1 *Corresponding author: wnt@ntu.edu.tw
Otolith Sr:Ca Ratios as Natural Mark to
Discriminate the Restocked and Naturally
Recruited European Eels in Latvia
Y
u-J
iaL
inInstitute of Fisheries Science, National Taiwan University, Taipei, Taiwan, R.O.C.
J
en-C
hiehS
hiaoInstitute of Oceanography, National Taiwan University, Taipei, Taiwan, R.O.C.
M
ariSP
LikShSLatvian Fish Resources Agency, State Agency, Ministry of Agriculture, Republic of Latvia
a
rtiSM
indeLatvian Fish Resources Agency, State Agency, Ministry of Agriculture, Republic of Latvia
Y
oShiYukii
izukaInstitute of Earth Sciences, Academia Sinica, Nankang, Taipei, Taiwan, R.O.C.
i
Saakr
aShaLInstitute of Biology, University of Latvia, Salaspils, Republic of Latvia
W
ann-n
iant
zeng*Institute of Fisheries Science, National Taiwan University, Taipei, Taiwan, R.O.C. and
Department of Life Science, National Taiwan Unversity, Taipei, Taiwan, R.O.C.
2
Introduction
European eel Anguilla anguilla is a facul-tative catadromous fish spawning in the Sar-gasso Sea and living in the freshwater, brack-ish and costal waters in Europe (Tsukamoto et al. 1998; Tzeng et al. 2000; Tesch 2003). In Latvia, A. anguilla was one of the most expensive fish that fishermen were most in-terested in. To enhance the local eel produc-tion in Latvia, the restocking plan has been conduct since 1927. Approximately 30 mil-lion of glass eels imported from France were regularly released in 51 Latvian lakes during 1960 to 1988 by the government. From 1990 to present, numerous glass eels were released in inland freshwaters by private enterprises, e.g. fishing rights or lakes leaseholder.
Although the restocking plans were con-ducted for a long time in Latvia, the follow-ing restockfollow-ing efficiency was seldom evalu-ated, nor was the contribution of restocked eels to the local population examined. This might be due to the lack of appropriate marks or tags to discriminate the restocked eels from naturally-recruited eels. Recent stud-ies indicated that the otolith Sr:Ca ratio is a useful natural mark to discriminate fish from freshwater and saltwater habitats (Campana
1999; Campana and Thorrold 2001), and has been used to reveal the migratory histories of A. anguilla between freshwater and saltwa-ter (e.g., Tzeng et al. 1997; Tsukamoto et al. 1998; Tzeng et al. 2000).
The glass eels being restocked were purchased in the estuaries in France, trans-ported across the Baltic Sea and released in the planned freshwater lakes in the middle reaches of the river. The naturally-recruited ones entered the brackish Baltic Sea and mi-grate across the Baltic Sea to reach the south eastern Baltic countries. The differences in route to Latvian waters resulted in different saltwater experience between restocked and naturally-recruited eels, which can be re-vealed by otolith Sr:Ca ratios. This approach has been applied to examine the restocking efficiency in Lithuania (Shiao et al. 2006) and the growth differences between restocked or naturally-recruited eels (Lin et al. (2007b). However, the restocking program efficiency was still unclear in Latvian waters, nor was the growth compared between migratory his-tory types and eel origins.
The objectives of this study were (1) to identify the origins of eels (restocked or nat-urally-recruited) from three Latvian waters: river mouth (Daugavgriva), nearby lakes
Abstract.—The restocking programs of the European eel Anguilla anguilla have
been conducted for nearly one century in Latvia. To evaluate the efficiency of the eel restocking program and reveal the migratory life histories of European eels in Latvian waters, a total of 75 individuals were collected from the mouth of River Daugava (Daugavgriva, brackish), nearby lake (Lake Ķīšezers, freshwater), and a coastal site (Mērsrags, brackish). The naturally-recruited eels consisted of two saltwater type in-dividuals (SW, 0–7%): eels lived in saltwater and did not enter freshwaters, and inter-habitat-shifter (IHS, 60–85%): eels had experienced both freshwater and saltwater. The restocked eels consisted of purely freshwater type (FW, 7–36.7%) that the eels permanently lived in freshwater without saltwater experience. Average otolith Sr:Ca ratios in the edge were different among sites and origins, and corresponded to the salinities of sampling sites. The mean age at first freshwater entry of IHS was 4.8 ± 2.3 years, similar to previous study in Lithuania (p = 0.188). The growth rates of FW eels in the river mouth and coast were significantly slower than that of IHS eels (p < 0.001 and 0.012), but not in the lake (p = 0.372), implying a site-origin interaction on the growth rate. The use of otolith Sr:Ca ratios as a natural mark to distinguish the restocked eels enabled the assessment of restocking efficiency in the future.
