國立台東大學 生命科學研究所
Institute of Life Science National Taitung University
碩士論文
Master Thesis
指導教授 : 李炎 博士
由鞘絲藍菌Lyngbya sp.中分離純化藻藍素之 方法及其抗氧化活性探討
The Purification of Phycocyanin from Lyngbya sp. and It’s Oxidation Resistance Activity
研究生 : 張智翔
Student: Chih-Hsiang Chang
中華民國 98 年 6 月
June 2009
由鞘絲藍菌Lyngbya sp.中分離純化藻藍素之 方法及其抗氧化活性探討
The Purification of Phycocyanin from Lyngbya sp. and It’s Oxidation Resistance Activity
研究生 : 張智翔 Student:Chih-Hsiang Chang 指導教授 : 李炎 博士 Advisor:Yen Lee Ph.D.
國立台東大學 生命科學研究所
碩士學位論文
Submitted to Department of Institute of Life Science National Taitung University
In Partial Fulfillment of the Requirements for the Degree of Master in
Life Science June 2009
Taitung, Taiwan, Republic o f China
中華民國 98 年 6 月
June 2009
謝 誌
奔奔波波跌跌走過多少曲折不知幾個彎 ,過程當中,經歷了無數的 風風雨雨,從一開始的懵懵懂懂,到論文順利完成的今天,由衷的感謝 恩師李炎老師在這段期間耐心的指導實驗上許多應注意的細節 ,使我從 中學習了許多以前未知的經驗,訓練了我獨立思考的能力,老師的認真 也改變了我對實驗的態度,讓我了解到求學問不可囫圇吞棗,必須腳踏 實地才能登峰造極,在此對老師致上最高的謝意 。
也要感謝李俊霖老師在這段期間內給予我的指導與 幫助,與老師亦 師亦友的互動,使我能將遇到的困難或瓶頸與老師討論 ,老師的意見總 是能啟發我新的想法,讓我順利解決實驗上的問題 ,非常感謝老師。
要感謝的人很多,但最要感謝的是我的哥哥也是我的學長張智超 , 若沒有他的幫忙,我的實驗不會這麼順利;另外本所的劉炯錫老師、彭 仁君老師,黃祥恩老師,中央研究院生化所蔡明道所長等,諸位師長於 在學期間的指導及鼓勵以及本所的助理張惠嵐小姐給予許多協助 ,同 學、學長姊及學弟妹們的幫忙,一併在此致上深深的感謝。
最後,要感謝我最親愛的父母,辛苦的栽培與教養,困擾中給我支 持,失意中給我鼓勵,你們的支持是我最大的動力,讓我能順利完成碩 士學業,謝謝你們。
在此也祝大家事事順心,健康平安!
中文摘要
本研究主要在探討以快速簡單之萃取方式從 鞘絲藍菌 Lyngbya sp.
中分離純化出藻藍素,並評估所萃取出之藻藍素抗氧化能力 。利用物理 方式先磨碎均質並離心,將上清液進行管柱層析分離或冷凍乾燥法可得 濃度為 3.021 mg/ml;透過光譜分析所測得最強吸收波長為 620nm、SDS page 分子量鑑定可分離出 α-subunit 及 β-subunit 兩主要次單元,分子 量分別為 17 kDa 及 18 kDa,對照標準品結果皆相同。
抗氧化能力分析上,首先建立細胞氧化模型,利用 H2O2 來誘導細 胞產生氧化濃度測試的結果為 0.03% 最適當,高於 0.03% 則細胞所受 氧化壓力過大,低於 0.03% 細胞無顯著影响,故最適濃度訂為 0.03%。
藻藍素作用 H2O2 誘導氧化之細胞模型並與抗氧化劑維生 素 C、維生素 E 以及 β-carotene 同時測試比較,結果發現,當濃度皆以 10 mM 進行測 試時,以藻藍素有最佳的抗氧化能力,相較於同濃度之維生素 C 可抑制 細胞承受氧化壓力之時間為其 6 倍,實驗結果也顯示,當細胞接受氧化 壓力所攻擊時,在三小時內加入藻藍素仍具有相當的抗氧化能 力,而同 濃度的維生素 C 與維生素 E 則否;β-carotene 在 10mM 時對細胞而言濃 度過高,會導致細胞產生皺縮現象,但在抗氧化測試方面,由結果顯示,
先加入 β-carotene 在一小時內對氧化攻擊有明顯的抑制作用,但隨時間 增長則對細胞會造成死亡,抗氧化效果亦不及同濃度的藻藍素來的佳 。 經過實驗的結果顯示,抗氧化劑加入的時間先後對細胞而言影響甚鉅 , 在細胞受到氧化攻擊之後的適當時間加入,才可達到抑制效果;但本實 驗結果顯示 β-carotene 在細胞受氧化壓力前一小時加入,有明顯的效
果,由此可知抗氧化劑若先加入或超過了有效時間,則無法有效抑制氧 化壓力,因種類而異。因此本實驗結果證明了由鞘絲藍菌中所萃取之藻 藍素有優於其他抗氧化劑的能力,效果可達至少 5 倍以上,對於日後保 健食品以及美妝保養品或抗氧化藥劑的研發上 ,有極佳的前瞻性。
關鍵字:鞘絲藍菌、藻藍素、抗氧化、抗氧化劑。
Abstract
This research used a fast and simple extraction method to purify the phycocyanin from Lyngbya sp., and tested the phycocyanin oxidation resistance ability. Lyngbya sp. cells were ground in distilled water and centrifuged, then the supernatant run through the column chromatography tube. The separated liquid had the concentration of 3.021 mg/ml. The liquid was then freeze dried. The liquid was analyzed by spectrophotometer. It showed the maxmum absorption wave length was 620 nm. The SDS p age result showed the protein had α - and β- two main subunits. Their molecular weights were 17 kDa and 18 kDa respectively, compared to Sigma standard.
For the oxidation resistance ability analyzation, cell oxidation model was firstly established. Different concentrations of H2O2 were used to treat the Vero cells. The most adequate concentration was found to be 0.03%. While 10mM of phycocyanin, vitamin C, vitamin E as well as β -carotene were used separately to treat the H2O2 stressed Vero cells, phycocyanin showed the best oxidation resistance ability. The experimen tal results also demonstrated, during 3 hours of Vero cells that treated by oxidization attack, then administrated the phycocyanin to the cells, phycocynin could help the cells to have suitable oxidation resistance ability. Nevertheless, the same concentration of neither vitamin C, vitamin E nor β-carotene could prevent the cells from shrinkage phenomenon. Experiment results also demonstrated thatβ-carotene added to Vero cells one hour before oxidization attack could
have obviously inhibitory effect. It was estimated that the phycocya nin extracted from Lyngbya sp. suppressed other oxidation inhibitor ability about 5-fold.
It is reasonablely to say, Lyngbya sp. has good ability for applications in the future in either health food or cosmic market.
