馬鬱蘭天然抗氧化成分(鼠尾草酸和熊果酸)預防肥胖導致之心血管疾病:抑制resistin誘發的人類臍靜脈內皮細胞黏附因子表現; Carnosic Acid and Ursolic Acid (Origanum Majorana L.) Prevents Cardiovascular Disease Caused by Obesity: Suppression of Resistin-induced Adhesion Molecule Expression in HUVECs
89
0
0
全文
(2) 中 國 醫 藥 大 學 營 養 學 系 碩 士 班 碩 士 論 文 馬鬱蘭天然抗氧化成分(鼠尾草酸和熊果酸) 預防肥胖導致之心血管疾病: 抑制 resistin 誘發的人類臍靜脈內皮細胞黏附因子表現 Carnosic Acid and Ursolic Acid (Origanum Majorana L.) Prevents Cardiovascular Disease Caused by Obesity: Suppression of Resistin-induced Adhesion Molecule Expression in HUVECs 研究生:林于婷 (Yu-Ting, Lin) 指導教授:曾政鴻 博士 (Jen-Horng, Tseng) 余雅美 博士 (Ya-Mei, Yu) 共同指導教授:江素瑛 博士 (Su-Yin, Chiang) 中. 華. 民. 國. 97. 年. 06. 月. 27. 日.
(3) 謝. 誌. 在這兩年的研究所生涯中,不管是學業或者生活上,我都受益良 多。首先我要感謝的是我的指導教授余雅美老師、曾政鴻老師與共同指 導教授江素瑛老師,感謝你們對於我實驗研究上的指導以及生活上的關 心與照顧,使本論文得以順利完成。同時感謝詹淑秦老師辛苦的幫我收 集臍帶,使我的實驗得以進行。師恩浩瀚,永銘於心。 文稿初成,感謝口試委員余雅美老師、曾政鴻老師與江素瑛老師專 業的指導及寶貴的建議,使論文內容更加完備。 在兩年的求學過程中,感謝所有給予我協助的人,包括芩卉學姊、 杏純學姊,感謝你們細心指導我實驗上的問題;同時感謝我的同窗好友 瑜妏、小孟、姤君、雯渟、依儒、智聖、宜儒的協助及支持;也感謝實 驗室之學妹郁婷、佩琪、榆茜的幫忙;再者,感謝我的好友大華、藝霖、 姿瑤、素月、芳羽、思穎,給予我滿滿的鼓勵與信心。 最後,我由衷的感謝我親愛的爸爸、媽媽以及兩個姊姊,謝謝你們 的支持及關心,使我得以順利完成學業。將此論文獻給所有幫助過我的 人,謝謝你們! 林于婷 中國醫藥大學. 謹致於 營養學系.
(4) Contents. 中文摘要……………………………………………………………………………..........1 Abstract …………………………………………………………………………………...3 Chapter 1 Introduction…………………………………………………………………...5 1-1 The purpose of this research..........................................................................................6. Chapter 2 background………………………………………………………………….7 2-1 The formation of atherosclerosis.............................................................................7 2-2 Obesity and metabolic syndrome……………………………………………………...9 2-3 Resistin and atherosclerosis………………………………………………………….11 2-4 Adhesion and atherosclerosis………………………………………………………...13 2-4-1 Selectins and atherosclerosis…………………………………………………..14 2-4-2 Integrins and atherosclerosis…………………………………………………..14 2-4-3 Immunoglobulin gene superfamily……………………………………………15 2-5 NF-κB translocation and atherosclerosis…………………………………………….17 2-6 Oxidative stress and atherosclerosis…………………………………………………19 2-7 Origanum Majorana L……………………………………………………………….20. Chapter 3 Materials and methods……………………………………………………….35 Chapter 4 Results………………………………………………………………………..47 Chapter 5 Discussion……………………………………………………………………67 Chapter 6 Conclusion……………………………………………………………………72 References………………………………………………………………………………..74.
(5) Figure of contents. Figure 2-1 Endothelial dysfunction in atherosclerosis…………………………………26 Figure 2-2 Fatty-streak formation in atherosclerosis…………………………………...27 Figure 2-3 Formation of an advanced, complicated lesion of atherosclerosis…………..28 Figure 2-4 Unstable fibrous plaques in atherosclerosis………………………………….29 Figure 2-5 Role of the different adhesion molecule n atherosclerosis………………...…30 Figure 2-6 The signal transduction pathway of NF-κB…………………………………..32 Figure 2-7 (A) (B) (C) The picture of Origanum majorana L...........................................33 Figure 2-8 Structure of carnosic acid…………………………………………………….34 Figure 2-9 Structure of ursolic acid……………………………………………………...34 Figure 4-1 Cytotoxic effect of carnosic acid on HUVECs with MTT test………………55 Figure 4-2 Cytotoxic effect of ursolic acid on HUVECs with MTT test………………..56 Figure 4-3 Effects of resistin-induced adhesion of U937 cells to HUVECs…………….57 Figure 4-4 Effects of carnosic acid and ursolic acid on resistin-induced adhesion of U937 cells to HUVECs……………………………………………………………..59 Figure 4-5 Effects of carnosic acid and ursolic acid on the protein levels of VCAM-1 in resistin-treated HUVECs……………………………………………………..60 Figure 4-6 Effects of carnosic acid and ursolic acid on the protein levels of ICAM-1 in resistin-treated HUVECs…………………………………………………….61 Figure 4-7 Effects of carnosic acid and ursolic acid on the protein levels of E-selectin in resistin-treated HUVECs……………………………………………………62 Figure 4-8 Effects of carnosic acid and ursolic acid on the protein levels of NF-κB p65 in resistin-treated HUVECs……………………………………………………..63. i.
(6) Figure 4-9 Effects of carnosic acid and ursolic acid on the protein levels of NF-κB p50 in resistin-treated HUVECs…………………………………………………….64 Figure 4-10 Effects of carnosic acid and ursolic acid on resistin-induced the production of ROS in HUVECs…………………………………………………………….66 Figure 6-1 The role of carnosic acid and ursolic acid in the prevention of atherosclerosis.. ……………………………………………………………………………….73. ii.
(7) Table of contents. Table 2-1 National Cholesterol Education Program (NCEP) Adult Treatment Panel III: The Metabolic Syndrome……………………………………………………24 Table 2-2 World Health Organization (WHO) 1999 definition Of metabolic (insulin resistance) syndrome…………………………………………………………25 Table 2-3 Vascular adhesion molecules in atherosclerosis……………………………….31 Table 4-1 Antioxidative capacities of carnosic acid and ursolic acid in vitro…………....54. iii.
(8) 中文摘要 肥胖是在全世界高度盛行的一種疾病。肥胖所導致的代謝反應是造 成動脈粥狀硬化的一個最普遍的危險因子,而胰島素阻抗也許是肥胖導 致心血管疾病的一個可能機制。近幾年發現一個新的脂肪細胞激素 -resistin,因其具有胰島素阻抗的特性,所以以此來命名。而且根據研究, 它可被視為肥胖與心血管疾病的連結因子。內皮細胞中黏附分子的表現 及單核球細胞黏附至內皮細胞的表現,在心血管疾病中,像是動脈粥狀 硬化的早期階段扮演一個重要的角色。馬鬱蘭是一種唇形花科的香草類 植物,它具有醫藥效果並且呈現抗氧化特性。先前有研究指出鼠尾草酸 (carnosic acid) 和熊果酸 (ursolic acid) 是馬鬱蘭中的主要化合物。在本 篇研究中,我們觀察鼠尾草酸和熊果酸在體外實驗中的抗氧化活性,也 觀察在以 resistin 誘發的人類臍靜脈內皮細胞中,鼠尾草酸和熊果酸對於 人類單核球細胞 (U937) 黏附至內皮細胞的效應,而一些黏附分子的表 現也同樣被檢測。再者,我們觀察這兩種化合物對於細胞內活性氧成份 的生成以及對於 NF-κB 表現的效應。我們的結果指出,鼠尾草酸和熊果 酸在體外實驗中具有抗氧化活性,且這兩種化合物皆能抑制 resistin 誘發 的人類單核球細胞黏附至人類臍靜脈內皮細胞,並且能夠抑制活性氧成 份的生成。而以西方墨點法檢測,鼠尾草酸和熊果酸皆能抑制因 resistin. 1.
(9) 誘 發 的 血 管 細 胞 黏 連 分 子 -1 (vascular cell adhesion molecule-1, VCAM-1)、細胞間黏連分子-1 (intercellular adhesion molecule-1, ICAM-1) 和內皮細胞中選擇素 (selectins) 的表現。除此之外,此兩種化合物能減 低或阻斷 resistin 誘發 NF-κB p50,p65 的表現。總而言之,鼠尾草酸和 熊果酸擁有抗發炎特性,它們能抑制 resistin 誘發的動脈粥狀硬化。此一 結果暗示馬鬱蘭在肥胖導致的心血管疾病中也許扮演一個保護的角色。 關鍵字:馬鬱蘭 (Origanum majorana L.),肥胖,黏附分子,抵抗素 (resistin),心血管疾病. 2.
