國立臺灣大學理學院海洋研究所 碩士論文
Institute of Oceanography College of Science
National Taiwan University Master Thesis
臺灣近海康氏馬加鰆之年齡成長與季節性分布
Age, growth and seasonal distribution of narrow-barred Spanish mackerel (Scomberomorus commerson) in the water off Taiwan
羅允信 Yun-Sin Lo
指導教授:蕭仁傑 博士 Advisor: Jen-Chieh Shiao, Ph.D
中華民國 107 年 1 月
January, 2018
致謝
能夠完成本研究首要感謝指導教授蕭仁傑老師,兩年前我對海洋一無所知,
進來海研所後才發現只有膚淺的好奇是不夠的,還好蕭老師願意收留我,讓只是 純粹抱著嘗試心態的我有這麼有趣的主題可以研究,並且不斷指引我前進方向,
當指導教授真的很辛苦。另外也非常感謝水試所翁博士幫我採集大量的樣本,讓 我只需要專心地做研究,同時也提供許多土魠魚的基礎知識。天俠學長也提供不 小的貢獻,要是沒有學長編寫的程式,也不可能完成研究!感謝口試委員,每個 老師給的意見,都讓我的研究更有價值。在分析上,謝謝賴玲雯學的幫助,我一 波三折的研究過程需要更多的分析來嘗試,有賴玲雯學姊百忙之中安排分析時間
。也謝謝民弦一直被我叨擾,幫無助的我寫了很多程式擷取我需要的資料。除了 我研究部分,謝謝我的父母一直以來站在我的背後,給我溫暖的避風港,讓我在 想休息時,可以好好的沉澱。好同學們基哥腦殘粉和東海鼻涕妹,也讓實驗室天 天都很歡樂,我一定不會忘記我們一起欺負玻璃閃光喵和九四八七錞的日子,每 天都很期待這兩個活寶耍憨和鬥嘴。在夜深人靜的時候,也謝謝竹友會的各位,
看著你們講幹話,甚麼煩惱都可以拋下。斗笠哲、小朋友、馬基威和胖雞也謝謝 你們在我煩惱時,我知道隨時可以找你們聊幾句。變態保羅和變態跑步團,每次 耍憨講屁話也讓我可以傾吐心中的鬱悶,最後要感謝胖皮的陪伴,每次只要看到 憨憨的你,受不了我變臉而大笑時,好像困難都不見了。謝謝大家!我畢業啦~
摘要
康氏馬加鰆,又名土魠魚,是臺灣重要的經濟魚種。根據漁業年報的資料顯 示,澎湖的康氏馬加鰆產量已明顯減少,需要強化基礎研究以利漁業管理。本研 究以康氏馬加鰆的耳石邊緣明暗帶比例於各月份的消長,驗證耳石輪紋的周期性,
再以耳石輪紋估計的年齡和體長資料建立成長曲線和Age-Length Key。結果顯示 康氏馬加鰆耳石沉積暗帶的高峰期在 12 月到隔年 3 月,而康氏馬加鰆 von Bertalanffy growth equation 的參數𝐿∞為117.4 公分、K 為 0.5 year-1、t0為-1.04 歲,
0 到 3 歲年級群在 2016 與 2017 年佔了超過 90%。另外以耳石核心至邊緣的氧同 位素含量變化,推測康氏馬加鰆的季節性分布。結果顯示康氏馬加鰆的稚魚於春 末分布在臺灣西部沿岸以及澎湖到臺灣淺灘一帶,接著分布區域會在臺灣海峽中 部分裂後分別繼續往南海及東海移動與擴張,秋季時分布區域又在臺灣海峽中部 重合為一連續分布帶,在冬季時分布區域往南移動至南海,在春季時沿臺灣淺灘 外側經澎湖水道延伸至臺灣西部沿岸。康氏馬加鰆的季節性分布和溫度 23-25℃
分布帶的變化有密切關係,推測土魠魚的季節性分布和中國沿岸流與黑潮的鋒面 有關。
關鍵字:康氏馬加鰆、年齡成長、耳石穩定性同位素、季節性分布
Abstract
Narrow-barred Spanish mackerel, Scomberomorus commerson, is a highly valued food fish in Taiwan and is widely distributed from South Africa to Southeast Asia, north to China and Japan and south to southeast Australia. According to the fishery statistics reports, the catch of S. commerson dramatically drop after 2003 and enhancing fishery research and management are imperative. Totally 324 otoliths were examined to validate the period of opaque zone formation, which was the basis for the studies of age, growth and age compositions of S. commerson landed in Taiwan. The generated von Bertalanffy growth equation had L∞ = 117.4 cm, K = -0.5 year-1, t0 = -1.04 years. The age classes of 0-3 years constitute more than 90% of the catch in 2016 and 2017. Otolith stable oxygen isotope, sea sanility and water temperature was also used in estimating the seasonal distribution of S. commerson catched near Penghu island. The results suggested that the juvenile S. commerson were likely to distribute near Taiwan Banks and western coasts of Taiwan in spring, followed by the movement to East China Sea or South China Sea from spring to summer and returned to Taiwan strait in autumn.
