山苦瓜改善高油高糖餵食大鼠腹部體脂堆積及胰島素抗性; Wild Bitter Melon Ameliorate Abdominal Fat Accumulation and Insulin Resistance in High Fat High Sugar Fed Rats
207
0
0
全文
(2) 目錄 縮寫對照表…………………………………………………………………..I 組別縮寫意義…………………………………………………………..…III 摘要………………………………………………………………………...IV Abstract…………………………………………………..………………..VI 第一章 前言………………………………………………………………...1 第二章 文獻回顧…………………………………………………………...2 一、. 過氧化體與過氧化體活化增殖接受器……………………….........2. (一) 過氧化體 Peroxisome……………………………………………………...………2 (二) Peroxisomal fatty acid β – oxidation………………………….…………………...2 (三) 過氧化體增殖劑 Peroxisome proliferators……………………………………….3 (四) 過氧化體增殖劑活化接受器 Peroxisome proliferator - activated receptor…....3. 二、 脂肪細胞生合成 Adipogenesis…………………………………......11 (一) 脂肪細胞生合成的 In vitro 模式………………………………………………….11 (二) 脂肪組織來自 Multipotent stem cells…………………………………………….12 (三) 脂肪細胞生合成的過程…………………………………………………………...13 (四) 脂肪細胞生合成的轉錄調控……………………………………………………...16 (五) 刺激脂肪細胞生合成的因子……………………………………………………...27 (六) 抑制脂肪細胞生合成的因子……………………………………………………...28. 三、 脂肪細胞是內分泌器官影響胰島素抗性………………………….33 (一) Secretory function of adipose tissue………………………………………………33 (二) Insulin resistance………………………………………………………………......34 (三) Adipocytokines……………………………………………………………………..36. 四、. 代謝症候群 Metabolic syndrome………………………………..43. (一) Characteristic of metabolic syndrome……………………………………………43 (二) Definition of metabolic syndrome………………………………………………...44.
(3) 五、 苦瓜 Momordica charantia..............................................................46 (一) 苦瓜的植物化學成分(Phytochemicals)…………………………………………..47 (二) 苦瓜的藥理特性........................................................................................................48 (三) 苦瓜與PPAR……………………………………………………………………….51 (四) 抗肥胖活性 Antiobesity activity………………………………………………….51 (五) 台灣山苦瓜…………………………………………………………………………53. 第三章 材料與方法……………………………………………………….55 實驗設計……………………………………………..…………………….55 實驗流程設計………………………………………..…………………….56 一、. 山苦瓜凍乾粉的製備……………………..……………………...57. 二、. 山苦瓜凍乾粉乙酸乙酯萃物……………..……………………...57. 三、. 試驗飼料之配製…………………………..……………………...57. 四、. 動物飼養…………………………………..……………………...60. 五、. 動物犧牲及樣品收集……………………..……………………...61. 六、. Glycerol-3-phosphate dehydrogenase (G3PDH)酵素活性測定 62. 七、. Lipoprotein lipase (LPL)酵素活性測定....……………………...66. 八、. 脂解速率測定....…………………….............................................70. 九、. 血清脂質分析…………….............................................................72. (一) 血清三酸甘油酯含量測定……................................................................................72 (二) 血清膽固醇含量測定…………................................................................................73 (三) 血清游離脂肪酸測定…………................................................................................74. 十、. 脂肪組織三酸甘油酯含量測定.....................................................75. 十一、 脂肪組織 DNA 含量測定...............................................................76 十二、 鋨酸染色固定……………….........................................................78 十三、 口服葡萄糖耐受測試 Oral Glucose Tolerance Test (OGTT)…81.
(4) 十四、 禁食全血葡萄糖含量測定……………………………………….81 十五、 禁食血清胰島素測定…………………………………………….82 十六、 禁食血清 Adiponectin 測定……………………………………...83 十七、 以 北 方 點 墨 法 (Northern blot) 分 析 PPARγ 、 Adiponectin 、 Leptin、LPL 及 ADD1 / SERBP1 c mRNA 表現量……………84 (一) 總 RNA 之抽取………………………………………………………..…84 (二) RNA 電泳……………………………………………………..………...85 (三) RNA 轉印.………………………………………………..……………...88 (四) 探針標誌.………………………………………………..………............89 (五) 雜交反應.………………………………………………..………............90. 十八、 統計分析……………………………………………………….... 92 第四章 結果…………………………………………………………….....93 第五章 討論……………………………………………………………...137 第六章 結論……………………………………………………………...150 第七章 參考文獻………………………………………………………...151 附錄……………………………………………………………………….185.
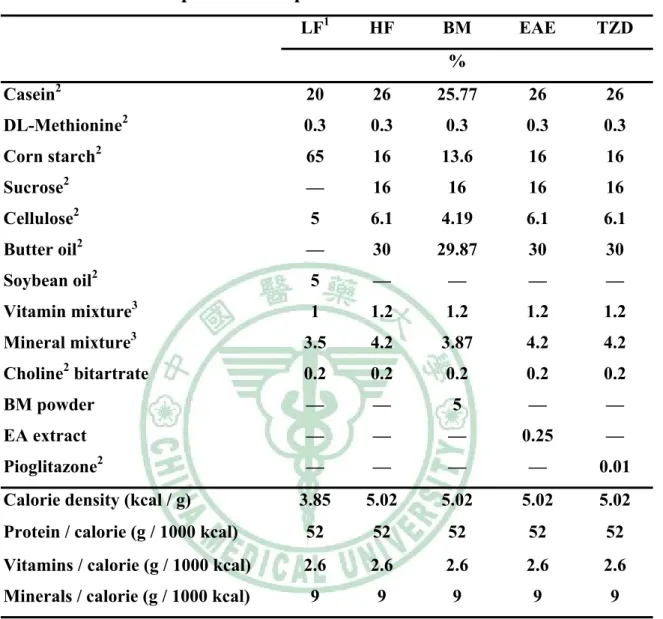
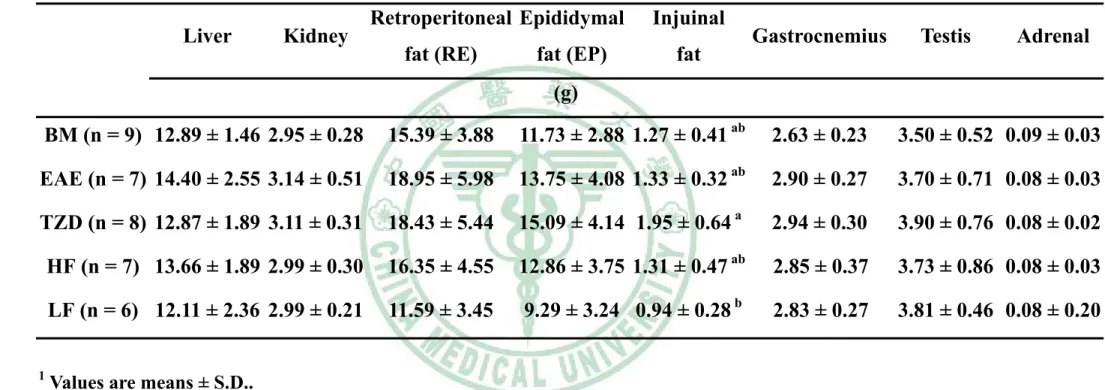
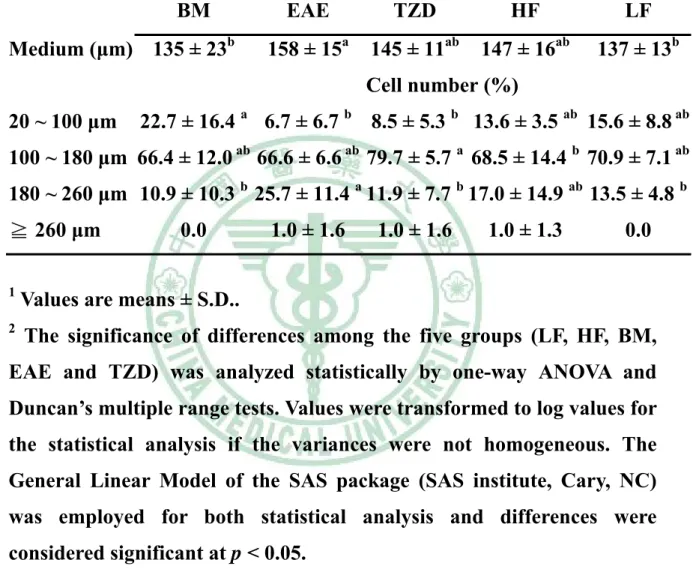
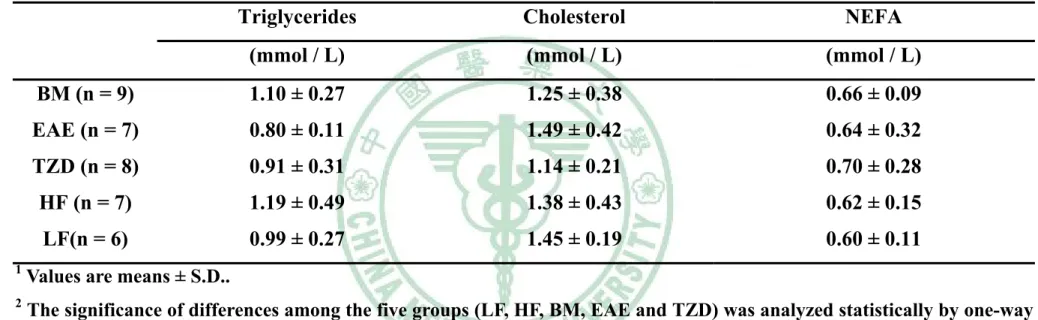
(5) 表目錄 表 3 - 1 山苦瓜凍乾粉成分分析………………………………………….58 表 3 - 2 大鼠試驗飼料組成……………………………………………….59 表 4 - 1 Wistar 大鼠餵食試驗飲食九週之初始體重、最終體重、攝食 量、能量攝取、飼料效率及能量效率………………………….100 表 4 - 2 Wistar 大鼠餵食試驗飲食九週之絕對組織重……………….111 表4-3. Wistar 大鼠餵食試驗飲食九週之相對組織重……………….112. 表 4 - 4 Wistar 大鼠餵食試驗飲食九週之腹膜後脂肪(RE)的脂肪細胞 直徑…………………………………………………………….113 表 4 - 5 Wistar 大鼠餵食試驗飲食九週之副睪脂肪(EP)的脂肪細胞 直徑…………………………………………………………….114 表 4 - 6 Wistar 大鼠餵食試驗飲食九週之腹膜後脂肪(RE)所含三酸甘 油酯、DNA 與每毫克 DNA 中所含三酸甘油酯含量……….115 表 4 - 7 Wistar 大鼠餵食試驗飲食九週之副睪脂肪(EP)所含三酸甘油 酯、DNA 與每毫克 DNA 中所含三酸甘油酯含量…………..116 表4-8. Wistar 大鼠餵食試驗飲食九週之禁食血脂質……………….117. 表 5 - 1 大鼠餵食試驗飲食脂肪組織中脂質生合成基因表現……….144.
