國立臺灣大學生命科學院生化科技學系暨研究所 碩士論文
Department of Biochemical Science & Technology College of Life Science
National Taiwan University Master Thesis
感覺型視紫質 SRM 和其傳導元 HtrM 對 嗜鹽古生菌光趨性之影響
The Functional Impact Of Photosensory Rhodopsin SRM-HtrM In The Phototaxis Response Of Haloarchaea
陳政良
Jheng-Liang Chen
指導教授 :楊啓伸 博士 Advisor : Chii-Shen Yang, Ph.D.
中華民國 106 年 7 月
July 2017
目錄
目錄………..…….…..………..……..………. i
圖目錄………..…….……..….……… vi
表目錄………...…………... viii
摘要………..………...………. ix
Abstract……….…..……… x
第一章 緒論………..………... 1
第一節 嗜鹽古生菌之介紹………..…..………... 1
第二節 微生物視紫質……….….….…...…….. 2
第三節 古生菌感覺型視紫質………...………..…….……. 7
I-3.1 紫外光/可見光譜……….……...………...….. 7
I-3.2 光週期………...…….………….…..8
I-3.3 質子傳輸………...…….…………...……9
I-3.4 重要胺基酸………...…….…...………11
I-3.5 演化探討………...…..….…………13
第四節 傳導元 (Transducer) 的結構與特性…………...….….……..…… 14
第五節 嗜鹽古生菌的光趨性………...……….….….…… 16
I-5.1 SRs 於嗜鹽古生菌中的光趨性………...…...…....….……… 16
I-5.2 嗜鹽古生菌的光趨性………...….…….….……16
I-5.3 SRs 調控光趨性之機制………...……….…..….….……... 18
第六節 研究動機及策略………...…….……….……..…………... 20
第二章 材料與方法………...……...………...……. 22
第一節 生物試劑………..……...…………...…… 22
II-1.1 菌種………...…….….……… 22
II-1.2 質體………....…….……….. 23
II-1.3 蛋白質藥品………...…..…...…...…………..…… 23
第二節 化學藥品………...…..…….……….………. 23
第三節 儀器設備………...…..….…..….………... 25
II-3.1 核酸電泳……….……...…..…….… 25
II-3.2 蛋白質電泳及轉印………...………….…….… 25
II-3.3 離心機……….…………..…...…...… 25
II-3.4 光學設備…………...…..……….…..…….…… 26
II-3.5 恆溫培養箱………...…..………...….…… 27
II-3.6 酸鹼度計………...…..……….…..….…… 27
II-3.7 其他……….……...…..…...…… 27
第四節 實驗方法………...…..……….…..…… 28
II-4.1 生物資訊學分析………...…..……… 28
II-4.1.1 基因資料庫………...…..………….…..…… 28
II-4.1.2 序列親緣比對……….……...…..…..…… 28
II-4.1.3 蛋白質結構分析……….………...………… 28
II-4.2 DNA 建構及轉形……...…..………..….….. 29
II-4.2.1 小量核酸萃取…………...…..……….…….…… 29
II-4.2.2 聚合酶鏈鎖反應 (PCR) …………...…..…….…..……….. 29
II-4.2.3 DNA 膠體純化………...…..…….……… 30
II-4.2.4 限制酶截切………...…..……...…… 30
II-4.2.5 DNA 黏合………...….…....….…. 30
II-4.2.6 大腸桿菌轉形………...…..…….…..… 30
II-4.2.7 嗜鹽古生菌 H. salinarum 轉形…………...…..….…..….… 31
II-4.2.8 轉形株鑑定...…..……….….. 32
II-4.3 重組蛋白質之表現及純化….…..……….……… 32
II-4.3.1 重組視紫質表現…………...…..……….……..… 32
II-4.3.2 重組可溶蛋白質表現………...…..……….…..… 33
II-4.3.3 重組視紫質純化………...…....…. 33
II-4.3.4 重組可溶蛋白質純化 (Hexa-His-tagged) ...…..…….….… 34
II-4.3.5 重組可溶蛋白質純化 (GST-tagged)……...…..…….…….. 34
II-4.3.6 H. salinarum 細胞膜之純化……...….……… 35
II-4.4 蛋白質定量及定性…...…………..……….…….… 35
II-4.4.1 蛋白質定量………...……….…… 35
II-4.4.2 蛋白質變性電泳………..…...…..………….……… 36
II-4.4.3 蛋白質原態電泳………..…………...…....……… 37
II-4.4.4 蛋白質轉印………...…..…………..……….……… 38
II-4.4.5 免疫呈色………...…..…..……….……… 38
II-4.5 感光蛋白質光學分析………....…..…….……… 38
II-4.5.1 吸收光譜測定………..………...…..…. 38
II-4.5.2 視紫質光週期量測………...…..…..………….……… 39
II-4.5.3 光驅動離子幫浦活性測定………...…..….…… 39
II-4.6.1 泳動菌株挑選………...…...…………....…… 39
II-4.6.2 顯微鏡觀察………...…..……...…..……… 40
II-4.6.3 光趨性分析……….…..……….……… 41
II-4.6.4 光照生長曲線……...……….……… 41
第三章 實驗結果………...…..……….….... 43
第一節 蛋白質性質…...…..………...…….….…… 43
III-1.1 序列比對…………...…..……….….….…… 43
III-1.2 結構模擬………...…..……...……….…..….……… 44
III-1.3 pI 及 pKa………...…..…….…...….…… 46
III-1.4 光誘發質子傳輸……….………...…...…… 49
III-1.5 光週期……...………...……. 50
第二節 微生物光趨性研究方法…...…..……….…....… 51
III-2.1 顯微鏡觀察………...…..….……….…...…..… 51
III-2.2 ELISA Reader 測定………..…………...………...………. 56
第三節 SRM 及 SRM-HtrM 之移植………....……...……...…. 60
III-3.1 DNA 確認…………..……….……..…….. 60
III-3.2 蛋白質表現確認………....…..………..……….……. 60
III-3.3 SRM 功能性測試………...…..………..…………...…… 62
第四節 嗜鹽古生菌之生理探討………...…..……...…..…… 64
III-4.1 細胞型態……….…...…...….….. 64
III-4.2 生長曲線………...…..………...…… 65
III-4.3 光趨性研究………...…..…………...………… 67
第四章 結論與探討………...…..……….…….….…….. 70
第一節 微生物之光趨性研究…………...…..………....……… 70
第二節 光趨性在生理上之意義…………...…..………..….….…… 71
第三節 H. salinarum 的 SRM-HtrM 功能獲得型轉殖株………...…... 72
第四節 嗜鹽古生菌的光趨性機制………...………...…… 73
第五章 未來展望………...…..………..….……….. 74
第一節 感覺型視紫質的轉介…………....…..………...…….………… 74
第二節 光趨性機制之研究方向………....…...………… 74
第六章 參考文獻………...…...… 76
附圖………..……...…...….83
圖目錄
圖1 : 微生物視紫質之共通特性………...…..…….……...… 2
圖2 : 嗜鹽古生菌中視紫質的分佈………...…..………...……….……... 4
圖3 : 嗜鹽古生菌中感覺型視紫質與其傳導元………...………...….… 6
圖4 : H. salinarum 感覺型視紫質的質子傳輸現象………...…..……...….…… 10
圖5 : 感覺型視紫質重要胺基酸比對……….…...…..………..… 12
圖6 : 傳導元之結構及比較………...…..……….………..…… 14
圖7 : 菌體泳動分析………...…..……….…….….……… 17
圖8 : NpSRII 傳遞避光反應的訊息路徑………...……..……… 19
圖9 : 實驗流程表………..………...…….…………..… 21
圖10 : 感覺型視紫質之間的蛋白質多序列比對……….……....………….… 45
圖11 : HmCheR 及 HmHtrM 之模擬結構 ……….………...……… 46
圖12 : SRM 及其複合蛋白質之 pKa 測定……….………..…….… 47
圖13 : 原態等電聚焦電泳……….………...….….. 48
圖14 : SRM 及 SRM-HtrM 的氫離子傳輸能力……….………….…..…… 49
圖15 : 顯微鏡之架設……….……….….……... 52
