國立臺中教育大學環境教育及管理研究所
碩士論文
以批次試驗及兩段式串聯反應槽提升
甘蔗渣厭氧產氫之可行性研究
指導教授〆林明瑞
博士
研 究
生〆吳宜曄 撰
中 華 民 國 一 ○ ○ 年 一 月
i
摘
要
傳統化石能源逐漸枯竭以及面臨溫室效應氣體管制的今日。以農業廢棄 物對進行厭氧醱酵產氫是相當重要的生質能源發展技術之一。本研究將利用 每年大量產生的農業廢棄物甘蔗渣作為基質,以了解這類基質厭氧醱酵產氫 的可行性及最佳的產氫操作條件。本研究必先設法提高這類基質的水解效率, 才得以進行後續的醱酵產氫反應,因此本研究由所購得的3株Bacillus subtilis 菌種作為水解菌,再以馴養的醱酵產氫菌做為產氫菌,進行一連串的批次試 驗及水解串連醱酵產氫的二段式反應槽連續流試驗,以評估甘蔗渣之厭氧產 氫可行性。 批次試驗研究結果顯示,由Bacillus substilus取得的3株水解菌中,紅甘 蔗渣COD濃度20,000mg/L,搭配Bacillus subtilis (C),起始pH為7.5時有最好 的水解效果。其溶解性COD增加率為27.2%々加入醱酵產氫菌後,起始pH 為5.25時,有最佳的產氫率為0.83 mmole H2/g-CODin。由Gompertz equation迴歸分析結果,r2 帄均為0.91,顯示迴歸式適合厭氧產氫之模擬。
在連續流兩段式反應槽試驗中,前段 SBR 反應槽之總循序時間為 7 hr、 起始pH 為 7.0 時,有最佳的水解效率,溶解性 COD 增加率為 28.1%々而後 段 CSTR 反應槽之 pH 為 5.25,HRT 為 18hrs 時有最佳每克 COD 及單位體 積反應槽產氫率為2.49 mmole-H2/g-CODin及22.6 mmole-H2/L〃day。在質量
帄衡方面,COD 總帄均回收率介於 93.0%。在微生物動力學模擬方面,各 個試程的r2帄均為0.88,各個試程的試驗在前段 SBR 反應槽基質利用率(P) 值均比後段段反應槽基質利用率(P)值明顯高出許多,由此可顯示前段 SBR 水解反應槽的COD 去除率較後段 CSTR 來的較好。 蔗渣經由兩段式反應槽反應後之產氫率。其總迴歸分別為
ii (1) 每克進流COD產氫率(mmole- H2 / g-CODin)= 有機負荷 (kg-COD / m3 . day)0.292 × 0.748│前段起始pH-7.0│ × 0.552│後段起始pH-5.25 │ (2) 單位體積反應槽產氫率(mmole-H2 /L〃day)= 有機負荷(kg-COD / m3 . day)0.943 × 0.747│前段起始pH-7.0│ × 0.665│後段起始pH-5.25 │ 本研究以水解 SBR 反應槽串聯醱酵產氫 CSTR 反應槽,其產氫量比單 段反應槽之醱酵產氫率較佳,可得知蔗渣基質經由 SBR 水解後,可以有效 提升產氫率,且蔗渣廢棄物取得容易,所以利用蔗渣基質來產氫的可行性 高。 關鍵字〆SBR、CSTR、水解、厭氧醱酵產氫、農業廢棄物、蔗渣
iii
The promoting of hydrogen production on bagasse via
batch tests and series-connected two-stages-reactors
Abstract
Due to the fossil fuel is exhausted gradually and we should face the pressure of eliminating on greenhouse gas now. It is one of the critical biomass energy technologies that is to utilize agricultural wastes to produce hydrogen by anaerobic microorganism. This study aimed at exploring the feasibility of bagasse of the agricultural wastes for fermentative hydrogenesis and to find out the feasibility of producing fermentative hydrogenesis from the bagasse and the conditions for the optimal operation.
First of all, the hydrolization for the substrate must be promoted before the fermentative hydrogenic reaction could be conducted in the study. Therefore, three strains of Bacillus subtilis used as the hydrolytic bacteria together with the fermentative hydrogenic bacteria cultivated by our lab. A series of batch-type tests and 2-stages-reactors continuous input tests were conducted for the hydrolysis and fermentative hydrogenesis.
According to the batch-type tests results, the maximum hydrolization was got with the increase of soluble COD up to 27.2% when Bacillus subtilis(C) among three strains was used as the hydrolytic bacteria, influent COD concentration of bagasse was 20,000mg/L and the initial pH was 7.5. The initial pH of fermentative hydrogenic bacteria being added was 5.25, the best fermentative hydrogen productions were got, 0.83 mmole H2/g-CODin for
bagasse. According to the results of the regression by Gompertz equation, the average of r-square was 0.91, which revealed that the regression Gompertz equation indeed fit the anaerobic hydrogen production of the study.
iv
In the continuous input tests of 2-stages-reactors, when the total progressive time of pre-stage aerobic SBR was 7hrs and the initial pH was 7.0, the best hydrolysis was got with the increase of soluble COD up to 28.1%. And the pH of post-stage CSTR was 5.25, and HRT was 18hrs, the best hydrogen productions in term of per gram influent COD, 2.49 mmole-H2/g-CODin and the hydrogen
production in term of per unit volume reactor, 22.6 mmole-H2/L〃day, were get,
respectively. For mass balance of COD, the average recovery was 93.0%. As to the microbial kinetics simulation, the average of r-square was 0.88, it was shown that substrate utilization rate (P) of pre-stage SBR was obviously higher than that of post-stage CSTR, which indicated that the COD removal of pre-stage SBR was better than that of post-stage CSTR.
The general regression equation of hydrogen production on bagasse via series-connected 2-stages-reactors:
(1) hydrogen production of per gram influent COD (mmole-H2/g-CODin) =
organic loading(kg-COD/m3〃day)0.292 × 0.748│SBR initial pH-7.0│× 0.552│CSTR pH-5.25│ (2) hydrogen production in term of per unit volume reactor (mmole-H2 /L〃day) =
organic loading(kg-COD/m3〃day)0.943×0.747│SBR initial pH-7.0│×0.665│CSTR pH-5.25 │ The hydrogen production of two-stages-reactors in this study was much higher than that of single stage reactor. The bagasse substrate hydrolysis can be effectively enhanced by the SBR to promote hydrogen production and bagasse available can be everywhere. Therefore, bagasse is a feasible solution for hydrogenesis.