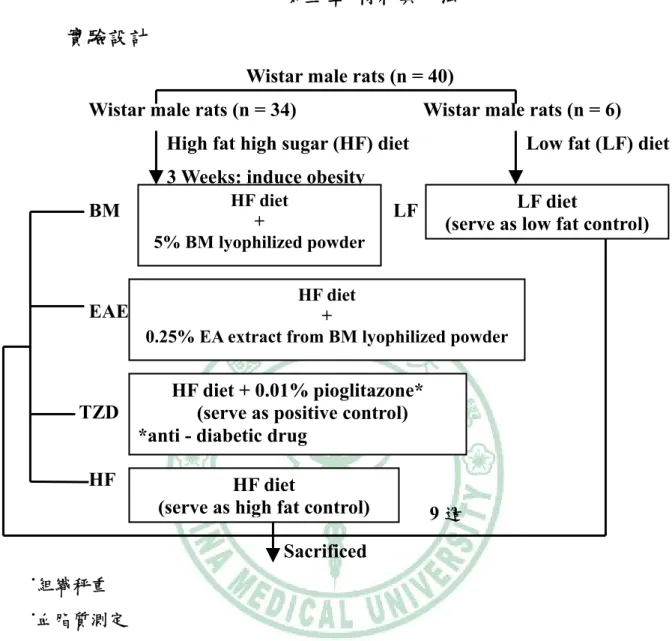
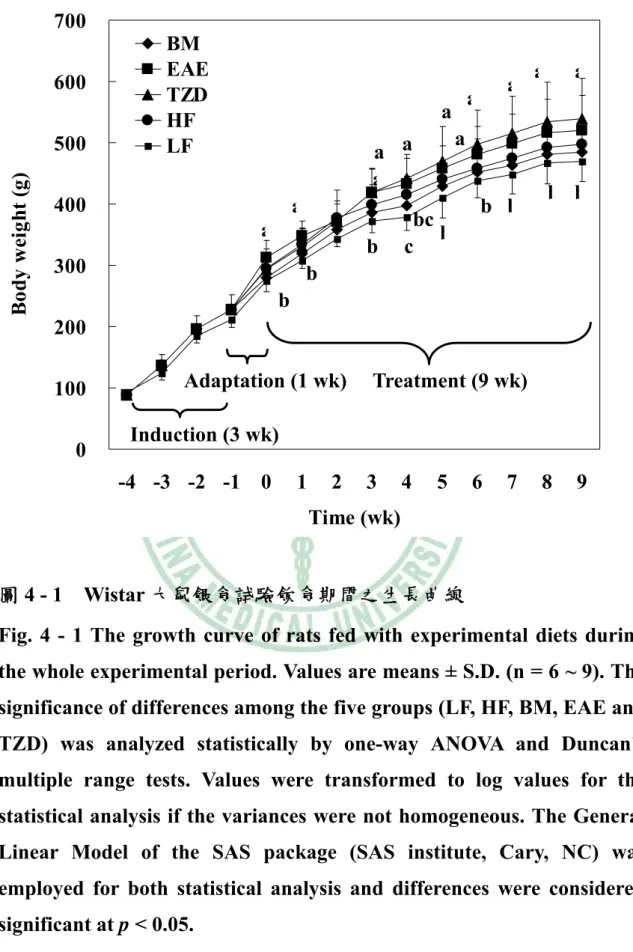
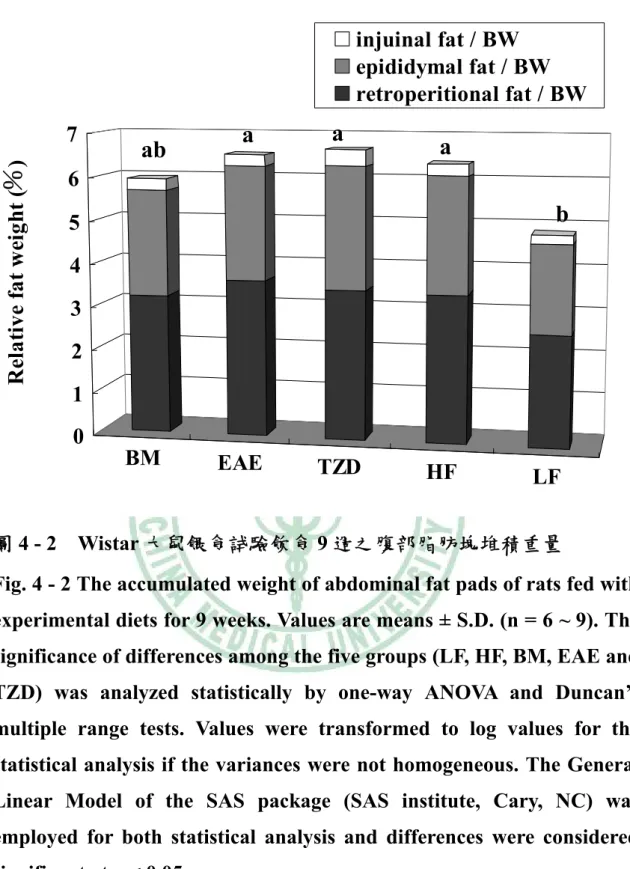
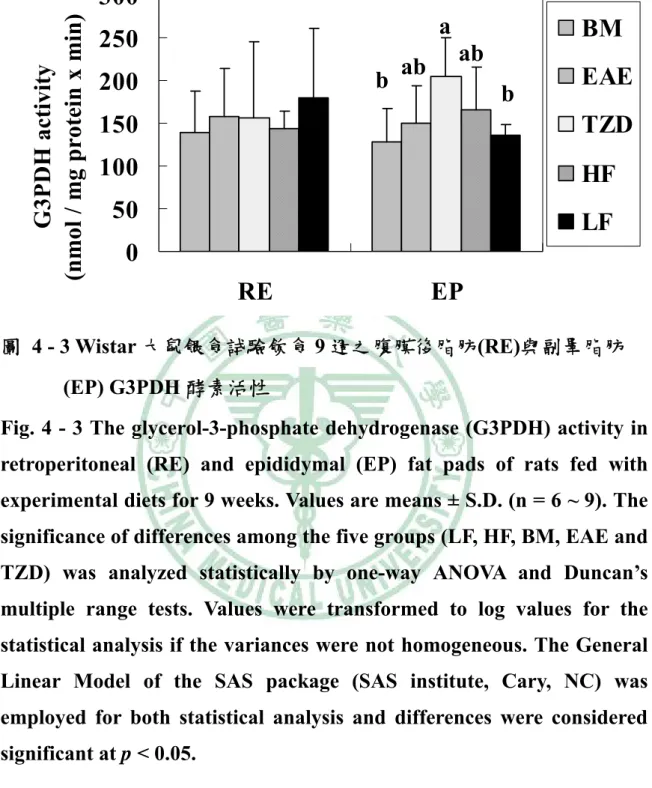
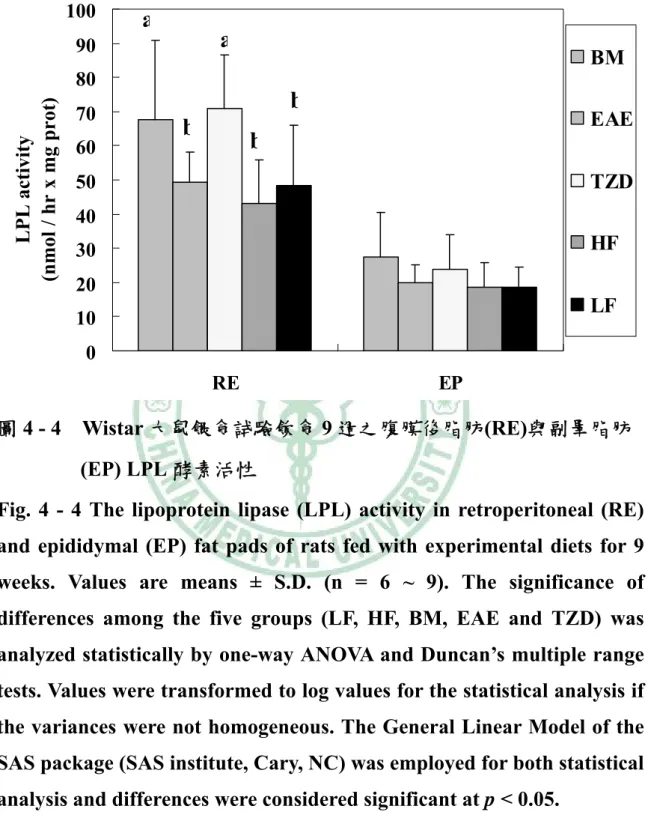
(6) 圖目錄 圖2 - 1. PPAR之結構特徵…………………..…………………………….6. 圖2 - 2. PPAR之基因轉錄作用機制…………..………………………….7. 圖2 - 3. 脂肪細胞生合成的過程………………………..………………..16. 圖2 - 4. A model of the transcriptional cascade leading to adipogenesis. ……………………………………………………………………………...25 圖2 - 5. 胰島素抗性、肥胖與代謝症候群之相關………………………44. 圖2 - 6 山苦瓜.…………………………………………………………….54 圖 3 – 1 山苦瓜改善高油高糖餵食大鼠腹部體脂堆積及胰島素抗性之實 驗大綱…………………………………………………………….55 圖 3 – 2 山苦瓜改善高油高糖餵食大鼠腹部體脂堆積及胰島素抗性 之實驗流程設計………………………………………………….56 圖4-1. Wistar 大鼠餵食試驗飲食期間之生長曲線………………….118. 圖4-2. Wistar 大鼠餵食試驗飲食 9 週之腹部脂肪塊堆積重量…….119. 圖 4 - 3 Wistar 大鼠餵食試驗飲食 9 週之腹膜後脂肪(RE)與副睪脂肪 (EP) G3PDH 酵素活性…………………………………………120 圖 4 - 4 Wistar 大鼠餵食試驗飲食 9 週之腹膜後脂肪(RE)與副睪脂肪 (EP) LPL 酵素活性……………………………………………121 圖 4 - 5 Wistar 大鼠餵食試驗飲食 9 週之腹膜後脂肪(RE)刺激與基礎 脂解速率……………………………………………………….122 圖 4 - 6 Wistar 大鼠餵食試驗飲食 9 週之副睪脂肪(EP)刺激與基礎 脂解速率……………………………………………………….123 圖 4 - 7 Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測腹膜後脂肪 (RE) PPARγ mRNA 相對量…………………………………..124.