Key word:Lyngbya SP.、phycocyanin、antioxidant
目 錄
中文摘要... I 英文摘要... III 目 錄... V 圖目錄... VIII
壹、緒論... 1
一、前言... 1
二、研究目標 ... 3
貳、文獻整理... 5
一、藍菌概述 ... 5
二、藻藍素萃取方法回顧 ... 13
三、測定抗氧化活性之方法原理 ... 14
四、自由基的來源 ... 19
五、氧自由基對生物體內所造成之危害 ... 21
六、抗氧化劑之種類 ... 23
七、藻藍素... 27
八、藻藍素的生理功效 ... 31
参、實驗流程... 33
肆、材料與方法 ... 34
一、實驗材料 ... 34
二、實驗儀器 ... 35
三、藍菌純化與培養方法 ... 35
3.1 培養基配製: ... 35
3.2 藍菌培養 ... 36
3.3 藍菌純化 ... 37
3.4 藍菌保存 ... 37
四、藻藍素分離前處理 ... 38
4.1 冷凍乾燥法 ... 38
4.2 膠體過濾層析法 ... 39
五、藻藍素萃取條件方法開發 ... 39
5.1 冷凍乾燥法 ... 39
5.2 膠體過濾層析法 ... 39
六、藻藍素特性分析 ... 40
6.1 藻藍素定量分析 ... 40
6.2 藻藍素定性分析 ... 41
七、藻藍素對細胞抗氧化能力分析 ... 42
7.1 細胞來源 ... 42
7.2 細胞培養 ... 42
7.3 過氧化氫誘導細胞氧化模型的建立 ... 45
7.4 藻藍素對過氧化氫誘導細胞氧化之影響 ... 45
7.5 維生素 C 對過氧化氫誘導細胞氧化之影響 ... 46
7.6 維生素 E 對過氧化氫誘導細胞氧化之影響 ... 46
7.7 β-carotene 對過氧化氫誘導細胞氧化之影響 ... 47
7.8 藻藍素抗氧化能力分析 ... 47
伍、結果與討論 ... 49
一、藍菌培養 ... 49
1.1 固態培養小量純化 ... 49
1.2 液態小量培養 ... 49
1.3 液態大量培養 ... 49
1.4 藍菌純化 ... 54
二、藻藍素萃取條件方法開發 ... 54
2.1 冷凍乾燥法 ... 54
2.2 膠體過濾層析法 ... 59
三、藻藍素特性分析 ... 59
3.1 藻藍素定量分析 ... 59
3.2 分子量 ... 61
3.3 吸收光譜 ... 61
四、藻藍素細胞抗氧化能力分析 ... 64
4.1 過氧化氫誘導細胞氧化模型建立 ... 64
4.2 藻藍素對過氧化氫誘導細胞氧化之影響 ... 68
4.3 維生素 C 對過氧化氫誘導細胞氧化之影響 ... 68
4.4 維生素 E 對過氧化氫誘導細胞氧化之影響 ... 73
4.5 β-carotene 對過氧化氫誘導細胞氧化之影響 ... 76
4.6 藻藍素抗氧化能力分析 ... 79
陸、結論... 83
柒、參考文獻... 85
附錄一、C-10 培養基配方 ... 97
C-10 培養基 (藥品單位為每公升蒸餾水中所含公克數 : g/L ) ... 97
微量元素 (藥品單位為每公升蒸餾水中所含公克數 : g/L ) ... 97
圖目錄
圖一.藍菌( Cyanobacteria ) ... 2
圖二.鞘絲藍菌 (Lyngbya sp.)... 4
圖三.藍菌細胞圖(Cyanobacterial cell structure) ... 6
圖四.類囊體 (Thylakoid)... 7
圖五.位於類囊體上之藻膽體(phycobilisome) ... 9
圖六.藻膽體結構圖... 10
圖七.藻膽體吸收光能產生能量路徑圖 ... 11
圖八.不同色素之吸收光譜 ... 29
圖九.藻藍素( C-phycocyanin, C-PC )... 30
圖十. Lyngbya sp.固態培養圖 ... 50
圖十一.顯微鏡下 Lyngbya sp. ... 51
圖十二.小量液態培養之 Lyngbya sp. ... 52
圖十三.大量液態培養之 Lyngbya sp. ... 53
圖十四.萃取出的藻藍素粉末... 56
圖十五.萃取出的藻藍素水溶液 ... 57
圖十六.萃取出的藻藍素水溶液 ... 58
圖十七.光譜強度之線性關係... 60
圖十八.Bovine serum albumin (BSA) 蛋白質定量標準曲線 ... 60
圖十九.10% 聚丙烯醯胺膠體電泳結果 ... 62
圖二十.藻藍素吸收光譜... 63
壹、緒論
一、前言
藍菌 ( Cyanobacteria )(圖一),在地球上出現約已 30 億年,是最早 的光合放氧生物,對地球表面大氣原始的無氧環境轉變為有氧環境 有重大 作用,原始大氣層主要是由氮氣及二氧化碳所組成 ,環境不適合生物生 長,地表溫度高,大氣中缺乏遮蔽紫外線之臭氧層,而生物經光合作用產 生之氧氣受紫外線照射後會形成具有毒性的活性氧 ( 氧氣分子多一個電 子O2
-‧),而藍菌因其有強抗氧化作用可消除毒性氧而生存,並藉釋出光 合作用產生之氧氣使地球漸形成臭氧層:氧氣分子可以吸收小於 240 μm 波長的太陽光中的紫外線,並分解形成氧原子。這些氧原子與氧分子相結 合生成臭氧,生成的臭氧對太陽的紫外輻射有很強的吸收作用,有效地阻 擋了對地表生物有傷害的紫外線,而使其他生物得以陸續生長。在分類上 因不具有細胞核與粒腺體 ( mitochondria ),高氏體 ( Golgi body ),或葉 綠體 ( chloroplast ) 等胞器,而被列為原核生物 ( Beams and Kessel 1977 )。
目前已發現的藍菌種類約 2000 多種,分佈廣泛,在淡水和海水中,
潮濕和乾旱的土壤或岩石上、樹榦和樹葉上,溫泉中、冰雪上,甚至在鹽 滷池、岩石縫中都可以發現藍菌;有些還可穿入鈣質岩石或介殼中或土壤 深層中。由於藍菌分佈廣,數量多,學界對其之研究也越來越多,已知的 研究已有應用在食品方面,其菌體內所提取之化學物質亦有被發現對於抗 細菌或病毒以及免疫調節等效果,對於生理醫學上的研究有極大前瞻性 ; 農業方面的肥料使用,甚至工廠廢水處理,廚餘臭味的消除;另外生產含
放射性同位素的化合物,利用其生理代謝方式產生氫氣等,都是目前藍菌 被廣泛應用的範疇。
圖一.藍菌( Cyanobacteria ) (Nikon YS100 目鏡 20x;物鏡 40x)
二、研究目標
由於藍菌普遍富含藻藍素,而藻藍素又被研究發現具有許多生理上之 功效,例如抗氧化、抗發炎作用;護肝以及抗腫瘤作用等(廖 2006),而 本實驗室由李炎教授分離純化出之藍菌 Lyngbya sp. (鞘絲藍菌)(圖二) 依 生物學上之分類,歸類於原核生物界 (Monear),藍菌門 (Cyanophyta),
藍 菌 綱 (Cyanophyceae) , 顫 藍 菌 目 (Oscillatoriales) , 顫 藍 菌 科 (Oscillatoriaceae),鞘絲藍菌屬 (Lyngbya ),培養時間短,成本低且於培養 液中生長快速,經篩選又為藍菌食品開發之優良菌屬(李 2005),而從已 知的 Lyngbya sp. 菌屬當中,發現許多抗細菌及病毒之化學物質,於生理 醫學以及保健食品的開發上具有相當之價值(李 2005),故本實驗決定以 Lyngbya sp. 為主要研究對象,研發從菌體中萃取分離純化出 藻藍素,進 行光譜吸收波長比對,分子量鑑定,並研究探討其細胞抗氧化能力。
圖二.鞘絲藍菌 (Lyngbya sp.) (Nikon YS100 目鏡 20x;物鏡 40x)
貳、文獻整理
一、 藍菌概述
藍菌如前述在生物學上的分類屬於原核生物界。其細胞膜組成屬格蘭 氏 陰 性 ( Gram-negative ) , 細 胞 膜 附 著 細 胞 壁 , 細 胞 壁 為 脂 多 糖 ( lipopolysaccharide-LPS ) 與肽聚糖 ( peptidoglycan ) 組成之複合體,細 胞膜內部則為細胞質 ( cytoplasm )( Ris and Singh 1961; Stolz 1991 )。細胞 大小由 1 μm 至 100 μm 不等,差異很大 ( Castenholz and Waterbury 1989 )(圖三)。而其與細菌及古生菌之區別 ,可由 16S 與 5S rRNA 的排 序鑑別出,另外,比較 23S rRNA 排序與延長因子 ( elogngation factor ) Tu 與 ATP 合成酶 ( ATPase ) 的 β 附屬單位 ( subunit ) 的蛋白質結構,可 更明顯分別出與一般細菌之不同 。類囊體(thylakoid)(圖四)是它們主要發 生光合作用之處,是一存在於細胞內的膜狀結構 ( Stolz 1991 ),類囊體會 隨著不同環境如光照、溫度、細胞生長時間長短、缺磷、氫化物、形成厚 垣孢子 ( akinete )或異形細胞 ( heterocyst ) 等情形下改變表現狀況,例如 扭曲、緊靠、鬆散、甚至消失等 ( Stolz 1991; Grossman and others 1993 )。
