0032-0889/83/73/0881/05/$00.50/0
Role
of
Ethylene
in the
Senescence of Detached Rice
Leaves'
Receivedfor publicationApril 15, 1983 and inrevised form July 21, 1983
CHING HUEI KAO AND SHANG FA YANG
Department
of
Agronomy,National
Taiwan University, Taipei, Taiwan,Republic
of China
(C. H. K.);and
Departmentof Vegetable
Crops, Universityof California,
Davis,California
95616(S.
F. Y.)ABSTRACT
The role of ethylene in the senescence of detached rice leaves in relation to their changes in 1-aminocyclopropane-1-carboxylic acid (ACC) content and ethyleneproduction wasstudied. Infreshlyexcised riceleaf segments, ACC level andethylene production rates were very low.Following incubation, theratesof ethyleneproductionincreased and reached a maximum in 12 h, and subsequently declined. The rise of
ethyleneproductionwasassociated with a 20- to30-foldincrease inACC
level.
Ethylene seems to beinvolved in theregulationof the senescenceof
detachedriceleaves.This conclusionwasbased on theobservationsthat
(a) maximum ethyleneproduction preceded chlorophyll degradation,(b) ACC application promoted chlorophyll degradation, (c) inhibitors of
ethyleneproductionandethyleneaction retardedchlorophylldegradation,
and (d) various treatments such aslight, cycloheximide, a,a-dipyridyl,
Ni2+,andcold temperature, which retardedchlorophylldegradation,also
inhibitedethyleneproduction.
Abscisic acid promoted senescence butsignificantlydecreasedethylene
production, whereas benzyladenine retarded senescence but promoted
ethylene production. This is interpreted to indicate that abscisic acid treatmentincreased thetissuesensitivitytoethylene,whereas
benzylad-enine treatment decreased it.
The senescence ofdetached riceleaves is characterized by a
decreasein Chl andproteincontentsandanincreaseina-amino nitrogen content (12). Senescence ofdetached rice leaves has beenreportedtoberetardedbyAg+,Co2+, Ni2+,orby anaerobic conditions(11, 15, 24). Sincethediscovery ofACC2as thekey intermediatein thepathwayofethylenebiosynthesis,our
under-standing oftheregulationof ethyleneproductionhas beengreatly clarified (1, 23). Cobalt ion and anaerobiosis are known to interfere withtheconversionof ACCtoethylene (23).Like
Co2+,
Ni2+inhibitsethyleneproduction in manyplanttissues (16). In contrast, Ag+ isa potent inhibitor of ethylene action in plants(4, 5). Recent workof Aharoni andLieberman (3)andGepstein
and Thimann (9)suggestedthatendogenous ethyleneplayed a significant role in the senescence process of detached tobacco
andoatleaves.
Inthepresent investigation,weexamined the roleof ethylene ininducing senescence of detached rice leaves and the changes 'Supported byaresearchgrant from the National ScienceFoundation (PCM-8 114933). This work was conductedwhileC. H. K.held a Repub-licof China National Science Council Fellowship and S. F. Y. held a
GuggenheimFellowship.
2Abbreviations:ACC, I-aminocyclopropane-l-carboxylicacid; AOA, aminooxyaceticacid; AVG, aminoethoxyvinylglycine; CHI,
cyclohexi-mide;DP,aa-dipyridyl.
of ethylene production in relationto theirsenescence as influ-enced by various environmental and external factors, such as
light, C02, cytokinin, and ABA treatments, which may either
accelerate or retard senescence. Since ACC is the immediate
precursorofethylene anditslevelregulates ethyleneproduction
rate (1), the changes ofendogenous ACC level in relation to
ethyleneproductionratesduringsenescence werealsostudied.
MATERIALS AND METHODS
Plant Materials and Incubation Conditions.Rice (Orvza sativa
L. 'Taichung' Native 1) seedlings were cultured as previously described (14). Theapical 3-cmsegmentsexcised fromthethird
leavesof8-d-old seedlings were used. A group of 10 segments was floated in a 50-ml flask containing 10 ml test solution. Incubation wascarriedout at 30°Ceitherunder light (80
4mol
quanta m-2s-')provided byamixture of cool-whiteandGrolux
lamps orin darkness. In experiments with Ag+ treatment, leaf
segmentswerepretreated with 10mg/l silver nitratefor 30 min,
and then transferred to 50-ml flasks containing test solution
without Ag+. For the experiments in which various
concentra-tionsofCO2 wereused,all flaskswereflushed withC02-free air
and sealedwithrubberserumcaps. Toobtain desired
concentra-tions of C02, known amount ofCO2 was injected into each sealedflask;aC02-free atmospherewasachieved byhanging in
theflaska centerwellcontainingafilterpaperwickwettedwith 0.2mlof 20% KOH.Allexperimentswererepeatedatleast three
times. Similar results and identical trends were obtained each
time.The datareported herewerefromasingleexperiment.