圖16 : LED 燈源之波譜……….……….………...… 53
圖17 : 樣品薄片……….……….…...….……… 53
圖18 : 顯微鏡刺激光源的照射範圍……….……….……… 54
圖19 : 光源刺激時間……….……… 55
圖20 : 光趨性影像分析………..…… 55
圖21 : 光趨性分析裝置架設……….……….………… 56
圖22 : 照射光源的波譜……….……….……… 57
圖23 : 偵測光源範圍……….………..……..….………… 58
圖24 : 光源限制片……….…………..………...…… 58
圖25 : C. reinhardtii 於光趨性分析裝置之分析……….……….…..…...….… 58
圖26 : C. reinhardtii 在白光下的巨觀避光現象……….………....…..… 59
圖27 : H. salinarum 轉形株的菌落 PCR………..….…… 61
圖28 : H. salinarum 轉形株之 Western Blot……….……….…..…...…..… 61
圖29 : H. salinarum 野生株及轉形株細胞膜的可見光吸收波譜…………...…… 62
圖30 : SRM 及 SRM-HtrM 於不同條件之光週期分析………...……… 63
圖31 : H. salinarum 野生株及轉形株型態……….…..……… 64
圖32 : 嗜鹽古生菌於不同光照下之生長曲線………….……….….…...…… 66
圖33 : 嗜鹽古生菌在顯微鏡之光趨性分析……….……….……....… 68
圖34 : 嗜鹽古生菌在 ELISA Reader 之光趨性測定………….……….…….…… 69
圖35 : E. coli BL21 表現 HmCheR ……….………...…………..…..… 75
圖36 : E. coli BL21 小量表現 GST 嵌合蛋白質……….…...………….…..… 75
附圖目錄
附圖1 : 光照培養箱之設計……….….…..….….… 83
附圖2 : H. salinarum 的趨氧現象……….………..………83
附圖3 : 嗜鹽古生菌於不同光照下之菌體狀況……….………….………… 84
表目錄
表1 : 各類視紫質在四種嗜鹽古生菌中的分佈及吸收峰…………....…..…… 8表2 : 各蛋白質長度及帶電胺基酸的占比……….…...……...… 44
表3 : HmSRs 在不同 pH 值下的光週期速率………..……… 50
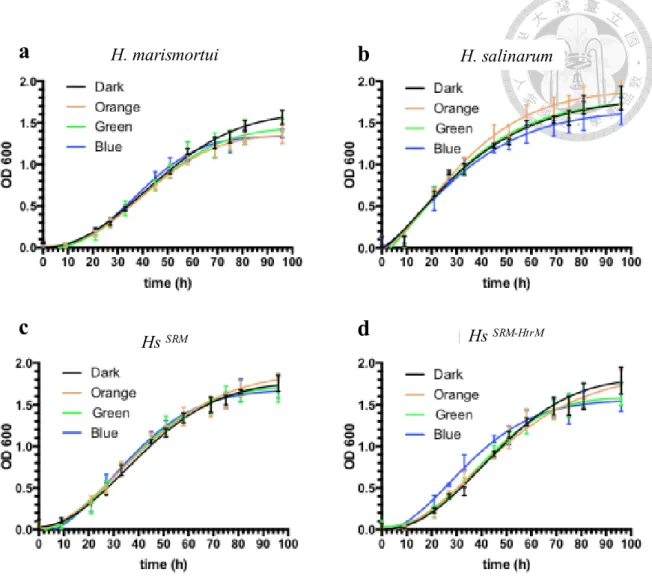
表4 : 嗜鹽古生菌於不同光照環境下之世代時間………….……...………… 65
表5 : 單隻嗜鹽古生菌對 RGB 光源刺激之反應……….……...………… 67
摘要
古生菌的感覺型視紫質 (Sensory Rhodopsin, SR) 可藉由感受不同可見光波 長之光源,並藉由其對應之傳導元 (Transducer, Htr) 傳遞訊息,以調控嗜鹽古生 菌之趨、避光反應。目前發現之 SRs 共有三種型態 : SRI, SRII 及 SRM;其感受 之 可 見 光 波 長 分 別 分 佈 於 紅 藍 綠 光 波 段 。 基 因 體 的 序 列 中 , 嗜 鹽 古 生 菌 Natronomonas pharaonis 僅具有調控吸收 ~498 nm 而驅動避光的 NpSRII、
Halobacterium salinarum 擁有 SRII 及吸收~590 nm 來調控趨光的 SRI 。本實驗 室先前發表基因體的 Haloarcula marismortui,為一三色感光系統的嗜鹽古生菌,
除了 SRI, SRII 以外,尚有一個功能未定的 SRM。SRI 及 SRII 之傳導元藉由類 同細菌中化學趨性的 two-component system ,將光訊號轉化為化學訊息傳遞至 鞭毛,進而調控菌體之泳動 ; 然而, SRM 的傳導元 HtrM 結構上缺乏了許多 構件,因此我們推測 SRM-HtrM 應由其他機制調控嗜鹽古生菌之光趨性。本篇 研究,先建立 SRM 及 SRM-HtrM 在二色感光系統 H. salinarum 菌株中的表現 及確立其功能,再以兩種本篇研究發展之光趨性研究方法 (ELISA Reader 之量 測及顯微鏡之觀察),量化 H. salinarum 及其轉形株的光趨反應。初步結果顯示,
SRM-HtrM 減少了 H. salinarum 在綠光及藍光下的避光反應; 並且,在 HtrM 缺 乏的轉形株中,該現象並沒有被觀察到。因此,我們認為 HtrM 的存在,對於 SRM-HtrM 複合蛋白質在 H. salinarum 中調控的趨光反應是重要的。未來,可以 嘗試將 SRM-HtrM 複合蛋白質嵌入磷脂中,分析其與化學趨性相關蛋白質之間 的交互作用,以解出其分子機制。
關鍵字: 光趨性、感覺型視紫質、Halobacteriuma salinarum、SRM、HtrM
Abstract
A group of photoreceptors, sensory rhodopsin (SR), regulates phototaxis in haloarchaea through absorbing diverse range of visible light and relaying the signals to the cell by their cognate transducer (Htr). To date, there are three types of SRs identified:
SRI, SRII and SRM, and they response to red, blue and green light, respectively.
Among the annotated archaeal genomes, Natromonas pharaonis solely own the NpSRII, which response blue light and mediates photorepellent; Halobacterium salinarum holds HsSRII and HsSRI, and HsSRI is known to absorb ~590 nm of light to mediate photoattractant. However, in Haloarcula marismortui, there exists three SRs, namely SRI, SRII and functionally unknown SRM. Previous studies showed that SRI and SRII transduce photo signal to flagellum through transducer and chemotaxis proteins, similar to two-component system of chemotaxis in bacteria. However, it is speculated that the SRM-HtrM complex regulates the phototaxis responses through new pathway as SRM- HtrM lacks many structural components seen in other transducers. In this study, we transplanted SRM and SRM-HtrM into H. salinarum cells and compare the phototaxis responses of H. salinarum and its transformants under different wavelengths of light through two new measurements developed in this study. It is found that SRM-HtrM decreased the photorepellent response of H. salinarum in green and blue light; but not in SRM transformant. The importance of HtrM in SRM-HtrM signaling was concluded.
In future, the molecular mechanism of SRM-HtrM can be examined through measurements of the interaction between lipid-reconstituted SRM-HtrM with related chemotaxis proteins.
Keywords: Phototaxis, Sensory Rhodopsin, Halobacterium salinarum, SRM, HtrM
第一章 緒論
第一節 嗜鹽古生菌之介紹
自然界的生命系統,藉由 16S rRNA 的演化分析,被分為細菌、古生菌及真 核生物;甚至可以藉此,區分出他們的種或屬 [1, 2]。其中,古生菌被相信是一群 生活在地球早期環境的微生物,能夠適應早期的嚴苛環境,如高溫、高壓、高鹽、
酸性及缺氧等等 [3]。其中,嗜鹽古生菌能夠忍受高光照、高鹽 (1.5 ~ 2.5 M NaCl 以上) 且乾旱的環境 [4, 5],包 括在 1919 年最早被發現並且被研究較多的 Halobacterium salinarum [6]、於死海發現的 Haloarcula marismortui [7, 8]、於埃及 鹽湖分離出的 Natronomonas pharaonis [9]及具有特殊扁方形狀且能忍受高濃度 鎂離子的Haloquadratum walsbyi [10, 11]。嗜鹽古生菌廣泛分佈在世界各地 [4],並 且隨著分子生物學的進步,嗜鹽古生菌中的模式生物 Halobacterium salinarum (Halobacterium sp. NRC-1) 及 Haloarcula marismortui 的全基因體已分別於西元 2000 及 2004 年被發表 [3, 12],目前已經有20 餘種嗜鹽古生菌的全基因體序列被 記錄於資料庫中。
