國立臺灣大學醫學院免疫學研究所 碩士論文
Graduate Institute of Immunology College of Medicine
National Taiwan University Master Thesis
慢性 B 型肝炎小鼠動物模式共抑制性受體所誘發 CD8
+T 細胞 耗竭之免疫代謝功能失調之研究
Study of immunometabolism dysregulation of co-inhibitory receptors induced T cells exhaustion in chronic HBV mouse model
劉威延 Wei-Yan Liu
指導老師:許秉寧 博士 Advisor: Ping-Ning Hsu, Ph.D.
中華民國 107 年 7 月
July 2018
謝辭
碩士班兩年過去了,兩年間在實驗室中不斷嘗試新的題目新的方法,從一開始進實驗室 對實驗懵懵懂懂,中途失敗檢討以及題目推展的過程,慢慢到了逐漸上手實驗技術以及完成 碩士論文,從一個知識的容器逐漸開始能應用所學進行實驗研究,遇到困難便想辦法突破 它,在人生當中是一個新的階段。
感謝實驗室的指導教授,許秉寧博士,在進入免疫所接受老師的指導,從實驗的觀念、
安排、建構,導正途中所發現的錯誤所並且建立新的實驗系統,正確的報告呈現方法,能在 免疫學的研究上有正確的觀念,並且在實驗室新的研究系統上老師也特別請了清華大學鄭世 進博士幫忙檢查實驗的方法以及提供意見,確認了我在這樣的研究題目下所做出的結果是否 正確。感謝三位口試委員,朱清良博士、楊宏志博士、鄭世進博士,三次的口試報告當中給 與許多導正以及意見,讓我的實驗題目能及時修正錯誤的地方,完成碩士班兩年的研究,相 當感謝老師們對於我實驗過程中各種錯誤並有耐心地進行指導以及幫助。
感謝實驗室同組的博士班學長奇璋、世鴻、致宏、以祖學長,教導對於小鼠 B 型肝炎 實驗的作法,以及隨時討論各種 B 型肝炎相關的研究、各種實驗疑難雜症和新的免疫學觀 念,以及聊聊各種日常知識、常識,也感謝同屆的同學怡孜、凱富、彥佑、羅筠,對於碩士 班的問題、研究上提供幫助,以及大學時期的同學宥均、念如、威稷,關於研究以及心煩時 幫助我、傾聽我的心裡話,最後感謝父母、兄長,一直都是我的心靈支柱,隨時支持著我,
各位的幫助讓我得以更順利的通過碩士班的各種考驗,完成學業。
Abstract
Hepatitis B virus is a double-stranded DNA virus. About 257 million people in the world are
infected with hepatitis B virus. Most adults recover from hepatitis B infection, but approximately
5% to 10% of the population will not completely eliminate HBV infection, develop chronic
infections, and are at high risk of cirrhosis or liver cancer. However, there are currently safe and
protective vaccines can prevent hepatitis B, but it has not yet been able to effectively remove the
virus from infected patients with hepatitis B. From previous studies, it has been found that PD-1
(Programmed Death 1) and TIM-3 (T-cell immunoglobulin and mucin-domain containing-3) can be
detected under the HBV chronic infected mouse animal model, and displayed dysfunctional glucose
metabolism. Subsequent studies in PD-1 knockout mice challenged with LCMV have shown that
glucose metabolism of CD8+ T cells can be partially restored compared to wildtype LCMV infected
mice, indicating PD -1 affects the function of glucose metabolism and becomes the key to the
development of exhaustion in the later period. In mouse cancer animal models, the degree, number,
and extent of expression of co-inhibitory receptors PD-1 and TIM-3 have also been found that
positively correlated with decreased mitochondrial mass and rate of glucose uptake. However,
whether co-inhibitory receptors affect mitochondrial function is still unclear.
In previous studies in our laboratory, treatment of anti-PD-1 antibodies in mice chronically
infected with HBV enhanced the ability of mice to remove HBV, but it could not be completely
eliminated, indicating that the ability to eliminate the virus may be affected by other pathways. Our
results show that in chronically HBV infected mice, intrahepatic CD8+ T cells expressed PD-1, Tim-
3 surface co-inhibitory receptors and displayed dysfunctional mitochondrial function. In vitro
treating CD8+ T cells from HBV chronic infected mice with anti-PD-1, anti-Tim-3 or antioxidant
can increase CD8+ T cells mitochondrial membrane potential and IFNγ production. In summary, the
expression of co-inhibitory receptors in chronic HBV infected mice is associated with the
dysregulation of mitochondrial function and cytokine production in CD8+ T cells and through
blocking PD-1 and Tim-3 or treating mitochondrial antioxidant can partially restore T cell response.
It has been shown that the phenomenon of sputum cell exhaustion caused by hepatitis B virus
infection may involve in cellular energy metabolism, and regulation of immune metabolism may
enhance the function of these sputum cells and serve as a direction of treatment.