(10) Abstract. Obesity is a highly prevalent disease over the world. The metabolic effects of obesity are one of the most common risk factors for atherosclerosis. Insulin resistance may be a potential mechanism whereby obesity leads to cardiovascular diseases. A novel adipokine, resistin, is named for its insulin resistance property, and it can be done linkage between obesity and cardiovascular diseases. Expression of cell adhesion molecules by the endothelium and the attachment of monocytes to endothelium may play an important role in the early stage of cardiovascular diseases, such as atherosclerosis. Origanum majorana L. has been known to possess medicinal effects and exhibit antioxidative properties. Previous studies indicated that carnosic acid and ursolic acid are the major compounds in Origanum majorana L.. In this study, we investigated the antioxidative effects of carnosic acid and ursolic acid in vitro. The effects of carnosic acid and ursolic acid on the adhesion of U937cells to resistin-induced human umbilical vein endothelial cells (HUVECs), as well as on the expression of adhesion molecules were examined. Furthermore, we also observed the effect of these two compounds on the formation of intracellular reaction oxygen spiecies (ROS) and the effect on nuclear factor-κB (NF-κB) p50, p65. Our results indicated that carnosic acid and ursolic caid had antioxidative activities in vitro. Both of them can suppress resistin-induced the adhesion of U937 to HUVECs and the production of ROS. Both of them also suppressed the expression of vascular cell adhesion molecule-1 (VCAM-1), intracellular. 3.
(11) cell adhesion molecule-1 (ICAM-1), and endothelial cell selectin by Western blot assay. Besides, they attenuated or blocked resistin-induced NF-κB p50/ p65. In conclusion, carnosic acid and ursolic acid have anti-inflammatory property and inhibit resistin-inducd atherosclerosis, suggesting that Origanum majorana L. may play a protective role in obesity-induced cardiovascular diseases. Key words: Origanum majorana L., Obesity, Adhesion molecule, Resistin, Cardiovascular disease. 4.
(12) Chapter 1 Introduction. Worldwide, the prevalence of obesity is increase dramatically, and previous studies show that nearly two thirds of the U.S. adult population is overweighted or obese (Katz, 2005). A lot of diseases are related to obesity, such as cancer, stroke, heart failure, atherosclerosis, and diabetes mellitus. Insulin resistance may be the cause of clustering of cardiovascular risk factors (Zanolin, ME, et al, 2006). Expression of cell adhesion molecules by the endothelium and the attachment of monocytes to endothelium may play a crucial role in the early stage of cardiovascular diseases, such as atherosclerosis (Ross, 1999). Furthermore, nuclear factor-kappa B (NF-κB), a redox-sensitive transcription factor regulating a battery of inflammatory genes, has been implicated as an eventful factor for initiation and progression of pathogenesis in atherosclerosis (Barnes and Adcock, 1997). ROS play a central role in the pathogenesis of endothelial dysfunction and atherosclerosis (Harrison et al., 2003). Previous studies indicated that the activation mechanisms of NF-κB nuclear translocation has been suggested that to involve ROS (Schreck et al., 1991&1992; Suzuki et al., 1993). Resistin is a recently reported noval adipokine that has been suggested to play an important role in the development of insulin resistance and obesity (Verma et al., 2003). On the other hand, Origanum majorana L., a Labiatae herb of mint family, is commonly used in traditional medicine as well as. 5.
(13) food and cosmetic industries (Vagi, et al., 2002&2005). Carnosic acid (CA) and ursolic acid (UA) are the major compounds in Origanum majorana L. (Vagi, et al., 2004). These different extracts of marjoram possess antioxidative, antimicrobial, and anti-inflammatory effects (Vagi, et al., 2002&2005).. 1.1 The purpose of this research. Resistin is an obesity-induced adipokine. Previous studies indicated that resistin can induce the expression of adhesion molecules and the attachment of monocytes to endothelial cells. Moreover, the anti-inflammatory properties of Origanum majorana L. were reported. In order to develop a potential functional food to protect obesity-induced cardiovascular disease, we investigated the potential protective effects of Origanum majorana L. in resistin-induced human umbilical vein endothelialcells (HUVECs).. 6.
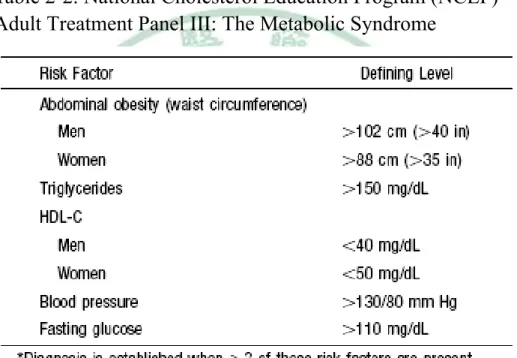
(14) Chapter 2 Background. 2-1 The formation of atherosclerosis. Cardiovascular disease, currently the leading cause of death and illness in developed countries, will soon become the pre-eminent health problem worldwide (Li et al., 2007). Atherosclerosis- a progressive disease characterized by the accumulation of lipids and fibrous elements in the large arteries-constitutes the single most important contributor to this growing burden of cardiovascular disease (Libby, 2002). Atherosclerosis has traditionally been viewed to simply reflect the deposition of lipids within the vessel wall of medium-sized and large arteries. This concept has changed. It is now assumed that a complex endothelial dysfunction induced by elevated and modified low-density lipoproteins (LDL), free radicals, infectious microorganisms, shear stress, hypertension, toxins after smoking or combinations of these and other factors leads to a compensatory inflammatory response (Ross, 1999) (Fig. 2-1). Endothelial dysfunction is characterized by decreased nitric oxide synthesis, local oxidation of circulating lipoproteins and their entry into the vessel wall (Davignon and Ganz, 2004). Intracellular reactive oxygen species similarly induced by the multiple atherosclerosis risk factors lead to enhanced oxidative stress in vascular cells and further activate intracellular signaling. 7.
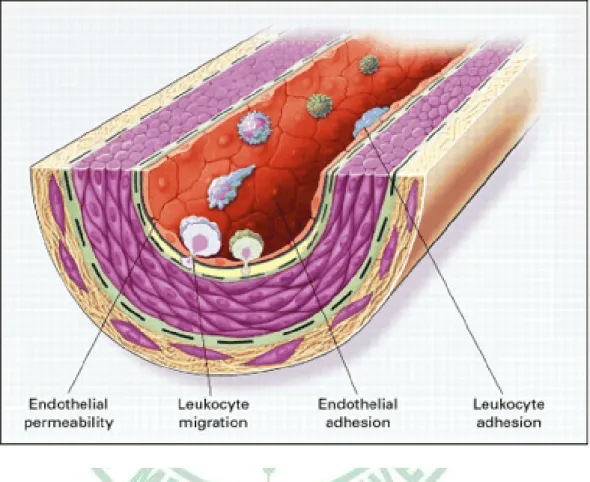
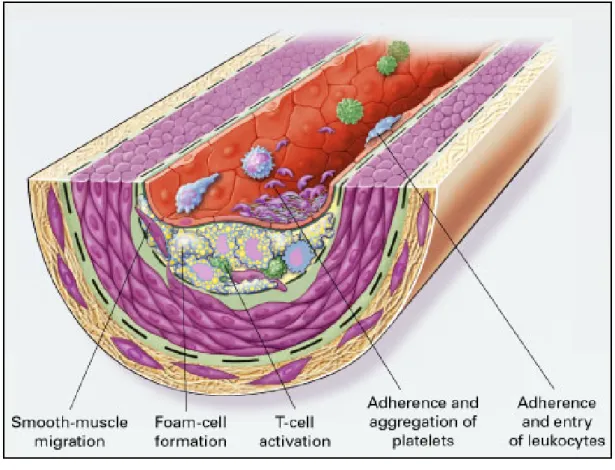
(15) molecules involved in gene expression (Fuster et al., 2005). Upregulation of cell adhesion molecules facilitates adherence of leukocytes to the dysfunctional endothelium and their subsequent transmigration into the vessel wall (Stoll and Bendszus, 2006) (Fig. 2-2). It is now widely accepted that the earliest stages of the development of atherothrombosis are mediated, in large part, by the inflammatory cascade (Libby, 2002). VCAM-1 expression increases recruitment of monocytes and T-cells to sites of endothelial injury; subsequent release of monocyte chemo-attractant. protein-1. (MCP-1). by. leukocytes magnifies. the. inflammatory cascade by recruiting additional leukocytes, activating leukocytes in the media, and causing recruitment and proliferation of smooth muscle cells (Crowther, 2005). In response to signals generated within the early plaque, monocytes adhere to the endothelium and then migrate through the endothelium and basement membrane by elaborating enzymes, including locally activated matrix metaloproteinases (MMP) that degrade the connective tissue matrix (Crowther, 2005). Recruited macrophages both release additional cytokines and begin to migrate through the endothelial surface into media of the vessel (Crowther, 2005) (Fig. 2-3). This process is further enhanced by the local release of monocyte-colony stimulating factor (M-CSF), which causes monocytic proliferation; local activation of monocytes leads to both cytokine-mediated progression of atherosclerosis, and oxidation of low-density lipoprotein (Crowther, 2005).. 8.
(16) Once initiated, many mediators of inflammation have been described to influence the development of the atherosclerotic plaque (Crowther, 2005) (Fig. 2-4). For example, CD40L elaborated within the plaque has been shown to increase the expression of tissue factor (and thus, presumably increase the likelihood of thrombosis) in atherosclerotic plaques; anti-CD40L abrogates evolution of established atherosclerotic lesions in animal models (Schonbeck et al., 2000). Inflammatory mediators expressed by smooth cells within the atherosclerotic plaque include, but are not limited to, interleukin (IL)-1ß, tumor necrosis factor (TNF) and ß, IL-6, M-CSF, MCP-1, IL-18 and CD-40L. The impact of these mediators is diverse and includes mitogenesis, intracellular matrix proliferation, angiogenesis and foam cell development (Crowther, 2005).. 2-2 Obesity and metabolic syndrome. Obesity is becoming a global epidemic, and in the past 10 years in the United States, dramatic increases in obesity have occurred in both children and adults. In adults, overweight is defined as a BMI of 25.0 to 29.9 kg/m2; obesity is defined as a BMI. 30.0kg/m2 (Ogden et al., 2006). The. prevalence of overweight and obesity is approximately 66% in the United States (Ogden et al., 2006). Obesity increased significantly from 23 to 30.5% between 1988 and 2000, and the increase occurred for both men and women and for all age groups (Flegal et al., 2002). Since then, the trend for. 9.