Then the fish moved into South China Sea again in winter. The fish might return to Taiwan Banks in spring of following year. The seasonal distribution of the juvenile S.
commerson was closely associated with the mean water temperature of 23-25℃, which might be influenced by the Kuroshio and China coastal current.
Keywords : narrow-barred Spanish mackerel (Scomberomorus commerson), age and growth, otolith stable oxygen isotope, seasonal distribution
目錄
致謝... i
摘要... ii
Abstract ... iii
目錄... iv
表格目錄... vi
圖目錄... vii
壹、前言... 1
貳、材料方法 ... 5
2.1 樣本來源 ... 5
2.2 耳石樣本處理及分析 ... 5
2.2.1 耳石樣本製備 ... 6
2.2.2 年輪週期驗證、年輪判讀和日輪樣本處理 ... 6
2.2.3 耳石穩定性碳、氧同位素分析 ... 7
2.3 年齡與成長 ... 8
2.3.1 年齡估計 ... 8
2.3.2 年輪判讀和年齡估計精準度分析 ... 9
2.3.3 成長方程式 ... 10
2.3.4 年齡與長度對應表 ... 10
2.4 季節性分布估計 ... 10
2.4.1 穩定性氧同位素值對應日期估計 ... 10
2.4.2 臺灣周邊海域之耳石穩定性氧同位素分布估計 ... 11
叁、結果... 13
3.1 年輪週期驗證 ... 13
3.2 年齡與成長 ... 14
3.2.1 耳石年輪判讀精準度 ... 14
3.2.2 成長方程式 ... 14
3.2.3 年齡與長度對應表 ... 14
3.3 氧同位素變化趨勢 ... 15
3.4 季節性分布 ... 15
肆、討論... 18
4.1 年輪週期驗證 ... 18
4.2 年齡與成長 ... 20
4.3 漁獲年齡組成 ... 21
4.4 等高線頻率分析、平均差分析與方均根誤差分析之差異 ... 22
4.5 康氏馬加鰆之季節性分布與生活史 ... 24
4.6 耳石穩定性氧同位素與魚隻分布動態 ... 25
五、結論... 28
參考文獻... 29
表格目錄
表 1、土魠魚耳石樣本於各月份採樣數量。... 33 表 2、各月份所量測的土魠魚數量、體長範圍及平均體長。... 34 表 3、土魠魚耳石各暗帶與第一暗帶之最大與最小的間距。... 35 表 4、2016 年 1 月到 2017 年 4 月所採樣的 324 尾土魠魚所繪製之 Age-
Length Key。... 36 表 5、土魠魚的分布估計樣本之體長、年齡、分布估計起終日期和捕獲位
