皮膚最內層稱之為皮下組織,主要由脂肪組織組成,並藉纖維膠 原與真皮層區隔,含有神經、血管、淋巴、毛囊及汗腺等。提供保暖、 儲存能量和吸收震動之功能(光井武夫, 2004; 張新湘, 1977)。 5 .
(18) . 第二節 膠原蛋白 (一)膠原蛋白生理功能 膠原蛋白(collagen)是細胞外基質含量最豐富的蛋白質,它是一群 具有相似特性及結構蛋白質的總稱,目前共發現 21 種膠原蛋白。膠 原蛋白是構成結締組織的主要成分,也可以在組織間隙發現其蹤跡, 其能穩定組織、器官架構,並維持結構之完整性。 (二)膠原蛋白結構 膠原蛋白是富含脯胺酸(proline)的三胜肽,由 Gly-X-Y 的重複片 斷所構成的三條螺旋狀聚胜肽鏈纏繞形成的螺旋狀纖維蛋白,三條 α-chains 以左旋方向纏繞,每 18 個胺基酸形成一個圈,以一個基團與 其他分子連結形成圍繞中心的的右旋三股螺旋。形成此結構乃因膠原 蛋白主要的胺基酸為甘胺酸,是最小的胺基酸,形成之立體障礙最 小,可使中心緊密有序排列。而 Gly-X-Y 的其他基團則位於螺旋外 部,X、Y 大多為脯胺酸或羥脯胺酸。依膠原蛋白的類型不同,胺基 酸會由酶做轉譯後修飾,特別是脯胺酸與離胺酸(lysine)的基團,接 上羥基,4-羥基脯胺酸而能形成分子內氫鍵,穩定三股螺旋結構,三 股螺旋之長度視膠原蛋白類型不同長短及功能而不相同。. 6 .
(19) . 圖二 Molecular structure of type I collagen with the various subdomains as well as the cleavage sites for N- and C-procollegenases. (Gelse et al., 2003) (三)第 I 型膠原蛋白 (Gly-X-Y)n 控制第 I 型膠原蛋白 domain 的長度(300 nm)由 1000 個胺基酸所組成,如圖二。第 I 型膠原蛋白佔骨骼之 90 %,也是肌 腱、皮膚、韌帶、角膜和組織間隙主要的膠原蛋白類型。第 I 型膠原 蛋白是由三條 α-chains 所組成,兩條 α 1(I) chain,一條 α 2 (I) chain, 其基因分別為 COL1A1 與 COL1A2,位於第 17 號染色體(17q21.31) 及第 7 號染色體(7q22.1)。主要分佈於皮膚真皮層,支撐皮膚的結構, 因而膠原蛋白於維持皮膚外觀扮演重要角色,少了膠原蛋白的支撐, 真皮間隙產生空洞,皮膚會變得鬆垮、無彈性,皺紋隨之出現,老化 的徵象一一出現。膠原蛋白從轉錄、轉譯、轉譯後修飾及分泌至細胞 外的過程:膠原蛋白的基因序列經剪接體(spliceosome)將序列中內含 子(intron)剪切後,外顯子(exon)黏合形成 mRNA,並將 mRNA 輸出 至細胞質的粗糙內質網與高基氏體進行轉譯及轉譯後修飾。經核醣體 7 .
(20) . 轉譯形成胜肽後,並接上訊號胜肽酶(signal peptidase),進行一連串的 後 修 飾 作 用 。 藉. hydroxylysyl. galactosyltransferase. 及. galactosylhydroxylysyl glucosyltransferase 將半乳糖與葡萄糖接於胜 肽 上 , lysyl hydroxylase 於 離 胺 酸 (lysine) 接 上 - OH 基 , prolyl hydroxylase 於脯胺酸(proline)接上- OH 基。纏繞形成膠原纖維後, oligosaccharyl transferase complex 於 C 端接上寡糖,protein disulfide isomerase 形成雙硫鍵,peptidyl-prolyl cis-trans-isomerase 改變蛋白質 結構,以 heat shock protein 47 幫助褶疊,傳送分泌至細胞外。再分別 以膠原蛋白原 N-proteinase、膠原蛋白原 C-proteinase 切除 N、C 端 之胜肽。經 lysyl oxidase 於纖維接上- OH 基,使膠原纖維共價連結形 成膠原蛋白(Gelse et al., 2003) 。. 8 .
(21) . 圖三 Schematic representation of collagen synthesis starting from the nuclear transcription of the collagen genes, mRNA processing, ribosomal protein synthesis (translation) and post-translational modifications, secretion and the final steps of fibril formation. (SP: signal peptidase; GT: hydroxylysyl galactosyltransferase and galactosylhydroxylysyl glucosyltransferase; LH: lysyl hydroxylase; PH: prolyl hydroxylase; OTC: oligosaccharyl transferase complex; PDI: protein disulphide isomerase; PPI: peptidyl-prolyl cis-trans-isomerase; NP: procollagen N-proteinase; CP: procollagen C-proteinase; LO: lysyl oxidase; HSP47: heat shock protein 47, colligin1). (Gelse et al., 2003) 9 .
(22) . 第三節 基質金屬蛋白酶(MMPs)(Wong et al., 2002) (一) 基質金屬蛋白酶生理功能 基質金屬蛋白酶(Matrix metalloproteinases;MMPs)可分解細胞外 基質及基底膜蛋白質,這些酶與胚胎發育、生長、傷口癒合的過程皆 息息相關。若MMPs活性長期失調會造成疾病的產生,不正常的細胞 外蛋白質水解,例如:關節炎(arthritis)、腫瘤形成(tumorgenesis)、 組織潰瘍(tissue ulceration)及動脈粥狀硬化(atherosclerosis)。 基質金屬蛋白酶於正常組織分泌量很低,發炎反應將促使其蛋白 表現量增加及活性提高,此亦因細胞激素、賀爾蒙、生長因子與致癌 基因的刺激而發生。MMPs活性在身體裡高度抑制避免失去控制,而 過度破壞細胞外基質。而調控的機制可於轉譯的層次或者是轉譯後的 修飾,抑制酶的活化與以TIMPs抑制活化的MMPs。 (二) 基質金屬蛋白酶分類 基質金屬蛋白酶至目前了解約近三十個成員,如表一,可以依據 結 構 的 特 色 分 為 五 大 類 : 膠 原 蛋 白 酶 (collagenases) 、 明 膠 酶 (gelatinases) 、 基 質 溶 素 (stromelysins) 、 膜 型 基 質 金 屬 白 酶 (membrane-type MMPs;MT-MMPs)、matrilysins以及其他。主要可以 歸納出幾項特徵: 1. MMPs以zymogen(latent form)分泌。 10 .
(23) . 2. MMPs皆能分解細胞外基質。 3. 酶的催化機轉為活化位的Zn2+/Cu2+ 4. MMPs的活性可以被tissue inhibitiors of metalloproteinases(TIMPs) 所抑制。 5. cDNA的序列皆與膠原蛋白酶相似。. 膠原蛋白酶專司分解結締組織中的膠原蛋白,主要是已被鬆開的 螺旋膠原纖維及第I、II和III型的膠原蛋白。形成變性的明膠,使其易 受其他金屬蛋白酶和蛋白水解酶的分解。明膠酶亦稱為第IV型的膠原 蛋白酶,可分解基底膜及變性的膠原蛋白。而Stromelysins的受質較 多樣,能夠分解細胞外基質,例如fibronectin,proteoglycans,laminin 和基底膜的第IV型膠原蛋白。 MT-MMPs的結構不同於其他的基質金屬蛋白酶,其具有穿膜的 domain,靠細胞質一端有25個胺基酸所形成的錨,使酶能固定於細胞 外膜上。膜型MMPs有一個獨特的辨識區域,即是位於propeptide區之 furin(與高基氏體有關的蛋白水解酶),此與其細胞內活性有關。. 11 .
(24) . 表一 The matrix metalloproteinase (MMP) family (Raffetto and Khalil, 2008). 12 .