(17) increased prevalence has continued, particularly for adolescents and men. Obesity is also on the rise in other industrialized countries (Ogden et al., 2006). Obesity, type 2 diabetes, and the metabolic syndrome are multifactorial diseases of considerable heterogeneity (Zimmet et al., 2001). However, whereas diagnostic criteria for obesity and for type 2 diabetes are clear cut, this is not the case for the metabolic syndrome (Haffner and Taegtmeyer, 2003). There have been a number of attempts to develop standardized criteria for the diagnosis of the metabolic syndrome. The World Health Organization (WHO) developed a definition in 1998 that stated that individuals need to show evidence of insulin resistance and at least 2 of 4 other factors (hypertension, hyperlipidemia, obesity, and microalbuminuria) (Alberti and Zimmet, 1998) (Table 2-1). Isomaa et al suggested that this definition of the metabolic syndrome strongly predicted cardiovascular disease in the Botnia, Finland population (Isomaa et al., 2001). In 2001, the National Cholesterol Education Program (NCEP) suggested another definition for the metabolic syndrome, which required at least 3 of 5 factors to be present for definition of the metabolic syndrome (Table 2-2). The 5 factors are the following: increased waist circumference, hypertriglyceridemia, low HDL cholesterol, hypertension, and a fasting glucose of 110 mg/dL or higher (NCEP, 2001). This definition is easier to use in clinical practice because glucose tolerance testing, insulin concentration measurements, and microalbuminuria testing are not required (Lemieux et al., 2000). Although the criteria differ slightly, current theories focus on insulin resistance as the prime factor linking. 10.
(18) obesity with adverse metabolic change. The evidence of chronic systemic inflammation is one of the most consistent, and numerous inflammatory markers are highly correlated with the degree of obesity and insulin resistance; many of these inflammatory markers are, in turn, highly predictive of vascular disease risk (Rader, 2000). Pathways leading from obesity to the manifestations of metabolic syndrome involve a number of metabolic risk factors, as well as adipokines, mediators of inflammatory response, thrombogenic and thrombolytic parameters, and vascular endothelial reactivity. Increased adipose tissue mass contributes to augmented secretion of proinflammatory adipokines, particularly tumor necrosis factor-α (TNF-α), along with diminished secretion of the "protective" adiponectin (Sonnenberg et al., 2004).. 2-3 Resistin and atherosclerosis. Over the past few years, much effort has been made to understand the interaction between insulin resistance and endothelial dysfunction, with particular emphasis on adipocyte-derived hormones (adipokines) and their effects on vascular homeostasis (Shuldiner et al., 2001; Cooke and Oka, 2002). The adipocyte has emerged as a key secretory organ, releasing a number of bioactive molecules such as leptin, adiponectin, tumor necrosis factor-α (TNF-α), plasminogen activator inhibitor type 1, and the recently. 11.
(19) described hormone resistin (Steppan et al., 2001). These adipokines not only function prominently in the pathogenesis of the insulin-resistant syndrome but may also serve as important vasoactive factors, directly affecting endothelial function and vascular health (Cooke and Oka, 2002). Previous studies thought that insulin resistance might be mediated by a TZD regulated adipocyte-derived factor. A screen for genes that are induced during adipocyte differentiation but downregulated in mature adipocytes exposed to TZDs led to the discovery of a protein that we have called resistin (Steppan et al., 2000). Resistin is a recently described novel adipokine that has been suggested to play a role in the development of insulin resistance and obesity (Verma et al., 2003). It belongs to a family of resistin-like molecules (RELM), and the resistin mRNA encodes a 114-amino acid polypeptide that contains a 20-amino acid signal sequence (Rajala et al., 2003). Furthermore, Resistin appears to have inflammatory activity; it is a member of a novel family of cysteine-rich secretory proteins called “found in inflammatory zone” (FIZZ) (Holcomb et al., 2000). Indeed, although resistin was originally found to be expressed in adipocytes, several reports have indicated that resistin is highly expressed in human peripheral blood mononuclear cells, and even in leukemia cells, suggesting that resistin may be importantly involved in inflammation and/or immune modulation (Savage et al., 2001; Patel et al., 2003; Yang et al., 2003). In addition, Verma et al. recently found that resistin activates human endothelial cells (ECs) in vitro; when incubated with ECs, recombinant resistin increases endothelin-1 mRNA expression and protein secretion,. 12.
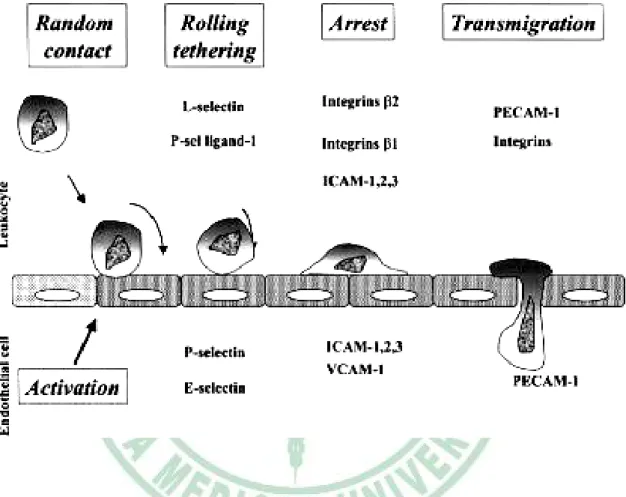
(20) increases vascular cell adhesion molecule-1 (VCAM-1) and monocyte chemoattractant protein-1 (MCP-1) expression, and impairs tumor necrosis factor receptor-associated factor-3 (TRAF-3) expression (Verma et al., 2003). Resistin concentrations were of higher values in obese and diabetic mice. Administration of resistin increased plasma glucose levels and stimulated endogenous glucose production in rodents (Steppan et al., 2001; Rajala et al., 2003). Moreover, previous study indicated that the role of resistin in the development of insulin resistance and diabetes in obese people does not seem to result from its direct effect on glucose metabolism but is closely connected with the intensification of inflammatory processes progressing in the adipose tissue (Janowska et al., 2006).. 2-4 Adhesion and atherosclerosis. Atherosclerosis results in cardiovascular death of approximately 16.7 million people around the world each year (WHO Health Report, 2003). Atherosclerosis is a chronic inflammatory process that is characterized by the formation of plaques consisting of foam cells, immune cells, vascular endothelial cells (ECs), smooth muscle cells (SMCs), platelets, extracellular matrix, and a lipid-rich core with extensive necrosis and fibrosis of surrounding tissues (Elena and Klaus, 2007 ). There are several steps in leukocyte recruitment into vascular tissues (Fig. 2-5) : (1) initial selectin-dependent tethering and rolling, (2) triggering. 13.
(21) of adhesion via chemokines and their receptors or through selectin binding to P-selectin glycoprotein ligand-1 (PSGL-1), adhesion. and. adhesion. strengthening. by. (3) integrin-dependent integrin clustering,. (4). transmigration across endothelium (Elena and Klaus, 2007 ) (Table 2-3).. 2-41-1 Selectins and atherosclerosis. L-, P-, and E-selectins are C-type lectins that bind sialylated and fucosylated carbohydrate ligands presented by sialomucins and mediate initial capture, tethering, and rolling along endothelium (McEver, 2002). L-selectin is expressed on most circulating leukocytes and mediates lymphocyte rolling in high endothelial venules (HEV) of secondary lymphoid organs and at sites of chronic inflammation, where HEV-like vessels are formed (Bistrup, 2004). L-selectin also participates in secondary capture, defined as leukocyte capture by adherent leukocytes (Eriksson et al., 2001; Sperandio et al., 2003). P-selectin is stored in Weibel-Palade bodies of endothelial cells and in intracellular α-granules of platelets and quickly released to the plasma membrane on endothelial cell activation P- and E-selectins are expressed in acute as well as in chronically inflamed endothelium and serve as rolling molecules for monocytes, neutrophils, effector T cells, B cells, and natural killer cells (McEver, 2002).. 2-4-2 Integrins and atherosclerosis. 14.
(22) Integrins are a family of 24 cell-surface receptors composed of 18α and 8β subunits that form αβ heterodimers (Hynes, 2002). Integrins mediate cell-cell, cell-extracellular matrix, and cell-pathogen contact. They regulate leukocyte homing, organize the immunologic synapse, participate in costimulation, migration, and phagocytosis (Luo et al., 2007). Integrins rapidly change the conformation of their extracellular domain structure (inside-outside signaling) and are able to cluster in response to activation (Hogg et al., 2002). In parallel, ligation of integrins leads to a signal cascade from the extracellular domain to the cytoplasm (outside-inside signaling) (Luo et al., 2007). The main integrins that participate in the regulation of leukocyte trafficking are β2 and α4 integrins (Kurzinger et al., 1981).. 2-4-3 Immunoglobulin gene superfamily. Increased expression of adhesion molecules by the activated endothelium is a critical feature of atherosclerosis. VCAM-1 is one of the immunoglobulin gene superfamily, and it is noteworthy that VCAM-1 upregulation is detected mainly at atherosclerosis-prone sites of the endothelium (Nakashima et al., 1998). In human coronary atherosclerotic plaques, elevated expression of VCAM-1 and ICAM-1 and increased numbers of plaque intimal macrophages and T cells were observed within regions of plaque neovasculazation, but less in the arterial luminal endothelium (O’Brien et al., 1996). These results suggest VCAM-1– and ICAM-1–dependent recruitment. of. 15. immune. cells. through. intimal.
