行政院國家科學委員會專題研究計畫 成果報告
豬與土雞繁殖力表現體及 cDNA 晶片研發計畫-高繁殖力豬胚
表現序列株系之建立
計畫類別: 個別型計畫 計畫編號: NSC91-3112-P-110-001-Y 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 國立中山大學生物醫學研究所 計畫主持人: 李水龍 共同主持人: 薛佑玲,李金龍,黃弘文 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 2 月 13 日
行政院國家科學委員會補助專題研究計畫
;
成 果 報 告
□期中進度報告
(計畫名稱)
豬與土雞繁殖力表現體及 cDNA 晶片研發計劃-高繁殖力豬胚表現序
列株系之建立
計畫類別:□ 個別型計畫
;
整合型計畫
計畫編號:NSC 91-3112-P-110-001-Y
執行期間: 91 年 8 月 1 日至 92 年 7 月 31 日
計畫主持人:李水龍
共同主持人:黃弘文,薛佑玲
計畫參與人員:
成果報告類型(依經費核定清單規定繳交):□精簡報告
;
完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:國立中山大學生物醫學研究所
中 華 民 國 93 年 2 月 5 日
中英文摘要:
中文摘要:
本子計畫主要工作為建立高產豬的 cDNA 基因庫.本年度我們已成功的由 50 個母豬的成 熟未受精卵建立一個基因庫。我們亦利用已知在老鼠的成熟卵中特異表現的基因來測試我 們的基因庫。我們目前已將此基因庫挑選菌落並加以保存,在本年度我們亦選殖了一些基 因定序,利用 NCBI 的 WWW 的 BLAST 分析結果。將來我們會篩除掉高重複表現的基因,並 將此基因庫作完整的定序研究。英文摘要:
The project is concerning in setup porcine cDNA library from high productive sows. We succeeded construction a mature unfertilized oocyte cDNA library from 50 oocytes. Mouse oocyte specific expression genes are used to verify the quality of the library. At last, we picked and stored the library. Some clones are sequenced and the result is analyzed with BLAST program in NCBI WWW site. We will screen out the high repetitively expressed genes and perform whole library sequence in the further years.
目錄
一、 前言...1
二、 研究目的...3
三、 文獻探討...4
四、 研究方法...7
五、 結果討論...10
六、
結論與建議
...14
七、
參考文獻
...15
八、
計劃成果自評
...16
行政院國家科學委員會專題研究計畫成果報告
豬與土雞繁殖力表現體及 cDNA 晶片研發計劃-高繁殖力豬胚表現序
列株系之建立
計畫編號:NSC 91-3112-P-110-001-Y
執行期限:91 年 8 月 1 日至 92 年 7 月 31 日
主持人:李水龍 國立中山大學生物醫學研究所
壹、 前言
豬與土雞為台灣目前農產價值最高的二種畜產動物,約佔農業產值 30%。其品種改 良及養殖方法均為研究的重點,因應目前分子生物及基因體學等快速的發展,利用新的 科學方法幫助解決傳統產業問題乃目前的重要課題,故行政院於民國九十年第五次 SRB 會議即規畫了產產動物基因體的研究方向及目標:1.免疫力、抗病、抗緊迫有關 SNP 及 分子遺傳標記開發, 2.肉質和繁殖力有關的 EST 表現標記的研發,3.生物資訊中心的設 立和生物晶片製作等。而本計畫則將目標對準在豬和土雞繁殖力的研究上,而在豬的研 究方面,則著重於高產母豬之胚的表現基因研究。 繁殖力的控制可由公、母豬共同控制,在此研究方面,農委會已有公豬 testis 之 EST 計畫執行,故本計畫著重於母豬方面。在這方面的研究上,除了本計畫著重的胚發 育外,共有二個主要的方向: 1. 營養與母豬生產力 2. 