(25) . 第四節 光老化 皮膚的老化主要可分為兩個原因:內因性老化(intrinsic aging)與 外因性老化(extrinsic aging),而外因性老化受外在環境,如紫外線、 汙染物、自由基等影響,最主要為紫外線所引起的光老化 (photoaging)。光老化是指慢性光損害的皮膚,長期暴露於陽光下而 造成皮膚光化。內因性老化皮膚的特色為平滑、乾燥、暗沉並伴隨著 細紋的產生。而光老化的皮膚則是有嚴重的皺紋和色素沉澱,例如: 曬斑和臉部、頸部與手臂斑點狀的色素沉著(Svobodova et al., 2006) 。 轉譯因子NF-κB會因為UV照射而活化,NF-κB會刺激發炎反應的 細胞激素產生,放大皮膚對於UV的反應,吸引已形成嗜中性白血球 膠原蛋白酶(MMP-8)的嗜中性白血球。NF-κB也能促進MMP-9的表 現。只要很小的UV劑量,低於造成紅斑的劑量,就可使MMP的表現 增加。隨著UV照射劑量的增加,MMPs的表現量也會隨之增加。 經人體試驗證實,光老化的皮膚膠原蛋白產生量會減少,照射UV 後膠原蛋白原產生量降低,於二十四小時後最為明顯(Chung, 2003; Fisher et al., 2000; Rabe et al., 2006; Rittie and Fisher, 2002) 。. (一) UV對於DNA所造成的傷害 皮膚經UV照射會產生直接或是間接的影響,大約有50%UV所造 成的傷害乃因形成自由基,自由基會直接造成細胞的傷害。而光致癌 13 .
(26) . 性(photocarcinogenesis)及UV照射所引起的DNA分子改變,已被廣泛 研究。皮膚組織中黑色素吸收能量達到激發態(excited states),產生化 學變化,將能量轉移至其他分子,或以光或熱的方式散發多餘的能 量。DNA吸收UV的波長範圍為245 nm至290 nm,因此UVB為誘導 DNA突變的最主要原因。UVB對DNA所造成的突變除了化學變化之 外,尚有形成環丁烷嘧啶二聚體(cyclobutane pyrimidine dimmers)與 Pyrimidine-pyrimidone (6-4) photoproducts(如圖四),會使DNA的螺旋 結構產生扭曲變形 (Ichihashi et al., 2003; Moriwaki and Takahashi, 2008)。 皮膚經UVA照射間接產生活性氧,包括超氧陰離子(superoxide anion)、過氧化物(peroxide)和單態氧(singlet oxygen)。活性氧不僅會 損害DNA也會傷害組織裡之脂質與蛋白質,UVA能使trans-urocanic acid產生變化,使單態氧產生與DNA nicks。8-Hydroxy guanine 也是 UVA藉由活性氧誘導產生。. 14 .
(27) . . 圖四 Adjacent pyrimidine bases on the same stand pyrimidine dimer (a) or (6-4) pyrimidine-pyrrimidone photoproduct (b) alter absorbing UVB light energy.(Ichihashi et al., 2003). (二) UV誘導的訊號傳遞 UV照射後產生活性氧(Reactive oxygen species,ROS),會引發下 游一連串的訊號傳遞途徑產生。暴露UV後15分鐘,角質細胞、纖維 母細胞的表皮生長因子、IL-1、TNF-α的受器會活化,而影響下游的 訊號傳遞。促使核轉譯因子AP-1(activator protein 1)被活化,進而調控 MMPs的表現。 活化細胞表面細胞激素與生長因子受器將產生調節下游訊號傳 15 .
(28) . 遞的蛋白質,匯集這些訊號傳遞將會活化small GTP-binding protein例 如: Ras、Rac與Cdc42,這三個small GTP-binding protein是ERK、JNK 與p38等mitogen activated protein kinase (MAP Kinase)的上游調控因 子。在人體皮膚中經UV照射後的30分鐘內,Ras就會被活化。與GTP 結合後的Ras會吸引Raf-1 kinase到細胞膜,Raf-1因而被活化,而Raf-1 是 ERk MAP Kinase 的 上 游 活 化 因 子 。 另 一 個 small GTP-binding protein,Rac-1,是由生長因子、細胞激素所活化,同樣地,在人體 皮膚裡照射UV後能夠快速的被活化。皮膚經UV照射15分鐘後會誘導 NADPH oxidase活性,因而提高超氧陰離子的活性。活性氧產生增加 時可能擴大訊號傳遞而活化MAP Kinases。Rac-1和Cdc42已被證實能 與調節MEKK(JNK、ERK和p38上游活化因子)的序列結合如圖四 (Jinlian et al., 2007; Rittie and Fisher, 2002)。 UV照射後細胞內神經醯胺(ceramide)含量增加,能活化MAPK pathway。神經醯胺是由神經鞘磷脂酵素(sphingomyelinases, SMases) 分解鞘磷脂(sphingomelin)所產生,能快速活化Raf-1、MEKK1和Erk。 由於神經醯胺與活性氧的量同時提高,推測UV誘導神經醯胺的產生 與活性氧產生的提高有相關性。而且UV所誘導神經醯胺的產生會被 自由基清除劑如維生素E所抑制。. 16 .
(29) . 圖五 UV-induced signaling cascades: UV activation of growth factor or cytokine receptors leads to the activation of MAP Kinase.(Rittie and Fisher, 2002) 17 .
(30) . 1. UV誘導AP-1活化 MAP kinase pathways最主要的效應產生於轉錄因子AP-1。AP-1是 由Jun和Fos家族蛋白(c-Jun,JunB,JunD,c-Fos,FosB,Fra1和Fra2) 所組成(Karin et al., 1997),以異雙元體(heterodimer)最為典型,由c-Jun 與c-Fos所組成,縱使組合的變化依據啟動子前後的序列而定,但 c-Jun/c-Fos具有最高的轉錄活性。與培養的細胞相比,人類皮膚會持 續高度表現c-Fos,且不受UV的影響。當皮膚未照射UV時,AP-1轉錄 因子通常由c-Fos和JunD所組成。在許多生長因子、細胞激素以及外 在環境刺激下(例如:UV)能誘導c-Fos和c-Jun基因的轉錄。MAPK pathways可以直接或間接地促進c-Fos和c-Jun基因的轉錄,除此之 外,JNK及p38可透過磷酸化的方式活化與穩定c-Jun。人體皮膚中, UV所誘導c-Jun的mRNA及蛋白質量的增加可在30分鐘及1小時分別 被偵測出,c-Jun蛋白質的量可以持續維持24小時,其能有效與JunD競 爭形成具有活性的AP-1(c-Jun:c-Fos)並遍及表皮與真皮細胞。. 2. AP-1調控MMPs與膠原蛋白生成 AP-1可以調控許多基因的表現,而這些基因可進一步調節細胞生 長與分化。有些MMPs家族成員也受到AP-1的調控,MMPs是依賴鋅 (zinc-dependent)的蛋白內切酶,可以有效降解所有的細胞外基質蛋 白,以不活化態(zymogens)分泌。經過酵素切斷前結構域(prodomain) 18 .
(31) . 方能成為活化態,而MMPs的活化可藉由三方面來調控:MMPs的合 成、zymogens的活化和抑制水解活性。MMPs活性可藉由內生性的蛋 白抑制劑TIMPs所抑制。而這些MMPs中可被AP-1所活化的包含: MMP-1、MMP-9和MMP-3。在人體試驗中,皮膚經UV照射會誘發 MMP-1、MMP-3和MMP-9基因的表現(Fisher et al., 1996; Fisher et al., 1997)。UV照射後的8小時可以觀察到MMP-1、MMP-3和MMP-9基因 的轉錄增加,這三個MMPs的基因表達增加進一步合成許多具有酵素 活性的MMPs(Fisher et al., 1998)。UV造成皮膚的傷害會誘發部分 MMPs的表現,而MMP-1、MMP-3和MMP-9具降解膠原蛋白纖維的 能力。人體試驗中觀察得知,UV照射後24小時內,可以發現膠原蛋 白的降解量增加(Fisher et al., 1997)。因此,皮膚暴露於UV下,將導 致AP-1轉錄因子增加,而提高MMPs的濃度,造成細胞外基質的降解。 UV不僅增加膠原蛋白降解速度,也會減少第I型膠原蛋白的生 成,形成膠原纖維的量也會降低(Fisher et al., 2000)。活性氧能直接攻 擊組織使其氧化,使膠原蛋白降解(Dong et al., 2008),干擾膠原蛋白 基因表現的訊號傳遞。在體外實驗(Bizot-Foulon et al., 1995; Fisher et al., 1997)與人體試驗(Bizot-Foulon et al., 1995; Fisher et al., 1996)中發 現,纖維母細胞照射UV後,細胞外基質的降解會增加。第I型的膠原 蛋 白 的 減 少 部 分 是 由 於 UV 所 誘 發 之 AP-1 造 成 的 , AP-1 可 降 低 19 .
