國立臺灣大學生命科學院生化科技學系 碩士論文
Department of Biochemical Science and Technology College of Life Science
National Taiwan University Master Thesis
EB 病毒蛋白質 Rta 對粒線體的影響
Role of Rta of Epstein-Barr virus in mitochondria
蔡曉涵 Hsiao-Han Tsai
指導教授:張麗冠 博士 Advisor: Li-Kwan Chang, Ph.D.
中華民國 105 年 6 月
June, 2016
謝辭
碩班的日子很快就過了,但同時卻又覺得很漫長,兩年多來跌跌撞撞地終於 也走到了這一步,還以為謝辭並不難打的,但在這當下看著游標閃爍卻頓時覺得 語塞。首先要最感謝張麗冠老師,不只在學業上不斷地引導我正確的研究方向,
在實驗室之外也從不減關心與照顧,能夠在老師這裡學習是非常幸運的一件事。
謝謝口試委員劉世東老師、莊健盈老師、張世宗老師和陳慧文老師,對這個研究 提出了許多討論與建議,並指出一些我忽略的細節,讓這本論文能更臻完善;尤 其謝謝劉世東老師還親自幫我們修改初稿。謝謝實驗室裡的助理、各位學長姊和 學弟妹,在我剛進實驗室時協助我上軌道、融入環境,在實驗進度停滯時和我一 起想點子、想方法,因為大家不吝嗇的幫忙我才有辦法做出這些實驗、完成這個 研究。謝謝朋友一直以來的扶持,半拖半拉、半哄半騙地讓我能持續往前走,若 沒有這些陪伴也許我無法順利走完畢業這條路。最後要謝謝我的父母和家人,在 這一段不算短的時間裡包容我所有的任性與懦弱,家庭永遠是我最堅強的後盾。
其實心底明白,拿到碩士學位只是人生中一個小小的里程碑,之後還有更長的路、
更多的關卡,期望自己能夠一直秉持著誠實與善良,保有信心,不被心中的不安 和恐懼所支配,直挺挺地迎接人生接下來的每一個路口。
ii
摘要
Epstein-Barr Virus (EB 病毒) 屬於人類疱疹病毒科,其生活史包括潛伏期 (latent life cycle) 與溶裂期 (lytic cycle),而極早期蛋白質 Rta 是促使 EB 病毒進 入溶裂期的關鍵轉錄因子之一。本實驗室在先前的研究中發現 Rta 蛋白質在 EB 病毒潛伏期時會分布於粒線體。粒線體參與了許多細胞生理途徑,如能量的產生、
細胞死亡、宿主免疫反應的誘導和老化等;而粒線體自噬 (mitophagy) 為維持粒 線體網絡健康的重要機制。本研究欲證實 Rta 蛋白質上是否具有粒線體導向序列 (mitochondria targeting sequence,MTS),並初步探討 Rta 蛋白質位於粒線體上的 意義及其扮演的角色。首先以具有病毒潛伏的 P3HR1 細胞、B95-8 細胞及 293-2089 細胞為材料,利用免疫螢光染色分析探討 Rta 於細胞中的分佈情形,結 果證實 Rta 蛋白質位於粒線體;同時利用粒線體分離分析結果指出,細胞中 Rta 蛋白質在細胞質與粒線體中皆有表現。接著利用 293T 細胞為材料,轉染入不同 Rta 片段之建構質體,可藉由觀察其進粒線體與否推定出 MTS 為 Rta 蛋白質之 N 端第 68 至 90 胺基酸片段,並由生物資訊軟體預測出其具有 α 螺旋體結構。
接著以共免疫沉澱分析證明了 Rta 蛋白質會與粒線體外膜蛋白導入受體 (import receptor) Tom20 結合。而後以 ATP 測定分析結果發現,在 293T 細胞表現 Rta 後 會使細胞中的 ATP 產量提高。最後,以 carbonyl cyanide m-chlorophenyl hydrazine (CCCP) 處理 293T 細胞,誘導粒線體損害進而引發粒線體自噬,結果發現 Rta 蛋 白質的表現可能會促進粒線體自噬。總和以上所述,本研究首度證實 Rta 含有一 α 螺旋體結構的 MTS,可位於粒線體並影響粒線體的功能。
關鍵字:EB 病毒、Rta、粒線體、粒線體導向序列、粒線體自噬
Abstract
Epstein-Barr virus (EBV) is a human herpesvirus with two distinct life cycles, latent
and lytic. Rta, an immediate-early protein, is an important transcription factor required
for initiating the lytic cycle of EBV. Our preliminary study found that Rta is localized at
mitochondria during EBV latency. Mitochondrium is a multifunctional organelle with
diverse roles including energy production, apoptosis, elicitation of host immune
response, and aging. Moreover, mitochondria autophagy (mitophagy) is a critical
mechanism to maintain mitochondrial health and homeostasis. This study aims to
investigate whether Rta contains mitochondria targeting sequence (MTS), and examine
roles of Rta in mitochondria. First, immunofluorescence analysis indicated that Rta
colocalizes with mitochondria in P3HR1 cells, B95-8 cells and 293-2089 cells during
both EBV latency and lytic cycle. Mitochondria isolation assay was performed and
indicated that Rta expresses both in cytosolic and mitochondrial fraction. Mutagenesis
and bioinformatic analyses demonstrated that an MTS is localized between amino acid
residues 68 to 90 in Rta, which forms an amphipathic α-helix structure.
Co-immunoprecipitation assay further indicated that Rta binds to Tom20, an import
receptor on mitochondrial outer membrane. ATP determination assay also revealed that
iv
overexpressing Rta increases ATP levels in 293T cells. Finally, treatment of carbonyl
cyanide m-chlorophenyl hydrazine (CCCP) to 293T cells with expression of Rta affects
mitophagy. Taken together, this study demonstrates that Rta is a mitochondrion-targeted
protein that contains an amphipathic -helical MTS, which may affect the functions of
mitochondria.
Key words: Epstein-Barr virus, Rta, mitochondria, mitochondria targeting sequence,
mitophagy
目錄
謝辭 ... i
摘要 ... ii
Abstract ... iii
第一章、 前言 ... 1
一、 Epstein-Barr virus (EB 病毒) 的發現及其致病性 ... 1
二、 EB 病毒的遺傳物質及結構 ... 2
三、 EB 病毒的生活史 ... 2
潛伏期 (Latency)....………...3
溶裂期 (Lytic cycle)………..3
四、 EB 病毒的極早期蛋白質 ... 5
Zta 蛋白質……….5
Rta 蛋白質……….7
Rta 轉錄活性的調控……….7
Rta 參與細胞生理途徑………...10
五、 粒線體 (mitochondria) 的生理重要性 ... 11
粒線體藉氧化磷酸化合成 ATP………...12
粒線體參與細胞凋亡 (apoptosis)………..12
粒線體與免疫系統的關聯………..14
粒線體自噬 (mitophagy)………....14
六、 粒線體導向序列 (mitochondrial targeting signal, MTS) ... 15
七、 病毒對粒線體功能的調節 ... 16
1. 病毒在宿主細胞中引發氧化壓力 (oxidative stress)……….16
vi
2. 病毒調節宿主的粒線體膜電位 (mitochondrial membrane potential,
MMP)………...17
3. 病毒可調控細胞凋亡 (apoptosis)………..18
4. 病毒影響粒線體相關的抗病毒免疫機制………..19
5. 病毒改變粒線體在細胞內的分布………..19
6. 病毒造成宿主粒線體 DNA 耗竭 (mitochondrial DNA depletion).19 八、 EB 病毒與粒線體間的關係 ... 20
九、 研究目的 ... 21
第二章、 材料與方法 ... 22
一、 細胞株 ... 22
二、 EB 病毒的溶裂期誘導 ... 22
三、 質體與抗體 ... 23
四、 質體 DNA 的萃取 ... 23
五、 細菌 ... 23
六、 細菌轉型作用 (transformation) ... 23
七、 細胞轉染 (Transfection) ... 23
八、 免疫螢光染色分析 (Immunofluorescence analysis) ... 24
九、 粒線體的分離分析 ... 25
十、 西方點墨法分析 (Western blot analysis) ... 25
十一、 免疫沈澱法 (Immunoprecipitation, IP) ... 26
十二、 加藥處理 ... 26
十三、 ATP 測定分析 (ATP Determination assay) ... 26
第三章、 結果 ... 28
一、 Rta 蛋白質存在於粒線體 ... 28
二、 Rta 蛋白質會與 Tom20 結合 ... 29
三、 Rta 的粒線體導向序列 (MTS) 分析 ... 30