(23) neovasculature that may participate in atherosclerosis (Bradfield et al., 2007). Another member of the immunoglobulin superfamily, ICAM-1, is also involved in atherosclerosis, presumably through the regulation of monocyte recruitment into atherosclerosis-prone areas (Kita et al., 2001). ICAM-1 expression is elevated in atherosclerosis-prone aortas and is regulated by proinflammatory stimuli (Kita et al., 2001). Not only oxLDL but also native LDL increase the expression of ICAM-1 on HUVECs and elevates monocyte adhesion to the activated endothelium, suggesting that ICAM-1 is more important in spontaneous atherosclerosis than in response to injury (Manka et al., 2001). Platelet endothelial cell adhesion molecule-1 (PECAM-1), also known as CD31, is a member of the immunoglobulin gene superfamily, a transmembrane glycoprotein with 6 extracellular immunoglobulin (Ig)-like domains (Newman et al., 1990). PECAM-1 is expressed at high density at the lateral borders of ECs and at lower density on the surface of hematopoietic and immune cells, including M , neutrophils, monocytes, mast cells, natural killer cells, lymphocytes, and platelets (Muller, 2003). PECAM-1 gene polymorphisms and elevated soluble PECAM-1 levels are associated with severe coronary artery disease (Elrayess et al., 2003). In common with other adhesion molecules, PECAM-1 has important signaling properties (Newman et al., 2003).. 16.
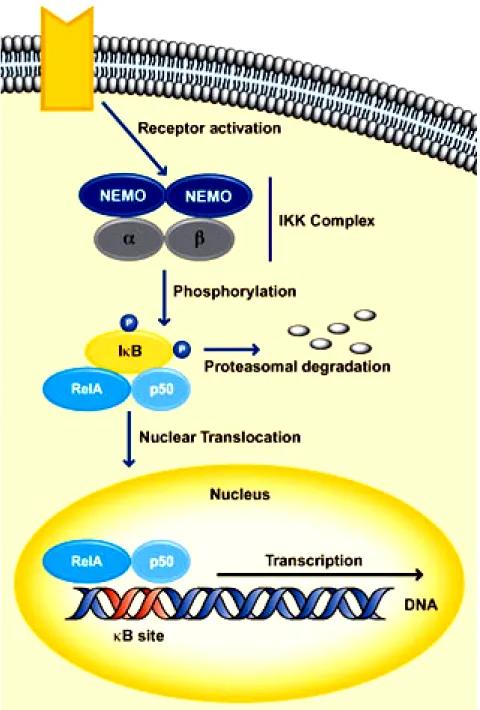
(24) 2-5 NF-κB translocation and atherosclerosis. In general, the family of nuclear transcription factor kappaB (NF-κB) proteins consists of five different members, which are namely p65 (RelA), c-Rel, RelB, p50 and p52. These proteins are all characterized by a structurally conserved N-terminal 300 amino acid region containing specific domains, which allow dimerization, nuclear localization and DNA-binding. Amongst these members of the NF-κB family, only p65, c-Rel and RelB are directly able to activate the transcription of target genes. The transcriptional capacity of p50 and p52, which are initially synthesized as large precursors called p105 and p100, are dependent on dimerization with p65, c-Rel or RelB (Atreya et al., 2008). The ability of NF-κB to regulate gene expression is controlled by chemical modifications such as the addition of phosphate groups, and by interactions with other proteins notably members of the IκB family (inhibitors of NF-κB) (Ting and Endy, 2002). The binding of NF-κB to IκB helps to localize NF-κB in the cytoplasm (Fig. 2-6). Upon activation of the NF-κB signaling pathway by tumor necrosis factor (TNF), IκB kinases target IκBs for degradation. This allows NF-κB to accumulate in the nucleus, where it binds to DNA, resulting in the expression of target genes (Schmid and Adler, 2000). One of the genes activated by NF-κB is that encoding IκBa. Newly synthesized IκBa binds to NF-κB and attenuates the pathway response to TNF, thereby creating a negative feedback loop within the NF-κB/IκB signaling pathway (Atreya et al., 2008).. 17.
(25) To activate NF-κB, there exist two different intracellular pathways – the classic and the alternative pathway – where both result in the release of NF-κB from its inhibitors and in the nuclear localization of NF-κB (Atreya et al., 2008). Classic activation of NF-κB can be initiated by a broad panel of different. stimuli. including. bacterial. cell. wall. components. like. lipopolysaccharide, pro-inflammatory cytokines like tumour necrosis factor (TNF)-α or interleukin (IL)-1, viruses and DNA damaging agents (Atreya et al., 2008). These triggering substances are able to induce intracellular signalling cascades, resulting in a subsequent activation of the IκB kinase (IKK) complex (Atreya et al., 2008). The IKK complex is composed of two catalytic subunits IKKα and IKKβ as well as a regulatory protein, named NFkappaB essential modulator (NEMO) (Atreya et al., 2008). Whilst NEMO serves as an essential adaptor molecule, both catalytic subunits are able to phosphorylate specific serine residues within the IκB molecules (Atreya et al., 2008). Phosphorylation of NF-κB-bound IκB, mainly mediated by IKKβ in case of classic NF-κB activation, subsequently initiates the proteosomal degradation of IκB and finally allows the nuclear localization of NF-κB (Atreya et al., 2008). Some inducers of the classical NF-κB pathway (for example the TNF receptor family member CD40) are additionally able to trigger the alternative NF-κB pathway, which aims at post-translational processing of the p100 precursor to mature p52 (Atreya et al., 2008). In contrast to the classical pathway, NEMO is not absolutely required for alternative NF-κB activation and instead of IKKβ, IKKα is of indispensable importance. Irrespective of the signalling cascade, which leads. 18.
(26) to its activation, activated and translocated NF-κB dimers in the nucleus are able to interact with regulatory NF-κB elements in promoters and enhancers, thereby inducing the expression of NF-κB target genes (Atreya et al., 2008). Generally, genes which are regulated by the transcriptional activity of NF-κB can be categorized into four functional groups: inflammatory and immunoregulatory genes, cell cycle regulating genes, anti-apoptotic genes and genes that encode negative regulators of NF-κB (autoinhibitory feedback loop) (Atreya et al., 2008).. 2-6 Oxidative stress and atherosclerosis. Reactive oxygen species (ROS) play a pivotal role in the pathogenesis of endothelial dysfunction and atherosclerosis (Harrison et al., 2003). Reactive oxygen species (ROS) are intermediary metabolites that are normally produced in the course of oxygen metabolism (Zalba et al., 2006). Under physiological conditions, ROS play a critical role as signal molecules, and ROS produced by activated leucocytes and macrophages are essential for defense against the invading micro-organisms (Zalba et al., 2006). In addition to a mitochondrial origin, ROS can be generated by a great number of enzymes including oxidases, cyclo-oxygenases and lipoxygenases (Zalba et al., 2006). Normally, ROS are contained by a wide array of antioxidant enzymes and endogenous and dietary antioxidants (Zalba et al., 2006). The excess production of ROS or impaired antioxidant defense capacity leads to. 19.
(27) oxidative stress,. in. which. uncontained. ROS cause. oxidation. of. macro-molecules, tissue damage and dysfunction (Zalba et al., 2006). The primary ROS produced in the body is superoxide anion (O2·-) generated from a one-electron reduction of molecular oxygen (Zalba et al., 2006). A number of findings support the notion that enhanced (O2·-) levels play an important role in the pathophysiology of atherosclerosis (Kojda and Harrison, 1999). For instance, (O2·-) may inactivate nitric oxide (NO) and diminish its bioavailability, thus inducing endothelial dysfunction (Kojda and Harrison, 1999). Alternatively, (O2·-) may promote oxidation of the endogenous NO synthase cofactor tetrahydrobiopterin, leading to NO synthase uncoupling with decreased NO production and increased (O2·-) production from the enzyme (Vasquez-Vivar et al., 2003). In addition, the reaction product between (O2·-) and NO, peroxynitrite, constitutes a strong oxidant molecule, which is able to oxidize proteins, lipids and nucleic acids, causing vascular cell damage (Beckman and Koppenol, 1996). Finally, (O2·-) facilitates oxidative modification of low-density lipoproteins (LDL) that play a key role in the formation of atherosclerotic lesions (Stocker and Keaney, 2004).. 2-7 Origanum Majorana L.. Antioxidants are added to food, cosmetic, and pharmaceutical products in tiny amounts to inhibit the oxidation of lipids, thereby greatly extending. 20.
(28) the shelf life of lipid-rich products (El-Ashmawy et al., 2005). Equivocal data on the long-term toxicology of commonly used synthetic antioxidants, such as butylated hydroxytoluene (BHT) and propyl gallate, have combined with consumer preference for “natural” ingredients to motivate extensive research into effective antioxidants derived from plants. Marjoram, Origanum majorana L. (Fig. 2-7 (A) (B) (C)), is a tender perennial herb of the mint family (Lamiaceae or Labiatae), which was formerly classified as Majorana hortensis Moench. Marjoram is commonly used as a herb culinary applications. Because marjoram has been known to possess medicinal effects, it can be used in the industries of cosmetics or pharmaceuticals (Bauer et al., 1990; Price, 1995; Vera, 1999). The plant has been noted to exhibit antioxidant and antifungal properties (Daferea et al., 2000; Deans and Svoboda, 1990). Typically, products identified as marjoram are the dried leaves and flowering tops of O. majorana L, which is found throughout the world. It contains phenolic terpenoids (thymol, carvacrol), flavonoids (diosmetin, luteolin, apigenin), tannins, hydroquinone, phenolic glycosides (arbutin, methyl arbutin, vitexin, orientin, thymonin), triacontan, sitosterol, acids (oleanolic acid) and cis-sabinene hydrate (Leung, 1980; Assaf et al., 1987; Novak et al., 2000; Roth, 2001). The antiviral, bactericidal, antiseptic and antifungal effects of marjoram are attributed to ursolic acid and essential oil and in particular to thymol and carvacrol (Roth, 2001; Bagamboula et al., 2003; Velluti, et al., 2003; Kelly, 2004; Nostro et al., 2004). Antioxidant properties of marjoram (Origanum majorana L.) herb and extracts obtained. 21.