(32) . COL1A1 and COL1A2 的 基 因 表 現 (Bornstein, 1996; Chung et al., 1996)。UV照射後的8個小時內,第I型膠原蛋白的表現會降低。在接 下來的24小時內,可以觀察到上層的真皮中第I型膠原蛋白原減少 (Chung, 2003; Fisher et al., 2000; Rabe et al., 2006)。 3. UV削弱TGF-β訊號傳遞 TGF-β能調節多種細胞功能,包括分化、增生與誘發細胞外基質 蛋白的生成,在皮膚TGF-β會抑制表皮角質細胞生長和促進真皮纖維 母細胞生長,並誘發產生細胞外基質膠原蛋白和彈力蛋白,抑制 MMP-1和MMP-3。TGF-β的受器遍及表皮與真皮。TGF-β是膠原蛋白 合成過程中很重要的啟動子(promoter),其中最占優勢為TGF-β2群 組。TGF-β的作用是經由與細胞表面受器複合體結合,複合體含3種 蛋白:Type I、II、III (TβRI、TβRII、TβRIII),TGF-β的接合是與兩 個TβRI及兩個TβRII的複合體結合,TβRII磷酸化TβRI,TβRI又磷酸 化receptor-activated Smad protein(R-Smad)-Smad2和Smad3形成雜聚 體並與Smad4(co-Smad)結合形成聚合體,此聚合體進入細胞核後 調節標的基因的轉錄。Inhibitory Smads(I-Smad),Smad6和Smad7 能夠阻斷R-Smad磷酸化,因此影響TGF-β的訊號傳遞(Gambichler et al., 2007)。經UV照射後TGF-β2的蛋白表現量降低和增加Smad7的表 現,造成第I型膠原蛋白減少,能夠藉由調控AP-1與TGF-β來調節UV 20 .
(33) . 造成的膠原蛋白合成降低。 CCN family是 cystein-rich proteins,其中的兩類connective tissue growth factor (CTGF) 和Cyr61,可以調控TGF-β所誘發的膠原蛋白原 產生。TGF-β藉由Smad這條路徑來刺激CTGF基因表達。以CTGF注射 老鼠皮膚或加至培養的纖維母細胞中,可以刺激第I型膠原蛋白原的 產生。也有文獻指出,利用anti-CTGF的抗體及RNAi的方式可以抑制 由 TGF-β 所 誘 導 的 第 I 型 膠 原 蛋 白 原 的 產 生 (Duncan et al., 1999; Kothapalli et al., 1997)。相反地,在培養的纖維母細胞中加入Cyr61則 可抑制膠原蛋白的產生,當外加TGF-β時可以克服此現象。CTGF和 Cyr61於人體皮膚的表皮及真皮中或是培養的角質細胞、纖維母細胞 中皆能持續表現。 UV 射 線 會 藉 由 減 少 TβRII 的 表 現 及 增 加 Smad7 的 表 現 來 削 弱 TGF-β 的 訊 號 傳 遞 , 其 中 TβRII 的 減 少 所 產 生 的 影 響 較 大 , 如 此 TGF-β/Smad的訊號傳遞在一開始的步驟就被阻斷。在動物實驗與細 胞實驗中,經UV照射會減低CTGF(Quan et al., 2002),此作用也可能 是UV所造成第I型膠原蛋白原生成減少的原因,但UV抑制CTGF基因 表現的機制目前尚未明瞭。 (三) 內因性老化 1. 內因性老化皮膚膠原蛋白生成降低 21 .
(34) . 內因性最主要的特徵就是第I型、第III型膠原蛋白原生成量降低, 使得皮膚變薄而脆弱,其不論是蛋白質或是mRNA的量皆降低。在體 外實驗中,老化的纖維母細胞TGF-β1、TβRI、TβRII基因表現降低, 而在老化的皮膚中TGF-β1和TβRII的量減少。TβRII的減少或許也是重 要的一環,因為如此會減低TGF-β所誘發的訊號傳遞,但TβRII的減 少不會影響一般蛋白質的合成,因為在老化皮膚中TβRI並未減少。除 此之外,CTGF也是減少的,這樣的現象與TGF-β誘導的反應一致, 皆會降低第I型膠原蛋白原的生成。 Decorin是富含白胺酸(leucine)的蛋白聚醣(proteoglycan)。在老化 的皮膚中TGF-β會與decorin結合且被隔離。Decorin與膠原蛋白間的作 用會影響到皮膚的張力強度,並調控膠原蛋白纖維形成的速率。 decorin基因缺陷的老鼠,皮膚因無張力而易損傷,大量膠原蛋白纖維 呈現不規則狀。與年輕老鼠相較,年長老鼠皮膚真皮中decorin的量會 上升,在體外實驗中,老化的人類纖維母細胞也有相同的情形。推測 老化的人類皮膚中可觀察到過量decorin,造成膠原蛋白纖維結構破 壞。然而decorin的量在老化與UV照射的皮膚中是否升高,仍是未知 的。 2. 內因性老化皮膚基質金屬蛋白酶表現增加 內因性的皮膚除了膠原蛋白生成減少外,MMP-1、MMP-2 22 .
(35) . (gelatinase A)、MMP-3和MMP-9的產生增加。除了MMP-2以外其餘 MMPs皆由AP-1所調節,也能由UV所誘發,不論在人體試驗或是體 外實驗中AP-1於老化的皮膚中的表現量皆是上升。c-Jun mRNA的表 現量與年輕的皮膚比較,老化的皮膚中增加了兩倍,c-Fos的量則是 不變。因此可知MMP-1、MMP-3和MMP-9的表現增加是由於AP-1的 緣故,MMP-2增加的機轉則不明。 3. AP-1表現增加 AP-1是MAP kinase訊息傳遞路徑中重要的效應因子。在老化皮膚 中,AP-1表現增加是由於JNK MAP kinase活性增加,JNK活性提高約 3.4倍,是因酵素活性增加而非蛋白量增加(Chung et al., 2000)。JNK 將c-Jun磷酸化,刺激下游基因作用及增加其半衰期。因此AP-1所調 節的MMPs表現增加及c-Jun蛋白量增加可能都歸因於JNK活性增加 (Chung et al., 2000)。與光老化皮膚不同的是Erk於自然老化皮膚的活 性是減少的,而蛋白質量未改變,推測其原因是上游訊號傳遞減弱。 Erk是由生長因子受器所活化,訊號降低可能是反應細胞增生的減 少。儘管EGF-G的表現不因老化而減少,但EGF-R-dependent的訊息 傳遞將會減少(Reenstra et al., 1996)。 4. 氧化壓力提高 氧化壓力被認為是老化過程中最主要的因素,老化的自由基理論 23 .
(36) . 在幾十年前被提出,老化起源於終生的累積有氧代謝產生的自由基, 細胞暴露於過量的活性氧時,造成氧化損傷(Hensley and Floyd, 2002)。皮膚雖擁有高效率的抗氧化系統,隨著年紀的增長,活性氧 量提高而皮膚抗氧化能力下降(Hu et al., 2000; Kohen and Gati, 2000)。活性氧是MAP kinase傳遞路徑中必要的參與者,當MAPK kinase活化將誘導MMP表現的上游調控者AP-1表現,在老化皮膚中膠 原蛋白降解增加可能是很重要的機轉(Rittie and Fisher, 2002) 。. 第五節 玉蘭花、檄樹與咖啡之文獻考察 (一) 玉蘭花 1. 概述 學名:Michelia alba DC. 白玉蘭為木蘭科之常綠喬木。樹皮灰 色,嫩枝被白色毛。葉互生,柄長 1.5~2 cm,葉片大型,革質,卵 狀橢圓形或長橢圓形,長 10~25 cm,寬 4~8 cm,先端長漸尖或尾 狀漸尖,基部稍鈍狀尖形,全緣或微波緣,兩面光滑無毛或背被疏毛。 花單生葉腋,白色,極芳香,長 3~4 cm。萼片長圓形,花瓣披針形, 長 3~3.5 cm;雄蕊多數,多輪,花絲扁平。雌蕊心皮多數,每心皮 具胚珠多於 2 枚,螺旋狀排列於花托柄上,子房被細毛,柱頭頭狀。 果實近球形或木魚狀,由多數開裂之心皮組成,為大多數不結果。花 期夏、秋間,果期秋季。分布於中國大陸浙江、福建、湖南、湖北、 24 .