四、 Rta 的 N 端第 68 至 90 胺基酸之二級、三級結構預測 ... 31
五、 MTS 上四個疏水性胺基酸突變使 Rta 無法進入粒線體 ... 32
六、 Rta 蛋白質影響細胞內 ATP 的產量 ... 32
七、 Rta 蛋白質影響粒線體自噬 (mitophagy) ... 33
第四章、 討論 ... 34
圖表 ... 41
表 1、 本研究所使用之質體 ... 41
表 2、 本研究所使用之抗體 ... 45
圖 1-1、EB 病毒的結構示意圖 ... 46
圖 1-2、EB 病毒的生活史 ... 47
圖 1-3、EB 病毒的溶裂期生活史 ... 48
圖 1-4、EB 病毒極早期蛋白質 Rta 與 Zta 的表現 ... 49
圖 1-5、Rta 蛋白質的功能區及入核訊號示意圖 ... 50
圖 1-6、粒線體參與的生理途徑 ... 51
圖 1-7、病毒可調節粒線體功能 ... 52
圖 3-1、Rta 蛋白質與粒線體在 EB 病毒潛伏期的免疫螢光染色分析 ... 53
圖 3-2、Rta 蛋白質與粒線體在 EB 病毒溶裂期的免疫螢光染色分析 ... 54
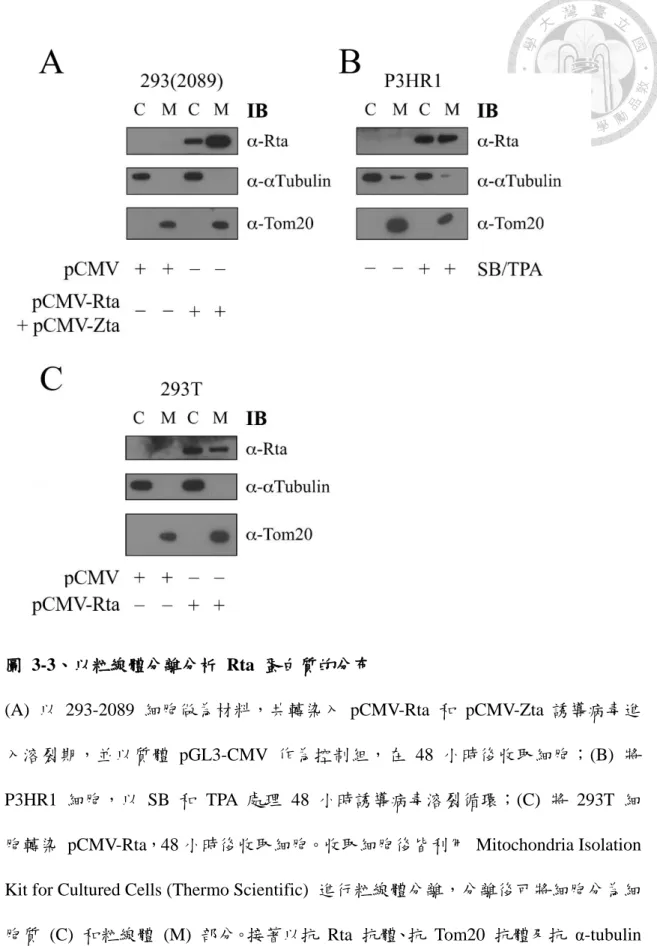
圖 3-3、以粒線體分離分析 Rta 蛋白質的分布 ... 56
圖 3-4、Rta 與 Tom20 在細胞內結合 ... 57
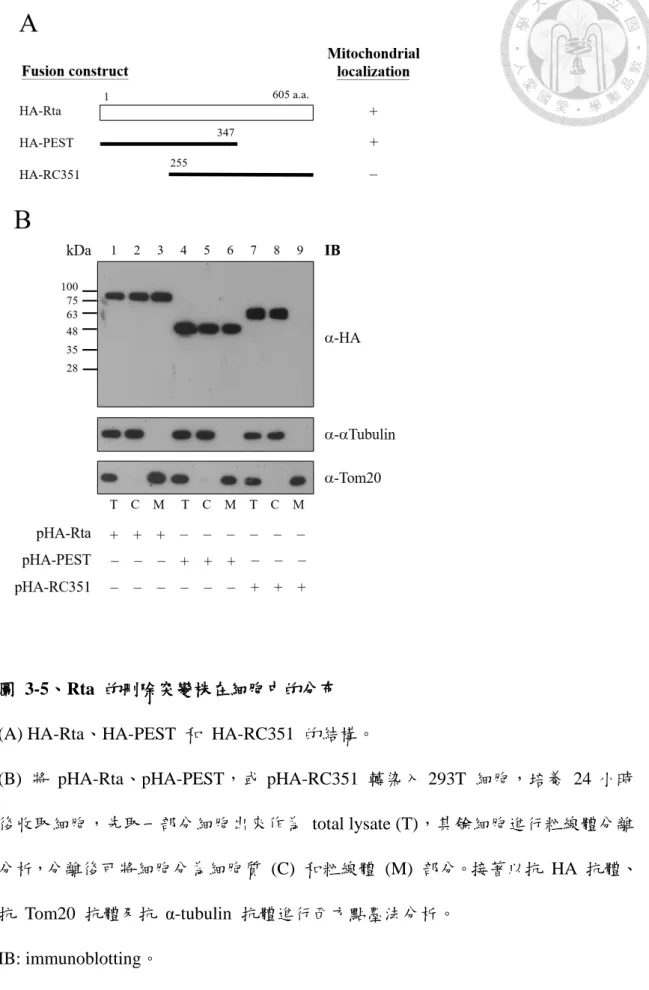
圖 3-5、Rta 的刪除突變株在細胞中的分布 ... 58
viii
圖 3-6、Rta 及其刪除突變株的粒線體導向序列分析 ... 60
圖 3-7、Rta 的突變株和粒線體在 293T 細胞中的免疫螢光染色分析 ... 62
圖 3-8、Rta 的 N 端片段之二級、三級結構預測 ... 63
圖 3-9、Rta 及 Rta 的 MTS 突變株在細胞中的分布位置 ... 64
圖 3-10、Rta 的 MTS 突變株和粒線體在 293T 細胞中的免疫螢光染色分析 ………..65
圖 3-11、Rta 對細胞產生能量的影響 ... 67
圖 3-12、Rta 對粒線體自噬的影響 ... 68
圖 4-1、Rta 蛋白質與粒線體的關係 ... 69
參考文獻………..70
附錄 ... 98
附錄 1、Rta 蛋白質在電子顯微鏡觀察下於 P3HR1 細胞內的分佈情形 (王文 宏, 未發表) ... 98
附錄 2、粒線體內膜上的電子傳遞鍊與 ATP 合成酶 ... 99
附錄 3、細胞凋亡的外在與內在途徑 ... 101
附錄 4、粒線體參與的抗病毒免疫機制 ... 103
附錄 5、粒線體的動態恆定以及質量管制 ... 104
附錄 6、粒線體蛋白質的運輸機制 ... 105
附錄 7、MTS 的結構特性及其與 Tom20 受體蛋白的相互作用 ... 106
第一章、 前言
一、 Epstein-Barr virus (EB 病毒) 的發現及其致病性
Epstein-Barr virus (EB 病毒) 屬於疱疹病毒科 (Herpesviridae),又稱為第四型 人類疱疹病毒 (human herpesvirus 4, HHV-4),為最常見的人類病毒之一。西元 1958 年,外科醫師 Denis Burkitt 在非洲孩童體內發現一種原發於上下頷骨的淋巴瘤,
其 病 徵 和 型 態 皆 和 已 知 腫 瘤 不 同 , 便 將 其 命 名 為 巴 克 氏 淋 巴 瘤 (Burkitt’s lymphoma) (Burkitt, 1958),並提出此腫瘤具有感染性物質的假說 (Shaper and Burkitt, 1962)。而在西元 1964 年,英國科學家 Anthony Epstein、Yvonne Barr 及 Bert Achong 成功從淋巴瘤樣本中建立細胞株,並以電子顯微鏡在此細胞內觀察到 結構類似疱疹病毒的病毒顆粒 (Epstein and Barr, 1964; Epstein et al., 1964)。接著在 西元 1966 年,Werner Henle 和 Gertrude Henle 利用免疫螢光分析等方法證實此 病毒與其他疱疹病毒不同,為新的病毒株,並將之命名為 Epstein-Barr virus (EB 病 毒) (Henle and Henle, 1966b, 1966a)。Henle 等人 (1966) 也在罹患巴克氏淋巴瘤的 病人血清中偵測到高含量的 EB 病毒抗原,證實了 EB 病毒與此腫瘤的直接關連 性。 (Henle et al., 1966)。
EB 病毒的感染率非常高,估計全球約有 90% 以上的成人受其感染 (Young and Rickinson, 2004)。EB 病毒主要藉由唾液傳播,可感染人類的 B 淋巴細胞和 口咽部上皮細胞 (Nilsson et al., 1971; Sixbey et al., 1984)。目前已知與 EB 病毒感 染相關的疾病包括了傳染性單核球增多症 (infectious mononucleosis) (Niederman et al., 1968)、鼻咽癌 (nasopharyngeal carcinoma) (Wolf et al., 1973)、巴克氏淋巴瘤 (Burkitt’s lymphoma) (Geser et al., 1982)、霍奇金氏症 (Hodgkin’s disease) (Weiss et al., 1989) 及胃腺癌 (gastric carcinoma) (Shibata and Weiss, 1992) 等疾病。
2
二、 EB 病毒的遺傳物質及結構
EB 病毒的遺傳物質為雙股的去氧核醣核酸 (deoxyribonucleic acid, DNA),包 含了八十五個基因,在 B95-8 細胞株內其基因體的全長為 172 kb (Baer et al., 1984)。病毒的基因體被包裹於由 162 個 capsomer 所組成的正二十面體蛋白質外 鞘殼 (capsid) 內,形成核蛋白質外鞘殼 (nucleocapsid);外鞘殼體外會再被外套膜 (envelope) 包覆,外套膜的來源為宿主細胞膜及核膜,上面嵌有病毒表現的膜蛋 白; 而在核蛋白質外殼和外套膜之間的空腔則填充了 間質蛋白質 (tegument protein),如此可構成完整的病毒顆粒,其直徑為 155-200 nm (Dolyniuk et al., 1976a;
Dolyniuk et al., 1976b; Johannsen et al., 2004) (圖 1-1)。
EB 病毒的基因體上有六段重複序列,分別為位於末端的兩個 terminal direct repeats (TRs),與位於基因體中間的 internal direct repeats (IRs) (Given and Kieff, 1979; Given et al., 1979)。在 EB 病毒的病毒顆粒中,病毒的基因體是以線狀形式 存在,但當病毒感染宿主細胞後,基因體兩端的 TR 序列會互相連接形成環狀的 質體構造。EB 病毒的基因是以 BamH1 限制酶圖譜來定義,B95-8 細胞株內的 EB 病毒基因體經 BamH1 限制酶切割後,依片段大小以英文字母 A-Z 與 a-g 的 順序編碼,而病毒基因便依據其所在的片段與 Open Reading Frame (ORF) 的方向 來命名,如 BRLF1 (BamHI, R fragment, Left Open Reading Frame 1) 即是基因體經 BamHI 限制酶切割後,R 片段朝左方向第一個表現的 ORF (Dambaugh et al., 1980)。
三、 EB 病毒的生活史
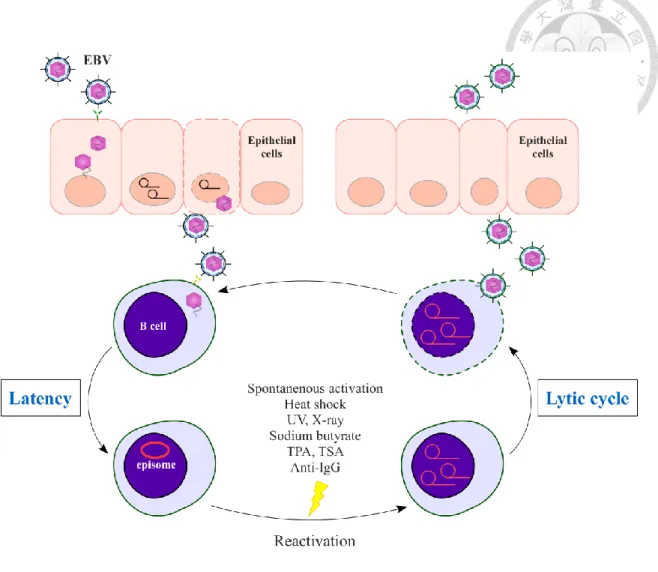
EB 病毒會感染人類的 B 淋巴細胞及上皮細胞,當初次被 EB 病毒感染時,
病毒會先在口腔上皮細胞中複製病毒顆粒,並於感染數週至數個月內釋出少量的
病毒顆粒以感染 B 細胞,並長期潛伏在淋巴組織中。EB 病毒的生活史可區分為 潛伏期 (latency) 及溶裂期 (lytic cycle) 兩個階段。此兩種生活史的轉換將有助於 病毒逃離免疫系統的攻擊,並能建立長期的感染 (Bajaj et al., 2007; Young and Rickinson, 2004) (圖 1-2)。
潛伏期 (Latency)
EB 病毒感染宿主後病毒會以三種潛伏模式存在於宿主細胞:latency Ⅲ、
latency Ⅱ 和 latency Ⅰ,依病毒感染的淋巴細胞類型或感染後的潛伏期進程而分 別表達不同的潛伏期基因 (Bajaj, et al., 2007; Amon and Farrell, 2005)。潛伏期時,
EB 病毒的基因體會形成一環狀質體 (episome),可藉由病毒製造的 EBNA1 蛋白 質結合上 viral origin of replication (oriP) 序列 (Adams, 1987; Kirchmaier and Sugden, 1995; Yates and Guan, 1991),隨著宿主進入 S-phase 而複製,並伴隨著細胞 染色體一起分裂而分布至每個子細胞中 (Hampar et al., 1974; Yates et al., 1985)。EB 病毒於潛伏期時僅表現少量基因,包括六種核抗原蛋白質 (nuclear antigens):
EBNA1、EBNA2、EBNA3A、EBNA3B、EBNA3C 和 EBNA-LP;三種潛伏期膜蛋 白質 (latent membrane proteins):LMP1、LMP2A 和 LMP2B;以及一系列的小片段 核醣核酸 (micro ribonucleic acid, microRNA) 如 EBER1、EBER2 (Allday et al., 1989; Cai et al., 2006)。EB 病毒藉由這些分子調控宿主的基因表現、訊息傳遞,以 幫助 EB 病毒基因體穩定存在於宿主體內,或進一步造成 B 細胞的轉型以及細胞 癌化 (Pfeffer et al., 2004; Yenamandra et al., 2009; Young and Murray, 2003)。
溶裂期 (Lytic cycle)
潛伏期的 EB 病毒受到紫外光 (ultraviolet, UV)、X 光 (X-ray) (Henderson and
4
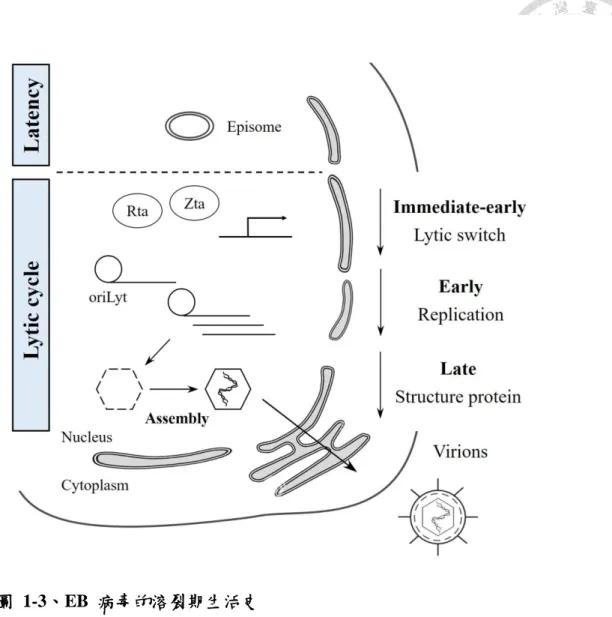
Long, 1981)、sodium butyrate (SB) (Kallin et al., 1979; Luka et al., 1979; Nutter et al., 1987)、12-O-tetradecanoylphorbol 13-acetate (TPA) (Flemington and Speck, 1990b; zur Hausen et al., 1978)、trichostatin A (TSA) (Chang and Liu, 2000)、免疫球蛋白質 (immunoglobulin) (Tovey et al., 1978) 及 transforming growth factor beta (TGF-β) (Schuster et al., 1991) 等誘導,或病毒蛋白質 Zta (BZLF1,EB1,ZEBRA 或 Z) 及 Rta (BRLF1 或 R) 自我調控時,EB 病毒會由潛伏期進入溶裂期。
