國立臺灣大學生物資源暨農學院食品科技研究所 碩士論文
Graduate Institute of Food Science and Technology College of Bio-Resources and Agriculture
National Taiwan University Master Thesis
不同品種甘薯葉乙醇萃取物抗氧化及 改善葡萄糖攝入之研究
Antioxidation and glucose uptake improvement by ethanol extracts from leaves of different sweet potato cultivars
劉斌雁
Penk-Yeir Low
指導教授:吳瑞碧博士 共同指導教授:施明智博士
Advisor: James Swi-Bea Wu, Ph. D.
Co-advisor: Ming-Chih Shih, Ph. D.
中華民國一百零二年七月
July, 2013
中文摘要
農產品及蔬果中所含之植物化學物質 (phytochemical),如花青素、類胡蘿蔔 素、類黃酮或酚類化合物 (polyphenols),具有很強的抗氧化能力,可以降低慢性 疾病的發生機率,例如心血管疾病、糖尿病、癌症等。甘薯葉富含花青素及多酚 類,可被廣泛利用及開發成各種商品。
本研究主要以三個品種的甘薯葉為原材料,針對其冷凍乾燥或 40 ℃ 乾燥處 理後之 70 % 乙醇萃取物的抗氧化特性加以探討。所用甘薯葉品種包括亞蔬 CN 1927-16、台農 64 號及 CYY 98-59。抗氧化功能測定包括螯合亞鐵離子、清除 DPPH 自由基與總抗氧化能力。之後再以小鼠肝臟上皮 FL83B 細胞進行細胞毒 性 (cell viability) 測試,以確定其安全使用劑量。同時,利用 TNF-α 誘導小鼠肝 臟上皮細胞產生胰島素阻抗模式,並以不同品種之甘薯葉萃取物處理前述細胞;
評估其對提升葡萄糖攝入能力及改善醣類代謝之效果。
研究結果顯示,總酚含量以台農 64 號 (380.59 mg GAE/g DW)以及 CYY 98-59 (603.09 mg GAE/g DW) 兩個品種的凍乾處理組較高。而在總黃酮類和總花 青素類的結果也和總酚含量的測定相近,分別以台農 64 號 (0.30 g 總黃酮類/g DW;0.25 g 總花青素/g DW)以及 CYY 98-59 (0.22 g 總黃酮類/g DW;0.18 g 總 花青素/g DW) 兩個品種的凍乾處理組較高。在抗氧化特性部份,具有螯合亞鐵 離子能力效果最佳的為 CYY 98-59 的 40 ℃ 乾燥處理後之 70 % 乙醇萃取物 (EC 50值為 3.98 mg/ml);而台農 64 號以及 CYY 98-59 兩個品種對 DPPH 自由基 具有較強的清除活性,EC 50值分別為 4.02 mg/ml 以及 4.11 mg/ml。在總抗氧化 能力測定的方面,也以台農 64 號以及 CYY 98-59 兩個品種的凍乾處理組最佳,
EC 50值為 7.64 mg/ml 以及 8.72 mg/ml。
由細胞試驗結果發現,三種品種甘薯葉之 70 % 乙醇萃取物在濃度 1 mg/ml 之內對小鼠肝臟上皮細胞均無毒性。胰島素阻抗細胞模式之葡萄糖攝入能力試驗,
結果顯示,處理濃度 1 mg/ml 時 ,台農 64 之 40℃乾燥處理組效果最好,與控
制組 (TNF-α 處理) 比較可增加 57.72 % 的葡萄糖攝入能力;其次為 CYY 98-59 之凍乾處理組及亞蔬 1927-16 之凍乾處理組,分別提升了 46.25 % 和 43.84 %。
之後探討不同品種之甘薯葉萃取物對 TNF-α 誘導小鼠肝臟上皮細胞之胰島素訊 息傳遞蛋白質及葡萄糖轉運蛋白表現量的影響。結果顯示,FL83B 細胞經 20 ng/ml TNF-α 誘導後其 IR、IRS-1 及 GLUT-2 之表現量與正控制組比較之下量 並無顯著差異,其明確機轉有待探討。
綜合上述結果,甘薯葉具有抗氧化力將可提供保健食品原料業者之新選擇,
並開發出符合市場需求並具保健功效之新產品,藉以提升甘薯葉加工利用多元性,
增加甘薯葉之附加價值,提高農民的收益。
關鍵字:花青素,甘薯葉,乙醇萃取物,抗氧化活性,葡萄糖攝入
Abstract
Sweet potato leaves have been consumed as a fresh vegetable in many parts of the world. They are rich in vitamin B complex, β-carotene, iron, calcium, zinc, and protein.
Recent experiments revealed that sweet potato leaves contain high contents of polyphenolics, namely anthocyanins and phenolic acids, compared with commercial vegetables.
This study was aimed to evaluate the antioxidation activities and alleviation of insulin resistance of 70 % ethanol extract of leaves of 3 sweet potato cultivars, TN 64, CYY 98-59 and CN1927. Sweet potato leaves were separated into 2 groups, lyophilized and 40 ℃ air-dried. Afterthat, they were extracted with 70 % ethanol at 25 oC. The antioxidation activities tests include total phenolics assay, α,α-diphenyl- β-picrylhydrazyl (DPPH) scavenging ability, ferrous ion chelating ability and scavenging ability on long-life radical anion ABTS˙+ were performed. The in vitro anti-hyperglycemic activity of the extracts was investigated using glucose uptake test in FL83B mouse hepatocytes. The fluorescent dye 2-(N-(7-nitrobenz-2-oxa-1, 3-diazol-4-yl) amino)-2-deoxyglucose was used to estimate the uptake ability of the cells.
The total phenolics of the extracts was determined spectrophotometrically according to the Folin Ciocalteau procedure and ranged from 198.49 to 603.09 mg GAE/g DW. For total flavonoids and total anthocyanins, the values were in the descending order: 0.30 > 0.22 > 0.20 > 0.05 > 0.03 > 0.01 g/g DW which were found in TN 64 > CYY 98-59 > CN 1927-16, respectively. With regard to the results of antioxidation activities of ethanol extracts from leaves of different sweet potato cultivars, the antioxidation activities of lyophilized of 70 % ethanol extract CYY 98-59 and TN 64 were significantly higher than Trolox in ABTS˙+ radical-scavenging activity
(p < 0.05), EC 50 values were 8.72 mg/ml and 7.64 mg/ml respectively. The evaluation of ability to chelate ferrous ion showed that 40 ℃ air-drying of 70 % ethanol extract of CYY 98-59 had lower EC 50 value (3.98 mg/ml), on the other hand, lyophilized of 70 % ethanol extract CYY 98-59 and TN 64 had higher scavenging activity on DPPH free radical with EC 50 values at 4.11 mg/ml and 4.02 mg/ml respectively.
The cell viability test showed that the ethanol extracts from leaves of different sweet potato cultivars have no cell toxicity in concentration of 1 mg/ml. Results of the glucose uptake test in the insulin resistance cell model indicated that the highest improvement can be achieved by the 40 ℃ air-dried of ethanol extract TN 64 in PBS buffer. Compared with the control group (TNF-α treated group), the ethanol extract could increase the glucose uptake ability by 57.72 %. Next is the lyophilized 70 % ethanol extract of CYY 98-59 and CN 1927-16, the percentage increase of the glucose uptake were 46.25 % and 43.84 % respectively. However, the results did not show that ethanol extracts from leaves of different sweet potato cultivars promote the expressions of insulin receptor substrate-1 (IRS-1), insulin receptor and glucose transporter-2 (GLUT-2).
In conclusion, 70 % ethanol extract from sweet potato leaves has potential on physiologic effects as antioxidation and anti-hyperglycemic agents.