(37) . 廣東、廣西、四川、雲南等地。台灣於 1661 年引進,現今全境普遍 栽種。白木蘭《福建藥物誌》,白蘭花(四川),白蘭(廣州),白緬花、 緬桂花(雲南),黃桷蘭(四川)。(李幸祥, 2007) 2. 化學成分 根及樹皮含黃心樹寧(ushiruninine),氧化黃心樹寧 (oxoushinsunine),白蘭花鹼(michelalbine),柳葉木蘭鹼(salicfoline), 掌葉防己鹼(palmatine),藥根鹼(jatrooohizine),尖刺鹼(oxyacanthine)。 葉含芳樟醇(linalool),甲基丁香油酚(methyleugenol),苯乙醇(phenyl ethyl alcohol)。花含揮發油,有順式-氧化芳樟醇(cis-linalooloxide), 反式-葛縷醇(trans-carveol),甲基異丁香酚(methyl-isoeugenal),芳樟 醇,β-蒎烯(β-pinene),月桂烯(myrcene),沉香醇(agarol),羅勒烯 (ocimene),檸檬烯(limonene)。. (二) 咖啡 1. 概述 學名:Coffea arabica,茜草科多年生常綠小喬木或灌木。高約4~9 m,枝條平展。葉對生,披針形,中肋與第一側脈交叉處具凸腺,葉 面具蠟質。花腋出,白色叢生。漿果簇生於側枝對生葉片間,長橢圓 形,紅色或深紫色;種子卵形。臺灣零星栽培於雲林、嘉義及南投等 地。(郭昭麟, 2006) 25 .
(38) . 2. 化學成分(Ranheim and Halvorsen, 2005) 咖啡果實含diterpenoid alcohols類,如cafestol, kahweol。生物鹼如 caffeine,及有機酸caffeic acid, chlorogenic acid。 3. 活性研究 A. 抗病毒 研磨咖啡熱水萃取物與即溶咖啡溶液皆能藉由殺病毒活性與抑 制傳染性病毒子代於受感染細胞內形成達到抑制第I型單純皰疹病毒 (HSV-1)增殖(Namba and Matsuse, 2002) 。咖啡中主要的成分 caffeine、quinic acid與chlorogenic acid,其中僅caffeine具抑制病毒增 殖的作用,但不具病毒毒殺作用,推測咖啡中尚有其他化合物具有殺 病毒活性。咖啡水萃取物也能夠抑制小兒麻痺病毒增殖,但不具毒殺 該病毒的活性。咖啡水萃取物能夠抑制B型肝炎病毒表面抗原表現 (Utsunomiya et al., 2008) 。 B. 抗氧化 咖啡水萃取物能抑制超氧自由基、脂質過氧化(Namba and Matsuse, 2002),咖啡酸塗抹裸鼠腹部皮膚能降低UVA所誘導產生之 活性氧,於口服給藥後亦可發現咖啡酸分布於皮膚中(Yamada et al., 2006)。咖啡之綠原酸於可清除NO2預防N2O3形成(Takahama et al., 2007)。 26 .
(39) . C. 其他 咖啡萃取物能抗腸桿菌(enterobacteria)(Almeida et al., 2006)。 caffeic acid與chlorogenic acid藉由提高S-adenosyl-L-homocysteine之生 成,進而抑制乳癌細胞DNA甲基化(Lee and Zhu, 2006)。於動物皮膚 實驗證實caffeic acid能抑制因照射UVB所誘導的IL-10表現與MAPK 的活化,進而達到防禦UVB所造成的免疫反應與光致癌性(Staniforth et al., 2006)。含綠原酸較多的即溶咖啡可降低體重(Thom, 2007)。於 動物實驗發現綠原酸具有抗發炎、止痛作用(dos Santos et al., 2006), 具抗輻射作用,能降低小鼠照射γ射線後的致死率(Hosseinimehr et al., 2008)。 (三) 檄樹 1. 概述 學名:Morinda citrifolia Linn.茜草科常綠小喬木,高可達 9 m以 上,小枝條具有4稜角,全株光滑無毛。單葉對生,具短柄,厚紙質, 深綠色,葉面光亮,葉片橢圓形或長橢圓形,長10~15 cm,寬5~8 cm, 大者長可達30 cm,寬15 cm,兩端均銳,全緣。托葉膜質,闊橢圓形 或半月形。花白色,簇生而成頭狀,總梗單一,長約3~4 cm,常與葉 對生。萼杯狀,邊緣截狀。花冠圓筒形,長約1.2 cm,先端5裂,冠 喉有毛。雄蕊5枚,著生於喉部。柱頭2裂。果實為聚合果,由肉質、 27 .
(40) . 擴大而合生的花萼組成,球形,徑約4~7 cm,漿質,果實剛開始時為 綠色,後轉變為黃色,成熟時幾乎變為白色,內有很多種子。全年可 以開花與結果。花期6~8月較盛。 2. 化學成分 檄 葉 含 roseoside II , deacetyl asperuloside , kaempferol-3-O-α -Lrhamnopyranosyl-1(1→6)-β -D-glucopyranoside , ursolic acid , quercetin-3-O-α -L-rhamnopyranosyl-1(1→6)-β -D-glucopyranoside , 13-hydroxy-9,11,15-octadecatrienoic acid,pteryxin,peucedanocoumarin III,5,15-dimethylmorindol (11),8) barbinervic acid,clethric acid, rotungenic acid,hederagenin,oleanolic acid,pheophorbide a,methyl pheophorbide β , methyl pheophorbide α , 151(S)-hydroxypurpurin-7 lactone dimethyl ester,132(R)-hydroxypheophorbide a methyl ester, 151(R)-hydroxypurpurin-7. lactone. dimethyl. ester. ,. 132(S)-hydroxypheophorbide a methyl ester,3-O-acetylpomolic Acid,13-epi-phaeophorbide a methyl ester和phytol等成分(Takashima et al., 2007)。 3. 活性研究 A. 降血糖 檄樹甲醇萃取物分離出之damnacanthol-3-O-beta-D-primeveroside, 28 .
(41) . lucidin 3-O-beta-D-primeveroside具有降低streptozotocin所誘導的糖尿 病小鼠血糖之效果(Kamiya et al., 2008)。並能提高肉芽組織重量,增 加組織內羥脯胺酸含量,促進糖尿病老鼠傷口癒合(Nayak et al., 2007)。 B. 保肝 檄樹果汁對於S.D.大鼠由四氯化碳所誘發造成的急性肝臟損傷具 保護的效果(Wang et al., 2008)。 C. 預防動脈硬化 檄 樹 果 汁 可 預 防 動 脈 硬 化 , 其 所 含 成 分 3,3'-bisdemethylpinoresinol, americanol A,morindolin和isoprincepin 有明顯抑制由銅所誘導的LDL氧化(Kamiya et al., 2004)。 D. 其他 檄樹葉含iridoids成分可抑制UVB所誘導的AP-1的活性(Sang et al., 2001b; Sang et al., 2003),其乙醇萃取物的正己烷層,具抗結核菌的活 性(Saludes et al., 2002)。檄樹果汁可藉活化CB2 receptors與抑制IL-4 達到調控免疫系統的作用並提高IFN-γ產生(Palu et al., 2008)。檄樹果 甲醇萃取物可與GABA競爭GABAa接受器,具有抗焦慮與鎮靜效果 (Deng et al., 2007)。. 29 .
(42) . 第三章 材料與方法 一、 實驗材料 (一)試藥與溶媒 Sigma Chemical Co. (St. Louis, MO, U.S.A.) Sigma Chemical Co. (St. 2-Mercaptoethanol Louis, MO, U.S.A.) 聯工化學廠股份有限公司 Acetic acid (Taiwan) Acryamide/Bis, 37.5:1, 40 % J.T. Baker, Inc. solution (Phillipsburg, NJ, U.S.A.) Sigma Chemical Co. (St. Agarose Louis, MO, U.S.A.) Sigma Chemical Co. (St. Albumin from bovine serum , BSA Louis, MO, U.S.A.) USB corporation Amido black (Cleveland, OH, U.S.A.) Sigma Chemical Co. (St. Ammonium persulfate Louis, MO, U.S.A.) Bio-Rad Laboratories Bradford Reagent (California,U.S.A) Sigma Chemical Co. (St. Bromophenol blue Louis, MO, U.S.A.) Sigma Chemical Co. (St. Caffeic acid Louis, MO, U.S.A.) Sigma Chemical Co. (St. Calcium Chloride (CaCl2) Louis, MO, U.S.A.) Sigma Chemical Co. (St. Chlorogenic acid Louis, MO, U.S.A.) Calbiochem (San Diego, Collagenase, Type I U.S.A) USB corporation Coomassie blue R-250 (Cleveland, OH U.S.A.) Dibasic sodium phosphate USB corporation. 1. 1, 2-Propandiol 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16.. 30 .