Otolith Sr:Ca Ratios as Natural Tags
(Lake Ķīšezers) and Baltic coastal region (Mērsrags) by otolith Sr:Ca ratios, (2) to ex-amine the growth differences of eels among different migratory history groups and ori-gins, and (3) to compare the results in Latvia with previous studies in Lithuania (Shiao et al. 2006; Lin et al. 2007b).
Methods
Study site
European eels A. anguilla were collected by fyke-nets from Daugavgriva, the mouth of River Daugava, during August to September, 2005, as well as from nearby Lake Ķīšezers and a coastal site, Mērsrags, during August to September 2006 in Riga Gulf (Figure 1). Riga Gulf is located in the eastern part of the Baltic with the salinity ranging from 2 to 7 ‰, depending on regions and seasons. The Latvian coastline of Riga Gulf is about 250 km and the average depth of the coastal zone (2 mi) is around 10 m. Bottom vegetation in
the coastal waters of the gulf is unevenly dis-tributed. The brown algae are dominant, fol-lowed by the filamentous green algae and red algae. In general, Riga Gulf can be consid-ered as highly eutrophic.
Lake Ķīšezers is located close to the coast and connected with the Baltic Sea through lower reach of River Daugava. It is a fresh-water lake but during strong western winds brackish water from Gulf of Riga can enter the lake for a short time. The area of this lake is 1,730 ha with an average depth of 2.4 m and a maximum depth of 4.5 m in the central part. Lake Ķīšezers is also characterized by brown color of water and low transparency, which is around 0.5 m in the summer.
River Daugava is the largest river in Lat-via with the largest river basin as well. The width of the lower reach of River Daugava is around 500 m and the depth is around 15 m. The sampling site in the river mouth is brackish with a mean salinity of 4.2 ‰. Three big hydroelectric plants were constructed on Daugava and one located just 28 km away
Figure 1. Sampling sites of European eel Anguilla anguilla in Gulf of Riga in Latvia. (1) Mouth of River
4
from the river mouth without equipped fish-pass. Therefore, the upstream migration of diadromous fish in River Daugava is cut off, but downstream migration is believed to be limited.
The total length (TL) and weight (TW) of the eel were measured to the nearest 1.0 mm and 1.0 g, respectively. Sexes were deter-mined macroscopically from the gross mor-phology of the gonads, where eels with thin, regularly lobed organs (Syrski’s organ) were considered males, while individuals with more broad and folded curtain-like gonads were females (Tesch 2003). Nearly all eels caught were females except for one male, which was excluded in the analysis.
Otolith Sr:Ca ratio measurement and age determination
The largest pair of otoliths (sagittae) was removed, dried in the air, embedded in Epo-fix resin, ground and polished until the pre-mordium was exposed. The polished otoliths were coated with carbon under a high-vacu-um evaporator. Sr and Ca concentrations in the otolith (wt %) were measured from the primordium to the edge at 10 µm intervals by electron probe microanalyzer (EPMA, JEOL JXA-8900R) with the same conditions and standards described by Lin et al. (2007b). The critical otolith Sr:Ca ratio indicating fresh-water residency was set at 2.24 × 10–3 (Shiao et al. 2006) and the region where two con-secutive points were lower than the critical value was regarded as freshwater experience (Jessop et al. 2007). If the EPMA transect passed through the vaterite inclusion, it was not used and an alternative one without pass-ing the vaterite inclusion was conducted to avoid the misidentification of the migratory group (Tzeng et al. 2007; Jessop et al. 2008). The otolith edge was deposited when the fish is caught, and thus contains environmental information about the sampling site. There-fore, otolith ratios of the last three points to
the otolith edge were averaged. To evaluate whether the different salinities among sites and eel origins can affect average otolith Sr:Ca ratios in the edge, the average values were compared by sites and origins.