Keywords: SBR, CSTR, hydrolysis, anaerobic fermentative hydrogenesis,
v
目錄
摘 要 ... i Abstract ...iii 目錄 ... v 表目錄 ... vii 圖目錄 ... xi 第一章 緒 論 ... 1 第一節 研究背景與動機 ... 1 第二節 研究目的 ... 4 第三節 名詞釋義 ... 5 第二章 文獻回顧 ... 7 第一節 能源危機與生質能源 ... 7 第二節 蔗渣的成份分析及用途 ... 11 第三節 各種裂解纖維素之水解微生物 ... 13 第四節 各種產氫微生物 ... 18 第五節 厭氧醱酵產氫反應機制 ... 21 第六節 厭氧醱酵產氫之影響因子 ... 26 第七節 各種厭氧產氫反應槽之探討 ... 36 第八節 SBR 能源化技術之探討 ... 39 第九節 厭氧醱酵產能反應動力學模式 ... 42 第十一節 厭氧醱酵產氫之模擬 ... 45 第三章 研究方法 ... 47 第一節 試驗材料與設備 ... 47 第二節 試驗設計及流程 ... 53 第三節 分析項目與方法 ... 59vi 第四章 結果與討論 ... 65 第一節 不同蔗渣種類及水解菌種批次試驗 ... 66 第二節 水解菌不同起始 pH 值之水解批次試驗 ... 69 第三節 不同基質 COD 濃度之水解批次試驗 ... 72 第四節 不同起始醱酵產氫pH 之醱酵產氫試驗 ... 74 第五節 水解菌培養液清洗與否對產氫效率之影響 ... 81 第六節 變化前段 SBR 不同總循序時間之產氫試驗 ... 83 第七節 後段 CSTR 不同 HRT 的產氫試驗 ... 87 第八節 前段 SBR 不同起始 pH 對產氫效率之影響... 91 第九節 後段 CSTR 中不同之 pH 對醱酵產氫效率之影響 ... 97 第十節 不同基質 COD 濃度對醱酵產氫效率之影響 ... 101 第十一節 各組試驗產氫率與各項水質分析之迴歸分析 ... 105 第十二節 質量帄衡 ... 106 第十三節 厭氧產氫之模擬 ... 110 第十四節 以蔗渣經厭氧醱酵之產氫效率總迴歸 ... 116 第十五節 不同基質之產氫率比較 ... 118 第五章 結論與建議 ... 121 第一節 結論 ... 121 第二節 建議 ... 123 參考文獻 ... 125
vii
表目錄
表2-1 台灣地區各種能源之供需預測表 ... 8 表2-2 厭氧產氫技術之 SWOT 分析 ... 11 表2-3 甘蔗渣的化學組成分析 ... 12 表2-4 目前已知可分解纖維素之厭氧性微生物 ... 15 表2-5 目前已知可分解纖維素之好氧性微生物 ... 16 表2-6 厭氧產氫微生物之種類 ... 20 表2-7 適合厭氧醱酵產氫之營養物質及濃度 ... 29 表2-8 氨氮濃度對厭氧菌分解之影響 ... 32 表2-9 各種輕離子濃度對厭氧消化之影響 ... 35 表2-10 各種厭氧生物廢水處理技術評估表 ... 38 表3-1 水解菌種培養條件 ... 48 表3-2 各水解菌種之培養基成份及濃度 ... 48 表3-3 厭氧發酵產氫菌訓養成份 ... 48 表3-4 不同蔗渣種類及水解菌種批次試驗中之控制及操作條件 ... 54 表3-5 水解菌不同起始 pH 值之水解試驗中之控制及操作條件 ... 54 表3-6 基質不同 COD 濃度之批次水解試驗中之控制及操作條件 ... 55 表3-7 醱酵產氫菌不同起始 pH 批次試驗中之控制及操作條件 ... 55 表3-8 不同 SBR 程序控制及循環時間試驗中之控制及操作條件 ... 56 表3-9 SBR 反應槽循序時間 ... 56 表3-10 不同 CSTR 程序控制及循環時間試驗中之控制及操作條件 ... 57 表3-11 SBR 不同起始 pH 試驗中之控制及操作條件 ... 57 表3-12 不同 CSTR 中之 pH 醱酵產氫試驗中的控制及操作條件 ... 58 表3-13 進流基質不同 COD 濃度之醱酵產氫試驗的控制及操作條件 ... 58 表4-1 紅、白蔗渣為不同水解菌分解後總 COD、溶解性 COD 變化情形 .. 68 表4-2 紅甘蔗渣基質之水質分析 ... 68viii 表4-3 不同起始 pH 下 Bacillus(C)分解甘蔗渣基質之 pH 變化情形 ... 71 表4-4 批次水解試驗不同起始 pH 值下,反應瓶中 θpH與pH 校正係數 θ .. 72 表4-5 加入醱酵產氫菌,後段厭氧曝氣後,不同起始 pH 批次試驗之氣 體組成及累積產氫量 ... 77 表4-6 後段封瓶使其厭氧狀,不同起始醱酵產氫 pH 之批次試驗的水質 分析 ... 77 表4-7 不同起始醱酵產氫 pH 的批次試驗之總固體物、懸浮固體的變化 情形 ... 79 表4-8 後段不同操作起始 pH 值下,反應瓶中 YpH與pH 校正係數 θ ... 80 表4-8 水解菌培養液是否清洗對前後水質之影響 ... 82 表4-9 水解菌培養液清洗與否對醱酵產氫之氣體產量與成份分析比較 ... 82 表4-10 SBR 不同循序時間對前段 SBR 之溶解性 COD 增加率的影響 ... 84 表4-11 SBR 不同總循序時間對後段 CSTR 之產氣組成及累積產氫量之 影響 ... 84 表4-12 前段 SBR 不同之總循序時間,其前後兩段反應槽之 COD 濃度、 溶解性COD 濃度變化情形... 85 表4-13 前段 SBR 不同總循序時間,其前後段反應槽之揮發酸濃度變化 情形 ... 86 表4-14 前段 SBR 不同之總循序時間,前後段反應槽 TS、SS 濃度變化 情形 ... 87 表4-15 後段 CSTR 不同 HRT,對 CSTR 產氣組成及累積產氫量之影響 ... 88 表4-16 後段 CSTR 不同 HRT,其前後段反應槽之總 COD 及 SCOD 濃度 變化情形 ... 89 表4-17 後段 CSTR 不同 HRT,其前後段反應槽之揮發酸濃度變化情形 ... 90 表4-18 後段 CSTR 不同 HRT,其前後段反應槽之 TS、SS 濃度變化情形 91 表4-19 SBR 不同起始進流水 pH,其前段 SBR 溶解性 COD 增加率 ... 92
ix 表4-20 前段 SBR 不同起始 pH 進流,對後段 CSTR 產氣組成及累積產 氫量之影響 ... 93 表4-21 蔗渣不同操作 pH 值下,反應槽中 YpH與 pH 校正係數θ ... 93 表4-22 蔗渣不同操作 pH 值下,反應槽中 YpH與 pH 校正係數θ ... 94 表4-23 前段 SBR 不同起始 pH,前後兩段反應槽進出流水之總 COD、 SCOD 濃度變化情形 ... 95 表4-24 前段 SBR 反應槽不同起始 pH,其前後段反應槽出流水中之揮發 酸濃度變化情形 ... 95 表4-25 前段 SBR 反應槽不同起始 pH,前後段反應槽之進出流水中 TS、 SS 濃度及去除率之變化情形... 96 表4-26 後段 CSTR 中不同之 pH 對產氣組成及累積產氫量之影響 ... 97 表4-27 蔗渣不同操作 pH 值下,反應槽中 YpH與 pH 校正係數 θ ... 98 表4-28 蔗渣不同操作 pH 值下,反應槽中 YpH與 pH 校正係數 θ ... 98 表4-29 變換後段 CSTR 不同起始 pH,其前後段反應槽之 COD、SCOD 濃度變化情形 ... 99 表4-30 變換後段 CSTR 不同起始 pH,其前後段反應槽之揮酸濃度變化 情形 ... 100 表4-31 後段 CSTR 不同起始 pH,其前後段反應槽 TS、SS 濃度變化情 形 ... 101 表4-32 不同進流基質 COD 濃度對產氣組成及累積產氫量之影響 ... 102 表4-33 不同基質 COD 濃度,其前後兩段反應槽中進出流水之總 COD、 SCOD 濃度變化情形 ... 103 表4-34 不同進流基質 COD 濃度試驗,其前後段反應槽中進出流水之揮 發酸濃度變化情形 ... 104 表4-35 不同進流基質 COD 濃度試驗,其前後段反應槽金出流水之 TS、 SS 濃度變化情形 ... 105
x 表4-36 各組批次水解、產氫試驗 COD 質量帄衡及回收率 ... 108 表4-37 各組連續流試驗之 COD 質量帄衡及回收率 ... 109 表4-38 不同起始 pH 批次產氫試驗 Gompertz equation 迴歸分析結果 ... 111 表4-39 以 Michaelis-Menten 模擬連續流各試驗基質利用率之計算值 ... 113 表4-40 以 Michaelis-Menten 迴歸求得動力學參數 ... 114 表4-41 各組連續流試驗之動力學方程式 ... 115 表4-42 蔗渣不同操作條件,各組試驗每克進流 COD 產氫率之迴歸係數 116 表4-43 蔗渣不同操作條件,各組試驗單位體積反應槽產氫率迴歸係數 .. 117 表4-44 不同農業廢棄物基質產氫之比較 ... 119
xi
圖目錄
圖2-1 微生物藉由 cellulose 附著在纖維素上並進行分解與代謝 ... 17 圖2-2 傳統厭氧消化反應過程 ... 22 圖2-3 厭氧醱酵路徑圖 ... 23 圖2-4 中溫消化與高溫消化之關係圖 ... 30 圖2-5 SBR 每一循環的典型操作程序流程圖 ... 39 圖2-6 菌種利用基質之遲滯期、產能潛勢、與反應速率示意圖 ... 46 圖3-1 批次試驗儀器設備示意圖 ... 51 圖3-2 SBR 反應槽中活性碳棉攪拌架與攪拌馬達 ... 51 圖3-3 二段式 SBR 水解槽串聯 CSTR 醱酵產氫反應槽之示意圖 ... 52 圖4-1 以不同水解菌種分解紅、白蔗渣之總 COD 殘留濃度比例變化情 形示意圖 ... 67 圖4-2 不同水解菌種分解基質之溶解性 COD 濃度變化情形示意圖 ... 67 圖4-3 不同起始 pH,Bacillus(C)分解蔗渣基質之總 COD 濃度變化情形 示意圖 ... 70 圖4-4 不同起始 pH,Bacillus(C)分解蔗渣基質之溶解性 COD 濃度變化 情形示意圖 ... 70 圖4-5 不同 COD 濃度下,甘蔗渣基質之總 COD 濃度的 C/C0變化曲線圖 73 圖4-6 不同 COD 濃度下,蔗渣基質之 SCOD 濃度的 C/C0變化曲線圖 ... 73 圖 4-7 後段不同起始醱酵產氫pH 對醱酵產氫之累積產氫圖 ... 75 圖 4-8 後段封瓶使其厭氧狀,不同起始醱酵產氫 pH 對醱酵產氫之累積 產氫圖 ... 76 圖4-9 不同起始醱酵產氫 pH 批次試驗之揮發酸變化情形 ... 78 圖4-10 後段不同起始醱酵產氫批次試驗中,各 pH 組反應過程中的 pH 與每日產氫量之變化曲線圖 ... 81 圖4-11 前段 SBR 反應槽不同總循序時間其揮發酸產物之組成 ... 86xii 圖4-12 後段 CSTR 反應槽不同 HRT,其揮發酸產物組成 ... 91 圖4-13 後段 CSTR 反應槽不同起始 pH,其前後段反應槽之揮發酸產物 組成 ... 100 圖4-14 SBR 不同總循序時間對產氫效率影響試驗,前段 SBR 反應動力 學迴歸直線 ... 115 圖4-15 SBR 不同總循序時間對產氫效率影響試驗,後段 CSTR 反應動 力學迴歸直線 ... 115