第二節 微生物視紫質 (Microbial Rhodopsins)
微生物視紫質 (Microbial Rhodopsin, MRho),是普遍存在於微生物中,能夠 感受可見光,並對微生物產生生理作用的一群穿膜蛋白質 [13, 14]。這群感光蛋白 質 (圖 1) 具有以下共同特性 : (1) 由七個 alpha 螺旋穿膜區域構成的筒狀蛋白 質,由N 端至 C 端分別被標示為 A-G helix ; (2) 由視黃醛 (retinal) 與 G-helix 上 的離胺酸 (Lysine) 形成共價鍵結的 Schiff base 構成一個感光基團 ; (3) 蛋白質 在基態時,發光基團為全反式視黃醛 (all-trans retinal),並且發光基團受光子激 發後,會變為13-順視黃醛 (13-cis retinal),進而改變蛋白質結構,使其變為數個 不同中間激發態 (intermediates) ; (4) 視紫質能夠依其 retinal binding pocket (RBP) 朝向細胞外或胞內,而分別定義為E 及 C conformer [15]。
圖1、微生物視紫質之共通特性。 a. 由 A 至 G 共七個 alpha 螺旋構成的穿膜區 域,虛線圈出感光基團 all-trans-retinal 與蛋白質 G 螺旋上的離胺酸形成 Schiff base 鍵結。 b. 以菌式視紫質 HsBR 為模板,展示視紫質的蛋白質結構,藍色為 N 端,紅色為 C 端。 c. 發光基團 retinal 在光照前後,於 all-trans 及 13-cis 型 態間之轉換。
統整目前在所有微生物中 MRho 的功能研究結果,MRho 主要可區分出光 驅動離子幫浦 (light-driven ion transporters) 及光趨性受體 (phototaxis receptors) 兩 大 類 [14, 16, 17]。 第 一 類 中 , 包 含 了 在 嗜 鹽 古 生 菌 中 的 菌 式 視 紫 質 (Bacteriorhodopsin, BR) 、氯視紫質 (Halorhodopsin, HR) [18],以及在海洋細菌中 的鈉視紫質(Sodium-pumping rhodopsin) [19]。而在第二類光趨性受體中,則包含 了在嗜鹽古生菌的感覺型視紫質 (Sensory Rhodopsin, SR) 、藍綠菌中的感覺型 視紫質 (Anabaena Sensory Rhodopsin) [20, 21]、藻類中的陽/陰離子通道 (Cation / Anion ChannelRhodopsin, ChR) [22, 23]調控微細綠藻的鞭毛等等。
在真菌,除了原先被發現的 Nop-1 的真菌視紫質 [24]外,近年藉由生物資訊 學的方式,也的確在許多真菌中發現視紫質的存在,並有其豐富的多樣性,如光 週期快的、慢的以及離子傳輸與否的都有被報導過 [25, 26],有些研究指出真菌視 紫質能夠調控真菌的光趨性 [27]。
若只針對嗜鹽古生菌來看,截至目前的研究,這類 MRho 主要功能有三種 (圖 2) :菌式視紫質、氯視紫質、以及感覺型視紫質 [28]。 值得一提的是,在 目前發現的微生物視紫質中,有兩個功能尚未確定的蛋白質,分別為: HwMR
[29] 及 HmSRM [30, 31]。
其中,HwMR 存在於缺乏感覺型視紫質的 H. walsbyi 中,該菌株藉由未知 機制調控液胞 (gas vesicles) ,而能在水域中改變可見光的獲取量 [11]。HwMR 在 序列比對上被歸屬於 BR,但其同時具有光週期慢的特性,這又比較類似感覺型
marismortui 中 [31, 32], 由 序 列 分 析 及 基 因 體 比 對 發 現 和 它 配 對 的 傳 導 元 (transducer) HtrM [31],推測其為感覺型視紫質,為本篇研究的重點。
圖2、嗜鹽古生菌中視紫質的分佈。將七株嗜鹽古生菌基因體中發現的微生物視
紫質,按照蛋白質序列比對親緣關係,並以不同色塊標出五大類的微生物視紫質:
BR (青色), HR (綠色), SRI (紅色), SRII (黃色), SRM (橘色)。標記方式為學名加上 蛋白質名稱。圖中數字標出 Bootstrap value (>0.5),數值越大表示統計資料的確 定性越高。BR: bacteriorhodopsin; HR: halorhodopsin; SR: sensory rhodopsin。
菌式視紫質 (Bacteriorhodopsin; BR) 以三聯體為單位,聚集在古生菌細胞膜 上,形成一塊連續性的特定區位,稱之為紫膜 (purple membrane) [35]。BR 是一 個光驅動質子幫浦,能夠吸收550 nm 波長附近的綠光波長,將質子從細胞內傳 輸至細胞外 [33, 34]。藉由此質子傳輸形成胞內外間的質子濃度梯度,使質子由胞
(Lynch EA, 2012)
外通過 ATPase 進入胞內時,以 10 : 3 的質子/ATP 分子比例,產生生物之能量 分子 ATP [35]。HsBR 是最早從 H. salinarum 被選殖出來表現並定性的 BR,
Pebay-Peyroula 更在 1997 年提出了解析度達 2.35 Å 的 X-ray 結構 [36]。其質子傳 遞路徑及過程,也已經藉由點突變、X-ray 結晶繞射及 NMR 等技術,完整的呈 現 [37]。
氯視紫質 (Halorhodopsin; HR) 為一光驅動氯離子幫浦,普遍最高吸收峰在 於570 nm 前後,能夠將氯離子從細胞外傳輸至胞內 [38]。於本實驗室2016 年出 版的研究中,發現其具有質子依賴 (NpHR, HwHR) 及非依賴 (HsHR, HmHR) 兩 種氯離子運送型態 [39]。生理上,因為他廣泛存在於目前發現的嗜鹽古生菌中,
並且和其他存活於高鹽環境的生物,以利用消耗ATP 驅動 Na/Cl transporter 維 持滲透壓 [40]不同,是利用光能運送氯離子,被認為與維持細胞內滲透壓有關。
嗜鹽古生菌的感覺型視紫質,在視紫質本身外,還有一個具兩個穿膜區域 (alpha helix) 的傳導元 (transducer) 以氫鍵及電荷配對的方式依附在其 F & G 螺 旋旁邊 (圖 3a) [41]。依其功能及吸收波段分為二類:吸收紅光波段並且調節藉趨 光反應的 Sensory Rhodopsin I (SRI) [42] ; 及吸收藍光波段並調節避光反應的 Sensory Rhodopsin II (SRII) [43]。本實驗室發表的Sensory Rhodopsin M (SRM) 則 會吸收綠光波段,推測能夠增加趨光反應。感覺型視紫質 (NpSRII) 訊息傳遞機 制,是由F, G alpha 螺旋上兩個胺基酸:tyrosine, threonine,往外推送,將此感受 訊號藉由氫鍵,物理性地傳導至旁邊的傳導元 (transducer) [44]。生理上,SRs 會 以三聯雙聯體 (trimer of dimer) 的形式存在於細胞膜上,而此形式能夠增加其酵 素活性達百倍 (圖 3b) [45, 46]。
圖3、嗜鹽古生菌中感覺型視紫質與其傳導元。 a 以 SRM 及 HtrM 的簡圖示意,
感覺型視紫質與其傳導元之位置關係。 b 以 NpSRII 的 X-ray 結構表示,生理上 感覺型視紫質以三聯雙聯體存在。SR 單元體之間距離約 9 nm 、 傳導元之長度 約為 35 nm 、 傳導元之間的距離則為 7-8 nm。不同之單元體以不同的顏色標 示。複合體胞內端以六個傳導元尖端,共同構成與 CheA/CheW 複合體結合之區 域。
第三節 古生菌感覺型視紫質
MRho 彼此之間具有相似的主要結構,然其間亦有許多的異同之處,以下分 成物理特性及演化觀點兩部分介紹古生菌的感覺型視紫質:第一部分 (3.1 – 3.3) 包含了,它們的吸收光譜、光週期及離子傳輸之現象; 第二部分 (3.4, 3.5) 則以 序列及蛋白質功能來探討它們在演化上的關係。
I-3.1 紫外光/可見光譜
如前節所提,所有的微生物視紫質,MRho,都是七穿膜的蛋白質,也皆使 用視黃醛作為它們的感光基團,但相同的視黃醛分子被被包在不同七穿膜蛋白質 時,其分子環境是由不同的胺基酸支鏈所組成,因此,視黃醛被激發的波長,也 就是其感受最敏銳的可見光波長 (最高吸收峰) 都不一樣。不同的最高吸收峰,
可能意味著不同的生理功能、不同光週期快慢、和相同蛋白質,但在不同的構形 狀態。因此,光譜分析可以幫助我們掌握蛋白質之特性及狀態。以感覺型視紫質 為例,依其感受的可見光波段以及其調控的生理功能,可分為 SRI (趨光 / S373
參與避光反應) 、SRM (推測為協助趨光反應) 及 SRII (避光) 三類,其於不同嗜 鹽古生菌中的特徵吸收峰整理於 (表 1)。此外,研究也發現在不同 pH 值、氯化 鈉濃度等條件下,也會造成特徵吸收波長的改變 [43, 47, 48]; 甚或透過胺基酸點突 變也可能造成吸收峰的藍移或紅移,可以進而推測出某些胺基酸對於 Retinal Binding Pocket 微環境的影響 [49-51]。
I-3.2 光週期
光週期的定義為,當視紫質的發光基團 (retinal) 受到一個光子激發後,會從 原本的all-trans retinal 異構化為 13-cis retinal。這個異構化 (isomerization) 的過 程,會帶動視紫質整體結構的改變,如前所述,發光團也隨著一系列蛋白質構形 變化,改變其最高吸收波長。於是從基態引發一連串型態的變化,最後再回到基 態,可以藉由測量特定狀態,最高吸收波長隨時間的變化而知,稱之為光週期 [52]。 菌式視紫質的光週期通常都在數十毫秒 (millisecond) 內完成,其光週期因為最 早被量測,因此由其定義出 K, L, M, N, O state,後續在視紫質光週期的研究中,
也都沿用這些名詞表述 retinal 在光週期中的狀態改變 [52]。氯視紫質的光週期 則稍微快一點,通常在數毫秒內完成,過程中Schiff base 維持在質子化的狀態,