After EPMA analysis, the otoliths were re-polished to remove the carbon layer and etched with 5% EDTA for 1–2 min to reveal annual rings. The age of the fish was estimat-ed by counting the number of annual rings and the growth rate (GR) was calculated as (TL – L0) × Age–1 where L
0 is mean length of elvers (Wang and Tzeng 2000). The age of first freshwater entry was estimated by counting the number of rings corresponding to where the otolith Sr:Ca ratio below the freshwater criterion.
Data Analysis
The proportions of restocked eels be-tween sites were compared by chi-square test. The average of otolith Sr:Ca ratios in the edge were compared by sites and origins using two-way analysis of variance (ANO-VA) with Turkey’s multiple comparison. The total length, age and growth rate were log-transformed to meet the assumption of normal distribution (KS Test, all p > 0.08) and homogeneity of variance among groups (Levene’s Test, all p > 0.29). They were then compared among sites and migratory history types by two-way ANOVA. The effects of sites, origins and site-origin interaction on log-transformed growth rates were compared by two-way ANOVA. Although the distribu-tion of the age of first freshwater entry was not normally distributed for eels from Latvia and Lithuania (KS test, all p < 0.001), they were homogeneous to each other (KS test, p = 0.64). Therefore, the age at first fresh-water entry of the eels between Latvia and Lithuania were compared by nonparametric Wilcoxon Rank Sum test. All the statistical computation were completed in SAS®
(Ver-Otolith Sr:Ca Ratios as Natural Tags
sion 8.01) and the significance level α was at 0.05.
Results
Migratory history types and origins of the eels
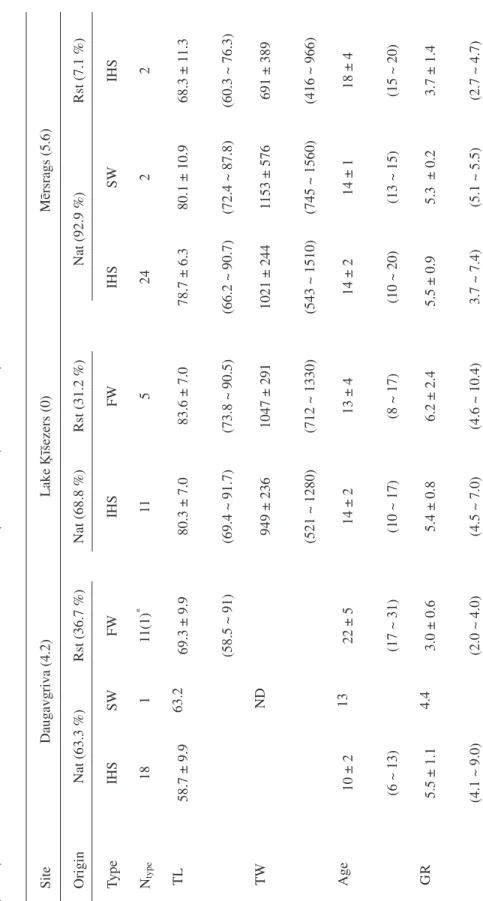
A total of 75 eels were collected, includ-ing 31 in Daugavgriva, 16 in Lake Ķīšezers and 28 eels in Mērsrags. All eels were fe-males except one caught in Daugavgriva, which was excluded in subsequent analysis. The Sr:Ca ratios of the eels after elver stage were classified into saltwater type (SW): the otolith Sr:Ca ratios of the eels were consis-tently higher than the critical value of 2.24 × 10–3, indicating that the eels lived in salt-water since elver stage until being caught (Figure 2a), inter-habitat- shifter (IHS): the ratios were initially higher and then varied between the critical value, indicating that the eels moved between saltwater and freshwater (Figure 2b) and freshwater type (FW): the ra-tios were consistently lower than the critical value, indicating they persistently stayed in the freshwater since elver stage until being caught (Figure 2c). The eels examined were mainly IHS, followed by FW with very few SW individuals. Eighteen IHS, 12 FW, and 1 SW eels were found in Daugavgriva, 11 IHS and 5 FW eels were found in Lake Ķīšezers, and 26 IHS, 0 FW, and 2 SW eels were identi-fied in Mērsrags (Table 1).