置。... 37 表 6、其他地區及本研究之土魠魚成長參數。... 38 表 7、其他地區與本研究之土魠魚 1-5 歲尾叉長估計和繁殖高峰期海表溫
度及葉綠素a 濃度。... 39
圖目錄
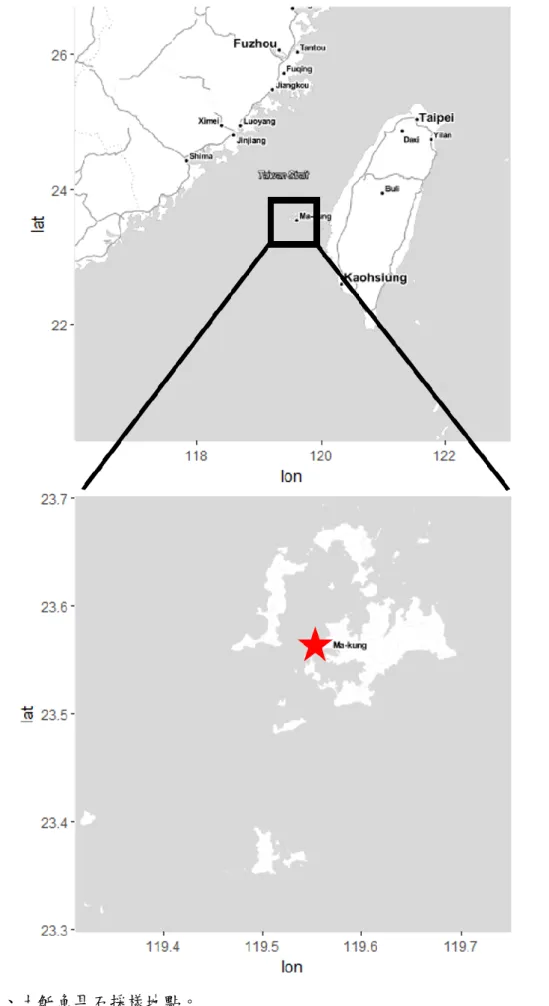
圖 1、土魠魚採樣地點... 41
圖 2、2016 年各月分體長頻度。... 42
圖 3、2017 年各月份體長頻度。... 43
圖 4、本研究樣本與 2016 及 2017 年度漁獲量測之體長頻度。... 44
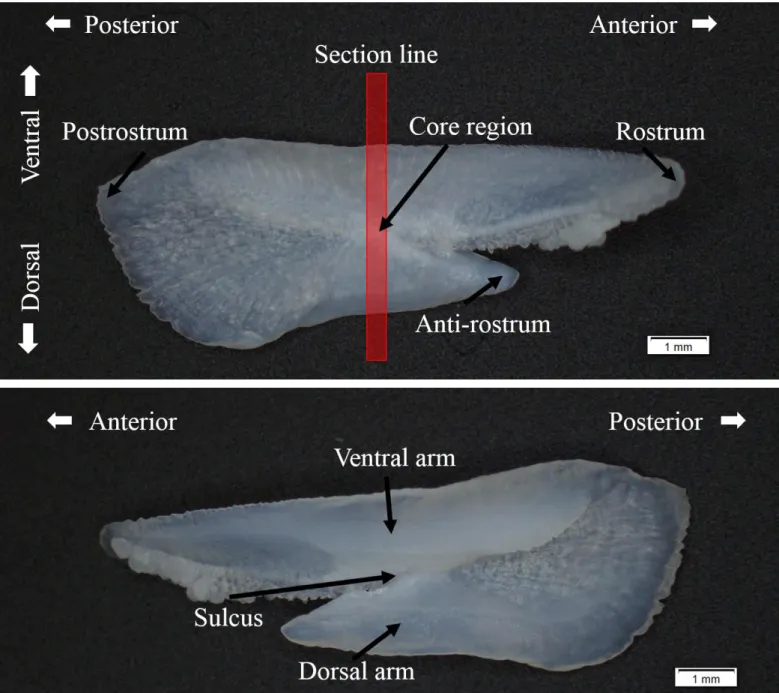
圖 5、土魠魚右側矢狀耳石構造。... 45
圖 6、土魠魚耳石橫切面構造。... 46
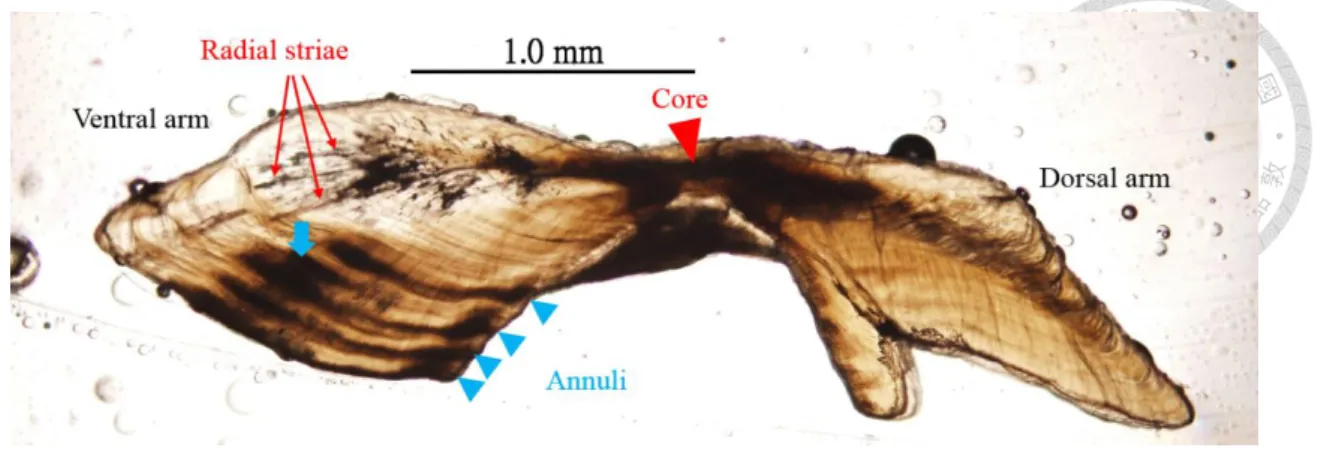
圖 7、土魠魚耳石輪紋分布測量。... 47
圖 8、土魠魚矢狀耳石橫切面核心結構。... 48
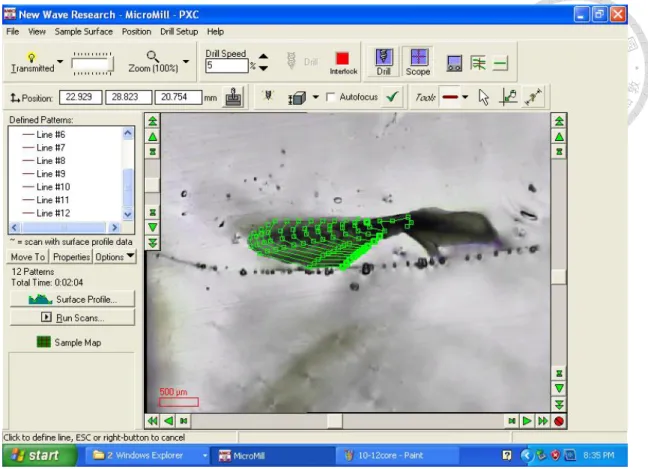
圖 9、耳石同位素分析取樣線。... 49
圖 10、10-12 月捕獲,邊緣為暗帶之土魠魚耳石樣本。 ... 50
圖 11、1-3 月捕獲,邊緣為暗帶之土魠魚耳石樣本。 ... 51
圖 12、4-6 月捕獲,邊緣為暗帶之土魠魚耳石樣本。 ... 52
圖 13、7-9 月捕獲,邊緣為暗帶之土魠魚耳石樣本。 ... 53
圖 14、4-9 月捕獲,邊緣為明帶之土魠魚耳石樣本。 ... 54
圖 15、1-3 月捕獲,邊緣為明帶之土魠魚耳石樣本。 ... 55
圖 16、10-12 月捕獲,邊緣為明帶之土魠魚耳石樣本。 ... 56
圖 17、土魠魚矢狀耳石橫切面的再礦化構造。... 57
圖 18、土魠魚耳石邊緣明暗帶分析(校正前)。 ... 58
圖 19、各輪紋與第一暗帶距離。... 59
圖 20、校正後(偽年輪不以暗帶處理)之邊緣明暗帶分析。 ... 60
圖 21、土魠魚成長曲線。... 61
圖 22、2016 年於澎湖海域捕獲土魠魚之各月份年齡組成。... 62
圖 23、2017 年於澎湖海域捕獲土魠魚之各月份年齡組成。... 63
圖 24、2016 及 2017 整年漁獲年齡組成... 64
圖 25、土魠魚耳石氧同位素變化趨勢。... 65
圖 26、土魠魚 3 號樣本之等高線頻率分布。... 67
圖 27、土魠魚 11 號樣本之等高線頻率分布。 ... 68
圖 28、土魠魚 200 號樣本之等高線頻率分布。... 69
圖 29、土魠魚 218 號樣本之等高線頻率分布。... 70
圖 30、土魠魚 219 號樣本之等高線頻率分布。... 71
圖 31、土魠魚 221 號樣本之等高線頻率分布。... 73
圖 32、土魠魚 274 號樣本之等高線頻率分布。... 74
圖 33、土魠魚 278 號樣本之等高線頻率分布。... 75
圖 34、土魠魚 304 號樣本之等高線頻率分布。... 76
圖 35、土魠魚 384 號樣本之等高線頻率分布。... 77