1
第一章
緒 論
蔗渣為原料甘蔗製糖、製燃料酒精時所產生的副產品,經常被視為一種 農業廢棄物。每年約生產70~80餘萬公噸的甘蔗產量,相對的也產出10餘噸 可觀的蔗渣量。近年來,除大部份用作鍋爐燃料外,其餘幾乎全部用於作紙 漿。但蔗渣的成分中含大量的微生物代謝與生長的基質來原-纖維素及半纖 維素,若利用以微生物進行厭氧醱酵產氫,從中獲得可用的能源。本研究先 以批次試驗來先期評估蔗渣用於產氫的可行性,之後將以好氧循序批次反應 槽( Sequencing Batch Reactor,SBR)來進行連續流試驗,以作為產氫相關研 究及後續研究者的參考依據。本章共分三節,第一節首先說明研究背景與動 機々第二節再敘述研究目的々第三節則是本研究的重要名詞釋義。第一節
研究背景與動機
能源是推動國家經濟發展的原動力,而全世界化石能源蘊藏量已逐漸匱 乏,依據現有資料顯示,預估石油可開採41年、天然氣67年、煤192年(曲 新生,2005)。近年來隨著經濟的發展,全球能源需求快速增加,導致化石 燃料的價格上漲,特別是石油價格飆漲々雖然目前化石燃料的開採量有逐年 下降的趨勢,卻仍佔總能源供應量的大部分,且燃燒化石燃料更會造成酸雨、 空氣污染及溫室效應等現象(王啟川,2007)。美國能源部能源署(Energy Information Administration, EIA)在2007年發表的「國際能源概觀」中也指 出,在未來的油價已無回跌的可能性。台灣自產能源有限,能源進口依存度 高達99.1%,因此基於上述原因,有必要發展一種乾淨、無污染且具有自主 性的再生能源。行政院於91年8月所通過的「再生能源發展條例(草案)」,對再生能源 定義為太陽能、生質能、地熱、海洋能、風力、水力(不含抽蓄水力)或其
2
他經中央主管機關認定可永續利用之能源。而在眾多再生能源技術中,生質 能是國際公認最被廣泛使用的能源,約占世界所有再生能源應用的三分之二, 因為其最大的優點除了可產生能源外,並且可利用及去除有機污染物(曾智 鉉,2007)。依據美國能源部能源署於2006年所做之全球初級能源消費 (Primary energy consumption)統計〆屬於太陽能、生質能、風力、地熱與 水力等再生能源者,共計37.245×1015BTU,佔全球初級能源消費472.274× 1015BTU之7.88%,略高於核能之6.49%,至於台灣再生能源則僅為1.56%, 遠低於石油、煤炭、核能與天然氣之使用,顯示台灣在再生能源之發展還有 很大之成長空間。生質能源轉化及生產技術方面主要有〆燃料酒精、生質柴 油以及生質氣體等三部份,在燃料酒精方面,目前農委會規劃以糖份或澱粉 質高的農業作物作為主要物料來源,以澱粉酵母或酵素將澱粉質轉化為酒精, 再以純化分離技術生產燃料酒精々在生質柴油部分,以高油份豆類(如大豆、 油菜籽…等炸油)加以轉酯化為生質柴油々而第三部份則是生質氣體(蘇忠 振,2006)。就生質能源開發技術中,以生質氣體-利用微生物進行厭氧醱 酵產能極具發展潛力,因為不僅能產能且能去除有機廢棄物達到資源化目的。 目前主要可回收使用的生質氣體有氫氣及甲烷々在產氫方面,以厭氧醱酵產 氫技術最具有發展潛力。目前多以使用有機污染物(如有機廢水、污泥、稻 桿、蔗渣等)再利用來產能的技術被視為最值得重視及發展潛力的生質能源 技術。 根據2005年6月20日全國能源會議之決議,政府的能源政策必頇從能源 自主性、農業、環保、經濟及水資源等層面做通盤考慮,會議中擬定以汽油 年耗量1,000萬公秉估算,計畫在2010年推動E5酒精汽油(即添加5%酒精) 所需之生質酒精量至少達50萬公秉以上。在台灣,生質酒精的原料主要是以 甘蔗為主,台糖公司則預估未來帄均單位甘蔗產量為80噸/公頃,屆時甘蔗 產量可達150萬噸之多(鄭作林、蕭耀基,2006),相對到時也將有百萬多
3 噸的蔗渣產生。蔗渣為甘蔗於製糖、製燃料酒精過程中經壓榨機壓榨後所得 的農業廢棄物,通常作為鍋爐燃料使用,少部份可做成養雞、養猪飼料。在 工業上亦有多種用途,如製造隔音絕熱板、家畜床墊、用紙之製造纖維版及 圕合板等,由此顯示蔗渣的利用非常廣泛。蔗渣的成分中含大量的纖維素及 半纖維素,還有少部分的木質素、糖分、碳水化合物等等,這些都是微生物 生長與代謝的重要基質來源,若能再利用以微生物進行厭氧醱酵產能,本研 究推估將有利於厭氧醱酵反應進行提高回收生質氣體的產量。 能源作物再利用的殘餘物如果可仿效其他農業廢棄物如稻桿、米糠、廚 餘等,可經由水解、醱酵產氫反應過程產能回收能源,例如曾智鉉(2006) 研究所使用酒廠廢酒糟及污水處理廠廢棄物污泥作為混合基質最佳的產能 效率有811 cal/ g-COD々郭倍甄(2007)研究中所使用米糠及污水處理廠廢棄物 污泥作為混合基質最佳產能效率有1,526 cal/g-COD々黃倩毓(2008)的研究 中,在最佳pH控制條件下,油菜籽粕為基質有最佳產氫率為1.13 mmole H2/g-CODin,以油菜為基質其最佳產氫率為0.432 mmole H2/g-CODin。而能源
作物利用後的全株及殘餘物再利用也是越來越普遍,因含大量的纖維素 (cellulose)、半纖維素(hemicellulose)及木質素(lignocellulose)等成分, 相當難以分解,所以利用能夠分解纖維素的菌種如: Clostridium、Bacillus及 Geobacillus等菌屬的特定菌株々在厭氧狀態下,利用Clostridium為產氫菌種 搭配純種水解菌種(兼氣菌)組成反應菌液,以能源作物為基質,於兼氣的環 境下經水解菌及Clostridium的水解、醱酵酸化反應過程得以回收H2能源(黃 倩毓,2008)。 黃介辰(2007)的研究指出 Bacillus 與 Clostridium 共培養下,氫氣為分 解纖維素反應中一項重要的生物性指標,因纖維素經由生物方式的水解轉化, 可變成氫氣和其他有機物如醣類、有機酸及醇類。而由共培養實驗結果發現, Bacillus 的存在可使產氫遲滯期明顯縮短,遲滯期最多可自 3~4 天縮短至 1~2
4 天,且可促進之後 Clostridium 將纖維素轉變成氫氣的作用。藉由實驗反應 後的溶解性COD 發現,纖維素分解率最高約可達 23%,且共培養下此外能 自分解纖維素的反應中提升氫氣的產生量。甘蔗渣為固形物,相當難以分解, 必頇以不穩態水解反應來處理,所以採用水解 SBR 反應槽來進行水解分解 以進行厭氧產氫試驗。另外葉明泰(2006)的研究中發現,在反應槽中,氣 密馬達攪拌支架上若密集懸掛活性碳棉作為固定化生物菌體之介質,則可以 有效蓄留菌體於反應槽中,提升水解、醱酵產氫之效果。故本試驗將先以批 次試驗的方式,評估產氫試驗中之可行性以及何種最佳操作條件。再以批次 試驗的結果,利用固定化技術以不穩態操作分離式反應槽(前段水解 SBR +後段醱酵產氫CSTR 反應槽)進行連續流試驗,求得最佳的操作條件,並 進行厭氧生物產能反應之動力學模擬以研究推估至工程上的可行性。
第二節
研究目的
本研究是用蔗渣為主要基質,以新竹食品工業發展研究所生物資源保存 及研究中心所購買的三株水解菌種,搭配台中市黎明污水處理廠終沉池污泥 所馴養的產氫菌,與蔗渣基質進行產氫批次試驗,探討評估在何種操作條件 下,產氫試驗有最佳結果。再以批次試驗的最佳條件,利用固定化技術以不 穩態操作分離式反應槽(前段水解SBR+後段醱酵產氫CSTR反應槽),進行 連續流反應槽來進行厭氧產氫試驗,以評估在何種操作條件下可以得到最佳 的產氫效率。 本研究之目的如下〆 一、以批次試驗評估蔗渣基質在不同操作條件下(水解菌種、水解起始pH、 COD濃度、醱酵產氫之pH)瞭解最佳的水解、產氫效率以及操作條件 為何。5 二、以批次試驗結果為基礎,進行以二段式反應槽 ( 前段水解SBR反應槽+ 後段醱酵產氫CSTR反應槽) 進行前後段不同之循序時間、HRT、pH、 COD濃度變化的連續流產氫試驗,以瞭解最佳操作條件及最佳產氫效 率為何。 三、藉由反應動力學方程式的模擬,以瞭解微生物生長及產氣的動力參數。
第三節
名詞釋義
一、生質能源(biomass energy) 我國將生質能定義為國內農林植物、一般廢棄物與一般事業廢棄物等直 接利用或經生物處理過程所產生之能源,即為「生質能源」(再生能源發展 條例草案,2002)。生質能是一種新能源,與風能、太陽能一樣具有取之不 盡、用之不竭的特性々與其他新能源比較,生質能因使用材料大多為廢棄物, 故兼具廢棄物的回收處理與能源生產的雙重效益(台電,2009)。 二、厭氧醱酵產氫反應 一般傳統厭氧消化反應可分為三階段,即水解、酸化、及甲烷化三階段, 厭氧醱酵產氫反應則發生在水解及酸化階段,也就是在傳統厭氧反應中設法 終止後面的甲烷化反應,並促使前面的水解產酸反應持續進行,以利於氫氣 的產生。醱酵產氫是利用特殊微生物及酵素,在水體環境中將複雜有機物水 解酸化,分解出中間產物氫氣,其原理為以有機物為電子提供者,經由醱酵 作用將有機質分解,伴隨產生的部份電子則藉由特定之電子傳遞系統與產氫 酵素,將電子傳遞給水體中的質子(H+)而產生氫氣(Sawyer, McCarty, Parkin, 1994)。6
三、循序批次反應槽(Sequencing Batch Reactor, SBR)
SBR法為廢水的生物處理程序,屬活性污泥法的範疇,而污泥物去除的 機制、BOD 負荷等參數均適合於SBR循序批次活性污泥法有、厭氣-好氣操 作、間斷進流的特點。而SBR法是一種半連續-間歇式反應槽,它與傳統的 曝氣池不同。進流方式可以是間歇式,也可以是連續式,基質進入反應槽後, 可曝氣、攪拌與生物污泥反應,或不曝氣僅攪拌與生物污泥反應,當反應完 後之放流階段一般是採間歇式或一次式抽離用過的基質廢水。SBR法在單一 反應槽中以時間為導向,控制每一個批次循環操作的廢水生物處理程序,此 操作程序介於傳統批次與連續操作之間,不但能連續操作馴養污泥,更能有 效截留菌體量(周家弘,2001)。 四、批次實驗(batch-type test) 批次試驗是指在一個密閉系統及有限的培養基中,植入微生物菌種進行 培養,使微生物生長繁殖,在特定的條件下只完成一個生長周期的微生物培 養方法,且培養系統沒有與外界有物質交換作用。批次培養過程中,隨著培 養基的營養物質的不斷減少,微生物的生長環境也隨之不斷變化,因此,微 生物批次培養是一種非穩定狀態的培養方法。在批次培養過程 ,隨著微生 物細胞和代謝物濃度的不斷變化,微生物生長可分為停滯期、對數生長期、 穩定期和衰退期四個階段(林逸群,2003)。