(29) with ethanol, n-hexane, and supercritical CO2 extraction is involved to ursolic acid and carnosic acid (Vági et al., 2004). Many plants have carnosic acid and ursolic acid and they also have anti-inflammatory effect, such as Rosmarinus officinalis and Salvia Officinalis (Albu et al., 2004; Kiyofumi et al., 2004). Carnosic acid (Fig. 2-8) is a phenolic diterpene considered to be the most important antioxidant molecule in extracts of plants. It is a quite labile molecule in hydrophilic media, it is shortly stable in the solvents in which it can be isolated or analysed, and its degradation is enhanced by light or high temperatures (Pelillo et al., 2004). Carnosic acid is an abundant abietane diterpene found in the widely used Lamiaceae herbs. Carnosic acid is a precursor of many other related diterpenes including carnosol, and both of these compounds are powerful antioxidants. The o-diphenol (catechol) structure of carnosic acid is responsible for its ability to inhibit lipid peroxidation and superoxide generation in isolated chloroplasts and microsomes versus chemically induced oxidative stresses. Some constituents of the plant, such as the triterpenes oleanolic and ursolic acids or the diterpene carnosol, were shown to present anti-inflammatory properties or related biological activities (Tokuda et al., 1986; Huang et al., 1994; Liu, 1995). Ursolic acid (UA; 3β-hydroxy-12-urs-12-en-28-oic acid) (Fig. 2-9), a pentacyclic triterpene carboxylic acid found in various plants in the form of an aglycone or as glycosides, is well known to possess many important. 22.
(30) biological functions such as anticancer, anti-inflammatory, hepato-protective, anti-ulcer, hypolipidemic, and anti-atherosclerotic activities (Huang et al., 1994; Manez et al., 1997; Nishino et al., 1998). Further, it has been reported that ursolic acid attenuates the expression of inducible nitric oxide synthase (iNOS) and COX-2 expression through nuclear factor-kappa B (NF-kB) repression in lipopolysaccharide (LPS) or interferon-γ (IFN-γ) activated mouse macrophage (Suh et al., 1998; Kim et al., 2000). Moreover, ursolic acid is able to inhibit key steps of angiogenesis in vitro, including endothelial cell proliferation, migration, and differentiation (Ardenas et al., 2004).. 23.
(31) Table 2-1. World Health Organization (WHO) 1999 definition Of metabolic (insulin resistance) syndrome11: impaired glucose tolerance,* diabetes mellitus,† or insulin resistance,‡ together with at least 2 of the components listed in the table. Haffner and Taegtmeyer, 2003. 24.
(32) Table 2-2. National Cholesterol Education Program (NCEP) Adult Treatment Panel III: The Metabolic Syndrome. Haffner and Taegtmeyer, 2003. 25.
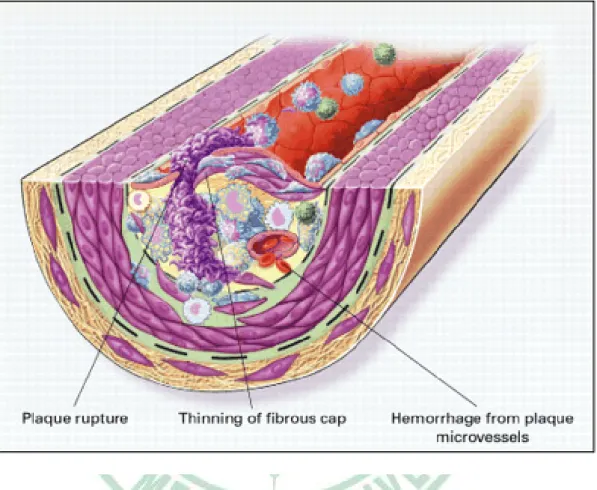
(33) Fig. 2-1. Endothelial dysfunction in atherosclerosis (Ross, 1999).. 26.
(34) Fig. 2-2. Fatty-streak formation in atherosclerosis (Ross, 1999).. 27.
(35) Fig. 2-3. Formation of an advanced, complicated lesion of atherosclerosis (Ross, 1999).. 28.
(36) Fig. 2-4. Unstable fibrous plaques in atherosclerosis (Ross, 1999).. 29.
(37) Fig. 2-5. Role of the different adhesion molecule in atherosclerosis (Blankenberg et al., 2003).. 30.
(38) Table 2-3. Vascular adhesion molecules in atherosclerosis. Galkina and Ley, 2007. 31.
(39) Fig. 2-6. The signal transduction pathway of NF-κB. 32.
(40) Origanum majorana L.. Fig. 2-7. (A) (B) (C) The pictures of Origanum majorana L... 33.
(41) Fig. 2-8. Structure of carnosic acid.. Fig. 2-9.Structure of ursolic acid.. 34.
(42) Chapter 3 Materials and methods 3.1 Instruments. CO2 incubator. NUAIRE, MN, USA. Laminar flow. NUAIRE, MN, USA. Haemocytometer. Boeco, Germany. Eppendorf centrifugator. Hamburg, Germany. Microscope. Nikon, Janpan. Waterbath tank. TKS, Taiwan. Microplate fluorescence reader. Bio-Tek, VT, USA. Shaking incubator. Orbital, VA, USA. Stirrer/Hotplate. Corning, Taiwan. Electrophoresis tank. Bio-Rad, CA, USA. Electrophoresis chamber. Bio-Rad, CA, USA. Power supply. Hoefer, CA, USA. Transfer system. Bio-Rad, CA, USA. PH meter. HANNA, RI, USA. ELISA plate reader. Bio-Tek, VT, USA. Pipetman. Gilson, France. 35.
(43) 3.2 Chemicals. ABTS (2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)). Sigma, MO, USA. BSA (Bovine serum albmin). Sigma, MO, USA. Bradford reagent. Bio-Rad, CA, USA. Carnosic acid. Sigma, MO, USA. CuSO4 (Cupric sulfate). Sigma, MO, USA. DCFH-DA (2’,7’-dichlorofluorscin diacetate). Molcular Probe, Oregon, USA. DMSO (Dimethyl sulfoxide). Sigma, MO, USA. DPPH (2,2-diphenyl-1-picrylhydrazyl). Sigma, MO, USA. DTT (1,4-dithio-D,L-thereitol). Bio-Rad, CA, USA. ECG (Endothelial cell growth supplement). Sigma, MO, USA. FBS (Fetal bovine serum ). Gibco, NY, USA. Gelatin (2% solution type B). Sigma, MO, USA. Glucose. Sigma, MO, USA. Glycine. PROTECH, Taiwan. HBSS (Hank’s balanced salts modified). Sigma, MO, USA. HCl (Hydrochloric acid). Wako, Osaka, Japan. Heparin. Sigma, MO, USA. Hepes buffer solution (1 mol/l). Gibco, NY, USA. H2O2 (Hydrogen peroxide). Wako, Osaka, Japan. KBr (Potassium bromide). SHOWA, Tokyo, Japan. KCl (Potassium chloride). SHOWA, Tokyo, Japan. KH2PO4 (Potassium dihydrogenphosphate). SHOWA, Tokyo, Japan. 36.
(44) L-Glutamine (200 mmol/l). Gibco, NY, USA. MTT (3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyl-terazoliumbromide) Sigma, MO, USA Medium 199. Sigma, MO, USA. Methanol. TEDIA, OH, USA. NaCl (Sodium chloride). Sigma, MO, USA. NaHCO3 (Sodium bicarbonate). Sigma, MO, USA. Na2HPO4 (Disodium hydrogenphosphate). SHOWA, Tokyo, Japan. Nuclear extract kit. TransAM, Active Motif, CA, USA. Penicillin-Streptomyscin. Gibco, NY, USA. Peroxidase (Type Ⅵ-A). Sigma, MO, USA. Recombinant human resistin. ProSpec-Tany, Rehovot, Isreal. RPMI-1640. Gibco, NY, USA. Trolox (6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid). Alderich, WI, USA. Trypan blue. Sigma, MO, USA. Trypsin-EDTA (0.25%). Gibco, NY, USA. SDS (Sodium dodecyl sulfate). Bio-Rad, CA, USA. 2-propanol. Sigma, MO, USA. 95 % Ethanol. Echo Chemical, Taichung, Taiwan. Goat anti-rabbit IgG antibody. Abcam, Cambridge, UK. Mouse anti-actin monoclonal antibody. CHEMICON, USA. Mouse anti-human ICAM-1. Abcam, Cambridge, UK. Mouse anti-human VCAM-1. Abcam, Cambridge, UK. Mouse anti-human E-selectin. Abcam, Cambridge, UK. Mouse anti-human nuclei & chromosomes monoclonal antibody. 37. CHEMICON, USA.
(45) Mouse anti-human P-selectin. Abcam, Cambridge, UK. Rabbit anti-human NF-κB p50. Abcam, Cambridge, UK. Rabbit anti-human NF-κB p65. Abcam, Cambridge, UK. Sheep anti-mouse IgG antibody. Abcam, Cambridge, UK. Tris. PROTECH, Taiwan. Tween-20. PROTECH, Taiwan. Ursolic acid. Sigma, MO, USA. 38.