圖 36、土魠魚 385 號樣本之等高線頻率分布。... 78
圖 37、土魠魚 386 號樣本之等高線頻率分布。... 79
圖 38、土魠魚 3 號樣本之平均差(MD)分布。 ... 80
圖 39、土魠魚 11 號樣本之平均差(MD)分布。 ... 81
圖 40、土魠魚 200 號樣本之平均差(MD)分布。 ... 82
圖 41、土魠魚 218 號樣本之平均差(MD)分布。 ... 83
圖 42、土魠魚 219 號樣本之平均差(MD)分布。 ... 84
圖 43、土魠魚 221 號樣本之平均差(MD)分布。 ... 85
圖 44、土魠魚 274 號 樣本之平均差(MD)分布。 ... 86
圖 45、土魠魚 278 號樣本之平均差(MD)分布。 ... 87
圖 46、土魠魚 304 號樣本之平均差(MD)分布。 ... 88
圖 47、土魠魚 384 號樣本之平均差(MD)分布。 ... 89
圖 48、土魠魚 385 號樣本之平均差(MD)分布。 ... 90
圖 49、土魠魚 386 號樣本之平均差(MD)分布。 ... 91
圖 50、土魠魚 3 號樣本之方均根差(RMSD)分布。... 92
圖 51、土魠魚 11 號樣本之方均根差(RMSD)分布。 ... 93
圖 52、土魠魚 200 號樣本之方均根差(RMSD)分布。... 94
圖 53、土魠魚 218 號樣本之方均根差(RMSD)分布。... 95
圖 54、土魠魚 219 號樣本之方均根差(RMSD)分布。... 96
圖 55、土魠魚 221 號樣本之方均根差(RMSD)分布。... 97
圖 56、土魠魚 274 號樣本之方均根差(RMSD)分布。... 98
圖 57、土魠魚 278 號樣本之方均根差(RMSD)分布。... 99
圖 58、土魠魚 304 號樣本之方均根差(RMSD)分布。... 100
圖 59、土魠魚 384 號樣本之方均根差(RMSD)分布。... 101
圖 60、土魠魚 385 號樣本之方均根差(RMSD)分布。... 102
圖 61、土魠魚 386 號樣本之方均根差(RMSD)分布。... 103
圖 62、臺灣周邊海域各月份海葉綠素 a 濃度變化。... 104
圖 63、阿曼與南波斯灣在土魠魚繁殖高峰海表溫度及葉綠素 a 濃度。 .. 108
圖 64、澳洲西岸 Pilbara 及澳洲東岸 Queensland 在土魠魚繁殖高峰月份之 海表溫度及葉綠素a 濃度。... 109
圖 65、臺灣在土魠魚繁殖高峰月份之海表溫度及葉綠素 a 濃度。... 110
圖 66、阿曼沿海與南波斯灣在繁殖期高峰後的海表溫度。... 111
圖 67、臺灣周邊各月份 80 m 以淺平均溫度變化。 ... 112
壹、前言
康 氏 馬 加 鰆 屬 鱸 目 (Perciformes) 、 鯖 科 (Scombridae) 、 馬 加 鰆 屬 (Scomberomorus),主要棲息於印度-西太平洋,西自非洲東岸和紅海,東到澳洲 而北至日本、韓國(Kailola et al, 1993)。康氏馬加鰆為高經濟價值之食用魚種,