Chl Determination. Chl was extracted anddetermined as
de-scribed before(12). ChlwasexpressedasA665 per 10 segments in 10 ml 80% ethanol.
Determination ofEthylene. Theflaskswereflushed withfresh air, sealed with rubberserum capsand incubated at 30C. For thetime-course experiment(Fig. 1), l-ml gassamplewas
with-drawn from the headspace ofthe flask at specified times. The
flaskswerethenflushed with fresh airandresealeduntil thenext
ethylene determination. Forthose experimentswith treatments
thatincludedABA,ACC, BA, CHI, DP,low temperature, inhib-itors of ethylene production or inhibitors of ethylene action, ethylene thataccumulated in the first 6 h ofincubation under
lightor dark condition was determined. Ethylene was assayed usinga gaschromatograph equippedwith an alumina column andaflame ionizationdetector.
Determination of ACC. ACCwasextractedanddetermined as
describedpreviously( 14).
Determination of Respiration Rates. Leaf segments were placed into 14-mltesttubescontaining0.5mlofdeionizedH20, and sealedwith rubberserumcapsafterflushingwith fresh air.
Respiration rates weredetermined bymeasuring theCO2 accu-mulated in darknessduringa 2-h period; respiration rateswere linear for4h.Gassamplesweretakenasdescribed for ethylene and determined with a gas chromatograph (model 800, Carle 881
Instruments, Inc.) equipped with a silica-gelcolumn and a ther-moconductivitydetector.
RESULTS
Changes in Ethylene Production, Respiration Rate, andChl and ACC Contents of Detached Leaves
during
Senescence inLight or Darkness. The senescenceof rice leaves was followed bymeasuring the decrease ofChl. Figure1 showsthe time courses of Chl and ACC contents and ethylene and C02 production rates of leafsegments floating on water in the light or dark. The decreaseof Chl contentwasevidentat24hafter leaf detachment under bothlight and dark conditions. It is also clear from Figure
1 that light was effective in retarding senescence of rice leaf segments. Under both light and dark conditions, the rates of
ethylene production increased immediately after excision and reached amaximum in 12 h, and subsequently declined. Light substantially inhibited the endogenous ethylene production. The increaseof ACC level in the leaf segments coincided closely with the increase of ethylene production under both light and dark conditions. The endogenous level of ACC rapidly increased and peaked at 9 and 12 h after excision in the light and dark, respectively, and thendeclined. Therise of ethylene production wasassociated with a 20- and 30-fold increase in ACC content inlight and darkness,respectively. During the first 9 h incuba-tion, no significant difference could be found in ACC level betweenlight and dark treatments, but subsequently ACC
con-tentsinthelightwerelower.
,2 0.8_-.-. 0. Y _ o 0.4_ 0 -J 10_ 5-1008 _ DARK ----LIGHT
7500--,3
500___C7,
Qw'~~~~~~0
Therelationship between leaf senescence andrespiration rate
is not a simple one. Ifsenescence is viewed as ageneral decline in function (20), a steady decline of respiration rate would be
expected.On the other hand,some senescence-related changes, such as thesynthesis ofdegradative enzymes (20),seem torequire higher respiratory activity. Indetached leaves, therise of respi-rationrateis usually preceded bya steepfallor a constant rate
(2, 17, 18). Itis also true in rice leaf segments as shown in Figure 1. Respiration rate decreases during early stage of incubation, butit started to increase after about 12 and 48 hincubation in the light and dark, respectively. Under both light and dark
conditions, respiration peaked at a later stage (about 72 hafter incubation), during which time leaf segments had low Chl level. It seems thattheriseof respiration in the presentsystem isthe
result rather than the cause of leaf senescence. In climacteric fruits, the onset of respiratory increase is accompanied by the
onsetofethylene production, whereas theethylene production in rice leaves occurs considerably earlier than maximal respira-tion rate. Thus, it is unlikely that ethylene plays any causative roleininducing the rise ofrespirationrateduring the senescence of rice leaf segments. Figure 1 also shows that light substantially promotedrespiration rate throughout the course of senescence. Since respiration was determined under dark condition, the higherrespiration rate is unlikely due to photorespiration.