缺乏 M, N state [53]。感覺型視紫質的光週期相對長很多,通常在一秒至數秒,
甚或更久 [54, 55],且具有以下特性 : 當 SRIs 與可以專一性作用的傳導元結合 後,光週期會變得對於pH 質較不敏感 (pH 4-8) ; 當 SRIIs 與專一性作用的傳導 元結合後,光週期則會變得較快 [56],這個特性也成為研究SRs 是否與 transducers 作用的依據之一 [57, 58]。
表1、各類視紫質在四種嗜鹽古生菌中的分佈及吸收峰。
表中的數字代表各個MRho,在 micelle (DDM) 環境下的最高吸收峰 (nm)。
Haloarchaea 𝜆𝑚𝑎𝑥
BR HR SRI SRM SRII
H. marismortui 552 576 578 503 483
H. salinarum 550 578 590 X 490
N. pharaonis X 589 X X 500
Haloferax sp. X X X X X
I-3.3 質子傳輸
雖然感覺型視紫質主要功能為調控古生菌光趨性,但先前的研究也發現,感 覺型視紫質在沒有傳導元的依附下,同樣會有類似 BRs 質子傳輸的現象。
HsSRI 及 HsSRII 已於前人的努力下,對於其質子傳輸現象有較完整的陳述 [59,
60]; HmSRM 則於近期研究中指出,未與傳導元結合的情況下, HmSRM 具有 微弱質子幫浦能力 [30]; 但其與傳導元 HtrM 作用後的質子傳輸及光週期並未 被研究。
目前的研究顯示,HsSRI 的質子傳輸機制較為複雜,但研究也認為質子傳輸 或循環對於 SRs 的 M state (實際將光趨性訊號傳遞至傳導元的 state) 之形成都 是重要的 [59]。在沒有傳導元的作用且環境 pH 值高於中性的條件下,因為胞外 側的Asp76 會被去質子化而帶負電 (對應至 BR 的 Asp85) ,成為 Schiff base 的 拮抗離子 (counterion) [61],因此導致 HsSRI 的吸收峰藍移至 552 nm。此時 HsSRI 能夠像質子幫浦般,將質子幫浦至細胞外,且不會伴隨調節避光反應的 M’ state 形成 (圖 4a);反之,若 pH 值處於酸性,則 HtrI-free HsSRI 的吸收峰會紅移至 587 nm,若只有 587 nm 的光子激發,並不會有從胞外拿取質子的 M’ state 形成,
所以並無淨質子傳輸; 但若同時給予 373 nm 波段的光子,則 HsSRI 能藉由 two- photon pathway 進入 M’s state 而調控避光反應,質子向胞內淨傳輸 [62]。然而當 HsSRI 與 HsHtrI 作用後,因為 Asp76 的 pKa 增加至 ~8.5 [62],質子僅能藉由胞 內側的 His166 循環,並不會改變淨質子傳輸量 (圖 4b) [63]。利用這個質子傳輸 抑制的現象,我們也可以判斷SRIs 是否與傳導元作用。
圖4、H. salinarum 感覺型視紫質的質子傳輸現象。a SRI 在沒有傳導元作用下,
吸收峰會藍移35 nm,並藉由 His166 > Schiff base > D76 之路徑,將質子幫浦至 胞外 b 當 SRI 與傳導元作用後,能夠進行 two photon pathway,進入可調控避 光反應的 M’ state。若僅有紅光刺激,則質子只會在胞內循環; 若進入 M’ state 則,整體質子往胞內淨傳輸 c SRII 與傳導元做用後,質子僅藉由 Asp73 在胞外 循環。方框斜線開口標示了該狀態下, Schiff base 朝向的方向:胞外端 (E conformer) 或胞內端 (C conformer)。
HsSRII 於光週期的特徵跟 BRs 很像,並且也具有 Asp73 做為 Schiff base 的拮抗離子,承接了 M state 形成時丟出的質子 [64]。然而,在實驗結果中並未 顯示出 (或說微弱至可忽略) 其具有質子幫浦的能力; 在更進一步的研究指出,
HtrI-deficient a
b
c
(Sasaki J, 2000)
Asp73 負責了在不同的 state 下,質子在胞外側的循環 (圖 4c) [60]。因為無實際 資料指出 SRIIs 具有質子傳輸功能,所以也無法以此為依據,判斷 HtrII 是否與 其作用。另外, HtrII 的結合,也不會提高 Asp73 及 Schiff base 的 pKa; 在吸 收峰的改變上,更沒有像SRIs 與傳導元作用後,紅移達 35 nm [56]。值得一提的 是,傳導元的作用會影響 SRs 進入 M state 時,F (G)-helix 向外的推出 (C conformer)。並且, HsHtrII 會加快 HsSRII – M, O state 的進行 (加速 Asp73 去 質子化) [60]。F, G 螺旋的向外推出,導致 E, C conformer 轉換的動作,也被視為 SRs 感受光子並傳遞訊號的重要指標之一。在一些研究中發現,只要能觸發 SRs 處於C conformer (對應至 SR 的 M state) ,就能將光趨性訊息傳遞下去 [65]。
I-3.4 重要胺基酸
於 NpSRII 的研究中,藉由 X-ray 晶體繞射,可以得知 SRII 藉由哪些胺基 酸與傳導元作用 [66, 67]。在 SRII 的 Tyr51 (B-helix), Tyr174 (F-helix), Aspp201 &
Thr204 (G-helix) 構成了感光基團的結合區域(RBP) [66, 68],利用氫鍵的網絡,將 視黃醛產生形變的物理作用力傳導至F 及 G-helix (圖 5c) [69]。Tyr51 對應至 SRM 為 Met54,但在 BRs 中同樣是 Tyrosine; Thr204 對應至 HsBR 及 NpSRII 都為 Alanine,並且 HsBR_A215T 具有與 HtrII 作用的能力 (圖 5a) [66]。傳導元藉由 兩個位置與 SRII 結合:Thr189 (F-G loop) / Glu43 (loop), Ser62 (TM2) 及Tyr199 (G-helix) / Asn74 (TM2) [NpSRII / HtrII] (圖 5b) [15, 67]。 Thr189 對應至 SRM 為 Glu189,在 HsBR 中則是 Proline; Tyr199 對應至 HsBR 及 NpSRII (199) 都 為Valine (圖 5a)。
圖5、感覺型視紫質重要胺基酸比對。a HmSRM, NpSRII & HsBR 蛋白質序列的比對。紫色方框標示出retinal binding pocket的重 要胺基酸; 綠色方框標示出,NpSRII朝外與傳導元作用的胺基酸; 旁邊標示的胺基酸編碼,為對應至NpSRII上的胺基酸位置。 b 結構上,NpSRII與傳導元胺基酸之間的氫鍵作用 (黃色虛線)。橘色箭頭表示,蛋白質形變後,Y199的移動方向。 c HsBR(紅), HsBR_A215T (黃) & NpSRII (藍) 的RBP在結構上的比較。
I-3.5 演化探討
嗜鹽古生菌的視紫質目前並沒有一個完整的演化理論,但從蛋白質序列比對、
點突變及功能特性上,可以找出在演化上可能的相關性 [70]。 HsBR_D85T (D85 為 BR Schiff base 的拮抗離子,對應至 HR 的位置),具有與 HR 相似的氯離子 幫浦功能 [71, 72]; 然目前仍無法以類似方法,將 HR 轉變為質子幫浦 [73, 74]。SRI 具有與BR 類似的質子幫浦功能,但 SRII 因為缺乏了胞內端的質子受體,質子 僅能在胞外端循環。在 NpSRII 的研究中,有個爭議的問題,在一些研究中顯示,
氯離子會跟 NpSRII 結合,並且會有吸收峰及光週期上的差異 [75]。BR 也可以 利用點突變的方式,將對應至 NpSRII 的 Thr189, Tyr199, Thr204 的胺基酸突變,
成功地利用 BR 變異株傳導光趨性的訊號 [66, 76]。本篇研究的 HmSRM 在蛋白 質序列的比對上 (圖 5a),相較於 SRI,其相似度與 SRII 較近; SRII 在序列比 對上又與 BR 相似度較高,因此目前認為 SRII 可能是由 BR 演化而來。
第四節 傳導元的結構與特性
傳導元 (Transducer) 是 SRs 將光訊號轉換成生理化學訊號的一個重要輔助 蛋白質,它專一性地依附在SRs 旁邊 [58],並以2 : 2 的分子比例形成雙聯複合體
[51]。依其野生型態結合的SRs 命名,如 HsSRI 的傳導元即為 HsHtrI , HmSRM 的 傳 導 元 即 為 HmHtrM 。 傳 導 元 的 結 構 上 ( 圖 6a) 具 有 兩 個 穿 膜 (transmembrane; TM) 的 alpha 螺旋: TM1, TM2、在胞內的 HAMP (Histidine kinases, Adenylyl cyclases, Methyl binding proteins, Phosphatases) domain 及 MCP (Methyl-accepting chemotaxis protein) domain。 HtrI 的胞外環 (loop between TM1, 2) 普遍都很短, HtrII 的胞外環通常都長很多; 然而, NpHtrII 及 HmHtrM 的 胞外環卻跟 HtrI 比較類似,並沒有很長。本篇研究的動機之一為,缺乏 MCP domain 及與 Che proteins 作用之尖端的 HmHtrM 如何調控光趨性(圖 6b)。
圖 6、傳導元之結構及比
較。 a 傳導元的基本結 構 。包含 兩個穿膜 螺旋 (藍)、胞外環 (上方灰)、
HAMP domain ( 綠 ) 及 MA domain (紅),下方的 尖 端 則 與 CheA/CheW 作用。然而,SRM 的傳導 元 HtrM 缺乏了虛線的 幾個區域。 b 不同種的 SRs 在吸收波譜上的分佈。
左邊橘黃色的為SRIIs,中間紅色的為 SRM ,右邊紫色的則為 SRIs 。可看出 傳導元的胞外環有長短之分,並且HtrM 沒有很長且向下延長的尖端 (紅色虛線)。
b
T M 2
a
HAMP
(Fu HY, 2010)