Naturally-recruited (Nat) eels have ex-perienced saltwater after metamorphosis to elvers when migrating across the brackish Baltic Sea, and the restocked (Rst) eels did not experience saltwater. So Nat eels were equivalently SW and IHS eels, and Rst eels were the FW eels. Although from brackish Mērsrags, there are two IHS eels considered as Rst eels, because they entered freshwater after metamorphosis. At radius about 750 and 1500 µm, they moved to saltwater and showed
elevated Sr:Ca ratios representative of brack-ish coast water (Figure 3). The proportions of Rst eels was 36.7% in Daugavgriva, 31.2% in Lake Ķīšezers and 7.1% in Mērsrags, sig-nificantly different among sites χ2 Test, p = 0.015) (Table 1). The temporal changes of otolith Sr:Ca ratios differed substantially be-tween Nat and Rst eels. The average otolith Sr:Ca ratios for the Nat eels were approxi-mately 5 × 10–3 at the elver stage and gradu-ally decreased to 2 × 10–3 at the edge. The average ratios of Rst eels remained at a low value of around 1.5 ×10–3 from the elver stage until the edge. The 95% confidence interval for the mean did not overlap until at radius of around 1000 µm (Figure 4), indicating their different migratory histories.
Average otolith Sr:Ca ratio in the edge among sites and origins
For Nat eels, the average ±SD) otolith Sr:Ca ratio in the edge were 2.69 ± 1.16, 1.35 ± 0.92 and 2.33 ± 1.26 × 10–3 in Daugavgri-va, Ķīšezers and Mērsrags, respectively. For Rst eels, they were 0.70 ± 0.41, 0.82 ± 0.74 and 2.29 ± 0.02 × 10–3 (Table 2). The average edge ratios of eels were significantly different by sampling sites and eel origins (Two-way ANOVA, p = 0.03 and <0.001, respectively). When compared between origins by sites, the Nat eels had significantly higher average edge ratios than Rst eels in Daugavgriva (p < 0.001). However, their edge ratios were not significantly different between origins in Ķīšezers and Mērsrags (p = 0.28 and >0.5, respectively). When compared among sites by origins, the edge ratios of Nat eels from Daugavgriva and Mērsrags were significant-ly higher than those of Nat eels from Ķīšezers (p = 0.01), corresponding to the salinity of sampling sites. The edge ratios of Rst eels from Mērsrags were significantly higher than those from Daugavgriva and Ķīšezers (p = 0.003), because the two Rst eels have moved to the coast (Figure 3).
6
Figure 2. Otolith Sr:Ca ratios of (a) saltwater type (SW), (b) inter-habitat-shifter (IHS) and (c)
freshwater-type (FW) eels. The grey area under the broken line (Sr:Ca ratio = 2.23 × 10–3) indicated the freshwater
Otolith Sr:Ca Ratios as Natural Tags
Otolith annulus patterns and age at first freshwater entry
The otolith annulus patterns of Nat and Rst eels were shown in Figue 5a and b. The region from the primordium (P) to the el-ver check (EC), which corresponded to the marine larval period from leptocephalus to the elver stage, was dark in appearance after etching by HCl. After EC, clear circular an-nuli were deposited in the otolith and some less-clear pseudo-annuli were found be-tween the success annuli. The otolith annu-lus patterns were generally similar between the Nat (Figure 5a) and Rst eels (Figure 5b) except that more annuli were found in Rst eels because of older ages. The mean ±SD) age at first freshwater entry of the IHS, de-termined by otolith Sr:Ca ratios (Figure 2b) , was 4.8 ± 2.3 years and it ranged from 1 to 11 years, which was not significantly differ-ent from the eels from Lithuania (Shiao et al. 2006; mean ±SD = 5.2 ± 2.1 year, Wilcoxon Rank Sum test, p = 0.19). There were seem-ingly two peaks in the age at first freshwater
entry in both countries that 55–57% at age 3–5 years and 23–28% at age of 7–9 years (Figure 6).