Effects of Inhibitors of Ethylene Production and ACC on the Senescence of Detached Leaves. If ethylene plays a regulatory role in senescence, it is expected that inhibitors of ethylene
DARK ---- LIGHT o* C2H4 40 A ACC 30 T E 20 '. \ Q~~~ - 0 24 48 72 96 TIME(h)
Table I. EffectsofACC,AVG,andAOA onEthyleneProduction and Chl Content ofDetached Rice Leaves in Light and Darkness Chlwasdeterminedafter3 and4din darknessandlight, respectively.
Darkness Light Treatment C2H Chl C2H4 Chl nlg-'h- A"5 nlg-'h-' A"s Control 7.88 0.383±0.021 3.62 0.403±0.034 ACC, 1 mM 15.95 0.334±0.002 9.37 0.292±0.008 AVG,0.2mM 1.22 0.452 ±0.031 1.23 0.521 ±0.006 AOA, 1 mM 2.22 0.458±0.019
biosynthesis wouldretardsenescence.AVGandAOA,whichare
known to inhibit ethyleneproduction by blocking ACC forma-tion (23),significantly retardedthesenescenceof detachedleaves incubated inthe lightordark(Table I).Table I also shows the effect of ACC, the immediate precursor ofethylene (1), which significantly promoted ethylene production, and accelerated
se-nescenceofdetached leaves under both dark andlightconditions.
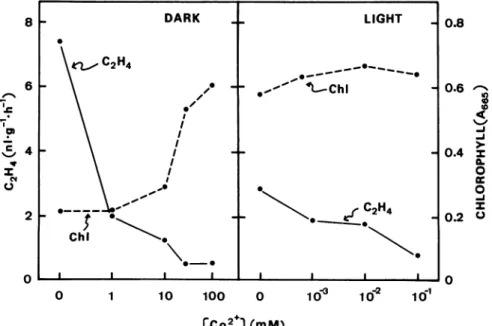
C02+,
an inhibitorofethyleneproduction (5), which blocks theconversion of ACC to ethylene (25), also retarded senescence
andethyleneproduction in rice leaves(Fig.2).
TheeffectsofCo2'aremorepronounced in darknessthan in
thelight. Itshould be notedthat theeffective concentration of
Co2+toretardsenescenceandtoinhibitethyleneproduction in thelight ismuchlowerthan that indarkness.
Effects of Inhibitors of Ethylene ActionontheSenescence of
DetachedLeaves.
Age
andCO2 are knownto inhibitethylene action in many physiological responses(4, 5, 7, 24). BothAge
andCO2effectively retardedsenescenceofdetachedriceleaves, although they promoted ethylene production (Table II). Thepromotiveeffect ofAg+andCO2onethyleneproduction inother plant systems has been well documented (3, 14). Ag+ was, however,lesseffective than CO2onretardation of Chl degrada-tion. Theseobservationsareincontrast tothoseof Aharoniand
Lieberman (3), who reported that Ag+was moreeffective than CO2 inretardingsenescenceintobaccoleaves.
Effects ofSenescenceRetardantsonEthylene Productionand
Senescence ofDetached Leaves. CHI and Ni2+ have been
re-portedtoretardsenescenceindetachedrice leaves(1 1, 15).The
8 6 0-'- 4 %S4 C., 2F 0 0
FIG. 2. Effects ofthe concentrations ofCo2+, determined after3 d.
10 100
chelator, DP, and low temperature (5C) were also known to
delaysenescenceinoatleaf tissues in thedark (19, 20).All these treatments areexpectedtoinhibit ethylene production, if ethyl-eneplaysarole inregulating senescence. Indeed, itis thecase. Table III shows theeffects ofCHI,DP,Ni2+,and lowtemperature
(5C) on leaf senescence and ethylene production. All these treatments notonlyeffectively retarded leafsenescence,but also,
inhibitedethyleneproduction.