在先前的研究中,傳導元與 SRs 的專一性由兩個穿膜區域及部分的胞內端 即可決定 [77]。在 HsSRI 的研究上,只需要 HsHtrI 由 N 端算起的 147 個胺基 酸,即可專一性地結合並穩定 HsSRI [58]。有趣的是,當把 NpSRII 接上一個複 合傳導元 (NpHtrII + E. coli Tsr) ,藉由 NpHtrII 的兩個穿膜區域及一個 HAMP domain,該傳導元可以傳遞 NpSRII 感受的藍光訊號,並且藉由大腸桿菌的化學 趨性受體 Tsr 的胞內端,改變化學趨性系統 (chemotaxis signaling pathway) 的活 性 [78]。在 NpSRII 與 HsBR 的結構比較上, SR 胞內端的表面電荷相較於 BR,
帶有許多正電荷,推測與傳導元胞內端負電荷胺基酸的電荷作用有關,因此 BR 無法與傳導元結合 [79]。
光子趨動 SRs 的 F-helix 推出後,造成了傳導元的 TM2 旋轉了 12-30 度
[80],這個旋轉的動作導致了下游各個元件,包含 TM2、HAMP1、HAMP2、甲基 化區域及尖端,能動性 (dynamic) 的交替轉換。於 NpSRII / HtrII trimer of dimer 的研究中,發現在活化訊息的傳遞狀態下, TM2、HAMP2 及尖端的活動性較 高 [81]。因此,以雙聯體形式結合在尖端的 CheA/CheW 能夠被活化,而進行生 化訊息的傳遞。另外,甲基化區域可逆的甲基化 (methylation) 反應也影響了該 區域的能動性,在 CheR 的幫助下,該區域會被甲基化,並且因為電荷減少的緣 故,彼此靠近造成該區域的能動性下降 [81]。此舉會增加尖端的能動性 (因為能 動性的高低,是各元件間交替起伏的) ,而活化與尖端結合的 CheA kinase ,將 訊息以生化路徑傳遞至鞭毛 (圖 8) [81]。
第五節 嗜鹽古生菌的光趨性
I-5.1 SRs 於嗜鹽古生菌中的分佈
感覺型視紫質顯然並非嗜鹽古生菌必要的蛋白質,並且在不同的嗜鹽古生菌 中,分布的數量也不同 (表 1,圖 6b); 然而,它可以將菌體帶至適合 BRs 作用 並且輻射傷害較少的光照環境。藉由這些具有不同數量 SRs 的嗜鹽古生菌 (包 含了在基因體鑑定上沒有發現視紫質的菌株Hfx),我們可以更廣泛地去探討,
HmSRM-HtrM 在調控光趨性的機制中,扮演的角色。
I-5.2 嗜鹽古生菌的光趨性
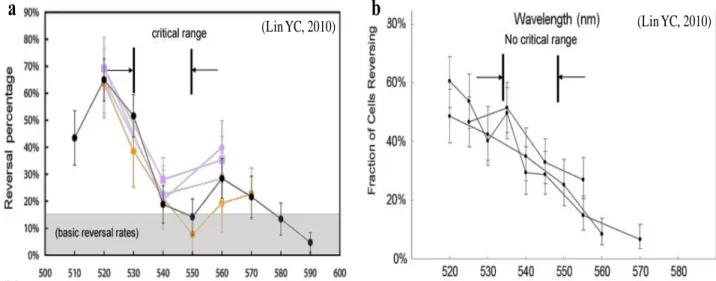
微生物感光的目的,通常為避開有害藍光或找到利用光源的最佳生態地理位 置。對於嗜鹽古生菌來說,生理上,他們必須在面對有害的藍光及利於 BRs 作 用,進而產生ATPs 的黃綠光,之間取得一個平衡。如前所述,目前主要以蛋白 質序列、感受的光源波段及其調控的生理功能,將感覺型視紫質分為三類:SRIs 可以感受橘至紅光波段光源,其M state 傳遞了趨光反應的訊息 [42]。 然而,如 果在 SRI 進入 M state (S373) 時,再給予另一個藍光光子,可以使其進入 M ’ state 則會傳遞避光反應的訊號 [82]。 SRMs 可以感受綠光 (𝜆𝑚𝑎𝑥 = 503 nm) , 在生理上的泳動分析,推測它的作用是增加趨光反應的效應 (圖 7; unpublished data from Lin Y.C.)。SRIIs 感受藍光光源,並且傳遞避光的訊號 [43],目前主要的 研究都在於,僅具有 SRII 的嗜鹽古生菌 Natromonas pharaonis 的 NpSRII 上。
目前僅有 Haloarcula 屬被發現,同時擁有這三種 SRs; 而 SRs 在嗜鹽古生菌中 也不是並非每個都是必要的 (表 1)。
先前於感覺型視紫質如何調控光趨性的機制,主要在於 NpSRII 的機制研究。
目前認為 SRI 及 SRII 共用了 Che (chemotaxis) proteins, (細菌調控化學趨性 的two-component system) 調控了嗜鹽古生菌的光趨性。SRs 受光激發後,藉由 推出的 F helix ,使傳導元的 TM2 位移。傳導元的整體結構有如蹺蹺板,有許 多元件間的節點,在活化及非活化的狀態下,各區域會在動態及靜態之間轉換
[81]。以此機制去調節CheA (藉由 CheW 的輔助而結合在傳導元尾端) 的活性,進 而磷酸化CheB 及 CheY 而使其活化, CheB-P 能將傳導元的 MCPs 去甲基化,
CheZ 則負責將 CheY-P 去磷酸化。 CheY-P 能夠調控菌體的鞭毛,使其產生 tumble 的反應而轉向,在整個系統重置後,菌體改為前進模式,最後產生避光的 折返現象 [81]。
圖 7、菌體泳動分析。a Haloarcula marismortui b Halobacterium salinarum,在 不同波長的光源脈衝(0.5 sec) 刺激後,菌體折返的機率 (偏移原路徑 > 90˚)。折 返率越高,代表越高的避光反應。 X 軸為使用光源之波長,單位: nm。
a b
(Lin YC, 2010) (Lin YC, 2010)
I-5.3 SRs 調控光趨性之機制
避光反應 (菌體折返) 的過程牽涉了許多的蛋白質,按順序為:Sensory rhodopsin (SR) 、 Transducer (Htr) 、 CheA/CheW 、 CheR 、 CheB 、 CheY 、 CheZ 及鞭毛。NpSRII/HtrII complex 本身就是一個感受光刺激的 two component system 。 CheR 是 一 個 持 續 活 化 的 酵 素 , 持 續 地 將 傳 導 元 甲 基 化 , 維 持 NpSRII/HtrII complex 在高光敏度,也是在一個週期的光刺激後,將感光系統重 置 (reset) 的酵素 [45]。訊息傳導的完整流程如下 : (1) NpSRII 受藍光激發,
retinal 構型由 all-trans 變為 13-cis,帶動 F(G)-helix 向外推出。 (2) 推出的 F- helix,造成傳導元 TM2 旋轉,改變該區域的能動性。 (3) 藉由交替的能動性改 變,傳導元尖端活化與其結合的 CheA/CheW 雙聯複合體。 (CheA 為 kinase;
CheW 為幫助 CheA 結合在傳導元尖端的蛋白質) (4) 被活化的 CheA,會將 CheB 及 CheY 磷酸化而使其活化。 (5) CheB-P 會將傳導元的甲基化區域去甲 基化,傳導元失去活性 (負回饋抑制) (6) Che-Y 則跑去活化鞭毛,使其產 生 ”tumble” (順時針轉動) 的生理反應,藉此將菌體轉向。 (7) CheZ 會將 CheY-P 去磷酸化,將其重置。 (8) 當整個訊息傳遞完 ”tumble” 的訊息並且重置 後,細胞轉為 ”Run” (逆時針轉動) 的模式,產生了一個折返的生理反應 (圖 8)
[81]。
圖 8、NpSRII 傳遞避光反應的訊息路徑。NpSRII 及 CheA/CheW 都是以雙聯 複合體的形式存在,CheR 是一直具有活性的甲基化酵素,CheR 及 CheY 則是被 CheA kinase 磷酸化後,才活化的去甲基化酵素及鞭毛調控因子。另外,有個未 標示出的 CheZ 酵素,會將磷酸化的 CheY 去磷酸化,以重置訊息的傳遞。
第六節 研究動機及策略
本實驗室的傅煦媛博士於2010 年,發表了一篇關於在 Haloarcula marismortui 這株於死海發現的嗜鹽古生菌中,定義出截至目前為止,具有最多視紫質的古生 菌 [31]。其中,有個視紫質因為藉由胺基酸序列比對,發現既沒有類似光驅動離 子傳輸受體的序列,也沒有能夠與傳導元交互作用的保守胺基酸,因此命名為未 知功能的opsin: xop2 (HmSRM)。於光週期的研究中,發現 SRM 的光週期很慢,
與 SRs 的特性類似。並且在後續的基因體研究中,發現該基因下游有一個 184 個胺基酸的 HAMP domain 蛋白質,是可以與 HmSRM 作用的傳導元 HmHtrM。
因此,目前認為 HmSRM 是一個感覺型視紫質,並且與僅具有兩個穿膜區域及 一個 HAMP domain 的傳導元 HmHtrM 作用。由於這個蛋白質在生物資訊學上 的分析,不同於以往定義出的視紫質 (BR, HR, SRI, SRII),因此我們對於這個蛋 白質在生理上,是否與先前研究的看法相同,有極大的興趣。
本篇研究著重於 HmSRM 的兩個層面: (1) 生理上,它在趨光反應中產生的效 應 (2) 機制上,HtrM 是否為光趨性調控的重要元件。
本實驗室在先前於SRM 的研究中,發現 H. marismortui 相較於只有 SRI 及 SRII
的H. salinarum,在面對接近綠光波段的光源時,會有一個明顯快速上升的趨光
反應 (圖 7)。並且由於 HtrM 的胞內端缺少許多結構,因此推測 SRM-HtrM 複合 體,可能會與CheR 或 CheB 結合,“間接”地調控光趨性系統的活性。本研究藉 由轉基因菌株 (HsSRM, HsSRM-HtrM) 及重新設計之光趨性分析系統,分析具有不同 數量 SRs 的菌株,觀察 SRM 對於嗜鹽古生菌光趨性的影響。
圖9、實驗流程表。本研究分為四大部分: HmSRM 蛋白質的性質、 HmSRM 在 H. salinarum 轉形株的建立、光趨性研究方法的建立及 H. salinarum 轉形株 之分理分析。
第二章 材料與方法
第一節 生物試劑
II-1.1 菌種
E. coli strain DH5𝛼,用以放大複製及保存質體。
E. coli strain BL21 (DE3),用以表現重組可溶蛋白質。
E. coli strain C43 (DE3),基因突變所致細胞膜增生之細胞株。用以表現 重組視紫質(穿膜蛋白質)。由中央研究院 王惠鈞博士實驗室提供。
Haloarcula marismortui ATCC 43049,於 2010 年由本實驗室 傅煦媛博士發表具 有六種視紫質之嗜鹽古生菌。本研究中之感覺型視紫質 HmSRM 及 HmSRM-HtrM 即為其視紫質之一。由國立陽明大學 生物技術研究所 吳韋訥 博士實驗室提供。