7
第二章
文獻回顧
由於目前能源短缺,生質能源將逐漸成為世界各國能源技術發展及應用 的主流々利用微生物來產能最具有發展潛力,是生質能源的重要發展技術之 一。本章將以能源危機與生質能源、蔗渣的成份分析及用途、水解能源作物 纖維素相關微生物、各種產氣微生物、厭氧醱酵產氫反應機制、各種厭氧產 氫反應槽之探討、SBR能源化技術探討、細胞固定化技術、厭氧醱酵產能之 影響因子、厭氧醱酵產氫反應動力學模式等十一節進行回顧探討。第一節
能源危機與生質能源
一、能源危機 化石燃料為人類目前主要的使用能源,而化石燃料主要有煤炭、石油及 天然氣等三項,估計占全球現在使用能源總量的百分之八十五以上,其中石 油是最具代表性的化石能源(李育明,2007)。在台灣有99.1%的能源需仰賴 進口,從表2-1台灣地區能源供需預測表中可以發現,雖然預估化石燃料的 需求會降低々另外也可以發現,再生能源在預定的目標中是被看好逐年成長 的,但化石燃料仍佔總供應量的80~90﹪以上。 人類的生活離不開石油,舉凡人造圕膠、產業需求、交通需求……等都 以其為原料。然而石油為不可再生的能源,在1980年代兩次石油能源危機, 以及2008年的金融海嘯也使的原油價格飆漲,造成全球性的影響,對經濟展 產有重大的衝擊及變化。根據英國石油公司「2008年版世界能源統計要覽」, 在技術與成本的限制下,預估世界石油蘊藏量只能再開採40年,天然氣可再 開採60年,煤炭可再開採133年(British Petroleum, 2008)。全球經濟仍持續 的發展,以目前石油之消耗速率,若無法取得大量、價廉之替代能源,能源 危機遲早會再度來臨。8 二、化石燃料對環境的衝擊 人類的經濟活動大量使用化石能源,雖然帶來便利,卻也讓大氣中CO2 等溫室氣體濃度急速增加,全球產生越來越明顯的增溫現象。由於全球氣溫 的急遽暖化,使得自然環境的氣候和生態、以及人文環境的社會和經濟都受 到了影響。根據IPCC(IPCC, 2007)第四次評估報告指出,全球氣候的改變 會導致的環境問題與影響可由三方面來看〆全球帄均溫度上升、海帄面上升 及異常氣候頻率增加。另外,除了溫室效應外,化石燃料之大量使用,燃燒 過程中產生SOx、NOx排放至大氣環境中,經氧化反應生成硫酸、硝酸等酸 性物質使得雨水之pH值降低,形成酸雨々還有燃煤產生的落塵也都會造成空 氣的污染,影響我們的健康。 三、再生能源 因應「京都議定書」生效後之CO2減量要求,94年6月21日「全國能源 會議」建議政府再生能源發電推廣目標,至2025年再生能源發電量應占5~ 7%。在「能源發展條例(草案)」中,對再生能源種類分為〆太陽能、生質 能、地熱、海洋能、風力、水力或其他經中央主管機關認定可永續利用之能 源,其中以生質能之利用最為廣泛(台灣電力公司,2009)。 表2-1 台灣地區各種能源之供需預測表 項目 2004年 2010年 2020年 2004年~ 2020年 百萬公秉油當 量MKLOE % 百萬公秉油當量 MKLOE % 百萬公秉油當 量MKLOE % 年帄成長率 煤炭 43.5 34 44.2 30 64.3 37 2.1 石油 63.5 49 63.8 43 56.3 32 -0.5 天然氣與液 化石油氣 10.7 9 18.9 12 29.7 17 6.7 其他 10.8 8 15.3 11 12.9 8 0.7 再生能源 0.6 0 5.0 4 10.7 6 17.4 資料來源〆行政院經濟部能源局(2009)
9 四、生質能源 生質能(Biomass energy)泛指由微生物所產生之有機物質,如農業、 畜牧業、工業、都市廢棄物以及能源作物,經過焚化、氣化、裂解、醱酵等 技術轉換成燃油(燃料酒精、生質柴油)、燃氣(甲烷、氫氣)與電力等可 用之能源,即係指利用生物質(biomass),經過轉換所獲得的可用能源(工 研院能資所,2009)。而根據國際能源總署(2006)的統計,目前生質能是 全球第四大能源,僅次於石油、煤及天然氣。生質能供應全球約14%的初級 能源需求,約佔世界所有再生能源應用的80%,是目前最廣泛使用的再生能 源。與其他再生能源比較,生質能的優勢包括技術較成熟、有商業化運轉能 力、可併用在傳統能源供應的架構、且因可使用廢棄物作為燃料,故還可兼 具廢棄物的回收處理與能源生產的雙重效益,由此可以得知生質能具有相當 大的發展潛力。 生質能利用的技術範圍相當廣泛,其轉換為能源的方式可概分為直接燃 燒技術、物理轉換技術、熱轉換技術與化學/生物轉換技術,其說明如下(吳 耿東、李宏台等,2004)〆 1.直接燃燒技術〆把廢棄物直接燃燒以產生熱能與電力,例如在大型垃圾焚 化廠中,以焚化垃圾發電。 2.物理轉換技術〆將廢棄物經破碎、分選、乾燥、混合添加劑及成型等過程, 製成易於運輸及儲存的固態衍生燃料,作為鍋爐、水泥窯的燃料,例如紙 廠把廢棄物製成錠型的固態燃料,作為燃煤鍋爐的輔助燃料。 3.熱轉換技術〆利用氣化與裂解(液化)等熱轉換程序,將廢棄物合成燃油 或燃氣(瓦斯),作為燃燒與發電設備的燃料。例如稻殼、能源作物或廢 紙渣可產製合成燃氣,進行燃氣發電。 4.化學/生物轉換技術,是指經醱酵、轉酯化等生物化學轉換程序以產生沼 氣、燃料酒精、生質柴油、氫氣等,作為引擎、發電機與燃料電池的燃料。
10 例如甘蔗、玉米經糖化醱酵後產製燃料酒精々又如廢食用油經轉酯化反應 可產製生質柴油,這些生質能都可作為汽車的替代燃料等。 目前燃料酒精、生質柴油、生質氣體這三種是最具市場發展潛力。燃料 酒精主要採用醱酵法生產酒精。除了巴西自 1970 年代石油危機起,以甘蔗 為主要能源作物從事酒精燃料之研發與生產外,另一主要國家為美國利用玉 米澱粉來醱酵以生產酒精。而以廢棄農作物來生產酒精最具潛力,廢棄農作 物含有相當量的纖維素,目前將纖維素由生質轉化成糖的技術一般可分為酸 水解法與酵素水解法兩種。酵素水解法已有商業化規模,可以用於一般農業 廢棄物如麥桿、玉米桿和硬木之處理。 利用油脂作物或廢食用油與甲醇(或乙醇)進行轉酯化反應,可產生脂 肪酸甲酯(或乙酯)及甘油等產物々經分離甘油後,以蒸餾去除未反應完全 的油脂,產生與一般柴油品質相當的液態燃料,稱為生質柴油。由於使用生 質柴油的引擎排氣不含鉛、二氧化硫、鹵化物,並能大幅降低碳煙、硫化物、 未燃碳氫化合物、一氧化碳及二氧化碳,目前已成為世界各國積極發展的生 質能。生質柴油可直接作為柴油的替代燃料,或以不同比例摻配於市售柴油 中(一般建議摻配20%)。 生質氣體的發展是相當被看好。目前主要回收的生質氣體 (biogas)有氫 氣、甲烷。氫氣是屬於乾淨的能源之一,以 Clostridium 等兼氣菌為主的醱 酵產氫技術最具有發展潛力,其產能效率每克的氫氣中約有122 千焦耳,約 為石油的3 倍,甲烷的 2.4 倍,且燃燒過後只產生能量和水,不會形成溫室 氣體二氧化碳而造成環境污染,因此氫氣是未來重要能源之一(毛宗強, 2008)。在基質利用方面,最有經濟且有開發效益,就是以有機廢水、廢棄 活性污泥、農業廢棄物、食品、酒類等廢棄物為基質,達到不僅產能且能去 除有機廢棄物達到資源化目的。經濟部能源局針對台灣的發展生質氣體的情
11 況,表 2-2 為產氫技術之 SWOT 分析,包括〆產氫技術在應用及商業運轉 時,所可能遭遇之優勢(Strength)、弱勢(Weakness)、機會(Opportunity)及威 脅(Threat)。 表2-2 厭氧產氫技術之 SWOT 分析 技術 厭氧醱酵/光合作用產氫技術 優勢 (Strength) *無污染。 *於常溫常壓下操作並不需提供額外能源,可消化廢棄 物以減少環境污染。 *氫氣熱值相當高。 *技術居於國際領先地位 弱勢 (Weakness) *技術仍在研發階段,未進入實用化階段,尚未有任何 實廠化案例。 機會 (Opportunity) *能源工業中氫氣是極好的傳熱載體。 *氫能可利用形式眾多。 *物料來源豐富。 *因應燃料電池動力的普及,氫能市場將有極大幅度的 擴張。 *台灣已有學術單位投入生物產氫的基礎研究,在基礎 技術開發上,台灣不遜於各先進國家。 威脅 (Threat) *日本、歐盟、美國等國家皆已投入大量經費進行全面 整合性之研究。 *各先進國家多已進入技術實用化開發階段,開始進行 大規模戶外生物產氫程序示範。受限於學術界研發規 模有限,台灣在生物產氫實用化上是較為落後的。 資料來源〆經濟部能源局(2009)
第二節
蔗渣的成份分析及用途
甘蔗為禾木科的多年生植物,適合於熱帶和亞熱帶繁殖,一直以來都是 台灣特用作物產量最高之一種作物,甘蔗之用途可分為生食用甘蔗和原料用 甘蔗。其中,生食用甘蔗在2001年時總產量曾高達260,491公噸,原料用甘 蔗之總產量更高達4,536,231公噸,爾後雖然逐年下降,仍有近上百萬噸的產 量。依據農業委員會97年之農業統計年報,2008年台灣製糖甘蔗產量為12 707,102公噸,生食用之甘蔗產量為58,400公噸(行政院農業委員會,2009)。 利用甘蔗榨汁所得之糖蜜(molasses)來進行醱酵產製酒精早已行之年,總產 量在2004 年已到達3811萬公秉且持續增加中,目前主要的生產國為巴西 (39.6%)、美國(35.1 %)、中國(9.5 %)與印度(4.5 %),可見世界各國對於利用 酒精來取代汽油都是抱持相當樂觀的態度 (RFA,2005) 。在台灣,預計在 2010年推動E5酒精汽油(即添加5%酒精)所需之生質酒精量至少達50萬公 秉以上,台糖公司則預估未來帄均單位甘蔗產量為80噸/公頃,屆時甘蔗產 量可達150萬噸之多(鄭作林、蕭耀基,2006)。 蔗渣係指甘蔗搾汁完畢後的殘留物,目前國內的甘蔗渣,是為製糖過 程產生的廢棄物,其廢棄物若不適時處理會帶來環境污染,以往多用在製 糖廠本身的燃料,或加工成建築用的隔音隔熱板與阻燃材料,也有作成堆 肥施於田園中、作成蔗渣紙漿或製作阻燃石膏纖維版等等。但以能源的觀 點來看,蔗渣是有再度被利用而用於產氫的可能性。蔗渣的基本性質如表 2-3所示,主要是以纖維素、半纖維素與木質素為主。這些元素對於微生物 生長與代謝是一重要基質來源,有利於生化反應之進行。藉由微生物的作 用,使蔗渣有利於轉化成能源,因此非常適合作為厭氧產能的基質來源之 一。 表2-3 甘蔗渣的化學組成分析 成份項目 紅蔗渣含量(%) 白甘蔗渣含量(%) 水份 7.08 1.3 纖維素 33.28 36.1 半纖維素 22.58 25.4 木質素 6.15 4.7 糖份 24.69 30.4 灰份 4.22 2.1 蛋白質 2.00 -