中文摘要
B 型肝炎病毒(Hepatitis B virus)是一種雙股 DNA 病毒,世界上大約兩億五千七百萬人處 於 B 型肝炎感染當中。大多數成人感染 B 型肝炎之後會康復,但是大約 5%~10%的比例會 無法完全清除 HBV 病毒的感染,形成慢性感染,並且為罹患肝硬化或是肝癌的高危險群,
然而現行已經有安全且有保護效力的疫苗存在可以預防 B 型肝炎,卻還沒有能有效清除已 感染 B 型肝炎病人病毒的醫療方法。從先前的研究中發現,在 HBV 慢性感染小鼠動物模式 之下,可以分析出表現 PD-1(Programmed Death 1)、TIM-3(T-cell immunoglobulin and mucin- domain containing-3)等 co-inhibitory 受器的肝內 CD8+ T 細胞族群,並且具有葡萄糖代謝作用 功能失調的現象,而之後在 PD-1 基因剔除的 LCMV 慢性感染小鼠研究中顯示葡萄糖代謝作 用功能可以被部分回復,顯示出 PD-1 影響了葡萄糖代謝功能的作用,成為發展至後期的 exhaustion 現象的關鍵,而在小鼠癌症動物模式當中,也發現了 co-inhibitory 受器 PD-1、
TIM-3 表現的程度、數量與粒腺體質量下降和葡萄糖的攝取呈正相關。然而是否共抑制性受 體會影響粒線體功能仍然不清楚。
在我們實驗室先前的研究當中,對於 HBV 慢性感染小鼠處理 anti-PD-1 抗體可以加強小 鼠對於 HBV 的清除能力,然而無法完全清除,顯示可能透過其它途徑影響了清除病毒的能 力。在我們的結果顯示,HBV 慢性感染小鼠中,肝內淋巴細胞 CD8+ T 細胞表現 PD-1、
TIM-3 表面受器,並且表現出粒線體呼吸作用功能失調的現象,體外處理 HBV 慢性感染小 鼠 CD8+ T 細胞抗 PD-1、Tim-3 抗體或是粒線體抗氧化劑回復 CD8+ T 細胞粒線體膜電位以 及IFNγ 的產生量。綜上所述,HBV 慢性感染中共抑制性受體的表現與 CD8+ T 細胞粒線體 呼吸作用失調現象關聯,影響 T 細胞的細胞激素的分泌,並且透過阻斷 PD-1 以及 Tim-3 或 者是處理粒線體抗氧化劑得以回復肝內淋巴細胞 CD8+ T 細胞粒線體膜電位。顯示出在 B 型 肝炎病毒感染中所造成 T 細胞衰竭的現象可能涉及細胞能量代謝的異常,經由免疫代謝的調 節可能可提升這些 T 細胞的功能,並作為治療的一個方向。
目 錄
謝辭………...1
英文摘要………...2
中文摘要………...4
第一章 引言...………..7
1. B 型肝炎………...7
2. T 細胞耗竭………...7
3. T 細胞免疫代謝………...8
4. B 型肝炎與 T 細胞免疫代謝功能之關聯………..8
5. 研究動機與目的………10
第二章 實驗材料與方法………...11
第三章 實驗結果………...16
1. HBV 慢性感染小鼠肝內 CD8
+T 細胞表達 PD-1 以及 Tim-3 共抑制性受 體並且 IFNγ 表達量下降……….16
2. HBV 慢性感染小鼠肝內 CD8+ T 細胞表現粒線體膜電位下降以及粒線體 ROS 累積量上升表徵………16
3. 表達共抑制性受體之肝內淋巴細胞 CD8+ T 細胞經過體外 CD3、CD28
刺激無法活化粒線體膜電位訊號並維持低電位表現……….17
4. 共抑制性受體配體降低粒線體膜電位之訊號,以及呼吸作用之氧氣消
耗速率而阻斷 PD-1、Tim-3 訊號處理之後回復粒腺體之膜電位訊號以 及呼吸作用的氧氣消耗速率……….17 5. 共抑制性受體配體降低 CD8
+T 細胞 IFNγ 產生而處理 PD-1、Tim-3 抗體或是粒線體抗氧化劑後回復 IFNγ 產生量………...17 6. 阻斷共抑制性受體之訊號回復 BV 慢性感染小鼠之肝內淋巴細胞 CD8+
T 細胞之粒線體膜電位訊號………...18
7. 粒線體抗氧化劑處理之 HBV 感染小鼠肝內 CD8+ T 細胞提高 IFNγ 表
現量………...18
第四章 討論………...19
1. HBV 高壓水注射小鼠動物模式 T 細胞免疫反應……….19
2. 肝內 T 細胞免疫代謝功能在 HBV 慢性感染所誘發之 T 細胞耗竭當中降
低粒線體膜電位、葡萄糖攝取效率……….…… 19
3. 共抑制性受體訊號降低 CD8+ T 細胞之粒腺體功能以及細胞激素的表現
量………..……20
4. 阻斷共抑制性受體訊號回復 HBV 慢性感染所誘發之 T 細胞耗竭之粒線
體功能失調……….……….21
結論………..23
Figures………..24
第一章 引言
1. B 型肝炎
B 型肝炎病毒(Hepatitis B virus, HBV)由諾貝爾獎得主巴魯克•塞繆爾•布隆伯格於 1965 年發現於澳大利亞原住民的血液樣本,當時命名為"Australia Antigen"(BS Blumberg, 1970)。B 型肝炎為一種部分雙股 DNA 病毒,感染到宿主細胞後會以病 毒的聚合酶修復為完整雙股 cccDNA 形式,並且可以嵌合到宿主的基因當中,複 製時經過 RNA 作為中間產物,表現基因時透過 RNA 產生病毒相關蛋白,包括 LHBsAg、MHBsAg、SHBsAg、HBcAg 等。B 型肝炎病毒可在體外存活大約 7 天,可再感染不受疫苗保護的人,B 型肝炎病毒可以藉由體液、輸血、性行為或 是母子垂直感染來傳染至別的人(Liang, 2009)。世界上大約有兩億五千七百萬的 人遭受 B 型肝炎病毒感染,大多數成人感染之後可以痊癒,然而有大約 5%~10%
的人會無法完全清除病毒,造成慢性感染,部分的病人將演變成肝硬化或肝癌,
而現行有有效保護且安全的 B 型肝炎疫苗,然而對於已經慢性感染的病人卻還沒 有有效的方法可以治療,而常見降低 B 型肝炎感染的方法為貝樂克與 IFNα,然 而皆只能抑制病毒的複製,仍然無法使慢性感染病人的病毒清除完全,在停藥之 後很容易出現復發的現象(World Health Organization, 2017),因此尋找可以醫治 B 型肝炎的有效療法為現在重要的課題所在。
2. T 細胞耗竭
在 2006 年首先在淋巴球性脈絡叢腦膜炎 clone 13 的小鼠動物模式當中定義一群細 胞激素分泌量下降、細胞增殖能力損失的 T 細胞為 exhaustion 的狀態(Barber et al., 2006)。在 C 型肝炎慢性感染病人樣本研究當中 CD4+、CD8+ T 細胞提高表現 TIM-3,並且分泌更少量的 IFNγ 以及失去增殖的能力,為慢性感染之下產生的 exhausted T 細胞(Jones et al., 2008)。之後另一篇研究透過將 TIM-3 與共表現的