極少數藍菌無類囊體,細胞膜是它們主要發生光合作用機制之處 ( Stolz 1991 )。
肝糖微粒
藻膽體 (側面) 藻膽體 (正面)
藻藍蛋白
類核體
細胞壁 細胞膜
類囊體
磷微粒
脂質微粒
羧化酶體 核糖體 氣泡
圖三.藍菌細胞圖(Cyanobacterial cell structure)(Hartwell T.Crim,1998 )。
圖四.類囊體 (Thylakoid)。
(http://www.biologie.uni-hamburg.de/b-online/library/webb/BOT311/Cyanobacteria/Cyano bacteria.htm)
藍 菌 菌 體 中 亦 含 有 藻 膽 體 ( phycobilisome )( 圖 五 ) , 富 含 藻 藍 素 ( phycocyanin, PC ) 與異藻藍素 ( allophycocyanin, APC );又因菌種的不 同,有的含有藻紅素 ( phycoerythrin, PE ) 有的則否(圖六)。藍菌均具藻 膽體,藻膽體位於類囊體面向細胞質外的部分,當光子被藻膽體吸收,可 利用所吸收的能,將水分解,形成 O2與 H+,並透過電子傳遞系統,在類 囊體膜兩側形成電化滲透壓 (chemiosmotic or electrochemical potential)。所 形成的能量,可使 ADP + Pi ATP (圖七)(Bryce and Hill 1999)。
其中藻藍素已被廣泛研究,於生理活性上已被發現具有抗氧化、抗發 炎以及能護肝且有細胞週期調節之作用 (廖 2006);在分離純化方面也有 許多的研究探討(Brejc et al., 1995;Minkova et al., 2003;Pinero et al., 2001),而藻類機能性保健食品的開發 ,如綠藻、螺旋藍菌等皆被認為具 有可預防生活習慣疾病的功效,且已逐漸受到人們所重視,被認為含有高 營養、抗氧化、降低膽固醇的成分 ( Bhat et al., 2000;Herrero et al., 2003;
Mendes et al., 2003 )。而其中藻藍素具有之鎮痛、抗氧化抗發炎能力,因 其結構與血紅素之降解產物膽紅素 ( bilirubin ) 極為相似 , 膽紅素 ( bilirubin ) 是人體中重要的抗氧化物,可抑制血漿蛋白質與芳香族胺基 酸的氧化並可清除血清蛋白的氧自由基如超氧陰離子 ( O2
-‧)與過氧化自 由基 ( Farrera et a., 1994; Romay et al., 2003 )。
圖五.位於類囊體上之藻膽體(phycobilisome)。
(http://www.biologie.uni-hamburg.de/b-online/library/webb/BOT311/Cyanobacteria/Cyano bacteria.htm)
圖六.藻膽體結構圖:藻藍素( phycocyanin, PC );異藻藍素( allophycocyanin, APC );藻紅素( phycoerythrin, PE )。
(http://www.biologie.uni-hamburg.de/b-online/library/webb/BOT311/Cyanobacteria/Cyano bacteria.htm)
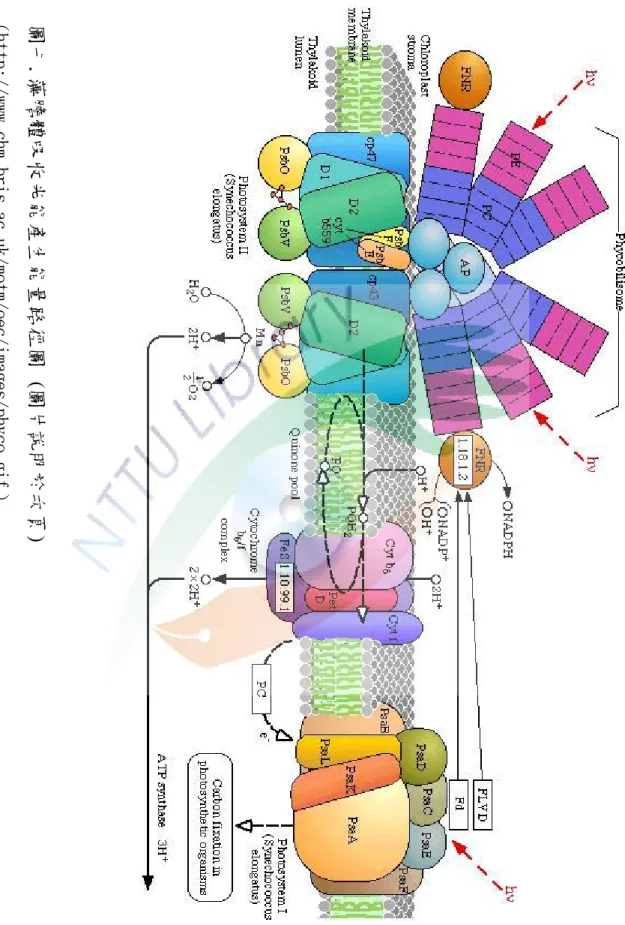
圖七.藻膽體吸收光能產生能量路徑圖
圖七.藻膽體吸收光能產生能量路徑圖(圖片說明於次頁)
(http://www.chm.bris.ac.uk/motm/oec/images/phyco.gif)
圖七中:
phycobilisome = 藻膽體,
hv = 可見光光子
PE = phycoerythrin 藻紅素 PC = phycocyanin 藻藍素
AP = allophycocyanin 異藻藍素 thylakoid membrane = 類囊體膜 D1、D2 = PS II 的核心蛋白質體
CP43,、CP 47= 分子量分別為 43 與 47 kD 的葉綠素蛋白質複合體 PsbO, PsbV = PS II 中與氧結合有關的蛋白質
Cyt b559 、Cyt b6 、Cyt f = 細胞色素 b559、細胞色素 b6、細胞色素 f PQ = Plastoquinone 質醌
FeS = 鐵硫氧化還原中心
方形 PC (Cyt c553) = 細胞色素 c553 ,ㄧ種類囊體囊腔中電子傳遞蛋白質 , thylakoid lumen = 類囊體囊腔
stroma = 基質(亦即細胞質的部份)
FNR = ferredoxin/flavodoxin NADPH oxidoreductase 鐵氧化還原蛋白/煙 鹼醯胺腺嘌呤二核苷酸磷酸,氧化還原酶
PsaA, PsaB, PsaD, PsaE, PsaL, PsaK = 在 PS I 中與葉綠素結合的蛋白質 Flvd = ㄧ種黃素氧化還原蛋白
Fd = 鐵氧化還原蛋白
二、藻藍素萃取方法回顧
藻膽蛋白皆為水溶性,可輕易與其他脂溶性色素分離。有關藻藍素之 研究已有很多,目前主要的純化方式仍是以硫酸銨沉澱後進行色層分離法 為主 , 但所選擇之分 離系統及純化條 件各有不同 , 亦 有以 rivanol (6,9-Diamino-2-ethoxyacridine-DL-lactate monohydrate) 進 行 前 處 理 者 (Minkova et al., 2003 );色層分離系統可選用膠體過濾法及離子交換法 , 或 再 以 羥 基 磷 酸 鈣 管 柱 (hydroxylapatite) 將 藻 藍 素 及 異 藻 藍 素 分 離 ( Pinero Estrada et al., 2001),也有使用了厭 水交互作用色層分析法 hydrophobic interaction chromatography (HIC) (Badrish Soni et al., 2007),和 離子交換色層分析法來萃取純化藻藍素(Julio Abalde et al., 1998)。
近幾年也有學者利用雙水相萃取法 aqueous two phase extraction (ATPE)搭配離子交換層析法進行藻藍素的萃取,利用聚合物之間的不相 溶性,即聚合物分子的空間阻礙作用,相互無法滲透,不能形成均勻一相,
從而具有分離傾向,在一定條件下即可分為二相。一般認為只要兩聚合物 水溶液的疏水程度有所差異,混合時就可發生相分離(Ganapathi Patil et al., 2008)。
除了傳統的蛋白質萃取與純化方式,Swanson 及 Glazer 利用反相高 效能液相層析儀對藻膽蛋白直接進行純化及分析,可得到高純度的藻藍素 ( Swanson and Glazer, 1990);但此種方式的缺點是成本高,不適合應用於 工業上的大量純化。
三、測定抗氧化活性之方法原理
抗氧化劑是能阻止或很大程度抑制易氧化物質氧化過程,能清除活性 氧或者阻止活性氧生成的一類物質,在病理學和飲食中都起著不可或缺的 作用。而一些人工合成的抗氧化劑有潛在的毒性,天然抗氧化劑越來越受 到人們的重視。
抗氧化劑的抗氧化活性主要表現在抑制脂質氧化降解、清除自由基、
抑制促氧化劑 (如螯合過渡金屬) 和還原能力等幾方面。現就較常用的一 些方法進行簡單的介紹。
l.還原力的測定
還原力的測定,實質上是檢驗物質是否為良好的電子供應者的過程 。 還原力強的物質供應的電子除可以還原氧化性物質外 ,也可與自由基反 應,使自由基成為穩定的物質。
還原力測定源於 Oyaizu(1986) 之方法,以 Fe4(Fe(CN)6 生成量作為 指標。將K3Fe (CN)6 還原成 K4Fe(CN)6,再利用 Fe2+ 形成 Fe4(Fe(CN)6。 在700 nm 吸光值反映 Fe4(Fe(CN)6 的生成量, 吸光值越大,樣品還原力 值越強,表示抗氧化效果越佳。Wang 等從煙葉中提取多酚(polyphenols),
以維生素E作對照,用此法測定了還原力。結果顯示,還原力對於同一物 質只是劑量回應,加大濃度對回應性質沒有影響 ( Wang et al., 2008)。
2.抗油脂過氧化 (Lipid peroxide) 法
生物體內細胞膜的流動性和滲透性由其磷脂等成分決定,過多的自由 基攻擊,油脂過氧化會導致細胞死亡。因此,抗氧化劑對脂類氧化的抑制 作用也顯得非常重要。Rathee 等用老鼠肝臟粒線體作為受質,利用一種
植物芽體 ( nagkesar ) 提取物抗脂質過氧化能力為 ︰α-tocopherol > 水-乙 醇萃取物 > 甲醇萃取物 (Rathee et al., 2006)。
3.金屬螯合