EB 病毒進入溶裂期之後,依照病毒基因表現的時間與功能,可將溶裂期的基 因分為極早期基因 (immediate-early genes)、早期基因 (early genes) 和晚期基因 (late genes)。EBV 的極早期基因,BZLF1 及 BRLF1,在溶裂期中最早表現,其所 表現的蛋白質 Rta 及 Zta 具有轉錄因子 (transcription factors) 的活性,可以活化 彼此和下游的早期及晚期基因,也能調控宿主的基因表現 (Chang et al., 1990; Liu and Speck, 2003; Manet et al., 1989; Quinlivan et al., 1993)。早期基因受極早期基因 誘導產生,其產物主要為病毒複製所需要的蛋白質,包括 DNA 聚合酶 (DNA polymerase) BALF5、解旋酶 (helicase) BBLF4 與引子合成酶 (primase) BSLF1 等 (Fixman et al., 1992; Liu et al., 1996a; Yokoyama et al., 1999)。晚期基因的產物則大 多為病毒的結構蛋白質,用以組裝病毒顆粒,如病毒的外鞘蛋白質 (capsid proteins) BDLF1、BORF1 (Henson et al., 2009; Wang et al., 2011),以及填充於外套膜和外鞘 殼之間的間質蛋白質 (tegument proteins) (Chiu et al., 2012; Johannsen, et al., 2004)。經由上述蛋白質的表現,EB 病毒能夠產生具有感染性的病毒顆粒,並由 宿主細胞中釋放後,藉由外套膜上的醣蛋白質,如 glycoprotein 350 (gp350)、
glycoprotein 42 (gp42) 或 glycoprotein H (gH) 等,附著於另一細胞表面上,進行下 一次的感染 (Molesworth et al., 2000; Nemerow et al., 1987) (圖 1-3)。
四、 EB 病毒的極早期蛋白質
EB 病毒的極早期蛋白質 Zta (BZLF1,EB1,ZEBRA 或 Z) 和 Rta (BRLF1 或 R) 在溶裂期中最先表現,為驅動 EB 病毒由潛伏期進入溶裂期的關鍵樞紐 (Amon and Farrell, 2005; Ragoczy et al., 1998; Speck et al., 1997; Tsurumi et al., 2005;
Zalani et al., 1996)。Zta 是由 BZLF1 基因所表現,BZLF1 基因在 EB 病毒基因 體上的位置緊鄰於 BRLF1 基因之後,其基因的活化主要透過上游啟動子 (Zp) 的 調控而轉錄出長度為 1 kb 的單基因 (monocistronic) mRNA,經轉譯作用後產生出 Zta 蛋白質 (Manet, et al., 1989)。Rta 是由 BRLF1 基因所表現,其基因透過上游 啟 動 子 (Rp) 的 調 控 , 可 轉 錄 出 長 度 為 4.0 kb 的 BRLF1-BZLF1 雙 基 因 (bicistronic) mRNA,透過截切作用 (splicing) 後可產生長度為 3.3 kb 或 0.8 kb 的 mRNA,其中 4.0 kb 和 3.3 kb 的雙基因 mRNA 皆能同時轉譯出 Rta 及 Zta 蛋 白質 (Manet, et al., 1989) (圖 1-4)。
先前研究指出,Zta 的表現能促進 EB 病毒溶裂期的進行 (Amon and Farrell, 2005; Speck, et al., 1997; Tsurumi, et al., 2005),而 Rta 的表現亦能啟動溶裂期生活 史,其對於溶裂期基因的活化能力較 Zta 弱 (Ragoczy, et al., 1998; Zalani, et al., 1996),但完整的溶裂期蛋白質表現仍需要 Rta 與 Zta 的共同存在下才能順利完 成 (Feederle et al., 2000),顯示出兩者在病毒溶裂期中的重要性。而 Zta 與 Rta 皆為轉錄作用的活化因子,可以活化彼此的啟動子並調控 EB 病毒溶裂期下游的 基 因 表 現 , 或 彼 此 形 成 複 合 體 共 同 調 控 基 因 轉 錄 (Chang et al., 2010;
Chevallier-Greco et al., 1986; Giot et al., 1991; Holley-Guthrie et al., 1990)。
Zta 蛋白質
Zta 蛋白質是由 245 個胺基酸所組成,其 N 端為轉錄活化區 (transactivation
6
domain),C 端則包含了 DNA 結合區 (DNA binding domain) 及同型二聚體形成 區 (dimerization domain)。Zta 是 bZIP (basic leucine zipper) 家族的轉錄因子,其 序列及結構與 c-Jun、c-Fos 等 activation protein-1 (AP-1) 轉錄因子具有同源性,
但具有其自身獨特的 DNA 結合區與同型二聚體形成區 (Chang, et al., 1990)。Zta 能夠與特定的 AP-1 結合序列或 Zta-responsive elements (ZREs) 序列結合,進而 活化自身或其他基因的表現 (Chang, et al., 1990; Farrell et al., 1989; Flemington and Speck, 1990a; Lieberman et al., 1990);其中典型的 ZRE 序列為 TGTGCAA 相似序 列 (T-G/T-T/A-G-T/C-G/C/A-A) (Lieberman, et al., 1990)。
Zta 偏好結合於受甲基化修飾的 ZREs 上 (Bergbauer et al., 2010; Bhende et al., 2004);一般而言,甲基化修飾會造成基因的靜默現象,然而 Zta 則對於某些 受甲基化修飾的 ZRE 序列有較高的親和力,如 Zta 偏好與 BRLF1 上被甲基化 的 ZRE 結合,使得在 EB 病毒潛伏期時被高度甲基化的基因能夠被活化,進而 促使 EB 病毒進入溶裂期 (Bhende, et al., 2004);此特性對於 EB 病毒生活史的轉 換十分重要。其中, Zta 對甲基化序列的選擇性是由 Zta 本身的 Ser186 及 Cys189 所調控,若將 Zta 上的 Cys189 突變為 Ser 會造成 Zta 無法與甲基化的 ZRE 結合,病毒則因此無法順利進入溶裂期 (Bhende et al., 2005; Karlsson et al., 2008)。Zta 也能與溶裂複製起始點 (oriLyt) 上的 ZRE 結合,以促進病毒基因體 的複製 (Fixman, et al., 1992)。除此之外,Zta 可藉由與宿主蛋白質相互作用,調 控其轉錄活性,如 Zta 與 Ku80 結合後可增進 Zta 對 BHLF1 啟動子的活化 (Chen et al., 2011);或其與 CREB-binding protein (CBP) 結合可提升組蛋白乙醯轉 移酶 (histone acetyltransferase) 的活性,進而影響轉錄作用 (Chen et al., 2001a)。
Zta 也會與 DNA 修補機制相關的蛋白質 53BP1 結合,進而增進病毒的複製能力 (Bailey et al., 2009)。另一方面,細胞內存在著能夠抑制 Zta 轉錄活性的蛋白質,
如 B 細胞特有的轉錄因子,Oct-2,其與 Zta 結合後會抑制 Zta 活化溶裂期基因 的能力,使病毒以潛伏期存在於 B 細胞中 (Robinson et al., 2012)。
Rta 蛋白質
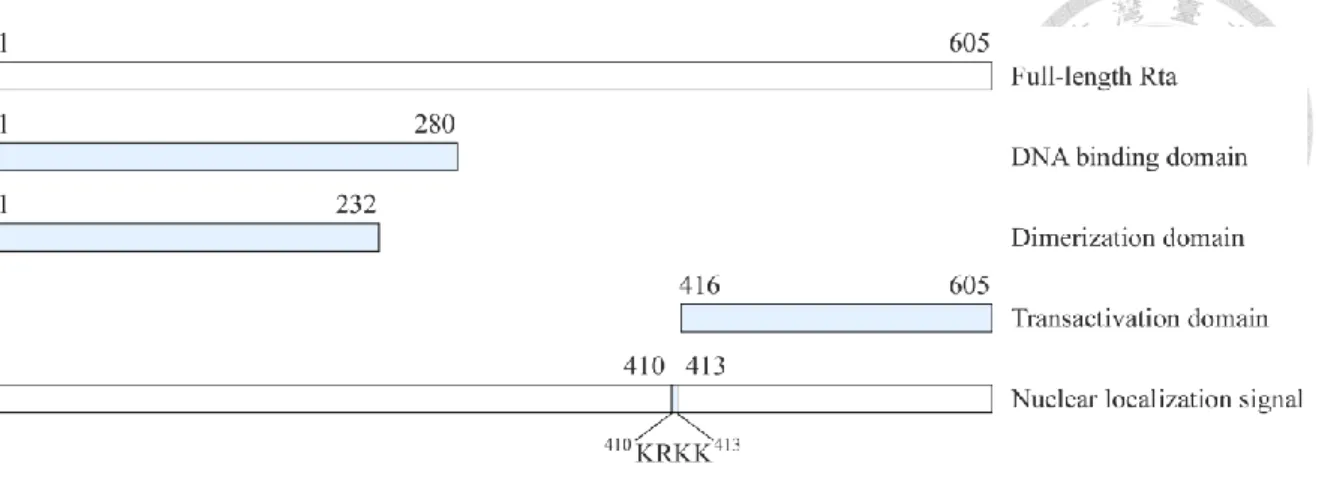
Rta 蛋白質是由 605 個胺基酸所組成,其 N 端為 DNA 結合區 (1-280 胺基 酸),C 端為轉錄活化區 (416-605 胺基酸),在第 1 至 232 個胺基酸的位置為形 成同型二聚體 (homodimer) 的區域 (Manet et al., 1991),且其具有進核序列 (nuclear localization signal,NLS),為 410KRKK413 (Hsu et al., 2005) (圖 1-5)。Rta 主 要藉由結合至啟動子上的 Rta-respoonse element (RRE) 而促進下游基因的表現。
標準的 RRE 序列模式為 GNCCN9GGNG,其中 N 表示任何鹼基,N9 則表示 C 與 G 之間須包含九個核苷酸 (Chen et al., 2005; Gruffat and Sergeant, 1994)。藉由 辨識這段保守性序列,Rta 會形成同型二聚體結合在 RRE 上 (Manet, et al., 1991),
進而活化下游基因 (Gruffat et al., 1990; Gruffat and Sergeant, 1994)。EB 病毒的 BHRF1 是最早被發現能藉由 RRE 序列而被 Rta 活化的基因 (Hardwick et al., 1988),之後發現 Rta 也能結合至 BMRF1、BMLF1、BHLF1 及 BALF2 等 EB 病 毒基因的 RRE 上,活化其基因的表現 (Gruffat et al., 1992; Holley-Guthrie, et al., 1990; Hung and Liu, 1999; Quinlivan, et al., 1993)。
Rta 轉錄活性的調控
Rta 可與其他蛋白質相互作用以調控下游基因。例如 Rta 與 CREB-binding protein (CBP) 的結合能增強 Rta 本身的轉錄活性 (Swenson et al., 2001)。Rta 能經 由宿主蛋白質 E2F 及 upstream stimulatory factor (USF) 的協助而活化 BALF5 啟動子 (Liu, et al., 1996a; Hung et al., 2015);往後研究證明其亦能與 TAF4 結合以
8