Halobacterium salinarum,具有 SRI 及 SRII 兩種感覺型視紫質的嗜鹽古生菌。
Chlamydomonas reinhardtii (137C),野生株的單細胞綠藻,具有兩根鞭毛及眼點,
調控光趨性反應。由國立臺灣大學 生物科技研究所 陳仁治博士提供。
II-1.2 質體
pET-21b: 用於表達重組蛋白質 (包含 xop2, CheB, CheR)
pET-Duet: 用於建構及表現兩目標重組蛋白質之結合蛋白質。(包含 xop2-xHtrII);
本研究中使用之基因為將 6His-tag 放在傳導元 xHtrII 兩個穿膜螺旋之間的胞外 環。
pGEX 4T-1: 用於表現 GST 蛋白質,並且建構 GST-CheR 及 GST-CheB 嵌合蛋白
質的載體。
以上載體皆由T7 promoter 所調控,藉由 IPTG 誘導,於 Escherichia coli 異源表 現系統中表現重組蛋白質。
pJS005: 使用菌式視紫質的啟動子 bop promoter 作為大量表現目標基因之啟動子,
於低氧高光照的條件下被誘導。此研究中使用,分別將 xop2 及 xop2-xHtrII 放 在pJS005 載體的兩個建構質體。
II-1.3蛋白質藥品
第二節 化學藥品
藥品名稱 生產廠商
Acrylamide/Bis-acrylamide, v/v = 37.5:1, 40% solution
J. T. Baker
Agar Bioshop
Agarose (Halo-plate) Bioman
酵素名稱 生產廠商
DNA T4 ligase Yeastern Biotech Restriction enzyme Fermentas fast digest 2x PCR Dye Master Mix II (Taq; red) GeneMark
GoTaq® Green Master Mix (2x) Promega 2x Pfu PCR MasterMix (blue) Genomics Penta His HRP Conjugate kit QIAGEN
all-trans retinal SIGMA
Ampicillin Bioman
Beta-mercaptoethanol Bioman Commassie Blue R-250 Biobasic Diaminobenzidine (DAB-HCl) SIGMA DNA ladder 100 bp marker Bioman Gel extraction kit Geneaid HealthView Nucleic Acid Stain Genomics High-speed plasmid mini kit Geneaid Hydrochloric Acid 36.5-38.0 % J. T. Baker 30% Hydrogen peroxide SIGMA
Imidazole Fluka Analytical Isopropyl 𝛽-D-1-thiogalactopyranoside
(IPTG)
Bioman
Luria-Bertani Broth Neogen Magnesium chloride hexahydrate SIGMA
MES hydrate SIGMA
Methanol GENEStar
N-dodecyl-beta-D-maltoside (DDM) Anatrace Ni-NTA resin GE Healthcare Oxido LP0034 peptone Oxoid
Potassium chloride J. T. Baker Prestain protein marker Geneaid Soldium chloride SIGMA
Sodium citrate Fluka Analytical Tetramethylethylenediamine (TEMED) SIGMA
Tris-HCl AMRESCO
Trisodium chloride J. T. Baker
第三節 儀器設備
II-3.1 核酸電泳
迷你水平電泳槽及鑄膠器:霈璟實業 MJ-105 電源供應器:Bio-Rad PowerPac Basic Power Supply
II-3.2 蛋白質電泳及轉印
蛋白質垂直電泳鑄膠器:Bio-Rad
蛋白質垂直電泳槽:Bio-Rad Mini-PROTEAN 3 Elextrophoresis cell 蛋白質轉印槽:Bio-Rad Mini Trans-Blot Cell
電源供應器:Major Science (MP-250V power supply)
II-3.3 離心機
桌上型低速離心機:CUBEE Mini Centrifuge 桌上型離心機:KUBOTA KM-15200
低溫桌上型離心機(置冰箱內): GyroSpin Mini (10-001-04-0007)
落地式高速離心機:HITACHI CR 21 GII
落地式超高ㄉ速離心機:HITACHI Ultracentrifuge CP80WX
II-3.4 光學設備
分光光度計 HITACHI U-1900
ELISA Reader Molecular Device Spectramax 96 孔黑盤 Greiner Bio-one
顯微鏡架設 光學顯微鏡 OLYMPUS BH-2 RGB LED 燈泡 DL 夢想地; DSL5W-16 恆溫熱盤 KITAZATO MR-10DMF CCD camera MINTRON MTV-62V6HN
樣品薄片 載玻片 (KIMBLE)、蓋玻片、透明膠帶 (北極 熊)、凡士林 (Vaseline)
光週期測量架設 可調式超短脈衝雷射 LOTIS TII LS 2137/3
光電倍增管 Photomultiplier; Hamamatsu R928 PMT Socket Hamamatsu E717
高壓電源供應器 Stanford research system PS325 訊號放大器 Stanford research system SR570 示波器 Tektronix DPO4032
單光儀 Optometrics DMC1-02 監測白光光源 StellarNet SL1-Cuvette
其餘光學元件 Mirror and holder from OptoSigma
II-3.5 恆溫培養箱
Tks orbital shaking incubator OSI500R Biometra OV3 (37˚C for swam plate)
II-3.6 酸鹼度計
SUNTEX SP-701; EUTECH CyberScan pH 2100(連接至電腦即時偵測,菌體胞 外pH 值變化量測用); HORIBA (電極: 9669-10D)
II-3.7 其他
超音波清洗器:Branson 2510
超音波破菌機:Misonix Sonicator 3000
聚合酶鏈鎖反應器:Labneet MultiGene; Thermo PCR Sprint 試管振盪器:Spire mixer 5100
乾浴槽:PANTECH firefox dry bath 6120; Major Science (MD-mini) 恆溫水浴槽:Firstek Scientific
第四節 實驗方法
II-4.1 生物資訊學分析
II-4.1.1 基因資料庫
基因序列之比對藉由National Center for Biotechnology Information (NCBI) 網站 上之 Nucleotide Basic Local Alignment Search Tool (BLASTx) 分析比對。
II-4.1.2 序列親緣比對
蛋白質之間之親緣與序列相似比對,為將蛋白質序列上傳至 Center for Integrative Bioinformatics VU 網站,以 PRALINE - Multiple sequence alignment 之公開資源 分析。
II-4.1.3 蛋白質結構分析
蛋白質結構主要以分析軟體 PyMol (maintained and distributed by Schrödinger) 分 析,包含結構胺基酸位置,表面電荷分佈及胺基酸變異策略分析。若已解出結構 之蛋白質,則以其公開之 protein data bank (pdb) 檔案呈現;若無結構之發表,
則以相似之結構分析 (視紫質以 bacteriorhodopsin 為模板) 。對於未知結構的蛋 白質,以 ExPASY Swiss Model 找尋最佳模板,及結構模型的模擬。
II-4.2 DNA 建構及轉形
II-4.2.1 小量核酸萃取
藉由實驗方法 4.2.6 將目標質體遞送至大腸桿菌株 DH5𝛼 中,並將菌液塗布至 具 50 µg/ml ampicillin 之 LB 培養基,隔夜培養於 37℃。將單一菌落接種至約 6 ml 添加 ampicillin 的 LB 培養液中,37℃ 培養 12-16 小時,使其生長至生長 穩定期。其後,使用 High-speed plasmid mini kit 將質體從菌液中抽出: (1) 以 12,000 rpm、 30 秒將菌液收集至菌塊; (2) 依序加入 PDI, II, III 試劑 200, 200, 300 𝜇𝑙,使其溶菌,破菌及沈澱蛋白質 ; (3) 以 15,000 rpm、5 分鐘,將蛋白質 沈澱離心成塊狀 ; (4) 將上清夜 (含質體 DNA) 吸取至核酸親和濾管,並以 12,000 rpm、30 秒將其流過濾膜; (5) 以 Wash buffer 清洗濾膜兩次,方法同(4);
(6) 最後,以 50 𝜇𝑙之 Elution buffer 及 12,000 rpm、2 分鐘將質體析出。
II-4.2.2 聚合酶鏈鎖反應 (PCR)
分成一般PCR 及菌落 PCR。一般 PCR,將目標基因片段放大,並賦予其限制酶 切位於兩端,通常會接續方法 4.2.3 將其純化出來:分別加入 DNA 模板, 順向 引子,反向引子, 2x Mastermix (<1 kbp, 用 Omega® Master Mix taq; >= 1 kbp, 用 2x Pfu PCR Master Mix),無核酸切酶之水 1, 3, 3, 15, 8 𝜇𝑙,於 PCR 反應管。