(7) 圖 4 - 8 Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測副睪脂肪(EP) PPARγ mRNA 相對量………………………………………...125 圖 4 - 9 Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測腹膜後脂肪 (RE) Adiponectin mRNA 相對量……………………………..126 圖 4 - 10. Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測副睪脂(EP) Adiponectin mRNA 相對量………………………………...127. 圖 4 - 11. Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測腹膜後脂肪 (RE) Leptin mRNA 相對量………………………………….128. 圖 4 - 12. Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測副睪(EP) Leptin mRNA 相對量………………………………………129. 圖 4 - 13. Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測腹膜後脂肪 (RE) LPL mRNA 相對………………………………………130. 圖 4 - 14. Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測副睪(EP) LPL mRNA 相對量…………………………………………131. 圖 4 - 15. Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測腹膜後脂肪 (RE) ADD1 / SREBP1c mRNA 相對量……………………132. 圖 4 - 16. Wistar 大鼠餵食試驗飲食 9 週以北方點墨法偵測副睪脂(EP) ADD1 / SREBP1c mRNA 相對量………………………….133. 圖 4 - 17. Wistar 大鼠餵食試驗飲食 9 週之禁食全血葡萄糖變化……134. 圖 4 - 18. Wistar 大鼠餵食試驗飲食 9 週之禁食血清胰島素變化……135. 圖 4 - 19 Wistar 大鼠餵食試驗飲食 9 週之血清 Adiponectin 濃度….136 圖5 - 1. Wistar 大鼠餵食試驗飲食 9 週之 GOT 和 GPT 酵素活性…149.
(8) 附圖 1 - 1 五種山苦瓜品種乙酸乙酯萃物處理 24 小時對 3T3 - L1 前脂肪 細胞增生之影響……………………………………………...193 附圖 1 - 2 五種山苦瓜品種乙酸乙酯萃物處理 48 小時對 3T3 - L1 前脂肪 細胞增生之影響……………………………………………...194 附圖 1 - 3 TZD(BRL 49653)處理 24 及 48 小時對 3T3 - L1 前脂肪細胞 增生之影響……………………………………………….......195 附圖 1 - 4 四種山苦瓜品種乙酸乙酯萃物處理 7 天對 3T3 - L1 前脂肪細 胞分化之影響………………………………………………...196 附圖 1 - 5 四種山苦瓜品種乙酸乙酯萃物處理 11 天對 3T3 - L1 前脂肪細 胞分化之影響………………………………………………...197 附圖 1 - 6. 18 : 1 游離脂肪酸與 Conjugated linolenic acid (CLN)脂肪酸 處理 7 天對 3T3 - L1 前脂肪細胞分化之影響……………..198. 附圖 1 - 7. 18 : 1 游離脂肪酸與 Conjugated linolenic acid (CLN)脂肪酸 處理 11 天對 3T3 - L1 前脂肪細胞分化之………………….199.
(9) 縮寫對照表 縮. 寫. 全. Acrp30. 名 Adiponectin. ACC. Acetyl - CoA carboxylase. ACS. Acyl - CoA synthetase. ACO. Acetyl - CoA oxidase. ADD1. Adipocyte determination and differentiation factor 1. ADSF. Adipocyte - specific secretory factor. ALBP. Adipocyte lipid binding protein. AMPK. AMP - activated protein kinase. AP – 1 aP2. Activator Protein 1 Adipocyte specific fatty acid binding protein. bHLH. Basic helix – loop - helix. BM. Bitter. C / EBP. melon. CCAAT / enhancer binding protein. CLA. Conjugated linoleic acid. CLN. Conjugated linolenic acid Cytochrome P4504A. CYP4A DBD. DNA binding domain. DEX. Dexamethasone. ES. Embryonic stem. ECM. Extracellular matrix. FAT. Fatty acid transporter. FATP. F a t t y a c i d t r a ns p o r t p r o t e i n. FA S FIZZ G3PDH. Fatty acid synthase Found in inflammatory zone Glycerol – 3 - phosphate dehydrogenase. GCs. Glucocorticoids. GLUT. Glucose transporter. HDL. High density lipoprotein. HSL. Hormone sensitive lipase. I.
(10) IL IRS. Interleukin -. 1. Insulin. receptor. substrates. 1. LBD. Ligand binding domain. LPL. Lipoprotein lipase. LPS. Lipopolysaccharide. L X R. L i v e r. X. r e c e p r o r. MADS. Multipotent adipose - derived stem. MAPK. Mitogen - activated protein kinase. MIX. Methylisobutylxanthine. MMP. Metalloproteinase. NF - κB PAI – 1. Nuclear factor - κB Plasminogen activator inhibitor - 1. PBP PEPCK. P PA R γ. Prostaglandin 15 – deoxy - delta12 – 14 - PGJ 2. 15d - PGJ 2 PI3K. protein. Phosphenolpyruvate carboxykinase. PG PGC. binding. Peroxisome proliferator - activated receptor γ coactivator Phosphatidylinositol. P K. -. 3. P y r u v a t e. kinase k i n a s e. PPAR. Peroxisome proliferator - activated receptor. PPRE. Peroxisome proliferator responsive element. Pref – 1. Pre - adipocyte factor 1. RARs. Retinoic acid receptors. RELMs. Resistin - like molecules. RXR. Retionoid X receptor. SCD. St e a r o y l - C o A d e s a t u r a s e. SREBPs SREs. St e ro l re g ul a t o r y e l e m e n t bi nd i n g p rot e i ns Sterol. TZD. regulatory. elements. Thiazolidinedione. TRs. Thyroid hormone receptors. TNFα. Tumor necrosis factor α. UCP. Uncoupling. II. protein.
(11) 組 別 縮 寫 BM. 組別縮寫意義 縮 寫 意 義 Wild bitter melon lyophilized powder. EAE. Ethyl acetate extract form wild bitter melon lyophilized powder. TZD. Positive control (Thiazolidinedione). HF. High fat control (High fat high sugar die). LF. Low fat control (Low fat diet). III.
(12) 山苦瓜改善高油高糖餵食大鼠腹部體脂堆積及胰島素抗性 中國醫藥大學營養學系碩士班 洪雅雯. 摘要 在亞洲,苦瓜(Bitter melon; Momordica charantia)為廣泛使用的食材。已有 研究指出,苦瓜具降血糖、降血脂和抗發炎的作用,但其作用機制與功能性成分仍 不明確。近來,有研究顯示苦瓜可活化 Peroxisome proliferator - activated receptor (PPAR)α 和 γ,暗示苦瓜在預防和治療代謝症候群上具有應用之潛力。目前臨床廣 泛使用之 Thiazolidinedione (TZD)治療糖尿病藥物雖可改善胰島素敏感性,但卻具 有肥胖的副作用。因此,本研究擬利用體內模式探討山苦瓜對於改善大鼠腹部體脂 堆積和胰島素敏感性之功效並與 TZD 藥物做比較。 實驗採用 40 隻 6 週齡雄性 Wistar 大鼠,其中 34 隻先以含 30%奶油的高脂飲 食餵食 3 週誘導肥胖後,分成 4 組。BM 組(含 5%山苦瓜凍乾粉之高脂飲食)、EAE 組(含 0.25%山苦瓜凍乾粉乙酸乙酯萃物之高脂飲食)、HF 組(高脂對照)及 TZD 組(含 0.01% Pioglitazone 之高脂飲食做為正對照)。另外 6 隻大鼠餵食含 5%新鮮大豆油 (LF 組)做低脂對照。分組餵食 9 週後犧牲,分別取腹膜後脂肪、副睪脂肪和腹 股溝脂肪秤重代表腹部脂肪堆積。取腹膜後脂肪和副睪脂肪進一步分析脂肪細胞大 小、三酸甘油酯和 DNA 含量、脂質生合成酵素活性和脂解速率與脂肪細胞分化相 關基因表現。在實驗期間,每隔 3 週採取尾巴血測量禁食血糖及胰島素。 結果顯示,TZD 組在體重上顯著高於 LF 組(p < 0.05),此結果與 TZD 藥物肥 胖的副作用相符合。在腹部脂肪重量方面,HF 組、EAE 組和 TZD 組都顯著高於 LF 組(p < 0.006),而 BM 組與 LF 組無顯著差異,表示山苦瓜凍乾粉可以改善高脂 飲食誘導的腹部體脂堆積。分離自腹膜後脂肪和副睪脂肪的脂肪細胞,其大小比例 分佈的結果顯示,BM 組與 HF 組相較之下,有較多小的以及較少大的脂肪細胞。 此外,BM 組脂肪組織三酸甘油酯堆積有減少的傾向。Glycerol – 3 - phosphate dehydrogenase (G3PDH)是脂肪細胞中重要的脂質合成酵素。BM 組與 LF 組副睪脂 肪中 G3PDH 活性,皆顯著低於 TZD 組(p < 0.05)。在北方點墨定量結果中,BM 組 副睪脂肪的 ADD1 / SREBP1c 調控脂肪酸生合成之轉錄因子 mRNA 顯著低於 TZD. IV.
(13) 組(p < 0.05)。HF 組在高脂飲食下會誘發高胰島素血症,但此現象不發生在 TZD 組 和 BM 組。 以上這些結果指出山苦瓜如同 TZD 藥物可改善高脂飲食誘發之高胰島素血 症,但相較於 TZD 有肥胖的副作用,山苦瓜反而抑制腹部體脂堆積。並且山苦瓜 的降體脂作用可能與抑制脂質生合成有關。然而,本研究中使用的山苦瓜凍乾粉乙 酸乙酯萃物也許劑量不足,並沒有呈現如山苦瓜凍乾粉之效用。 關鍵詞:山苦瓜、Peroxisome proliferator - activated receptor (PPAR)、 Thiazolidinedione (TZD)、腹部脂肪、胰島素抗性. V.
(14) Wild Bitter Melon Ameliorate Abdominal Fat Accumulation and Insulin Resistance in High Fat High Sugar Fed Rats Ya - Wen Hong. Abstract In Asia countries, bitter melon (Momordica charantia; BM) is a commonly used vegetable. It had been reported that the bitter melon has hypoglycemic, hypolipidemia and anti - inflammatory effects. But its mechanism and functional component are still unknown. Recently, bitter melon was found to activate both peroxisome proliferator - activated receptor (PPAR) α and γ which may be valuable in prevention and amelioration of metabolic syndrome. The clinically used antidiabetic drug TZD (thiazolidinedione) is effective in ameliorating insulin resistance, but has side effect of obesity. In this study, the effect of wild BM lyophilized powder on anti - adiposity and insulin sensitivity in high fat diet fed rats was evaluated and compared with the antidiabetic drug TZD. We used forty Wistar male rats at the age of 6 weeks. Thirty-four rats were fed with high fat high sugar diet containing 30% butter to induce obesity, then divided into four groups: BM (a high fat diet containing 5% wild BM lyophilized powder), EAE (a high fat diet containing 0.25% EA extract from wild BM lyophilized powder), HF (a high fat diet serve as a high fat control), and TZD (a high fat diet containing 0.01% clinically used antidiabetic drug pioglitazone, serve as a positive control). For comparison, six rats fed with a low fat diet containing 5% soybean oil serve as LF group. After 9 weeks, rats were sacrificed and abdominal fat including retroperitoneal, epididymal, and injuinal fat were excised and weighted. Epididymal and retroperitoneal fat were also used for determination of adipocyte size, triglycerides and DNA contents. The activities of enzymes participate in lipogenesis and lipolysis, and the genes expression of adipocyte differentiation markers were also determined in epididymal and retroperitoneal fat pads. At wk 0, 3, 6, 9, fasting blood was collected from tail for glucose and insulin determination.. VI.