13 因富含大量的纖維素、半纖維素及木質素等成分,根據李傳華(2003) 的研究指出利 用 水 解 技 術 , 以甘 蔗 渣 、 樹 葉 和 菜 皮 三 種 物 質 為 材 料 , 並 加 入 催 化 劑 Na2CO3 的 條 件 下,上 述 廢 棄 物 可 轉 化 得 到 含 有 大 量 的 腐 植 酸 物 質,最 高 能 達到45%左右,而 這 類腐 植 酸 物 質 可 以 作為 生 態 肥 料,具 有 良 好 的 肥 效 和 經 濟 價 值。而 臧金燦、樊國燕(2003)研究中 指出,稻桿的鹼化處理可使植物細胞壁變得鬆散,易於分解處理。因此若能 選擇適當的前處理技術,將可有效分解像纖維素等難以分解有機物質,而經 分解後之有機物可經由後段的能源化技術來產生能源,使能源作物之產能效 益提升。許超傑(2006)以厭氧程序轉換蔗渣為生質能源之研究中指出,當 蔗渣被微生物水解,所得的揮發酸作為鹼性燃料電池基質的測試中顯示厭氧 醱酵的酸液確實可以發電々本研究利用甘蔗渣主要是探討其木質纖維素經水 解醱酵後的產能之效率。
第三節
各種裂解纖維素之水解微生物
纖維素不溶於水且架構穩定難以分解。一般植物成份內不僅只含纖維素, 還包含半纖維素及木質素等主要成分,因此利用植物原料產生氫氣的過程中 需先適當的前處理方法處理半纖維素及木質素後,再進行纖維素水解醣化及 醱酵作用,最後產生氫氣。而纖維素酵素水解是經由纖維素分解微生物,將 不溶解性纖維素轉換成可溶解性醣類(主要醣類為纖維雙醣及葡萄糖)。圖 2-1為微生物藉由cellulose附著在纖維素上並進行分解與代謝的過程。在自然 界中可以分泌纖維水解脢之微生物,包括細菌與真菌(Fungi)。這些微生 物可能是好氧的(Aerobic)或厭氧的(Anarerobic)、中溫的(Mesophilic) 或高溫的(Thermophilic)々表2-4、2-5為纖維素水解微生物種類。不同微生 物合成之纖維水解脢組成結構不同,對纖維素之水解能力也不同。細菌為單 細胞生物,能獨立生存與增殖,其中可產製屬纖維水解脢者,包括Clostridium、
14
Cellulomonas、Bacillus、Thermomonospora、Ruminococcus、Bacteriodes、
Erwinia、Acetovibrio、Microbispora與Streptomyces。目前,以Cellulomonas fimi
與Thermomonospora fusca之研究與應用最多。雖然可水解纖維素之細菌,特
別是厭氧菌如Clostridium thermocellum 與Bacteroides cellulosolvens所產製
之纖維水解脢具有較高之活性(Duff & Murray, 1996々Sun & Cheng, 2002), 其生長環境,Clostridium thermocellum JW20可生長在纖維素、木質素中,而 Clostridium thermocellum生長在纖維素中的一個特徵是會產生一種黃色色素
物質(Ljungdahll & Erikssonk, 1985)。除此之外,亦可生長在葡萄糖、果糖、 木糖中。在最適生長環境方面,pH約介於6.1-7.5之間,溫度為28-69℃間, 而當以纖維素及半纖維素為基質時,其生長的適應時間分別為6.5 hrs及2.5 hrs。而Clostridium cellulolyticum在pH 7.5、溫度35℃時,有最佳的生長狀況。 王馨怡(2006)的研究中指出乳牛所排放出之糞便中,培養出能降解纖維 素產生乙醇的嗜熱厭氧穩定菌群。研究菌群以濾紙為纖維素基質,其最佳乙 醇生成溫度55-60℃左右及最佳pH 7.7,乙醇的產率約為0.29g ethanol/g cellulose。黃倩毓(2008)以油菜及油菜籽粕為基質時,都在水解菌/醱酵產 氫菌配比均為1/4,也有最佳的產氫率,以油菜組為基質產氫率為0.441 mmole H2/g-CODin 々以油菜籽粕組為基質之產氫率為1.10 mmole H2/g-CODin。
由以上文獻可以得知,利用水解纖維素的微生物菌種,可以得到較好的 纖維素水解,本研究將嘗試以不同水解菌與蔗渣基質以了解何種操作條件有 較佳水解效果及產氫效率 。
15
表2-4 目前已知可分解纖維素之厭氧性微生物 Anaerobes / Facultative aerobes
Acetivibrio cellulolyticus
Bacteroides (Fibrobacter) succinogenes Bacteroides cellulosolvens Bacteroides xylanolyticus Butyrivibrio fibrisolvens Caldocellum saccharolyticum Clostridium acetobutylicum Clostridium cellobioparum Clostridium cellulolyticum Clostridium cellulovorans Clostridium josui Clostridium papyrosolvens Clostridium stercorarium Clostridium thermocellum Clostridium thermocopriae Erwinia carotovora Erwinia chrysanthemi
Eubacterium (cillobacterium) cellulosolvens Micromonospora propionici a Micromonospora ruminantium a Pseudonocardia thermophila Ruminococcus albus Ruminococcus flavefaciens Thermoanaerobacter cellulolyticus
a No longer recognized as true members of genus, or by this name or by inclusion in
Bergey’s Manual (1984-1989).
16
表2-5 目前已知可分解纖維素之好氧性微生物 Aerobes / Facultative anaerobes
Acidothermus cellulolyticus Cytophaga johnsonae
Actinoplanes brasiliensis Cytophaga krzemieniewskae a Actinoplanes phillipinensis Cytophaga pectinovora Actinopolyspora halophila Cytophaga rubra a Archangium spp. Cytophaga tenuissima a Arthrobacter spp. Cytophaga winogradskii a Bacillus brevis Frankia sp
Bacillus cereus Herpetosiphon geysericolus Bacillus coagulans Microbispora bispora
Bacillus firmus Micromonospora aurantiaca a Bacillus licheniformis Micromonospora chalcae Bacillus pasteurii Micromonospora inositola Bacillus polymyxa Micromonospora melanosporea Bacillus pumilus Micromonospora olivasterospora Bacillus stearothermophilus Polyangium spp.
Bacillus substilus Pseudomonas fluorescens var. cellulosa Cellulomonas biazotea Pseudomonas solanacearum
Cellulomonas cellulans (formerly C. cartae) Serratia marcescens Cellulomonas cellasea Sorangium spp.
Cellulomonas fermentans a Sporocytophaga myxococcoides Cellulomonas fimi Staphylococcus saprophyticus Cellulomonas flavigena Streptomyces albogriseolus Cellulomonas gelida Streptomyces celluloflavus Cellulomonas turbata a Streptomyces cellulosae Cellulomonas uda Streptomyces flavogriseus Cellvibrio flavescens a Streptomyces lividans Cellvibrio fulvus a Streptomyces nitrosporeus Cellvibrio gilvus a Streptomyces olivochromogenes Cellvibrio mixtus a Streptomyces thermodiastaticus Cellvibrio ochraeus a Streptomyces thermovulgaris Cellvibrio viridus a Streptomyces viridosporus Cellvibrio vulgaris a Thermoactinomyces cellulosae Corynebacterium michiganense Thermomonospora curvata Cytophaga aprica Thermomonospora fusca Cytophaga aquatilis Xanthomonas albilineans Cytophaga aurantiaca Xanthomonas campestris Cytophaga hutchinsonii
a No longer recognized as true members of genus, or by this name or by inclusion in Bergey’s
Manual (1984-1989).