(43) . (Cleveland, OH U.S.A.) 17. Dimethyl sulfoxide 18. DL-Dithiothreitol 19. Disodium dehydrate (EDTA) 20. Doxycycline hyclate 21. Dulbecco’s Modified Eagle Media(DMEM) 22. Donkey anti-goat IgG-HRP: sc-2020 23. ECL western blotting kit 24. Elastase substrate IV 25. Elastase, Porcine 26. Ethyl alcohol. 聯工化學廠股份有限公司. 27. Ethyl acetate 28. 29. 30. 31. 32. 33. 34.. (Taiwan) Santa Cruz Biotechnology, ERK 1 (C-16): sc-93 Inc. (California) GIBCOTM Fetal Bovine Serum (FBS) Invitrogen Co.(NY, U.S.A) Full-Range rainbow molecular Amersham Biosciences weight makers (England) Sigma Chemical Co. (St. Gelatin Louis, MO, U.S.A.) Sigma Chemical Co. (St. Glycerol Louis, MO, U.S.A.) USB corporation Glycine (Cleveland, OH, U.S.A.) Santa Cruz Biotechnology, Goat anti-mouse IgG-HRP: sc-2005 Inc. (California) 31 . . Sigma Chemical Co. (St. Louis, MO, U.S.A.) Sigma Chemical Co. (St. Louis, MO, U.S.A.) USB corporation (Cleveland, OH U.S.A.) Sigma Chemical Co. (St. Louis, MO, U.S.A.) GIBCOTM Invitrogen Co.(NY, U.S.A) Santa Cruz Biotechnology, Inc. (California) Amersham Biosciences (England) Calbiochem (San Diego, U.S.A) Calbiochem (San Diego, U.S.A) 台灣糖業股份有限公司 (Taiwan).
(44) . 35. Goat anti-rabbit IgG-HRP: sc-2004 36. Hydrochloric acid 37. lgepalTM CA-630 38. JNK1 (G-13): sc-46006 39. Leupeptin 40. Methyl alcohol 41. MMP substrate I 42. MMP-1(L-20): sc-12348 43. MMP-3 (1B4): sc-21732 44. MMP-9 (6-6B): sc-12759 45. Monobasic potassium phosphate 46. Monobasic sodium phosphate. Santa Cruz Biotechnology, Inc. (California) J.T. Baker, Inc. (Phillipsburg, NJ, U.S.A.) USB corporation (Cleveland, OH, U.S.A.) Santa Cruz Biotechnology, Inc. (California, U.S.A) Sigma Chemical Co. (St. Louis, MO, U.S.A.) J.T. Baker, Inc. (Phillipsburg, NJ, U.S.A.) R&D systems (U.S.A) Santa Cruz Biotechnology, Inc. (California) Santa Cruz Biotechnology, Inc. (California, U.S.A) Santa Cruz Biotechnology, Inc. (California, U.S.A) USB corporation (Cleveland, OH, U.S.A.) USB corporation (Cleveland, OH, U.S.A.). 47. N, N, N’, N’-tetra Invitrogen Co.(NY, U.S.A) -methylethylene-diamine (TEMDE) New Zealand Milk. Ltd. 48. Non-fat milk (New Zealand) GIBCOTM 49. Penicillin-Streptomycin-Glutamine Invitrogen Co.(NY, U.S.A) Santa Cruz Biotechnology, 50. p38 (A-12): sc-7972 Inc. (California, U.S.A) 51. p-p38 (Thr 180/Tyr 182)-R: Santa Cruz Biotechnology, sc-17852-R Inc. (California, U.S.A) Santa Cruz Biotechnology, 52. p-JNK (Thr 183/Tyr 185): sc-12882 Inc. (California, U.S.A) 53. p-ERK 1/2 (Thr 202/Tyr 204): Santa Cruz Biotechnology, sc-16982 Inc. (California, U.S.A) 54. Phenylmethanesulfonyl fluoride Sigma Chemical Co. (St. 32 .
(45) . (PMSF). Louis, MO, U.S.A.). USB corporation (Cleveland, OH, U.S.A.) Santa Cruz Biotechnology, Procollagen Type I (N-17): sc-8782 Inc. (California, U.S.A) J.T. Baker, Inc. Sodium Chloride (NaCl) (Phillipsburg, NJ, U.S.A.) Sigma Chemical Co. (St. Sodium deoxycholate Louis, MO, U.S.A.) USB corporation Sodium dodecyl sulfate (SDS) (Cleveland, OH, U.S.A.) Sigma Chemical Co. (St. Sodium orthovanadate (Na3VO4) Louis, MO, U.S.A.) Thiazolyl Blue Tetrazolium USB corporation Bromides (MTT) (Cleveland, OH, U.S.A.) USB corporation Tris (Cleveland, OH, U.S.A.) Sigma Chemical Co. (St. Trypan blue solution (0.4 %) Louis, MO, U.S.A.) Trypsin-EDTA (0.5 % Trypsin, GIBCOTM EDTA‧4Na) Invitrogen Co.(NY, U.S.A). 55. Potassium chloride (KCl) 56. 57. 58. 59. 60. 61. 62. 63. 64.. USB corporation (Cleveland, OH, U.S.A.). 65. Tween 20. (二)儀器設備、耗材 1. Electrophoretic transfer cell 2. Mini Vertical Gel Electrophoresis Apparatus 3. Luminescent image analyzer (LAS-4000) 4. Power supply MP500V 5. Forma Direct Heat CO2 Incubator HEPA class 100 33 . Major Science (Taiwan) Major Science (Taiwan) Fujifilm Corporation (Japan) Major Science (Taiwan) Thermo Electron Corporation (Vantaa, Finland).
(46) . 6. Ultraviolet crosslinkers CL-1000M 7. Luminescence spectrometer LS 50B 8. Microplate reader (Multiskan Ascent) 9. 高速離心機 Hsiangtai CFT-6000 10. 渦旋振盪器 Vortex Genie G-560 11. 超音波振盪器 Soniclean 2000HT 12. 氮氣濃縮裝置 N-EVAP 112 R-MT 13. 分析天平 AB 104 14. 水壓抽氣機 Eyela Aspirator A-2S. UVP, Inc. (Upland, California, U.S.A) PerkinElmer Inc. (Massachusetts, U.S.A) Thermo Electron Corporation (Vantaa, Finland) Hermle Labotechnik GmbH (Wehingen, Germany) Scientific Industries Inc. (Bohemia, NY, U.S.A.) Soniclean.Pty.Ltd (Australia) Organomation Associates Inc. (Berlin, MA, U.S.A.) Mettler Toledo (Switzerland) Tokyo Rikakikai Co. Ltd. (Tokyo, Japan). 15. 高壓滅菌鍋 TM-329. Tomin Medical Equipment CO., Ltd. 16. 電器水浴鍋 WB-24. Shin Kwang (Taiwan). Gilson S.A.S. (Entrepreneurs, 17. 微量移液管 pipette 2-20 μL, 10-100 μL, 20-200 μL, Villiers Le Bel, France) 100-1000 μL Sartorius-Stedim Biotech 18. 0.2 μm Filter sterile-EO (Goettingen, Germany) Terumo Medical Corporation 19. 拋棄式注射針及針筒 (Elkton, MD, U.S.A.) 1.0 mL syringe (0.45×13 mm) Axygen Scientific, Inc. (Union City, CA, U.S.A.). 20. 微量吸管尖 tips (200 μL, 1000 μL) 21. 微量離心管 Microtubes (1.5 mL). Axygen Scientific, Inc. (Union City, CA, U.S.A.) Corning Incorporated Costar® (U.S.A). 22. 96-Well plate. 34 .
(47) . Corning Incorporated Costar® (U.S.A) Corning Incorporated Costar® (U.S.A) Corning Incorporated Costar® (U.S.A) Amersham Biosciences (England). 23. 96-Well plate (non-sterile) 24. 75 cm2 Cell Culture Flask 25. 10 cm2 Cell Culture dish 26. PVDF transfer membrane. (三)藥材 1. 咖啡葉. 採自雲林縣林內鄉. 2. 玉蘭花葉. 採自彰化縣田尾鄉. 3. 檄樹葉. 採自彰化縣花壇鄉. 二、 實驗方法 (一) 咖啡、玉蘭花及檄樹之葉甲醇萃取物之製備 1. 甲醇萃取物之製備 秤取咖啡、玉蘭花及檄樹的乾燥葉各 10 g,分別加入甲醇 300 mL,浸潤 60 分鐘後,以超音波震盪器震盪 1 小時,經布氏漏斗抽氣 過濾。同上步驟再萃取一次,合併濾液,減壓濃縮至浸膏狀,置於抽 氣櫃揮去溶媒後,乾固物儲存於乾燥箱備用。 2. 酸水解物之製備 取上述咖啡、玉蘭花及檄樹甲醇萃取物 10 mg,分別各加入 1.2 N、2.4 N HCl 2 mL 混勻後,置於 80℃水浴鍋中反應 30 分鐘後,以 35 .