活化一些不含 TATA box 的啟動子,包括 BALF5、BNLF1 和人類雄性激素受體 (human androgen receptor) 的啟動子 (Yang and Chang, 2013)。此外,Rta 也能與宿 主蛋白質 MBD1-containing chromatin-associated factor 1 (MCAF1) 及 Specific protein (Sp1) 形成 Rta-MCAF1-Sp1 複合體,間接性地結合至含有 Sp1/Sp3 結合 序列的 Rta 啟動子 (Rp),或是具有 Sp1 結合序列的宿主基因啟動子上,以活化 自身或宿主基因的表現 (Chang et al., 2005)。Rta 與 Zta 也可藉由協同作用 (synergy) 活化病毒基因的表現 (Cox et al., 1990; Liu and Speck, 2003; Quinlivan, et al., 1993),本實驗室先前的研究證明,Rta 是透過中介者 MCAF1 的連結,與 Zta 形 成 Rta-MCAF1-Zta 複 合 體 共 同 結 合 在 ZRE 序 列 上 以 產 生 協 同 活 化 作 用 (Chang, et al., 2010);宿主蛋白質 Ran-binding protein M (RanBPM) 也可作為 Rta 與 Zta 的中介蛋白質,此 Rta-RanBPM-Zta 複合體可結合至病毒基因如 BHLF1 和 BHRF1 基因的 ZRE 上,活化其轉錄活性 (Yang et al., 2015)。而 Rta 透過 MCAF1 與 ATF2 所形成的 Rta-MCAF1-ATF2 複合體也可活化 Zta 基因表現,並 可結合於 BMRF2 啟動子上的 AP-1 典型結合序列 (Lin et al., 2014)。Rta 除了可 透過與宿主蛋白質結合而啟動下游基因的表現,其也能活化 p38、mitogen-activated protein kinase (MAPK)、Jun amino-terminal kinases (JNK) 或 extracellular-regulated kinase (ERK) 等訊息傳導路徑,促使轉錄因子 ATF2 磷酸化使之能結合在 Zta 啟 動子 (Zp) 上,進而活化 Zta 的表現 (Adamson et al., 2000; Lee et al., 2008)。
許 多 研 究 指 出 , Rta 的 功 能 可 受 到 後 轉 譯 修 飾 的 調 控 。 後 轉 譯 修 飾 (post-translational modifications, PTMs) 是指蛋白質在轉譯合成之後所發生的共價 鍵結化學修飾,其能藉由改變蛋白質的化學性質、結構或代電性等特性而影響蛋 白質的功能,其中包括甲基化、乙醯化、磷酸根修飾、泛素修飾 (ubiquitination) 及 SUMO 修飾 (sumoylation) (Seo and Lee, 2004)。Rta 蛋白質的 Lys19、Lys213 和
Lys517 可 受 到 SUMO-1 的 修 飾 而 使 Rta 的 轉 錄 活 性 提 升 (Chang et al., 2004a),而人類的 SUMO E3 連接酶 (E3 ligase) PIAS1、PIASxα 和 PIASxβ 能穩 定 Rta 的 SUMO-1 修飾同時促進其轉錄活性 (Chang, et al., 2004a; Liu et al., 2006)。此外,宿主蛋白質 RanBPM 的過量表現也會增加 Rta 被 SUMO-1 修飾 的程度,進而增強 Rta 對 BMLF1 及 P21 的轉錄活化 (Chang et al., 2008)。Rta 蛋 白 質 上 的 Lys426 、 Lys446 、 Lys517 和 Lys 530 則 會 被 SUMO-2/3 修 飾 (Heilmann et al., 2010),而在 B95-8 EB 病毒株中所缺少的蛋白質 LF2 (Parker et al., 1990) 可與 Rta 結合,增加 Rta 被 SUMO-2/3 修飾的程度 (Calderwood et al., 2008; Calderwood et al., 2007)。本實驗室先前的研究發現,屬於 SUMO-targeted ubiquitin ligase (STUbLs) 家 族 的 RING-finger protein 4 (RNF4) 會 辨 認 被 SUMO-2 修飾的 Rta,並對 Rta 進行泛素化修飾而降低其穩定性,降低 Rta 對下 游基因的轉錄活性 (Yang et al., 2013)。另外,在 EB 病毒溶裂期啟動約 12 小時 之後,可觀察到 Rta 上有磷酸化的修飾,但其功能目前仍尚未釐清 (Zacny et al., 1998)。
Rta 大多位於細胞核內以行使其轉錄活性,但仍有少量的 Rta 存在於細胞質 中 (Cox, et al., 1990)。Rta 上含有進核序列 (NLS),若將 Rta 的 NLS 突變則 Rta 無法進入細胞核,其將不能活化 BGLF5 的啟動子,然而此突變株卻依然能活化 BRLF1 及 BZLF1 啟動子,並且在含有 EB 病毒的細胞株中具有活化溶裂期的能 力 (Hsu, et al., 2005)。另外,LF2 能改變 Rta 的分布使其由細胞核移動至細胞質,
因而導致 Rta 的轉錄活性下降,然而此抑制作用與前述中 LF2 所造成的 Rta 的 SUMO-2/3 修飾並無關聯 (Heilmann, et al., 2010)。
10
Rta 參與細胞生理途徑
除了活化 EB 病毒本身的下游基因,Rta 也可調控宿主的基因表現,其可間 接地活化宿主的 c-Myc 啟動子 (Gutsch et al., 1994)、蛋白質激酶 c-Mer (Li et al., 2004a) 、 脂 肪 酸 合 成 酶 (fatty acid synthase, FAS) (Li et al., 2004b) 以 及 phosphatidylinositol-3 kinase (PI3K) 路徑 (Darr et al., 2001) 和 ERK 路徑 (Lee, et al., 2008),這些信號途徑的活化大多對於維持 EB 病毒的溶裂期十分重要。Rta 具 有調控細胞週期的能力,若將含有 EB 病毒的 Akata 細胞誘導進入溶裂期,可觀 察到 Rta 會與細胞週期的關鍵調控子 Rb (retinoblastoma protein) 相互結合,造成 Rb 將原本結合的轉錄因子 E2F1 釋出,使細胞週期重新啟動 (Zacny, et al., 1998)。若將 Rta 表現於與接觸性抑制的纖維母細胞 (contact-inhibited fibroblast) 或人類骨癌 (osteosarcoma) 細胞株中,則會偵測到 E2F1 與 E2F4 增加,且 Rb、
p107、及 p130 減少,使得細胞重新進入 S phase (Swenson et al., 1999)。此外,
Rta 可活化 EB 病毒促轉型基因 latent membrane protein 1 (LMP-1) (Chang et al., 2004b),LMP-1 為協助原代 B 細胞 (primary B cell) 轉化及建立 EB 病毒潛伏期 的關鍵蛋白質,其能在 EB 病毒潛伏的細胞中促進干擾素 (interferone, IFN) 的生 成,協助病毒潛伏期的建立與維護 (Xu et al., 2006; Zhang et al., 2004)。Rta 也可藉 由直接與 RRE 結合而活化 Decoy receptor 3 (DcR3) 基因 (Ho et al., 2007),DcR3 屬於腫瘤壞死因子受體家族 (tumor necrosis receptor superfamily),常過度表現於不 同類型的惡性腫瘤中;在 EB 病毒感染的淋巴癌細胞株中,Rta 能活化 DcR3 以 抑制細胞凋亡,並避免 EB 病毒受免疫系統攻擊 (Ho, et al., 2007),而在 EB 病毒 感染的鼻炎癌細胞中,Rta 及 LMP-1 會透過 PI3K 及 nuclear factor-kappaB (NF-κB) 活 化 DcR3 進 而 抑 制 細 胞 凋 亡 , 此 現 象 有 助 於 EBV 相 關 癌 細 胞 (EBV-associated cancer cells) 躲避宿主免疫系統而獲得生存優勢,並增強了腫瘤細
胞的遷移 (migration) 及侵襲 (invasion),因而增加了癌症轉移 (metastasis) 的機率
(Ho et al., 2009)。另一方面,Rta 可藉由增加 E2F1 結合至干擾素調節因子 3 (interferon regulatory factor 3, IRF-3) 的啟動子上,進而抑制 IRF-3 的表達,削弱 干擾素的產生 (Zhu et al., 2014)。Rta 也能經由活化 ERK 路徑而促進自噬作用 (autophagy),並影響病毒的溶裂期進程 (Hung et al., 2014)。
五、 粒線體 (mitochondria) 的生理重要性
粒線體 (mitochondria) 為一雙層膜胞器,普遍存在於真核生物的細胞質中 (Henze and Martin, 2003)。其直徑範圍為 0.5 至 1.0 微米 (μm),不同真核細胞種類 中的粒線體在大小、數量和外觀等方面上皆有不同,然而所有的粒線體結構皆相同,
其是由粒線體外膜 (outer membrane)、粒線體內膜 (inner membrane)、膜間腔 (intermembrane space) 以及基質 (matrix) 所組成。粒線體為半自主胞器,具有自 身獨特的的 DNA,稱為粒線體 DNA (mitochondrial DNA, mtDNA),其為 16 kb 的 雙股環狀基因組,共包含 37 種基因,可表現 13 種蛋白質、22 種粒線體 tRNA,
以及 2 種 rRNA (Shadel and Clayton, 1997; Shoubridge, 2002)。其可合成粒線體內 少數呼吸作用所不可或缺的蛋白質 (Burger et al., 2003; Wallace, 2005)。
粒線體為細胞內合成三磷酸腺苷 (adenosine triphosphate, ATP) 的主要場所,
供應了大部分細胞活動所需的能量,因此有「細胞動力工廠」 (the powerhouse of the cell) 之稱。除了為細胞供應能量之外,粒線體也參與了其他許多生理途徑,例如 細胞信息傳遞 (signal transduction) (Bossy-Wetzel et al., 2003)、細胞老化 (aging) (Chan, 2006; Wallace, 2005)、細胞凋亡 (apoptosis) (Gradzka, 2006; McBride et al., 2006)、抗病毒反應 (antiviral response) (Seth et al., 2005) 等過程 (圖 1-6),並擁有 調控細胞代謝、細胞週期與細胞發育的能力 (McBride, et al., 2006)。若粒線體功能
12
失調 (dysfunction) 將影響細胞內許多生理功能,受損的粒線體會影響生物合成路 徑,並製造過量的活性氧化物 (reactive oxygen species) 以及活化細胞死亡途徑。
目前粒線體失調已被報導與許多疾病相關,例如:癌症、神經肌肉退化性疾病、