CEACAM1 (carcinoembryonic antigen cell adhesion molecule 1)做阻斷的情形之下可 以增加 T 細胞抗腫瘤的能力(Huang et al., 2015),另外在 B 型肝炎的感染中,慢性 感染的病人血液樣本中可以偵查到表達 TIM-3 的 exhausted T 細胞族群,這群 T 細胞也產生了IFNγ 與 TNFα 分泌量下降的特徵(Nebbia et al., 2012)。
3. T 細胞免疫代謝
T 細胞的活化以及細胞激素的分泌都需要依靠能量來源,T 細胞可以依靠氧化磷 酸化階層產生以及糖解作用產生的 ATP 作為能量,不同狀態、時期皆能切換到適 當的能量供應形式,然而若是在正常培養及刺激的情形之下阻斷了糖解作用,可 以直接讓 T 細胞產生 IFNγ 的能力喪失,顯示出代謝作用對於 T 細胞功能運作的 重要性(Chang et al., 2013)。有研究顯示在於 LCMV 的小鼠動物模式早期中,受到 慢性感染病毒株感染的小鼠體內 T 細胞在主要的 exhaustion 特徵表達前代謝作用 產生轉變,透過阻斷 PD-1 訊號、增加葡萄糖取得以及加強代謝調節因子 PGC- 1α 表現能部分改善早期 exhausted 的情形,阻止部分細胞轉變成晚期 exhausted 的 型態,作者以此推論早期的代謝機制會影響到 T 細胞轉換成完整 exhausted 的型 態,代謝失調比如粒線體堆積無法清除的活性氧物質(ROS)將會造成 T 細胞功能 上的損壞(Bengsch et al., 2016)。
4. B 型肝炎與 T 細胞免疫代謝功能之關聯
而在 B 型肝炎的人類檢體實驗當中,也在慢性感染 B 型肝炎的血液樣本中偵測到 葡萄糖代謝功能的失調現象,表現如粒線體膜電位去極化、葡萄糖攝入量下降,
T 細胞出現功能上的損失(Schurich et al., 2016),而另一篇研究則是對於慢性 B 型 肝炎病人血液檢體中的 exhausted T 細胞施以抗氧化劑,能透過清理因堆積大量活 性氧物質而受損的粒腺體內部 ROS 而恢復粒腺體的運作功能,而發現在粒線體 功能回復可以再使的 T 細胞在經過病毒蛋白刺激後能提高表現 IFNγ 以及
TNFα(tumor necrosis factor-α),隱射 T 細胞的能力回復並可能得以增加病毒清除
的能力(Fisicaro et al., 2017),然而共抑制性受體與粒線體途徑之間的影響方式以 及詳細機制並且導致 T 細胞產生 exhaustion 現象尚無詳細研究報告。
5. 研究動機與目的:
關於 B 型肝炎小鼠動物模式之病毒清除能力研究,我們實驗室以
Hydrodynamic injection 方式,透過從小鼠尾靜脈打入建構在腺相關病毒之 B 型肝 炎病毒的質體 DNA,以高壓打入轉染到小鼠的肝細胞當中,模擬病毒感染之情 形,血液當中可以檢驗出 B 型肝炎所產生之 HBsAg 用以追蹤感染病程,並且在 慢性感染當中可以偵測到表現 co-inhibitory 受器的 T 細胞族群。而之前研究當中 對於慢性感染小鼠施打 anti-PD-1 抗體處理結果發現可以增加小鼠對於 B 型肝炎 病毒的清除能力,然而猶未能完全清除病毒的感染,而另外我們實驗室也有研究 TIM-3 阻斷的情形下,亦能增加慢性感染小鼠對於 B 型肝炎病毒的清除能力,並 且在另一篇研究報導指出在小鼠癌症動物模式研究當中,T 細胞表面表現的 co- inhibitory 受器數量與葡萄糖代謝的失調相關,PD-1、TIM-3 共表現對比只有 PD- 1 表現的 T 細胞族群更是呈現葡萄糖代謝功能損失的程度加強,葡萄糖攝取量下 降以及粒線體總體質量的減少,進而暗指 T 細胞的共抑制性受體表現與葡萄糖代 謝的關聯性(Scharping et al., 2016),然而共抑制性受體對於 T 細胞粒線體功能的 下降關聯性還尚未有明確的研究。
第二章 實驗材料與方法
實驗方法:
1. 實驗動物
國家實驗動物中心 SPF 等級代養飼育之 6 週齡 C57BL/6 老鼠,實驗動物相關實驗 以及使用方法皆經過實驗動物照護及使用委員會(IACUC)之檢閱以及核可。
2. 高壓水動力注射
六週齡大之 C57BL/6 小鼠,使用小鼠保定器固定之後以五秒內透過尾部側邊靜脈 注射相當於小鼠十分之一體重之含有 10μg HBV 質體之 DPBS,B 型肝炎轉染之 模擬感染小鼠以小鼠臉頰採血方式檢驗血液當中所呈現的 B 型肝炎表面蛋白抗原 濃度追蹤感染情形。表現 HBV 基因之腺相關病毒載體(pAAV)藉由無內毒素質體 DNA 純化試劑純化獲得。
3. 分離肝內淋巴細胞
實驗鼠經由靜脈注射舒泰麻醉,實驗鼠之肝臟以 10 c.c.之 DPBS 灌注清洗並且全 部切除。取下之實驗鼠肝臟以活塞磨碎並通過70μm 之尼龍細胞濾網,以 HBSS 清洗並配至體積 50c.c.,50xg 轉速 4 度溫度之下離心兩次用以去除溶液中的肝臟 細胞以及大型細胞團,再由 400xg 離心含有肝內淋巴細胞之上清液並去除上清 液,離心下來之細胞溶於 40%之 Percoll 層析液中,並緩緩加入到 70%之 Percoll 層析液上方,之後以 1200xg 在 25 度環境下 25 分鐘離心,肝內淋巴細胞在離心 完之後會在 40%/70%的 Percoll 層析液界面上,吸取之後加入 10c.c.的 HBSS 清洗 以 4 度離心 400xg 10 分鐘,取出細胞沉澱供分析或培養使用。
4. 流式細胞儀
小鼠細胞表面受體以 BV421-conjugated anti-mouse CD3, BV510-conjugated anti- mouse CD8, PE-conjugated anti-mouse PD-1, PE-Cy7-conjugated anti-mouse Tim-3, APC-conjugated anti-mouse Tim-3, PE-Cy7-conjugated anti-mouse PD-1 染色;細胞 激素以 PE-conjugated anti-mouse IFNγ 染色。細胞表面受體染劑以 1:500 稀釋並於 含有 2% FBS 之 DPBS 細胞懸浮溶液當中染色 30 分鐘,細胞激素的染色在細胞表 面受體染色結束後以eBioscience™ Intracellular Fixation & Permeabilization Buffer Set 固定並且打破細胞進行細胞內染 IFNγ,染色結束分析細胞表面受體、細胞激 素之表現情形,以 FlowJo 軟體分析所得到之流式細胞儀檔案。