初期胚的存活及產胚數目 在營養與母豬生產力方面,加強懷胎期間的母豬營養及管理會影響其生產力,如高 清澤(2001),從留種新母豬起到母豬不同繁殖時期,供餵營養素和飼料量不同。影響母 豬繁殖加和生產力。種豬由於遺傳學和育種之進步,供給營養素需要略加調整。另外由 飼料原料中(大豆、玉米)由於基因改變,提升營養成份,所以調製飼料時也要加以調整 之。 重點提示留種豬需及早選取,在體重一○○公斤前予以任食,選定留種用種豬改餵 種豬後期飼料,供餵肉豬肥育料和懷孕前期料均不適當。配種前七-十四天新母豬和老母 豬需充分餵料和供給哺乳期飼料。配種後廿一日內宜限食,不可過食或任食。哺乳期母 豬隻日飼料量為四-五公斤,不可限食在二•五公斤以下,雖然任食可促進繁殖性能表現 (肯定作用),但不符合經營和利益原則。 新母豬 P2 點背脂厚度在一•四○公分以下者, 不宜留種。因為不利繁殖性能表現。也不宜例行使用性荷爾蒙製劑催情,它限用於繁殖 問題時。 70 年代即提出母豬的生產力受幾個主要與母豬繁殖性能管理有關的因素之決定,初 次成功配種懷孕時早齡化,每窩分娩有高生產數,短哺乳期,縮短斷奶至下次發情配種 的間隔期是獲得高生產力的主要因素。目前母豬之生產管理亦已受到重視,戴卓見(2001) 提到,在豬的整個生命期中,必須應進行的飼養強度與哺乳期發生的大量攝食量推薦值 相一致。還建議注意包括適宜溫度控制和光照供給的環境條件作為主要推薦值。通過分 批分娩進行評定,為保證最佳生產力和最成功的豬場,在連續生理階段還必須將分批生 產制引入到母豬的管理中。 在初期胚的存活及產胚數目上,豬從排卵至離乳,通常只能保有 50%不到的窩仔豬, 而這些損失中,哺乳期間的損失約 15 ~ 20%,其餘的 80 ~ 85%發生出生前,而未受精 的損失只有 0 ~ 10%,因此有 70 ~ 75%是在懷孕期間損失,其中懷孕前 25 天 ( 胚階 段 ) 的損失佔 80 ~ 90%,只有 10 ~ 20%係發生胎兒階段, 在排卵率方面,由於窩仔數之上限決定於排卵的數目,母豬的排卵數遠超過其可孕育的胚胎數,因此增加排卵率也 是提高出生活仔豬數的方式之一。而母豬之排卵率主要受到品種、年齡 ( 胎次 )、營養 等因素之影響 (阮喜文, 1996)。 由上述可知,初期胚的存活在豬的繁殖力上扮演重要的角色,除了可以用営養、管 理上改良外,本計畫以功能性基因體學的方式,研究高產族群母豬胚的表現基因,藉此 來發現控制因子。 工作的主要內容為: (主計畫) 1.樣品:將包括卵(豬)、不同發育階段胚與生殖道(卵巢、輸卵管、子宮等)及繁殖相 關組織(如腦下垂體與肝臟)(雞)由採樣團隊提供,與其他分項計晝一共同進行。 2.製備咸高品質蛋白質樣品庫(台灣動物科技研究所團隊負責),並安全保存樣品備份 於兩個以上之不同實驗室內(核心與協力實驗室)。 3.核心實驗室在第一期研究計晝中之任務包括設立蛋白質體研究所需之技術平台與 基本儀器設備(如雙向電泳設備、質譜儀與蛋白質點挖取機)、蛋白質表現及純化、 抗體製備、以及抗體與蛋白質晶片之設計與開發技術平台。 4.協力實驗室則將承接核心實驗室委託之研究分析項目,預期在兩到三年內可確認 3 -5 千個與豬和土雞繁殖力有關之蛋白質,所得到的蛋白質序列資訊經初步確認後 即傳至生物資訊實驗室,以供建立豬和土雞繁殖力蛋白質資料庫• 5.研究計晝亦將應用經由生物資訊實驗室整理與註解後之資訊,選取與繁殖力有關之 蛋白質進行抗體與蛋白質晶片之設計與製作。 本分項計畫: 建立高繁殖力母豬之未受精卵的基因庫,並對其定序及保存。
貳、研究目的、
由高繁殖力的母豬之未受精卵,分離其 mRNA 並製作 EST 基因庫,並保存及定序此基 因庫。
參、文獻探討、
一、卵的早期發育 原始生殖細胞進入生殖腺原基後,不斷地進行有絲分裂來增加數目,這些細胞稱為 卵原細胞(oogonia),所以增值期的特點為卵原細胞的數目不斷地增加。哺乳類和鳥類的增 值期是在胚胎或胎兒時期完成。從這以後直到成體,卵原細胞不再進行有絲分裂。魚類 和兩棲類在成體時,每當生殖季節,卵巢裡的卵原細胞活躍地進行有絲分裂以增加卵原細 胞的數目。 並不是所有的卵原細胞都能夠發育成有功能的卵子,以人類女性胎兒來說,第一個 月的胎兒大約有 1700 各原始生殖細胞移到生殖腺裡,它們活躍的繁殖,,所以在第二個月 胎兒的生殖腺內就含有六十萬個左右的卵原細胞,第五個月胎兒的生殖腺具有了卵巢的 特點,內含七百萬個卵原細胞。這個時期有絲分裂往往停止,其中許多卵原細胞開始進入 減數分裂Ⅰ的前期,這時它們轉變為初級卵母細胞(primay oocyte)。 