(15) Our results showed, the body weight gain of TZD group of rats was significantly higher than that of LF group of rats (p < 0.05), and this was in accordance with the well - known TZD side effect of obesity. HF, EAE and TZD group of rats also showed a significantly (p < 0.006) higher abdominal fat accumulation (retroperitoneal, epididymal and injuinal fat) than LF group, but there was no significant difference between BM and LF groups of rats. This result indicated that wild BM lyophilized powder was effective in inhibiting abdominal fat accumulation induced by high fat diet. The distribution of adipocytes size, isolated from retroperitoneal and epididymal fat, showed there were more smaller cells and less larger cells in BM group of rats when compared with those of HF group of rats. In addition, the triglycerides contents in adipose tissue of BM group of rats tended to be lowered. In epididymal fat, the activity of glycerol – 3 - phosphate dehydrogenase (G3PDH) which is an important lipogenic enzyme in adipose tissue, was comparable between BM and LF, and significantly lower than TZD group of rats (p < 0.05). Northern blot showed that ADD1 / SREBP1c mRNA, a transcription factor for regulating fatty acid synthesis, was significantly lower in BM group of rats than TZD group of rats (p < 0.05) in epididymal fat. Hyperinsulinemia induced by high fat diet was only observed in HF , but not in TZD and BM group of rats. These results indicate wild bitter melon, as well as TZD, could ameliorate insulin resistance induced by high fat diet. In contrast to the well - known obesity side effect of TZD, wild bitter melon is more beneficial in inhibiting abdominal fat accumulation. The anti - adiposity effect of wild bitter melon may be related to the inhibition of lipogenesis. However, the dose of EA extract from wild BM lyophilized powder used in this study (0.25%) may be too low to show its effect. Keywords: Wild bitter melon, Peroxisome proliferator - activated receptor (PPAR), Thiazolidinedione (TZD), Abdominal fat, Insulin resistance. VII.
(16) 第一章 前言. 第一章 前言. 代謝症候群(Metabolic syndrome)意指腹部肥胖、血脂紊亂、高血糖及高血壓, 已成為是世界性主要的公共健康問題,並且是第二型糖尿病與心血管疾病的危險因 子。雖然導致代謝症候群的原因還未清楚,但目前認為高胰島素血症與腹部肥胖是 主要原因。 苦瓜(Momordica charantia) 是常被用做藥物使用的植物之一。在亞洲,苦瓜主 要是栽培作為蔬菜食用;在印度,常作為治療糖尿病的傳統用藥。雖然許多研究指 出苦瓜有許多藥理特性,例如抗癌、降血糖及降三酸甘油酯等的作用,其中以降血 糖的作用最常被研究,但這些作用機制與功能性成分並不清楚。 近來,轉活化研究顯示苦瓜可活化 Peroxisome proliferator - activated receptor (PPAR) α 和 γ,PPAR 是細胞核荷爾蒙接受器之轉錄因子。已知 Fibrate 類降血脂 藥物與 Thiazolidinedione (TZD)類抗糖尿病藥物分別是 PPARα 和 PPARγ 的活化 劑,雖然 TZD 可改善胰島素敏感性,但卻具有肥胖的副作用。畢竟這些都作為藥 物治療使用,若是能由飲食來源中來找尋出可活化 PPAR 的天然食物,即可同時改 善高血脂與高血糖。這也暗示了苦瓜可能具有同時調節葡萄糖利用與脂質代謝的潛 力,可應用於預防或治療代謝症候群。 苦瓜降低血糖是已被確認的作用,近來對苦瓜的研究轉移集中在抗肥胖的潛在 作用。在 2003 年香港大學首度證實了苦瓜可抑制腹部體脂的堆積,並推測可能藉此 改善血糖,但苦瓜對脂肪細胞型態或生合成影響並未被研究。因此,本研究擬利用 餵食高油高糖飲食誘發肥胖大鼠探討山苦瓜對於改善腹部體脂堆積和胰島素敏感性 之功效,並且與已知抗糖尿病藥 TZD 比較。. 1.
(17) 第二章 文獻回顧. 第二章 文獻回顧. 一、過氧化體與過氧化體增殖劑活化接受器. (一) 過氧化體 Peroxisome. 過氧化體是一小的單層膜胞器,含有各種代謝反應如能量代謝的酵素,而過氧 化體至少含有 50 種不同酵素參與不同細胞型態的各種生物路徑(Biochemical pathways)。 1954 年 Rhodin 在腎細胞中發現 Microboies 此胞器(1),之後有學者利用細胞分 離與顯微鏡觀察對此新胞器做鑑定(2)。de Duve 將此胞器命名為過氧化體,即為含 有氫氧代謝酵素的胞器(1),此酵素含有分解 H2O2 (Catalase)與生成 H2O2 (H2O2 – generating oxidases)的酵素。過氧化體在脂肪酸 β - oxidation 中扮演重要角色(3), 當 過 氧 化 體 生 合 成 中 基 因 突 變 時 , 就 會 造 成 Zellweger disease 、 X - linked adrenoleukodystrophy (X - ALD)等基因突變疾病。這些患者因過氧化體中脂肪酸的 氧化作用受損,而血液和組織中大量堆積極長鏈脂肪酸(Very – long - chain fatty acids, VLCFA)(4, 5),因此可見過氧化體在人類生理角色上是必需的。. (二). Peroxisomal fatty acid β – oxidation 雖然脂肪酸能藉由不同機制發生氧化作用,但大部分的脂肪酸是經由β -. oxidation的方式而被降解。過氧化體與粒線體一樣都具有脂肪酸β - oxidation的機 制,包括了4個反應階段:Dehydrogenation、Hydration (of the double bond)、 Dehydrogenation again與Thiolytic cleavage (6)。透過此4步驟,從每個脂肪酸水解 下來的2 - carbon unit會形成Acety l - CoA unit,接著進入Citric acid (Krebs) cycle 生成二氧化碳和水。過氧化體與粒線體的β - oxidation system催化不同脂肪酸及其衍 生物的β – oxidation,粒線體催化衍生自飲食包括Palmitate、Oleate、Linoleate和 Linolenate的脂肪酸,而過氧化體是催化Hexacosanoic acid (C26:0)的極長鏈脂肪. 2.
(18) 第二章 文獻回顧. 酸 、 Pristanic acid (2, 6, 10, 14 - tetramethylpentadecanoic acid) 以 及 Di - 和 Trihydroxycholestanoic acid (DHCA和THCA)。 因為過氧化體缺乏呼吸鏈與Citric acid (Krebs) cycle,所以脂肪酸β - oxidation 的最終產物如Acetyl - CoA、Propionyl - CoA和Medium - chain acyl - CoA esters, 必需以Carnitine esters的形式運送到粒線體中。另外,在粒線體中是由Carnitine cycle攜帶脂肪酸橫跨粒線體細胞內膜,而在過氧化體中是藉由ATP - binding cassette transporter (ABC transporter) 催化脂肪酸橫跨細胞膜的運送。. (三) 過氧化體增殖劑 Peroxisome proliferators. 1965 年 Hess 等 學 者 首 次 發 現 到 大 鼠 肝 細 胞 中 過 氧 化 體 增 殖 (Peroxisome proliferation)的現象(7),隨後在1966年Svoboda與Azarnoff也觀察到此現象(8)。降 血脂藥物Clofibrate的主要作用在降低血漿中三酸甘油酯與膽固醇濃度(9),有研究 指出降血脂藥物Clofibrate會增加大鼠肝臟過氧化體中Fatty acyl - CoA oxidizing system (10)。 過氧化體增殖劑是一群包括有降血脂藥物、除草劑和Industrial plasticisers的化 學物質(11-13),當給予齧齒類動物這些物質會造成明顯肝臟的過氧化體增殖如同肝 細胞的增生(Liver hyperplasia)一樣(14)。此現象會增加長鏈脂肪酸Peroxisomal β oxidation以及Cytochrome P450 IV family基因的轉錄作用(9, 12, 15)。. (四). 過氧化體增殖劑活化接受器. Peroxisome proliferator - activated receptor, PPAR 1990年,Isseman和Green從小鼠肝臟cDNA library篩選出與其它細胞核接受器 的DNA binding domain中,具高度保留序列相似的新類固醇荷爾蒙接受器,並以其 過氧化體增殖的特性命名之(16)。之後其它的PPAR在不同物種與組織中被鑑定出 (17, 18),這些PPAR在序列上具同源性(Homology)以及組織分佈差異。在過去十年, PPAR的研究揭開了對脂質代謝調控的新機制,以及對包括第二型糖尿病和肥胖這 些代謝性疾病的可能分子因子。. 3.
(19) 第二章 文獻回顧. 1.. PPAR的種類. PPAR是屬於細胞核接受器的Superfamily,這些細胞核接受器包括有Retinoic acid receptors (RARs)、Thyroid hormone receptors (TRs)以及Steroid receptors (19) 。 到 目 前 為 止 以 不 同 基 因 轉 殖 並 鑑 定 出 三 種 PPAR 種 類 , 分 別 為 PPARα (NR1C1)、PPAR - δ / β (NR1C2)和PPARγ (NR1C3)。已知天然的脂肪酸以及Fibrate 類降血脂藥物是PPAR的活化劑(Activators),PPAR調控許多對脂質與葡萄糖代謝相 關的基因表現(20)。在一些研究中發現PPARγ agonists在脂肪細胞分化上扮演重要決 定角色,以及是抗糖尿病藥物TZD的作用位置(21, 22)。. (1) PPARα 已知Fibrate類降脂血藥是PPARα的活化劑,PPARα在齧齒類動物和人類的許 多組織中表現,包括有肝臟、腎臟、心臟、骨骼肌和棕色脂肪(18, 23),而且也有研 究 指 出 PPARα 在 血 管 細 胞 如 內 皮 細 胞 (24) 、 VSMCs (vascular smooth muscle cells)(25, 26)和單核細胞 / 巨噬細胞(27)也有表現。. (2) PPAR – δ / β 目前對PPAR – δ / β的功能鑑定仍然不明確,已知PPAR – δ / β廣泛表現在許多組 織和細胞中,而在腦、脂肪組織和皮膚是高度表現(18, 28)。. (3) PPARγ 目前有七種PPARγ mRNA已被鑑定出(29, 30),它們都是由不同的Initiation promoter和5 - terminal region (A1、A2、B、C和D)的五個Exons所剪輯,而產生相 同的PPARγ基因。PPARγ2在N - terminus比PPARγ1多了30個氨基酸。PPARγ1廣泛 表現在許多組織中如結腸、肝臟和血管細胞,但PPARγ2專一表現在脂肪組織中 (31),PPARγ3大量表現在巨噬細胞、大腸和脂肪組織(32),PPARγ4和PPARγ5只表. 4.