17
圖2-1 微生物藉由 cellulose 附著在纖維素上並進行分解與代謝 (Haigler & Weimer, 1991)
18
第四節
各種產氫微生物
本研究為評估以甘蔗渣為基質用於醱酵產氫的可行性,在本研究試程中 氫氣的產生主要是厭氧醱酵菌在厭氧反應中所產生的,因此必頇瞭解醱酵產 氫菌的生長特性,才能有效發揮微生物產能的作用。 二十世紀初,外國學者Toerien、Iannotti等人曾利用Vibrio succinogenes 菌株,將葡萄糖等碳水化合物,於厭氧環境下,醱酵產生氫氣(毛宗強,2008)。 因此,由過去許多的研究中發現,有很多微生物均具有產氫能力,如表2-6 (Das & Veziroğlu, 2001)。厭氧產氫微生物種類繁多,其生理特性與產氫機制 皆不同,早期,C. T. Gray and H. Gest 將產氫微生物分為四類(Twarog, 1965)〆 1.不含細胞色素系統的厭氧異體滋養菌(Heterotrophs),例如梭狀芽孢桿菌 屬(Clostridium)、細球菌屬(Micrococci)、甲烷菌(Methanobacteria)等。 2.含細胞色素的異體滋養的兼性厭氧菌(Facultative Anaerobics),如大腸埃 希氏大腸菌(Escherichia Coli)、脾桿菌(Enterobactor)。 3. 含 細 胞 色 素 的 厭 氧 異 體 滋 養 菌 , 如 脫 硫 弧 菌 屬 (Desulforibrio desulfricans)。 4.可在光線照射下還原的NADH 產生氫氣的光合菌,例如含硫紫菌(Sulfur purple bacteria)、不含硫紫菌厭氧適應性藻類等。 近年來的研究則是把分類做了改變,根據 Yokoi、Ohkawara、Hirose、 Hayashi and Takasaki(1995)的研究指出,產氫菌主要分為光合作用微生物 (Photosynthetic microorganisms) 和 醱酵 產 氫 微生 物 (Fermentative hydrogen producing microorganisms)二大類。光合作用微生物部份包括〆紫色非硫菌、 藻類等。醱酵產氫微生物又依其對氧需求性分為兼性厭氧菌(如 Enterobacter19
aerogenes 等)及絕對厭氧菌( 如 Clostridium 等)。醱酵產氫微生物中以 Clostridium 菌屬為主,此菌屬大部分都可以產氫,其中以 Clostridium butyricum 菌種的產氫效率為較佳(Yokoi, 1997々Odom, 1983)。本研究將
以蔗渣為基質,先以水解菌水解纖維素,再以 Clostridium 菌種進行醱酵產 氫。 Clostridium菌屬為目前已知產氫菌中產氫能力較佳的菌種, 由學者 Prazmoski在1880年發現々此菌屬適合生長於厭氧、中溫(30-37℃)及中性pH 的環境下(Minton, 1989),在較不適合生長的環境下會產生內孢子而進入休眠 的狀態。Clostridium的孢子屬內孢子菌(endospores),產生於菌體內為此菌屬 特有之結構,每一株菌體可生成一個孢子,孢子的特徵為位於菌體的末端, 一般較菌體寬,與其它產胞菌較小且位於菌體中央不同(李國鏞、游若荻, 1992)。Clostridium的產氫能力已廣受注目,主要是因為針對混合產氫系統的 菌種進行鑑定結果發現〆主要的菌群是以Clostridium為主(Ueno, Haruta, Ishii
& Igarashi, 2001々Fang, Zhang & Liu, 2002a)。且此菌對有機物的利用亦相 當廣泛,但Clostridium屬的細菌之高產氫能力,常對有機物的分解利用具專 一性 (曾怡禎,2000),如食品工廠、啤酒製造廠等具有高糖份、高澱粉、易 酸化、水解之高濃度有機廢水。 因此,若在厭氧甲烷化系統中,提供一個較為惡劣的環境,如加入強酸、 強鹼、加熱煮沸等方式以殺死甲烷菌並保留已經形成孢子的Clostridum,藉 此達到菌種篩選之目的,以利於進行之後的厭氧生物產氫程序。根據葉明泰 (2006 ) 的 研 究 中 發 現 黎 明 污 水 處 理 廠 終 沉 池 底 泥 中 含 有 相 當 量 之 Clostridium 醱酵產氫菌。所以本研究在連續流試驗中所使用污水處理廠終 沈池底層污泥,經由熱篩、酸篩將甲烷形成菌篩除或抑制,並維持適合生長 環境進行馴養,做為本試驗發酵產氫菌來源。
20
表2-6 厭氧產氫微生物之種類
分類 微生物名稱
Green algae Scenedesmus obliquus Chlamydomonas reinhardii
Chlamydomonas movewusii Cynobacteria Heterocystous Anabaena azollae Anabaena CA Anabaena variabilis Anabaena cylindrical Nostoc muscorum Nostoc spongiaeforme Westiellopsis prolifica Non-heterocystous Plectonema boryanum Oscillotoria Miami BG7 Oscillotoria limnetica Synechococcus sp. Alphanothece halophytico Mastidocladus Phormidium valderianum Photosynthetic bacteria Rhodobacter sphaeroides Rhodobacter capsulatus Rhodobacter sulidophilus Rhodopseudomonas sphaeroides Rhodopseudomonas palustris Rhodopseudomonas capsulata Rhodospirillum rebnum Chromatium sp. Miami PSB 1071 Chlorobium limicola Chloroflexu aurantiacus Thiocapsa roseopersicina Halobacterium halobium Fermentative bacteria Enterobacter aerogenes Enterobacter cloacae Clostridium butyricum Clostridium pasteurianum Desulfovibrio vulgaris Magashaera elsdenii Citrobacter intermedius Escherichia coli
21
第五節
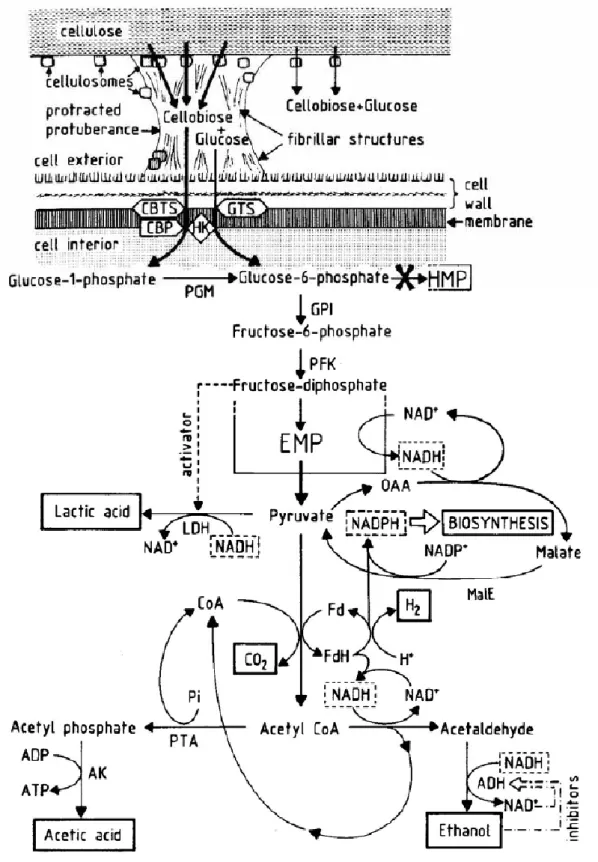
厭氧醱酵產氫反應機制
控制厭氧微生物生化反應在適合產氫的途徑,以獲得較大的產氫量,是 重要的關鍵之一。一般厭氧醱酵指在厭氧環境下,利用厭氧微生物之代謝作 用將高分子有機物分解轉換為甲烷和二氧化碳等最終產物的過程々此程序可 分為水解(Hydrolysis)、酸化(Acidogenesis)及甲烷化(Methanogenesis)三個階 段(劉安琪,1996),流程如圖2-2所示,各階段的主要微生物都有其特定 的產物,首先,由兼氧性及厭氧性細菌透過水解、產酸的反應 (即厭氧醱酵 階段),將廢水中有機物 (如不溶性有機物質、碳水化合物、脂肪或蛋白質 等)轉化成揮發酸、氫氣、醇類或醛類等中間代謝產物後々而這些中間代謝 產物再轉化為甲烷及二氧化碳等最終穩定產物。在反應過程中,氫氣產生後 很快被甲烷形成菌轉化成甲烷與二氧化碳,所以氫氣在傳統的厭氧反應過程 中殘留量非常少,可以當作傳統厭氧反應操作不良的重要指標 (林明瑞, 1989),但又依黃國豪、黃耀輝、陳致君、梁德明(1990)的研究顯示,該 反應無法單獨作為系統控制的依據。 厭氧產氫的機制依控制環境的不同,參與的微生物不同而有差異々即使 參與反應的微生物相同,不同的控制環境,也會產生不同的生化反應,造成 產氫效率上的不同。以圖2-3說明Clostridium acetobutylicum醱酵分解葡萄糖 之代謝路徑,不同的反應路徑所生成的代謝產物就有所不同,包括〆乳酸 (lactate)、乙酸(acetate)、丁酸 (butyrate)、乙醇 (ethanol)、丙酮 (acetone)、 丁醇 (butanol)等,而最佳的代謝路徑就是產乙酸的路徑。從圖中可看出在轉 換到丙酮酸(pyruvate)的過程中可產生NADH,NADH在氫化(Hydrogenase) 及鐵氧化還原蛋白(Ferredoxin)的作用下轉換回NAD+並釋出H2々pyruvate 再往下轉換,在產乙酸路徑不會消耗NADH,也就是說分解葡萄糖所產生的 NADH不會被消耗,而全部經由Hydrogenase及Ferredoxin的作用釋放出H2。