Keywords: Sweet potato leaves, phenolic acids, anthocyanins, antioxidation activities, glucose uptake test
目 錄
中文摘要 ... i
Abstract ... iii
目 錄 ... v
圖 次 ... viii
表 次 ... ix
第一章、前言 ... 1
第 二 章 、 文 獻 整 理 ... 3
一、 自由基與健康之關聯性 ... 3
(一) 自由基與活性氧的產生及作用機制 ... 3
(二) 自由基與氧化壓力 ... 4
(三) 過氧化對人體造成的傷害 ... 5
(四) 抗氧化劑 ... 6
二、 糖尿病 ... 9
(一) 糖尿病流行病學 ... 9
(二) 糖尿病簡介 ... 10
(三) 糖尿病分類 ... 11
(四) 糖尿病藥物治療 ... 14
三、 胰島素 ... 17
(一) 胰島素信號傳導 ... 17
(二) 葡萄糖代謝與胰島素阻抗之關係 ... 19
(三) 胰島素阻抗 ... 20
(四) 胰島素分泌缺失 ... 21
(五) 腫瘤壞死因子 tumor necrosis factor-α (TNF-α) 與胰島素阻抗 ... 22
(六) 以細胞模式探討胰島素阻抗 ... 23
四、 甘薯 ... 26
第三章、 研究動機與實驗架構 ... 30
第四章、 不同品種甘薯葉乙醇萃取物抗氧化特性之研究 ... 32
一、 前言 ... 32
二、 實驗材料 ... 33
(一) 原料 ... 33
(二) 試藥級藥品 ... 33
(三) 儀器設備 ... 33
三、 實驗方法 ... 36
(一) 樣品 70 % 乙醇粗萃物製取 ... 36
(二) 總酚化合物含量測定 ... 36
(三) 總類黃酮類測定 ... 36
(四) 總花青素類測定 ... 36
(五) 甘薯葉乙醇粗萃物之抗氧化能力探討 ... 37
(六) 統計分析 ... 38
四、 結果與討論 ... 39
(一) 不同品種甘薯葉之乙醇萃取物之萃取率 ... 39
(二) 總酚化合物含量測定 ... 39
(三) 總黃酮類測定 ... 40
(四) 總花青素類測定 ... 40
(五) 甘薯葉乙醇粗萃物之抗氧化能力探討 ... 41
五、 結論 ... 52
第五章、 不同品種甘薯葉乙醇萃取物對葡萄糖攝入能力之探討 ... 54
一、 前言 ... 54
二、 實驗材料 ... 55
(一) 原料 ... 55
(二) 實驗細胞 ... 55
(三) 試藥級藥品 ... 55
(四) 實驗樣品配製 ... 56
(五) 儀器設備 ... 58
三、 實驗方法 ... 59
(一) 樣品 70 % 乙醇粗萃物製取 ... 59
(二) 小鼠肝細胞 FL83B 之保存與培養 ... 59
(三) FL83B 細胞存活率試驗 (MTT assay) ... 59
(四) 甘薯葉乙醇萃物對具胰島素阻抗性 FL83B 細胞之葡萄糖攝入試驗 (Glucose uptake) ... 59
(五) 西方轉印分析 (Western blot)... 60
(六) 統計分析 ... 62
四、 結果與討論 ... 63
(一) 不同品種甘薯葉之乙醇萃取物對 FL83B 細胞生長存活率 (cell viability)63 (二) 甘薯葉乙醇萃物對具胰島素阻抗 FL83B 細胞葡萄糖攝入試驗 (glucose uptake) ... 64
(三) 不同品種甘薯葉乙醇萃取物對胰島素阻抗 FL83B 細胞胰島素訊息傳遞 蛋白質表現之影響 ... 65
五、 結論 ... 75
第六章、 總結 ... 76
第七章、 參考文獻 ... 77
圖 次
圖 2-1. 101 年主要死因死亡率。 ... 10
圖 2-2. 肝臟中胰島素代謝作用之調節。 ... 18
圖 2-3. 氧化壓力活化絲胺酸激酶誘導胰島素阻抗。 ... 21
圖 2-4. 腫瘤壞死因子與胰島素阻抗。 ... 23
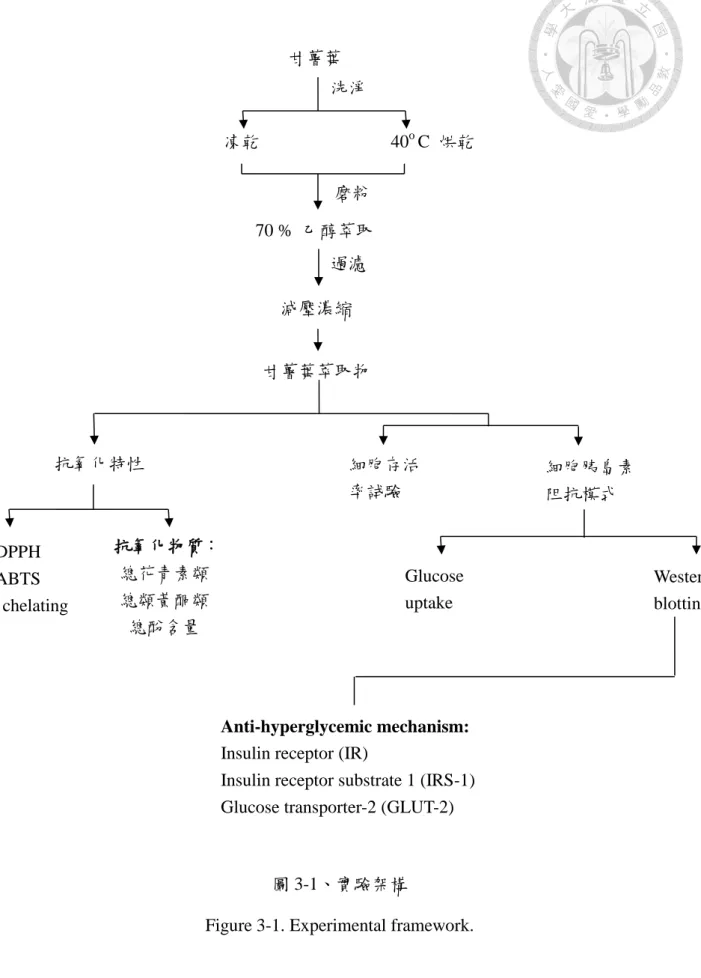
圖 3-1. 實驗架構。 ... 31
圖 4-1. 本實驗所分析的 3 個甘薯品種,(a)亞蔬 CN 1927-16 (黃葉種) ,(b) 台農 TN 64 (紅紫色心藏形大葉) 及 (c) CYY 98-59 (缺刻形紫葉) 。。 ... 35
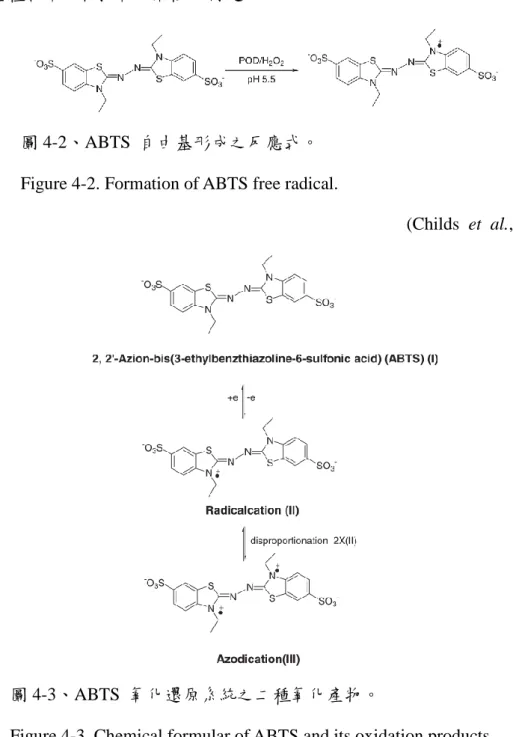
圖 4-2. ABTS 自由基形成之反應式。 ... 43
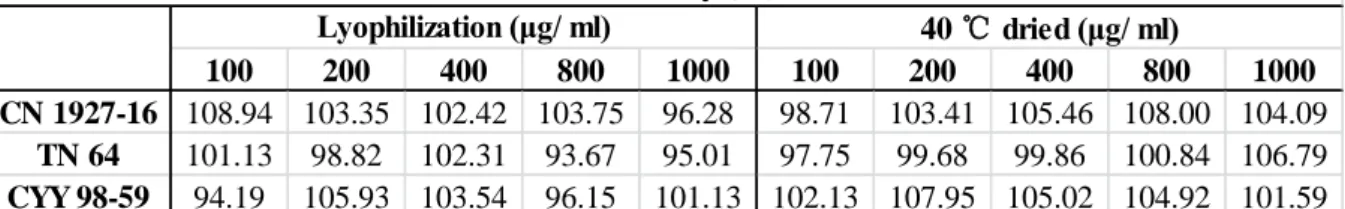
圖 4-3. ABTS 氧化還原系統之二種氧化產物。 ... 43
圖 4-4. 不同品種甘薯葉乙醇萃取物可能參與的抗氧化機制。 ... 53
圖 5-1. 不同品種冷凍乾燥處理甘薯葉之乙醇萃取物 (1 mg/ml) 對具胰島素阻抗 FL83B 細胞葡萄糖攝入之影響。 ... 70
圖 5-2. 不同品種 40 ℃ 乾燥處理甘薯葉之乙醇萃取物 (1 mg/ml) 對具胰島素阻 抗 FL83B 細胞葡萄糖攝入之影響。 ... 71
圖 5-3. 不同品種甘薯葉之乙醇萃取物 (1 mg/ml) 對具胰島素阻抗 FL83B 細胞 胰島素受器表現之影響。 ... 72
圖 5-4. 不同品種甘薯葉之乙醇萃取物 (1 mg/ml) 對具胰島素阻抗 FL83B 細胞 胰島素受器受質蛋白質表現量之影響。 ... 73
圖 5-5. 不同品種甘薯葉之乙醇萃取物 (1 mg/ml) 對具胰島素阻抗 FL83B 細胞 葡萄糖轉運蛋白質表現量之影響。 ... 74
表 次
表 2-1. 糖尿病的分類 ... 12
表 2-2. 糖尿病診斷標準 ... 13
表 4-1. 不同品種甘薯葉萃取率 ... 45
表 4-2. 不同品種甘薯葉之總酚類含量 ... 46
表 4-3. 不同品種甘薯葉之總黃酮類含量 ... 47
表 4-4. 不同品種甘薯葉之總花青素類含量 ... 48
表 4-5. 不同品種甘薯葉乙醇萃取物 DPPH 自由基清除能力之 EC 50 值 ... 49
表 4-6. 不同品種甘薯葉乙醇萃取物 ABTS 自由基清除能力之 EC 50 值 ... 50
表 4-7. 不同品種甘薯葉乙醇萃取物螯合亞鐵離子能力之 EC 50 值 ... 51
表 5-1. 不同品種甘薯葉乙醇萃取對 FL83B 細胞之細胞存活率... 68
表 5-2. 不同品種甘薯葉乙醇萃取物 (1 mg/ml) 對具胰島素阻抗性 FL83B 細胞 葡萄糖攝入之改善情形 ... 69
第一章、前言
甘薯 (Ipomoea batatas(L.) Lam., sweet potato) 又名地瓜,適合在台灣氣候條件 下生產,不受季節限制。地瓜葉在許多報告中皆被提及其營養價值很高;不但維 生素 A、B1 與 C 含量高,也含有豐富的蛋白質與礦物質 (鈣、磷、鐵) ,除此 之外;其所含的膳食纖維柔細,可促進胃腸蠕動,幫助消化,預防便秘、痔瘡及 大腸癌之罹病率;又能增加飽食感,具有減少熱量攝取的間接優點,有助於糖尿 病患者之血糖控制,多食降低膽固醇,有益人體之健康。
除此之外,根據亞洲蔬菜研究發展中心的分析報告,甘薯葉的抗氧化活性為 蔬菜類的第一名,其次分別為薑、莧菜、菠菜、茄子、青江菜、白菜類、花椰菜、
苦瓜、蕃茄等 (http://avrdc.org/)。甘薯葉具有高含量的類黃酮及胡蘿蔔素,是抗 氧化能力重要來源,可抑制過氧化的連鎖反應,其抗氧化能力和熟知的維生素 E 或維生素 C 互補。甘薯葉尚含有綠原酸,是一種重要的生物活性物質,具有抗菌、
抗病毒、增高白血球、保肝利膽、抗腫瘤、降血壓、降血脂、清除自由基和興奮 中樞神經系統等作用。甘薯葉含有多種花青素,其中矢車菊素 (cyanidin),不但 較穩定,抗突變和抗氧化力還遠超過其他花青素。花青素可預防糖尿病引起的視 網膜病和胸部纖維囊腫病、保護視力、抗腫瘤、保護幅射傷害、保護血管、抗發 炎、降低化療毒性、保護肝臟於四氯化碳傷害及保護抗氧化酶 (SOD、glutathione peroxidase、catalase)的功能。
甘薯葉病蟲害少,高營養價值,以其為原料製作的保健食品可用於血糖、血 脂、體脂肪的控制,具有保健功效,長期使用可以調控新陳代謝症後群,減少慢 性疾病的發生。
第二型糖尿病也稱非胰島素依賴型糖尿病,常出現在成年人 (特別是肥胖症 患者) 身上,其病症會導致消瘦。病因包括胰島素抗性 (insulin resistance),使到 身體不能有效地使用胰島素,或是胰島素分泌的減少,無法滿足身體所需。
糖尿病主要的臨床表現為多飲、多尿、多食和體重下降 (三多一少) ,以及