Fenton之氧化原理:Fenton 氧化原理於 1894 年由 Fenton 氏提出 H2O2/Fe2+氧化機制,直至 1934 年由 Haber 及 Weiss 氏證實 Fenton 可 用來氧化有機化合物(Carberry and Yang, 1994)。 Fenton 法乃是過氧化氫 與亞鐵離子於酸性條件下反應,衍生具強氧化能力的氫氧自由基 (OH‧) 氧化有機物,亞鐵離子被氧化成三價鐵離子,故 Fenton 程序同時兼具氫 氧自由基氧化與鐵鹽混凝雙重功能 (Lidia et al., 2001),其反應機制如下所 示:
Fe2+ + H2O2 → Fe3+ + OH- + OH‧ (1)
Fe2+ + OH‧→ Fe3+ + OH- (2)
2Fe2+ + H2O2 → 2 Fe3+ + 2OH- (3) H2O2 + OH‧→ H2O + HO2‧ (4)
Fe3+ + HO2‧→Fe2+ + H+ + O2 (5)
由方程式(1)顯示,亞鐵離子被過氧化氫氧化成三價鐵並生成氫氧自 由基(OH‧ ),亞鐵離子亦會與氫氧自由基產生如(2)式之反應,綜合(1) ~ (2) 式可得到如(3)式之反應,氫氧自由基氧化能力強於過氧化氫 ,與過氧化 氫反應產生水及HO2 其反應如(4)式,(5)式為三價鐵與 HO2 反應還原 為亞鐵離子,若過氧化氫劑量足夠,則反應可循環不斷進行。
在 Fenton 反應之前,金屬螯合降低過渡金屬酵素的濃度,還原性樣
品與 H2O2 競爭,因而在抗氧化性能測定中顯得非常重要 。Lopes等在研 究中發現,單寧酸的抗氧化能力與亞鐵離子濃度成反比 ,相對於 O H‧
其抗氧化能力歸因於鐵的螯合 (Lopes et al., 1999)。
4.自由基清除
自由基是含有未成對電子的原子、原子團或分子。活性氧如超氧陰離 子、羥基自由基和過氧化氫由正常的生理過程和各種外在 因素造成。它們 一旦產生,就會引發油脂過氧化,與人體很多疾病相關。自由基損傷生物 大分子,是造成衰老的主因。因此,自由基清除能力的測定對生物研究非 常重要。
4.1 DPPH‧(α,α-diphenyl-β-pricrylhydrazyl) 和 ABTS +‧(2,2’-azino-bis -[3-ethylbenzthiazoline sulfonic acid] )清除
大部分自由基性質活潑,壽命非常短,但 DPPH‧和 ABTS +‧例外,
是兩種穩定的自由基。DPPH‧乙醇溶液呈深紫色,在 517 nm 處有一強 吸收。若有單電子配對,則吸收消失,其褪色程度與接受的電子數成定量 關係,因而可用分光光度法進行定量分析 。Rathee 等用 DPPH 法測 nagkesar 的甲醇萃取物和水-乙醇萃取物的抗氧化性 ,結果顯示甲醇提取 物的清除能力 (IC508.33 ± 0.391 g/ml) 優於水-乙醇萃取物的清除能力 (Rathee et al., 2006)。
而 ABTS 能被各種氧化劑生成藍綠色的自由基陽離 子,而抗氧化劑 的存在又可使 ABTS +‧還原,顏色褪去。特徵吸收峰下波長比對,清除 能力用 TEAC( trolox equivalent antioxidant capacity,trolox 當量抗氧化能 力)表示。Mathew 等用過硫酸鉀氧化羥自由基,測玉桂葉對 ABTS +‧的
清除能力,得沒食子酸> 抗壞血酸> BHT > 玉桂葉(Mathew et al., 2006)。
4.2 羥自由基( OH‧)的清除
OH‧是體內最活潑的活性氧,也是已知的對生物分子破壞能力最強 的自由基之一(Mathew and Abraham, 2006)。
4.2.1 去氧核糖法
利用 Fe3+-EDTA-VitC-H2O2 體系產生 OH‧,然後去氧核糖受 OH‧
攻擊裂解,產物在加熱、酸性條件下與硫代巴比妥酸 (thiobarbituric Acid -TBA)回應生成一粉紅色化合物。因而樣品對去氧核糖裂解的抑制作用就 很好的展現了其抗氧化性。
反應過程為:
Fe3++EDTA +VitC→ Fe2+-EDTA
Fe2+ -EDTA +H2O2→OH‧+ Fe3+-EDTA OH‧+去氧核糖+TBA→粉紅色化合物
Rathee 等用此法對 nagkesar 進行抗氧化研究發現 ,其甲醇萃取物和水- 乙醇萃取物均對去氧核糖裂解的抑制作用很明顯,其中若不加引發劑抗壞 血酸則幾乎沒什麼效果(Rathee et al., 2006)。
4.2.2 Fe(phen)3法
此法是利用 H2O2/Fe2+透過 Fenton 反應產生 OH‧,Fe2+(phen)3(phen = 1,10-phenanthroline) 溶液被 OH‧氧化成 Fe3+(phen)3。Fe2+(phen)3 (橙紅色) 的褪色程度可用來衡量 OH‧的清除量。金鳴等建立了此法,並用此法證 明甘露醇、抗壞血酸和硫脲清除 OH‧作用呈良好量效關系(金 1996)。
4.3 超氧陰離子( O2
-‧)清除
4.3.1 NBT (nitroblue tetrazolium) 法 xanthine-xanthine oxidase 體系產 O2
-‧,NBT 被 O2-‧還原成藍紫色 formazane 。比色 formazane 即間接測定 O2
-‧。Sakanaka 等用此法對幾 種醋進行抗氧化性測定,它們都有 O2-‧清除能力,且柿子醋和米醋的清 除能力比加工過的米醋和蘋果醋的 要高(Sakanaka and Ishihar, 2008)。
4.3.2 APS(ammonium persulfate)/TEMED(tetramethylethylenediamin e)法 此法是透過過硫酸銨/TEMED 體系產生氧自由基,然後可採用不同的 方法測定。反應機轉為 O2-‧與羥胺溶液反應生成 NO2-,NO2-經對氨基苯 磺酸和 α-naphthylamine 顯色在 530 nm 附近有特徵吸收峰,透過檢測 NO2- 間接檢測 O2-‧。此法由蕭華山等建立,並驗證了抗壞血酸對 O2-‧的清 除作用呈明顯量效關係(蕭 1999)。
4.3.3 MTT (3-(4,5-cimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide )測 定法
MTT 在鹼性條件下自動氧化,不斷釋放出 O2
-‧,O2
-‧又可進一步 促進自身氧化。在 MTT 自氧化 30~40s 後,中間物累積濃度與時間呈線 性關係,一般線性時間可維持 4 min 左右,可求得自身氧化速率來顯示 O2
-‧的清除能力(鐘 2004)。
四、自由基的型式
自由基與活性氧的來源廣泛,分為內在與外在兩大來源,內在生理效 應 產 生 自 由 基 的 途 徑 為 O2 自 粒 線 體 電 子 傳 遞 鏈 、 細 胞 色 素 P-450 ( cytochrome P-450 )的解毒反應、過氧化體 ( peroxisome ) 代謝反應等系 統中,連續接受 4 個電子和 4 個氫原子還原成水。在此過程中經由電子 轉移會依序產生 O2-‧、H2O2 和 OH‧( Imlay and Fridovich, 1991 );另 ROS (reactive oxygen species) 亦會經由許多生理作用產生 ,包括吞噬作用 ( phagocytosis )、訊息傳遞 ( signal transduction ) 與細胞凋亡( apoptosis ) 等 ( Moncada and Higgs, 1991 )。外在環境來源則包括:空氣污染物、高 氧環境、離子化輻射、化學藥劑如四氯化碳、農藥或防腐劑等均會產生自 由基,引起中毒或慢性病。活性氧的種類包括:
1. 超氧陰離子自由基 ( superoxide anion radical )
以 O2-‧表示,為體內最先產生的自由基,可再轉變為 H2O2、OH‧
等,是由體內吞噬細胞 ( phagocyte ) 及粒線體中的電子傳遞鏈產生 。當 吞噬細胞遇到外來物質時,即產生 O2-‧,以抵抗外來的入侵者;而粒線 體為人體製造能量的場所,經過電子傳遞鏈產生 ATP 以供身體所需,
但是電子傳遞鏈產能的步驟會產生 O2
-‧,若產生過多,將對人體細胞造 成傷害,引發疾病 ( 丁 1996;增山 1997 )。
2. 過氧化氫 ( hydrogen peroxide, H2O2)
由 O2-‧轉變而來,或吞噬細胞的吞噬作用及其他氧化還原循環中產 生。H2O2 本身雖比其他的自由基穩定 ,但對細胞的傷害較大,主要是因
為 H2O2 可以通過細胞膜,和體內運轉中的鐵或銅發生費敦反應 ( Fenton reaction ),因而產生 OH‧ ( 丁 1996;增山 1997 )。
3. 氫氧自由基 ( hydroxyl radical )
以 OH‧表示,由前述的自由基轉變而來,或過多 X-ray 及日光的 照射均會在體內產生。OH‧是破壞力最強的自由基,造成體內脂質過氧 化。一個 OH‧與不飽和脂肪酸產生過氧化反應,因連鎖反應而造成上百 個不飽和脂肪酸的氧化,使體內的細胞膜遭到破壞。OH‧與體內醣類、
胺基酸、磷脂質、核醣體、有機酸等任何物質反應,造成細胞死亡或突變,