阿茲海默症 (Alzheimer’s disease) (Yao et al., 2009; Eckert et al., 2008; Manczak et al., 2006)、帕金森氏症 (Parkinson’s disease) (Saiki et al., 2012)、病毒性肝炎、心臟 疾病 (cardiac disease) (Lesnefsky et al., 2001)、以及粒線體疾病 (mitochondria disease/disorder) (McFarland et al., 2007) 等。
粒線體藉氧化磷酸化合成 ATP
在真核細胞中,儘管有一些 ATP 是經由糖解作用 (glycolysis) 在細胞質中產 生,絕大部分的 ATP 都是由粒線體以氧化磷酸化 (Oxidative phosphorylation) 合 成,其為最高效率的產能方式。氧化磷酸化期間,電子從電子供體轉移至最終的 電子受體,分子氧,期間所釋放的能量用於合成 ATP。這些氧化還原反應在一系 列 位 於 粒 線 體 內 膜 上 的 蛋 白 質 複 合 體 的 參 與 下 完 成 , 其 中 包 含 電 子 傳 遞 鍊 (electron transport chain, ETC)。此五種主要的蛋白質複合體為 complex Ⅰ、Ⅱ、
Ⅲ、Ⅳ、Ⅴ。在電子傳遞過程中,電子轉移所產生的能量可被 complex Ⅰ、Ⅲ、
Ⅳ 用於將質子泵入粒線體膜間腔,粒線體內膜兩側產生質子梯度後使質子通過 complex Ⅴ,即 ATP 合成酶 (ATP synthase),從膜間腔回到粒線體基質,其電勢 能將 ADP 和磷酸合成為 ATP (Mitchell, 1967; Mitchell and Moyle, 1967) (附錄 2)。
粒線體參與細胞凋亡 (apoptosis)
細胞中有兩個主要途徑可誘發細胞凋亡,一為外在途徑 (extrinsic pathway),
另一為內在途徑 (intrinsic pathway) (Sanfilippo and Blaho, 2003; Borutaite, 2010)。其
中內在途徑為壓力信號直接影響粒線體膜的通透性所引發,主要由粒線體所調 控。當凋亡訊號產生,將會刺激促細胞凋亡的蛋白質的表達,如 Bax 及 Bak,其 導致粒線體內的細胞色素 C (cytochrom C, Cyt-C) 釋放 (Castanier and Arnoult, 2010; Liu et al., 1996b)。而 Cyt-C 會與細胞質中的 Apaf-1 以及 caspase 9 共同組 成凋亡體 (apoptosome),接著引發下游一連串的 caspase 級聯反應,進而促使細 胞凋亡。除了 Cyt-C,Smac/DIABLO 和核酸內切酶 G (endonuclease G) 也可作為 caspase 的活化物;另外有抑制細胞凋亡的蛋白質如 Bcl-2 和 Bcl-XL 則會阻斷此 細胞凋亡的過程 (Mayer and Oberbauer, 2003) (附錄 3)。
粒線體所涉及的細胞凋亡可由 B cell lymphoma-2 (Bcl-2) 家族蛋白嚴格控管 (Danial et al., 2010; Soriano and Scorrano, 2010)。其上具有 Bcl-2 同源性結構區 (Bcl-2 homology, BH domain),由 N 端至 C 端依序為 BH4-BH3-BH2-BH1;而在 C 端則有疏水性的穿膜 (transmembrane, TM) 結構區,可將蛋白質錨定於粒線體 外膜 (Scorrano and Korsmeyer, 2003)。BH1 和 BH2 結構區主要負責抗凋亡 (antiapoptotic) 活性和 Bcl-2 家族蛋白質之間的多聚化 (multimerization),BH3 結 構區負責促凋亡 (proapoptotic) 活性,而 BH4 結構區則為 Bcl-2 和 Bcl-XL 蛋白 質的抗凋亡活性所需 (Danial, et al., 2010; Scorrano and Korsmeyer, 2003)。大部分的 抗凋亡蛋白質具有多結構區,包括全部四個 BH 結構區和 TM 結構區;相反地,
促凋亡蛋白質則可具有其中三個結構區 (BH1 至 BH3) 或僅具單獨結構區 BH3 (Galluzzi et al., 2008)。這些 Bcl-2 家族蛋白質可調控粒線體膜電位 (mitochondrial membrane potential, MMP),而 MMP 為細胞凋亡末期的標誌,可決定細胞是否走 向死亡 (Crompton, 2000; Waterhouse et al., 2002; Zamzami and Kroemer, 2003)。
14
粒線體與免疫系統的關聯
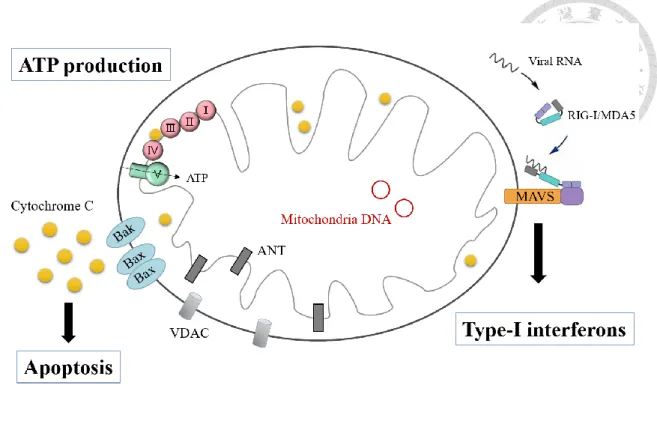
先天免疫 (innate immunity) 是宿主對抗病原體感染的第一道防線。近期的研 究認為,粒線體在先天免疫的訊息傳導路徑中扮演著「抗病毒平台 (antiviral platform) 」的角色,許多重要的訊息分子需被招募至粒線體上組裝成抗病毒複合 體,進而引發下游轉錄因子的活化及干擾素 (interferon, IFN) 產生 (Arnoult et al., 2011; Zemirli and Arnoult, 2012)。當宿主受到病毒感染時,細胞內的病毒遺傳因子 會被模式辨識受體 (pattern recognition receptors, PRRs) 所辨認 (Akira et al., 2006),粒線體與其中的 RIG-I like receptor (RLR) 有關,包括 RIG-I 和 MDA5。
位於細胞質的 RLRs 辨認到外來遺傳物質時會因此被活化並易位至粒線體,與粒 線體外膜上的抗病毒信息蛋白質 (mitochondrial antiviral signaling, MAVS) 相互作 用 (Seth, et al., 2005)。MAVS 接著招募許多下游因子如:TRF3、TBK1 和 IKKε,
共同形成抗病毒複合體來促進干擾素調控因子 IRF3 的活化,使第一型干擾素產 生,引起抗病毒作用 (Kawai and Akira, 2008; Seth et al., 2006) (附錄 4)。
粒線體自噬 (mitophagy)
粒線體為一動態的胞器,其融合 (fusion) 與分裂 (fission) 的循環週期被認為 可調控粒線體網絡的健康;若粒線體遭受外來壓力或受損,其將被標記並被選擇 性地降解,以防止其與其它健康粒線體聚集融合。為了維持粒線體的健康,細胞 發展出一質量控管機制,可藉由粒線體自噬 (mitophagy) 此途徑,將受損粒線體 經溶酶體 (lysosome) 隔離、隨後降解 (附錄 5)。粒線體自噬可被粒線體相關激酶 PINK1 和 E3 泛素連接酶 (E3 ubiquitin ligase) parkin 所調控。在健全的粒線體 上,PINK1 會被粒線體的蛋白酶降解;而當粒線體損傷時 PINK1 會累積於其外 膜,並招募 parkin 至外膜表面 (Narendra et al., 2008; Narendra et al., 2010),parkin
隨後泛素化些許粒線體蛋白質,進而引發粒線體自噬信號以移除受損粒線體 (Chan et al., 2011; Ordureau et al., 2014)。
六、 粒線體導向序列 (mitochondrial targeting signal, MTS)
細胞中 99% 以上的粒線體蛋白質是由細胞核基因所表現,其 N 端胺基酸序列中 含有靶向信號 (targeting signal),稱為前序列 (presequence),可與粒線體外膜上的受 體蛋白質相互作用。受體蛋白質為 TOM 複合體 (translocon of the outer mitochondrial membrane, TOM complex) 的一部分,此多蛋白複合體包含 Tom20、Tom22、Tom40、
Tom70 等 (Neupert and Herrmann, 2007)。粒線體前蛋白 (preprotein) 可藉由其前序 列先後與粒線體受體蛋白 Tom20 和 Tom22 結合 (Brix et al., 1997; Saitoh et al., 2007)。這些受體將前序列轉移至 Tom40 易位子 (translocon),其形成的通道可使 前蛋白從 N-到 C-末端 (N→C) 的線性模式通過外膜 (Model et al., 2008)。在膜間 腔中,前序列藉由驅動蛋白 PAM (presequence translocase associated motor, PAM) 的幫忙,與 TIM 複合物 (translocon of the inner mitochondrial membrane complex, TIM complex) 中的 Tim23 相互作用,而後前蛋白得以通過內膜 (Neupert and Herrmann, 2007; Truscott et al., 2001)。前蛋白進入粒線體基質後,其前序列會因粒 線體肽酶 MPP (mitochondrial processing peptidase, MPP) 的作用而被截切釋放 (Teixeira and Glaser, 2013),經修飾過後的蛋白質將折疊為正確的三級結構,最後 呈現其功能 (Schmidt et al., 2010) (附錄 6)。
粒線體前序列多位於前蛋白的 N 端,可為 10 個至 80 個胺基酸的長度。研 究顯示,粒線體前序列所編碼而成的粒線體導向序列 (mitochondrial targeting signal, MTS),其胺基酸序列並不具有典型的保守性,因此 MTS 並無共有序列 (consensus sequence)。然而,這些序列仍有一些共有的物理或化學特性,例如其主
16
要由帶正電荷的胺基酸組成,尤其為 arginine,同時幾乎不存在帶負電荷的個別胺 基酸 (Huang et al., 2009; von Heijne et al., 1989)。此外,此序列傾向形成兩親性 (amphipathic) 的 α 螺旋體,螺旋體的一側為疏水性 (hydrophobic),另一側則為正 電荷殘基組成 (附錄 7, A)。大鼠乙醛去氫酶 (aldehyde dehydrogenase, Aldh2) 的 前序列中含有一 α 螺旋體可與受體 Tom20 的細胞質部分結合,其由六個胺基酸
14RLSRLL19 作為辨認 Tom20 之核心 (附錄 7, B),經粒線體前序列比對之後揭露 了 Tom20 辨認保守模式為 σφχβφφ,或更具廣泛性的 φχχφφ,其中 σ 表示親水 性胺基酸,φ 表示一較巨大的疏水性胺基酸,χ 表示任何胺基酸,β 表示鹼性胺 基酸 (Obita et al., 2003)。經由 Tom20 及其結合物的共同三維結構解析,可得知 前序列的兩親性螺旋體會位於一廣且淺的結合凹槽中,其螺旋體疏水性的一側與 Tom20 的結合槽接觸,而帶正電的一側則與結合槽邊界的負電荷相互作用 (Abe et al., 2000; Saitoh, et al., 2007) (附錄 7, C)。