Growth rate comparisons between sampling sites and migratory history types
FW (equivalently Rst) eels had signifi-cantly smaller total length and older age than the IHS (Nat) eels (two-way ANOVA, p = 0.028 and <0.001) in Daugavgriva, but the dif-ferences in total length and age between FW and IHS eels were not significant in Mērsrags (p = 0.09 and 0.13) and Lake Ķīšezers (p = 0.40 and 0.60). The growth rates were signifi-cantly influenced by sampling sites, migratory history types and their interaction (two-way ANOVA, all p < 0.001). When the sites were separated, the growth rate of FW eels was significantly slower than that of IHS eels (p < 0.001) in Daugavgriva and Mērsrags (p = 0.01). However the growth rate was not signif-icant difference between FW and IHS eels in Lake Ķīšezers, (F = 0.85 p = 0.37) (Table 1).
8 T a b l e 1. Anguilla anguilla collected in Daugavgriva, Lake Ķī šezers and M ē rsrags in Latvia with mean salinities (‰) in the brackets. Per centage by origins (Nat: natural-r ecruited, Rst: restocked eels) and numbers of eels by migrator y histor y types (Ntypes, IHS: inter -habitat- shifters, SW : saltwater type, FW : fr eshwater type eels). Mean (± SD, ranges in par enthesis) total length (TL, cm) and weight (TW , g), age (years) and gr owth rate (GR, cm year -1). ND = data wer
e not available.*: All wer
e females except one male (TL = 55.9 cm).
Site D au ga vg riva (4.2) L ak e Ķ īšezers (0) M ēr sr ag s (5 .6 ) O ri gi n Nat (63.3 %) Rst (36.7 %) Nat (68.8 %) Rst (31.2 %) Nat (92.9 %) Rst (7.1 %) Ty pe IH S SW FW IH S FW IH S SW IH S Nty pe 18 1 11 (1 ) * 11 5 24 2 2 T L 58.7 ± 9.9 63.2 69.3 ± 9.9 (58.5 ~ 91) 80.3 ± 7.0 (69.4 ~ 91.7) 83.6 ± 7.0 (73.8 ~ 90.5) 78.7 ± 6.3 (66.2 ~ 90.7) 80.1 ± 10.9 (72.4 ~ 87.8) 68.3 ± 1 1.3 (60.3 ~ 76.3) T W N D 949 ± 236 (521 ~ 1280) 1047 ± 291 (712 ~ 1330) 1021 ± 244 (543 ~ 1510) 1153 ± 576 (745 ~ 1560) 691 ± 389 (416 ~ 966) A ge 10 ± 2 (6 ~ 13) 13 22 ± 5 (17 ~ 31) 14 ± 2 (10 ~ 17) 13 ± 4 (8 ~ 17) 14 ± 2 (10 ~ 20) 14 ± 1 (13 ~ 15) 18 ± 4 (15 ~ 20) G R 5.5 ± 1.1 (4.1 ~ 9.0) 4.4 3.0 ± 0.6 (2.0 ~ 4.0) 5.4 ± 0.8 (4.5 ~ 7.0) 6.2 ± 2.4 (4.6 ~ 10.4) 5.5 ± 0.9 3.7 ~ 7.4) 5.3 ± 0.2 (5.1 ~ 5.5) 3.7 ± 1.4 (2.7 ~ 4.7)
Otolith Sr:Ca Ratios as Natural Tags
Figure 3. Mean (± 95 % CI) otolith Sr:Ca ratios of naturally-recruited (black line, n = 56) and restocked
(grey line, n = 18) eels.
Figure 4. Sagittal plan of otoliths of (a) naturally-recruited with Sr:Ca profiles in Figure 2b and (b)
restocked eels in Figure 2c. P = primordium, EC = elver check. 1st FE = 1st freshwater entry. Black cycles are annuli. The white line from the primordium to edge was the track where Sr:Ca ratios were measured. Scale bar = 1 mm.