使得組織不能發揮應有的功用 ,而產生疾病 (丁 1996;增山 1997;
Halliwell and Cutteridge, 1986 )。
4. 單線態氧 ( singlet oxygen )
以 1O2 表示,是由一般的三線態氧經紫外線的照射 ,使得電子軌域的電 子由基態 ( ground state ) 轉變到激發態 ( excited state ),而單線態氧比三 線態氧每莫耳高出 22 Kcal 的能量,故單線態氧的活性非常高 ,比一般 氧氣的反應力更強,也較不穩定 ( 丁 1996;增山 1997 )。
5. 過氧化脂質 ( lipid hydroperoxide )
脂質和脂蛋白的不飽和脂肪酸對自由基引起的氧化特別敏感,因不飽 和脂肪酸具有特殊之化學結構,烯丙基的 ( allylic ) 雙鍵以 CH2基分開,
導致 CH2 基上的氫易受自由基的攻擊而生成碳中心自由基 ,再與基態氧 分子結合形成過氧化自由基與脂質過氧化物。不飽和脂質氧化成帶有不成 對電子的脂質,而一連串連鎖反應,造成更多的脂質被破壞 ( 增山 1997;
Giese 1996 )。另有研究顯示,脂質過氧化物和活性氧在許多癌症及促進 老 化 的 發 展 上 相 當 重 要 , 脂 質 氧 化 的 二 級 產 物 , 如 丙 二 醛 ( malondialdehyde, MDA ) 是相當具有活性的,易與生物體的蛋白質、胺 基 酸 、 DNA 反 應 , 和 老 化 、 致 突 變 性 ( mutagenesis ) 和 致 癌 性 ( carcinogenesis ) 等皆有關連 ( Tamura et al., 1991; Wang and Liehr, 1995;
Hamilton et al., 1997 )。
五、氧自由基對生物體內所造成之危害
人體內以活性氧之自由基最為重要,活性氧 ( reactive oxygen species, ROS ) 形成有許多來源,如:黃嘌呤氧化酶 ( xanthine oxidase );( Kukreja and Hess, 1992 ) 、 活 化 之 吞 噬 細 胞 ( Weiss 1989 ) 、 粒 線 體 ( mitochondria )( Luft 1994 )、溶酵素體( peroxisome )( Kasai et al., 1989 )、內質網或微粒體之電子傳遞系統的 cytochrome P450 oxidase 及 reductase ( Bachur et al., 1979 )、epinephrine 與 dopamine 之自身氧化作用 ( autooxidation ) 相伴引起氧化還原循環作用 ( redox cycling ) 均會產生 活性氧 ( Ames et al., 1993;Dix and Aikens 1993 )。一般經呼吸進入體內 的氧氣約有 1-3 % 的活性氧,包括 過氧化脂質(LOO‧)與活性氧。環境 中的自由基主要來自離子輻射、空氣污染、紫外線照射、抽菸、生理與心 理的壓力及暴露於化學物質等 ( Schechter 1996 )。生物體中具有許多防禦 系統來控制過氧化作用,但若吸收或體內的活性氧與自由基過多時,體內 的抗氧化系統無法維持在一個平衡狀態。超氧自由基在酸的環境下,易質 子化形成過氧化氫活性氧 ( H2O2 ) 或在含有大量一氧化氮 ( NO‧ ),
在環境中易與 NO‧形成peroxynitrite(ONOO-),此物氫基化裂解會放出 HO‧,造成周圍生物分子損傷 ( Halliwell and Gutteridge, 1990 )。因有不 成對電子的存在,賦予自由基極不穩定的特性而具有強化學反應性,易與 細 胞 產 生 一 連 串 的 化 學 反 應 , 造 成 脂 質 氧 化 性 傷 害 (Halliwell and Gutteridge, 1990 )、蛋白質氧化性傷害 ( Stadtman 1992 )、核酸氧化性傷 害 ( Janssen et al., 1993 )、醣類氧化性傷害( Woff and Dean, 1987 )。
此外,隨著年齡的增長,抗氧化防禦系統亦會降低,導致體內抗氧化 還原失調,引起許多病變。增山( 1997 ) 指出,促使脂質過氧化的自由基 以及經由脂質過氧化產生的自由基,不僅會攻擊細胞膜、細胞組成物而且 危害細胞核內的細胞物質,最後導致細胞死亡及致癌性,此與許多疾病的 發展有重要的相關性。目前已知和自由基有關的疾病包括心臟循環系統以 及發生在肺臟、腎臟、紅血球、消化系統、腦、眼球、皮膚、癌症所產生 的疾病。最近亦發現人體的老化與體內自由基過多有關。體內活性氧或自 由 基 的 增 加 或 抗 氧 化 防 禦 能 力 的 降 低 將 導 致 氧 化 壓 力 ( oxidative stress )。氧化壓力可以說是疾病的第二性狀 ,也是疾病所導致的一個結 果。氧化壓力與組織傷害互為因果關係,白血球在慢性發炎的部位產生過 量的 O2
-‧、H2O2 及其他自由基會導致組織劇烈的傷害 ,此時若抗氧化 系統無法提供適當的保護,將促使疾病的惡化。然而自由基在許多人類疾 病上扮演的重要角色,適量的自由基在人體內有生理作用,若不正常過量 的自由基或活性氧卻可直接或間接造成人類疾病,許多證據顯示自由基與 人體病變有密切的關係 ,但自由基在人體的生理角色卻是非常重要 ( Sundaresan et al., 1995 )。
而陽明大學最新研究成果表示,以證實位在第四號染色體上的 Cisd2 基因可以控制哺乳類動物壽命並 有效調控細胞老化,由於不論低等或高等
生物同樣皆具有Cisd2基因。研究顯示,剔除Cisd2基因的小黑鼠,在四到 八週(約人類十五至二十五歲)後,不僅體型比同胎兄弟瘦小,還長出白 毛、不停顫抖、喘不過氣;晚期眼睛凸出白濁、骨質疏鬆、駝背、皮膚鬆 弛。研究的小黑鼠當中有 40% 在七個月死亡,比野生鼠平均能活兩年的 生命周期,明顯短命許多。Cisd2 是影響粒線體結構完整的關鍵,粒線體 是細胞製造能量的工廠,對肌肉和神經細胞尤其重要。一旦缺少Cisd2時,
粒線體破損、功能失調,老化徵狀接踵而來,而使用抗氧化劑活化Cisd2 的小黑鼠,代謝功能好,雄鼠還能在十二個月後罕見地讓雌鼠受孕,證實 了抗氧化劑可調控生物體內的 Cisd2基因,抑制氧化,並進一步達到防止 老化,延長壽命的功效(Chen et al., 2009)。
六、抗氧化劑之種類
可分為酵素型抗氧化劑與非酵素型抗氧化劑 。
1. 酵素型抗氧化劑
(1) 超氧岐化酵素 ( superoxide dismutase, SOD ) 於體內中以兩種型式存 在,以銅/鋅為輔因子 ( Cu, Zn/SOD ) 存於細胞質中;或以錳為輔因子 ( Mn/SOD ) 存於粒腺體間質 ( Kurechi et al., 1980 )。其反應為歧化反應 ( dismutation reaction ),主要在清除超氧陰離子 ( O2
-‧),產生的 H2O2 會 被觸酶及麩胱甘肽過氧化酶代謝成 H2O。反應式如下:
SOD 2 O2
-+ 2 H+ → H2O2 + O2
(2) 麩胱甘肽過氧化酶 ( glutathione peroxidase, GSHPx )
GPx 的反應中心為硒 ( Se ),在體內負責清除 H2O2,藉催化還原態 glutathione ( GSH ) 將 H2O2 或 ROOH 還原成 H2O 或 ROH,其反應式 為 ( Bondorden and Pariza, 1994 ):
2GSH + H2O2 → GSSG + 2H2O 或
2GSH + ROOH → GSSG + ROH + H2O
可降低細胞內氧化壓力的形成 ( Halliwell 1994 ),亦可避免紅血球受到 H2O2 的攻擊,抑制血紅素的鐵離子釋放。
(3) 觸酶( Catalase, CAT )
觸酶為存在於細胞膜內過氧化小 體 ( peroxisome ) 中的含鐵酵素 ( Meir et al., 1995 ),在細胞內會催化 H2O2 分解成 H2O 及 O2,其反應 式( Bondorden and Pariza, 1994 ):
Catalase
2 H2O2 → O2 + 2 H2O
2. 非酵素型抗氧化劑
此類型的抗氧化劑,主要有天然抗氧化劑、人工抗氧化劑及其他。
其分類如下:
(1) 天然抗氧化劑
生物體中含有多種清除活性氧或自由基的抗氧化物質 ,包括維生素 E、維生素 C、類胡蘿蔔素、微量元素、類黃酮素、藻藍素及其他一些酚 類物質,可當作還原劑、金屬螯合劑或活性氧清除劑而終止自由基引起的 油脂過氧化反應。
A、 維生素 E
維生素 E 化學名為:tocophero1,是一種脂溶性的抗氧化物 ,可以 有效地抑制脂質過氧化作用,一般認為 d 型最具抗氧化性,抗氧化作用 是發生在細胞膜或脂肪組織,主要是利用苯環上的羥基,使羥基的氫原子 易被自由基奪走而成為a-tocophero1 radical,此複合物穩定性高攻擊其他 分子機率大幅降低(Sridevi et al., 2007)。