血糖高、尿液中含有葡萄糖 (正常的尿液中不應含有葡萄糖) 等。早期第二型糖 尿病患者可以通過改善生活方式 (如健康飲食,適量運動等) 來控制糖尿病。大 多數患者可通過口服降血糖藥來幫助控制體內血糖。而一些第二型糖尿病患者需 要胰島素注射。糖尿病會引發多種慢性疾病產生,如高血壓、神經病變、全身動 脈硬化 (Shen et al., 2008) 等癥狀,故為避免此現象發生,有效調節血糖或延緩糖 尿病的發生即顯得格外重要。
本研究主要針對其冷凍乾燥或 40 ℃ 乾燥處理後之 70 % 乙醇萃取物的抗 氧化特性加以探討。所用甘薯葉品種包括亞蔬 CN 1927-16、台農 64 號及 CYY 98-59。抗氧化功能測定包括螯合亞鐵離子、清除 DPPH 自由基與總抗氧化能力。
之後利用 TNF-α 誘導小鼠肝臟上皮細胞產生胰島素阻抗模式,並以不同品種之甘 薯葉處理前述細胞;評估其對提升葡萄糖攝入能力及改善醣類代謝之效果。
第 二 章 、 文 獻 整 理 一、自由基與健康之關聯性
由於人體疾病與老化的發生,近年來醫學界開始重視自由基 (free radical) 與 抗氧化物質的研究。人體內無時無刻都在產生自由基,在呼吸代謝過程中約有 2
%~3 % 氧會轉換成超氧自由基的活性氧 (superoxide),尤其到了中年後,身體機 能、免疫力及對於自由基的清除能力隨著年齡增長而逐漸降低,容易導致各種疾 病產生。若經由適當服用抗氧化劑可將油脂在氧化過程產生的自由基清除,藉以 中斷油脂氧化的連鎖反應,阻止氧化反應的進行 (Kaur and Kapoor, 2002;
Halliwell, 2009);或藉由抗氧化劑對金屬離子螯合能力 (chelating) 來抑制金屬離 子促氧化的功能,而得以彌補身體抵抗氧化傷害之不足、達到抗氧化之目的,更 可延緩老化而增加壽命 (Ames et al., 1993; Sang et al., 2002) 。
(一)自由基與活性氧的產生及作用機制
自由基是指含有一個或多個不成對電子而獨立存在之原子或分子 (Halliwell, 1989),不同型態的自由基其化學反應性差異相當大,大部分的自由基是處於較不 穩定狀態且半衰期短,其反應性較非自由基分子高,易與其他分子發生反應,使 自己形成穩定電子對狀態,而生成自由基,並進一步發生連鎖反應 (廖,2004) 。 氧是生物體維持生命所必須的基本要素,在大氣中是以安定的三重態氧 (triplet state oxygen; 3O2) 存在。而氧經由電子及能量轉移等作用,生成反應性較三重態 氧分子強的含氧分子稱為活性氧 (activated oxygens),活性氧中亦有一些自由基,
這些含氧之自由基亦稱含氧自由基 (oxyradicals),故活性氧可分為 (1) 含氧自由 基,如超氧陰離子、氫氧自由基、過氧化自由基、烷氧自由基及過氧化氫自由基 等;(2) 非自由基的活性氧,如過氧化氫、次氯酸、臭氧、單重氧及次溴酸。當 體內的自由基與活性氧過量時則易與細胞膜上之多元不飽和脂肪酸作用而引發脂 質過氧化 (lipid peroxidation)的作用,造成細胞膜的傷害及擾亂 (陳,2004) 。
活性氧族具有很高的反應性、半衰期很短且極不穩定,可能攻擊生物分子而 引發自由基連續反應。在生理所需的濃度下,活性氧族可行使正常細胞功能;但 若活性氧族過度產生,則可能造成細胞中 DNA、脂質、蛋白質等分子過度氧化,
而造成細胞傷害 (Wei, 1998)。相關之研究證據顯示,活性氧族可進一步造成多種 退化性疾病、老化、突變以及癌症的形成。而在所有好氧生物體 (aerobic organisms)
內,生物體內約有 95 %的氧氣應用在粒線體內膜的呼吸鏈 (Cadenas and Davies, 2000),在產生 ATP 的過程約有 1-2 % 的氧會轉變成超氧陰離子 (superoxide anion, O2
-)、過氧化氫 (H2O2)、及其他活性氧自由基 (Cadenas and Davies, 2000)。
活性氧族會在氧化磷酸化中不斷的形成及分解。而體內亦有防禦與清除自由基的 系統來抵禦自由基的攻擊,並具有修補系統來修復受到自由基攻擊的組織 (Nordberg and Arner, 2001)。
正常細胞中,活性氧族可調節訊息傳遞與生理狀態 (Fleury et al., 2002),過多 的活性氧族則藉由清除系統來清除。主要透過體內抗自由基氧化系統如 Mn- superoxide dismutase (Mn-SOD)、Zn/Cu-SOD、catalase、glutathione peroxidase等酵 素可清除自由基。但若抗自由基氧化系統與自由基的產生有失衡現象,便會造成 氧化壓力 (Wei, 1998)。
(二)自由基與氧化壓力
在生物體中,抗氧化防禦機制可使活性氧形成的量保持在最少 (MeKee, 2003)。
自由基可能涉及人體多種生物功能,例如調控細胞生長及訊息傳遞,但是當自由 基的生成超出人體本身的防禦範圍時,就會有氧化壓力 (oxidative stress)的形成,
氧化壓力是指體內抗氧化系統與氧化傷害失去平衡而造成生物體受損的現象,主 要是因酵素失活,多醣類去聚合作用、DNA 裂解和分子變性而導致損害,在某 些情況下,例如某些代謝異常、過度使用某些藥品、曝露在強烈的輻射或反覆的 接觸如二手煙等環境汙染物,皆會導致嚴重的氧化傷害。
氧化壓力可能造成細胞內訊息傳遞混亂、細胞膜損害、細胞離子溝通損傷、
引發細胞膜脂質過氧化 (Halliwell and Chirico, 1993)、影響細胞內所有含SH基的 分子 (sulfhydryl-containing molecules),包含蛋白質、DNA等皆會產生影響。最近 的研究證據顯示活性氧族作為次級訊息傳遞物 (second messengers),活化轉錄因 數 (transcription factor), 如 AP-1 及 NF-κB 與誘導免疫發炎相關基因表現 (Clement and Pervaiz, 1999; Desagher and Martinou, 2000)。除此之外,不同細胞種 類,活性氧族可作為 mitotic stimuli、cellular senescence inducer 與 cell death mediator (Cadenas and Davies, 2000; Finkel and Holbrook, 2000; Nordberg and Arner, 2001)。
氧化傷害至少與 100 種人類疾病有關,包括癌症、循環系統失調,例如動脈 硬化症、心肌梗塞、高血壓和神經失調,例如肌萎縮硬化症、柏金森氏症及阿茲 海默症等。
(三)過氧化對人體造成的傷害
在正常的情況下,細胞組成的抗氧化防禦機制會將氧化傷害減至最小。在氧 分子的還原過程中,第一個形成的活性氧是 O2,大部分的 O- 2-都是由 complex Ⅲ 的 Q cycle 或黃素蛋白 NADH 去氫酶 (complex I) 而來的電子所形成的。O2- 扮 演親核物質 (nucleophile),常是氧化劑,但在特殊條件下,也有可能是還原劑,
因為它的溶解特性,O2-會導致細胞膜磷脂質相當大地傷害。當O2-在水溶液的環 境中產生,O2-會與它本身反應產生 O2 和 H2O2。
2 H++ 2 O2-
→ O2 + H2O2
H2O2 不是自由基,因為它沒有未成對的電子,但其有限的反應性會因它通過 細胞膜散佈更廣而增強。H2O2 隨後與 Fe2+ (或其他轉移金屬)反應產生一個具高 度反應性的物質---羥自由基 (hydroxyl radical, .OH)。
Fe2+ + H2O2 →Fe3++.OH + OH-
羥自由基是具高度反應性的分子,且是特別危險的自由基,在其與任何生物 分子碰撞反應前,只會短距離的散佈,一旦開始反應,其可啟動自體催化自由基 鏈反應。當雙氧吸收了充足的能量去轉移一未成對電子至較高軌域,超氧陰離子 (O2-
)就會形成單重態氧 (1O2):
2 O2-
+ 2H+ →H2O2 + 1O2
單重態氧雖然不是自由基,但因為它是強力氧化劑,甚至比氫氧自由基更具 反應性。
1. 破壞 DNA
氫氧自由基 (OH·) 會破壞 DNA 的每個部份,造成 DNA 雙股的切斷或鹼基 結構,導致 DNA 永久性的損傷 (Cameron et al., 1976; Valko et al., 2004) 。DNA 遭到切斷,人體在修補的過程中,可能會因為無法正常修復而產生突變。自由基 攻擊鹼基後會衍生一些副產品,這些副產品會使遺傳發生錯誤而有致癌的可能。
人體若長期遭受這一連串的傷害,所造成的結果就是各種慢性疾病發生和老化。
2. 氧化脂質
體內的多元不飽和脂肪酸受到氧化後會產生有毒的醛類 (malondialdehyde, MDA),MDA 會引起突變及致癌的危險 (Carr et al., 2000)。細胞膜上脂質被氧化,