10
Figure 5. The otolith Sr:Ca ratios of two restocked eels from Mērsrags. The grey area under the black
broken line (Sr:Ca ratio = 2.23 × 10–3) indicated the freshwater experiences
Table 2. Mean (± SD, ranges in parenthesis, × 10–3) of the average otolith Sr:Ca ratio of last three points
in the edge in naturally-rectuited (Nat) and restocked eels (Rst) by sites. Different superscripts indi-cated significant difference in otolith Sr:Ca ratios among sites. The asterisk in Daugavgriva indiindi-cated that the average edge ratios of Nat eels significantly differed from the ratios of Rst eels.
Site Daugavgriva* Ķīšezers Mērsrags
Nat 2.69 ± 1.16a 1.35 ± 0.92b 2.33 ± 1.26a
(0.00 ~ 4.24) (0.13 ~ 3.03) (0.50 ~ 4.59)
Rst 0.70 ± 0.41a 0.82 ± 0.74a 2.29 ± 0.02b
Otolith Sr:Ca Ratios as Natural Tags
Discussion
Implication of different migratory history types
The dominance of IHS indicated that the movement of the eels from brackish Baltic waters to freshwater habitats was frequent in Latvian waters that a majority of the eels have entered freshwaters at least once. This was also in consistent with the eels in Lithu-anian (Shiao et al. 2006). The sampling sites were either near or in the coastal waters in Latvia, suggesting that the freshwaters con-necting to the Baltic Sea appeared to be as crucial as the coastal waters for the European eels, which have flexible and plastic habitat uses (Daverat et al. 2006).
Proportion of Rst eels
If some of the Rst eels in Latvian waters came from other countries, such as Lithuania, they had to move though the brackish
Bal-tic, leaving elevated otolith Sr:Ca ratios. All but two in Mērsrags entered freshwater and stayed there until being caught, so the Rst eels are probably not from other countries. The higher proportions of Rst eels in Lake Ķīšezers and Daugavgriva than in Mērsrags appeared reasonable because Rst elvers were released in some midland lakes which have connection with the Gulf of Riga through River Daugava. However, the outlets of these lakes were equipped with eel weirs, which covered practically most eel escapements. In addition, downstream migration of eels which have overcome the eel weirs was believed to be further limited by the hydroelectric power station dams without fish ladders.
The existence of Rst eels in Daugavgriva and Lake Ķīšezers implied that at least some Rst eels have passed these obstructions suc-cessfully and they were able to be caught in the waters in the lower reach of River Daugava and the Baltic coast. Sampled eels were com-posed of both yellow and silver eels and there-fore the spawning migration of silver Rst eels
Figure 6. Age of first freshwater entry for the naturally-recruited eels from Latvia (grey, mean (± SD) =
12
(Limburg et al. 2003; Westin 2003) appeared not as limited by the obstructive constructions as previously considered. It was unclear how these eels passed these artificial obstructions, probably through the period of water dis-charge. Although the eels can pass these pow-er plants, they also suffpow-er additional mortality from 15 to 38% (ICES 2007). Therefore, the contribution of Rst eels to downstream sites in Latvia is still constrained.
On the other hand, the proportions of Rst eels in river mouth (36.7%) and coastal lake (31.2%) in Latvia were substantially higher than that in the Curonian Lagoon in Lithuania (20%, Shiao et al. 2006). It was also slightly higher in coastal waters in Latvia (7.1%) than in Lithuania (2%). Numerous factors might result in this phenomenon, such as the number and frequencies of glass eels released, days, months, seasons and years of releasing, the geographic features, current systems, behav-ioral differences, productivity of the waters where the eels were released and the degree of connectivity between upper and lower reach-es. So far, it has been premature to conclude that the restocking efficiency was different between the two countries examined.
Average otolith Sr:Ca ratios in the edge
The average otolith Sr:Ca ratios in the edge of Nat eels corresponded to the salinities of sampling sites, indicating that the average Sr:Ca ratios of last three points were able to distinguish the brackish sites of 4.2–5.6 ‰ from the freshwater sites (Lin et al. 2007a). Insignificant differences in the average edge ratios between Nat and Rst in Lake Ķīšezers and Mērsrags also suggested that the average edge ratios can be used to represent the salini-ty of sampling site. However in Daugavgriva, the edge ratios of Rst eels were significantly lower than those of Nat eels. It suggested that other factors, such as eel origins, should still be considered when applying the otolith Sr:Ca ratio in the edge as an empirical
indica-tor of habitat salinity. This might also imply that these Rst eels have stayed in midland freshwaters for a certain time, and they were caught soon after moving to Daugavgriva.