5. T 細胞醣類攝取效率測定、粒線體膜電位、超氧化物測定
實驗鼠肝內淋巴細胞、脾臟細胞在 37 度環境中 DPBS 溶液內加入 2-(N-(7-
Nitrobenz-2-oxa-1,3-diazol-4-yl)Amino)-2-Deoxyglucose (2-NBDG) 20μM 培養 30 分 鐘,流式細胞儀偵測同時間內細胞攝取之 2-NBDG 螢光強度分析細胞從外界攝取 醣類之相對效率;肝內淋巴細胞在 37 度環境中含 2% FBS 之 DPBS 溶液內以 JC- 1 線粒體膜電位探針 20μM、tetramethylrhodamine ethyl ester (TMRE)試劑 200nM 或MitoTracker™ Deep Red FM 粒線體深紅色螢光探針 100nM 染色 30 分鐘並以流 式細胞儀偵測整體螢光強度分析粒腺體膜電位變化。
6. T 細胞共抑制受體配體體外處理、粒線體抗氧化劑處理以及細胞激素產生測 定
實驗鼠脾臟淋巴細胞以 Biolegend MojoSortTM Mouse CD8 T Cell Isolation Kit 分離 出 CD8+ T 細胞,並於 37 度環境中含有培養於 10% FBS、外加 glutamax 1mM、
sodium pyruvate 1mM、IL-2 之 RPMI1640 於 CD3 抗體 Pre-coated 的 96 孔盤中 48 小時,之後置換於 CD3 抗體、PD-L1 或 Ceacam-1(10μg/ml) Pre-coated 的 96 孔盤
中 48 小時,經過抗 PD-1、抗 Tim-3 抗體(10μg/ml)或粒腺體抗氧化劑 10 μM 之處 理,以流式細胞儀測定粒線體膜電位、T 細胞 IFNγ 產生之比例。
7. 海馬微量細胞代謝分析儀
實驗鼠脾臟淋巴細胞以 Biolegend MojoSortTM Mouse CD8 T Cell Isolation Kit 分離 出 CD8+ T 細胞,於 37 度培養於含有 10% FBS、外加 glutamax 1mM、sodium pyruvate 1mM 之 RPMI1640,實驗處理之後轉移至 96-well polystyrene Seahorse plates 於 RPMI1640(不含 FBS)並以 Seahorse XFp Analyzer 機台處理 Mito Stress kit 試劑並偵測在 oligomycin (1 mM), FCCP (1.5mM) and antimycin A/rotenone (1 mM/0.1mM)處理之下細胞的氧消耗速率(OCR)變化圖。
8. 細胞免疫斑點法(ELISpot)
實驗小鼠肝內淋巴細胞加入 ELISpot 白色盤中,以 CTL 無血清培養基加入 glutamax 1mM、sodium pyruvate 1mM、IL-2 培養並以 B 型肝炎核心蛋白胜肽再 刺激以及抗氧化劑 MitoTEMPO、共抑制受體抗體 anti-PD-1, anti-Tim-3 處理 37 度 24 小時,之後以 ImmunoSpot® Kits Mouse IFN-γ Single-Color ELISPOT 進行偵測 IFN-γ 產生之班點數,以細胞免疫斑點影像分析儀計算 ELISpot 白色盤每格斑點 形成之數目。
實驗材料:
BV421-conjugated anti-mouse CD3(17A2) Biolegend, San Diego, CA, USA BV510-conjugated anti-mouse CD8(53-6.7) Biolegend, San Diego, CA, USA PE-conjugated anti-mouse PD-1 Biolegend, San Diego, CA, USA PE-Cy7-conjugated anti-mouse Tim-3 Biolegend, San Diego, CA, USA APC-conjugated anti-mouse Tim-3 Biolegend, San Diego, CA, USA PE-Cy7-conjugated anti-mouse PD-1 Biolegend, San Diego, CA, USA PE-conjugated anti-mouse IFNγ Biolegend, San Diego, CA, USA Percoll GE Healthcare
HBSS Corning, Arizona, USA
DPBS 0.1M Na2HPO4, 0.018M KH2PO4, 0.8% NaCl, 0.02% KCl, pH7.4 MitoTracker Deep Red FM ThermoFisher, Waltham, Massachusetts, USA JC-1 Dye ThermoFisher, Waltham, Massachusetts, USA
TMRE Abcam, Cambridge, England 2-NBDG Abcam, Cambridge, England Seahorse XFp FluxPak Agilent Santa Clara, CA, USA PD-L1 Biolegend, San Diego, CA, USA Ceacam-1 Sino biological, Wayne, PA, USA MitoTEMPO Sigma, St.Louis, USA
Anti-PD-1 Biolegend, San Diego, CA, USA Anti-Tim-3 Biolegend, San Diego, CA, USA RPMI-1640 Gibco, USA
Glutamax Gibco, USA
Sodium pyruvate Sigma, St.Louis, USA
ImmunoSpot® Kits Mouse IFN-γ Cellular Technology Limited (CTL), Single-Color ELISPOT Cleveland, USA
第三章 實驗結果
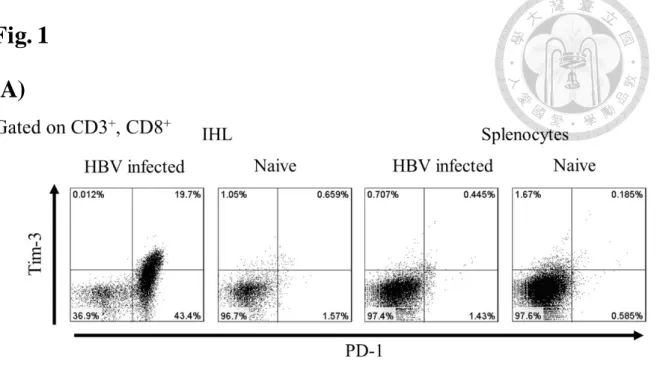