會使細胞膜流通性改變,養分無法進入細胞內造成細胞壞死,若細胞膜被破壞的 速度大於細胞再生的速度,組織器官的功能就會受到影響而產生老化。低密度脂 蛋白 (LDL) 被氧化,會增加血小板的凝集、內皮細胞的脫落,而造成動脈粥狀 硬化。
3. 攻擊蛋白質
自由基攻擊胺基酸的側鏈,令蛋白質失去功能,或形成大分子,甚而斷裂,
使細胞無法正常運作 (Stadtman et al., 2004)。發生在皮下之膠原組織,會令皮膚 失去彈性、筋骨僵硬等。
(四)抗氧化劑
抗氧化劑具有抑制低密度脂蛋白氧化效果,故攝取含抗氧化的物質可預防心 血管疾病的發生 (Salleh et al., 2002; Zin et al., 2002) 。因此,抗氧化劑的應用引 起世界廣泛關注,更引起一連串使用抗氧化劑作為養生保健食品及化妝品原料的 潮流,促使「抗氧化物質」在目前各國的保健食品中有極大的市場佔有率。
近年來由於食品工業的發達,食品添加物之種類與數量逐年增加,為了消費 者的健康,從自然界中尋求安全又有效的天然抗氧化劑乃成為食品界之一項重要 研究。天然的抗氧化劑已被大量使用在食品工業上,其安全性高,但抗氧化性沒 有人工合成的抗氧化劑有效且價格昂貴,因此尋找便宜又有效的天然抗氧化劑已 成為重要的課題。
自然界中天然抗氧化物質一般可分為:
1. 酵素類
a. 超氧化物歧化酶 (superoxide dismustase)
SOD 是人體內最重要的抗氧化酵素系統,將活性較大的超氧化物 (superoxide, O2
-) 作用成活性較小的過氧化氫 (H2O2),反應式為:2O2
- + 2H+→ H2O2 + O2,而 SOD 需要與金屬離子結合,如:Cu2+、Zn2+ 或 Mn2+,當作電子的傳 遞者,才有作用,SOD 在體內經氧化還原反應後可循環再利用 (McCord et al., 1969; Valko et al., 2007)
b. 過氧化氫酶 (Catalase)
Catalase 主要是將 SOD 產生的 H2O2 作用成無害的氧及水,反應式為:
2H2O2→ O2+2H2O,但 catalase 需要 Fe3+ 來催化整個反應的進行。
c. 麩胺基硫過氧化酶 (glutathione peroxidase)
主要是活化 glutathione (GSH),直接將過氧化脂質 R-OOH 還原成醇 R-OH 或分解 H2O2,但 glutathione peroxidase 也需要硒的催化 (Pastore et al., 2003)。
2. 非酵素類 a. 維生素 C
維生素 C 是水溶性維生素,可利用分子上的氫氧基 (OH) 和自由基作用,OH 會釋出一個 H+和一個電子,自由基得到電子後,成為較穩定的的化合物,而 維生素 C 上的 OH 則變成 O-,但由於其可產生共振,所以活校較低、較無 破壞性。維生素 C 被氧化後會被 glutathione peroxidase 系統還原,而可以一 再循環重複利用 (Cameron et al., 1976; Sies et al., 1992; Carr et al., 1999) 。 b. 維生素 E
維生素 E 是其中一個脂溶性的維生素,有酚的結構,可利用其上的 OH 基釋 出一個 H+ 和一個電子,使自由基變成較穩定的化合物,維生素 E 上的 OH 則變成 O-,因旁有苯環可以產生共振,成為較穩定的自由基。作用後的氧化 態維生素 E,會被維生素 C 還原,因此可重複再利用 (Sies et al., 1992) 。 c. β 胡蘿蔔素
β 胡蘿蔔素是維生素 A 的前驅物質,不過維生素 A 並不像 β 胡蘿蔔素具有 良好的抗氧化作用。β 胡蘿蔔素在人體內有二種抗氧化能力:一、可以與脂質 過氧化自由基結合而中斷脂質過氧化連鎖反應。二、吸收氧氣因為光線照射 (例如在眼睛) 而變成的激發氧氣的過多能量,阻止氧化作用的進行。β 胡蘿蔔 素的主要食物來源是深綠、黃色的蔬菜和藻類,其中以紅蘿蔔最具代表性 (Burton et al., 1984) 。
d. 其他植物化學物 (phytochemicals) 中的多酚類、帖類、類黃酮類及有機酸等成 分
花青素屬於多酚類化合物中的類黃酮代謝所衍生的化合物,常見者包括天竺葵 色素 (pelargonidin)、矢車菊素 (cyanidin)、花翠素 (delphinidin)、芍藥花素 (peonidin)、矮牽牛素 (petunidin) 及錦葵色素 (malvidin) 等 (Albert et al., 2009;
詹, 2010) 。花青素在植物界中是很大一群以 C6-C3-C6 芳香環為結構骨架之 水溶性色素,主要累積在植物細胞的液泡,常存在於葉片、花瓣、種子和果實 中,隨液胞中酸鹼度的不同而使其表現出不同的顏色,花青素為植物各種顏色 (紅、紫、紫紅) 之主要來源,其結構與抗氧化力有相當大的關係,一般與醣類 形成醣苷 (glycosylated) 後也具有抗氧化活性。最常見的花青素是在 A、C 環 C-3或 C-3, 5 之位置發生醣苷鍵結 (Francis, 1989),而矢車菊色素之抗氧化性 來自 B 環的 3, 4- dihydroxybenzoic acid 氧化成 3,4- diketobenzoic acid (楊,
1994) 。花青素的醣化受到不同醣取代基的影響,且捕捉 OH 的能力與黃酮 類類似,會因 B 環OH 基含量的增多而增加其捕捉自由基的能力,及因 C 環 結構 C-4 位置甲基化或 C-3 位置醣化作用,而減弱其抗氧化活性 (Zhan, 1996;鍾,2006)。
目前有許多研究集中於植物抗氧化潛能的開發,發現抗氧化能力與植物內 多酚類含量多寡成正相關 (Yoshinaga et al., 2000; Malkeet and Picha , 2008),也 有相關報告 (Tsuda et al., 1996; Goda et al., 1997; Huang et al., 2004; Yin et al., 2004; 高等, 2005; Saigusa et al., 2005; Huang et al., 2006; Wang et al., 2006;
Teow et al., 2007) 指出花青素在人體內的抗氧化及清除自由基的能力為維他 命 E 的 50 倍、維他命 C 的 20 倍。其抗氧化能力可維持正常的細胞連結、
穩定內皮細胞上的磷脂質,避免動脈與靜脈的細胞被自由基破壞。亦可增加膠 質及黏多糖的合成及預防聚集物依附在血小板表面產生過度凝集現象,以維持 動脈壁的完整性及避免膽固醇被氧化,增強血管彈性,增加循環,預防心血管 疾病、延緩細胞老化、減緩糖尿病、改善視力及抗癌等功能。因此,花青素相 關產品受到食品工業青睞,目前最常見之花青素商品有藍莓萃取物及葡萄萃取 物等 (Lu et al., 2001; Saijusa et al., 2005) 。黃 (2005) 研究指出,紫色甘薯葉 含豐富的 quercetin 和 cyanidin,並且可有效避免淋巴球細胞 DNA 氧化損傷,
所以紫色甘薯葉為良好抗氧化材料,故選為本研究的材料其中一項來源。
二、糖尿病
(一)糖尿病流行病學
隨著醫藥的發達,傳染性疾病的控制,世人生活型態的靜態化,肥胖之高度 盛行,糖尿病已成為二十一世紀全世界最重要的公共衛生課題之一。在開發中以 及新興工業化國家,它的發生率 (incidence) 及盛行率 (prevalence) 正在快速的增 加中。預測至 2025 年世界衛生組織有三億三千萬名病患,其中絕大多數為第 2 型糖尿病。用以治療糖尿病及其併發症的社會和經濟成本也越來越高,可能會使 國家的健康照護預算陷入潛藏性的癱瘓。目前,根據衛生署統計 (圖2-1) ,台灣 2012 年十大主要死因中,原排名第 4 的糖尿病與排名第 5 的肺炎對調,死於糖 尿病的人口達 9281 人,平均每天有 25 人死於糖尿病。雖然這個快速的增加可 能和疾病本身的增加以及和國內的 ICD 編碼方式造成的高估都有關係,不可忽 視的是糖尿病所造成的社會負擔相當可觀。以美國為例,1992 年有 700 萬名診 斷確立的糖尿病人,他們雖然只佔總人口的 2.8 %,但卻耗掉所有保健經費的 5.8
%。另外,估計當年花在糖尿病的總費用約為 920 億美元,其中直接的醫藥支出 為 452 億美元,而間接的費用則為 466 億美元。再以加拿大為例,1998 年一年 間糖尿病及其併發症的花費分別為 48 % 和 52 %;其中,已確診糖尿病的直接 花費約為 57 億,和糖尿病相關的併發症以及心臟血管疾病的花費反而高達 64 億。國內的研究顯示糖尿病患者的洗腎花費 (每年美金兩萬七千元) 較之非糖尿 病患的花費高 12 %。主要是由於有糖尿病的洗腎病人的住院花費較高。
圖2-1、101年主要死因死亡率 (行政院衛生署,2013) Figure 2-1. The top 10 causes of death in 2012.