Age at first freshwater entry
The similarity in age at first freshwa-ter entry between eels in Latvia and Lithu-ania implied the similar migration behavior of IHS eels from the opening of the Baltic Sea to the freshwaters. Because the eels were sampled in different years (2003 to 2004 in Lithuania and 2005 to 2006 in Latvia), it also indicated that the migration of the eels into freshwater was in some degree spatially and temporally persistent in the studied sites dur-ing investigated period. However, this mi-gration behavior exhibited certain degree of individual variation, as indicated by the large range from 1 to 11 years. Around the 1910s to 1920s, the elvers were found to enter fresh-water in the northeastern and eastern regions of Sweden and southern regions of Lithuania (Figure 6 in Dekker 2003a), suggesting that the eels were able to cross the Baltic Sea with a young age. But the drastic decrease in re-cruitment of European eels in recent years (Dekker 2003b) might discourage the eels from migrating eastward (Shiao et al. 2006). Two modes at age of 3–5 and 7–9 implied that the migration of the eels toward freshwa-ters might be composed of two subtypes, one moved to freshwater quickly and the other did slowly. This bimodality might be partly explained by the different types of eel fresh-water movement, namely founders, pioneers, home range dweller and nomads (Feunteun et al. 2003).
Differences in growth rates between migratory history types and eel origins
Because the SW eels were few, the growth comparison was focused on IHS and FW eels, equivalently between Nat and Rst eels. The
Otolith Sr:Ca Ratios as Natural Tags
growth rates of FW (Rst) eels were smaller than those of IHS (Nat) eels in Daugavgriva and Mērsrags, but they did not differ in Lake Ķīšezers. The productivity was similar among the three studied sites, and the slower growth rate of FW eels in Daugavgriva might be due to that the FW eels have escaped from mid-land lakes with low productivity, in which the growth of the eel was slow (Lin et al. 2007b). Previous studies in Lithuania suggested that the sampling sites might be the major fac-tor influencing the growth of the eels rather than origins (Lin et al. 2007b). The signifi-cant site-type interaction in this study further suggested that the influences of sampling site and eel origins on the growth of the eels might be more complicated and appeared to be site-specific.
In conclusion, the eels collected from three Latvian waters were composed of main-ly IHS eels, followed by the FW and the SW eels were the least. The Rst eels contributed around 30% of the total eel caught, suggest-ing the substantial contribution of Rst eels to the eel production in three sampling sites. The average otolith Sr:Ca ratios of last three measuring points were able to represent the salinity of sampling site. The otolith annulus pattern was similar between Nat and Rst eels except more annuli found in the otoliths of Rst eels. The age at first freshwater entry of the eels was similar between eels from Lat-vian and Lithuanian waters, indicating the migration across the Baltic Sea was spatially and temporally persistent during investigated period. The growth of FW (Rst) eels was not consistently slower than the IHS (Nat) eels in the sampling sites, indicating a complicated site–origin interaction on the growth of the eel. This study further suggested the impor-tance of otolith Sr:Ca ratio as natural mark to discriminate the restocked European eels from naturally-recruited eels in the Baltic countries. The eels in the Baltic regions were still less studied and future research efforts were still needed.
Acknowledgment
We are grateful to Lithuania-Latvia-Tai-wan (Republic of China) Mutual Fund for the financial support of the collaborative research project between the countries.
References
Campana, S. E. 1999. Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Marine Ecology Progress Series 188:263–297.
Campana, S. E., and S. R. Thorrold. 2001. Oto-liths, increments, and elements: keys to a comprehensive understanding of fish popu-lations? Canadian Journal of Fisheries and Aquatic Sciences 58:30–38.