其餘的卵原細胞逐漸退化,所以胎兒出生時僅有二百萬個初級卵母細胞。胎兒出生 後初級卵母細胞的數目繼續下降,到青春期僅剩下三十萬個初級卵母細胞。此後每個月 有幾個初級卵母細胞因受到激素刺激而發育成熟,但往往只有一個完成了減數分裂和排 卵,其他幾個都退化了。初級卵母細胞經過第一次減數分裂成為一個次級卵母細胞 (secondary oocyte)、一個極體,而次級卵母細胞再經過第二次減數分裂就產生了一個卵子 (ovum)以及第二個極體。 豬之卵母細胞於體內(in vivo)之成熟歷程,約在發情後(即激素刺激)之 36 hr 左右發展 至第二次減數分裂之中期(metaphaseⅡ; MetⅡ),並在發情後 40∼42 hr 排卵(Hunter and Polge, 1966)。而卵的成熟基本上與這些外圍細胞的調節有相當的關係,這些卵丘細胞(cumulus cells)以 gap junction 來調節卵的發育(Heifz, 1987, Martin et al., 2002)。
Martin et al.(2002)對卵在滤泡中的成熟(folliculogenesis)作了以下的重點描述: Oocyte 藉著分泌一些 factors 來調控 granulosa cells 的生長(在 oocyte 旁的稱為 granulosa cumulus cells, 在 follicule 的稱為 mural granulose cells),
1. 老鼠 12 天時(postnatal),oocyte 分泌 FIGa (factors in the germline a),促進 primordial follicles 生成。
2. KL,activin, AMH (somatic cell derived anti-Mullerian hormone)在形成 primary follicle (只有一層 cumulus cell)很重要。
3. 在 primary follicle 中,oocyte 和 cumulus cell 進行雙向調節,並發育為多層 4. 多層 cumulus cells 形成後,須有激素刺激才可形成 preovulatory follicle
(antral),gonadotropin hormones and require follicle-stimulating hormone (FSH) 為必需的激素,此外,oocyte 的 zona proteins, 如 ZP-3, ZP-2 (oocyte-derived zona pellucida proteins)的 knockout mice 均會導致不孕,顯示這些 zona proteins 亦有 調節 granulosa cells 的功能。
i. GDF-9(growth differentiation factor-9)由 oocyte 分泌,調節 cumulus cells. ii. Oocyte 上表現 KIT receptor,
iii. KITL(KIT ligand)由 granulosa cells 表現。
上述基因缺乏均會使老鼠卵停留在 2nd follicle (prenatral) stage. 5. GDF-9在排卵期的功能:
(i) induce hyaluronan synthase 2, pentraxin 3, and tumor necrosis factor–induced factor 6, and suppressing urokinase plasminogen activator (Elvin et al., 2000; Varani et al., 2002)
(ii) stimulates prostaglandin and progesterone synthesis and/or signaling pathways in preovulatory cumulus granulosa cells (Elvin et al., 2000);and
(iii) suppresses the luteinization of cumulus granulosa cells by inhibiting LH receptorexpression (Elvin et al., 2000).
oocyte-secreted TGF-β superfamily protein, bone morphogenetic protein 15 (BMP-15), 可 能和GDF-9合作來調節ovulation (Yan et al., 2001).