(二)糖尿病簡介
糖尿病是一種因體內胰島素絕對或者相對不足所導致的一系列倂發症,與遺 傳基因有著非常密切的關聯。糖尿病的主要臨床表現為多飲、多尿、多食和體重 下降 (三多一少) ,以及血糖高、尿液中含有葡萄糖 (正常的尿液中不應含有葡萄 糖) 等。正常人進食後,食物在腸胃道被消化分解後,產生葡萄糖,葡萄糖經由 血液運送至全身,會刺激胰臟釋出胰島素,胰島素可幫助葡萄糖進入身體各組織 細胞,轉變成能量,或貯存在肝、肌肉以及脂肪細胞中。如果沒有胰島素,或者 身體各組織對胰島素不發生反應,則葡萄糖就不能被利用,於是血液中葡萄糖含 量便會上升。當血糖升高到某一程度,超過腎臟所能回收的極限時,葡萄糖便會 從尿液中 "漏" 出,所以稱之為糖尿病。其實血糖超過訂定的標準便算是糖尿病,
不一定要有尿糖存在才算糖尿病,由於血糖要高到某程度才會有尿糖出現,所以 測定血糖較尿糖準確。因此診斷糖尿病是以測定血糖為主。
胰臟是一個位於胃後下方的橫置扁長型器官,重量不到 1/4 公斤,較大的一 端稱為頭部 (內側) ,較細長的一端稱為尾部室 (外側) 。主要的功能包括外分泌 和內分泌兩部分。胰臟外分泌部分的主要功能是直接將消化酵素分泌到腸道中,
協助消化食物。在胰臟的尾部中有許多細胞聚集而成的小島,稱為胰島,主要的
功能是分泌賀爾蒙到血液中,屬於內分泌系統,也就是與糖尿病有關的部分。在 胰島中有一種細胞叫做貝他細胞,主要分泌的賀爾蒙是胰島素。它可感應血中葡 萄糖濃度的變化,並適時適量地分泌出胰島素到血液中,循環全身,協助血糖進 入身體各細胞,維持血中葡萄糖濃度在正常範圍之內。胰島素的主要功能是幫助 我們的養分進入身體組織細胞,提供細胞正常運作所需要的能量。它的作用就像 一根門匙,打開葡萄糖進入細胞的通道,讓葡萄糖能夠順利進入細胞。當我們身 體不能分泌胰島素時 (如胰島素依賴型糖尿病) ;或體內細胞對胰島素反應不好 時 (如非胰島素依賴型糖尿病) ,除了因葡萄糖不能進入細胞所產生之血糖上升 外,體內的脂肪、蛋白質和碳水化合物的代謝都會發生異常現象。
(三)糖尿病分類
糖尿病的分類包括:第 1 型糖尿病、第 2 型糖尿病、其它型糖尿病及妊娠 型。
表2-1、糖尿病的分類
Table 2-1. Etiologic classification of diabetes mellitus
第 1 型糖尿病 (β 細胞破壞,通常會造成胰島素絕對缺乏) 自體免疫 (autoimmune)
特發性 (idiopathic)
第 2 型糖尿病 (範圍可從胰島素阻抗為主合併胰島素相對缺乏,至以胰 島素缺乏為主合併胰島素阻抗)
其它型糖尿病
β 細胞功能的基因缺陷 胰島素作用的基因缺陷
胰臟外分泌的疾病:胰臟炎、受傷/胰臟切除、腫瘤、囊性纖維化 內分泌疾病:Cushing’s syndrome,acromegaly,pheochromocytoma,
hyperthyroidism 等 藥物或化學試劑所誘發 感染
罕見的免疫性疾病
其它遺傳性症候群相關的糖尿病 妊娠型糖尿病
(American Diabetes, 2012)
目前診斷糖尿病,主要依據血糖質高低來判斷。通用的有世界衛生組織標準 和美國糖尿病學會的標準。以美國糖尿病學會的標準,正常 (無懷孕) 成人空腹 血漿中葡萄糖的濃度 (簡稱血糖) 是 < 115 mg/dl;飯後兩小時血糖是 < 140 mg/dl。
若出現下列的情況,就可診斷為糖尿病:
1. 隨時測得的血糖 (無論空腹與否的所謂隨機血糖) 是 200 mg/dl或以上,同時 合併出現糖尿病典型症狀 (包括多喝、多尿、多吃、體重下降等症狀) 。
2. 空腹靜脈血糖有兩次是 140 mg/dl或以上。
3. 空腹靜脈血糖 < 140mg/dl,但給予口服 75 公克葡萄糖 (葡萄糖耐量試驗) , 2 小時的靜脈血糖是 200 mg/dl或以上,加上試驗過程中有一次血糖是 200 mg/dl 或以上。
表2-2、糖尿病診斷標準
Table 2-2. Criteria for the diagnosis of diabetes A1C ≧ 6.5 %.
The test should be performed in a laboratory using a method that is NGSP certified and standardized to the DCCT assay.*
FPG ≧ 126 mg/dl (7.0 mmol/l).
Fasting is defined as no caloric intake for at least 8 h.*
2 hrs plasma glucose ≧ 200 mg/dl (11.1 mmol/l) during an OGTT.
The test should be performed as described by the World Health Organization, using a glucose load containing the equivalent of 75 g anhydrous glucose dissolved in water.*
In a patient with classic symptoms of hyperglycemia or hyperglycemic crisis, a random plasma glucose ≧ 200 mg/dl (11.1 mmol/l).
* In the absence of unequivocal hyperglycemia, criteria 1–3 should be confirmed by repeat testing.
(American Diabetes, 2012)
至於一些病人給予口服75公克葡萄糖 (葡萄糖耐受試驗)後,2 小時的靜脈血 糖介於 140 至 200 mg/dl間,加上試驗過程中有一次血糖是 200 mg/dl或以上;
就可說是葡萄糖耐受不良。至於婦女在懷孕時第一次發現糖尿病,分娩後又恢復 正常耐糖狀態的,稱作妊娠性糖尿病。近年來,美國糖尿病學會建議,婦女在懷 孕 24-28 週時,做妊娠性糖尿病篩檢。初步檢查不必空腹,在喝過 50 公克葡萄 糖沖成的糖水後一小時驗血,如果血漿糖值超過140 mg/dl,就應該進一步進行
100 公克葡萄糖耐量試驗。此時,孕婦在空腹及喝糖水後每隔一小時抽血一次,
前後共三小時,如果四項血糖值中有二項超過判定標準確 (0、1、2 及 3 小時值 分別是:105, 190, 165, 145 mg/dl),就可診斷為妊娠性糖尿病。妊娠性糖尿病病人 產後六週至三個月內,應該重新驗血 (75 公克葡萄糖耐量試驗),確定自己的耐 糖狀態分類。所以,葡萄糖耐量試驗並不是診斷糖尿病的必要手段,實施的時機,
不外乎:
(1) 空腹或隨機血糖不足以診斷糖尿病
(2) 懷孕期間或分娩後用來確定妊娠性糖尿病診斷 (3) 用來診斷葡萄糖耐受不良。
(四)糖尿病藥物治療
第 2 型糖尿病的治療包括營養、運動、藥物和衛教。營養與運動為基本步驟,
血糖較高者則視病情不同而給予不同的藥物治療;降血糖藥物治療包括口服抗糖 尿病藥與注射胰島素。症狀嚴重者,需要立即使用胰島素,否則可先行試用口服 抗糖尿病藥。口服抗糖尿病藥的使用,需要依據病人的病情,包括:血糖的高低、
肝、腎、心臟功能,藥物的療效以及副作用等因素而定,當然需要同時接受營養 與運動治療。如果未達到治療目標,則可單獨使用一種或同時使用多種口服抗糖 尿病藥,和/或注射胰島素。目前口服抗糖尿病藥依其作用機轉可分為下列五大 類:
1. 雙胍類 (biguanide) :目前最常使用的雙胍類製劑是二甲雙胍 (dimethyl biguanide, metformin) ,主要作用是減少肝臟葡萄糖的輸出和降低空腹血糖,
但不會刺激胰島素的分泌。它不會增加體重。大約 20 % 的病人服用 metformin 後會產生食慾不振、腹部不適、甚至腹瀉等副作用。Metformin雖會影響維生 素 B12 的吸收,但很少會造成貧血 (Bailey et al., 1996)。Metformin 和其它的 口服抗糖尿病藥相較,其不會增加體重,且單獨服用時很少發生低血糖。
2. 促胰島素分泌劑 (insulin secretagogues) :目前此類藥物有二種: (a) 磺醯脲素 (sulfonylureas) :主要作用是與胰臟 β 細胞的磺醯脲素受體結合,刺激胰島素 分泌。磺醯脲素大部份在肝臟代謝,其代謝產物活性大為減弱,和其它未經代 謝的原型藥物一起由腎臟和/或膽道排出。因此肝、腎功能不全的病人,使用磺 醯脲素時會出現磺醯脲素作用增強、時效延長的現象,此時須要特別小心低血 糖的發生。腎功能不全時,應儘量避免使用經由腎臟排泄的磺醯脲素 (例如:
chlorpropamide) ,如果必須使用這類藥物,則應減少其劑量、經常檢測血糖以 減少低血糖的發生。磺醯脲素的不良反應以體重增加和低血糖最為常見;低血 糖可能持續數日,對老年人、腎功能不全或食無定時者更是危險。 (b) 非磺醯 脲素 (meglitinides 或 glinides) :現有 repaglinide 和 nateglinide 二種。主要 的作用機轉是與胰臟β 細胞上磺醯脲素受體內的不同部位結合,刺激胰島素的 分泌;和磺醯脲素相較,其吸收速率較快且作用時間較短,因此需隨餐服用。