Daverat, F., K. E. Limburg, I. Thibault, J. C. Shiao, J.J. Dodson, F. Caron, W. N. Tzeng,, Y. Iizuka, and H. Wickström. 2006. Phenotypic plasticity of habitat use by three temperate eel species, Anguilla anguilla, A. japonica and A. rostrata. Marine Ecology Progress Se-ries 308:231–241.
Dekker, W. 2003a. On the distribution of the Euro-pean eel and its fisheries Canadian Journal of Fisheries and Aquatic Sciences 60:787–799. Dekker, W. 2003b. Epilogue: worldwide decline
of eel resources necessitates immediate ac-tion Fisheries 28:28–30.
Feunteun, E., P. Laffaille, T. Robinet, C. Briand, A. Baistez, J. M .Oliver, A. Lagault, and A. Acou. 2003. Movements of anguillid eels in fresh-water habitats. Pages 191–213 in K. Aida, K. Tsukamoto, and K. Yamauchi, editors. Ad-vances in eel biology. Springer, Tokyo. ICES. 2007. Report of the 2007 Session of the
Joint EIFAC/ICES Working Group on Eels. ICES CM 2007/ACFM:23.
Jessop, B. M., J. C. Shiao, Y. Iizuka, and W. N. Tzeng. 2007. Effects of inter-habitat migra-tion on the evaluamigra-tion of growth rate and habitat residence of American eels Anguilla
rostrata. Marine Ecology Progress Series 342:255–263.
Jessop, B. M., J. C. Shiao, Y. Iizuka, and W. N. Tzeng. 2008. Prevalence and intensity of
oc-14
currence of vaterite inclusions in aragonite otoliths of American eel Anguilla rostrata. Aquatic Biology 2:171–178.
Limburg, K. E., H. Wickström, H. Svedäng, M. Elfman, and P. Kristainsson. 2003. Do stocked freshwatr eels migrate? Evidence from the Baltic suggests “Yes.” Pages 275–284 in D. A. Dixon, editor. Biology, management, and protection of catadromous eels. American Fisheries Society, Symposium 33. Bethesda, Maryland.
Lin, S. H., C. W. Chang, Y. Iizuka, and W. N. Tz-eng. 2007a. Salinities, not diets, affect stron-tium/calcium ratios in otoliths of Anguilla
japonica. Jounal of Experimental Marine Biolology and Ecology 341:254–263. Lin, Y. J., L. Ložys, J. C. Shiao, Y. Iizuka, and W.
N. Tzeng. 2007b. Growth differences between naturally recruited and stocked European eel
Anguilla anguilla from different habitats in Lithuania. Journal of Fish Biology 71:1–15. Shiao, J. C., L. Ložys, Y. Iizuka, and W. N. Tzeng.
2006. Migratory patterns and contribution of stocking to the population of European eel in Lithuanian waters as indicated by otolith Sr:Ca ratios. Journal of Fish Biology 69:749– 769.
Tesch, F.W. (2003) The eel, 3rd edition. Black-well Scientific Publications, Oxford, U.K. Tsukamoto, K., I. Nakai, W. V. Tesch. 1998. Do all
freshwater eels migrate? Nature 396:635–636. Tzeng, W. N., K. P. Severin, H. Wickström. 1997.
Use of otolith microchemistry to investigate the environmental history of European eel
Anguilla anguilla. Marine Ecology Progress
Series 149:73–81.
Tzeng, W. N., C. H. Wang, H. Wickström, M. Reizenstein. 2000. Occurrence of the semi-cat-adromous European eel Anguilla anguilla (L.) in Baltic Sea. Marine Biology 137:93–98. Tzeng, W. N., C. W. Chang, C. H. Wang, J. C.
Shiao, Y. Iizuka, Y. J. Yang, C. F. You, L. Lo[icirc]zys. 2007. Misidentification of the migratory history of anguillid eels by Sr/Ca ratios of vaterite otoliths. Marine Ecology Progress Series 348:285–295.
Wang, C. H., and W. N. Tzeng. 2000. The tim-ing of metamorphosis and growth rates of American and European eel leptocephali: A mechanism of larval segregative migration. Fisheries Research 46:191–205.
Westin, L. 2003. Migration failure in stocked eels
Anguilla anguilla. Marine Ecology Progress