6.許多LH response factors 在 periovulatory period 以knockout mouse models得知: prostaglandin and progesterone pathways (e.g., CCAAT/enhancer binding protein b, cyclooxygenase 2, and progesterone receptor)(Richards, et al., 2002).
7.此時oocytes 累積maternal effect factors:
(i) MATER (“maternal antigen that embryos require”): 在two-cell stage非常重要。(Tong, et al., 2000), and
(ii) DNMT1o, oocyte-specificDNA methyltransferase: maintains genomic imprinting crucial for viability of the developing fetus (Howell et al., 2001).
二、卵發育基因體學相關文獻
在老鼠方面,針對早期胚胎發育的表現基因,已有數篇論文發表,Ko et al.(2000), 發表老鼠在胚胎早期的表現,將各時期的老鼠卵、受精卯、2 cell embyro、4 cell embryo、 8 cell embryo,及 Morula (16 cells)、Blastocyst (32-64 cells)等時期表現的基因做成 plasmid library,並且對其定序,結果顯示早期卵及胚(0-16 cells)約有 3000 多 ESTs,至 Blastocysts 增加至約 5000 多 ESTs,並且亦從中找到一些 stage specific gene groups,如:約 291 oocyte specific genes,241 genes (fertilize cell specific),282 genes(2 cells specific),248 genes (4 cells specific),347 genes (8 cells specific)。然而,由於 ESTs 定序為 PCR 放大且 random 取樣定 序,較真實的基因表現的分析可能須由 microarray 發展後才較清楚。
Stanton et at. (2001),利用 NIH databases 上已有的 15 個 mouse early embryo expressed EST libraries (unfertilized, fertilized, 2,4,8,16 cell stages),共有 71346 ESTs,以這個資料進 行分析,發現共有 11483 unique genes,此外亦有未知的 15067 ESTs 無法判定為 unique genes,其中 1585 個 unique 基因並未在其他 libraries 中發現。此外,其中有 109 個基因 為 stage-specific.Stanton 亦利用相似的方法,找到共約 4 千多個 unique genes 在 oocyte and fertilized oocyte 中表現。
在其他的生物早期胚表現基因的研究上,已有一些實驗室從事豬及牛的早期基因表 現的工作,如 Punsuksilli et al.(2002)即以 substrative libraries 的方式,做了牛的早期性胚 表現基因的研究, 在豬的方面,除了中國大陸對其 genomic project 的興趣外,許多畜產 公司亦有相關的研究,但由於其商業應的的價值,其結果並未釋出, 在人類方面,雖然人類的 microarry 已經可以方便的取得,但人類的胚胎來源嚴格, 較無法做完整有系列的取得。然而這方面的研究,必然會對人類的體外受精技術改進等, 具有相當重要的意義,本研究的結果,亦可提供指標基因供將來人類研究的應用。 事實上,我們已建立豬的成熟卵的 substraction library,並從以老鼠的體內成熟的卵 子基因做比對,發現老鼠某些特異表現的基因依然可以在豬卵中找到(see prilimary