(46) 3.3 Antioxidatve capacites of carnosic acid and ursolic acid in vitro. 3.3.1 DPPH scavenging assay. Free radical scavenging effect was determined using the free radical generator DPPH (2,2-diphenyl-1-picrylhydrazyl) by a similar method to Yamaguchi et al. 1998. Briefly, the reaction mixture contained 500 μl of carnosic acid and ursolic acid concentration (0-200 μmol/l) and 500 μl of DPPH (0.5 mmol/l in methanolic solution). The DPPH radical scavenging activity was evaluated by measuring the decrease of DPPH radical detected at 517 nm and by determining the difference in the peak area between control and reaction mixture. Inhibition % was calculated via Eq. Inhibition % = (B0-B1/B0) × 100 (where B0: is the absorbance of control. B1: is the absorbance of reaction mixture). The decoloration was plotted against the sample extract concentration in order to calculate the IC50 values, which is the amount of sample needed to decrease the absorption of DPPH by 50%.. 3.3.2 Trolox equivalent antioxidant capacity (TEAC) assay. The assay was carried out using a Spectronic Genesys 5. 39.
(47) spectrophotometer by the improved ABTS·+ method as described by Miller et al., (1993) with slight modification. Briefly, ABTS·+ radical cation was generated by a reaction of 500 μmol/l 2,2’-azino-bis(3-ethylbenzthiazoline -6-sulfonic acid) (ABTS) and 450 μmol/l hydrogen peroxide. The reaction mixture was allowed to stand in the dark for 6 min at room temperature and used within 2 days. All samples were diluted appropriately to provide 20-80% inhibition of the blank absorbance. Fifty microliters of the diluted sample were mixed with 1.9ml of diluted ABTS·+ solution. The assay with the mixture was carried out in triplicate, the mixture was allowed t stand for 6 min at room temperature and the absorbance was immediately recorded at 734 nm (Li et al., 2007). Trolox solution (final concentration 0~15 μmol/l) was used as a reference standard. A value of 1 TEAC in a sample is defined as a concentration equivalent to 1 μmol/l trolox, a water-soluble analog of α-tocopherol.. 3.4 Cell culture. Human umbilical vein endothelial cells (HUVECs) were isolated from human umbilical cords using collagenase type Ⅱ (Jaff, 1973), and cultured on 0.1% gelatin-coated culture dishes in medium M199 (Sigma, MO, USA) supplemented with 10% FBS, 1% Antibiotic-Antimycotic, Glutamine (2 mmol/l), Heparin (10 U/ml), Hepes (10 mmol/l), Endothelial cell growth supplement (ECG) (12.5 μg/ml) at 37 ℃ in a humidified atmosphere of 5%. 40.
(48) CO2 and 95% air. After 3 days, the medium was replaced by fresh medium and subcultured at 1:4 ratio one time per week. All experiments were performed with HUVECs from passage two to five.. 3.5 Cell viability assay (MTT test). The viability of the cells was assessed by MTT (3-[4,5-Dimethylthiazol -2-yl]-2,5-diphenyl-terazoliumbromide) assay (Mosmann,1983), which is based on the reduction of MTT by the mitochondrial dehydrogenase of intact cells to a blue formazan product. Briefly, 1× 104 cells/well were dispensed within 96-well culture plates and incubated with various concentrations of carnosic acid and ursolic acid (with was dissolved in dimethyl sulphoxide) for 24 h. Four hours before the end of the treatment 10 μl MTT (5 mg/ml) was added to each well. At the end of the treatment the incubation medium was removed and the formazan crystals were dissolved in 100 μl of solution of SDS. MTT reduction was quantified by measuring the light absorbance with a ELISA pate reader (μQUANT, Bio-Tek, USA) at 590 nm. The reduction in optical density caused by carnosic acid and ursolic acid were used as a measurement of cell viability, normalized to cells incubated in medium with DMSO only, which were considered to be 100% viable.. 3.6 Adhesion of U937 cells to endothelial cells. Adhesion was evaluated using the human leukemia pro-monocytic. 41.
(49) U937 cells which were labeled with calcein AM (10 nmo/l; Molecular Probe; Invitrogen) (Yu et al., 2007). HUVECs (2×105 cells/well) were distributed into 6-well plates and HUVECs (1×104 cells/well) were distributed into 96-well before the assay and allowed to reach confluence. Then the growth medium was supplemented with 10 and 20 μmol/l carnosic acid or ursolic acid, followed by incubating 100 ng/ml resistin for 24 h in the continued presence of carnosic acid or ursolic acid. U937 cells were grown in RPMI-1640 medium (Gibco, New York, USA) containing 10 % FBS and subcultured at a 1:5 ratio three times per week. U937 cells were incubated with 10 nmol/l calcein AM in RPMI-1640 medium for 30 min at 37℃, then washed with PBS to remove free dye and resuspended in M-199 medium containing 10 % FBS. Labeled U937 cells (2×105) or U937 cells (1×104) were added to each HUVEC-containing well and incubation continued for 30 min. Non-adherent cells were removed by two gentle washes with PBS, then the number of bound U937 cells was determined by a fluorescence plate reader at an excitation wavelength of 485 nm and emission at 530 nm; HUVECs cell monolayers with DMSO only were served as the control.. 3.7 Bradford assay for quantitative measurement of protein. The Bradford assay (Bradford, 1976), a colorimetric protein assay, is based on an absorbance shift in the dye Coomassie when bound to arginine and hydrophobic amino acid residues present in protein. The anionic (bound) form of the dye is blue and has an absorption spectrum maximum. 42.
(50) historically held to be at 595 nm is proportional to the amount of bound dye, and to the amount (concentration) of protein present in the sample. Standard solution contain a range of 0 to 15 micrograms protein (BSA) in 800 μl H2O, followed by adding 200 μl dye reagent and incubate 5 min. 1 μl of sample solution add into 799 μl H2O, followed by adding 200 μl dye reagent and incubated for 5 min. The absorbance was read at 595 nm. The results made a standard curve and the protein concentration of sample was determined by standard curve.. 3.8 Western blot assay for VCAM-1, ICAM-1, E-selectin and NF-κB p65/50. For Western blotting, 3×106 cells were seeded in 10 cm dishes and treated 10 and 20 μmo/l carnosic acid or ursolic acid, followed by incubating 10 ng/ml resistin for 24 h in the continued presence of carnosic acid or ursolic acid, afterwards cells were scraped in PBS and centrifuged at 1,200 rpm for 10 min. Cells were lysed for 1 h at 4℃ with lysis buffer (0.32 mol/l Sucrose, 10 mmol/l Tris, 5 mmol/l EDTA, 2 mmol/l DTT, 1 % Triton X-100, and 1mmol/l PMSF) and centrifuged at 1,2000 g for 30 min at 4℃. The protein concentration of cell extracts was determined with a Bradford-based assay (Bradford, 1976). Cell extracts were loaded per lane, resolved by SDS-PAGE and transferred at room temperature by blotting to polyvinylidene difloride (PVDF) membrane (Shishodia et al., 2003).. 43.
(51) Nonspecific binding was blocked by soaking the membrane in PBST-Tween 20 buffer containing 50 g/L fat-free milk and separately incubated for 24 h at 4℃ with mouse anti-human-VCAM-1, ICAM-1, E-selectin antibodies and rabbit anti-human NF-κB p65, p50 antibodies. Subsequently, the membrane was incubated with a sheep anti-mouse IGg antibody or Goat anti-rabbit IgG antibody for 1 h. The protein levels was determined with the enhanced chemiluminescence. (Upstate,. USA). and. High. performance. chemiluminescence film (Amersham biosciences, USA). Incubation with mouse anti-human β-actin antibody or mouse anti-human nuclei & chromosomes monoclonal antibody were also performed as an internal control. Results were quantified with scanning densitometer using an image analysis system with software.. 3.9 Nuclear extract preparation. Nuclear protein extract were prepared using a nuclear extract kit (TransAM nuclear extract kit, CA, USA) from HUVECs to assay the NF-κB activity. Nuclear protein was prepared as described previously (Dschietzig et al., 2001). 3×106 cells were seeded in 10 cm dishes and treated 10 and 20 μmol/l carnosic acid or ursolic acid, followed by incubating 100 ng/ml resistin for 24 h in the continued presence of carnosic acid or ursolic acid, afterwards cells were scraped in PBS, collected and centrifuged at 1,200 rpm for 10 min. Cells were lysed in hypotonic buffer and centrifuge suspension. 44.
(52) for 30 seconds at 1,4000 g in a microcentrifuge pre-cooled at 4℃. Then resuspend nuclear pellet in 25 μl complete lysis buffer containing 10 mmol/l DTT, lysis buffer AM1, and protease inhibitor cocktail by pipetting up and down. The suspension was incubated for 30 min on ice, and centrifuged for 10 min at 1,4000 g in a microcentrifuge pre-cooled at 4℃. Transfer supernatant and stored at -80℃. The protein concentration was determined with a Bradford-based assay (Bradford, 1976).. 3.10 Measurement of reactive oxygen species. ROS generation in cells was assessed using the probe 2’,7’dichlorofluorscin (DCF) (Molecular Probes Europe BV, Leiden, The Netherlands) according to the method reported by Wang and Joseph (1999). Confluent HUVECs (1×104 cells/well) in 96-well plate and HUVECs (2×105 cells/well) in 6-well plate were pretreated with 10 and 20 μmol/l carnosic acid or ursolic acid, followed by incubation of 100 ng/ml resistin for 24 h in the continued presence of carnosic acid or ursolic acid. After the removal of carnosic acid or ursolic acid from wells, cells were incubated with 10 μmol/l DCFH-DA for 30 minutes. The fluorescence intensity (relative fluorescence units) was measured at 485 nm excitation and 530 nm emission using a fluorescence microplate reader.. 3.11 Statistical analysis. 45.