1. HBV 慢性感染小鼠肝內 CD8+ T 細胞表達 PD-1 以及 Tim-3 共抑制性受體並且 IFNγ 表達量下降
以 hydrodynamic injection 轉染之小鼠動物模式當中,經過感染六周之後,取 出小鼠的肝內 T 細胞族群分析,T 細胞族群表現了 PD-1+ TIM-3lo 以及 PD-1+ TIM-3hi 次群(Fig. 1A),經由體外以 CD3/CD28 抗體額外刺激一天的情況之下,
IFNγ 產生量相較於未轉染 HBV 之小鼠組別下降(Fig. 1B)。在 HBV 慢性感染情形 之下共抑制性受體的表現與 CD8+ T 細胞之粒線體功能關聯性在下一個粒線體訊 號偵測實驗來證明。
2. HBV 慢性感染小鼠肝內 CD8+ T 細胞表現粒線體膜電位下降以及粒線體 ROS 累積量上升表徵
JC-1 染劑偵測 HBV 慢性感染六週之實驗小鼠肝內淋巴細胞表達共抑制性受 體的 T 細胞表現出粒腺體膜電位去極化現象,對照不表達共抑制受體之 T 細胞膜 電位訊號較低(Fig. 2A),以 JC-1 及 MitoTracker Deep Red 染劑偵測表達共抑制性 受體的 T 細胞對比於其餘 T 細胞族群呈現更高比例的粒線體膜電位去極化比例,
葡萄糖的攝取量亦在表達共抑制性受體 T 細胞當中表現出更低量攝取效率的特徵 (Fig. 2B),同時表達比較多的共抑制性受體 T 細胞也在以 MitoSOX 染色並以流式 細胞儀分析之中偵測到表現出更多量的粒線體 ROS 訊號(Fig. 2C)。
3. 表達共抑制性受體之肝內淋巴細胞 CD8+ T 細胞經過體外 CD3、CD28 刺激無 法活化粒線體膜電位訊號並維持低電位表現
肝內淋巴細胞經過 CD3、CD28 體外刺激實驗一天之後,粒線體膜電位偵測 染劑 JC-1 染劑以流式細胞儀偵測發現粒線體無法再度被活化,維持高比例低粒線 體膜電位之表徵,不表現共抑制性受體之 CD8+ T 細胞在刺激之後可以提升粒腺 體之膜電位,降低去極化之粒線體比例(Fig. 3)。共抑制性受體訊號對於粒線體功 能以及 CD8+ T 細胞反應能力的影響須進一步的研究。
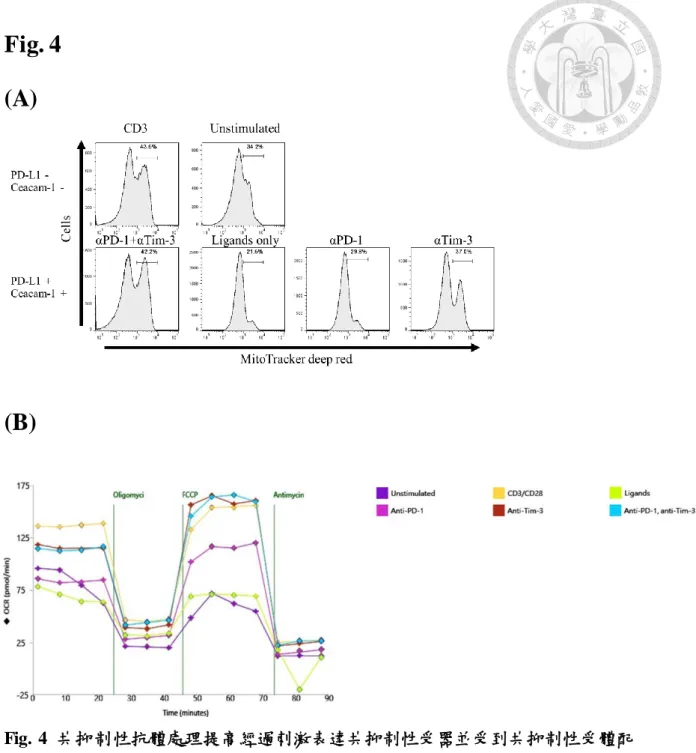
4. 共抑制性受體配體降低粒線體膜電位之訊號,以及呼吸作用之氧氣消耗速率 而阻斷 PD-1、Tim-3 訊號處理之後回復粒腺體之膜電位訊號以及呼吸作用的 氧氣消耗速率
實驗小鼠脾臟分離出之 CD8+ T 細胞以體外 PD-L1、Ceacam-1 處理,以流式 細胞儀偵測發現粒線體膜電位訊號降低的現象(Fig. 4A),以海馬生物能量測定儀 亦發現粒腺體的氧氣消耗速率下降(Fig. 4B),然而在加入 anti-PD-1 以及 anti-Tim- 3 抗體處理之後,粒線體的膜電位訊號得以回復(Fig. 4A)以及呼吸作用氧氣消耗 速率可以獲得回升(Fig. 4B)。
5. 共抑制性受體配體降低 CD8+ T 細胞 IFNγ 產生,而處理 PD-1、Tim-3 抗體或 是粒線體抗氧化劑回復IFNγ 產生量
實驗小鼠脾臟分離出之 CD8+ T 細胞透過抗 PD-1 和 Tim-3 抗體的處理,或是 抗氧化劑 MitoTEMPO 處理之後,得以在體外刺激之後回復 IFNγ 的產生量,比起 未處理抗體之組別能提高IFNγ 的產量(Fig. 5)。在 HBV 感染情形之下,HBV 病 毒所引起之 CD8+ T 細胞耗竭對於共抑制性受體訊號阻斷後回復 T 細胞功能以及
6. 阻斷共抑制性受體之訊號回復 HBV 慢性感染小鼠之肝內淋巴細胞 CD8+ T 細 胞之粒線體膜電位訊號
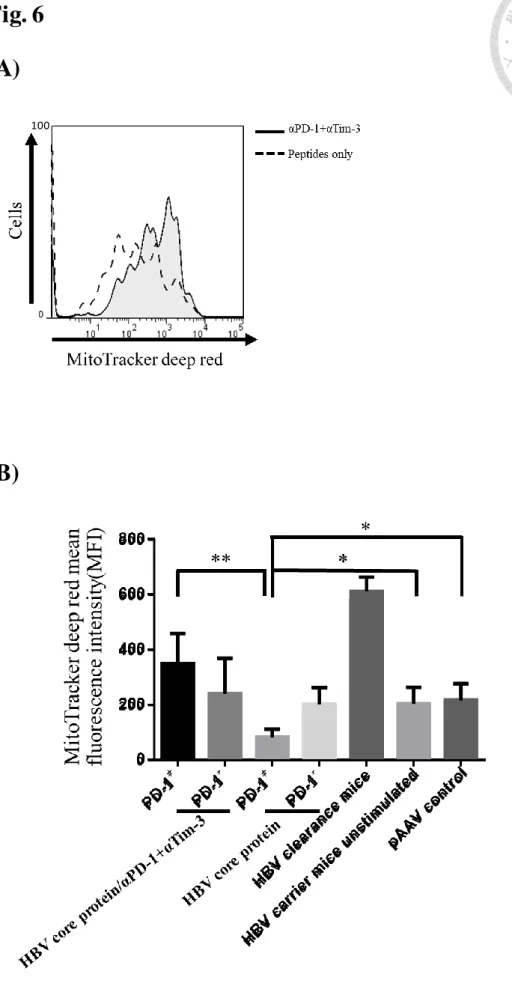
將 10μg 帶有表現 HBV DNA 之質體轉染之老鼠感染六週之後,取出肝內 淋巴細胞並且在體外以 HBV 核心蛋白做再刺激,以流式細胞儀偵測
MitoTracker Deep Red 之訊號強度,顯示經由 anti-PD-1 以及 anti-Tim-3 抗體處 理之組別表達共抑制受體之 CD8+ T 細胞粒線體膜電位得以在核心蛋白刺激之 中提高,而相較於不處理抗體組呈現較低的粒線體膜電位(Fig. 6A),以直條圖 顯示流式細胞儀偵測之 MitoTracker Deep Red 粒線體膜電位訊號之 MFI 值 (Mean fluorescence intensity),並且以學生 t 檢定(Student's t-test)雙尾檢驗 P 值 顯示出在抗體處理之後表達 PD-1 之 CD8+ T 細胞對比於物處理抗體之組別擁 有顯著差異(P 值小於 0.01)的粒線體膜電位 MFI 值的上升,而處理 HBV 核心 蛋白在刺激之 CD8+ T 細胞對比於不處理核心蛋白之組別具有顯著性的 MFI 值 下降(P 值小於 0.05) (Fig. 6B)。在表達共抑制性受體之 CD8+ T 細胞中影響了 粒線體的功能,而直接對於粒線體的功能回復在共抑制性受體訊號作用之下 對於 CD8+ T 細胞反應的改善為下一個我們想測試的實驗。
7. 粒線體抗氧化劑處理之 HBV 感染小鼠肝內 CD8+ T 細胞提高 IFNγ 表現量 透過抗氧化劑 MitoTEMPO 處理兩天之後,MitoTracker 訊號顯示表現共抑制 性受體的 T 細胞粒線體膜電位產生較大比例的回復(Fig. 7A),表現 IFNγ 上升量 較高(Fig. 7B, 8A, 8B),隱射出表現共抑制性受體的 T 細胞粒線體功能失調而造成 T 細胞的 exhaustion 狀態產生,表達共抑制性受體的 T 細胞比起不表達 PD-1 的 T 細胞呈現下降的粒線體膜電位狀態,而透過限制粒線體 ROS 產生的方法回復粒 線體功能得以回復 T 細胞的 IFNγ 產生量。綜合所有研究,共抑制受體可能透過 影響 CD8+ T 細胞的粒線體功能進而導致 IFNγ 產生量下降降低 T 細胞的反應能 力。
第四章 討論
1. HBV 高壓水注射小鼠動物模式 T 細胞免疫反應
每隻實驗鼠以 2 毫升 DPBS 帶有 10μg HBV 質體從尾靜脈在五秒之內打入,