(20) 第二章 文獻回顧. 現在巨噬細胞中而PPARγ6和PPARγ7則是在脂肪組織中可偵測得到(33)。 有研究指出Prostaglandin (PG) - related compounds如15 – deoxy - delta12 – 14 - PGJ2 (15d - PGJ2) 是 PPARγ 有 效 的 活 化 劑 (34) 。 另 外 在 Primary human trophoblasts的研究指出,氧化脂質如9 - HODE、13 - HODE和15 - HETE也是 PPARγ有效的活化劑(35)。已知Thiazolidinedione (TZD)類藥物如Troglitazone、 Ciglitazone、Pioglitazone和Englitazone對PPARγ是專一性的活化劑(36)。 近來也鑑定出一些新的PPARγ agonists和antagonists,Triterpenoid 2 – cyano – 3、12 – dioxooleana - 1, 9 – dien – 28 - oic acid (CDDO)是PPARγ agonists (37, 38), 而Bisphenol diglycidyl ether (BADGE)和LG - 100641是PPARγ antagonists (39, 40)。雖然這些化合物具較少的臨床意義,但可能對於了解PPARγ的生理作用以及新 活化劑的鑑定是有幫助的。除了利用化學合成方式外,在天然產物的研究中也發現 一 些 有 效 的 PPARγ agonists , 例 如 一 些 藥 用 植 物 中 , 從 Saururus chinensis (Saururaceae)來的Saurufuran A,一些黃酮類像是Chrysin、Apigenin和Kampferol 以及Glycyrrhiza uralensis (Fabaceae)中的酚類化合物,都是近來被發現新的PPARγ agonists (41-43)。 2.. PPAR之結構特徵. 三 種 PPAR isoforms 在 結 構 和 功 能 上 有 相 似 的 特 性 , 是 由 四 個 Functional domains中的6個Structural regions所組成,包括有A / B、C、D和E / F。N端的A / B domains含有PPAR磷酸化反應的Ligand - independent activation function 1 (AF 1)(44)。DNA binding domain (DBD)或C domain是促進PPAR與目標基因上Promoter region的Peroxisome proliferator responsive element (PPRE)結合(45)。D site是 Cofactors的Docking domain。E domain或Ligand binding domain (LBD)對Ligand 是專一性反應,並活化PPAR結合到PPRE上增加目標基因的表現(28)。位在E / F domain 的 Ligand - independent activation function 2 (AF - 2) 可 吸 引 PPAR cofactors,來促進基因轉錄的活性(46)。. 5.
(21) 第二章 文獻回顧. 圖2 - 1. PPAR之結構特徵. 資料來源:Guo and Tabrizchi., 2005 (47). 3.. PPAR之基因轉錄作用機制. 除了天然與合成的Ligands可活化PPAR外,其它像是Retionoid X receptor (RXR)、PPRE和Coactivators也在轉錄作用上扮演重要角色。. (1) Coactivators / Corepressor (Cofactors) 許多做為Coactivators或Corepressor的蛋白質可調節細胞核接受器的起始或抑 制轉錄作用的能力,這些蛋白質是以Ligand - dependent的方式與細胞核接受器來作 用(17)。在沒有Ligand的情況下,Heterodimer形式的細胞核接受器與含有Histone deacetylase activity的Multicomponent co - repressors,像是Nuclear receptor co – repressor (NCoR) 和 Silencing mediator for retinoid and thyroid hormone receptor(SMRT)相連結(48, 49),在Deacetylated下的Histone會抑制轉錄作用(50)。 相反的,像是Steroid receptor co - activator (SRC) - 1和PPAR binding protein (PBP) 這些具有Histone acetylase activity的Coactivators,會啟動並誘導Ligand與基因轉錄 序列的結合(51, 52)。. 6.
(22) 第二章 文獻回顧. (2) PPRE與PPAR目標基因. PPRE序列是由2組Hexanucleotides以單一核酸間隔的AGGTCA sequence所組 成的Direct repeat (DR) - 1 elements (17, 53)。不同於其它細胞核接受器的Responsive element DR - 3和DR - 4,此DR - 1 elements具PPAR - RXR heterodimer專一性。當 有Ligand結合在LBD時,會促進PPAR與RXR形成Heterodimer,並與Coactivators 結合到PPRE上,啟動基因轉錄作用。在一些調節脂質代謝相關基因中如Fatty acid transporter (FAT)、Adipocyte lipid binding protein (ALBP)、Acyl - CoA synthetase (ACS)和Acetyl - CoA oxidase (ACO)都含有PPRE序列(54)。. 圖2 - 2. PPAR之基因轉錄作用機制. 資料來源:Kota et al., 2005 (55). 7.
(23) 第二章 文獻回顧. 4.. PPAR生理角色. (1) PPARα PPARα在脂質代謝、發炎反應與動脈粥狀上都具有作用,已知PPARα agonists 對脂肪酸β - oxidation的調節扮演重要的角色(56)。此外,PPARα agonists藉由增加 Fatty acid transport protein (FATP)和Fatty acid transporter (FAT)的表現,刺激細 胞對脂肪酸的汲取(57)。PPARα的Ligands如Fibrates和其它Peroxisome proliferator agents會促進Cytochrome P4504A (CYP4A)的表現,CYP4A subclass的Cytochrome P450 enzyme催化脂肪酸的ω-hydroxylation (58),這些機制有益於降低三酸甘油酯的 生合成。另外,PPARα的活化會增加Lipoprotein lipase (LPL)的表現(59)和抑制肝 臟中的Apolipoprotein (apo) C – III (22),而降低三酸甘油酯濃度。 PPARα藉由調節對脂肪酸汲取和氧化反應的基因來供給能量給心肌(60),在心 臟PPARα會降低脂肪酸氧化作用和抑制LPL(61)。另外,PPARα agonists已被指出 可活化ApoA - I的表現(62),ApoA - I是高密度脂蛋白(High density lipoprotein, HDL) 的主要Apolipoprotein,在周邊細胞的Reverse cholesterol transport (RCT)扮演重要 的角色。PPARα的活化也影響巨噬細胞其 Cholesterol efflux “pump”中的 ATP binding cassette A - 1 (ABCA1)之表現(63),這是ApoA - I調節RCT路徑的重要角 色。一些研究顯示PPARα的活化會增強Liver X recepror (LXRα)來促進ABCA1之表 現(64)。因此,PPARα和LXRα agonists可能具有預防或減輕動脈粥狀作用。 PPARα agonists在血管細胞中有抗發炎作用,這些作用包括抑制細胞激素誘導 Vascular cell adhesion molecular – 1 (VACAM – 1)的表現(65),以及增加Endothelial nitric oxide synthase (eNOS)的表現(66)。在內皮細胞中,PPARα agonists降低血漿 中Interleukin – 6 (IL – 6)濃度以及藉由Nuclear factor - κB (NF - κB) signaling repression降低VSMCs中Cyclooxygenase - 2的表現(26)。PPARα也誘發Apoptosis(27) 和向下調節細胞激素活化的巨噬細胞中其Tissue factor (TF)和Metalloproteinase (MMP)的表現(67, 68)。. 8.
(24) 第二章 文獻回顧. (2) PPAR – δ / β PPARβ agonists已發現對高血脂症、動脈粥狀硬化、肥胖、Cholesterol efflux 和肌肉中能量消耗是有益的(69, 70)。Selective PPARβ agonists會活化PPARβ,顯著 降低第二型糖尿病動物模式中三酸甘油酯和葡萄糖濃度(71);PPARβ活化棕色脂肪 組織中的Heat - producing uncoupling enzymes (Uncoupling protein, UCP1和3)以及 肌肉中的UCP2,此作用可能有益於肥胖的控制(72)。 近來有研究指出PPARβ與癌症的關係,在Gastrointestinal neoplasias的形成 中,PPARβ做為β – catenin和Sulindac (NSAID) - sensitive factor直接轉錄目標(73), 表示PPARβ與小腸和結腸腫瘤形成有關(74)。相反的,在缺乏PPARβ小鼠的研究中 指出,PPARβ抑制直腸致癌物的生成(75),因此PPARβ對直腸癌的作用仍是需要進 一步的研究。 PPARβ在神經系統上也具有角色,先前研究表示PPARβ大量表現在中樞神經系 統 , 特 別 在 前 額 皮 質 、 神 經 節 以 及 丘 腦 核 仁 這 些 地 方 中 具 高 度 表 現 (76) 。 Bromopalmitate和L - 165041這些PPARβ agonists被發現可增加寡狀神經膠質細胞 的分化,顯示出PPARβ參與軸突的Myelination,也指出了PPARβ可能在腦中脂質 代謝上扮演重要角色(77, 78)。近來,Smith等人培養大鼠小腦粒狀神經細胞的研究 指出,PPARβ有神經保護特性,顯示了PPARβ在神經退化過程中具潛在作用(79)。. (3) PPARγ PPARγ對發炎反應與動脈粥狀有密切關係,PPARγ在內皮細胞、血管平滑肌細 胞以及巨噬細胞中的表現,對脂質代謝的影響是值得注意的問題,因而促進對 PPARγ其抗發炎特性的研究。另外,PPARγ在慢性發炎疾病如動脈粥狀硬化、關節 炎和Inflammatory bowel syndrome也被研究,PPARγ agonists在抗動脈粥狀硬化上 也扮演重要角色的,這些Agonists會促進單核球上Scavenger receptor CD36之表 現,增加併吞氧化型LDL而增加Foam cells的形成(80)。在內皮細胞中,TZD抑制黏 附因子VCAM - 1和Intercellular adhesion molecule (ICAM - 1)的表現,使動脈內膜 的單核細胞堆積減少(81)。在人類單核細胞衍生的巨噬細胞中,PPARγ agonists抑制. 9.