(53) All results are shown as mean ± S.D. Statistical analysis of the results were performed using One-way ANOVA followed by Dunnet’s test and Duncan’s Multiple Range Test. A value of P<0.05 is considered statistically significant.. 46.
(54) Chapter 4 Results. 4.1 Antioxidative capacities of carnosic acid and ursolic acid in vitro. We used the scavenging effect of free radical DPPH (2,2-diphenyl-1 -picrylhydrazyl) and TEAC (Trolox equivalent antioxidant capacity) to detect the antioxidative activities of CA and UA in vitro. DPPH is a stable free radical, and we can calculate the IC50 of carnosic acid and ursolic acid by the degree of scavenging effect. The IC50 of carnosic acid was 32.31±2.28 μmol/l. The TEAC assay is based on the reduction of ABTS (2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)) radical cation by antioxidants. The TEAC value of carnosic acid was 6.31±0.61 and the TEAC value of ursolic acid was 1.29±0.33 (Table 4-1). A value of 1 TEAC in a sample is defined as a concentration equivalent to 1 μmol/l Trolox, a water-soluble analog of α-tocopherol.. 4.2 The cell viability of carnosic acid for HUVECs. In order to carry on various experiments, we must detect the cell viability of carnosic acid for cell by the MTT test. We seeded 1×104 HUVECs into 96 well plate overnight, and we treated cells with 1% FBS medium 199 for 24h. After 24 h incubation with 10, 20, 30, 40 and 50. 47.
(55) μmol/l of carnosic acid, the cell viabilities of carnosic acid were 100.8±2.36, 99.78±7.54, 94.1±8.27, 91.39±5.11 and 85±6.92 % of control level (Fig 4-1). The 40 and 50 μmol/l of carnosic acid caused a significant reduction of cell viability, respectively. Therefore, we choose 10 and 20 μmol/l of carnosic acid to do all the experiments according to the MTT test.. 4.3 The cell viability of ursolic acid for HUVECs. In order to carry on various experiments, we must detect the cell viability of ursolic acid for cell by the MTT test. We seeded 1×104 HUVECs into 96 well plate first of all, and we treated cells with 1% FBS medium 199 for 24h. After 24 h incubation with 10, 20, 30, 40 and 50 μmol/l of ursolic acid, the cell viabilities of ursolic acid were 98.69±8.49, 93.42±8.6, 92.22±7.72, 76.15±15.32 and 70.75±11.64 % of control level (Fig 4-2). The 40 and 50 μmol/l of ursolic acid caused a significant reduction of cell viability, respectively. Therefore, we choose 10 and 20 μmol/l of ursolic acid to do all the experiments according to the MTT test.. 4.4 The effect of various concentration of resistin induce the adhesion of monocytes (U937) to HUVECs. In order to choose an applicable concentration of resistin to do all. 48.
(56) the experiments, we used different concentration of resistin to do the adhesion test. U937 cells are cultured with 10% FBS RPMI-1640 medium, and maintained the density of cell to 105~106/ml until experiment. We seeded 1×104 HUVECs into 96 well plate first of all, and we treated cells with 1% FBS medium 199 for 24h. After 24 h incubation with 10, 50, 100 and 200 μmol/l of resistin, the percentages of cell adhesion were evaluated by the quantification of calcein AM. The results indicated that 100 ng/ml of resistin has the most powerful ability to induce the adhesive reaction (Fig 4-3). Therefore, we choose this concentration of resistin doing all the experiments.. 4.5 Carnosic acid and ursolic acid inhibit the adhesion of U937 cells to resistin-induced HUVECs. In order to determine the effects of carnosic acid and ursolic acid on the adhesion of U937 cells to endothelial cells, HUVECs were treated with 10 and 20 μmol/l of carnosic acid or ursolic acid, followed by incubation of 100 ng/ml of resistin for 24 h. Then, the percentages of cell adhesion were evaluated by the quantification of calcein AM. The adhesion of U937 cells to HUVECs also was photographed. The control group showed that minimal binding of U937 cells to HUVECs, but adhesion significantly increased when the HUVECs were treated with 100 ng/ml of resistin. Treatment of 10 and 20 μmol/l of carnosic acid or ursolic acid could significantly reduce the number of U937 cells adhering. 49.
(57) to resistin-stimulated HUVECs. 20 μmol/l of CA or UA was more effective on cell adhesion than 10 μmol/l of CA or UA (Fig 4-4 A & B). These results indicate that carnosic acid and ursolic acid can inhibit the adhesion of monocytes to endothelial cells.. 4.6 Carnosic acid and ursolic acid decrease the expressions of resistin-induced VCAM-1、ICAM-1 and E-selectin in HUVECs. Carnosic acid and ursolic acid can inhibit the adhesion of monocytes to endothelial cells. Next, we want to know if the inhibition of cell adhesion by carnosic acid and ursolic acid was due to decrease the expressions of adhesion molecules, such as VCAM-1、ICAM-1 and E-selectin. Before the addition of 100 ng/ml of resistin, we treated endothelial cells with 10 and 20 μmol/l of carnosic acid or ursolic acid. After 24h coincubation, we use western blots assay to determine the effect of carnosic acid and ursolic acid on the protein level of VCAM-1、 ICAM-1 and E-selectin in resistin-induced HUVECs. The expressions of VCAM-1、ICAM-1 and E-selectin were significantly increased by the treatment of resistin (Fig 4-5, 4-6, 4-7). Pretreatment of 10 μmol/l of carnosic acid could significantly decrease the expressions of E-selectin in resistin- stimulated cells, but not VCAM-1and ICAM-1 (Fig 4-5, 4-6, 4-7). Pretreatment of 10 μmol/l of ursolic acid could significantly decrease the expressions of E-selectin in resistin- stimulated cells, but not. 50.
(58) VCAM-1 and ICAM-1 (Fig 4-5, 4-6, 4-7). Pretreatment of 20 μmol/l of carnosic. acid. could. significantly. decrease. the. expressions. of. VCAM-1and E-selectin in resistin- stimulated cells, but not ICAM-1 (Fig 4-5, 4-6, 4-7). Pretreatment of 20 μmol/l of ursolic acid could significantly decrease the expressions of VCAM-1, ICAM-1 and E-selectin in resistin- stimulated cells (Fig 4-5, 4-6, 4-7). These results indicated that the suppression of cell adhesion by carnosic acid and ursolic acid were due to the suppression of adhesion molecules.. 4.7 Carnosic acid and ursolic acid decrease the expressions of resistin-induced NF-κB p65 and p50 in HUVECs. According to the previous results, carnosic acid and ursolic acid could inhibit the level of cell adhesion and the expressions of adhesion molecules in resistin-induced HUVECs. Therefore, we further examine whether the inhibition effect of carnosic acid and ursolic acid was mediated via the activation of translocative mechanism of NF-κB. Before the addition of 100 ng/ml of resistin, we treated endothelial cells with 10 and 20 μmol/l of carnosic acid or ursolic acid. After 24h coincubation, we use western blots assay to determine the effect of carnosic acid and ursolic acid on the protein level of NF-κB p65, p50 in resistin-induced HUVECs. The expressions of NF-κB p65, p50 were significantly increased by the treatment of resistin (Fig 4-8, 4-9). Pretreatment of 10. 51.
(59) μmol/l of carnosic acid could not decrease the expressions of NF-κB p65 and p50 (Fig 4-8, 4-9). Pretreatment of 10 μmol/l of ursolic acid could significantly decrease the expressions of NF-κB p65, but not NF-κB p50 (Fig 4-8, 4-9). Pretreatment of 20 μmol/l of carnosic acid could significantly decrease both the expressions of NF-κB p65 and p50 (Fig 4-8, 4-9). Pretreatment of 20 μmol/l of ursolic acid could significantly decrease both the expressions of NF-κB p65 and p50. (Fig 4-8, 4-9).. These results suggested that the suppression of cell adhesion and adhesion molecules by carnosic acid and ursolic acid was due to the suppression of translocation of NF-κB.. 4.8 Carnosic acid and ursolic acid inhibit the resistin-induced production of ROS in HUVECs. To observe the effects of carnosic acid and ursolic acid on the production of ROS in resistin-induced HUVECs, cells were treated with 100 ng/ml of resistin for 24 h after treatment of 10 and 20 μmol/l of carnosic acid or ursolic acid. HUVECs were labeled with H2O2-sensitive fluorescent probe and detected by fluorescence microplate reader. The producton of ROS in HUVECs also was photographed. The production of ROS induced by resistin was increased significantly (Fig 4-10 A & B). Pretreatment of 10 μmol/l of ursolic acid could significantly decrease the production of ROS, but not 10 μmol/l of carnosic acid (Fig 4-10 A & B). Both 20 μmol/l of carnosic acid and ursolic acid could significantly. 52.
(60) decrease the production of ROS (Fig 4-10 A & B). In addition, pretreatment of HUVECs with 20 μmol/l of carnosic acid or ursolic acid were more effective on decreasing the production of ROS than 10 μmol/l of carnosic acid or ursolic acid (Fig 4-10 A & B). All the results hinted that the inhibitory effect of carnosic acid and ursolic acid may due to the reduction the ROS production.. 53.
(61) Table 4-1. Antioxidative capacities of carnosic acid and ursolic acid in vitro.. DPPH radicals scavenging ability (IC50, μmol/l). carnosic acid ursolic acid. TEAC. 32.31+2.28. 6.31+0.61. N.D.. 1.29+0.33. All values are mean ± S.D. IC50 values were obtained from the dose response curve; IC50 is the amount of sample needed to decrease the absorption of DPPH by 50%. n=3 independent experiments.. 54.