data),顯示哺乳類細胞在早期卵的發育可能相似,然而老鼠的胚與豬在早期即可能有差 異,如老鼠的母源基因在 2-cells 時期即大量消失,人在 4cells, 豬在 8cells,而這些母源 性基因的大量消失的機制,是否在更早期的卵就已經排定呢? 這也是由基因體學的比較 研究中,可能可以看出線索。
此外,由基因體學的研究,可以找出評估成熟卵品質的指標,如 Bilodeau-Goeseels S. 指出一些高表現基因如: 2 transcripts for housekeeping proteins (beta-actin and ribosomal protein L30) and 3 transcripts for growth factor ligand or receptors (platelet derived growth factor receptor alpha (PDGFRalpha), basic fibroblast growth factor (bFGF)),並不能代表成熟 卵的品質,這指出可能較少表現的基因扮演更重要的角色,顯示較有系統的研究才有機 會找到真正的指標基因。
肆、研究方法、
所得 EST 株系都要定序。核酸分別由兩端定序,以得正確之表現體序列。本計畫 將採修飾後之 Sanger 法,由中興大學與台灣大學畜產系的兩個核心定序中心為主要負 責單位,而成功大學(孫孝芳教授) ,中山大學(李水龍 黃弘文教授) 和畜試所(張秀 鑾主任)為協力實驗室。台灣大學畜產系的核心定序中心(鄭登貴/丁詩同/朱有田教授) 將購置 ABI3100,以供快速定序之用;中興大學畜產系的核心定序中心(黃木秋教授), 已有 ABI3100 可以協助定序;另成功大學核酸自動定序中心(孫孝芳教授)成立至今已邁 入第四年,該核酸自動定序中心配備了 ABI3100 及 ABI377-96 型的核酸自動定序儀, 可以協助定序。所以我們將使用 ABI3100 及 ABI377-96 型的核酸自動定序儀,並採用 目前最先進之第三代試劑 ABI BigDye Terminator 試劑,從事快速、精確、及長距離的定 序工作。預期配合整體計畫每年 2-4 萬個 cDNA 定序的總目標,必能完成定序工作。第 一年由三個定序實驗室合力,預計配合 EST 株系之建立,完成 2 萬條以上(台灣大學 一萬條,中興大學一萬條,中山大學一千條)的 cDNA 定序。 動物樣品將先作成高品質且表現樣式豐富之 cDNA libraries (興大邱繡河和中 山李水龍教授為兩核心實驗室),各 library 並應妥善保存其株系(clones),每物種組 織株系分析所得數目將達 5∼10 萬個,並應安全保存在標記清楚之至少 2 個不同冰 櫃內。負責定序 cDNA 株系之核心實驗室(台大丁詩同教授及興大黃木秋教授)和協 力實驗室(興大邱繡河、成大孫孝芳、中山李水龍 黃弘文教授和動科所劉世華博 士實驗室)將以最快速(以 ABI377-96 或 ABI3100 定序儀計算每天至少 2 輪,120 個 株系,每週至少 4 天,每年至少 40 週;即每年每個主要定序實驗室的能量為 160 天×120 個株系=19,600 個;ABI3700 具有 3∼5 倍 ABI3100 之能量)完成序列之分析 工作。台大將負責豬卵及胚 cDNA/EST (Expression Sequence Tag), 興大負責土雞生 殖道組織 cDNA/EST, 其他協力實驗室則接受核心實驗室之轉委託或送往國家基因 體中心去執行。兩三年之內可將豬和土雞繁殖力有關之 EST 全部序列得出,預計 得到 3∼5 萬個不重覆(unique)之 EST 序列。此部份之序列除各定序實驗室需執行 初步校正,編刪及排列之工作外,其他如表現基因資料庫建立、長序列比對、演化 樹分析、比較序列學、生物路徑排列等之工作則由生物資訊實驗室(如下述)負責。 由生物資訊實驗室整理所得欲列入 cDNA microarray 之株系將由各核心研究室提供 至興大黃木秋教授(土雞)和動科所劉世華博士(豬)實驗室進行 cDNA 晶片之製作, 並結合本團隊各研究室所送來因研究旨趣而製備之樣本以進行基因表現樣式之研 究,得出對特定胚胎發育階段、產蛋階段、營養影響因子、溫度影響因子等之生物 路徑以提供本國畜產界相關研究主題(如 SNP、遺傳標記、基因轉殖、複製科技等) 之參考。以上所需軟體及工具將由基因及蛋白體資訊實驗室開發及提供,所需基因 表現 mRNA 之確認工作將由興大邱繡河、動科所劉世華博士和中山李水龍教授實 驗室執行。豬卵細胞 cDNA 庫之建立、保存、EST 定序等工作,基本上 cDNA 庫之建立工作預 定於第一年完成,EST 定序則分年完成。