俗稱土魠魚,在臺灣主要漁具漁法為流刺網、曳繩釣及延繩釣,漁業統計年報 顯示1993 年到 2001 年之間的年捕獲量約在 2000-4000 公噸之間,在 2002-2003 年達到高峰,年捕獲量約達6000 公噸,隨後快速下降,從 2006 年到 2015 年間 的年捕獲量均未超過2000 公噸。
在中東的阿曼海域、澳洲東、西部及臺灣的繁殖期在春季,此時海溫逐漸 上升中(McPherson, 1993; Claereboudt et al., 2005; Mackie, 2005; 翁等, 2016),在 波斯灣的繁殖季稍長,跨越春夏兩季(Grandcourt et al., 2005)。土魠魚在一歲前 成長快速,以成長方程式估計,約可成長至60–70 cm 之間(Grandcourt et al., 2005;
Ballagh et al., 2006; Newman et al., 2012),一歲後成長明顯減緩,五歲之後成長 更為緩慢(Newman et al., 2012),土魠魚的壽命最大可以超過 20 歲(McIlwain et al., 2005)。過去研究中所估計的理論極限體長和成長參數有明顯差異,理論極 限體長在阿曼為 146.4 cm (McIlwain et al., 2005),在澳洲西岸的 Pilbara 則為 121.3 cm (Newman et al., 2012),而成長參數在南波斯灣為 0.21 year-1,而在澳洲 西岸的Pilbara 則高達 0.64 year-1 (Grandcourt et al., 2005; Newman et al., 2012)。
土魠魚的生活史和季節性分布似乎在不同區域有不同表現,土魠魚被認為 會進行沿岸移動(Kishinouye, 1923; Munro, 1942),但在澳洲東岸的族群同時存在 洄游和定居的個體(McPherson, 1981)。先前的研究根據臺灣沿岸的土魠魚漁獲 分布變化,推論土魠魚在秋季時來到臺灣北部並沿臺灣西岸移動至澎湖西南方,
在春季可能沿著中國沿岸往北移動或到中國南部沿海(曾等, 1971)。儘管土魠魚 是高度移動性的表層魚類,在澳洲北部仍存在三個土魠魚基因群(Buckworth et
al., 2007),前人也透過耳石年輪的可讀性、耳石穩定性同位素和寄生蟲的調查,
認為澳洲西部沿岸包含在空間上分離的土魠魚成魚族群(Lester et al., 2001;
Newman et al., 2009; Newman et al., 2012)。
儘管土魠魚初期成長速度快,成熟年齡低,因此有高生殖潛能(Newman et al., 2012),但 Shojaei et al. (2007)認為伊朗的土魠魚資源已被過度開發、Al-Hosni and Siddeek (1999)和 Govender et al. (2006)同樣報告土魠魚資源在阿曼也已經過 度開發。臺灣的土魠魚漁獲在漁業統計年報的資料中,從2002-2003 的高峰(約 6500 噸)之後就呈現連續 10 年低迷的漁獲量(均在 2000 噸以下)。漁業年報也顯 示,同為馬加鰆屬的高麗馬加鰆(Scomberomorus koreanus),在 2012 年之後幾乎 沒有野生的魚獲。因此在土魠魚的漁業管理與基礎研究十分急迫,需盡快透過 研究土魠魚的漁業生物學與生態資料,訂定保護臺灣土魠魚的漁業管理政策。