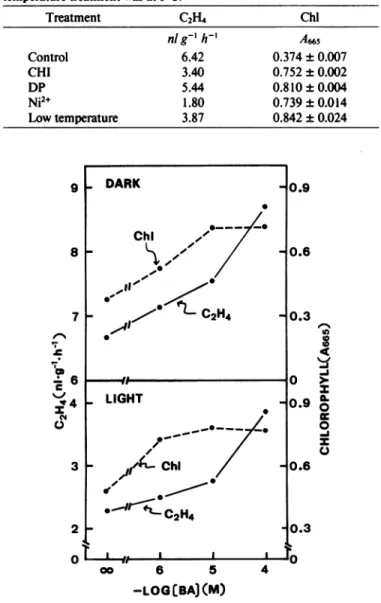
Effects ofBA and ABA on Ethylene Production and Senes-cence of Detached Leaves. It has long been recognized that
cytokininsare effective in retarding the senescenceofmost, if
notall, leaves.Theeffect of cytokinins in retardingsenescenceis species-orvariety-specific; fortherice varietyused inthis inves-tigation, BAhasbeen foundtobe the mostactivecytokinin in retardingsenescencein thedark(11). The effect ofBA on leaf
senescence in relation to ethylene production is presented in Figure 3. Although BA effectively retarded senescence in the
light,aswell asinthe dark, BAsignificantlypromoted ethylene
production. Among the known promotersofsenescence, ABA
hasbeenstudiedmostwidely. Recently, Gepstein and Thimann (8) claimed thatABAisanendogenousfactor inleafsenescence.
Figure 4 showsthe effect ofABA on ethylene production and
leaf senescence in the light. Ethylene production was greatly decreasedbyABA. In contrast,leafsenescenceincreasedasthe
concentration ofABA were increased. In dark, ABA also
pro-motedleafsenescence anddecreasedethyleneproduction(data notshown).
DISCUSSION
The changes ofACC content and ethylene production have beenstudied in ripening fruitand senescing flowers (6, 10). In
senescing rice leaves,therelationshipbetweenACCandethylene
production isgenerally similar tothat found in ripening fruits andsenescing flowers.Infreshly excised rice leafsegments,both
ACCcontentand ethyleneproduction rate were verylow.
Sub-sequently, ethyleneproductionrateincreased whichwas
accom-panied byan increase in ACCcontent, suggestingthat ACC is an intermediatein thebiosynthesis of ethylene throughout the
senescenceprocess.Thus, thebiosyntheticpathway insenescing rice leaves appears to be the same asthat established in pple tissue(1). 0 io0' I10 161 0.8 0.6 ^-J 0.4 to 4
I-0,
-I -i 0.4 x 0 0 -J 0.2 0 0 [Co2 ) (mM)appliedas COCI2, on Chl content and ethylene production ofdetached rice leaves. Chl was
DARK LIGHT \n-- C21-4 ._- -_ -.- 1L.Chl .____ L-- + \ g C2H4 Chi U I l l l l
Table II. EffectsofAgNO3orCO2 TreatmentonEthylene Production
andChlContentofDetached Rice Leaves in Light and Darkness
Chl was determined after 3 d in darkness or 4 d in the light in
experiment I and after3 d in experiment 2. For
AgC
treatment, thedetachedleaveswerepretreatedfor 30minin 10mg/l AgNO3 solution; forCO2 treatment, the leaves were incubated in 5% CO2. Ethylene
accumulated in the first 6hof incubationwasdetermined.
Darkness Light Treatment C2H4 Chi C2H4 Chi nl g-' h-' A"s nlg-'h' A"s Exp. I Control 7.04 0.348±0.004 2.40 0.385±0.005 Ag+ 15.58 0.491±0.031 9.75 0.544±0.055 Exp. 2 Control 5.81 0.216±0.021 4.64 0.494±0.034 CO2 8.03 0.570±0.022 16.76 0.650±0.009
Table III. Effects ofCHI, DP,Ni2+, and Low TemperatureonEthylene Production and Chi Content ofDetached Rice Leaves in Darkness Chlwasdetermined after 3 d in darkness. The concentrations ofCHI,
DP, and Ni2+ employed were0.5, 0.1, and 10 mM, respectively. Low temperature treatmentwasat5°C.
Treatment C2H4 Chl nlg-' h-' A"s Control 6.42 0.374±0.007 CHI 3.40 0.752 ±0.002 DP 5.44 0.810±0.004 Ni2+ 1.80 0.739±0.014 Lowtemperature 3.87 0.842±0.024 9 8 7 *l% 6 C) 2 DARK _0. 0
,/11/0--
/
Chl #,.. le00 -0 .00 .01II~ ~ - / C2H4 _04 0 -1 1I0 LIGHT/Af
Chi *-/ t C2H4 I I l I .. .9 .6 .3 0.9 0.6 uN D 2 oti 0 0 Q 0.3 10% GO 6 5 4 -LOG[BAJ(M)FIG. 3. Effects of BA concentrationsonethylene productionand Chl
contentof detachedriceleaves.Chlwasdetermined after3 d.