菌落 PCR ,目的為初步檢測大腸桿菌轉形株之質體內含之目標基因大小:先於 PCR 反應管中加入順向引子,反向引子,2x PCR Dye Master Mix II,無核酸切酶 之水 1, 1, 7, 5 𝜇𝑙,並以牙籤將單一菌落先接種至新鮮 LB 培養皿後,再於反應 管中攪拌,使其懸浮。
II-4.2.3 DNA 膠體純化
使用 Gel Extraction kit 將 DNA 片段從瓊脂膠體內純化,流程如下: (1) 將具 有核酸片段之膠體切下,並放入微量離心管中; (2) 加入 500 𝜇𝑙 Gel/PCR solution,
並置60℃至膠體溶解; (3) 將混合液吸取至核酸親和濾管,並以 12,000 rpm, 30 秒 離下; (4) 以 600 𝜇𝑙 Wash Buffer 清洗兩次,方法同(3); (5) 以 20 𝜇𝑙 Elution buffer 靜置2 分鐘,離心 2 分鐘析出。
II-4.2.4 限制酶截切
(1) 將以下試劑依序加入 PCR 反應管中: DNA 載體,目標基因片段,10x buffer, 限制酶I, 限制酶 II 2.5, 10, 1.5, 0.5, 0.5 𝜇𝑙。(2) 靜置於 37℃中 2.5 小時。(3) 85℃、10 分鐘,使酵素失去活性。
II-4.2.5 DNA 黏合
(1) 將以下試劑依序加入 PCR 反應管中: 截切過之 DNA 載體及目標基因片段混 合液,10x buffer A, 10x buffer B, T4 ligase 7.5, 1, 1, 0.5 𝜇𝑙。(2) 靜置於 4℃中 12 小時或16℃中 4 小時。
II-4.2.6 大腸桿菌轉形
勝任細胞 (competent cell) 製作。 (1) 從凍菌,於 LB 培養皿中取出單一菌落,
接種至 3 ml LB broth, 37℃ 隔夜培養。(2) 次日,接種 500 𝜇𝑙至 500 ml LB,於 37℃培養至 OD600 = 0.3-0.5。(3) 以離心管收取,並置冰上 10 分鐘。(4) 以 3,500
rpm 離心後去除上清,再以 10 ml TB buffer 回溶後靜置冰上 10 分鐘。(5) 在離 心去除上清夜後,以2 ml TB buffer 回溶。(6) 每 100 𝜇𝑙分裝至微量離心管中,
並置-80℃存放備用。*TB (Transformation buffer): 15% glycerol, 2 mM NaOH, 10 mM MOPS, 85 mM CaCl2, 0.5& D-glucose
大腸桿菌轉形。(1) 將勝任細胞至於冰上 10 分鐘,待其退冰。(2) 將 1 𝜇𝑙質體加 入100 𝜇𝑙勝任細胞中,置冰上 15 分鐘。(3) 於 42℃,90 秒 (Heat shock)。(4) 靜 置冰上1 分鐘。(5) 加入 500 𝜇𝑙 LB,並於 37℃ 振盪培養 30 分鐘。(6) 取 100- 150 𝜇𝑙菌液塗布至具篩選標記之培養基。(7) 於 37℃隔夜培養,接續方法 4.2.8。
II-4.2.7 嗜鹽古生菌 H. salinarum 轉形
(1) 將嗜鹽古生菌 H. salinarum 畫線於 Halomedium plate 上 ; (2) 挑取單一菌落,
並接種至 2 ml CM+ medium,於 42˚C, 200 rpm,培養至 O.D.600 = 0.4 – 0.6 ; (3) 放大培養150 µl 菌液至 15 ml CM+ medium,於 42˚C, 200 rpm,培養大約 18 - 24 小時至O.D.600 = 0.4 – 0.5 ; (4) 以 15 ml falcon tube, 750 xg, 15 min,收 2 ml 菌 液 ; (5) 去除上清液後,以 200 µl Spheroplast solution (SPS) 溫和地搖盪懸浮菌 體,並於 2 小時內使用完畢 ; (6) 準備下列材料: a. 加 10 µl 0.5 M EDTA 到 falcon tube 中 b. 10 µl DNA + 20 µl SPS 30 µl c. 120 µl PEG600 + 120 µl SPS
50% PEG600 SPS ; (7) 將 200 µl 懸浮菌液 直接加於 10 µl 0.5 M EDTA 上 ; (8) 馬上接著加入 30 µl DNA,等待 5-10 分鐘 ; (9) 將 240 µl 50% PEG SPS 從管壁 緩慢流下,溫和振盪20-30 週期,等待 30 分鐘,溶液應該要沒有絲狀雜質 ; (10) 準備CM+ sucrose medium (CM+ medium:75% sucrose = 4:1) ; (11) 加入 5 ml CM+
sucrose medium 至管中,以去除 PEG ; (12) 2,000 rpm (750 xg), 15 min ; (13) 去除 上清,再以 5 ml CM+ sucrose medium 懸浮 ; (14) 37˚C, 130 rpm, 24 h,recover 菌 體 ; (15) 將 50 µl 菌液塗布於 SRMEV plate (10 µg/ml MEV) 上,42˚C 培養 ; (16) 24 小時後,再重複步驟 15 (10 天可以知道有或無,15 天可以收穫菌體)。
II-4.2.8 轉形株鑑定
初步挑選3 個單一菌落做菌落 PCR。並挑選出可能含有成功建構之質體的菌體,
做DNA 定序服務 (由基龍米克斯生物科技股份有限公司提供)。最後以西方墨點 法,確認目標蛋白質在轉形株中的表現。
II-4.3 重組蛋白質之表現及純化
II-4.3.1 重組視紫質表現
按方法4.2.6 將建構好之目標載體送入大腸桿菌株 C43 中。取單一菌落接種至 3 ml LB broth (50 𝜇g/ml ampicillin) 中,37℃ 培養 12 小時。再將 2 ml 菌液放大培 養至100 ml (50 倍稀釋),37℃ 隔夜培養至穩定生長期。於每一大瓶 LB 培養液 (800 ml) 中加入 16 ml 菌液,37℃培養約 1.5-2.0 小時至 OD600 = 0.4-0.6。加入 250 µM IPTG 及 5-10 nM all-trans retinal。37℃ 避光培養誘導 4 小時,進行純 化。
II-4.3.2 重組可溶蛋白質表現
按方法2-6 將建構好之目標載體送入大腸桿菌株 BL21 中。取單一菌落接種至 3 ml LB broth (50 𝜇g/ml ampicillin) 中,於 37℃ 培養 12 小時。再將 2 ml 菌液放大 培養至100 ml (50 倍稀釋),37℃ 隔夜培養至穩定生長期。於每一大瓶 LB 培養 液 (800 ml) 中加入 16 ml 菌液,於 37℃ 培養約 1.5-2.0 小時至 OD600 = 0.4-0.6,
靜置於 25˚C 半小時。加入 50 µM IPTG 誘導 1 小時。(GST 蛋白質於 37℃ 誘導 2 小時)
II-4.3.3 重組視紫質純化
將菌液以 6,000 rpm, 4˚C 離心 10 min,並用 30 ml 預冷過的 Lysis buffer (4 M NaCl, 50 mM Tris, pH 7.8) 回溶菌塊。以 lysis buffer 將體積補至 40 ml 後,加入 1.43 mM 2-ME 及 100 µM PMSF。設定超音波破菌機 (5 sec pulse, 5 sec rest, 共 5 min; 能量:69 W),將回溶後的菌液置入燒杯,放於冰上,啟動超音波破菌機。
將破菌後的均質液以 12,000 rpm, 4˚C 離心 10 min; 上清液進行超高速離心 (48,000 rpm, 70 min, 4˚C) 將細胞膜離下。以 2% DDM, lysis buffer 旋轉回溶(4˚C, 20 rpm, 16 h) 超高速離心後的膜塊。再以 18,000 rpm、離心 45 min, 於 4˚C 將未 回溶完全的殘骸去除,將上清液與預先以 20 mM imidazole, lysis buffer 洗過的 Ni-NTA 膠體混合,進行 Resin binding (4˚C, 20 rpm, 6 h)。在重力膠體管柱中,將 混合液中的液體流去 (flow through)。再分別以 20 mM, 50 mM, 250 mM imidazole, lysis buffer 流洗膠體。收集各個流洗液,並將 I250 的流洗液以 30 kD Amicon 濃縮後,透析至下一步試驗所需的溶液環境中。
II-4.3.4 重組可溶蛋白質純化 (Hexa-His-tagged)
將菌液以 6,000 rpm, 10 min, 4˚C 離心下來,並用 30 ml 預冷過的 Lysis buffer (50 mM NaCl, 50 mM Tris, pH 7.8) 回溶菌塊。以 lysis buffer 將體積補至 40 ml 後,
加入 1.43 mM 2-ME 及 100 µM PMSF。設定超音波破菌機 (5 sec pulse, 5 sec rest, 共5 min; 能量:69 W),將回溶後的菌液置入燒杯,必放於冰上,啟動超音波破 菌機。將破菌後的均質液以15,000 rpm, 20 min, 4˚C 離心; 上清夜與預先以 20 mM imidazole, lysis buffer 洗過的 Ni-NTA 膠體混合,進行 Resin binding (4˚C, 20 rpm, 1.5 h)。在重力膠體管柱中,將混合液中的液體流去 (flow through)。再分別以 20 mM, 50 mM, 250 mM imidazole, lysis buffer 流洗膠體。收集各個流洗液,並將 I250 的流洗液以 10 kD Amicon 濃縮後,透析至下一步試驗所需的溶液環境中。
II-4.3.5 重組可溶蛋白質純化 (GST-tagged)
將菌液以 6,000 rpm, 10 min, 4˚C 離心下來,並用 20 ml 預冷過的 Lysis buffer (50 mM NaCl, 50 mM Tris, pH 7.8)回溶菌塊,加入 1.43 mM 2-ME 及 100 µM PMSF。