B、 維生素 C
維生素 C 是一種水溶性抗氧化劑又稱抗壞血酸。因抗壞血酸容易氧 化,所以如有其它易氧化的化合物共同存在,抗壞血酸會先行貢獻自身之 氫原子給對方而被氧化,因而保護其他化合物,其天然型式可當做還原劑 或氧去除劑。抗壞血酸在人體內生化氧化過程中扮演輔助還原劑,會與亞 硝酸鹽反應,避免亞硝酸鹽與體內蛋白質反應產生致癌性之亞硝胺化合 物 。流行病學的研究亦顯示多食用維生素 C 具有防癌效果 ( Kitts 1997 )。
C、 類胡蘿蔔素
類胡蘿蔔素 ( carotenoid ) 是一種脂溶性、含 40 個碳之植物色素,
可分為兩群:含 C 和 H 之胡蘿蔔素及含氧之胡蘿蔔素 ( 葉黃素 ) 做 為抗氧化劑,類胡蘿蔔素可終止自由基的連鎖反應,它可與苯氧自由基作 用終止自由基的連鎖反應,並與苯氧自由基作用而形成較安定的類胡蘿蔔 素自由基,以防止蛋白質和 DNA 等之氧化變異(Toni et al., 2008)。
D、 葉綠素
葉綠素 ( chlorophyll ) 是植物體各器官含有之綠色色素,可吸收太陽 能而進行光合作用,有四種型式:chlorophyll a,chlorophyll b,phenophytin a 及 phenophytin b。在黑暗條件下,其在甲基亞麻油酸中之抗氧 化性大 小依序為 chlorophyll a > chlorophyll b > phenophytin a > phenophytin b,且 chlorophyll a 之抗氧化性比 BHT(Butyl hydroxytoluene) 之效果還好( 王 2002 );其抗氧化作用可能是藉由提供氫原子來清除過氧化自由基而抑制 脂質氧化。
E、 藻藍素 ( C-phycocyanin, C-PC )
藻藍素是一種藻膽色素蛋白質 ( phycobiliprotein ),為藍菌中主要成 分。此ㄧ蛋白質色素已被證實具有抗癌、自由基清除、抗氧化及抗發炎的 特性(廖 2006),並於後詳述。
七、藻藍素
藻藍素 ( C-phycocyanin, C-PC ) 為藍菌中的主要成分之一,屬於光合 作用色素 ( photosynthetic pigment ),是一種可溶於水且具有螢光的藻膽 色素蛋白質 ( phycobiliprotein )。最大吸收波長 ( absorbance maximum ) 約為 615 ~ 620 nm ( MacColl and Guard-Frair 1987; Bryant 1982 ),最大螢 光 散 射 波 長 ( fluorescence emission maximum ) 約 為 637 ~ 650 nm ( MacColl and Guard-Frair, 1987; Oi et al., 1982 )(圖八)。許多藍菌都含有 C-PC 的成分。
C-PC 單元體 (αβ-monomer ) 結構與葉綠素、血紅素類似,主要 由兩個次單元 ( subunit ) 構成:α 次單元,約 15,900 ~ 18,500 daltons;β 次單元,約 18,300 ~ 20,000 daltons。一般 C-PC 單體會以( αβ )3-trimer 或 ( αβ )6-hexamer 的型式存在。每一個 C-PC 單元體中都含有三個發色基 團 ( chromophore ),一個位於 α 次單元 ( α-84 position ),兩個位於 β 次 單元 ( β-84 和 β-155 positions )( Stec et al., 1999 )。此發色基團在藍菌中 稱 之 為 膽 素 ( bilin ) , 而 位 於 C-PC 單 元 體 中 的 又 稱 之 為 phycocyanobilin 。其主要結構為 open chain linear tetrapyrrole ,並利用 thioether bond 以 共 價 鍵 結 的 方 式 與 各 個 C-PC 次 單 元 之 輔 基 蛋 白 ( apoprotein ) 上的半胱氨酸 ( cysteine residue ) 相連結 ( Jacobi et al., 2000; Padyana et al., 2001 )( 圖 九 ) , 且 其 結 構 中 並 未 螯 合 金 屬 元 素 ( Duerring et al., 1991 )。一般而言,C-PC 除了單獨存在於藍菌細胞內,
也會和其他的藻膽色素蛋白質結合形成胞器 ( organelle ),稱之為藻膽體 ( phycobilisome )( Gantt and Conti 1965, 1966a, b; Gantt et al., 1976 )。藻膽 體由兩個主要區域 ( domain ) 組成:一個核心區域(core domain)和多個桿
狀區域(rods domain)。核心區域含有異藻藍素 ( allophycocyanin ),並且依 附在類囊膜上;桿狀區域含有藻藍素和藻紅素 ( phycoerythrin ),位於類 囊膜的外側,而藻藍素則介於藻紅素及異藻藍素之間(圖六)。各區域內的 藻膽色素蛋白質及各區域之間以多功能性聚胜肽 ( polypeptide ),稱之為 linker,組織起來形成核心及桿狀區域並將兩個區域相互連接 ( Tandeaude Marsac and Cohen-Bazire, 1977 )。除此之外,linker 亦可將藻膽體固定依 附在類囊膜上。
圖八.不同色素之吸收光譜。(source: www.egbeck.de)
圖九.藻藍素( C-phycocyanin, C-PC ) (A) α-subunit 以及 (B) β-subunit (C) 藻藍素發色團 Structure of open chain linear tetrapyrrole of
phycocyanobilin in C-phycocyanin.(Jacobi et al., J Org Chem. 2000; 65:
8478-8489;Padyana et al., Biochem Biophys Res Commun. 2001; 282: 893-898).
在藍菌中 C-PC 主要的功能是幫助葉綠素吸收來自外界的光 能。藍 菌一般都生長在水中,所以行光合作用需要的光線會被環境中 的水所阻隔 而改變波長並且影響了葉綠素吸收光能的效力。因此藍菌細胞就衍生出一 套補救的機制,也就是利用藻膽色素蛋白來補強光能 的吸收。以鞘絲藍菌 為例,藻藍素 ( C-phycocyanin ) 位於細胞內類囊膜 ( thylakoid membrane ) 的外側幫助葉綠素a ( chlorophyll a ) 吸收來自外界或上游光合色素傳遞 下來的光能,接著在將光能轉移到類囊膜內側的葉綠素。最後,葉綠素便 藉由電子傳遞鏈的機轉將光能轉換成化學能以進行光合作用 。
八、藻藍素的生理功效
由於 C-PC 屬於天然色素,使用在日常生活中已有多年的歷史 。在 日本、泰國及中國大陸就有將 C-PC 使用在食品 ( 如:口香糖、乳製品、
果凍等 ) 及化妝品 ( 如:口紅、眼線膏 ) 中當作著色劑的例子 ( Sarada et al., 1999 )。另外,C-PC 也有發展為螢光探針 ( phycofluor probes ) 應 用在免疫診斷 ( immunodiagnostics ) 上的例子 ( Kronik et al., 1983 )。除 此日常生活與技術上的應用 ,亦有文獻及研究報告指出 C-PC 具有許多 醫療上的功效(廖 2006)。
目前已知藻藍素具有抗氧化能力 (Romay and Gonzalez, 2000 ),能清 除活性氧自由基 ( Romay et al., 1998a; Romay et al., 2001 ),亦能抑制四氯 化 碳 和 Fe2+-ascorbic acid 所 引 起 的 粒 腺 體 脂 質 過 氧 化 ( Bhat and Madyastha, 2000 )。對 carrageenan 誘導之老鼠足蹠浮腫和花生四烯酸誘 導老鼠耳朵發炎反應下,藻藍素具有抗發炎的效果 ( Romay et al., 1998b;
Romay et al., 2001 ),其透過的可能作用機轉為清除氧自由基 ,抑制 cyclooxygenase-2 ( COX-2 ) 的表現 ( Reddy et al., 2000 ),並進一步抑制 Prostaglandin E2 ( PGE2 )及Leukotriene B4 ( LTB4 )的生成( Romay et al., 2000 ) 和降低 myeloperoxidase ( MPO ) 的活性等。藻藍素也會抑制經由 Lipopolysaccharide ( LPS ) 感染的小鼠血清中發炎性 tumour necrosis factor-α ( TNF-α ) 的含量 ( Romay et al., 2001 )。
参、實驗流程
Lyngbya sp.