透過此種方式模擬感染 HBV 病毒之情形。從實驗結果得知在此種情形之下感染 六週以上的小鼠肝臟當中可以分離出提高表現共抑制性受體之 CD8+ T 細胞族 群,並且在經過體外 CD3、CD28 刺激之後也相較於正常 T 細胞表達比較低量的 IFNγ,與之前對於 LCMV 感染小鼠動物模式(Barber et al., 2006)以及癌症小鼠動 物模式(Pauken and Wherry, 2015)當中 T 細胞耗竭情形吻合,然而在此種小鼠模式 當中 HBV 並無法進行正常感染過程,HBV 病毒無法藉由細胞表面受器進入老鼠 的肝臟細胞當中,在病毒免疫方面缺乏了從細胞外到細胞內並且病毒 DNA 穿插 到宿主肝臟細胞的染色體當中的過程,無法完全模擬真實 HBV 的感染過程,在 共抑制受體的偵測方面亦有更多的種類接續被發現(Ye et al., 2015),都是有可能造 成 T 細胞反應能力低落的可能性。體外的 CD3、CD28 的 T 細胞刺激以流式細胞 儀分析細胞內染之IFNγ 表現量可以得知進入耗竭狀態的 T 細胞無法在額外刺激 當中再被活化,而對於病毒免疫方面亦須進一步確認。
2. 肝內 T 細胞免疫代謝功能在 HBV 慢性感染所誘發之 T 細胞耗竭當中降低粒 線體膜電位、葡萄糖攝取效率
從 HBV 慢性感染 6 週之老鼠肝臟中分離之肝內 CD8+ T 細胞,以流式細胞儀 偵測粒線體探針 JC-1 以及 MitoTracker Deep Red 之訊號顯示出 CD8+ T 細胞粒線 體在慢性感染當中表現粒線體膜電位下降的情形,並且在偵測 2-NBDG 訊號亦顯 示出葡萄糖的攝取效率在此狀態之下降低,與先前 LCMV 感染小鼠動物模式之研 究符合(Bengsch et al., 2016),並且在表現更多的共抑制性受體的 T 細胞族群在粒 線體膜電位以及葡萄糖攝取效率有更加下降的趨勢,與先前癌症小鼠動物模式之
結果相似(Scharping et al., 2016),表達越多的共抑制性受體與葡萄糖攝取效率以及 粒線體質量下降成正相關代表在表達共抑制性受體的 T 細胞族群當中呈現相對降 低的細胞代謝能力,與細胞的反應能力以及分化能力等可能有相關,然而免疫代 謝作用對於 T 細胞是否能清除 HBV 感染之細胞的能力以及對於小鼠體內 HBV 整 體免疫能力的改變仍需要更多的實驗來做確認。
3. 共抑制性受體訊號降低 CD8+ T 細胞之粒腺體功能以及細胞激素的表現量 透過直接對於脾臟分離之未活化 T 細胞先以 CD3、CD28 訊號刺激再加入共 抑制性受體配體,可以發現此種情形之下 T 細胞的粒線體功能以及細胞激素產生 能力受到抑制,而透過再加入共抑制性受體抗體阻斷共抑制性受體的訊號的情形 之下,則可以再將 T 細胞的粒線體功能以及細胞激素產生量回升,顯示出共抑制 性受體對於 T 細胞的粒線體功能具有影響能力,並且也影響了 T 細胞的細胞激素 產生能力,造成類似於慢性病毒感染之 T 細胞耗竭狀態,而針對是否降低的粒線 體功能會影響到 T 細胞反應能力面,則使用了粒線體抗氧化劑 MitoTEMPO 處 理,先前研究指出透過清除、限制粒線體 ROS 產生可以回復一部份粒線體功 能,過量的 ROS 堆積將會造成粒線體的損壞(Fisicaro et al., 2017; Guo et al., 2013),而在粒線體抗氧化劑處理之後在共抑制性受體訊號持續情形之下也能回升 T 細胞的細胞激素的產生量,說明在 T 細胞接收共抑制性受體訊號可能影響了粒 線體功能進而降低 T 細胞反應能力,可以透過調整粒線體功能的方法回復 T 細胞 的細胞激素產生能力。在先前研究認為 T 細胞在活化並執行胞殺作用、分化以及 細胞激素產生的過程中主要以糖解作用為主要的能量來源(Pearce et al., 2013),然 而在之後的研究提出粒線體功能對於 T 細胞的正常活化以及抗原特異性 T 細胞的 再活化為重要角色(Klein Geltink et al., 2017; Sena et al., 2013),在 T 細胞耗竭當 中,共抑制性受體降低了粒線體膜電位亦可能是對於 T 細胞反應能力的抑制的原 因。不同的共抑制性受體對於 CD8+ T 細胞粒線體功能之影響也有所差異,從不 同的共抑制性受體配體 PD-L1 以及 Ceacam-1 處理對於 CD8+ T 細胞觀察到對於粒
線體膜電位的影響程度不同,Tim-3 訊號對於 CD8+ T 細胞粒線體膜電位變化影響 具有較大的趨勢,再經由 Tim-3 抗體處理之後恢復的程度也相對多,隱涉出 Tim-3 可能對於 CD8+ T 細胞的粒線體功能的影響性相對較多,Tim-3 相較於 PD-1 可能經由調控粒線體功能而影響 T 細胞細胞激素生成能力並且影響 T 細胞的反應 能力,在先前研究中 Tim-3 亦被認為可能影響了 CD8+ T 細胞的粒線體功能 (Scharping et al., 2016; Siska et al., 2014; Yin et al., 2016),作為 CD8+ T 細胞粒線體 功能失調中可能的原因,Tim-3 經由何種機制調控 CD8+ T 細胞粒線體功能以及反 應能力仍需要進一步的研究來確認。
4. 阻斷共抑制性受體訊號以及回復 HBV 慢性感染所誘發之 T 細胞耗竭之粒線體 功能失調
HBV 慢性感染之下所誘發之肝內 CD8+ T 細胞耗竭,經實驗鼠離體處理共抑 制性受體抗體得以回復粒線體膜電位訊號,經過 HBV 病毒之核心蛋白刺激之中 相對於不處理抗體組整體可以有更高的粒線體膜電位產生,顯示可以利用阻斷共 抑制性受體的方式來回復被 HBV 慢性感染所誘發之 T 細胞耗竭中的粒腺體功能 失調的情形,並且透過處理粒線體抗氧化劑之肝內 CD8+ T 細胞在 CD3、CD28 刺 激之後可以提高IFNγ 的表現量,顯示透過抑制共抑制性受體的訊號途徑或是調 整粒線體功能的方式可以改善 HBV 慢性感染所誘發之免疫代謝功能失調現象以 及回復一部分的 T 細胞反應能力。其他研究當中透過阻斷共抑制受體訊號 PD- 1、Tim-3 對於小鼠癌症模式當中可以增加對於腫瘤的清除能(Huang et al., 2015;
Sakuishi et al., 2010),而利用抗氧化劑限制粒線體 ROS 生成量則能在 B 型肝炎人 類血液樣本當中回復 CD8+ T 細胞的 IFNγ 表現量(Fisicaro et al., 2017),共抑制性 受體如 PD-1、Tim-3 可能透過抑制粒線體呼吸作用方式,降低粒線體膜電位以及 氧氣消耗速率,進而導致 T 細胞在產生 IFNγ 的能力下降。未來針對是否 CD8+ T 細胞免疫代謝失調對於老鼠 HBV 的清除是否有直接影響,以及共抑制性受體造
T 細胞粒線體功能以及反應的影響並可能作為新的治療目標,是需要進一步實驗 去證實的。
結論