設定超音波破菌機 (5 sec pulse, 5 sec rest, 共 4 min; 能量:69 W),將回溶後的菌 液置入燒杯,放於冰上,啟動超音波破菌機。將破菌後的均質液以48,000 rpm, 1 h 10 min, 4˚C 離心; 上清夜與預先以 lysis buffer 洗過的 GST-beads 膠體混合,進 行Resin binding (4˚C, 20 rpm, 2 h)。在重力膠體管柱中,將混合液中的液體流去 (flow through)。以 6.8 mg/ml glutathione, elution buffer (50 mM Tris, pH 8.0) 回沖 膠體後,進行蛋白質的溶出 (4˚C, 20 rpm, 12 h)。收集流洗液,並以 10 kD Amicon 濃縮後,加入 20 mM DTT 貯存。* GST-CheR 蛋白質必須在 4˚C 下進行純化,
避免大量沈澱生成。
II-4.3.6 H. salinarum 細胞膜之純化
挑出H. salinarum 及轉形株的單一菌落至 20 ml Halomedium,42˚C, 180 rpm, 光 照培養至 stationary phase。再次培養 16 ml 至 800 ml halomedium 中,同條件培 養約4 天,至 OD600 值為 1.0 - 1.4。將菌體以 6,000 rpm, 4˚C, 10 min 收下,並 用 20 ml cold MES buffer (4 M NaCl, 50 mM MES, pH 5.8) 回溶。超音波振盪 (“ON”: 5 sec; “OFF”: 5 sec; total 4 min; energy: 69 W) 將菌體破碎後,以超高速離 心機將細胞膜離下(48,000 rpm, 4˚C, 1 h 10 m)。之後以 2 ml cold MES buffer 再次 將細胞膜回溶 (嗜鹽古生菌細胞膜不需要界面活性劑即可回溶),4˚C, 12,000 rpm;
多餘殘骸去除。可見光譜的分析,以 MES buffer 稀釋 10 倍測量,光週期的分 析則以4 M NaCl, 50 mM (Tris/MES), pH (8.5/6.2) 稀釋五倍,最後以 pH meter 量 測pH 值。
II-4.4 蛋白質定量及定性
II-4.4.1 蛋白質定量
將蛋白質以12,000 rpm, 4˚C 將將沈澱及雜質離下。再將蛋白質存在的溶液 (Blank) 加 150 µl 在 cuvette 中,以拭鏡紙擦拭其觀測窗。在 Spectrometer U-1900 進 行 ”Baseline” (測量範圍:250 – 750 nm) 校正。將溶液倒出後,以去離子水洗 淨,再將蛋白質溶液150 µl 加入 cuvette 中,進行 ”Measure”。將目標蛋白質序 列,在ExPASy tool: ProtParam 中計算蛋白質的 Extinction coefficient。在按所測 得的280 nm 吸收值,以下列方程式計算蛋白質濃度:
𝐶𝑜𝑛𝑐𝑒𝑛𝑡𝑟𝑎𝑡𝑖𝑜𝑛 (𝑚𝑔
𝑚𝑙) = 𝑂𝐷 (𝑚𝑒𝑎𝑠𝑢𝑟𝑒𝑑) ∗ 𝑀𝑊 𝜀 (𝑒𝑥𝑡𝑖𝑛𝑐𝑡𝑖𝑜𝑛 𝑐𝑜𝑒𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑡)
II-4.4.2 蛋白質變性電泳
按下列表格配製聚丙烯醯胺膠體。
分離膠體 焦集膠體
膠體百分比 10% 12.5% 4%
A sol.(40%) 2.5 3.125 0.5
B solution 2.5 2.5 -
C solution - - 1.24
ddH2O 4.85 4.225 3.11
10% SDS 0.1 0.1 0.05
10% APS 0.05 0.05 0.1
總體積 (ml) 10 10 5
* Solution A : commercial acrylamide and bis-acrylamide mixture。Solution B : 1.5 M Tris, TEMED, pH 8.8。Solution C : 0.5 M Tris, TEMED, pH 6.8。
將Bio Rad 垂直電泳鑄膠器架好,玻璃板以 95% 酒精擦拭。將溶液分別配好後,
最後加入 APS 後,馬上將 4.5 ml 的分離膠體混合液注入玻璃板之間,再以 100 µl isopropanol 將膠體壓平。待約 30 min 分離膠體凝結後,以濾紙將 isopropanol 吸出。加入APS 後,將焦集膠體注滿玻璃板間,並放入尺梳。待其凝結後使用,
若不馬上使用,則保存於4˚C, TBE buffer (90 mM Tris, 80 mM Boric acid, 2.5 mM EDTA) 中。
將電泳槽架好,注入足夠的TBE buffer 於陰陽極槽,並將尺梳平行拿出。以微量 吸管P200 清洗樣品槽,並預跑 20-30 min。將蛋白質樣品 1:1 與 2x sample buffer (8% SDS, 15% glycerol, 100 mM Tris, 2 mM EDTA, 160 mM DTT, pH 6.8, 0.2 mg/ml bromophenol blue) 混合。若為可溶蛋白質,則以 95˚C 加熱 10 min; 若為膜蛋白 質,則以55˚C 加熱 30 min (或 42˚C, 1 h)。若為全細胞裂解分析,E. coli 以 95˚C 加熱菌液與 sample buffer 之混合液 10 分鐘; 嗜鹽古生菌則以 120 µl ddH2O 回 溶從 3-5 ml 菌液收集之菌體 (回溶即破菌),以超音波 microtip 破碎基因體 (“ON”: 1 sec; “OFF”: 1 sec; total 10 sec; energy: 6 W),,再與 2x sample buffer 混 合後,置於 55˚C, 30 分鐘。將 15 µl 的樣品及 2.5 µl protein marker 依序注入樣品 槽。以 4˚C, 60 V 執行電泳至追蹤染劑通過焦集膠體後,以 4˚C, 150 V (若含有膜 蛋白質樣品,則以100 V 執行) 繼續跑電泳至追蹤染劑跑至膠體底端。完成電泳 後,以 CBR 染劑 (1 g Coomassie brilliant blue R-250 in 250 ml ddH2O, 250 ml methanol, 50 ml acetic acid) 染色膠體 30 min,並以脫色液 (20% Methanol, 10%
Acetic acid) 脫色至蛋白質條帶出現。以玻璃紙將完成染色的膠體封起,並護貝 保存。
II-4.4.3 蛋白質原態電泳
同上表格配製膠體。 Solution B & C without TEMED (配製膠體時再加入 1%
TEMED)。* Running Buffer: 25 mM Tris, 192 mM glycine,2x sample buffer: 62.5 mM Tris, pH 6.8, 25% glycerol, 1% bromophenol blue。電泳及染色步驟同上,惟以 4˚C, 60 V 跑完電泳全程。
II-4.4.4 蛋白質轉印
將轉印 cassette 依序如下組裝:黑夾 海綿 濾紙 膠體 以 methanol 預先活化的PVDF 膜 濾紙 海綿 白夾(y94transfer buffer 中進行,並避 免氣泡產生)。將 cassette 置入電泳槽,並放入降溫冰塊。以 Transfer buffer (25 mM Tris, 192 mM glycine, pH 8.3, 10% methanol) 填滿,並以 400 mA, 1 h 執行轉 印。完成後,將 PVDF 膜取出,進行免疫呈色。
II-4.4.5 免疫呈色
將轉印後的 PVDF 膜取出,浸泡於 Urea-PBST, 1 h 至隔夜。以 TBS buffer 清 洗三次 (清洗步驟皆為 10 分鐘)。以 BSA 及 TBS 配製 Blocking buffer,並進行 blocking 至少至少 1 h。再以 TBS-Tween/Triton 清洗兩次,以 TBS 清洗一次。接 著浸泡於含 1/3000 Penta-His HRP 抗體的 Blocking buffer 搖晃 1 h。再來用 TBS-Tween/Triton 清洗兩次。於避光環境下加入 HRP staining solution 呈色 5 – 10 min,風乾後護貝。
II-4.5 感光蛋白質光學分析
II-4.5.1 吸收光譜測定
同方法4-1.中的 ”Baseline” 及 ”Measure”。
II-4.5.2 視紫質光週期量測
將蛋白質樣品定量至特徵吸收峰 = 0.3 - 0.6,取 150 µl 置入三面透光的石英比色 管中。比色管單一光徑上只有一個開口的那端接受20 U 焦耳強度的 6 ns 雷射脈 衝 (可調整波長,若未特別說明則為 532 nm)。相對的兩個開口分別連接至:白 光監測光源; 單光儀 (選擇偵測波長) 光電倍增管 (放大訊號) 示波器 (擷取訊號)。如此重複數個週期,並以平均模式計算。
II-4.5.3 光驅動離子幫浦活性測定
將誘導表現視紫質的大腸桿菌200 ml,以 3,500 rpm, 15 min, 25˚C 收集。再用 50 ml unbuffered solution (10 mM NaCl, 10 mM MgSO4, 100 µM CaCl2)清洗兩次。再 以25 ml unbuffered solution 懸浮後,保存於黑暗中。使用時,以 unbuffered solution 調整OD600 至大約 2.0,並置於裝有攪拌磁石的玻璃瓶中,置於磁攪拌台上。攪 拌的同時,以pH meter 測量 pH 值,紀錄有照射光源及黑暗的條件下,細胞外環 境的pH 值變化。
II-4.6 嗜鹽古生菌生理分析
II-4.6.1 泳動菌株挑選
將H. marismortui, H. salinarum 及轉形株 HsSRM, HsSRM-HtrM於高鹽培養基劃四區,
並挑取單一菌落接種至1 ml 液態培養基中。待生長至 stationary phase 後,取 10 µl 菌液,接種至 20 ml 泳動培養基 (swam plate; halo-medium w. 0.3% agar) ;重