3. 葡萄糖苷酶抑制劑 (α-glucosidase inhibitors) :主要作用是在近端小腸抑制澱粉 和雙醣類的分解,進而延緩其吸收。此藥不被腸胃道吸收,可降低餐後血糖和 胰島素濃度,但降血糖的效果不如磺醯脲素和 metformin,單獨使用並不會發 生低血糖。使用本藥後,醣類大都在遠側端腸道吸收,因此會產生脹氣和其它 腸胃道的副作用。葡萄糖苷酶抑制劑可預防葡萄糖失耐的病人發生糖尿病 (Chiasson et al., 2003),但其對心血管疾病的預防則有待更進一步的研究。病人 服用阿爾發葡萄糖苷酶抑制劑如果出現低血糖,治療時必須使用單醣 (例如:
葡萄糖或牛奶) ,而不能使用雙醣或多醣,例如:蔗糖 (因乳糖酶 (lactase) 不 受本藥抑制,但蔗糖酶 (sucrase) 則會受到抑制) 。
4. 胰島素增敏劑 (insulin sensitizers) :現有 pioglitazone 和 rosiglitazone 二種,
作用機轉為活化PPAR-γ (peroxisome proliferative-activated receptor-γ) ,增加肌 肉、脂肪和肝臟對內生性或是外源性胰島素的敏感度,降低空腹血糖和血中胰 島素濃度 (Yki-Järvinen, 2004)。常見的副作用有體重增加、體液滯留、周邊水 腫和心臟衰竭等。過去的研究顯示,胰島素增敏劑相較於磺醯脲素與雙胍類藥 物可維持較長期血糖的穩定效果 (Kahn et al., 2006),對糖尿病前期的病人則可 降低其疾病進展至第 2 型糖尿病的比例 (Yusuf Set al., 2006)。胰島素增敏劑 可和雙胍類、促胰島素分泌劑和胰島素合併使用。但在某些國家如加拿大,則 不建議胰島素和胰島素增敏劑合併使用。
5. 二肽基酶-4 抑制劑 (dipeptidyl peptidase 4 inhibitors, DPP 4 inhibitors) :腸促胰 液素 (incretins) 可調節體內葡萄糖代謝,包括類升糖素肽-1 (glucagon-like peptide-1,GLP-1) 和葡萄糖依賴性胰島素刺激多肽 (glucose-dependent
insulinotropic polypeptide,GIP) 二種。在生理情況下,GLP-1 和 GIP 會快速 的被 DPP 4 水解成不具活性的產物。DPP 4 是位於細胞膜上的蛋白質,能夠 在很多的組織中表現,包括免疫細胞 (Richter et al., 2008)。DPP 4 抑制劑是一
小分子,和其藉由提高活性腸泌素的濃度,可促進胰島素的釋出和抑制升糖素 的分泌,達到降低血糖的作用。
三、胰島素
第 2 型糖尿病屬於多重病因的複雜性疾病,一般認為基因遺傳與外在環境皆 扮演重要角色。其病理生理學上的特色為胰島素分泌缺失 (β 細胞數量和/或功能 受損) 、胰島素利用障礙 (肝臟、肌肉和脂肪組織產生胰島素阻抗) 、腸道分泌的 腸促胰液素 (incretin) 下降或功能異常、胰臟 α 細胞分泌升糖素增加、腎臟增加 糖類再吸收和腦部的胰島素阻抗,而引起以高血糖和其它代謝異常為主要特徵的 疾病 (Defronzo RA, 2009),其核心的病理生理學缺陷是胰島素分泌缺失和胰島素 阻抗。胰島素是一種由胰臟β 細胞所分泌的胜肽,它扮演著促進周邊組織對葡萄 糖吸收和抑制肝臟葡萄糖新生的角色。大多數病人表現出程度不等的周邊胰島素 阻抗和相對的胰島素分泌不足,二者皆可導致第 2型糖尿病 (Taylor, 2008)。這些 缺陷常發生於疾病的早期,甚至在尚未出現高血糖前就已經存在。此外第 2 型糖 尿病經常伴隨著高血壓、三酸甘油酯上升、低密度膽固醇脂蛋白上升和高密度膽 固醇脂蛋白降低,這些都會增加心血管疾病發生的風險 (Ecke et al., 2002)。
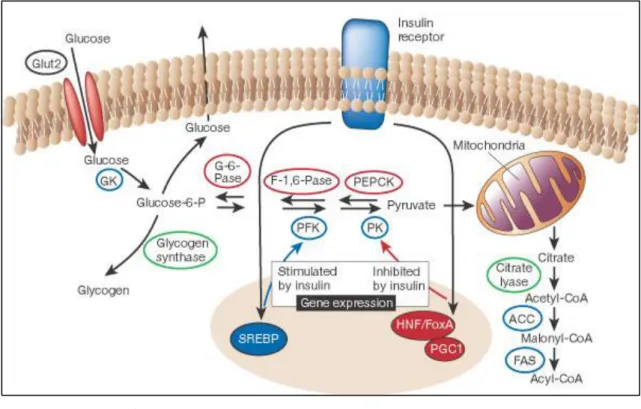
(一)胰島素信號傳導
信號傳導有前段與後段,即上游 (upstream),與下游 (downstream)。胰島素 信號的上游需要胰島素接受器 (insulin receptor)。包括兩個 α次單元與兩個 β次單 元,α次單元接受器在細胞外胰島素先與之結合,透過細胞膜後,再活化細胞內 的 β次單元,其含有 tyrosine kinsae 成分,結合後的複體再進行一連串的分子反 應,即在 tyrosine 分子上面,產生磷酸化反應 (phosphorylation)。在 COOH 端 上的 tyrosine 產生自體磷酸化 (autophosphorylation),促進胰島素對細胞分裂的 作用。胰島素接受器表現基因,在 tyrosine 分子上面產生磷酸化,才能表現其下 游的認證角色。
胰島素與胰島素接受器是胰島素信號傳導的上游,下游則與胰島素傳導器 (insulin transporter) 有關,與胰島素阻抗有關的胰島素傳導器是 4 號,簡稱 GLUT4 (glucose transporter 4),缺乏 GLUT4,則葡萄糖的吸收不良,會產生胰島 素阻抗 (Pessin et al., 2000)。
下游的認證功能之一是透過胰島素刺激下,使 PI3-kinase
(phosphatidylinositol- 3- kinase) 的活化,GLUT4 由細胞內移至細胞膜,再帶著葡 萄糖由細胞內排到細胞外。胰島素阻抗,缺少 PI-3-kinase 活化,葡萄糖耐受性
便下降,就是這個原理。PI3-kinase 作用的下游之一是 protein kinase B (PKB,也 稱 AKT),使胰島素接受器在 serine/threonine 上面產生磷酸化。
胰島素與胰島素接受器之傳導,類似神經細胞之突觸傳導, 前者係由接受器 前傳到接受器後,與突觸前傳到突觸後類似。胰島素阻抗不是單一基因所決定,
牽連很廣,係由多種基因與環境因素,造成肥胖或感染,才產生胰島素阻抗的特 徵。胰島素信號傳導異常,造成葡萄糖的吸收與代謝異常,即葡萄糖耐受性不良。
圖2-2. 肝臟中胰島素代謝作用之調節。
Figure 2-2. The regulation of glucose metabolism in the liver.
(Saltiel and Kahn, 2001)
1. 胰島素受器 (Insulin receptor, IR)
胰島素受器為一種鑲嵌於細胞膜上的醣蛋白大分子,具有 tyrosine kinase 活 性,包括 insulin-like growth factor (IGF) 與 insulin receptor-related receptor (IRR) (Patti and Khan, 1998)。胰島素受器為受體酪胺酸激酶 (receptor tyrosine kinases) 中的一個次家族 (subfamily),具異四構體之醣蛋白,主要有兩個 α 次單元及 β 次單元所構成。α 次單元位於細胞膜外,當與胰島素結合後其構形發生改變而聚 集,造成 β 次單元上 tyrosine 的自體磷酸化 (auto-phosphorylation) 而引發細胞 質內的一系列訊息傳遞。細胞質中與葡萄糖攝入相關之主要胰島素接受器的受質
為 insulin receptor substrate 1-4 (IRS 1-4)、β-adrenergic receptor 及 adipocyte lipid-binding protein (ALBP)。其中 IRS-1、2參與胰島素受器 auto-phosphorylation 而被磷酸化,造成下游訊息傳遞的啟動序號 (Satiel and Kahn, 2001)。
2. 胰島素受器基質 (insulin receptor substrate, IRS)
目前已知胰島素受器的受質至少有九種,為親水性的蛋白質,其結構在胺基 端有保護作用的 pleckstrin homology (PH) domain,並且鄰接著 phosphotyrosine binding (PTB) domain (Klaus et al., 2004)。其中較普遍的為IRS 1、IRS 2、IRS 3及 IRS 4 (Pessin and Satiel, 2000),其他則是 Gab-1、p60dok、Cb1、APS 及 Shc 的同 型體。這些蛋白質磷酸化後便可以連接下游的效應分子,而活化不同的下游路徑 (Marchand-Brustel et al., 2003)。
3. 葡萄糖轉運蛋白 (glucose transporter, GLUT)
葡萄糖轉運蛋白是由一個基因群 (gene family) 轉錄合成而來,在人體中表現 的葡萄糖轉運蛋白,其結構為 α - 螺旋結構 (α - helix) 往返穿越細胞膜間;在細 胞外側形成六個環狀結構;在細胞膜內側形成五個環狀結構。葡萄糖轉運蛋白的 C 端與 N 端皆位於細胞膜內側 (Olson and Pessin , 1996)。依據其 DNA 序列發 表順序來命名,目前已知的葡萄糖轉運蛋白共有 12 種,依序分別爲 GLUT 1、