(25) 第二章 文獻回顧. MMP - 9 gelatinolytic activity,而MMP - 9為具有斑塊脫落(Plaque rupture)的酵素 (82)。VSMCs在動脈粥狀硬化的形成中是最重要的角色,而近來研究顯示其為 PPARγ agonists的主要作用目標,TZD藉由降低Retinoblastoma protein的磷酸化以 及增加Cyclin dependent inhibitor p27的表現,而抑制VSMCs的增生(83)。 PPARγ agonists 在 巨 噬 細 胞 脂 質 恆 定 中 會 調 節 一 些 基 因 的 表 現 如 Apolipoprotein E (apoE)(84),而PPARγ agonists也會調節巨噬細胞的細胞激素生 成,但其作用仍然不清楚。較早之前研究指出TZD或15d - PGJ2會活化PPARγ,使 培養的巨噬細胞大量表現各種前發炎細胞激素(Pro - inflammatory cytokines)如 Tumor necrosis factor α (TNFα)或IL – 6 (85),但這些作用在LPS - challenged mice 中並未發現(86)。此外,也有研究曾指出巨噬細胞的分化是不需要PPARγ,顯示出 PPARγ沒有專一性抑制巨噬細胞生成細胞激素(87)。一些其它研究對此指出的機制 1 PPARγ抑制巨噬細胞中ApoB - 48 receptor,○ 2 PPARγ增加巨噬細胞的 包括有○ 3 PPARγ藉由降低Activator Protein 1 (AP - 1)、Signal Transducer and Apoptosis,○. Activator Transcription (STAT)和Nuclear Factor κB (NF - κB),來抑制Nitric oxide synthase、Gelatinase B和Scavenger receptor A的基因表現(88-90)。綜合而言,PPARγ agonists在控制發炎反應與動脈粥狀硬化上具有益處。 另外也有報告指出PPARγ與癌症的關係,在一些In vivo和In vitro的動物模式中 發現,PPARγ促進Apoptosis和細胞分化作用的特性對不同癌症如乳癌、結腸癌、前 列腺癌、胰臟癌和胃癌的治療是有益的(91)。另外還有PPARγ影響Retinal disorders 的文獻指出,在Retinal pigment epithelial (RPE) cells和Bovine choroidal endothelial cells (CECs)中,Troglitazone抑制血管內皮生長因子(Vascular endothelial growth factor, VEGF)誘導的Choroidal angiogenesis,TZD也抑制大鼠和猴子中Choroidal neovascularization的形成 (90)。PPARγ 調節 TZD對 大 鼠 的 Retinal ganglion cells (RGC - 5)之抗氧化特性(92)。這些作用顯示PPARγ ligands對Age - related和Diabetes - induced retinal disorders具潛在的作用。. 10.
(26) 第二章 文獻回顧. 二、脂肪細胞生合成 Adipogenesis. 脂肪組織(Adipose tissue)的形成與身體能量恆定調節相關,當能量攝取超過能 量消耗時,會使脂肪組織重量增加最後造成肥胖(Obesity)。在細胞層次上,正的能 量平衡會刺激成熟脂肪細胞(Adipocytes),經由增加三酸甘油酯的儲存來調節過多的 能量,當脂肪細胞達到一個臨界大小閾值時,會傳送訊息給前脂肪細胞 (Preadipocytes)去刺激新的脂肪細胞產生,來儲存過多的能量。在幼年期,主要是 經由新的脂肪細胞形成或脂肪細胞增生(Hyperplasia)使脂肪組織擴大,人與囓齒類 動物在生命期中具有從前脂肪細胞形成新脂肪細胞的能力,與瘦的動物相比較下, 肥胖的動物有較多且較大的脂肪細胞。脂肪組織重量在肥胖和 Lipodystrophy 中扮 演重要角色,而肥胖又與第二型糖尿病和胰島素抗性息息相關。因此,對於了解脂 肪細胞生合成的正向與負向調節是必要的。. (一) 脂肪細胞生合成的 In vitro 模式. 在研究脂肪細胞生合成的 In vitro 模式中,最常用的是分離自 Swiss 3T3 cells 的 3T3 - L1 和 3T3 - F442A 齧齒類前脂肪細胞株(Preadipocyte cell lines)(93, 94)。另 外,衍生自 ob / ob mice 副睪脂肪的 ob17 cell line 也是常被使用到的(95)。在 1970 年間,Green 與其團隊發展出 3T3 - L1 system 來研究脂肪細胞生合成(94),這些鼠 科細胞的形態與增殖類似纖維母細胞(Fibroblasts)。研究顯示,細胞在長滿之前會在 擴 大 生 長 的 階 段 , 當 長 滿 時 接 著 給 予 Proadipogenic agents 如 胰 島 素 、 Dexamethasone 和 Phosphodiesterase inhibitor,就會停止生長然後進入分化階段。 在分化非常早期時,因為細胞外基質和 Cytoskeleton proteins 使細胞從纖維狀變成 圓 形 , 未 成 熟 的 脂 肪 細 胞 需 要 有 絲 分 裂 信 號 (Mitogenic signals) 的 Clonal expansion,此作用可增加最後已分化的脂肪細胞比例,之後接著進入最後成熟階段 變成脂肪細胞。此分化作用的 In vitro 模式與 In vivo 的過程極為相似,因為培養的 細胞會堆積脂質,表現出成熟脂肪細胞的特徵並且具有胰島素敏感性。相反的,因 為 Catecholamine - sensitive lipolytic pathway 的調節,在能量喪失時脂肪細胞也會. 11.
(27) 第二章 文獻回顧. 水解三酸甘油酯,最後成熟的脂肪細胞具有生合成並分泌許多 Factors 的能力,其 中這些 Factors 與 Endocrine control 的能量恆定有關。 在 In vivo 研究指出,注射 3T3 - L1 或 Ob17 preadipocyte cell lines 到 Nude mice 體內中,形成的脂肪組織與正常小鼠體內的脂肪組織是無法區分的(96),此結果表 示 In vitro – differentiated adipocytes 與 In vivo adipocytes 具有相似的機制來發生脂 肪細胞生合成。近來也發展出 Human preadipocyte cell lines,例如有分別衍生自脂 肪肉瘤以及 Simpson – Golabi - Behmel syndrome 嬰孩的 Human preadipocyte cell strains (97, 98),還有新生兒肩胛骨棕色脂肪的 SV40 T - Ag transformation 來的 Brown preadipocyte cell line PAZ1 (99)。將重組的 Telomerase activity 結合 Human papillomavirus - E7 表現,可使 Human preadipocyte 在 Immortalization 之後保留 其分化能力(100)。 更近來,也發展出分離自年幼捐贈者脂肪組織的Multipotent adipose - derived stem (MADS) cells,這些細胞可以維持Ex vivo的特性,表示在體內有Self - renewal 的能力,因此可表現出正常的二倍體染色體(Diploid karyotype),且保持有分化為許 多Mesenchymal cell types作用的能力(101, 102)。 In vivo 中,脂肪組織分布在許多部位,包括在生殖器周邊的、腹膜後的以及皮 下的脂肪,分離自不同部位的前脂肪細胞有不同的脂質生合成潛力,這個問題至今 仍是未知的。除此之外,不同部位的成熟脂肪細胞其代謝反應也不同,例如內臟脂 肪與皮下脂肪對腎上腺素所刺激的脂解反應就有所差異。因此,建立前脂肪細胞的 細胞株來研究脂肪細胞生合成是必要的。. (二) 脂肪組織來自 Multipotent stem cells. 進來研究指出脂肪組織來自是Multipotent stem cells,根據接收不同的信號 Multipotent stem cells可分化為不同的細胞類型。在In vitro中Mouse embryonic stem (ES) cells的分化是脂肪細胞的來源之一,可用來了解早期脂肪細胞生合成以及 對調節基因和環境因子的鑑定(103)。Multipotent stem cells也存在於成熟的組織 中 , 最 早 是 從 骨 髓 中 被 分 離 出 來 (104) 。 在 現 在 , 脂 肪 組 織 也 可 分 離 出 Stem. 12.
(28) 第二章 文獻回顧. cells(105)。從人脂肪組織來的Multipotent stem cells可被誘導分化成為其它細胞類 型,包括有軟骨(Cartilage)、骨頭和肌肉(106, 107)。hMADS cells也可形成脂肪細胞, 將hMADS cells培養在無血清促脂肪細胞生成的培養基下,會有脂質油滴堆積以及 脂肪細胞分化指標和轉錄因子的表現(101),特別的是會像人的脂肪細胞一樣有 Lipolytic pathway, 而 能 分 泌 出 Leptin和 Adiponectin。人脂肪組織中的 Stroma vascular fraction可衍生出具功能的Cardiomyocyte - like cells、Endotheliallike cells 和Insulin - secreting cells (108-110)。脂肪組織衍生的細胞也能參與骨骼肌的再生以 及促進新血管形成(111, 112),這些顯示出衍生自脂肪組織的Multipotent stem cells 可能對疾病的治療是重要的。. (三) 脂肪細胞生合成的過程. 前脂肪細胞在轉變成脂肪細胞前具維持生長的能力,在脂肪細胞分化期間,脂 肪細胞的表型(Phenotype)是許多基因的表現,此反映在早期、中期和晚期 mRNA / protein markers 的表現以及三酸甘油酯堆積(113, 114)。這些變化主要是發生在轉錄 層次(Transcriptional level)上,除了基因的活化之外,這些抑制脂肪細胞生合成的或 對脂肪細胞功能不需要的基因會被抑制。. 1. Growth arrest 前脂肪細胞株和初代前脂肪細胞一樣,當長滿時就會停止生長。已知的兩個轉 錄因子,CCAAT / enhancer binding protein - α (C / EBPα)和 PPARγ 是有轉錄活性 的脂肪細胞專一性基因(Transactivate adipocyte specific genes) ,C / EBPα 和 PPARγ 似乎都與停止生長有關且對於脂肪細胞分化是需要的。McKnight 和其團隊 利用 C / EBPα - estrogen receptor fusion protein 使 C / EBPα 具抗有絲分裂活性 (Antimitotic activity),在給予 Estrogen 的活化作用下,觀察細胞數目和 DNA 合成 的結果顯示出有細胞會停止生長(115)。Darlington 等人的研究中發現 C / EBPα 會 增加 p21 / SDI - 1 mRNA 和蛋白質的表現,因為 Antisense p21 / SDI - 1 可削減 C / EBPα 的生長抑制作用,這表示 C / EBPα 具有增加 p21 / SDI - 1 表現的功能。. 13.