七、 病毒對粒線體功能的調節
為了在細胞中生存及複製,病毒需要抵抗細胞中的防禦和免疫系統,它們也 需要能量來進行複製及由細胞脫逃。只要在宿主細胞中,病毒便會調節各種細胞 信號通路及胞器,其中包含了粒線體。不同於粒線體蛋白質,大多數的粒線體導 向病毒蛋白質並沒有典型可被截切的 N 端前序列,然而其大多呈現兩親性的 α 螺旋體構造,位於蛋白質上不同的位置。病毒蛋白質將藉由此 MTS 而易位至粒 線體,從許多方面影響粒線體的功能以利病毒的生存 (圖 1-7)。
1. 病毒在宿主細胞中引發氧化壓力 (oxidative stress)
粒線體的有氧呼吸鏈為細胞中活性氧化物 (reactive oxygen species, ROS) 最
主要的來源。在正常的狀態下,電子傳遞鍊的複合物之間幾乎不會有電子洩漏 (leakage)。然而在壓力條件下,一小部分的電子會過早地遺漏至氧 (Niizuma et al., 2009),將導致超氧化物 (superoxide, O2-∙) 的形成。
一些病毒可引起宿主的氧化壓力,間接或直接地幫助病毒生存。人類腺病毒-5 (human-adenovirus-5, HAdV-5) 可引發溶體的細胞溶解酶 (lysosomal cathepsins) 釋放,使粒線體膜破裂而促進 ROS 的產生與釋放 (McGuire et al., 2011)。C 型肝 炎病毒 (hepatitis C virus, HCV) 的核心蛋白 (core protein) 也可誘導細胞產生氧化 壓力並改變細胞凋亡路徑 (Nishina et al., 2008)。而 B 型肝炎病毒 (hepatitis B virus, HBV) 的 X 蛋白可在病毒複製的不同階段發揮功能,導致氧化壓力的增高 或下降,利於病毒在宿主細胞裡生存 (Hu et al., 2011; Kim et al., 2010)。
2. 病毒調節宿主的粒線體膜電位 (mitochondrial membrane potential, MMP) 粒線體膜電位 (mitochondrial membrane potential, MMP) 可提供合成 ATP 的 能量。電壓依賴性陰離子通道 (voltage dependent anionic channel, VDAC) 位於粒線 體外膜,其可形成外膜通道使代謝物可進行穿膜運輸。外界刺激可誘導非特異性 的粒線體滲透性通透孔道 (mitochondrial permeability transition pores, MPTP) 產 生,其是由外膜的 VDAC、 內膜的腺嘌呤核苷酸轉位酶 (adenine nucleotide translocase, ANT) 和基質中的 cyclophilin D (CyP-D) 所組成,可負責維持 MMP。
在一般情況下,MMP 的下降將誘導細胞凋亡,而 MMP 增高則會阻止細胞凋亡 的發生。
許多病毒蛋白質可改變 MMP 以利於病毒於細胞內的存活。HCV 的 p7 蛋 白質為疏水性的膜內嵌蛋白,其位於粒線體 (Griffin et al., 2004),藉由控制陽離子 的膜滲透性促進細胞存活,以利於病毒複製 (Gonzalez and Carrasco, 2003)。人類
18
免疫缺乏病毒 (human immunodeficiency virus, HIV) 的 R 蛋白 (Vpr) 也位於粒 線體,其可與 ANT 相互作用,調節 MPTP,並誘導 Cyt-C 的釋放進而導致細胞 死亡 (Deniaud et al., 2004; Jacotot et al., 2000)。流感 A 型病毒的 PB1-F2 蛋白質 則在粒線體上與 VDAC1 和 ANT3 相互作用,使 MMP 下降而誘導促細胞凋亡 蛋白質的釋放,使細胞死亡 (Chen et al., 2001b; Gibbs et al., 2003)。後期研究則報 導 PB1-F2 也可藉由形成非選擇性的蛋白通道孔洞而使粒線體型態改變,造成 MMP 耗損,最終也導致細胞死亡 (Henkel et al., 2010)。
3. 病毒可調控細胞凋亡 (apoptosis)
病毒會產生 Bcl-2 蛋白質的同源物 (vBcl-2),其可促進或抑制細胞凋亡的發 生,以幫助病毒在宿主細胞中完成生命週期。在初次感染時,vBcl-2 與其他蛋白 質的相互作用可增長宿主細胞的壽命,使病毒後代高效產生;其也使潛伏感染的 病毒可度過轉換期,有利於病毒在細胞中的持久性 (Thomson, 2001)。
許多病毒會產生抗凋亡 Bcl-2 蛋白質的同系物,其偏好定位於粒線體,並可 與其他促凋亡蛋白例如 Bax 相互作用。HAdV-5 所產生的 E1B19K 蛋白質含有類 似 BH1 和 BH3 的結構區,可藉由抑制 Bax 蛋白質的作用使 caspase 的活化中 斷,進而阻擋細胞死亡信號 (Perez and White, 2000)。卡波西肉瘤疱疹病毒 (Kaposi’s sarcoma-associated herpes virus, KSHV) 的 Bcl-2 蛋白質則含有 BH1 和 BH2 結構區,可與單獨具有 BH3 結構區之蛋白質相互作用,具抗凋亡活性 (Flanagan and Letai, 2008)。病毒也會產生促細胞凋亡的 Bcl-2 蛋白質同源物。HBV 產生的 X 蛋白質含有 BH3 結構區,其位於粒線體並可與 VDAC 相互作用,使 MMP 下降而造成細胞凋亡 (Lu and Chen, 2005; Rahmani et al., 2000)。
4. 病毒影響粒線體相關的抗病毒免疫機制
由前述可知,粒線體為抗病毒複合體的組裝提供了一個適當的環境,使 MAVS 得以介導經 RLR 所傳遞的 IFN 誘導路徑 (RLR-mediated IFN induction),可視為 一個抗病毒訊息傳遞的重要樞紐,因此也成為了病毒逃避宿主的免疫反應時主要 目標。HCV 的非結構蛋白質 3 (nonstructural protein 3, NS3) 和非結構蛋白質 4A (NS4A) 會與粒線體 MAVS 蛋白質共同存在,導致 MAVS 斷裂,從而大幅降低 其誘導干擾素反應的能力 (Li et al., 2005; Meylan et al., 2005)。其他病毒如流感 A 型病毒在受感染的人類原發性巨噬細胞中,可將 RIG-I/MDA5 分子易位至粒線體 上,進而調控宿主的抗病毒和凋亡信號,增加病毒的生存能力 (Ohman et al., 2009)。
5. 病毒改變粒線體在細胞內的分布
病毒會改變粒線體在細胞內的分布,可使其集中至病毒附近以提供病毒複製 時的能量所需,或將粒線體於細胞質內封鎖隔離,以避免細胞凋亡的介質釋放。
HBV 的 X 蛋白質會藉由 p38 誘導動力蛋白 (dynein) 活化,使粒線體向細胞核 周邊聚集 (Kim et al., 2007)。HCV 的 NS4A 可單獨或與 NS3 一起累積於粒線 體,並改變粒線體於細胞內的分布 (Nomura-Takigawa et al., 2006)。HIV-1 的感染 會導致粒線體於感染的細胞中聚集 (Radovanovic et al., 1999)。而非洲豬瘟病毒 (African swine fever virus, ASFV) 會造成粒線體聚集於病毒組裝的區域附近,使其 可在病毒釋放時提供能量 (Rojo et al., 1998)。
6. 病毒造成宿主粒線體 DNA 耗竭 (mitochondrial DNA depletion)
哺乳動物粒線體含有一環狀小基因組,其可合成氧化磷酸化的酵素 (Burger, et al., 2003)。由於粒線體為細胞能量的主要來源,並其在抗病毒免疫機制裡扮演著重
20
要角色,粒線體 DNA 的損壞也許可幫助病毒躲避粒線體的抗病毒免疫反應 (Saffran et al., 2007)。哺乳動物細胞於體外感染期間,人類單純疱疹病毒 (Herpes simplex virus 1, HSV-1) 會誘導宿主粒線體 DNA 快速且完全地降解 (Saffran, et al., 2007)。HSV-1 的 UL12.5 蛋白質位於粒線體,而其在沒有其他病毒基因產物 的環境下仍可誘導 DNA 耗竭 (DNA depletion) (Corcoran et al., 2009; Saffran, et al., 2007)。而 HCV 則會誘導活性氧化物和氧化氮的產生,進而引發宿主 mtDNA 的損害 (Machida et al., 2006)。
八、 EB 病毒與粒線體間的關係
許多病毒會調節粒線體功能以利自身的生存,EB 病毒也不例外。EB 病毒在 感染宿主細胞後的 48 小時內會造成氧化壓力的增加,氧化壓力會活化 EB 病毒 的極早期基因 BZLF1,進而誘導病毒溶裂期的發生 (Lassoued et al. 2010);此現象 被 認 為 是 EB 病 毒 致 病 機 制 的 重 要 因 素 , 其 中 包 括 惡 性 轉 化 (malignant transformations) (Gargouri et al. 2009; Lassoued et al. 2008)。極早期蛋白質 Zta 會與 粒線體單股 DNA 結合蛋白 (mitochondrial single-stranded DNA binding protein, mtSSB) 相互作用,在抑制粒線體 DNA 複製的同時促進本身病毒基因的複製 (Wiedmer et al., 2008)。而 EB 病毒的 BHRF1 和 BALF1 皆為 Bcl-2 蛋白質的同 源物 (vBcl-2),其中 BHRF1 蛋白質含有 BH1 和 BH2 結構區,可與單獨具有 BH3 結構區之蛋白質相互作用,具抗凋亡活性 (Flanagan and Letai, 2008);而 BALF1 蛋 白 質 含 有 BH1 和 BH4 結 構 區 , 可 與 Bax-Bak 蛋 白 質 相 互 作 用 (Marshall et al., 1999),其被認為可作為 BHRF1 的拮抗物 (Bellows et al., 2002)。
此外,EB 病毒在感染宿主細胞後,其溶裂期複製的誘導會造成粒線體形態上的改 變 (LaJeunesse et al., 2005)。當 EB 病毒的溶裂期複製發生後,粒線體膜電位會下
降,其微管細胞骨架 (microtubular cytoskeleton) 會大幅增加,並有粒線體聚集的 現象產生且同時易位至細胞核周圍 (LaJeunesse, et al., 2005)。而此機制與 EB 病 毒的極早期蛋白質 Zta 及 Rta 有關,Zta 被認為可引發粒線體的融合,而 Rta 則 會造成粒線體易位至細胞核附近 (LaJeunesse, et al., 2005)。近期研究指出,乳腺癌 的遷移與侵襲與粒線體的動態恆定失調有關,而 EB 病毒的 LMP2A 蛋白質可經 由 Notch 路徑提高粒線體分裂蛋白質 dymanin-related protein 1 (Drp1) 的表現程 度,進而增強胃腺癌和乳腺癌細胞中粒線體的裂變,影響腫瘤細胞的轉移 (Pal et al., 2014)。