(62) 120. cell viability % of control. 100. *. *. 40. 50. 80 60 40 20 0 0. 10. 20. 30. carnosic acid (μmol/l). Figure 4-1. Cytotoxic effect of carnosic acid on HUVECs with MTT test. HUVECs were treated with various concentrations (0~50 μmol/l) of carnosic acid for 24h in 10% FBS-M199. Values are mean ± S.D., n=3 independent experiments. *P < 0.05 compared with control.. 55.
(63) 120 *. cell viability % of control. 100. *. 80 60 40 20 0 0. 10. 20. 30. 40. 50. ursolic acid (μmol/l). Figure 4-2. Cytotoxic effect of ursolic acid on HUVECs with MTT test. HUVECs were treated with various concentrations (0~50 μmol/l) of ursolic acid for 24h in 10% FBS-M199. Values are mean ± S.D., n=3 independent experiments. *P < 0.05 compared with control. 56.
(64) a d h e s io n % o f c o n tro l. 200 150. a b. ab. 10. 50. ab. c. 100 50 0 0. 100. 200. resistin (ng/ml). Figure 4-3. Effects of resistin on the adhesion of U937 cells to HUVECs. HUVECs were treated with various concentrations. of resistin for 24. hours. Fluorescence-labeled U937 cells were added to the HUVECs monolayer and allowed to adhere for 30 minutes. Values are mean ± S.D., n=3 independent experiments.. a-c. means with different letters are. significantly different at P< 0.05.. 57.
(65) C. R+CA10. R+UA10. R. R+CA20. R+UA20. Figure 4-4. (A) Effects of carnosic acid and ursolic acid on resistin-induced adhesion of U937 cells to HUVECs. (A) Fluorescent images showed the suppression of resistin-induced adhesion of U937 cells to HUVECs after pretreatment with 10 and 20 μmol/l of carnosic acid and ursolic acid, respectively.. 58.
(66) a. 180. adhesion % of control. 160 140. b c. c. c. c. 120 100 80 60 40 20 0 C. R. R+CA10 R+CA20 R+UA10 R+UA20. Figure 4-4. (B) Effects of carnosic acid and ursolic acid on resistin-induced adhesion of U937 cells to HUVECs. (B) HUVECs were induced by resistin for 24 hours after pretreatment of 10 and 20 μmol/l carnosic acid and ursolic acid. Fluorescence-labeled U937 cells were added to the HUVECs monolayer and allowed to adhere for 30 minutes. Values are mean ± S.D., n=3 independent experiments.. a-c. means. with different letters are significantly different at P< 0.05. Abbreviation : C ( control ), R (resistin), CA10 ( carnosic acid 10μmol/l ), CA20 ( carnosic acid 20μmol/l ), UA10 ( ursolic acid 10μmol/l ), UA20 ( ursolic acid 10μmol/l ).. 59.
(67) VCAM-1 β-actin. 160. % of control. VCAM-1. 140 120. a. ab. abc bc. c. c. 100 80 60 40 20 0 C. Figure 4-5.. R. R+CA10 R+CA20 R+UA10 R+UA20. Effects of carnosic acid and ursolic acid on the protein. levels of VCAM-1 in resistin-treated HUVECs. HUVECs were induced by resistin (100ng/ml) for 24 hours after treatment of 10 and 20 μmol/l carnosic acid and ursolic acid immediately. Representative images of the suppression of resistin-induced the expression of VCAM-1 by carnosic acid and ursolic acid in HUVECs. Values are mean ± S.D., n=3 independent experiments. significantly different at P< 0.05.. 60. a-c. means with different letters are.
(68) ICAM-1 β-actin a. 140. % of control. ICAM-1. 120. ab b. ab. ab. b. 100 80 60 40 20 0 C. Figure 4-6.. R. R+CA10 R+CA20 R+UA10 R+UA20. Effects of carnosic acid and ursolic acid on the protein. levels of ICAM-1 in resistin-treated HUVECs. HUVECs were induced by resistin (100ng/ml) for 24 hours after treatment of 10 and 20 μmol/l carnosic acid and ursolic acid immediately. Representative images of the suppression of resistin-induced the expression of ICAM-1 by carnosic acid and ursolic acid in HUVECs. Values are mean ± S.D., n=3 independent experiments. significantly different at P< 0.05.. 61. a-b. means with different letters are.
(69) E-selectin β-actin a. 140. % of control. E-selectin. 120. b. b. bc. bc c. 100 80 60 40 20 0 C. Figure 4-7.. R. R+CA10 R+CA20 R+UA10 R+UA20. Effects of carnosic acid and ursolic acid on the protein. levels of E-selectin in resistin-treated HUVECs. HUVECs were induced by resistin (100ng/ml) for 24 hours after treatment of 10 and 20 μmol/l carnosic acid and ursolic acid immediately. Representative images of the suppression of resistin-induced the expression of E-selectin by carnosic acid and ursolic acid in HUVECs. Values are mean ± S.D., n=3 independent experiments. significantly different at P< 0.05.. 62. a-c. means with different letters are.
(70) NF-κB p65 histone. a. 160. NF-κB p65 % of control. 140 120. a c. cd. bc d. 100 80 60 40 20 0 C. Figure 4-8.. R. R+CA10 R+CA20 R+UA10 R+UA20. Effects of carnosic acid and ursolic acid on the protein. levels of NF-κB p65 in resistin-treated HUVECs. HUVECs were induced by resistin (100ng/ml) for 24 hours after treatment of 10 and 20 μmol/l carnosic acid and ursolic acid immediately. Representative images of the suppression of resistin-induced the expression of NF-κB p65 by carnosic acid and ursolic acid in HUVECs. Values are mean ± S.D., n=3 independent experiments. are significantly different at P< 0.05.. 63. a-d. means with different letters.
(71) NF-κB p50. % of control. NF-κB p50. histone. 200 180 160 140 120 100 80 60 40 20 0. a bc. bc c. C. Figure 4-9.. a abc. R. R+CA10 R+CA20 R+UA10 R+UA20. Effects of carnosic acid and ursolic acid on the protein. levels of NF-κB p50 in resistin-treated HUVECs. HUVECs were induced by resistin (100ng/ml) for 24 hours after treatment of 10 and 20 μmol/l carnosic acid and ursolic acid immediately. Representative images of the suppression of resistin-induced the expression of NF-κB p50 by carnosic acid and ursolic acid in HUVECs. Values are mean ± S.D., n=3 independent experiments. are significantly different at P< 0.05.. 64. a-c. means with different letters.
(72) C. R+CA10. R+UA10. R. R+CA20. R+UA20. Figure 4-10. (A) Effects of carnosic acid and ursolic acid on resistin-induced the production of ROS in HUVECs. (A) Fluorescent images showed the suppression of resistin-induced ROS production in HUVECs after treatment with 10 and 20 μmol/l of carnosic acid and ursolic acid, respectively.. 65.
(73) a. 140. ROS % of control. 120. a. b. b. b b. 100 80 60 40 20 0 C. R. R+CA10 R+CA20 R+UA10 R+UA20. Figure 4-10. (B) Effects of carnosic acid and ursolic acid on resistin-induced the production of ROS in HUVECs. (B)HUVECs were induced by resistin for 24 hours after treatment of 10 and 20 μmol/l carnosic acid and ursolic acid immediately. HUVECs were labeled with H2O2-sensitive fluorescent probe and detected by fluorescence microplate reader. Values are mean ± S.D., n=3 independent experiments. a-b means with different letters are significantly different at P< 0.05.. 66.
(74) Chapter 5 Discussion. Obesity is a highly prevalent disease over the world and the status of it is get worse in western countries. Numerous diseases resulted from obesity, such as hypertension, diabetes mellitus, stroke and cardiovascular disease (Poirier, 2006). Therefore, more and more studies are focus on this area of research to realize the physiological relationship between obesity and various diseases. In this study, we attempted to find out a phytochemical compound to prevent atherosclerosis caused by obesity. It is well known that obesity is a chronic, low-grade inflammation state. The intensity of adipocytokine expression and production is related to the grade of obesity and insulin resistance of human obesity (Peraldi et al., 1998; Azuma et al., 2002). Resistin is a recently described novel adipokine that has been suggested to play a role in the development of insulin resistance and obesity (Steppan, 2001). Although the link between resistin, obesity, and insulin resistance appears to be strong in mice, the exact physiological relevance of resistin in humans remains debatable (Stejskal et al., 2002). Atherosclerosis, a kind of cardiovascular disease, is a progressive disease characterized by the accumulation of foam cell in narrow artery (Libby P, 2002). Expression of cell adhesion molecules by the endothelium and the attachment of monocytes to endothelium may play a crucial role in the early stage of atherosclerosis (Ross, 1999). The interaction between. 67.
數據

+7


相關文件
Other vegetables, prepared or preserved otherwise than by vinegar or acetic acid, frozen, Other than products of heading No. adzuki bean, all varieties and red long bean),
• The order of nucleotides on a nucleic acid chain specifies the order of amino acids in the primary protein structure. • A sequence of three
Reading Task 6: Genre Structure and Language Features. • Now let’s look at how language features (e.g. sentence patterns) are connected to the structure
S3: And the products were the lipase fatty acid…no, no, fatty acid and glycerol and the enzyme remained unchanged. S1: Our enzyme was amylase and our substrate
Microphone and 600 ohm line conduits shall be mechanically and electrically connected to receptacle boxes and electrically grounded to the audio system ground point.. Lines in
In the citric acid cycle, how many molecules of FADH are produced per molecule of glucose.. 111; moderate;
This database includes antigen’s PDB_ID, all sites (include interaction and non-interaction) of a nine amino acid sequence of primary structure and secondary structure.. After
– A.butylparaben; B.benzoic acid; C.methyl paraben; D.sorbic acid.. 碳酸鎂 (magnesi