制定適合的漁業保護政策需要對目標物種的生活史與棲息環境有足夠的了 解,在研究洄游物種時,自浮式衛星標籤已用在大型洄游魚類如鮪魚和鯊魚,不 過此標籤體積較大,因此在研究體型較小的個體,則以傳統感應式或非電子標籤 作為研究方法(Thomasson et al., 2000),而 Thibault et al., (2007)曾以無線電標籤追 蹤美洲鰻在河川內的移動,化學或生物標籤則以不同於自然界的同位素比值的溶 液浸泡個體,在魚體組織,例如耳石上留下特殊元素訊號(Thorrold et al., 2002, 2006; Williamson, 2009),注射或浸泡氧化四環黴素(OTC, Oxy-tetracycline)或茜素 (ALC, Alizarin complexone)則可以在耳石留下螢光記號,但此標示方式須將標識 魚再捕獲才能進行後續分析。
耳石為硬骨魚內耳系統中控制魚類平衡和聽覺的器官 (Campana, 1999;
Parmentier et al., 2001),從胚胎時即開始形成直到死亡,耳石主要成分為碳酸鈣,
一經沉積後不會受到生理因素被再吸收而有所改變(Campana and Neilson, 1985)。
耳石沉積的微量元素可以反應魚體所經歷的環境(Radtke et al., 1998; Schwarcz et al., 1998),且許多硬骨魚類的耳石會因日夜變化形成日周輪,或在特定季節形成
的年輪,因此配合固定周期的輪紋和耳石從核心到邊緣的化學成分變化,可用於 硬骨魚類的生活史研究,如Shiao et al., (2010)透過在代表第一個冬季的耳石氧同 位素變化,辨認孵化自不同產卵場的太平洋黑鮪。Shephard et al., (2007)也發現在 不同深海地形捕獲的大西洋胸棘鯛在仔稚魚時期有不同的氧同位素變化,藉由耳 石氧同位素估計魚體經歷的溫度,再以水溫對應的深度,可以看出不同的深海魚 在仔稚魚時期有不同的深度變化,重建了不同魚類的環境利用策略(Lin et al., 2012)。
由於耳石中的穩定性氧同位素受到海水中的氧同位素和溫度影響(Thorrold et al., 1997; Campana, 1999),Hanson et al., (2010)利用東北大西洋的海水穩定性氧 同位素及溫度資料,推算出大西洋鮭魚在東北大西洋海域的耳石氧同位素估計值,