培養於固態 C-10 培養基 小量純化
培養於液態 C-10 培養基 大量培養
分離前處理
藻藍素萃取條件 方法之開發
藻藍素特性分析 定量分析
1.蛋白質定量 2.吸收光譜分析 定性分析
1.分子量鑑定 2.光譜特性分析
藻藍素細胞抗氧化能力分析
1.過氧化氫誘導氧化細胞模型的建立 2.藻藍素對過氧化氫誘導細胞氧化之影響 3.維生素C對過氧化氫誘導細胞氧化之影響 4.維生素E對過氧化氫誘導細胞氧化之影響 5.β-carotene 對過氧化氫誘導細胞氧化之影響 6.藻藍素抗氧化能力分析
肆、材料與方法
一、實驗材料
試藥
品名 廠商
Bradford reagent、Protein marker Bio-Rad Laboratories
Bovine serum albumin (BSA) Sigma
Disodium hydrogen phosphate(Na2HPO4) Shimakyu Dulbecco,s modified eagle medium(DMEM) GIBCO
Methanol Shimakyu
Fetal bovine serum(FBS) GIBCO
Hydrogen peroxide(H2O2) 誠意興
Ampicillin + Streptomycin (AS) GIBCO
Potassium dihydrogenphosphate (KH2PO4) Shimakyu Phycocyanin Sigma Sodium carbonate anhydrous(Na2CO3) Shimakyu Sodium Dihydrogenphosphate(NaH2PO4) Shimakyu Sodium dodecyl sulfate(SDS) Sigma
Sodium bicarbonate(NaHCO3) Sigma
Sodium pyruvate Sigma
Trypsin Biowest
Trypsin-EDTA Biowest
Coomassie blue R250 Merck
二、實驗儀器
分光光度計 (UV-160A, Shimadzu, Tokyo,Japan; Shimadzu) 恆溫振盪培養箱 (S300R, Firstek)
高速離心機 (Himac CR 21,Hitachi, Tokyo, Japan; Hi tachi) 蛋白質電泳槽 (Bio-Rad Laboratories)
直立式顯微鏡 (Nikon YS100)
倒置式顯微鏡 (Nikon eclipse TS100)
細胞培養箱 (Nuaire US autoflow CO2 water-Jacketed incubator) 光合作用培養箱 (Firstek Model-TG5)
冷凍乾燥機 (Kingmech FD3-12P)
三、藍菌純化與培養方法(李 2005)
3.1 培養基配製:
培養基方面,採用淡水藍菌培養基培養,此種培養基材料取得容易,
製作時不麻煩且較為廉價,保存時間久,對一般藍菌生長效果良好等,故 選之使用。
3.1.1 淡水藍菌培養基:
本實驗使用之培養基為 C-10 培養基 (附錄一),將之配置好後,
以每 L 中加入 1 ml 之比例加入微量元素 (附錄一),再進行高溫高壓 消毒,可於 4 oC 保存 3 個月;微量元素配置完成後,以 10 % NaOH 調整 pH 至 8.2 ,並定量分裝,高溫高壓滅菌後,可於 -20oC 保存 一年以上。
3.2 藍菌培養
3.2.1 實驗室培養:
實驗室中以固態以及小量液態培養為主 ,固態培養可用於純化 藍 菌,而液態培養可用於將純化後的藍菌大量增殖 。
3.2.1.1 固態培養基培養法:
培養藍菌時將 C-10 培養基加入 1.5 % 的 agar,配製好後,以 autoclave 進行高溫高壓滅菌,121oC、15min,待其滅菌完成後,冷 卻倒入培養皿中,配製好的培養基作為純化藍菌使 用。
接種菌原,取菌種 100 μl,用平面培養基以 L 型玻棒沾酒精後 過火消毒,待冷卻後將菌種均勻塗布於培養基上,接種完成後將培養 基倒放置入光合作用培養箱,光源採用日光燈,因日光燈的光波波長 多在藍菌的各種色素吸收範圍內 。
3.2.1.2 液態培養基培養法 (小量培養):
將固態培養基生長一定數量之鞘絲藍菌,取些許樣本於顯微鏡下 觀察是否為預期菌種以及純度,若菌種純度夠且為預期的鞘絲藍菌 , 即可於無菌操作台中以圓形接種環將固態培養基上的鞘絲藍 菌刮下 並接種菌原至內含已配置好且滅菌消毒之液態 C-10 培養基之三角 錐形瓶中 ( 500 ml 錐形瓶盛 150 ~ 250 ml;1000 ml 錐形瓶盛 300 ~ 500 ml,即培養液約錐形瓶標式容量之 30 % ~ 50 % ),接種完瓶口 塞以棉花以便通氣,再放置到光合作用培養箱 (培養箱需具有震盪裝 置,震盪之轉速約為100 rpm / min)。光線強度,以一般日光燈照明,
光線強度為 500-2000 lux。於室溫下震盪培養,使其生長約一至二個 月,待其生長數量足夠,即可進行下一步實驗。
3.2.1.3 大量培養:
若將純菌分離出之後,可先增量,再移至大型培養槽中培養大 量,小型增量之方式:
膠桶打氧式:準備 20 L 透明塑膠方桶,於內倒入培養液,再加入已 純化出之藍菌 ( 最終濃度為 2g[紗布過慮濕重]/L 以上 ),並以小馬 達打空氣 ( 使水流動並使空氧中 CO2 進入水中 ),並以 300 lux 燈 光照之長期培養。若欲其快速生長可使用 1500 lux 燈光。
3.3 藍菌純化
取欲純化之藍菌菌種,以前述之 L 型玻棒於固態平面培養基塗勻 , 待其菌群生長之後,以燒紅消毒之無菌接種環 ,取出小量於培養液中稀 釋,再塗於平面固態培養基,如此反覆數次,可得到較純之藍菌菌群。
3.4 藍菌保存
藍菌的保存,可將液態培養基培養之菌體,先離心濃縮,再加入總體 積 10 % 的抗凍劑DMSO (dimethyl sulfoxide)或15 %的甘油(glycerol),裝 入冷凍保存細胞用的 冷凍小管中 (plastic ampule),再保存於液態氮中 ( -196 oC );另外亦可保存於 -70 oC,但需注意,無論保存在 -70 oC 或 -196 oC,均得逐漸降溫,先將冷凍小管置於 4oC、1 小時,再置 -20 oC、
1 小時,再置入 -70oC或 -196oC長期保存,或用自動逐步降溫冷凍儀降 溫後凍存。而前述之斜面固態培養法,待菌群生長好後,以蠟將蓋口封緊,
置光合作用培養箱,並將照度減低為正常培養的 1/2,約能保存 3 個月 以上。
四、藻藍素分離前處理
藻藍素分離的前處理步驟有許多方式 ,首先必須打破藍菌細胞,再 進行藻藍素的萃取 ,打破藍菌細胞 有幾種主要方式 :以高壓破碎裝置 (French Press)將細胞打破(Tandeau and Cohen, 1977 );以十二烷基二甲基 氧化胺(Lauryl Dimethyl Amine Oxide)處理( Barbara et al., 1985 );日本學 者也有人以超音波震碎的方式萃取( Takao et al., 2003 );在Jayant 的實驗 中,則是先以高溫250oC將菌體乾燥並以磷酸緩衝液萃取 ( Jayant 2005 );
而近幾年較為常見的方式是先以溫差反覆凍融造成細胞破裂,再以硫酸銨 分餾、Sephadex G-150 進行大小排除層析法以及DEAE (diethylaminoethyl) 纖維素進行離子交換層析法( Badrish et al., 2006 );另外也有以溫度影響 以及生物質量與溶劑比值 (biomass-solvent ratio) , 以不同條件來處理 ( Silveira et al., 2007 );而在Priya的實驗中,以研缽研磨的方式利用玻璃 均質器均質後,再加入聚丙烯處理(Priya and Christopher, 2007 )。
而本實驗則是選擇先以研缽將藍菌磨碎後 ,離心取得深藍色的上清 液,先進行冷凍乾燥,再以藻藍素不溶於甲醇之特性,進行後續純化的步 驟。
4.1 冷凍乾燥法
取大量培養後的藍菌,以抽氣過濾法去除培養液並抽乾,將過濾抽乾 後的藍菌秤乾重,再將乾燥後的藍菌置入研缽,加入少量蒸餾水磨碎,將 磨碎後的藍菌,倒入離心管中,離心 ( 10000 rpm,30 min ),離心之後取 上清液置入新的離心管中,再冷凍乾燥濃縮成粉末。
4.2 膠體過濾層析法
取大量培養後的藍菌,連同培養液倒入離心管中離心 ( 5000 rpm,10 min ),倒掉上清液,取離心後的藍菌置入研缽中加入適量蒸餾水磨碎 , 將磨碎後的藍菌,倒入離心管中,離心 ( 10000 rpm,30 min ),離心之後 取長 30cm 內徑 1.5cm 內含 Silica gel (Merck silica gel 60)之管柱並將 上清液注入層析管柱。
五、藻藍素萃取條件方法開發
5.1 冷凍乾燥法
取前處理過後得到的乾燥藍菌粉末,加入甲醇萃取,震盪混勻,倒出 萃取液留下沉澱物,再加入甲醇反複震盪萃取數次 ,最後離心 ( 5000 rpm,10 min ),倒出上清液,得到藍色沉澱物,再加入適量蒸餾水即可將 之溶解,溶解之深藍色液體即為藻藍素水溶液 。
5.2 膠體過濾層析法
取一 2.5 x 30 cm 的層析管柱,內填充矽膠 ( Merck silica gel 60 ),
約管柱的 2/3 高,將管柱裝置好,以滴管沿管壁緩緩加入前處理好的藍 菌上清液,待其與矽膠作用約 10 分鐘之後,利用中壓層析儀分離,分離 條件移動相使用分析級的甲醇,流速設定為 10 ml/min,將藻紅素、藻膽 蛋白等物質透過甲醇分離出,藻藍素則留於矽膠層中,待沖提結束後,以 刮勺將矽膠取出,置入三角錐型瓶中,加入適量蒸餾水與矽膠混勻,並以