九、 研究目的
粒線體對許多細胞功能扮演極重要的角色,例如能量的產生、代謝和細胞死 亡。在受病毒感染的細胞中,許多病毒蛋白質會藉由調節粒線體,以利於病毒自 身的生存與複製。EB 病毒的極早期蛋白質 Rta 為 EB 病毒由潛伏期進入溶裂期 的關鍵因子,其除了具有轉錄因子活性,更參與了 EB 病毒至晚期的生活史;其 也能調控宿主的基因表現,並與許多宿主蛋白質接觸,與 EB 病毒溶裂期的維持 及 B 細胞惡性轉化有很大的關聯。先前研究指出,EB 病毒感染宿主細胞後,其 溶裂期複製的誘導會造成粒線體形態上的改變,而 Rta 則會造成粒線體易位至細 胞核附近 (LaJeunesse, et al., 2005)。本實驗室在先前的研究當中,在電子顯微鏡的 觀察下,發現於病毒潛伏的細胞當中 Rta 蛋白質會集中分佈於粒線體上 (王文宏, 未發表) (附錄 1),使本研究欲探討 EB 病毒中重要的極早期蛋白質 Rta 蛋白質 對於粒線體的影響。首先本研究想釐清 Rta 蛋白質上是否具有粒線體導向序列 (mitochondria targeting sequence,MTS),接著進一步探討 Rta 如何影響粒線體的 功能。
22
第二章、 材料與方法
一、 細胞株
293T 細胞株是由人類胚胎腎臟細胞所分離出來 (Graham et al., 1977),廣泛地 被認為是腎臟上皮細胞,而近期研究則指出其與神經細胞間的密切關係 (Shaw et al., 2002),293-2089 細胞株為穩定轉染 Maxi-EBV 質體 p2089 之人類胚胎腎臟 細胞 (Hettich et al., 2006) ,兩者皆以含 10% 胎牛血清 (Fetal bovine serum, FBS)、100 μg/mL penicillin G (Sigma)、100 μg/mL streptomycin (Sigma) 及 2 mM L-Glutamine 的 Dulbecco’s modified Eagle’s medium (DMEM) 培養液貼壁培養,置 於 37°C、5 % 二氧化碳的環境當中。P3HR1 細胞株為含有 EB 病毒的巴克氏淋 巴癌細胞株 (Klein et al., 1975; Maurer et al., 1970; Hinuma et al., 1967),B95-8 細胞 株是以引起美國起源傳染性單核球增多症的 EB 病毒感染狨猴淋巴細胞所建立 (Miller and Lipman, 1973),兩者皆以含 10% 胎牛血清、100 μg/mL penicillin G (Sigma) 、 100 μg/mL streptomycin (Sigma) 及 2 mM L-Glutamine (Sigma) 的 RPMI-1640 培養液懸浮培養在 37°C、5% 二氧化碳的環境中。
二、 EB 病毒的溶裂期誘導
本研究中所使用的 P3HR1 細胞能以加藥的方式誘導細胞中的 EB 病毒進入 溶裂期,本實驗於培養液中加入 30 ng/mL 12-O-tetradecanoylphobol-13-acetate (TPA) (zur Hausen and Fresen, 1978) 及 3 mM sodium butyrate (SB) (Davies et al., 1991; Luka, et al., 1979) 依實驗需求作用 24 至 48 小時誘導細胞中的病毒進入溶 裂期。而 2089 則以質體 pCMV-Rta 與 pCMV-Zta 共轉染 24 至 48 小時誘導病 毒進入溶裂期。
三、 質體與抗體 如表 1、表 2。
四、 質體 DNA 的萃取
本研究所使用的質體 DNA 使用 Presto Mini Plasmid Kit (Geneaid),方法參照使 用手冊說明。
五、 細菌
本研究中主要使用的大腸桿菌 (Escherichia coli) 為 EPI300,其用於質體的大 量複製,培養於 37°C 含特定抗生素的 Luria Broth (LB) (Bertani, 1951) 中。
六、 細菌轉型作用 (transformation)
首先將大腸桿菌 EPI300 勝任細胞 (comptent cells) (Jensen and Haas, 1963) 與適量 DNA 混合,於冰上作用 15 分鐘,接著以 42°C 熱休克 (heat shock) 處 理 90 秒,之後加入 500 μL LB 培養液,於 37°C 培養箱震盪培養 30 分鐘,最 後取適量菌液均勻塗盤在含有特定抗生素的 LB 固態培養基 (Cohen et al., 1972) 上,於 37°C 培養箱靜置培養隔夜。
七、 細胞轉染 (Transfection)
將 2x105 的 293T 細胞以 2 mL DMEM 培養一天後取適量質體 DNA 及 Turbofect in vitro transfection reagent (Fermentas) (1 μg DNA/1.5 μL Turbofect),以不 含血清的 DMEM 培養液充分混合並靜置於室溫 20 分鐘,而後將混合液加入細 胞中,於 37°C 培養 24 至 48 小時。
24
八、 免疫螢光染色分析 (Immunofluorescence analysis)
本研究用於免疫螢光染色分析的細胞抹片,根據使用的細胞株而有不同製 程。若目標細胞為貼壁細胞,先在培養盤中置入蓋玻片,加入培養液與細胞使細 胞直接貼附其上。製作懸浮細胞的細胞抹片則需先將玻片進行前處理,將蓋玻片 以酸酒精 (3% HCl, 97% ethanol) 浸泡,使用前再以 95% 酒精將酸酒精洗掉,靜 置風乾後以 200 μL poly-L-lysine solution (Sigma P8920) 平鋪於玻片上使玻片帶正 電,作用 10 至 20 分鐘後將液體吸乾,並靜置風乾後備用。收取 P3HR1 或 B95-8 細胞,以 phosphate-buffered saline (PBS) 清洗並以 PBS 回溶 400 μL,平 鋪於 poly-L-lysine 作用過的蓋玻片上約 10 分鐘,使細胞吸附其上,後以 PBS 清 洗三次洗去多餘的細胞。貼附細胞後的蓋玻片以 4% paraformaldehyde 於 4°C 作 用 30 分鐘以固定細胞,接著以 PBS 清洗三次後以含 0.1% Triton X-100 的 PBS 處理 10 分鐘,增加細胞膜的通透性。PBS 清洗三次後,以 1% Bovine Serum Albumin (BSA) 在室溫下反應 1 小時,進行 blocking。將一級抗體以 1:50-1:200 的比例稀釋於 1% BSA 中平鋪於玻片上,室溫下作用 1 小時。後以 PBS 清洗三 次,接著以二級螢光抗體以 1:200 比例稀釋於 1% BSA 中,平鋪於玻片上在室溫 作用 45 分鐘,另取 5 μg/ml 4,6-diamidino-2-phenylindole (DAPI) 直接加於玻片上 作用 15 分鐘,將細胞核進行染色。以 PBS 清洗四次洗去多餘抗體,完成細胞的染 色。最後在載玻片上滴 10 μL 的 Citifluor Antifadent Mountant Solution (Agar Scientific, R1302),將蓋玻片以 45°角傾斜方式蓋上,吸去多餘水分後以指甲油進 行封片。玻片製作完成後使用共軛焦螢光顯微鏡 (Zeiss LSM780, Germany) 進行觀 察與拍攝。(Chang, et al., 2004a)
九、 粒線體的分離分析
將 P3HR1 細胞或 293-2089 細胞誘導進入溶裂期,或在 293T 細胞中轉染入 不同的建構質體後,於 24 小時後收取 1.2x106 至 2x106 的細胞,以 PBS 清洗 兩次,將五分之一的細胞取出作為 total lysate,其他部分則做粒線體的分離,可將 細胞區分為細胞質部分 (cytosol fraction) 及粒線體部分 (mitochondrial fraction)。
粒 線 體 的 分 離 是 利 用 Mitochondria Isolation Kit for Cultured Cells (Thermo Scientific),方法參照使用手冊說明。首先,Reagent A 及 Reagent C 在使用前須 先加入 Thermo Scientific Halt Protease Inhibitor Cocktail, EDTA-Free (100X),收取 下來的細胞以 200 μL Mitochondria Isolation Reagent A 均勻回溶,並於冰上作用 2 分鐘,而後加入 2.5 μL 的 Mitochondria Isolation Reagent B,並強烈震盪回溶液 5 秒。接著於冰上作用 5 分鐘,期間細胞回溶液每分鐘皆以強烈震盪處理 5 秒鐘。
之後加入 200 μL 的 Mitochondria Isolation Reagent C,上下倒置數次,均勻混勻 後以 700 xg 於 4°C 下離心 10 分鐘,而後轉移上清至新的離心管,以 12,000 xg 於 4°C 下離心 15 分鐘,上清即為細胞質部分,而沉澱物為粒線體部分。沉澱的 粒線體以 Reagent C 清洗兩次,去除上清後即可於 80°C 下保存或用於往後的實 驗。
十、 西方點墨法分析 (Western blot analysis)
樣 本 中 加 入 sample buffer (62.5 mM Tris-HCl pH 6.8, 2% SDS, 5%
β-mercaptoethanol, 10% glycerol, 2 mM EDTA, 0.01% bromophenoblue) 將蛋白質樣 本 溶 出 , 並 置 於 95°C 作 用 五 分 鐘 。 接 著 將 樣 本 注 入 配 置 好 的 8-13% 的 SDS-polyacryamide 膠體中,以 168 V 電壓進行蛋白質分離電泳。完成後將膠體 以 Nitrocellulose 轉印膜 (Whatman) 覆蓋並用濾紙夾住 ,於轉印液 (25 mM
26
Tris-Hcl, 5.5% glycine, 20% methanol, pH 7.3) 中以 94 V、 400 mA 轉印 1.5 小 時。完成轉印後將轉印膜浸於含有 5% 安佳脫脂奶粉的 TBST (20 mM Tris-HCl, 150 mM NaCl, 0.05% Tween 20, pH 7.5) 中進行 blocking 20 分鐘以避免非專一性 結合。接著以 TBST 洗淨後加入一級抗體於室溫下反應 1 小時。反應結束後 以 TBST 清洗三次,每次 3 分鐘,再加入接有 horseradish peroxidese (HRP) 的二級 抗體,室溫下反應 30 分鐘。而後以 TBST 清洗三次將多餘抗體洗去,加入受質 chemiluminescent HRP substrate (Millipore) 反應 3 分鐘後呈色,最後以 X-ray 底 片偵測呈色訊號。
十一、 免疫沈澱法 (Immunoprecipitation, IP)