高產豬群由台南畜產所由資料庫中篩選而來,母豬經由產道沖刷出卵子或受精卵後,立 即建立 cDNA 庫。由於每隻母豬僅能得到約 10 來個受精卵,故 cDNA 庫來源的 RNA 量 很少,須經 Clontech 之 SMART 放大技術後再植入 Lambda 病毒載體,另亦可考慮使用 殖體(參見邱教授之建構)。但由於殖體載體所須之 RNA 較多且轉殖效率較差,然而其保 存及 DNA 純化操作上有優勢,須經實際操作評估後再決定使用那一種方法。若可能的話,
亦可以採用二者並用的方式。以下就各相關實驗作描述: 1. cDNA 庫的建立
分離不同時期的豬卵細胞之 Total RNA (oocyte、2-cell、8-cell 及 blastocyst stage). Total RNA 的 純 化 及 cDNA 庫 的 建 立 主 要 依 據 Clontech SMART cDNA library construction kit。 Total RNA 及 mRNA 以 absolutely RNA Nanoprep kit. (Strategene)純 化。 mRNA 以反轉錄脢反轉後,以 Super Smart PCR cDNA synthesis kit (Clontech)放 大。放大後的 cDNA 的品質可用各種 embryogenesis marker genes 的 PCR 反應的結果來 做 檢 驗 。 最 後 cDNA 轉 殖 入 PCR2.1 質 體 (C.TA cloning Kit; Invitrogen) , 以 electro-poration 進入 E.coli,挑出菌落培養(3 重複),並取其一,培養 2ml, overnight, 質體加以純化後可從事定序,其餘則培養並分開貯存於-80°C 冰箱中。 2.cDNA 庫的保存及操作 因為須要的菌落甚多,為了避免人為的錯誤及污染,預定採購 QPix-II 之機器,可 自動將菌落挑點並分裝至 96 孔培養盤。單菌落的菌株可由機器挑點後放入 96 孔培養 盤後,預計將挑 100,000 個菌落,由機器作成 3 重覆,一份貯存於中山大學生醫所, 一份置於台灣科技研究所(ATIT),另一份則以 PCR 放大純化後分送各定序實驗室定 序。 3.EST 定序與分析
插入質體的 DNA 以 SMART 或質體本身的 PCR primer sets 放大後,以電泳分析 確定後,再以 Viogene plasmid purification kits 純化。 定序反應使用 ABI big dye sequencing kit. 最後以 96 wells ABI 377 automated sequencing machine(中山大學)或 ABI 3700(台大)分析。定序的結果送至各單位本身設立的生物資訊中心進行分析。 分 析後的資料再經核心資訊中心進行整合。
實驗流程如下:
5 500mmaattuurreeooooccyytteess((畜畜產產所所)) N NaannooRRNNAAeexxttrraaccttiioonn P PoollyyAA++ 3 3’’ C CDDSSIIIIIIPPrriimmeerr GGG 5’ 5’ O OlliiggoonnuucclleeoottiiddeessFirst-strand synthesis coupled
CCC 5’ GGG 5’ GGG CCC Poly A+ Poly A+
Template switching &
T TAAcclloonniinngg P Piicckkuuppccoolloonniieessaanndd i innsseerrttcchhaarraacctteerriizzaattiioonn T Trraannssffoorrmm((EE..ccoollii::TTOOPPOO1100)) D DNNAAsseeqquueenncceerr 9
伍、結果與討論(含結論與建議)