超音波震盪機震盪分離,藻藍素即可與矽膠分離溶於蒸餾水中,反覆加入 蒸餾水混勻震盪,收集藻藍素溶液,收集後的藻藍素溶液以濾紙過濾除去 矽膠殘渣,再將過濾後的藻藍素溶液,進行冷凍乾燥,即可得到藻藍素粉 末。
六、藻藍素特性分析
6.1 藻藍素定量分析
6.1.1 蛋白質定量 ( Bradford 1976 ) 蛋白質標準品檢量線製作:
以 Bradford 方法,以胎牛血清白蛋白 (bovine serum albumin;
BSA) 為蛋白質標準品,精確稱量 50 mg,用二次水 1 mL 溶解 後,稀釋成一系列濃度 ( 500 µg/mL、400 µg/mL、300 µg/mL、200 µg/mL、100 µg/mL ),取 20 µL 的蛋白質樣品與 1 mL 的 Bradford 染劑(稀釋 5 倍),混勻 5 分鐘後,放入分光光度計 590 nm 測定 其吸光值平均值,依並 OD value (Y) 對蛋白濃度 µg/mL (X),求出 趨勢線方程式 y = 0.359x + 0.0751 R2 = 0.9992。
樣品蛋白質定量:
取 20 µL 的蛋白質樣品與 1 mL 的 Bradford 染劑 (稀釋 5 倍),混勻 5 分鐘後,放入分光光度計 590 nm 測定其吸光值平均 值,利用檢量線公式則可求得實際蛋白質樣品濃度 。
6.1.2 吸收光譜分析
將藻藍素標準品以磷酸緩衝液稀釋成終濃度 100, 200, 500, 700, 1000 µg/ml 之溶液,並將藻藍素樣品稀釋 5 倍後,測定標準品及樣品 在 620 nm 處之吸光值,作出標準曲線後即可求出樣品中之藻藍素濃 度。
6.2 藻藍素定性分析 6.2.1 分子量鑑定
本實驗採用 SDS 聚丙烯胺膠體電泳( SDS-polyacrylamide gel electrophoresis ) 之方法分析。利用 bio-rad SDS-PAGE 標準品進行分 子量估算,膠片採用Bio-rad Ready gels 4-20% Tris-HCl。
6.2.1.1 電泳
將藻藍素標準品 (C-phycocyanin, Sigma P-2172) 及藻藍素樣品 約 10 μl 和 10 μl 之 sample buffer 均勻混和後,置於沸水中五分 鐘 , 再注 入膠體中 。 電泳 分析所使 用的緩 衝溶液為 SDS-PAGE running buffer。以 80 V 設定使樣品移動至堆積凝膠及分離凝膠之界 面後,同樣以 120 V 使樣品移動至分離凝膠末端時 ,即可取下分離 凝膠進行染色。
6.2.1.2 染色及退染
將分離凝膠取下後,放入塑膠盒中。先倒入適量 SDS-PAGE 染 劑 (Coomassie blue),於震盪器上搖晃 30 分鐘進行染色,之後將染 劑倒掉,再以適量退染液 (destain solution),進行退染,將其置於震
盪器上搖動至藍色背景退去後將分離凝膠 保存。
6.2.2 光譜特性分析
將藻藍素標準品與樣品以 磷酸緩衝液稀釋至濃度為 100 µg/ml,
測定其在波長 250-700 nm 之吸收,並與標準品之最大吸收波長比對。
七、藻藍素對細胞抗氧化能力分析
本實驗利用 Vero cell:非洲綠猴腎臟上皮正常細胞 (African green monkey kidney) 來探討藻藍素對細胞抗氧化能力 的影響,Vero cell生長快 速且易於培養,為藥理實驗上常用之細胞株 ,故選其使用。
7.1 細胞來源
本實驗室所採用之細胞株購自食品工業發展研究所 ,BCRC 60013 (BCRC Number:60013)。
7.2 細胞培養
7.2.1 製備培養基
Vero 細胞培養基為 MEM (Minimum Essential Medium) ,不含 L-glutamine、sodium pyruvate,購自Sigma公司。將 MEM (10g) 溶於 900 ml 的蒸餾水,攪拌均勻後加入 1.5g Na2HCO3 以及 1g sodium pyruvate 混勻,然後用 1N HCl 滴定至 pH 7.3~7.4,在無菌操作台內 用 0.22 μm 過濾膜無菌過濾,平均分裝到 2 個 500 ml 血清瓶,每
500 ml 培養基再加入 200 mM L-glutamine 2.5 ml,取 5 ml 於細胞培 養箱做空白測試 ,24小時候若無汙染再將 Fetal bovine serum 加入 MEM 中 佔 總 體 積 的 10% , 存 於 4℃ , 要 使 用 之 前 再 加 入 抗 生 素 Ampicillin 50 µg/ml 以及 streptomycin 30 µg/ml。
7.2.2 冷凍細胞活化
冷凍細胞之活化原則為37oC快速解凍,以避免冰晶對細胞造成傷 害。細胞活化後,約需數日,或繼代一至二代,其細胞生長或特性表 現才會恢復正常。
將新鮮培養基置於 37°C 水槽中預熱,以 70% ethanol 擦拭外 部,移入無菌操作台內。自液態氮容器中取出冷凍管,立即放入 37°C 水槽中快速解凍 ,輕搖冷凍管使其在 1~2分鐘內全部融化 ,以70%
ethanol 擦拭保存管外部,移入無菌操作台內。於無菌操作台內取適量 培養基加入細胞培養皿中,緩慢加入已解凍之細胞懸浮液 ,一般稀釋 比例為 1:10~1:15,與培養基混合均勻後,放入培養箱培養。可另取 樣少許解凍細胞懸浮液作存活測試 。解凍後是否立即去除冷凍保護劑 Dimethyl sulfoxide (DMSO),依細胞種類而異。本實驗之細胞株,不需 要立即去除冷凍保護劑,待第二天確定細胞生長或貼附良好後再去除 即可。惟極少數因對 DMSO 敏感或會造成細胞分化之細胞 ,需立即 去除 DMSO,若要立即去除,則將解凍之細胞懸浮液加入含有 5~10ml 培養基之離心管內,離心 1000 rpm、5 分鐘後,移去上清液,加入新 鮮培養基,混合均勻,放入培養瓶內,置於適當之培養箱培養。若不 需立即去除冷凍保存劑,則在解凍培養隔日後更換培養基即可 。
7.2.3 細胞繼代培養
細胞生長至高密度時,即須收集細胞,分殖至新的培養皿中,其 稀釋比例 1:2 或 1:6。收集細胞方法 係用 trypsin-EDTA 及離心處 理。Trypsin-EDTA 為收集吸附型細胞常用之方法。Trypsin 為胰蛋白 酵素,其作用為分解細胞與盤底之附著蛋白 ,EDTA 其作用為去除 Ca2+、Mg2+ 等離子,trypsin 與 EDTA 二者共同作用,可使附著之細 胞自盤壁脫落。Trypsin-EDTA 作用時間勿太久,否則可能對細胞造成 傷害或死亡。敲拍培養瓶不可太過猛烈 ,避免對細胞造成傷害或造成 培養瓶裂痕而污染。每一種細胞在繼代培養時,都須先測試 trypsin 對 細胞的影響,假如可以只用 trypsin 就可使附著之細胞自盤壁脫落 , 就不需要用 Trypsin-EDTA,避免對細胞傷害過大。
首 先 吸 除 舊 培 養 液 , 用 PBS 洗 滌 細 胞 一 至 二 次 , 再 加 入 trypsin-EDTA 2 ~ 3 ml,移至細胞培養箱,待數分鐘後細胞自盤底脫落 後,加入適量含血清之新鮮培養基 ,以終止 trypsin 作用,因血清中 含有 trypsin inhibitor,可以終止 trypsin 作用,離心後去除上清液,
移至新的培養盤中,依正常條件繼續培養之。
7.2.4 細胞計數方法
計算細胞數目用紅血球計數盤,血球計數盤有二個 chamber,每 個 chamber 中細刻 9 個 1 mm2 之大正方形,其中位於 4 個角落之 正方形再細刻 16 個小格,深度均為 0.1 mm。當 chamber 上方蓋上 蓋玻片後,每個大正方形之體積為 1 mm2x 0.1 mm = 1.0 x 10-4ml 。使 用時,計數每個大正方形內之細胞數目,乘以稀釋倍數,再乘以 104, 即為每 ml 中之細胞數目。利用染劑 (例如 trypan blue) 會滲入死細
胞中而呈色,而活細胞因細胞膜完整,染劑無法滲入而不會呈色。取 20 µl 細胞懸浮液與 20 µl trypan blue 等體積混合均勻。取少許混合液 (約10 µl) 自紅血球計數盤 chamber 上方凹槽加入,蓋上蓋玻片,於 100 倍倒立顯微鏡下觀察,活細胞不染色,死細胞則為藍色。計數五 個大方格之細胞總數,再除 5,乘以稀釋倍數,最後乘以 104,即為 每毫升中細胞懸浮液之細胞數 。
細胞存活率的算法公式如下:
細胞存活率%=活細胞數/(活細胞數+死細胞數) × 100%
7.3 過氧化氫誘導細胞氧化模型的建立
將繼代二次以後的細胞,吸除培養皿中原細胞培養液,用 PBS 洗滌 細胞一至二次,再加入 trypsin-EDTA 2 ~ 3 ml,移至細胞培養箱,待數分 鐘後細胞自盤底脫落後,移至 24 well 培養盤中培養,每 well 中細胞數 為 1 x 105 以上,培養至隔日後,移除原培養液,加入含不同濃度的 H2O2
之 MEM 培養液,H2O2 濃度設定分別以 9%、7%、5%、3%、1%、0.5%、
0.1%、0.08%、0.05%、0.04%、0.03%、0.02%、0.01%、0.025、0.0125%、
0.00625%,於37℃,5% CO2 條件下培養,培養12小時,每種濃度培養細 胞進行4重複,觀察細胞在不同農渡之 H2O2 作用下的形態變化,測試細 胞氧化誘導之 H2O2 最適濃度(可使細胞產生氧化壓力之最低濃度 )。
7.4 藻藍素對過氧化氫誘導細胞氧化之影響
找到最適誘導細胞氧化條件後,參考(Liu et al., 2008)分兩部份進行藻