(29) 第二章 文獻回顧. Spiegelman 和其團隊指出 PPARγ 可誘發纖維母細胞和 Adipogenic simian virus 40 large T antigen – transformed cells 停止生長,在這些表現有 PPARγ 的細胞中,因 為 DP – 1 的磷酸化和 Serine / threonine phosphatase PP2A catalytic subunit 表現減 少而使細胞週期退去,並伴隨 E2F / DP - 1 complex 的轉錄活性和 DNA 結合活性的 減少(116)。因此,C / EBPα 和 PPARγ 可能共同作用來使生長停止,雖然 C / EBPα 和 PPARγ 在脂肪細胞分化期間顯著地增加表現,但在前脂肪細胞中的低表現能夠 在分化之前調節使細胞停止生長。. 2. Clonal expansion 當長滿時而停止生長之後,前脂肪細胞必須受到適當的 Mitigenic signals 和 Adipogenic signals 的組合來繼續之後的分化步驟。在前脂肪細胞株的研究顯示,停 止生長的細胞會進行細胞分裂發生 DNA 複製和細胞增殖,來使細胞複製放大。對 3T3 - F442A 和 Ob17 細胞而言,細胞分裂增加與晚期 mRNA markers 的表現有關, 而抑制細胞分裂會防止脂肪細胞的形成,但是 Primary human preadipocytes 即使 培養在含有絲分裂抑制劑條件下,還是能正常發生分化作用(117)。這表示了對培養 的 Cell line system 而言,需要依賴外來的刺激使長滿後的細胞進行有絲分裂,但對 分離自脂肪塊的前脂肪細胞,其在 In vivo 就已發生了與之後脂肪細胞發展階段相關 的細胞分裂。. 3. Early changes in gene expression Lipoprotein lipase (LPL) mRNA 的表現通常作為脂肪細胞分化的早期指標, LPL 是由成熟的脂肪細胞所分泌且在控制脂質堆積上扮演重要的角色(118)。C / EBPs 和 PPARγ 這兩個轉錄因子在脂肪細胞分化早期就被誘導,C / EBPs 和 PPARγ 的早期表現與之後參與最後分化的 Adipocyte specific genes 有關(119)。PPARγ 的表 現在荷爾蒙誘導分化作用之後會快速地增加,在 3T3 - L1 脂肪細胞分化的第 2 天期 間就能測得 PPARγ 而且在成熟脂肪細胞中達到最大的表現。PPARδ 的誘導似乎在 PPARγ 之前,但是 PPARδ 的表現是廣泛分佈的,在各種組織中和一些培養的細胞. 14.
(30) 第二章 文獻回顧. 株中包括 CH310T1 / 2、3T3 - C2 和 NIH 3T3 都能測得。C / EBPβ 和 C / EBPδ 短 暫的增加是在 PPARγ 表現增加之前,C / EBPβ 和 C / EBPδ 在早期和中期分化階段 之後會減少並伴隨誘導 C / EBPα mRNA 的表現,在 Adipocyte specific genes 表現 之前 C / EBPα 會輕微地增加表現(120)。 在脂肪細胞分化期間,細胞從纖維狀變成圓形並且在形態上、Cytoskeletal components 和 Extracellular matrix (ECM) components 的濃度和型式有明顯的改 變。許多 Cytoskeletal 和 ECM components 對脂肪細胞分化影響的研究指出脂肪細 胞轉錄因子的特徵,這些變化可能是影響脂肪細胞分化期間 PPAR 或 C / EBPs 的表 現和作用。. 4. Late events and terminal differentiation 在分化的最後階段,培養中的脂肪細胞主要增加 De novo lipogenesis (脂質生合 成)和對胰島素的敏感性。一些參與三酸甘油酯代謝的酵素包括有 ATP citrate lyase、Malic enzyme、Acetyl - CoA carboxylase (ACC)、Stearoyl - CoA desaturase (SCD1)、Glycerol – 3 - phosphate acyltransferase、Glycerol – 3 - phosphate dehydrogenase (G3PDH) 、 Fatty acid synthase (FAS) 和 Glyceraldehyde – 3 phosphate dehydrogenase,它們的活性、蛋白質和 mRNA 表現增加了 10 ~ 100 倍 (121),Glucose transporters、Insulin receptor 的數量和胰島素敏感性也都增加。在 脂肪細胞分化期間,β1 - adrenergic receptors 的減少以及 β2 - 和 β3 - subtypes 的 增加,增加了 Total adrenergic receptor 的數量。除了與脂質代謝相關蛋白質的 mRNA 增加外,脂肪細胞也合成其它脂肪組織專一性產物,包括 Adipocyte specific fatty acid binding protein (aP2),FAT / CD36 這個 Fatty acid transporter 以及 Perilipin 這種 Lipid droplet - associated protein。除此,脂肪細胞還生成一些 Secreted products 包括有作為 Angiogenic agent 的 Monobutyrin、屬於 Serine protease complement factor D 的 Adipsin、Acrp30 / AdipoQ、PAI - 1 和 Angiotensiongen II (122)。PPARγ 和 C / EBPα 共同活化一些基因的表現包括了 aP2、GLUT4、SCD1、 Phosphenolpyruvate carboxykinase (PEPCK)和 Leptin (123)。. 15.
(31) 第二章 文獻回顧. 圖2 - 3. 脂肪細胞生合成的過程. 資料來源:Feve., 2005 (120). (四) 脂肪細胞生合成的轉錄調控. 已知有許多促進或抑制脂肪細胞生合成的轉錄因子會影響脂肪細胞生合成,一 些 In vitro 和 In vivo 的 研 究 顯 示 出 , 脂 肪 細 胞 生 合 成 的 調 控 是 一 連 串 的 Transcription cascade。在脂肪細胞發展過程中,有三個主要直接影響脂肪細胞生合 成的轉錄因子,分別是 PPARγ、C / EBPs 和 The basic helix – loop - helix family (ADD1 / SREBP1c)。. 16.
(32) 第二章 文獻回顧. 1.. 轉錄因子. (1) PPARγ 1 Role of PPARγ in adipocyte differentiation ○. 先前已知TZD是PPARγ的Ligands,而Kletzien等人發現TZD可以啟動脂肪細胞 生合成(124)。PPARγ最早是在哺乳動物中被發現(125),但Spiegelman發現脂肪細胞 專一的PPARγ並在其團隊的研究也顯示出PPARγ主要在脂肪細胞生合成期間被誘 導,而且大量表現在脂肪組織中(126, 127),因此Lehmann等人鑑定出TZD是PPARγ 高親和力的 Ligand (21)。 Tontonoz等人將纖維母細胞利用 Retrovirus異位表現 PPARγ的Gain – of – function研究最先指出,PPARγ在脂肪細胞生合成中的重要角 色(128)。除此,在脂肪細胞生合成的過程中需要有PPARγ的存在,當C / EBPα在沒 有PPARγ的存在下是無法誘發脂肪細胞的分化,但PPARγ在沒有C / EBPα下能誘導 脂肪細胞分化(129),此表示PPARγ對脂肪細胞分化是主要的調節者。 在Adipose cell - selective fatty acid binding protein aP2的模式中,其5’ flanking region被鑑定是轉錄的起始,此Enchancer可開啟培養的與基因轉殖小鼠的脂肪細胞 中Promoter的基因表現(130)。Tontonoz等人鑑定出此Region是ARF6與ARE6和 ARE7結合的Cis - acting elements,並指出PPARγ與RXR這個Heterodimer可與aP2 基因的5’ flanking region結合(126, 131)。PPARγ除了誘導aP2基因的表現外,也會活 化許多其它Adipocyte specific genes的Promoter,在脂肪細胞中PEPCK的表現需要 有PPARγ的結合(132)。在培養的脂肪細胞與基因轉殖小鼠In vivo的研究也發現,一 些脂肪細胞中其Promoter的活化是需要PPARγ,但在其它組織中並不需要(133)。 許多Loss – of - function研究也指出PPARγ對脂肪細胞生合成的重要性。最早在 1999年,完全剔除胚胎PPARγ的小鼠在胚胎期10 ~ 10.5天期間,因為不正常的胎盤 形成而胎死腹中(134, 135)。由於囓齒類動物的脂肪細胞生合成大部份是發生在出生 之後,為了解決PPARγ. -/-. embryo無法存活的問題,Rosen等人利用Wild - type ES. cells與完全剔除PPARγ的ES cells製造出Chimeric mice,這些小鼠可存活下來並有正 常的組織發育(136)。因為ES cell genotype可干擾PPARγ對形成其它特異細胞類型或. 17.
(33) 第二章 文獻回顧. 器官發展的需要,因此得知PPARγ對於早期胎盤的形成是需要的。Rosen等人也觀 察了Wild - type和PPARγ - / - ES cell的In vitro分化發現到,PPARγ + / + cells可發生脂 肪細胞生合成,而PPARγ - / - cells不會堆積脂質、表現分化指標或有胰島素敏感性。 另 外 Barak 等 人 利 用 正 常 的 Tetraploid cells 來 校 正 胎 盤 的 缺 失 來 製 造 Chimeric embryos,這些小鼠在出生後可短暫存活下來,且在解剖後發現沒有棕色或白色脂 肪組織(137),Heterozygous PPARγ - deficient mice與高脂飲食的Wild - type mice相 較下,其副睪脂肪、腹膜後脂肪和周邊脂肪塊重量是減少的,且有增加的胰島素敏 感性。 Cre - loxP策略是用來產生專一性剔除組織PPARγ的表現,這些小鼠都有相似的 特徵,像是全身脂肪塊重量減少的脂肪萎縮(Lipodystrophy)以及胰島素抗性。目標 專一性降低脂肪組織中PPARγ活性的Knockout小鼠,皮下和副睪脂肪塊會逐漸減 少,血漿游離脂肪酸與三酸甘油酯增加,且血漿Leptin和Adiponectin減少(138)。選 擇性破壞PPARγ2會使腹膜後脂肪、腹股溝脂肪和生殖器周邊脂肪減少(139)。利用 Neomycin - resistant gene cassette對序列轉錄干擾,來降低脂肪組織中PPARγ2和 PPARγ1表現的PPARγ Hypomorphic mice,也發現有脂肪萎縮的現象(140)。目標專 一性剔除肌肉組織中PPARγ的小鼠,會慢慢形成胰島素抗性且會增加脂肪組織重量 (141),上述這些小鼠模式清楚的指出PPARγ對脂肪細胞生合成的重要性。. 2 Adipose as the target tissue for insulin sensitizing effects of PPARγ ligands ○. 一些囓齒類與人類的基因研究指出PPARγ的活化具有胰島素敏感性作用。在 PPARγ基因上有Pro12Ala基因多型性的人有較好的胰島素敏感性(142),在一些少數 病人身上發現,Heterozygous mutation的PPARγ會造成嚴重的胰島素抗性(143)。此 外,抑制小鼠PPARγ的Serine122磷酸化可增加PPARγ活性,使其不會發生因肥胖所 致的胰島素抗性(144),出乎意料地發現缺少其中一個PPARγ對偶基因的小鼠,比正 常小鼠有較好的胰島素敏感性(145),但這個原因尚不清楚,有可能是削減了基因抑 制作用,因而降低在缺乏Ligands時PPARγ的表現。不管如何,PPARγ最佳活性的 表現決定於其作為胰島素敏感性轉錄調節者的作用(146)。. 18.