(一)、豬卵之收集: 材料由台南畜產(陳立人主任)所提供, 已收集材料如下表: Date 耳號 冷凍數 92.04.02 1484-2 4 cell x 57 (25 + 32) 1449-3 8 cell x 11 92.05.02 927-11 8 cell x 7 901-12 16 cell x 5 92.06.06 92712 8 cell x 12 (5+7) 4 cell x 12 2 cell x 2Total: 2 cellx4, 4 cellx37, 8 cellx30, 16 cellx5 92.07 Oocyte: 89
材料封存於毛細管,且置液態氮中保存,其中 oocytes 取出適當量(約 50 顆卵)用於建立基 因庫。
(二)、基因庫之建立
基因庫之建立採用 1.mRNA extraction: absolutely RNA Nanoprep kit. (Strategene), 2.Library construction: Super Smart PCR cDNA synthesis kit. (Clontech); 3.TA cloning Kit. (Invitrogen)等步驟:
B.基因庫之建立:
上述之 50 顆成熟 oocytes,以 absolutely RNA Nanoprep kit 純化後,約可得 25ng total RNA,經由 RT-PCR 之後,由 kit 中的 LD-PCR 放大,並跑膠決定其放大 PCR cycle 數及 size,本次實驗取 cycle 數 30 的產物,進行 TA-cloning。
上述的 LD-PCR 產物,經由 1kb fraction size column(Spin column-1000)純化後,與 PCR2.1 質體進行 TA cloning。再 transform 至 E. coli TOP10 中。
(三)、基因庫品質檢測
利用已知會表現在老鼠 oocyte 的基因(Ko et al.,2000),設計 primers,測試在豬卵中是否 表 現 , 選 擇 表 現 量 高 的 基 因 :oocyte-407, oocyte-603, 及 表 現 量 低 的 基 因 :oocyte-618, oocyte-3973.並設計引子測試是否在豬卵中表現。
Primer name Sequence(5’ to 3’) Product size (bp) 表現高低 (相對值) Oocyte-407 F:TGXTTCTTCACGATXTCCTTT R:GCCAGTTTXGTGTTGGTTTT 235 16 Oocyte-603 F:CCATTGACAACTCGCATCAT R:TGGAAGTTCAAACACCCTTTG 154 13 Oocyte-618 F:TCCTTCTTAAGGCCCCTCTC R:GGAAAGGGAGGGAACAGAAAA 206 1 Oocyte-3973 F:CCTTAATTACTTCCATGCACA R : GGAATXTGTATTTTGGAGGTGCT 170 5 表: 設計引子測試豬卵表現基因。
結果顯示,在老鼠中表現的 oocyte-603 及 oocyte-3973 均在豬卵中亦有表現。而 oocyte407 及 oocyte-618 則沒有預期的產物,可能原因為:
1. 鼠與豬的 mature oocyte 所表現的基因可能有差別。
2. oocyte-407, oocyte618 引子區域的序列,豬及老鼠的差異過大,使 PCR 無法成功。 然而,這個結果亦支持了我們做出的基因庫的品質,因為如 oocyte-168 這種在表現量低的 基因亦可測到。
(四)、定序結果
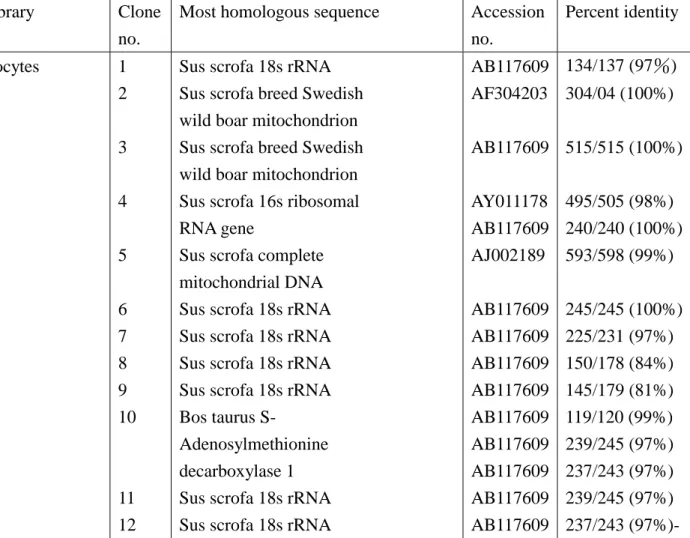
本年度主要工作在建立基因庫,故只做少量定序以測試往後大量定序工入得以進行,目前 以隨機選取 12 菌落的結果做為初步報告:
1. Sus Scrota 18s rRNA 【58.3%】 2. Sus Scrota Mitochondria【25%】
3. Sus Scrota 16s ribosome RNA gene【8.3%】
4. Bos taurus S-adenosylmethionine decarboxylase 1【8.3%】 Table 2. Sequence analysis of randomly chosen cDNA clones Library Clone
no.