3 .C - 2 1 co 6 5 4 0.8 e e 0 0.6 < IL 0 0.4 ° 0 0. 0.2
-LOG(ABA]
(M)FIG. 4. Effects ofABAconcentrationson ethylene production and
Chlcontentof detachedriceleavesinlight. Chlwasdetermined after2 d inlight.
Light substantially inhibited ethylene production throughout
the senescence process. During the first 9-h incubation, there
was nosignificant difference in ACCcontentbetween light and
dark conditions(Fig. 1), indicatingthat the inhibition ofethylene production by lightisexertednot atthe level of ACC butatthe conversionof ACCtoethylene. Using detachedrice and tobacco
leaves, Kao and Yang (14) found that inhibition of ethylene production by lightwas mediatedthrough the internal level of
C02, which directlymodulated theactivityof the enzyme
con-verting ACCtoethylene.
Sincethe increase ofethylene productioninrice leaves is rapid following excision,one mayarguethattheethylene production is probably a wound orother stress response. When detached
leaves were used to study senescence and determine ethylene production, wounding is always a problem. However, in the
present study, each long and narrow rice leafwas cut once
transversely, thearea ofwounding wasverysmall. It has been
reported that there was a slight but significant increase in the
freshweight of rice leafsegmentsfloatedonwater 13). Therefore,
therapidincrease ofethylene productioncouldnot resultfrom
water stress. The onset of ethylene production is therefore thoughttobeanexcision-relatedresponse.
In the study of hormonal control of leaf senescence, two approachesarecommonly employed.The first andmostwidely
used is external application ofplant hormones or chemicals
whichareknown toaffectthe synthesisortoexert an effecton
the action of the hormone. The second istocorrelatechangesin
endogenoushormonecontentwithsenescence.Thepresentstudy
indicated thatethylene participatesintheregulationof rice leaf
senescence. This conclusion was based on observations(under
both light and dark conditions) that (a) endogenous ethylene production preceded Chl degradation (Fig. 1); (b)inhibitors of
ethylene productionsuchasAVG and
CO2"
retardedsenescence(Table I; Fig. 2);(c)ACC,the immediateprecursorofethylene, promotedsenescence(Table I); (d)inhibitorsofethyleneaction
as Ag+ andCO2 retarded senescence (Table II); and (e) other treatments suchaslight, CHI, DP, Ni2+, and lowtemperature,
which retarded senescence, also inhibited ethylene production (Fig. 1; Table III). Recently, the importance of ethylene in regulatingsenescenceof detached leaveshas also been reported byAharoni and Lieberman(3)andGepsteinand Thimann(9).
Usingoatleaves, Gepsteinand Thimann(9)found kinetindid notaffectethylene production under darkconditions, but
pro-moted ethylene production under light conditions. They also
found that ABA promoted ethylene production, which is in contrasttoourresults (Fig. 4). The effect ofABA onethylene
productionfoundbythemwasprobablyawoundresponse,since
0
'11*
*.. Chi %*11
.
~~~%0
C2H4 '-S ' 1 I I' I-theyused fineabrasivetoscrub leaftissues in ordertoenhance
the entry of ABA. Wright (22), however, reported that ABA
significantly inhibited ethylene production from both excised wiltedandnonstressedwheatleaves.
WhenABAandcytokininsareappliedexogenously,theyare
effective in promoting and retarding, respectively, leaf
senes-cence.Furthermore,theendogenouslevelofABA and cytokinins
increases and decreases, respectively, before theonsetof senes-cencesymptoms(20). Thus, it is probablethatleafsenescenceis
regulated bymanyfactorsincluding interactionsamong
cytoki-nins,ABA, and ethylene. Recently, Trewavas (21) developeda
concept ofusing variations of quantitative tissue sensitivity to
explain the developmental phenomena. It seemsthat ethylene
effect is governednot only bythe ethylene production rate of thetissue, but alsoby the tissuesensitivity toethylene. In the present system,ABApromotedsenescencebutinhibitedethylene
production, whereasBAretardedsenescencebutpromoted
eth-yleneproduction.Asimpleexplanationwhichaccountsforthese
seemingly contradictoryobservations is thatABAincreases tissue
sensitivity toethylene,whereas BAdecreases it. Consequently,
eventhough ABA inhibits and BA promotes ethylene produc-tion, the overall ethylene effects may not parallel with their
ethylene productionrates.