(48) . 等容之乙酸乙酯分配萃取兩次,以 6000 rpm 離心 5 分鐘後,分取上 層,合併上層液體,以減壓濃縮設備濃縮至乾,置於- 20℃下備用。 (二) 明膠分解試驗 (gelatin digestion assay)(Kim et al., 2006b) 本研究以細菌膠原蛋白酶進行初篩,即以明膠分解試驗(gelatin digestion assay)評估其活性,進而測定對於細菌膠原蛋白酶的抑制率。 1. 試劑之配製 (1) 2倍緩衝溶液之配製(50 mM Tris–HCl, 10 mM CaCl2, 0.15 M NaCl, pH 7.8) 取1 M Tris–HCl (pH 7.8) 5 mL, 1 M CaCl2 1 mL, 4 M NaCl 3.75 mL,加二次水至100 mL。 (2) 1 % 明膠培養基之製備 秤取 agarose 1 g,加入2倍緩衝溶液45 mL,二次水45 mL以及以2 倍緩衝溶液配製的1.5 % 明膠溶液10 mL,置於微波爐中加熱至起泡 溶解。趁熱倒入13 mL於製膠容器(8 cm × 6 cm)中,靜置凝固,冷藏 備用。 (3) 細菌膠原蛋白酶之配製 取膠原蛋白酶 100 mg 以緩衝溶液(50 mM Tris-HCl pH7.8,3.6 mM CaCl2)少量溶解的,待完全溶解後定容至 10 mL。並分裝至微量離心 管中,儲存於-20℃備用。 (4) Doxycycline溶液之配製 36 .
(49) . 秤取doxycycline 0.05 g,加1 mL 水溶解;置於-20℃備用。 (5) 檢品溶液之配製 咖啡、玉蘭花及檄樹萃取物及其水解物以50 %丙二醇溶解,使成 檢品濃度為10 mg/mL。 (6) 染色液之配製 取 Coomassie blue R-250 1.25 g,amido black 0.5 g,加入甲醇 227 mL及冰醋酸 46 mL,溶解後加水至500 mL。 (7) 退染液之配製 取甲醇 50 mL、冰醋酸 75 mL混合後,加水至1000 mL。 2. 實驗操作步驟 於微量離心管中加入2倍緩衝溶液50 μL、水 30 μL、膠原蛋白酶 (0.1 mg/mL) 10 μL、檢品溶液或 doxycycline 溶液(做為正對照) 10 μL 混合均勻,於室溫下反應60分鐘。將反應產物以微量移液管吸取40 μL 滴於明膠培養基上的濾紙片,置於37℃,溫孵18小時後,移除濾紙片, 以染色液染色。再以退染液退染至適當顏色對比。拍照後,影像以 TINA軟體分析,計算其抑制率。. 3. 抑制率之計算. 抑制率(%) = . . ( C-B). . ( A-B). × 100% 37 .
(50) . A : 無酶,無檢品 B : 無檢品 C : 有檢品 進行三次獨立試驗後,計算平均值及標準偏差 (standard deviation, S.D.)。 (三) 膠原蛋白酶抑制試驗(Kim et al., 2006b) 細菌膠原蛋白酶,以不同濃度檢品處理後,再以螢光受質偵測其 抑制酵素活性及其是否具濃度依存性。 1. 試劑之製備 (1) 10倍緩衝溶液 1 M Tris (pH 7.8). 5 mL. 1 M CaCl2. 1 mL. 4 M NaCl. 3.75 mL. H2O. 0.25 mL. (2) Fluorogenic Peptide Substrate I 溶液之製備 螢光受質粉末 1 mg加入水1 mL溶解,置於-20 ℃備用。使用前再 以水稀釋至適當濃度。 2. 實驗操作步驟 取水 480 μL注入微量離心管中,再加入10倍緩衝溶液 80 μL、檢 品 80 μL、膠原蛋白酶 80 μL (0.01 mg/mL)、螢光受質 80 μL (10 38 .
(51) . uM),混合均勻後,放入37℃培養箱中溫孵20小時。分別於328 nm測 激發光,393 nm測放射光。 3. 抑制率之計算 抑制率(%) =. (A-B)-(C-D) ( A-B). × 100%. A : 有酶,無檢品 B : 無酶,無檢品 C : 有酶,有檢品 D : 無酶,有檢品 進行三次獨立試驗後,計算平均值及標準偏差 (standard deviation, S.D.)。 (四) 彈力蛋白酶抑制試驗(Kim et al., 2007; Tsukahara et al., 2006) 本試驗以猪胰臟的彈力蛋白酶進行初篩。 1. 試劑之製備 (1) 彈力蛋白酶之製備 取500 U彈力蛋白酶以10 mM Tris pH 6.0緩衝溶液 5 mL溶解,分 裝至微量離心管中,置於-20 ℃備用。 (2) 彈力蛋白酶受質 IV 溶液之製備 取彈力蛋白酶受質IV粉末5 mg,以100 mM Tris pH 8.0緩衝溶液 5 mL溶解,分裝至微量離心管中,置於-20 ℃備用。 39 .
(52) . 2. 實驗操作步驟 取100 mM Tris pH 8.0緩衝溶液100 μL加入96孔盤中,再依序加入 彈力受質IV溶液(2.5 mg/mL) 25 μL、檢品50 μL、彈力蛋白酶(1 U/mL) 25 μL。混合均勻,靜置於室溫20分鐘,以ELISA reader於波長405 nm 讀取OD值。 3. 抑制率之計算 抑制率(%) = 1 -. . ( C-D) × 100% ( A-B). A : 有酶,無檢品 B : 無酶,無檢品 C : 有酶,有檢品 D : 無酶,有檢品 進行三次獨立試驗後,計算平均值及標準偏差 (standard deviation, S.D.)。 (五) 細胞培養 (1) Hs 68細胞培養條件 BCRC編號. 60038. 組織來源. Human foreskin fibroblast. 細胞來源. 食品工業發展研究所. 40 .
(53) . 培養基. 90% Dulbecco's modified Eagle's medium with 4 mM L-glutamine adjusted to contain 1.5 g/L sodium bicarbonate and 4.5 g/L glucose + 10% fetal bovine serum. 培養條件. 37℃, 5% CO2. 冷凍培養基. 93% culture medium + 7% DMSO. 細胞型態. (六) 纖維母細胞之UVB照射實驗 先確立UVB照射纖維母細胞誘發基質金屬蛋白酶試驗模式之 UVB照射劑量,再據以UVB光源照射經不同濃度檢品處理之纖維母 細胞,供後續活性抑制試驗探討。 1. 檢品製備 取檢品100 mg,以1 mL DMSO復溶。於無菌操作台內以0.2 μm無 菌濾膜過濾。再以培養液稀釋至所需濃度。 2. 實驗操作步驟 (1) 纖維母細胞之檢品處理與UVB照射 待纖維母細胞長至緻密度80%後,將培養液換成不同濃度檢品培 養液。15分鐘後,將培養液吸除後,加入PBS溶液潤洗2次,再加入 PBS溶液。將細胞置於UV燈源下,以UV照射後,吸除PBS溶液再加 入無血清培養液繼續培養24小時。 41 .
(54) . (七) 分離細胞內細胞質之蛋白質 本試驗以破碎緩衝溶液破碎細胞再以離心的方式,取得位於細胞 質之蛋白質,以備蛋白質分析用。 1. 緩衝液之製備 (1) RIPA 緩衝液 取1 M Tris-HCl (pH 7.4)25 mL,NaCl 4.38 g,DL-dithiothreitol 0.0771 g,Sodium deoxycholate 2.5 g,0.5 M EDTA 1 mL,IgepalTM CA-630 5 mL及10 % SDS 5 mL,置於血清瓶中,加二次水至500 mL, 混溶滅菌備用。. (2) 破碎緩衝溶液 將 1 mL RIPA緩衝溶液新鮮加入10 mM Na3VO410 μL,Leupeptin 2 μL (10 mg/mL)及10 μL PMSF(10 mg/mL)即得。 2. 實驗操作步驟 (1) 萃取蛋白質 將培養液吸除,以預冰之PBS溶液清洗。後續的步驟皆於冰上操 作。加入破碎緩衝溶液適量,以刮棒將細胞全數刮下,移至微量離心 管。以12100×g 於4℃離心10分鐘後,吸取上清液,上清液進行蛋白 質定量。 (2) 蛋白質定量 42 .