(34) 第二章 文獻回顧. TZD 是 PPARγ專一性的 Ligands,而脂肪組織是一內分泌器官且大量表現 PPARγ,在低血糖且嚴重脂肪萎縮的小鼠中,TZD是沒有作用的(147),這顯示出脂 肪組織是TZD作用的主要位置。TZD可改善肌肉和肝臟中的胰島素敏感性(148),雖 然PPARγ在骨骼肌中表現很低,但專一剔除肌肉中PPARγ的小鼠有肝的胰島素抗性 現象,然而,在給予TZD之後能有效反轉其胰島素抗性(141)。相反的,另一個專一 剔除肌肉中PPARγ模式的小鼠有肌肉的胰島素抗性現象,但給予TZD並未改善其肝 臟的作用(149)。雖然其它組織可能也具角色,但目前研究顯示脂肪組織是TZD作用 的主要位置,藉由改變Adipose factors的分泌,在肌肉和肝臟中有間接的胰島素敏 感性作用。. 3 PPARγ regulation of adipocyte genes that modulate insulin sensitivity ○. 一些脂肪細胞的內分泌產物會改變周邊組織特別是肌肉和脂肪的胰島素敏感 性,已知PPARγ ligands TZD影響胰島素敏感性作用中的多種脂肪細胞基因。脂肪 細胞會分泌一些多胜肽,這些與葡萄糖恆定和胰島素敏感性有關,包括有TNFα、IL - 6、Leptin、Adiponectin和Resistin,這些基因表現會受到TZD的調節(150)。另一 個TZD主要的作用是降低循環的游離脂肪酸,而游離脂肪酸顯示與肥胖所致的胰島 素抗性有關(151)。TZD同時向上調節脂肪細胞中游離脂肪酸汲取相關基因如LPL和 FATP (152) , 以 及 游 離 脂 肪 酸 再 回 收 相 關 基 因 如 PEPCK 和 Glycerol kinase (GyK)(132)的表現。因此,TZD藉由對增加脂肪細胞汲取游離脂肪酸以及降低釋出 到循環中,因而降低血清游離脂肪酸濃度。. (2) CCAAT / enhancer binding protein (C / EBP) family C / EBPs是屬於Basic - leucine zipper的轉錄因子,已知有六種Isoforms,每一 種都有高度保留的bZIP domain以Homo - and / or heterodimers的類型作用(119),其 Basic DNA - binding domain與Regulatory element結合在下游基因的Promoters或 Enhancers上。C / EBPs gene的表現會調節三種蛋白質(α、β、δ),它們的組織分佈 並不侷限於脂肪組織中,大部分表現在脂肪與肝臟。它們有相似的DNA - binding. 19.
(35) 第二章 文獻回顧. activities,而且在它們的Promoters上有C / EBP regulatory elements的Trans – activating reporter gene。C / EBP protein在顆粒白血球(Granulocytes)和肝細胞的最 後分化作用上也扮演角色,另外C / EBPs對感染反應的抗性也有重要角色(114)。C / EBPs可調節許多層次包括了細胞中mRNA表現的轉錄作用,In vitro中cAMP已知是 脂肪細胞生合成的Inducer,能夠同時增強C / EBPα和C / EBPβ的表現。 在脂肪細胞生合成期間,一些C / EBP family membranes的表現是受到調節, 近來在Gain - and loss – of - function研究中指出,這些蛋白質深深影響脂肪細胞的 發展。在培養的前脂肪細胞株誘導分化時,C / EBPβ和δ其mRNA和蛋白質是最早且 短暫的表現,C / EBPα則在分化後期的階段被誘發出來,並可轉活化一些Adipocyte specific gene的Promoters (119)。 In vitro的Gain – of - function研究清楚表示了C / EBPβ和δ的Proadipogenic角 色,在沒有荷爾蒙的誘導下,異位表現C / EBPβ能夠誘發3T3 - L1 cells的分化作用。 對C / EBPδ而言,仍是需要有促分化藥劑的存在,但C / EBPδ可加速脂肪細胞生合 成。C / EBPβ也能促進一些細胞分化成為脂肪細胞,異位表現C / EBPβ的NIH - 3T3 纖維母細胞在荷爾蒙的誘導下可進行脂肪細胞生合成,當胚胎的纖維母細胞缺少C / EBPβ或δ時,會輕微降低脂肪細胞生合成的潛力,但是當細胞同時缺少C / EBPβ和δ 時,會嚴重阻礙纖維母細胞發展成脂肪細胞(120)。 In vitro的研究也強烈顯示出C / EBPα在脂肪細胞生合成之角色,Retroviral表 現C / EBPα的3T3 - L1前脂肪細胞或纖維母細胞可被誘導分化成為成熟的脂肪細 胞,而當這些細胞表現有C / EBPα antisense RNA時,會抑制其分化作用(153)。. (3) ADD1 / SREBP1c Adipocyte determination and differentiation factor 1 (ADD1)是Basic helix – loop - helix (bHLH)的轉錄因子,bHLH protein有組織專一性的調節作用,特別是 在肌肉中,與脂肪有共同的Mesodermal origin。ADD1是根據與E - boxes motif (CANNTG)這個bHLH protein的DNA recognition site結合能力,從大鼠脂肪細胞 cDNA中分離出來的(154),ADD1高度表現在棕色脂肪,其次是白色脂肪,肝臟和腎. 20.
(36) 第二章 文獻回顧. 臟。Sterol regulatory element binding proteins (SREBPs)調節膽固醇與脂肪酸代謝 相關基因的轉錄作用(155),Yokoyama等人依據與Sterol regulatory element結合能力 所獨立選殖出來與ADD1相同的轉錄因子,命名為Sterol regulatory element binding protein 1 (SREBP1)。在哺乳動物中有三種SREBPs,分別是SREBP1a、SREBP1c 和SREBP2。ADD1與SREBP1c的Amino terminus是相同的(156),ADD1 / SREBP1c 有同時與Sterol regulatory elements (SREs)和E - boxes結合的Dual DNA - binding specificity。 Kim與Spiegelman將ADD1 / SREBP1c mRNA表現在前脂肪細胞株中發現可明 顯誘導刺激分化作用,表示了ADD1 / SREBP1c在脂肪細胞生合成之角色(157)。高 度表現ADD1 / SREBP1c的3T3 - L1 cells在有荷爾蒙的誘導分化下,與控制組相比較 有增加的Adipocyte marker表現以及脂質堆積。此外,異位表現ADD1 / SREBP1c 的纖維母細胞在強力誘導分化下會有一些脂肪細胞的轉變,這些都指出ADD1 / SREBP1c與脂肪細胞生合成有關。 ADD1 / SREBP1c的脂肪細胞生合成功能的機制目前還是不清楚,而一些研究 指出與PPARγ有關。Kim與Spiegelman在PPARγ reporter system中共同表現ADD1 / SREBP1c與PPARγ發現有更大的轉錄活性,但單獨表現ADD1 / SREBP1c則是少的 作用(157),另一解釋為ADD1 / SREBP1c會生成一些因子而增加了PPARγ的活性。 Kim等人利用顯性抑制ADD1 / SREBP1c來阻斷脂肪細胞的分化,但當給予PPARγ ligands TZD時可完全回復其分化。綜合而言,這些結果顯示ADD1 / SREBP1c促進 內生性PPARγ ligand的生成(158)。 雖然ADD1 / SREBP1c對脂肪細胞生合成的影響尚不清楚,但已知ADD1 / SREBP1c刺激許多脂質生合成與醣解酵素基因的表現,這些基因包括有LPL、Fatty acid synthase (FAS)、ACC、Spot 14或Pyruvate kinase (L - PK)和Glycerol phosphate acyltransferase (159, 160)。當飢餓後再餵食會刺激脂肪組織中ADD1 / SREBP1c的表 現而促進脂質生合成,ADD1 / SREBP1c剔除的小鼠會對餵食後脂質生合成的反應 遲鈍,而胰島素可調節再餵食的作用,因此強力地調節脂肪與肝臟中ADD1 / SREBP1c的表現(161, 162)。. 21.
(37) 第二章 文獻回顧. (4) 其它轉錄因子. 近來有一些其它轉錄因子可能與脂肪細胞生合成有關,但是這些轉錄因子並沒 有像PPARγ與C / EBPs一樣是影響脂肪細胞形成的主要調節者,它們可能是做為對 脂肪細胞發展與功能上某些層面之調節者。. 1 PPARδ ○. PPARδ是廣泛表現的,一些研究顯示利用非專一的PPAR活化劑給予某些異位 表現PPARδ的纖維母細胞,可促進脂肪細胞生合成(163),但其它的研究指出,以各 種Ligands來活化異位表現PPARδ的細胞並不會促進脂肪細胞生合成。Berger等人用 PPARδ ligands給予內生性表現有PPARδ的3T3 - L1 cells,發現並沒有誘導脂肪細胞 的形成(71)。由這些研究可知PPARδ對脂肪細胞生合成的角色仍然是不明確的。. 2 GATA family ○. GATA - 2和GATA - 3這兩個GATA family的轉錄因子,是參與發展過程的 Zinger - finger DNA-binding protein,它們表現在前脂肪細胞中並在最後成熟作用 期間被向下調節。GATA - 2和GATA - 3的重組表現會抑制脂肪細胞分化,此對脂肪 細胞生合成的抑制作用可能是透過降低PPARγ活性來調節的。此外,缺乏GATA – 3 的EC cells可增強分化為脂肪細胞的能力;在基因的肥胖小鼠,其脂肪組織中GATA - 2和GATA - 3的表現顯著地被向下調節(164)。. 3 FOXC2 ○. 轉錄因子FOXC2在脂肪細胞中對基因表現是Pleiotropic action,一些研究指出 這個蛋白質可能在脂肪細胞發展與代謝中扮演重要角色。過度表現FOXC2的小鼠其 脂肪組織有瘦的表型且會增加胰島素敏感性,並增加了脂肪細胞中β - adrenergic respronsiveness以及氧消耗(165)。. 22.
數據

+7
相關文件
The multi-task learning problem comes from our biological application: Drosophila gene expression pattern analysis (funded by NSF and
The multi-task learning problem comes from our biological application: Drosophila gene expression pattern analysis (funded by NSF and
“ Numerical studies of hyperbolic IBVP with high-order finite difference operators satisfying a summation by parts rule.”.. “High-order finite difference methods,
According to the Heisenberg uncertainty principle, if the observed region has size L, an estimate of an individual Fourier mode with wavevector q will be a weighted average of
It’s not chocolate that causes problems, but the highly processed nature of the products containing chocolate and high levels of sugar and fat.. While it’s great that chocolate
– During each time unit, the backlight device at none(C), low light display (DL), and high light display (HL) consumes E C , E DL , and E DH of energy, respectively.. • Of course
• Concept of High Conflict Complainants, their characteristics and relationship with Personality Disorders.. • Tips in handling High Conflict
An alternative activated-state model was generated by substituting the single ligand-biased receptor refinement protocol with the following procedure: (1) rotation of TM VI by