Acknowledgment-The assistance of DaoHungLiu isgratefullyacknowledged. LITERATURE CITED
1. ADAMSDO, SFYANG 1979Ethylenebiosynthesis: identification of
1-amino-cyclopropane-l -carboxylic acid as an intermediate in the conversionof methioninetoethylene.Proc NatlAcadSciUSA76: 170-174
2. AHARONIN,M LIEBERMAN 1979Patternofethylene productioninsenescing leaves.Plant Physiol 64: 796-800
3. AHARONI N, M LIEBERMAN 1979 Ethyleneas aregulatorofsenescencein tobacco leaf discs. PlantPhysiol64: 801-804
4. BEYER JR EM 1976 Silver. A potentantiethylene agent incucumber and
tomato.HortScience11: 195-196
5. BEYER JR EM 1976 A potentinhibitor ofethyleneaction inplants. Plant
Physiol 58: 268-271
6. BUFLERG,YMOR,MSREID,SF YANG 1980Changesin I-aminocyclopro-pane-l-carboxylicacidcontentofcutcarnation flowers in relationtotheir
senescence.Planta 150: 439-442
7. BURGSP,EHBURG 1967Molecular requirements forthebiologicalactivity
ofethylene.PlantPhysiol42: 144-152
8. GEPSTEINS, KV THIMANN 1980 Changes in the abscisic acidcontentofoat
leaves duringsenescence.ProcNatl Acad Sci USA 77: 2050-2053
9. GEPSTEINS, KV THIMANN 1981 The roleof ethylene in thesenescenceofoat
leaves. Plant Physiol 68: 349-354
10. HOFFMANNE, SFYANG1980Changesof Iaminocyclopropane-l-carboxylic acidcontentinripening fruit in relationtotheir ethylene productionrates.
JAmSoc Hortic Sci 105: 492-495
11. KAo CH 1978Senescenceofrice leavesII.Antisenescentaction of cytokinins.
ProcNatl Sci Counc ROC 2:391-398
12. KAoCH 1980 Senescence of riceleaves IV. Influence of benzyladenine on chlorophyll degradation.Plant CellPhysiol21: 1255-1262
13. KAo CH1981 Senescence of rice leavesVI.Comparative study ofthe metabolic changesofsenescing turgid and water-stressedexcised leaves. Plant Cell Physiol22:683-388
14. KAoCH, SF YANG 1982 Light inhibition oftheconversion of l-aminocyclo-propane-l-carboxylic acidtoethyleneinleaves is mediated through carbon
dioxide. Planta155: 261-266
15. KAoCH, SM Yu 1981 Senescenceof riceleavesV.Retardation ofsenescence
bynickelousorcobaltous ions. Proc NatlSci Counc ROC 5: 45-48
16. LouOL, SF YANG 1976 Inhibitionofethylene productionby cobaltous ion.
PlantPhysiol58: 114-117
17. MCGLASsON WB,BWPOOVAIAH,HCDosITAL1975Ethylene productionand
respirationinagingleafsegmentsand in disks of fruittissueof normal and
mutant tomatoes.PlantPhysiol56: 547-549
18. TETLEYRM,KV THIMANN1974 The metabolism ofoatleavesduring senes-cenceI.Respiration, crabohydrate metabolism,andthe action ofcytokinins. PlantPhysiol54:294-303
19. TETLEYRM, KV THIMANN1975 The metabolism ofoatleavesduring senes-cenceIV.The effects ofa,a-dipyridyland othermetalchelatorson senes-cence.PlantPhysiol56: 140-142
20. TuIMANN KV1980 Thesenescenceof leaves. InKVThimann, ed,Senescence in Plants.CRC PressInc,BocaRaton,FLpp85-115
21.TREWAVAS A 1981 How doplant growth substances work? PlantCell Environ 4:203-238
22. WRIGHTSTC 1980 The effect of plantgrowth regulatortreatmentsonthe levels ofethylene emanatingfromexcisedturgidandwilted wheat leaves. Planta 148: 381-388
23. YANG SF 1981Biosynthesisofethyleneand itsregulation.InJFriend,MJC
Rhodes, eds,Recent Advances in theBiochemistryof Fruits andVegetables.
AcademicPress,NewYork,pp89-106
24. YuSM, CH KAo 1980 Antisenescentactivityofsilver ion. Proc Natl Sci Counc ROC 4: 335-337
25. YuY-B, SFYANG 1979Auxin-induced ethylene productionand its inhibition
byaminoethoxyvinylglycineand cobalt ion. PlantPhysiol64: 1074-1077