GLUT 2、GLUT 3 到 GLUT 12,共有 12 個具有功能性的同分異構型 (functional isoforms),且他們於組織的分佈、調控、轉運能力及受器的專一性上皆有極大的 差異。GLUT 1 和 GLUT 3 負責基礎葡萄糖攝入,它們在細胞膜上的數量不受胰 島素刺激而增加,而 GLUT 2 主要存在於肝臟、胰臟 β 細胞、小腸黏膜細胞與 腎小管上皮細胞中。在肝臟和胰臟β 細胞中,GLUT 2 與同樣具有高 Km 值的葡 萄糖激酶 (glucose kinase) 共同形成「葡萄糖感受器」,當組織正常分泌胰島素時 則共同進行葡萄糖的代謝,但若只在胰臟 β 細胞中的 GLUT 2 的作用則可直接 受血糖調節 (趙, 1997)。
(二)葡萄糖代謝與胰島素阻抗之關係
正常人胰島素濃度與血糖濃度,飯前與飯後都維持類似或固定的水準,飯後 的胰島素與血糖都增加時,靠的是肌肉合成 glycogen 來消化;糖尿病患飯後,
肌肉合成 glycogen 減少 50 %,飯後靠肌肉合成 glycogen 的速率減少,與第 2 型糖尿病人產生胰島素阻抗有關。第 2 型糖尿病人葡萄糖運轉程序較慢,使合成 glycogen 減少 50 %。葡萄糖的運轉先受到胰島素的刺激,結果不是胰島素決定,
是 glycogen 的合成,胰島素不夠敏感,使 glycogen 合成變慢。血中的葡萄糖無 法利用,導致血糖增加,即所謂葡萄糖耐受性不良 (glucose intolerance)。第 2 型 糖尿病人且胰島素阻抗者,脂肪酸濃度是異常的,。最近的研究證實,胰島素阻 抗,肌肉的三酸甘油脂濃度異常 (特別是脂肪細胞) ,原理是脂肪增加時,細胞 內的粒線體,忙著燃燒脂肪,顧不了葡萄糖的運轉;脂肪燃燒氧化後,細胞內產 生許多的檸檬酸 (citrate),它會進一步抑制 PFK 造成 G-6-P 的堆積增加,使葡 萄糖代謝受阻,原因是胰島素阻抗。細胞消耗熱量,主要是燃燒脂肪,約佔 90 %,
其次是葡萄糖,再其次才是蛋白質,這個模式早在 1963 年,由 Randle 等人提 出。不過,這個理論,1996 年被 Roden 等人推翻,他們發現脂肪酸濃度增加,
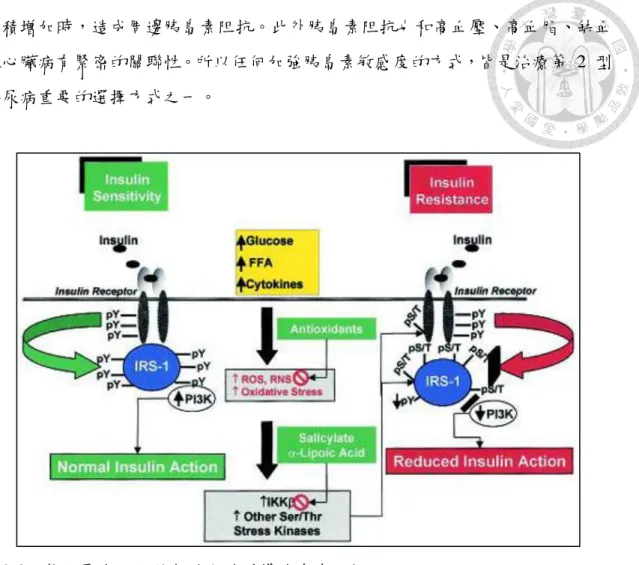
胰島素刺激下的肌肉的 glycogen 合成約減少 50 %,葡萄糖的運轉初期確實受到 抑制,接著是肌肉組織 glycogen 合成被抑制,葡萄糖氧化也被抑制。肌肉組織 堆積過多的脂肪酸,或者是細胞內脂肪酸代謝太慢,產生脂肪酸代謝物,如 diacylglycerol、fatty acid Co A 與 ceramides,這些物質進一步抑制 PI3-kinase 的 活化,使得胰島素接受器的磷酸化作用被抑制,造成使葡萄糖耐受性不良 (三)胰島素阻抗
產生胰島素阻抗後,會逐漸增加胰島素的分泌,所以在第 2 型糖尿病的自然 進程初期時,血中胰島素濃度甚至可增加為非糖尿病人的 2-4 倍。雖然有此一代 償反應來克服胰島素阻抗,但終究隨著疾病的進展,病人的血糖會逐漸上升,甚 至導致β 細胞葡萄糖毒性 (glucotoxicity) 或對一些相關基因表現產生影響,並影 響到β 細胞分泌胰島素的功能。此外在骨骼肌細胞對於餐後葡萄糖的吸收,會有 胰島素受體訊號傳遞途徑、葡萄糖運送蛋白及其磷酸化、肝醣生成和葡萄糖氧化 等多重功能的障礙,導致餐後血糖上升,而且骨骼肌的胰島素阻抗可解釋超過 85
%的全身葡萄糖利用 (glucose disposal) 障礙 (Pendergrass et al., 2007)。
至於肝臟胰島素阻抗主要特色是葡萄糖激酶活性顯著下降,和胰島素抑制肝 糖輸出功能下降 (Gulli G., 1992)。儘管血中胰島素代償性升高,但肝臟葡萄糖產 生 (hepatic glucose production) 的基礎速率還是過高,再加上血中升糖素的增加且 加強肝臟對升糖素的敏感度,都會導致餐後血糖或空腹血糖增高 (Bonora E et al., 2006)。因此,第 2 型糖尿病人的肝臟會過度製造和未充分利用葡萄糖,而且過 高的游離脂肪酸 (free fatty acid) 也會增加肝糖的輸出 (Charles MA et al., 1997)。
第 2 型糖尿病人多數是屬於肥胖的身材,但在非糖尿病人也會因肥胖或腹內脂肪
沉積增加時,造成周邊胰島素阻抗。此外胰島素阻抗也和高血壓、高血脂、缺血 性心臟病有緊密的關聯性。所以任何加強胰島素敏感度的方式,皆是治療第 2 型 糖尿病重要的選擇方式之一。
圖2-3、氧化壓力活化絲氨酸激酶誘導胰島素阻抗。
Figure 2-3. The role of serine kinase activation in oxidative stress-induced insulin resistance.
(Evans et al., 2003)
(四)胰島素分泌缺失
胰島素分泌缺失 (β 細胞衰退) 的原因包括年齡老化、基因影響、胰島素阻抗、
脂肪毒性 (lipotoxicity) 、葡萄糖毒性、類澱粉沉積 (amyloid deposition) 和腸促 胰液素分泌的缺失或作用障礙,初始的特色是當葡萄糖刺激時,其第 1 階段胰島 素分泌喪失 (Pratley RE et al., 2001)。近期研究發現β 細胞功能在診斷第 2 型糖 尿病時就有超過 80 %的喪失,這現象比原先的認知更早且更嚴重 (Defronzo RA, 2009)。除 β 細胞功能下降外,在第 2 型糖尿病人的解剖研究中發現β 細胞的總 量 (mass) 減少,且與β 細胞凋零 (apoptosis) 的增加相關 (Butler AE et al., 2003)。
第 2 型糖尿病人血液中過高的游離脂肪酸會損害 β 細胞分泌胰島素的能力,又
稱為脂肪毒性。慢性高血糖也會損害β 細胞分泌胰島素的功能 (Patanè G et al., 2002),包括第 1 階段和第 2 階段胰島素的分泌,此稱為葡萄糖毒性。總之,雖 然疾病初期肝臟和肌肉的胰島素阻抗早已被証實,但造成第 2 型糖尿病發病漸進 性的 β 細胞衰退 (failure) 則不可能缺席。
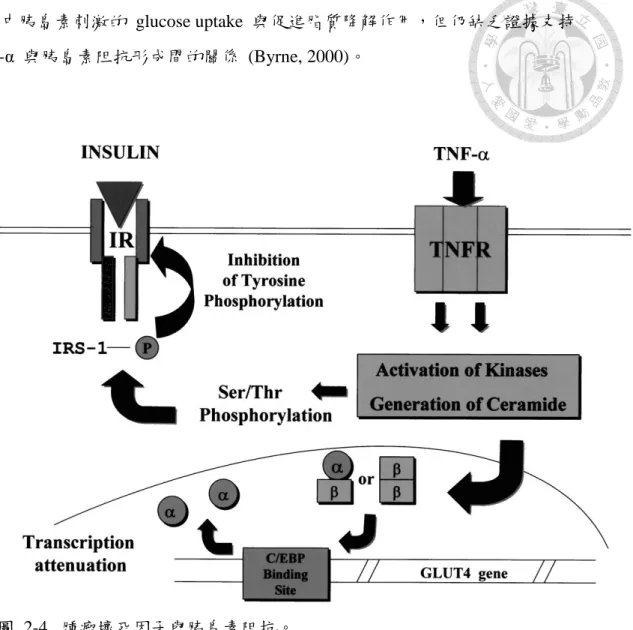
(五)腫瘤壞死因子 tumor necrosis factor-α (TNF-α) 與胰島素阻抗
TNF-α 是分子量為 26 kDa 的細胞穿透蛋白,會與細胞膜上兩個 TNF-α 受 器,p55 (55 kDa)及 p75 (75 kDa) isomers,結合引發 TNF-α 之訊息傳遞。TNF-α 與 TNF-α 受器 p55 (55 kDa) 結合後,會促使 sphingomyelinase 分解鞘髓磷脂 (sphingomyelin) 產生神經醯胺 (ceramide) 及膽鹼 (choline),ceramide 可能會增 加 IRS-1 serine phosphorylation,導致胰島素受體 (IR) 受到抑制而干擾胰島素訊 息傳遞,但詳細機制未明。除此之外,TNF-α 也會誘導其他促發炎細胞激素的產 生,包括 interleukin-1 (IL-1)、interleukin-6 (IL-6) 及 interferon-γ (IFN-γ) 等 (Tracey and Cerami, 1993; Vandenabeele et al., 1995),而這些促發炎細胞激素也與 胰島素阻抗之誘發有關 (Peraldi et al., 1996; Senn et al., 2002)。
近來研究發現,TNF-α可能與肥胖型胰島素阻抗或第二型糖尿病的形成有關 (Moller, 2000),也已有文獻證實 TNF-α 會干擾肝細胞 (hepatocytes)、纖維母細 胞 (fibroblasts) 及骨髓細胞 (myeloid cells) 的胰島素訊息傳遞 (Feinstein et al., 1993; Kroder et al., 1996; Peraldi et al., 1996) 。Moller (2000) 將 TNF-α 可能造成 胰島素阻抗的原因歸納如下:
1. 加速脂解與提高遊離脂肪酸濃度。
2. 降低 GLUT-4、胰島素受器 (IR) 與 IRS-1 合成速率。
3. 抑制胰島素誘發之葡萄糖轉移。
4. 抑制 peroxisomeproliferation-activated receptor (PPAR-γ) 合成。
5. 抑制 IR 磷酸化及 IRS-1 tyrosine 磷酸化。
6. 增加 IRS-1 serine/threonine 磷酸化。
7. 提升 protein tyrosine phosphatase (PTPase) 活性。
TNF-α 引起的胰島素阻抗機制仍有爭議,例如:雖然 TNF-α 會促使胰島素 受器 (IR) 上 tyrosine kinase 活性受損,造成 IRS-1 磷酸化程度下降
(Hotamisligi et al., 1993),但另有研究指出 TNF-α 會造成 IR、IRS-1 與 GLUT-4 含量降低 (Stephens et al., 1997)。另一派學者認為,雖然 TNF-α 會降低脂肪細胞
組織中胰島素刺激的 glucose uptake 與促進脂質降解作用,但仍缺乏證據支持 TNF-α 與胰島素阻抗形成間的關係 (Byrne, 2000)。
圖 2-4. 腫瘤壞死因子與胰島素阻抗。
Figure 2-4. TNF-α induced insulin resistance.