將表現特定蛋白質的質體 DNA 轉染至 293T 細胞中,於 24 小時後收取細 胞。以含有 0.5% IGEPAL 的 PBS 打破細胞,以 13,800 xg 在 4°C 下離心後取上 清液,加入 anti-FLAG-M2 agarose (Sigma) 於 4°C 平衡作用 1 小時 30 分鐘,接 著使用等體積含有 0.5% IGEPAL 的 PBS 清洗膠體三次,最後以西方點墨法偵測 anti-FLAG-M2 agarose 上所吸附的蛋白質。
十二、 加藥處理
為了誘導粒線體損害,將 293T 細胞加入 20 μM 羰基氰化物間氯苯腙 (carbonyl cyanide 3-chlorophenylhydrazone,CCCP) (Sigma, C2759),共同培養 1 至 24 小時。
十三、 ATP 測定分析 (ATP Determination assay)
將 293T 細 胞 培 養 於 24-well 培 養 盤 , 轉 染 質 體 pGL3-CMV 或
pCMV-Rta,48 小時後離心收取細胞,加入 30 μL lysis buffer (25 mM Tris-phosphate, pH 7.8, 2 mM EDTA, 2 mM DTT, 10% glycerol, 1% Triton X-100),震盪後於冰上反 應 5 分鐘以打破細胞。反應完成後以 13,800 xg 在 4 °C 下離心五分鐘,離心完成 後取10 μL 上清液至冷光分析盤中,接著加入 ATP Determination Kit (Invitrogen, A22026) 中 的 冷 光 反 應 溶 液 90 μL (0.5 mM D-luciferin, 1.25 μg/mL firefly luciferase, 25 mM Tricine buffer, pH 7.8, 5 mM MgSO4, 100 μM EDTA, 1 mM DTT),
最後立即放入冷光受質儀 (Orion II, Berthod, Bad Wildbad, Germany) 測量冷光數 值。其數值大小即代表樣本中的 ATP 含量。
28
第三章、 結果
一、 Rta 蛋白質存在於粒線體
本實驗室在先前的研究當中,利用電子顯微鏡觀察 Rta 蛋白質在 EB 病毒潛 伏期時於細胞內的分佈位置 (王文宏, 未發表) (附錄 1)。實驗當中以金粒子標定 Rta 蛋白質,結果發現在 EB 病毒潛伏期的 P3HR1 細胞中,Rta 蛋白質的分佈 除了集中於核仁外,也集中於粒線體 (附錄 1)。
首先,為佐證此結果,本研究利用免疫螢光染色分析,觀察在具病毒潛伏的 細胞中 Rta 蛋白質與粒線體的分佈情形。將含有 EB 病毒的 P3HR1 細胞 (圖 3-1 A) 和 B958 細胞 (圖 3-1 B) 固定於玻片上後,對於 Rta 蛋白質和在此作為粒線 體 marker 的 Tom20 蛋白質進行免疫螢光染色,螢光圖結果中顯示,在 EB 病毒 潛伏期時仍有微量的 Rta 蛋白質表現 (圖 3-1 b, g),細微的綠色螢光在局部放大圖 中清楚可見與紅色螢光重合,顯現黃色點狀螢光,結果顯示在病毒潛伏的細胞中,
潛伏期之 Rta 蛋白質與 Tom20 在細胞質中位於相同位置。之後本研究也接著檢 驗溶裂期的 Rta 蛋白質於細胞中的位置,將含有 EB 病毒的 P3HR1 細胞以 SB 和 TPA 作用 48 小時,誘導細胞中的病毒進入溶裂期,在細胞固定後對於 Rta 蛋 白質和粒線體 marker Tom20 進行免疫螢光染色,結果顯示 Rta 蛋白質與 Tom20 在細胞質中影像重合 (圖 3-2 B);於另一株也具 EB 病毒潛伏的 293-2089 上皮 細胞,以共轉染質體 pCMV-Rta 和 pCMV-Zta 的方式,誘導病毒進入溶裂期,並 以質體 pGL3-CMV 作為控制組,在 48 小時後收取細胞進行免疫螢光分析,結果 顯示 Rta 與 Tom20 分布於相同位置 (圖 3-2 A)。綜合以上之免疫螢光分析結 果,可推測 Rta 蛋白質無論在 EB 病毒潛伏期或溶裂期時皆會位於粒線體。
另一方面,本研究也利用粒線體分離的分析方法,將具 EB 病毒潛伏的 293-2089 上皮細胞,以共轉染質體 pCMV-Rta 和 pCMV-Zta 的方式,誘導病毒進 入溶裂期,並以質體 pGL3-CMV 作為控制組,在 48 小時後收取細胞進行粒線體 分離分析 (圖 3-3 A);而另一株也含有 EB 病毒的 P3HR1 細胞,以 SB 和 TPA 處理 48 小時誘導病毒溶裂循環,收取細胞後進行粒線體分離分析 (圖 3-3 B)。此 外,也在 293T 細胞中轉染 pCMV-Rta,48 小時後收取細胞,同樣進行粒線體分 離 (圖 3-3 C)。分離後可將細胞分為細胞質和粒線體部分,接著以西方點墨法觀 察 Rta 蛋白質的表現。結果顯示,Rta 蛋白質在細胞質與粒線體中皆有表現,證 實 Rta 確實會位於粒線體,且在無病毒潛伏的細胞株內仍有相同的分佈情況。
二、 Rta 蛋白質會與 Tom20 結合
Tom20 為 粒 線 體 外 膜 蛋 白 , 也 為 粒 線 體 上 的 蛋 白 質 導 入 受 體 (import receptor)。已知多數的粒線體內蛋白質皆為由細胞核 DNA (nuclear DNA) 在細胞 質產生,而典型的粒線體蛋白質上具有粒線體導向序列 (mitochondria targeting sequence,MTS),在蛋白質輸送往粒線體後 MTS 將與粒線體膜上的 TOM 蛋白 複合體接觸,進而導入粒線體。因此本研究欲探討 Rta 是否會與 Tom20 結合。
首先將質體 pTag2A 或 pTag-Rta 分別轉染至不含病毒的 293T 細胞中,培養 24 小時後收取細胞,先以抗 Flag 抗體膠體 (anti-FLAG-M2 agarose) 進行共免疫沉 澱,再利用抗 Tom20 抗體進行西方點墨法分析 (圖 3-4)。結果顯示,表現 Tag-Rta 的組別可觀察到被免疫沈澱下來的 Tom20 蛋白質 (圖 3-4 lane 2),而表現 pTag2A 的對照組則無 Tom20 蛋白質被免疫沈澱下來 (圖 3-4 lane 1)。由此結果可證實 Rta 蛋白質會與 Tom20 在細胞內結合。
30
三、 Rta 的粒線體導向序列 (MTS) 分析
經前述結果證實 Rta 確實位於粒線體,且會與粒線體膜上的導入蛋白 Tom20 結合,接著本研究欲探討 Rta 蛋白質上是否具有粒線體導向序列 (mitochondria targeting sequence,MTS),並找出其確切位置。首先,將 HA-Rta 及 Rta 的刪除 突變株 HA-PEST (1-347 a.a.)、HA-RC351 (255-605 a.a.) (圖 3-5A) 轉染入 293T 細 胞,培養 24 小時後收取細胞進行粒線體分離分析,最後利用西方點墨法以抗 HA 抗體偵測其訊號,結果顯示做為正對照組的 HA-Rta 在細胞質部分和粒線體部分 皆有分布 (圖 3-5 B lanes 1-3),只保留 Rta N 端 347 個胺基酸的 HA-PEST 在細 胞質部分和粒線體部分皆有分布 (圖 3-5 B lanes 4-6),然而只保留 Rta C 端的 HA-RC351 在粒線體部分則無分布 (圖 3-5 B lane 9),表示帶有 Rta N 端的 HA-PEST 會入粒線體,而不帶有 Rta N 端的 HA-RC351 則不入粒線體,因此推 測 Rta 的 MTS 應 位 於 Rta 蛋 白 質 之 N 端 。 接 著 於 293T 細 胞 轉 染 入 pTag-Rta、pTag-Bal5 (68-605 a.a.) 或 pTag-Stu (190-605 a.a.) (圖 3-6 A) 後經粒線 體分離分析,再利用西方點墨法以抗 Rta 抗體偵測其訊號,結果顯示做為正對照 組的 Flag-Rta 與 Flag-Bal5 在粒線體部分可偵測到其分布 (圖 3-6 B lanes 1-3, 7-9),而 Flag-Stu 在粒線體部分則無分布 (圖 3-6 B lane 6),可推測出 MTS 應位 於 Rta 蛋白質之 N 端第 68 至 190 胺基酸片段之間。同樣利用 293T 細胞為材 料,再經轉染入 pEGFP-N190、pEGFP-N120 或 pEGFP-N66~190 (圖 3-6 A) 後進 行粒線體分離分析,利用西方點墨法以抗 GFP 抗體偵測其訊號,結果顯示 GFP-N190、GFP-N120 和 GFP-N66~190 皆會進粒線體 (圖 3-6 C),故 MTS 之 位置可縮小推測範圍至 N 端第 68 至 120 胺基酸片段之間。最後,利用本研究 新構築的 Rta N 端刪除突變株 Flag-N80d (81-605 a.a.)、Flag-N90d (91-605 a.a.) (圖 3-6 A) 使用相同方法檢測其是否位於粒線體,結果顯示做為正對照組的 Flag-Rta
位於粒線體,而在粒線體部分則偵測不到 Flag-N80d 及 Flag-N90d 的分布 (圖 3-6 D lanes 6, 9)。MTS 之位置可進一步縮小推測範圍至 N 端第 68 至 90 胺基酸 片段之間。
另外本研究也以免疫螢光染色分析觀察 Rta 及其刪除突變株在細胞中的分布 位置。將 pTag-R 或 pTag-Stu 分別轉染入 293T 細胞中,於 24 小時後收取細胞,
以免疫螢光標定法標定 Rta 和 Tom20,結果顯示 Flag-Rta 與 Tom20 在細胞質 中會分布在相同位置 (圖 3-7 A, d),然而 Flag-Stu 則和 Tom20 明顯錯開 (圖 3-7 A, i),與粒線體分離分析的結果相符。另外將 GFP 或 GFP-N190 分別轉染入 293T 細胞中,24 小時後收取細胞,以免疫螢光標定法標定 Tom20,結果顯示 GFP 遍 佈於細胞各處 (圖 3-7 B),然而 GFP-N190 在細胞質中的分布位置明顯與粒線體 重合 (圖 3-7 B, i),表示 MTS 位於 Rta 蛋白質的 N 端 190 個胺基酸內,再次 佐證了先前粒線體分離分析的結果。
四、 Rta 的 N 端第 68 至 90 胺基酸之二級、三級結構預測
本研究使用 PredictProtein 和 PSIPRED 軟體預測 Rta 蛋白質 N 端片段 (68-100 a.a.) 的二級結構,結果顯示此片段由一個主要的 α 螺旋體和兩旁的環狀 結構 (loops) 所構成 (圖 3-8 A)。利用 SCRATCH 蛋白質預測建模網站和 Pymol 軟體對 Rta (68-90 a.a.) 建模獲得三級結構模型,該結果顯示此片段為一 α 螺旋體 結構,其範圍為 Leu73 至 Arg83 (圖 3-8 B 之 b,紅色部分);兩旁具有環狀結構 (圖 3-8 B 之 b,綠色部分);而螺旋體的一側為疏水性胺基酸所組成,其中包含 Leu73、Ile75、Phe78、Ile81、Leu82 (圖 3-8 B 之 c,橘黃色部分)。此模型為 MTS 的 典型結構,結果與二級結構預測結果基本一致。