再由耳石分析所得氧同位素實測值,對應耳石氧同位素估計值的分布,得到該個 體在不同時間的分布海域。從該嘗試性的研究中可以看到,利用海水氧同位素及 溫度資料可以估計魚類於海洋中可能的棲息範圍,不過該研究在耳石氧同位素分 析值對應時間的方式較為簡略,以及過大的估計範圍仍難以看出大西洋鮭魚的洄 游動態。由於海水中的氧同位素比值受到鹽度的影響,因此本研究以臺灣周遭鹽 度換算成海水氧同位素(張, 2000),再透過太平洋黑鮪的耳石氧同位素分化公式 (Kitagawa et al., 2013),計算土魠魚在臺灣周遭可能的氧同位素估計值分布,再以 耳石氧同位素分析值對應的區域,作為土魠魚可能分佈的區域,其中耳石氧同位 素分析值的子樣本代表的日期以日輪估計,可能分佈的區域透過三種不同精準度 的計算,推測稚魚時期土魠魚的季節性分布,並移除於較不合理地形出現的雜訊,
嘗試提升以耳石氧同位素估計魚隻分布的準確性。
土魠魚在不同區域展現不同的生態習性和成長速度與極限體長,耳石年輪形 成的時間也有差異,如在昆士蘭的土魠魚耳石約在7–10 月形成年輪(McPherson, 1992),而在澳洲西部約在 11 月到隔年 3 月(Newman et al., 2012),通常耳石年輪
由耳石暗帶定義,而目前對於土魠魚耳石形成暗帶的機制尚不明確,且前人研究 中也指出有較為細窄的暗帶出現在年輪之間(McPherson, 1992; Govender, 1994)。
本研究的目的為驗證臺灣近海土魠魚耳石的輪紋週期,讓後續研究可以更精 準的判斷土魠魚的年齡與成長,提供資源評估的重要生物參數。同時以有限的耳 石樣本推估成長方程式,建構臺灣近海土魠魚的年齡和體長關係,並推估臺灣近 海捕獲的土魠魚各月份年齡組成。最後嘗試以耳石穩定性氧同位素估計臺灣近海 土魠魚的季節性分布,並推測年輕個體的棲息海域與生活史。
貳、材料方法
2.1 樣本來源
土魠魚的耳石樣本由行政院農委會水產試驗所翁進興副研究員提供,採樣方 法如下: 從 2016 年 1 月至 2017 年 4 月,每月定期向在臺灣海峽內作業、裝有 (Voyage data recorder, VDR)之流刺網標本船競標土魠魚標本,並將得標的樣本攜 至水產試驗所澎湖海洋生物研究中心進行尾叉長(精確至公分)、體重(精確至公斤) 測量,受限於經費,大部分樣本則是在漁市場或是漁民家中直接測量與採集耳石,
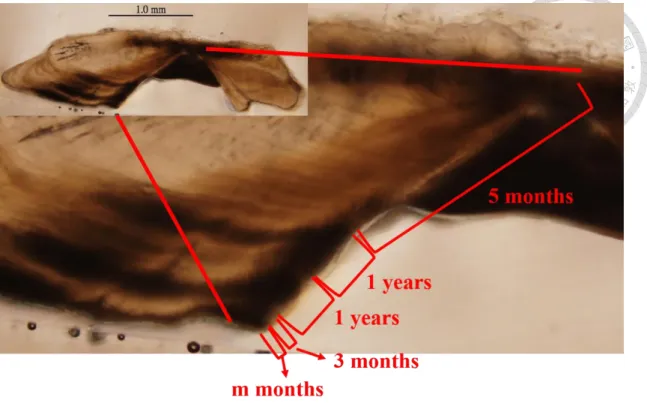
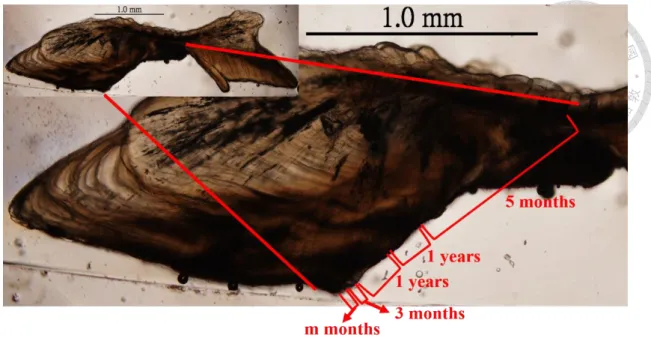
並未購買魚隻。各月份樣本數量如表1,尾叉長範圍為 34.5-128 cm,平均體長為 80.30±11.71 cm,體長頻度如圖 4,並取生殖腺及耳石,由於耳石末端較為細長 及脆弱,剖開時往往斷裂或遺失,因而選擇去除頭蓋骨方式進行,尋找最大的矢 狀石,頭蓋骨掀開後再以水清洗乾淨,發現矢狀耳石(sagitta)後以鑷子夾出,再以 清水洗淨自然烘乾後保存作為日後分析用,耳石構造如圖5。此耳石樣本將用在 耳石輪紋週期驗證、年齡評估、季節性分布估計上。
雌雄魚之判定乃是根據取出的生殖腺,以肉眼觀察性別。生殖腺為左右成對,
大小幾乎相同,外表為一卵膜所覆蓋,生殖腺上有生殖動脈,中央為靜脈,再分 出許多小動脈、靜脈及微血管,卵巢未成熟時成細條狀,成熟時卵巢飽滿呈黃色,
血管清晰可見,卵粒呈黃色或半透明色,肉眼明顯可觀察出。精巢則為白色或淡 紅色,生殖季節時呈飽滿且富彈性,輕押腹部時會有精液流出。