22 但由於Hydrogenase及Ferredoxin的反應速率不夠,造成NADH的累積,促使 代謝路徑移向消耗NADH 的產丁酸或產醇的路徑,造成氫氣產率的降低々 另外,大量的乙酸將使這種情形更加嚴重,若反應移往產丁醇的路徑將停止 產氫々就最大理論產氫量而言,每莫耳的葡萄糖可產生4莫耳的氫氣,此時 乙酸是醱酵的副產物而當醱酵的副產物為丁酸時,產氫量則僅為產乙酸時的 一半而已(Dabrock, Bahl & Gottschalk, 1992々Girbal, Crox, Vasconcelos & Soucaille, 1995々白明德,1999)。 複合性聚合體 (纖維素、多醣體、蛋白質) 單體(Oligomers) (醣、胺基酸) 醋酸 丙酸、丁酸 H2+CO2 水解 纖維素或其他水解性細菌 醱酵 醱酵性細菌 醋酸 產醋酸作用 產醋酸菌 產氧脂肪 醱酵 酸氧化菌 醋酸 H2+CO2 CH4 產甲烷菌 產甲烷菌 產甲烷作用 圖2-2 傳統厭氧消化反應過程(謝哲松譯,1995)
23
(1)hydrogenase (2)ferredoxin-NAD reductase (3)NADH-ferredoxin reductase (4)phosphotransacetylase
(5)acetate kinase (6)phosphotransbutrylase (7)butyrate kinase
24
在傳統厭氧反應分解有機物的過程中,厭氧微生物群先將有機物經水解 產酸反應,將大部分有機物轉化為揮發酸、醇、醛、氫氣等中間代謝產物, 若生長條件適當,這些中間代謝產物又會被甲烷生成菌分解或轉換成甲烷。 而厭氧醱酵產氫則是設法終止後面的甲烷化反應,使厭氧反應仍持續進行前 面的水解產酸反應而有利氫氣的產生。由Das and Vezirolu(2001)的研究中 指出厭氧醱酵產氫反應有非常高的氫氣轉換率、穩定的氫氣產生量及良好的 微生物生長率等優點,因此在微生物共培養狀況良好的系統中,電子、氫與 能量在水解、酸化與甲烷化反應之間傳遞著,氫氣很難被釋放至氣相之中, 但是反應槽中微生物共培養系統一旦失去帄衡,氫氣未被甲烷化反應快速消 耗,則將會被釋放至氣相中。在厭氧醱酵產氫機制方面,由於能量是微生物 存活所必頇的原動力,而微生物細胞內能量的來源,主要來自可釋放能量的 化學反應,故此能量為一種化學能。此種化學能貯存在許多的化合物內,當 分子分解時,可將貯存的能量釋出(楊美桂,2003)。而其中最主要的反應為 貯能化合物為腺嘌呤核苷三磷酸(Adenosine Triphosphate, ATP)水解成腺嘌 呤核苷二磷酸(Adenosine Diphosphate, ADP)而釋放出能量。
由上述可知,一般傳統厭氧消化反應可分三階段,即水解、酸化、及甲 烷化三階段。而厭氧醱酵產氫反應則發生在水解及酸化階段,以下就這二階 段進行討論。 (一)水解階段 此階段是利用細胞外水解酶(extracellular enzymes)將複雜的有機物 (如 蛋白質、脂肪、多醣類等)分解成簡單的化合物 (如胺基酸、甘油、單醣等), 因為能源作物大多屬於半固體基質物,所以需要經過水解階段,加以分解成 氨基酸、單糖類、長鏈脂肪酸即甘油等可溶性的小分子物質,以便能被微生 物利用,加強後需的產氫效率。而水解的效率受到 pH 值、水力停留時間 (HRT)及基質種類(Henze & Harremöes, 1983)等因素影響。而進流基質為
25
一般性有機廢水,在水解階段不易形成速率限制,但若進流基質含有芳香族 類有機化合物、纖維素等難水解物質時,在水解階段將成為限制反應速率階 段(Kennedy & van den Berg, 1982)。
水解階段可以使用具有分解纖維素的能力厭氧細菌如 Clostridium、 Bacillus 等,在厭氧狀態下,纖維素經分解菌分解反應後,其分解產物可 被加以利用達到好的產能效果。因此本研究將以批次實驗選出水解菌最佳水 解效果,再利用蔗渣基質在連續流試驗中,前段水解 SBR 反應槽把纖維素 分解後,後段醱酵產氫CSTR 反應槽來升產氫量。 (二)產酸階段 厭氧發酵產氫過程中,除了主要產物氫氣的產生之外,也會伴隨著其他 副產物的生成。當開始生長時,細胞內需大量的能量以供生長,所以會偏向 於生成乙酸的路徑,以利於能量的累積。而高氫氣分壓時會抑制乙酸根之生 成,而使基質大部分轉化為丙酸、丁酸、乙醇,因此傳統厭氧消化必需監控 氫氣之濃度。在厭氧生物產氫代謝機制方面,Taguchi, Chang, Takiguchi & Morimoto(1992)以Clostridium butyrium為菌種,反應槽溫度控制在36℃、 pH 值 控 制 在 5.5 的 條 件 下 操 作 , 理 想 的 產 氧 效 率 為 2.0 ~ 4.0 mole-H2/mole-Glucose,但實際產氫效率大約只有理想產氫效率的60%。其產 氫反應方程式如下所示〆 C6H12O6→ 2H2 + butyrate + 2CO2………(式1) C6H12O6 + 2H2O → 4H2 + acetate + CO2………(式2) 反應過程(式1)的產氫效率為33%,反應過程(式2)的產氫效率為 67%,其比值為1〆2,這說明了Clostridium butyricum如果能進一步將葡萄 糖分解到乙酸階段則其產氫量約為分解到丁酸階段的2倍。Lay et al. (1999) 研究中指出,目前利用有機廢污水以連續式試驗進行醱酵產氫其效果並不 佳,原因是氫氣會被甲烷轉換及消耗。因此如能將反應槽控制在厭氧產氫
26 菌最佳生長條件的操作下,可大幅提昇反應槽的厭氧產氫效率,而目前常 使用的方法有〆(1)pH值控制於較酸的環境々(2)加大有機負荷或縮短 水力停留時間々(3)經常變動操作狀況,如:pH值、溫度、有機負荷、ORP 值等,以對系統造成突增負荷(shock loading)々(4)完全混合反應培養々 (5)儘量將系統操作在醱酵產氫菌適合生長,而甲烷菌不適合生長的環 境。
第六節
厭氧醱酵產氫之影響因子
厭氧產氫過程中,產氫菌將大分子有機物分解成為小分子,除了產氫之 外,往往扮隨著揮發酸和醇類的產生,但產氫的代謝路徑往產酸或產醇的方 向是受到許多環境因素的影響,包含基質、營養源、溫度、pH值、抑制物 質、水力停留時間和食微比等(Chudoba, Capdeville & Chudoba, 1992々 Elefsiniotis & Oldham, 1994々Banerjee, Elefsiniotis & Tuhtar, 1998々Lay, 2000)。 一、基質(substrate) 微生物生長中的細胞必頇利用外來的基質及營養源進行生長並獲取能 量(謝哲松譯,1999)。而當微生物若無法得到充分基質及營養源會影響其體 內的生化反應,甚至會死亡。 生物厭氧產氫之進料基質供給主要以碳水化合物為主,而基質要有足夠 的濃度才可促使醱酵反應發生和產生大量的生質能源 (賴俊卲,2003)。碳水 化合物的濃度會影響產氫總量與代謝途徑(Mizuno et al., 2000),林明瑞、盧 重興、邱毓明、賴欣宏(2000)的研究證明產氫效能也隨著基質濃度的增加 而增加。鄭昭暉(2005)指出,以木糖為基質的批次試驗中,於20g COD/L 有最佳的產氫率1.3 mol H2/mol xylose,低於或高於此濃度則有較差的產氫量27 和產氫率,顯示適當的基質濃度有較佳的產氫效果。 而基質種類亦會影響優勢微生物的組成,以蔗糖為進流基質的生物產氫 反應槽,有76%與Clostridium之親緣相近之菌群々而以麥粕為基質的生物產 氫反應槽,只有0.7%與Clostridium之親緣相近,顯示生物產氫反應槽的微生 物社會結構會受到進流基質的影響 (簡青紅、曾怡禎,2003)。 鄭幸雄等(2001) 所進行厭氧生物產氫機制及程序控制之整合型研究,其研究發現,不同的基 質對產氫菌產氫效能有直接的影響,碳水化合物單位基質的產氫量約有9 mmole-H2/g-glucose,遠大於蛋白質的0.68 mmole-H2 /g-peptone及生物污泥的
0.08 mmole-H2/g-sludge。 在嵇本賢、陳文欽、曾治乾、陳志堅(2001)的研究中發現,以易水解 的glucose為基質,最易達成生物產氫的目標,相同單位的碳水化合物的產氫 量約為蛋白質的數倍,而脂肪與纖維質的產氫量最少。 利用農業廢棄物、 能源廢棄物當基質是未來的趨勢,因纖維質含量非常的高,所以在水解階段 可能難以分解利用,而可分解纖維素之生物在自然界中到處存在,包含單細 胞動物、真菌、細菌、好氧性微生物與厭氧性微生物以及嗜溫性細菌、嗜熱 性細菌等。而其生長環境包括土壤、沼澤濕地、河川湖泊、海水沈積物、腐 壞的葉子、枯木及污泥等。而在許多厭氧細菌中也具有分解纖維素的能力如 Clostridium、Bacillus等菌種,在厭氧狀態下,纖維素經水解菌分解反應後, 其分解產物有H2、乙酸、CO2及醇類。 二、營養源(Nutrients) 微生物生長的需求因子主要為水、碳源、氮源和一些無機鹽類,除此之 外,還包括許多厭氧菌無法合成的必需維他命及氨基酸,此為微生物的必需 品,以供其代謝生長所用。近年來國內外學者在探討厭氧產氫菌種之反應槽 形式之選擇及操作條件,馴養所使用之營養物質配方種類繁多,在混合基質 及混合微生物系統中,要建立特定必需的營養鹽資料,是相當不容易的一件