(55) . 以不同濃度之Albumin from bovine serum (2、4、6、8、10 mg/mL) 與檢品加入Bradford’s reagent (使用前以預冰之二次水稀釋5倍) 200 μL,反應10分鐘後 (不可超過1小時),於波長595 nm測定吸光值。 (八) 西方墨點法 以從細胞質分離出之蛋白質,以電泳法分離後,再測定特定蛋白 質量。 1. 緩衝液與試劑之製備 (1) Upper gel buffer-stacking buffer (0.5 M Tris-HCl, pH 6.8) 稱取Tris 30.3 g 置於燒杯中,加入水約300 mL,以磁石攪拌機混 勻後,以12 N HCl 調整 pH值 至 6.8,再倒入量筒中,加水至500 mL,倒入血清瓶中,高壓滅菌後備用。 (2) Lower gel buffer-running buffer (1.5 M Tris-HCl, pH 8.8) 稱取Tris 90.6 g置於燒杯中,加入水約300 mL,以磁石攪拌機混 勻後,以12 N HCl 調整 pH 至 8.8,再倒入量筒中,加水至500 mL, 倒入血清瓶中,高壓滅菌後備用。 (3) 10倍 Laemmli tank buffer 稱取 Tris 15.1 g及 glycine 93.8 g置於燒杯中,倒入量筒中,加水 至500 mL,倒入血清瓶中,高壓滅菌後備用。此貯備溶液使用前需稀 釋成1倍,稀釋方式如下:取Laemmli tank buffer (10倍)50 mL及10% SDS 5 mL,加水至 500 mL即得。 43 .
(56) . (4) 8倍 Transfer buffer 稱取 Tris 24.2 g及 glycine 115.4 g置於燒杯中,倒入量筒中,加 水至1 L (pH 8.3),倒入血清瓶中,高壓滅菌後備用。使用前需稀釋成 1倍,稀釋方式如下:取貯備溶液 250 mL,甲醇 400 mL,加水至2 L, 貯於4℃中備用。 (5) 5倍 TBST a. 1 M Tris-HCl (pH 7.5):稱取Tris 60.6 g,加入約300 mL水,以 磁石攪拌機混勻,以 12 N HCl 調整 pH 至 7.5,加水至500 mL,經 高壓滅菌後備用。 b. 4 M NaCl:稱取NaCl 233.8 g,加水至500 mL,經高壓滅菌後 備用。 c. 取 4 M NaCl 溶液312.5 mL、1 M Tris-HCl 50 mL、Tween 20 2.5 mL及135 mL H2O混合均勻,即成5倍TBST,冰存於4℃冰箱,使用前 以水稀釋成1倍即得。 (6) 4倍檢品緩衝液 取 glycerol 4 mL、bromophenol blue 40 mg、upper stacking buffer 5 mL、SDS 400 mg 混合均勻後,分裝至微量離心管中,每管0.9 mL, 貯存於 -20℃,使用前加 2-mercaptoethanol 100 μL,須保存於4℃冰 箱。 44 .
(57) . (7) 10% SDS溶液 取SDS1 g溶於10 mL 水中。 (8) 10% APS溶液 取APS 0.1 g 溶於1 mL水中。 (9) MTBST (Tris-Buffered Saline Tween-20 contain 5% non-fat milk) 取TBST 50 mL 加入2.5 g脫脂奶粉。 2. 實驗操作步驟 取各細胞檢品相當於30 μg之蛋白質與4倍檢品緩衝液以3:1比例 混合後,以二次水調整體積,置沸水中水浴鍋5分鐘。以10% 之 SDS-PAGE,電壓70伏特,進行電泳30分鐘,再以電壓140伏特,進 行電泳2小時。以400毫安培通電100分鐘將其轉印至 PVDF 轉印膜 上。將MTBST加於轉印膜,搖盪2小時後,以1倍TBST清洗數次,再 加入一級抗體 (以含5 % Albumin from bovine serum之TBST稀釋, β-actin 稀釋 1000 倍;MMP-1、MMP-3、MMP-9、type I procollagen、 ERK、JNK、p38、p-ERK、p-JNK、p-p38 稀釋 500 倍),置於 4℃ 冰 箱中搖盪至隔夜(至少16小時) ,回收一級抗體,再以TBST清洗數次 後,加入二級抗體 (以含5%BSA之TBST稀釋 5000 倍),置於室溫搖 盪 2 小時,回收二級抗體,再以1倍TBST清洗數次。最後移去1倍 TBST 加入ECL試劑,以LAS-4000顯影,multi Gauge V2.2定量分析。 (九) 細胞毒性試驗(MTT assay) 45 .
(58) . 本實驗以不同濃度之檢品處理纖維母細胞,以了解其細胞毒性。 1. 試劑之製備 (1) MTT 溶液 (5 mg/mL) 之配製 稱取MTT 100 mg溶解於PBS溶液 20 mL中,使成5 mg/mL,以褐 色微量離心管分裝,置於-20℃備用。 (2) SDS 溶液 (10 % SDS in HCl)之配製 稱取SDS 100 g 溶於0.01 N HCl 1 L中即得,避光存放。 2. 實驗操作步驟 將細胞接種於96孔盤中,每孔接種 104 cells/well 待24小時後,加 入待測檢品50 μL,經24小時培養後,加入 MTT 溶液15 μL,置37℃ 培養箱中3小時,加入 SDS溶液75 μL,隔夜於波長570 nm下測定吸 光值。 3. 抑制率之計算. 細胞存活率 (%) =. 實驗組吸光值 ×100 % 控制組吸光值. (十) 皮膚一次刺激性試驗(日本化粧品工業連合会, 2001) 1. 檢品之製備 秤取咖啡、玉蘭花萃取物0. 1 g(低劑量) , 0.5 g(高劑量)檢品, 以1 mL生理食鹽水復溶備用。 2. 實驗操作步驟 46 .
(59) . 將紐西蘭大白兔固定於兔架上,剃除背毛,以色筆畫出塗抹範圍 每格2.5 cm × 2.5 cm,共6格。於格內以無菌針頭於皮膚上輕劃4條平 行線,破壞角質層,不可見血。將檢品均勻塗佈於格內。24小時後, 以生理食鹽水輕拭,除去檢品,於24與72 小時兩個時間點觀察、評 分(如Table 13),若有刺激性產生則需延長觀察時間。 (十一) 眼球刺激性試驗(Wilhelmus, 2001; 日本化粧品工業連合会, 2001) 1. 檢品之製備 取咖啡、玉蘭花萃取物300 μg溶於1 mL生理食鹽水,以6000 rpm 離心5分鐘,取其上清液供作檢品。 2. 實驗操作步驟 取檢品100 μL,滴入白兔結膜囊,將白兔眼睛輕輕闔上。於1、5、 15、30分鐘,1、2、24、48、72小時等時間點觀察記錄、評分(如附 錄Table 14),若具刺激性須延長觀察時間。 (十二) 統計方法 以ANOVA及Student’s t test進行數據分析,p<0.05表示於統計上 有顯著差異,且每個實驗進行三次以上獨立實驗,實驗結果以 mean ± S.D. 表示。. 47 .