綜合所有實驗,實驗小鼠在模擬感染 HB 的過程當中,六週慢性感染過程造 成小鼠肝內 CD8+ T 細胞表達出共抑制性受體 PD-1、Tim-3,並且在經過體外 CD3、CD28 或者是 HBV 核心蛋白再刺激的情況之下,相比於不表達共抑制性受 體或是健康各體分離之 CD8+ T 細胞 IFNγ 的產量明顯較低,並且在粒線體功能以 及葡萄糖攝取效率方面亦有所降低。從共抑制受體配體 PD-L1、Ceacam-1 的處理 以及阻斷共抑制性受體訊號之實驗當中隱射共抑制受體可能影響 CD8+ T 細胞的 粒線體呼吸作用而導致 T 細胞的 IFNγ 產量下降,在 HBV 小鼠動物模式當中也顯 示出類似的結果,T 細胞的粒線體功能可以在阻斷 PD-1、Tim-3 後回復,並且一 部分 T 細胞的 IFNγ 產量能在維持粒線體功能中得到提升,CD8+ T 細胞的粒線體 功能作為可能的 T 細胞反應能力的原因及研究目標。
Figures
Fig. 1 (A)
(B)
Fig. 1 B 型肝炎慢性感染小鼠 CD8+ T 細胞表現 PD-1、TIM-3 以及 IFNγ 表現量 以 hydrodynamic injection 轉染 6 周之小鼠,取出肝內淋巴細胞族群以流式細胞儀 分析觀察 T 細胞的 co-inhibitory 受器的表現(A)以及內染 IFNγ(B), IFNγ 的表現 量在 HBV 慢性感染的小鼠當中的 CD8 T 細胞在 CD3/CD28 刺激一天後也比較少 量上升
Fig. 2 (A)
(B)
(C)
Fig. 2 肝內 T 細胞表達共抑制性受體次群相對於不表達共抑制受體次群粒線體膜 電位表現更為去極化現象,以及產生更多粒線體 ROS,降低葡萄糖的攝取速度 (A) 取出 hydrodynamic injection 轉染 6 周之小鼠肝內 T 細胞以流式細胞儀分析 JC-
1 染色訊號,偵測粒線體膜電位變化的情形
(B) 取出的 hydrodynamic injection 轉染 6 周之小鼠肝內 T 細胞以流式細胞儀偵測 MitoTracker Deep Red 以及 2-NBDG 分別分析粒線體膜電位以及葡萄糖攝取速 率
(C)以 MitoSOX 偵測慢性 HBV 感染小鼠的肝間 T 細胞之粒線體 ROS 產生量
Fig. 3
Fig. 3 肝內淋巴細胞表達共議性受體之 CD8+ T 細胞經過體外 CD3/CD28 抗體刺 激之後粒線體膜電位維持低訊號
對於取出的 hydrodynamic injection 轉染 6 周之小鼠肝內 T 細胞進行體外 CD3、
CD28 刺激 24 小時,以流式細胞儀偵測 JC-1 red 以及 green 訊號分析染色粒線體 膜電位變化
Fig. 4 (A)
(B)
Fig. 4 共抑制性抗體處理提高經過刺激表達共抑制性受器並受到共抑制性受體配 體處理之 T 細胞之粒線體膜電位比例、氧氣消耗速率
(A) 取出小鼠脾臟之未活化 T 細胞先行刺激 48 小時表現出共抑制性受器,並在之 後額外 48 小時刺激過程中以 Tim-3、PD-1 ligand 處理,以 MitoTracker Deep Red 偵測粒線體膜電位變化
(B) 以 Seahorse XFp analyzer 偵測 T 細胞在經過體外 PD-L1、Ceacam-1 處理並且 加入 anti-PD-1 以及 anti-Tim-3 抗體後的氧氣消耗速率變化
Fig. 5
Fig. 5 共抑制性抗體或粒線體抗氧化劑處理提高經過刺激表達共抑制性受器並受 到 Tim-3、PD-1 ligand 處理處理之 T 細胞之 IFNγ 的產生
實驗鼠脾臟分離出之 CD8+ T 細胞以 PD-L1 及 Ceacam-1 處理並加入 anti-PD-1、
anti-Tim-3 抗體和粒線體抗氧化劑 MitoTEMPO 10μM,細胞內染並以流式細胞儀 分析 T 細胞內 IFNγ 表現量
Fig. 6 (A)
(B)
PD-1
+
PD-1
-
PD-1
+
PD-1
-
HBV clearance m ice
HBV carrier mice unstimulated pAAV control 0
2 0 0 4 0 0 6 0 0 8 0 0 100
Fig. 6 抗共抑制性受體抗體 anti-PD-1 以及 anti-Tim-3 處理提高 HBV 感染實驗鼠 之肝內 T 細胞粒線體膜電位訊號
(A) 實驗鼠肝內淋巴細胞經過體外 HBV core 蛋白刺激,對照共抑制性受體抗體 anti-PD-1 以及 anti-Tim-3 處理以及不處理組以流式細胞儀偵測 MitoTracker Deep Red 訊號之強度
(B) 計算流式細胞儀偵測 MitoTracker Deep Red 訊號強度之 MFI 值,以直條圖顯 示 MFI 值(n=3),並以學生 t 檢定(Student's t-test)雙尾檢驗 P 值
Fig. 7 (A)
(B)
Fig. 7 MitoTEMPO 處理減少 CD8+ T 細胞粒線體中 ROS 以回復粒線體呼吸作用 能力並且提升IFNγ 的表現
(A) 從慢性感染小鼠取出肝內 T 細胞,處理 MitoTEMPO 並以體外 CD3、CD28 刺 激 48 小時,以流式細胞儀偵測 PD-1+ TIM-3- 和 PD-1+ TIM-3+ T 細胞族群 MitoTracker Deep Red 訊號,檢測粒線體的膜電位變化。
(B) 流式細胞儀偵測 CD8+ T 細胞內染 IFNγ 的表現
hi lo
Fig. 8 (A)
(B)
Fig. 8 抗氧化劑 MitoTEMPO 處理提高 HBV 感染之實驗小鼠之肝內 T 細胞 IFNγ 產量
(A)分離實驗小鼠肝內淋巴細胞,以 B 型肝炎病毒核心蛋白胜肽再刺激 24 小時以 及加入抗氧化劑、共抑制受體抗體處理,並以 ELISpot 偵測 IFNγ 分泌形成之斑 點數。
(B)計算細胞免疫斑點影像分析儀所計算之 ELISpot 斑點形成數目,並以學生 t 檢 定(Student's t-test)雙尾檢驗 P 值
參考資料
Barber, D.L., Wherry, E.J., Masopust, D., Zhu, B., Allison, J.P., Sharpe, A.H., Freeman,
G.J., and Ahmed, R. (2006). Restoring function in exhausted CD8 T cells during chronic
viral infection. Nature 439, 682-687.