Most homologous sequence Accession no. Percent identity Oocytes 1 2 3 4 5 6 7 8 9 10 11 12
Sus scrofa 18s rRNA Sus scrofa breed Swedish wild boar mitochondrion Sus scrofa breed Swedish wild boar mitochondrion Sus scrofa 16s ribosomal RNA gene
Sus scrofa complete mitochondrial DNA Sus scrofa 18s rRNA Sus scrofa 18s rRNA Sus scrofa 18s rRNA Sus scrofa 18s rRNA Bos taurus S-
Adenosylmethionine decarboxylase 1 Sus scrofa 18s rRNA Sus scrofa 18s rRNA
AB117609 AF304203 AB117609 AY011178 AB117609 AJ002189 AB117609 AB117609 AB117609 AB117609 AB117609 AB117609 AB117609 AB117609 AB117609 134/137 (97%) 304/04 (100%) 515/515 (100%) 495/505 (98%) 240/240 (100%) 593/598 (99%) 245/245 (100%) 225/231 (97%) 150/178 (84%) 145/179 (81%) 119/120 (99%) 239/245 (97%) 237/243 (97%) 239/245 (97%) 237/243 (97%)- 結果顯示,未經 normalize 的基因庫,含有大量 18s rRNA,往後挑選時須以 18s rRNA 為 probe, 挑選不能被其雜交的菌落進行定序工作。
六、結論與建議
綜合本年度的工作,得到以下結論:
1. 成功的建立品質不錯的高產豬之成熟未受精卵的基因庫。 2. 挑選菌落並加以保存。
七、參考文獻
高 清 澤 , 以 營 養 改 善 母 豬 繁 殖 表 現 之 最 近 進 展 研 究 , 現 代 養 豬 2001 年 2 月 號 P39~47
戴 卓 見 , 歐 洲 高 生 產 力 母 豬 管 理 制 度 , 現 代 養 豬 2001 年 2 月 號 P63~72, Pig News and Information 2000 Vol.21 No.3 89N-98N
阮喜文, 母豬生產力的影響因素與提昇途徑, 中國畜牧雜誌 1996,第 28 卷 (96),第 1 期 , 9~20.
Bilodeau-Goeseels S., (2003) Effect of oocyte quality on the relative abundance of specific gene transcripts in bovine mature oocytes and 16-cell embryos. Can J Vet Res. 67:151-156.
Elvin, J. A., Yan, C., Matzuk, M.M., (2000), Oocyte-expressed TGF-beta superfamily members in female fertility., Mol. Cell. Endocrinol., 159:1-5.
Heikinheimo, O and Gibbons, WE, (1998), The molecular mechanisms of oocyte maturation and early embryonic development are unveiling new insights into reproductive medicine, Mol Hum Reprod 4: 745-756.
Hafez, E.S.E., (1987), Folliculogenesis, egg maturation and ovulation., Reproduction in farm animals. 5th, edition. Lea & Febiger, Philadelphia. pp. 130-167.
Howell, C.Y.. et al. (2001), Genomic imprinting disrupted by a maternal effect mutation in the Dnmt1 gene, Cell, 104:829-838.
Ko, M.S. et al., (2000) Large-scale cDNA analysis reveals phased gene expression patterns during preimplantation mouse development. Development 127:1737-1749.
Martin M. Matzuk, Kathleen H. Burns, Maria M. Viveiros, and John J. Eppig , (2002), Intercellular Communication in the Mammalian Ovary: Oocytes Carry the Conversation, Science 296: 2178-2180
Richards, J.S., et al. (2002), Ovulation: new dimensions and new regulators of the inflammatory-like response, Annu. Rev. Physiol., 64:69-92.
Stanton, J.L., and Green D.P.L., (2001) Meta-analysis of gene expression in mouse preimplantation embryo development, Mol. Human Reprod. 7:545.
Stanton JL, Green DPL., (2001) A set of 840 mouse oocyte genes with well-matched human homologues., Mol. Human Reprod. 7:521.
Tong, Z.B., et al. (2000), Mater, a maternal effect gene required for early embryonic development in mice, Nature Genet., 26:267-268.
Varani, S. et al. (2002), Knockout of pentraxin 3, a downstream target of growth differentiation factor-9, causes female subfertility., Mol Endocrinol., 16:1154-67.
Yan, C. et al., (2001) Synergistic roles of bone morphogenetic protein 15 and growth differentiation factor 9 in ovarian function., Mol Endocrinol. 15:854-66.
八、計劃成果自評
1. 成功的建立建立高繁殖力豬的基因庫, 保存高繁殖力表現基因, 除了供本次實驗 使用外, 往後對豬的相關研究亦可由此得到相當大的助益.
2. cDNA 庫建立及 cDNA 庫之質量, 由 cDNA 庫中是否有 oocyte 發育中指標基因的表現 來評估,發現此基因庫已含包含高表現型及低表現型之指標基因。
3. 由於大量 18S rRNA 的出現,為避免大量重複的定序,須先以 probe 篩除高度重複 的基因。