(Chen and Philip, 2000)
(六)以細胞模式探討胰島素阻抗
TNF-α 已被證實會干擾胰島素訊息傳遞,而減少大鼠體內胰島素涉及的全身 葡萄糖代謝利用 (Hsu et al., 1996)。另外亦有學者利用體外方式誘導細胞胰島素 阻抗,並利用此模式探討抗糖尿病有效成份對胰島素阻抗細胞之影響。
1. 脂肪細胞 (adipocytes)
前脂肪細胞分化為成熟的脂肪細胞後才具有胰島素敏感性與受胰島素調控的 葡萄糖攝入能力。誘發脂肪細胞胰島素阻抗模式中,較廣泛被使用的如高劑量胰 島素刺激、長期胰島素刺激、飽和脂肪酸刺激 (Palmitic acid)、TNF-α、dexamethaone 等,皆可誘發胰島素抗性 (Hoehn et al., 2008) 。其中以胰島素與 TNF-α 誘發的
模式被認為會減低 IRS-1 的表現量,因而削弱了胰島素訊息傳遞路徑,以 dexamethaone 誘發阻抗的機制則為減低 Akt 磷酸化而減少 GLUT 4 轉位。已有 許多文獻指出,脂肪細胞以飽和脂肪酸誘發胰島素抗性 是透過活化 TLR-4 (toll-like receptor 4) 而啟動發炎反應,TLR-4 在脂肪細胞分化的第 5 天開始會大 量表現 (Song et al., 2006); TLR-4 被活化後會接續活化 NF-κB 而刺激下游基因 如:TNF-α、IL-6、COX-2、MCP-1 等表現量增加,而若將 TLR-4 基因剔除則 發炎反應被抑制,減少胰島素抗性。Iwata 等人 (2001) 以另外一種胰島素敏感藥 物 pioglitazone (TZD 的一種) 處理 TNF-α 誘發胰島素阻抗之 3T3-L1 脂肪細 胞,結果發現 pioglitazone 可使脂肪細胞 IR、IRS-1 tyrosine 磷酸化與 PI3-kinase 活性恢復正常,進而改善葡萄糖攝入的能力。
2. 骨骼及細胞 (skeleton muscles)
在人體口服葡萄糖耐受性試驗中,骨骼肌負載了 70~90 % 的葡萄糖攝入以維 持血液中葡萄糖的恒定,而進入肌肉細胞的葡萄糖則以肝醣的形式儲存;肝醣合 成的過程中需要胰島素的刺激,當肌肉細胞或肝細胞接收到胰島素的刺激後活化 下游的 IRS-1、PI3K、PDK-1 及 Akt/PKB (protein kinase B),活化的 PKB 會將 GSK 3 (glycogen synthase kinase 3) 磷酸化,當 GSK 3 被磷酸化後即失去活性而 不會將 GS (glycogen synthase) 自未磷酸化的活化態轉變成磷酸化的失活態,因 此 GS 持續活化而將葡萄糖以肝醣的方式儲存在肝細胞或是肌肉細胞中。
TNF-α 對骨骼肌細胞的影響仍不清楚,Del Aguila 等人 (1999) 報告指出 TNF-α 處理肌肉細胞 C2C12 會使胰島素刺激的 IRS-1 與 IRS-2 調控 PI3K 活 化能力受損,使胰島素調控的 IRS-1 tyrosine 磷酸化降低,並抑制胰島素誘發的 p42MAPK 與 p44MAPK tyrosine 的磷酸化,而發展成胰島素阻抗的細胞模式,
Schmitz-Peiffer 等人 (1999) 將 C2C12 肌肉細胞經 palmitate 處理後,結果發現飽 和脂肪酸會提升細胞內 ceramide (PBK 的抑制擠) 含量及 MAPkinase 活性,並 抑制肝醣合成與 PKB 的磷酸化,最後引起肌肉細胞產生胰島素阻抗;Cusi 等人 (2000) 則指出,以 palmitate處理 C2C12 肌肉細胞株實惠產生胰島素阻抗,但非 不影響其 IRS-1 tyrosine 磷酸化。另外有研究指出,C2C12 經 interleukin-6 (IL-6) 處理後,會降低 IR 表現量及 IR tyrosine autophosphorylation、IRS-1 含量及 IRS-1 tyrosine phosphorylation、PI3K 活性、Akt serine phosphorylation 以及葡萄 糖轉運蛋白 (GLUT 4) 表現,造成 C2C12 細胞葡萄糖攝入降低捯飭胰島素阻抗現
象,而 loperamide 則可以恢復因 IL-6 誘導所造成的胰島素減低 (Tzeng et al., 2005)。
3. 肝細胞 (hepatocytes)
肌肉細胞與脂肪細胞為常應用於評估糖尿病成份的體外細胞模式,但此兩株 須經分化後才可以獲得對胰島素的感受度,而且操作步驟複雜且耗時,不利應用 於抗糖尿病成份之篩選 (Cheng et al., 2008)。而肝細胞不需經分化即可具有對胰 島素的感受度,近年之研究將肝細胞誘導成胰島素阻抗,並以此體外模式評估抗 糖尿病有效成份。Cheng 等人 (2008) 先以 TNF-α誘導小鼠肝臟細胞 FL83B 胰 島素阻抗,再經胰島素處理後,以葡萄糖分析套組測定細胞培養液中葡萄糖含量,
發現培養液葡萄糖含量於相同處理時間下會隨 TNF-α濃度增加而增加,表示 TNF-α 會減少細胞經胰島素刺激的葡萄糖攝入,並以此模式快速篩選出植物中降 血糖之有效成份,而這些有效成份亦可以增加 IRS-1 表現,恢復胰島素阻抗細胞 中胰島素之訊息傳遞。另外,Senn 等人 (2002) 以 IL-6 處理小鼠初代肝細胞及 人類肝癌細胞株 Hep G2,發現會減少經胰島素刺激之 IRS-1 及 PI3K 表現,此 外亦會抑制調控胰島素訊息下游代謝活動之 Akt 的活性,最後導致小鼠初代肝細 胞中肝醣合成受到抑制。因此由以上得知 TNF-α及 IL-6 可以誘導肝細胞造成細 胞層次之胰島素阻抗,並應用於降血糖及抗糖尿病成份之評估。
四、甘薯
甘薯 (Ipomoea batatas (L.) Lam.) 又叫地瓜、山芋、蕃薯,為旋花科
(Convolvulaceae)、甘薯屬 (lpomoea),匍匐地面生長特性的一年生或多年生蔓生 草本植物。根據 FAO (Food and Agriculture Organization) 2005年之調查,甘薯為 世界第五大經濟作物,依序為小麥、稻米、玉米、樹薯及甘薯 (Rumbaoa et al., 2009)。
甘薯易於栽培管理,其塊根與莖、葉具有多種用途,是少數全株皆可被利用的雜 糧與蔬菜作物,更是天災人禍時之救援作物。甘薯塊根的營養成分相當豐富,含 澱粉、糖、蛋白質、脂肪、纖維、維生素 A、B 群、C和鐵、鈣等多種營養元素,
對人體保健裨益甚大,是非常好的膳食纖維來源,堪稱為現代物美價廉的保健食 品;此外,亦被視為一種生理鹼性食品,可中和一些生理酸性食品,調整人體代 謝機能 (黃,2002)。過去甘薯主要作為雜糧、飼料、澱粉等用途,近年來由於國 人飲食習慣及膳食結構改變,食用或是食品加工用甘薯及葉菜甘薯漸受消費者青 睞,甘薯為國產糧食作物中地位僅次於稻米者,種植一次可連續採收兩年,同時 整年均能收穫,2012 年全省栽培面積約 3171.3 公頃左右,產量約為 78931 公 噸 (台灣農業年報,2013) 。
根據台灣省農業試驗所嘉義分所的資料,現今台灣省 13 種栽培最多之甘薯 品種特性介紹如下 (台灣省農業試驗所,2013) :
1. 食味特性的甘薯品種 - 台農 57 號
莖黃綠色,節間帶淡紫色,毛茸少,葉形五深裂或三深裂,背面葉脈中助及 其基部均為深紫色,葉脈為淡紫色,頂葉黃綠色,塊根表皮棕黃色,無縱溝,
肉色橙黃,塊根紡錘形。
2. 紅心豐産的甘薯品種 - 台農 62 號
莖黃綠色,節間帶暗紫色,葉為心臟形,背面葉脈中肋及其基部均為赤紫色,
頂葉黃綠色,塊根表皮淡紅褐色,肉色橙紅,切面易氧化變為綠橙紅色,塊 根為下膨紡錘形。
3. 高胡蘿蔔素的甘薯品種 - 台農 63 號
莖為黃綠色,葉為心臟形,頂葉為黃綠色,背面葉脈中肋及其基部均為綠色,
塊根表皮鮮紅色,肉色深橙紅色,塊根為短紡錘形。
4. 用途廣泛的甘薯品種 - 台農 64 號
莖中大為紫色,葉為心臟形,頂葉為綠色,葉背基及葉脈均為紫色。塊根表 皮淡棕黃色,光滑美觀,塊根肉色深橙紅,塊根為紡錘形。
5. 含量蛋白質的甘薯品種 - 台農 65 號
莖粗大為紫紅色,無毛茸,葉為心臟形,頂葉淡紫紅色,葉背面、葉脈及其 基部均為淡紫色。塊根表皮棕紅色,光滑,塊根肉色橙紅,塊根為短紡錘形。
6. 適應不同期作栽培的甘薯品種 - 台農 66 號
莖粗大且強硬,綠色,無毛茸,葉為盾形,頂葉亦為綠色,葉背面、葉脈及 基部均為綠色,塊根表皮淡棕紅色,表皮尚光滑,肉色橙紅。
7. 澱粉消化性優良的食用及飼料用品種 - 台農 67 號
短蔓,莖淡紫紅色,無茸毛。葉為心臟形,綠色,葉背基部紫色,頂葉綠色,
塊根紡錘形,表皮淡紅,肉色橙黃,表皮光滑無脈紋。
8. 適合食品加工、製餡、油炸物兼作夏季蔬菜用品種 - 台農 68 號
短蔓,莖綠色,無茸毛。葉為心臟形,綠色,葉脈及葉背中脈基部為綠色,
頂葉綠色,塊根紡錘,表皮尚光滑淡黃色,肉色淡白色。
9. 豐産的食用紅肉甘薯品種 - 台農 69 號
短蔓,莖綠色,無茸毛。葉為長心臟形,綠色,葉脈及葉背中脈基部為綠色,
頂葉綠色,塊根紡錘形,表皮脈少,無瘤目及縱溝,深紫紅色,肉色深橙紅。
10. 耐低肥性的食用品種 - 台農 70 號
長蔓,莖綠色,無茸毛。葉為盾形,綠色,葉表中脈黃綠,葉背中脈紅紫,
頂葉綠色,葉柄綠色,具茸毛。塊根紡錘形,表皮紅褐色,根痕較深,皮層 薄,肉色深橙紅。
11. 葉菜用甘薯 - 台農 71 號
莖蔓長度中等,株型半直立,莖色深綠色,無茸毛。頂葉淡綠色,成熟葉呈 深綠色,新生葉之葉形為戟形,成熟葉呈鼻形,葉柄為綠色,且光滑無茸毛。
12. 優良食用加工用夏作品種 - 桃園 1 號
短蔓,淡綠色,無茸毛。葉為心臟形,淡綠色,葉脈及葉背中脈為綠色,葉 基部帶紫色,頂葉綠色,塊根紡錘形,表皮紫紅色,根痕淡,肉色橙黃微帶 紫暈。
13. 葉菜用甘薯 - 桃園 2 號