Bengsch, B., Johnson, A.L., Kurachi, M., Odorizzi, P.M., Pauken, K.E., Attanasio, J.,
Stelekati, E., McLane, L.M., Paley, M.A., Delgoffe, G.M., et al. (2016). Bioenergetic
Insufficiencies Due to Metabolic Alterations Regulated by the Inhibitory Receptor PD-1
Are an Early Driver of CD8+ T Cell Exhaustion. Immunity 45, 358-373.
Chang, C.-H., Curtis, J.D., Maggi, L.B., Faubert, B., Villarino, A.V., O’Sullivan, D.,
Huang, S.C.-C., van der Windt, G.J.W., Blagih, J., Qiu, J., et al. (2013).
Posttranscriptional Control of T Cell Effector Function by Aerobic Glycolysis. Cell 153,
1239-1251.
Fisicaro, P., Barili, V., Montanini, B., Acerbi, G., Ferracin, M., Guerrieri, F., Salerno, D.,
Boni, C., Massari, M., Cavallo, M.C., et al. (2017). Targeting mitochondrial dysfunction
can restore antiviral activity of exhausted HBV-specific CD8 T cells in chronic hepatitis
B. Nat Med 23, 327-336.
Guo, C., Sun, L., Chen, X., and Zhang, D. (2013). Oxidative stress, mitochondrial
damage and neurodegenerative diseases. Neural Regeneration Research 8, 2003-2014.
Huang, Y.-H., Zhu, C., Kondo, Y., Anderson, A.C., Gandhi, A., Russell, A., Dougan,
S.K., Petersen, B.-S., Melum, E., Pertel, T., et al. (2015). CEACAM1 regulates TIM–3–
mediated tolerance and exhaustion. Nature 517, 386-390.
Jones, R.B., Ndhlovu, L.C., Barbour, J.D., Sheth, P.M., Jha, A.R., Long, B.R., Wong,
J.C., Satkunarajah, M., Schweneker, M., Chapman, J.M., et al. (2008). Tim-3 expression
defines a novel population of dysfunctional T cells with highly elevated frequencies in
progressive HIV-1 infection. The Journal of Experimental Medicine 205, 2763-2779.
Klein Geltink, R.I., O’Sullivan, D., Corrado, M., Bremser, A., Buck, M.D., Buescher,
J.M., Firat, E., Zhu, X., Niedermann, G., Caputa, G., et al. (2017). Mitochondrial
Priming by CD28. Cell 171, 385-397.e311.
Liang, T.J. (2009). Hepatitis B: The Virus and Disease. Hepatology (Baltimore, Md) 49,
S13-S21.
Nebbia, G., Peppa, D., Schurich, A., Khanna, P., Singh, H.D., Cheng, Y., Rosenberg, W.,
Dusheiko, G., Gilson, R., ChinAleong, J., et al. (2012). Upregulation of the Tim-
3/Galectin-9 Pathway of T Cell Exhaustion in Chronic Hepatitis B Virus Infection.
PLOS ONE 7, e47648.
Pauken, K.E., and Wherry, E.J. (2015). Overcoming T cell exhaustion in infection and
cancer. Trends in Immunology 36, 265-276.
Pearce, E.L., Poffenberger, M.C., Chang, C.-H., and Jones, R.G. (2013). Fueling
Immunity: Insights into Metabolism and Lymphocyte Function. Science (New York,
NY) 342, 1242454-1242454.
Sakuishi, K., Apetoh, L., Sullivan, J.M., Blazar, B.R., Kuchroo, V.K., and Anderson,
A.C. (2010). Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and
restore anti-tumor immunity. The Journal of Experimental Medicine 207, 2187.
Scharping, N.E., Menk, A.V., Moreci, R.S., Whetstone, R.D., Dadey, R.E., Watkins,
S.C., Ferris, R.L., and Delgoffe, G.M. (2016). The Tumor Microenvironment Represses
T Cell Mitochondrial Biogenesis to Drive Intratumoral T Cell Metabolic Insufficiency
and Dysfunction. Immunity 45, 374-388.
Schurich, A., Pallett, Laura J., Jajbhay, D., Wijngaarden, J., Otano, I., Gill, Upkar S.,
Hansi, N., Kennedy, Patrick T., Nastouli, E., Gilson, R., et al. (2016). Distinct Metabolic
Requirements of Exhausted and Functional Virus-Specific CD8 T Cells in the Same
Host. Cell Reports 16, 1243-1252.
Sena, Laura A., Li, S., Jairaman, A., Prakriya, M., Ezponda, T., Hildeman, David A.,
Wang, C.-R., Schumacker, Paul T., Licht, Jonathan D., Perlman, H., et al. (2013).
Mitochondria Are Required for Antigen-Specific T Cell Activation through Reactive
Oxygen Species Signaling. Immunity 38, 225-236.
Siska, P.J., Weinberg, J.B., Rao, A.V., Bain, J.R., Muehlbauer, M.J., Liu, T., and
Rathmell, J. (2014). Acute and Chronic Lymphocytic Leukemia Induces Exhaustion and
Suppresses Metabolic Reprogramming in T Cell Activation. Blood 124, 4121.
Ye, B., Liu, X., Li, X., Kong, H., Tian, L., and Chen, Y. (2015). T-cell exhaustion in
chronic hepatitis B infection: current knowledge and clinical significance. Cell Death
Dis 6, e1694.
Yin, Y., Metzger, T., and Bailey-Bucktrout, S. (2016). Tumor infiltrating T cells have
abnormal lipid metabolism that can be modulated by PD-L1 blockade. The Journal of
Immunology 196, 144.121.