28 事,必頇針對各種情況加以決定,才能得到最適合的營養鹽配方。 營養源是可供細胞利用而合成細胞物質之化學元素,而為了使厭氧產氫 反應能夠持續進行,除了提供基質之外,還必頇要有足夠的營養源。對微生 物而言營養物質的主要功能有三個(林秋裕,1995)〆(一)供予碳及氮來源 來合成細胞物質和代謝產物々(二)產生生物合成和再生過程中所需的各種能 量々(三)調節細胞之新陳代謝。 為保持厭氧消化之進行,對厭氧菌而言,最佳之C〆N〆P比值為700〆5〆 1 (Sahm,1984)。而一般厭氧反應對N、P的需求較高,其中氮主要是被用來 合成蛋白質、酵素,磷則主要是用於合成儲存能量的化合物,如ATP 等, 所以細菌生長所需的N/P 比為5-7。厭氧消化反應除了一般大量的營養源如 氮、磷之基本需求外,由於厭氧菌大多無法合成一些必需維生素或胺基酸, 因此必需提供特定的胺基酸與營養源供細胞生長及代謝用。有關Clostridium 菌屬培養的研究中,發現Clostridium菌屬的培養,除了酸鹼度緩衝劑與基質 外,還需加入鎂、鈣、鈷、鉬等微量元素。相關研究指出有13種營養源會影 響產氫效率,最適合產氫之營養物質及濃度如表2-7,其產氫效率可從190 ml-H2/g sucrose consumed提升至253 ml-H2/g sucrose consumed,效率提升34
%(賴奇厚,2001)。
此外厭氧生物產氫反應中,hydrogenase 為其重要酵素之一,厭氧產氫 菌利用hydrogenase 和ferredoxin 等酵素系統,將體內氧化有機物多餘的電 子經由此酵素系統轉移給氫離子形成氫氣而排除體外(Chen & Mortenson, 1974々Adams、Mortenson & Chen,1980),減少體內多餘電子的壓力。鐵 離子為構成hydrogenase重要金屬元素之一,每分子的hydrogenase 中含有12 個原子的鐵離子。根據文獻的結果得知(Peguin & Soucaille,1995々Lee, 2001), 不同的Clostridium種類對於鐵離子濃度的需求大不相同,但是鐵離子的存在
29 三、溫度(temperature) 微生物的生長過程與其體內生化反應有關,而此生化反應速率又受到溫 度所控制,根據凡得霍夫定律,在一定的溫度範圍內,每升高10℃反應其速 率增加一倍,因此溫度對微生物的生長會有直接的影響。 表 2-7 適合厭氧醱酵產氫之營養物質及濃度 營養鹽 濃度(mg/L) MgCl2〃H2O 40 NaCl 1000 ZnCl2 0.1 FeSO4〃7H2O 0.2 KI 12.5 MnCl2〃6H2O 2.5 CoCl2〃6H2O 0.1 NH4Cl 50 Na2MoO4〃2H2O 0.1 NiCl2〃6H2O 2.5 MnSO4〃4H2O 2.5 CaCl2〃2H2O 10 CuSO4〃5H2O 0.5 資料來源〆賴奇厚(2001) 而微生物依生長溫度可以分為嗜低溫菌(15-20℃)嗜中溫菌(30-35℃)、嗜 高溫菌(50-55℃)三種。溫度不僅會影響微生物的代謝作用,也影響基質、氣 體的轉移速度及生物固體的沉降性能等。而以溫度與有機物負荷、產氣量的 關係圖可發現在厭氧產氫系統中,中溫消化反應範圍,最佳的消化溫度為 30-35℃間(Henze & Harremöes, 1983)々高溫消化反應範圍內,以50-55℃之間 的效果最好々而在40-45℃之間,甲烷菌生長速率會大幅減退,主要的原因 是在此溫度範圍內,甲烷形成菌之衰退速率(decayrate)相當快,導致生長速 率降低。所以在一般之生活污水處理廠在操作污泥厭氧消化時,若採用 25-40℃之中溫條件,則最佳操作條件為35℃(Koster, 1988)。
30
本研究以Clostridium sp.為菌種,25-45℃為Clostridium 較佳生長環境,
但對產氫速率而言,則以37-40℃為最佳產氫溫度。吳建忠(2005)在CSTR 連續流操作實驗中則發現以高溫50 ℃時的操作最佳,其產氫率、產氫速率 (HPR) 和比產氫速率(SHPR)分別可達1.3 molH2/ mol xylose、245.5 mmol
H2/l/day 及104.9 mmol H2/g-VSS/day々另中溫操作具較佳的系統穩定性,對
於溫度之遽升遽降有較好的緩衝能力(李國興、陳惠莉、吳季芳、羅泳中、 羅泳勝、張益盛、林屏杰、張嘉修,2003),因此本研究將把溫度控制在 35±0.5℃內,以求得最佳試驗條件。 圖2-4 中溫消化與高溫消化之關係圖 (Koster,1988) 四、酸鹼值(pH) 多數的微生物均有其最適生長的pH值,在厭氧產氫的過程中如果沒有 適當調整pH值,則容易使系統失敗。通常揮發酸的累積會使系統酸化,而 過多的氨氮則會使系統的pH值升高,同樣會有抑制的情形發生。厭氧系統
31 在低pH值及過量揮發酸累積的情況下,將取代中性pH之碳酸根系統,對甲 烷菌及醱酵產氫菌產生一定的抑制作用々此外微生物在pH值較高的環境其 適應性較差,較不容易生長,因此pH控制對於厭氧產氫醱酵來說是非常重 要的,適當控制pH值將有助於厭氧產氫的正常操作。一般而言,依據pH 值 的需求可將微生物分為酸性菌(pH=1~5.5)、中性菌(pH=5.5~8.0)及鹼性 菌(pH=8.5~11.5),大部分的產氫微生物最適合生長的pH 值範圍在6.5~8.5, 屬於中性菌。 在厭氧生物產氫程序中,由於大量的酸生成將形成一個低pH的環境, 低pH的環境將造成hydrogenase的活性下降或微生物代謝途徑的改變,甚至 在極端的環境下,會直接毒害微生物。環境的酸化主要影響微生物的增殖, 對於有機物的裂解及攝取影響則較小,Clostridia相較與其他low-G+C革蘭氏
陽性厭氧菌(主要為乳酸菌,lactic acid bacteria)而言,一般被認為屬於生活於 偏中性的環境中。實際上,對於厭氧產氫微生物而言,最佳的生長pH值範 圍為pH=9 ~ 4。所以,當以混合菌群進行生物產氫時,由於每種微生物對於 環境的需求不同,如此,當環境有微小變化時,對於系統的穩定並不會有明 顯的影響(張仕旻,2001)。另外Yokoi et al.(1995)研究中指出,Clostridium 菌屬最適生長的pH值為6.5-8.5之間々但其生長環境在pH值為5.0-5.5之間時, 也可以獲得不錯的產氫效果(Taguchi et al.,1992)。Eva and Cao(1995) 研究發現,在批次實驗中,有機酸的過度累積會抑制氫氣的生成,所以批次 實驗試程最佳pH值為6.0至7.0間。在中性pH範圍進行厭氧產氫,因非常適合 後續的甲烷反應之進行,因此在中性負荷都會進行甲烷化反應々而本研究為 能有放抑制甲烷化反應之進行,長期在pH5.0-5.5馴養,亦培養出在此pH範 圍,可以直效應產H2的Clostridium菌種。
32 五、抑制物質 在整個厭氧反應抑制機制中,有機酸、硫化物 氨氮以及金屬離子濃度 是主要的抑制物質。當濃度低時,有助於微生物之分解速率々但當到達某一 濃度後,會造成生物反應速率的減退,以至終止。 (一)氨氮 厭氧生物處理系統中,氨氮主要以NH4+與NH3兩種形態存在,兩者存在 之比例受水中鹽類濃度、pH值及溫度等因素之影響。在厭氧系統中,高pH 值時氨氮以NH3形態存在,而低pH值時則較有利於NH4+之形成。高pH值時, 高濃度NH3對微生物有毒性 (張鎮南、曾四恭、鄭幸雄、趙家珍、謝永旭、 Zimpro lnc.,2001)々pH為中性時,氨的毒性非常低。氨氮為一必頇的營養 物質,當濃度在50-200mg/L時,被認為可促進微生物的成長。McCarty(1964) 研究中指出,總氨氮濃度1,500-3,000 mg/L之間且pH>7.4時,氨就會對系統 產生抑制作用,但總氨氮濃度大於3,000mg/L時,無論pH值之高低,氨對系 統皆有毒性(表2-8)。Onodera, Miyahara & Noike (1997)的研究指出,氨濃度 愈高時,厭氧產氫菌的產氫能力與基質分解率都會受到抑制,特別是產氫能 力受到抑制的影響最大。 表2-8 氨氮濃度對厭氧菌分解之影響 影響 氨濃度 (mg/L) 有利 50~200 無不利影響 200∼1000 pH 高時,抑制分解 1500∼3000 具毒性 3000 以上 資料來源〆McCarty(1964) (二)有機酸 厭氧生物產氫程序中,有機物經由微生物反應後,會生成大量的揮發酸, 尤其是乙酸及丁酸。酸的生成會造成反應熱力學上的不利,且除了導致pH





![HPSH [ 氧化數平衡反應式係數 ]](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)