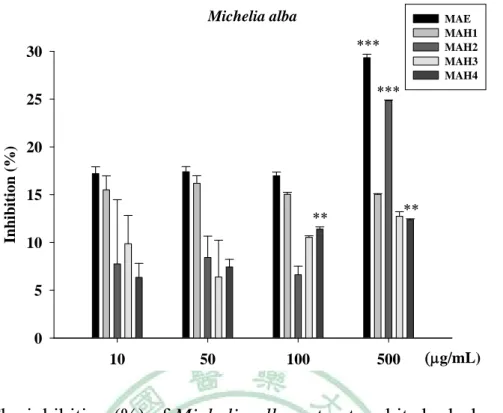
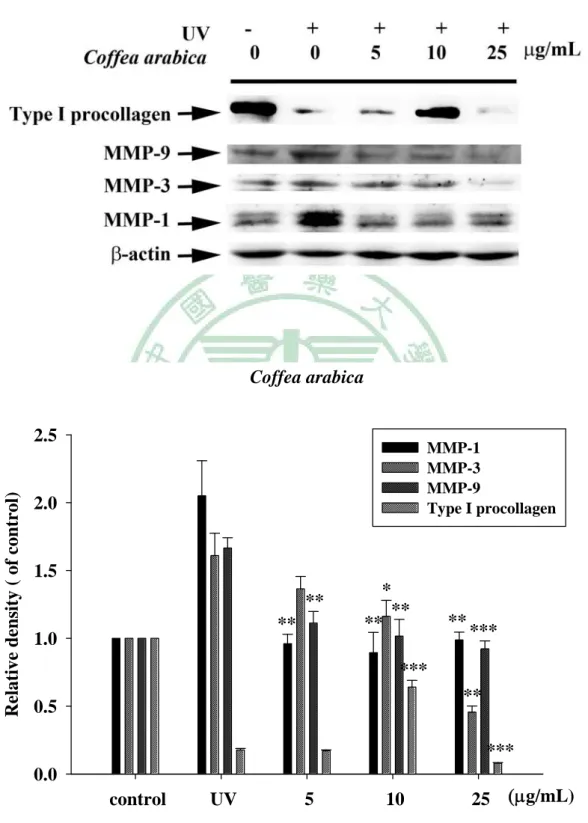
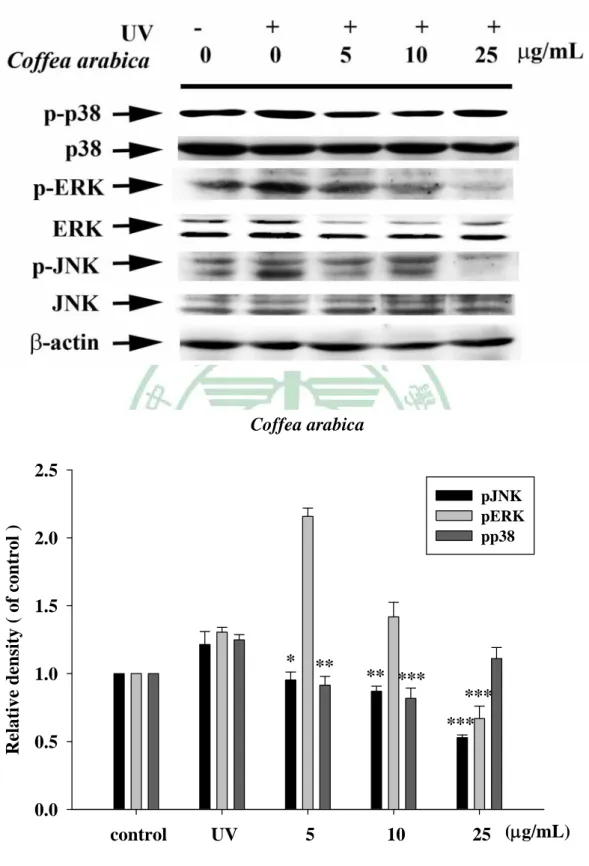
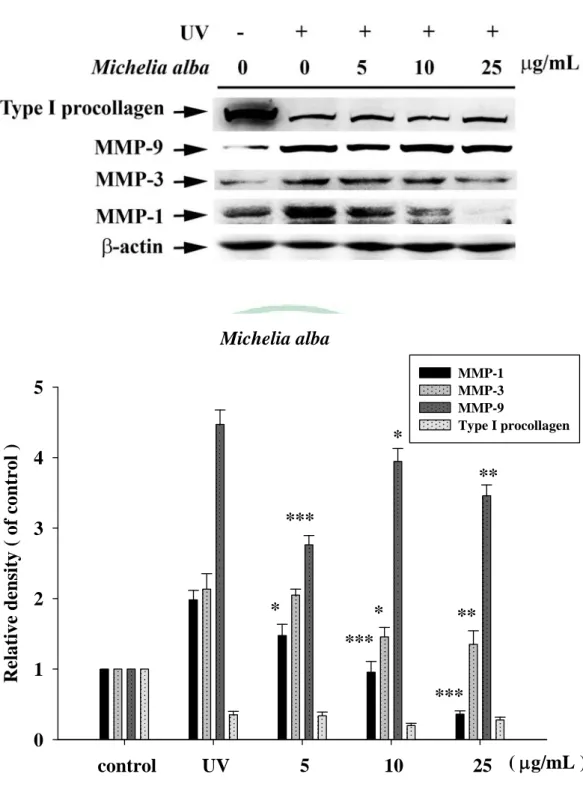
(60) . 第四章 結果 一、. 植物萃取及水解物製備. (一) 植物甲醇萃取物之製備 本試驗利用甲醇製備咖啡(CA)、玉蘭花(MA)及檄樹(MC)之葉萃 取物,所得萃取物萃取率分別為:CAE, 10.4%;MAE, 16.9%;MCE, 15.0%。 (二) 水解物之製備 水解物之水解條件,分別均於80℃水浴鍋,以1.2 N, 0.5 h;1.2 N, 1 h;2.4 N, 0.5 h及2.4 N, 1 h等四種條件處理,其代號分別為H1, H2, H3 及H4。文內各植物水解物,再冠以各植物之代號,如咖啡之不同水 解物之代號為CAH1, CAH2, CAH3及CAH4。其餘類推。 二、. 膠原蛋白酶活性抑制之試驗. (一) 明膠分解試驗 本試驗利用含明膠之洋菜培養基,評估植物萃取物及其水解物、 咖啡酸與綠原酸等對膠原蛋白酶的抑制效果,其結果如Fig. 1 (a)、 Table 2所示,正對照doxycycline (100 μg/mL ) 之抑制率為97.3± 0.02%,空白對照 (50% 1, 2-propandiol) 抑制率0.4±0.92%。咖啡萃取 物及其水解物(1000 μg/mL)之抑制率分別為CAE 100.0±0.05%,CAH1 99.5±0.06%,CAH2 96.7±0.08%,CAH3 97.0±0.07%及CAH4 95.1± 48 .
(61) . 0.23%。玉蘭花萃取物及其水解物之抑制率結果,如Fig. 1 (c)、Table 2 所示,分別為MAE 100.0±0.03%,MAH1 99.3±0.13%,MAH2 99.1± 0.14%,MAH3 92.2±0.12%及MAH4 93.7±0.40%。兩者水解物之抑制 率皆未優於其萃取物。如Fig. 1(e)、Table 2所示,檄樹萃取物及其水 解物之抑制率分別為MCE 74.3±1.04%,MCH1 -2.7±2.11%,MCH2 69.9±1.26%,MCH3 60.5±17.8%及MCH4 -3.3±0.97%。水解物之抑制 率與萃取物相比,抑制率皆無提高之情形,甚至喪失活性。以咖啡中 主要成分咖啡酸與綠原酸進行膠原蛋白酶活性抑制篩選如Fig. 1(f)所 示,抑制率分別只有26.1±0.53%與35.9±0.92%。 (二) 膠原蛋白酶抑制試驗 為更進一步確認抑制膠原蛋白酶活性效力,各植物萃取物及水解 物進行序列稀釋濃度(10 ~ 500 μg/mL)進行本試驗,惟檄樹、咖啡酸及 綠原酸於明膠分解試驗初篩效果不佳,未再進行本試驗。咖啡萃取物 及水解物對膠原蛋白酶之抑制率如Fig. 2、Table 3所示,咖啡萃取物 水解前後抑制效果皆具濃度依存性,於高濃度時(500 μg/mL)CAE及任 何條件水解物皆具有極佳的抑制效果(>95%) ,但隨著稀釋倍數增 加時,CAE(10 μg/mL)仍有84.6±1.46%的抑制率,而水解物抑制率皆 未大於60%,水解物伴隨酸的濃度增加及水解時間的延長,抑制率隨 之降低。玉蘭花萃取物及其水解產物抑制效果如Fig. 3、Table 4所示, 49 .
數據


+7

相關文件
In order to apply for a permit to employ Class B Foreign Worker(s), an Employer shall provide reasonable employment terms and register for such employment demands with local
Should an employer find it necessary to continue the employment of the Class A Foreign Worker(s), the employer shall, within four (4) months prior to the expiration of the
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
straight brown hair dark brown eyes What does he look like!. He has short
While Korean kids are learning how to ski and snowboard in the snow, Australian kids are learning how to surf and water-ski at the beach3. Some children never play in the snow
I am writing this letter because I want to make a new friend in another country.. Maybe you will come to Gibraltar
豆類和奶類 魚、肉 動物性蛋白質、植物性蛋白質 豬油、奶油.
Consistent with the negative price of systematic volatility risk found by the option pricing studies, we see lower average raw returns, CAPM alphas, and FF-3 alphas with higher