從 2016 年 1 月到 2017 年 12 月,各月份至少一天至馬公第三魚市場進行漁 獲體長量測(除了 2016 年 11 月僅有得標樣本的體長資料),從凌晨漁船進港開始,
將當天所有在馬公第三魚市場卸魚的土魠魚進行尾叉長量測,此尾叉長資料將配 合年齡與體長關係(Age-Length Key),評估臺灣土魠魚漁獲年齡組成。
2.2 耳石樣本處理及分析
2.2.1 耳石樣本製備
完整的土魠魚耳石外觀呈現於圖 5,Rostrum 往魚頭方向成長,往前方延伸 沉積。多數耳石樣本從 distal side 能較清楚的觀察到核心,核心大致位於長軸 (ventral arm)和短軸(dorsal arm)分岔的基部。將環氧樹脂和硬化劑(Struers, Demark) 以體積15 : 2 的比例進行灌注適量在模具作為底層,底層硬化後再置入耳石和樹 脂進行第一次包埋。以垂直rostrum 和 postrostrum 沿線的面,將經過一次包埋的 耳石連同核心以慢速切割機將350 μm 的厚度切下薄片,再以 permount 封片膠將 耳石切片封埋在載玻片與蓋玻片之間,接著觀察耳石年輪及判別邊緣明暗帶。
2.2.2 年輪週期驗證、年輪判讀和日輪樣本處理
本研究參考澳洲東岸 Queensland 對土魠魚的研究,以邊緣明暗帶在各月份 的比例變化作為週期驗證(McPherson, 1992),土魠魚耳石切片樣本呈現領結狀(圖 6),以核心連接長軸(ventral arm)和短軸(dorsal arm),年輪是根據長軸上的明暗帶 進行判讀,從核心往長軸邊緣300-400 μm 之後出現含有放射狀紋路的透明帶,
隨著在約600 μm 之後出現第一暗帶,接著成長變慢、方向改變,緊接著第二暗 帶出現,Lewis and Mackie (2002)指出土魠魚出生半年到一年間常有耳石暗帶連 續生成,本研究於輪紋觀察中,時常發現第二暗帶與第一或第三暗帶重疊,而第 三暗帶到第五暗帶之間,並沒有發現較明確的暗帶有重疊現象,如果將第二暗帶 判讀做年輪,將會出現二、三輪間距比一、二輪間距長,或是三、四輪間距比二、
三輪間距長的現象,而土魠魚的成長是逐年減緩的,這和過去研究認為耳石和個 體體型呈正相關的現象相反(Kasapoglu and Duzgunes , 2013),因此第二暗帶可能 為偽年輪,由於部分年輕個體並無出現和第一或第三暗帶重疊的第二暗帶,且無 第四或第五暗帶可做參考,故先以年齡較大且輪紋清楚的 19 片耳石樣本,以 image j 軟體測量第一暗帶、第二暗帶和第三暗帶的分布情形(圖 7),藉此判別偽 年輪出現的位置,幫助週期驗證的進行,並藉此減少年齡小的樣本,因較少輪紋 可做參考而把偽年輪判讀為年輪,造成高估年齡的誤差。
拍照記錄標本後,如該標本被選作季節性分布研究,將繼續從標本兩面小心 輪流研磨至核心露出標本表面如圖8,標本厚度同時也控制在 150-200 μm 之間,
此厚度的樣本在光學顯微鏡下所呈現的日輪最為清楚。
2.2.3 耳石穩定性氧同位素分析
將已完成一次包埋的耳石同樣連同核心以慢速切割機切下 350 μm 的薄片進 行二次包埋,再裁切成適當大小後以熱熔膠固定在載玻片上,以研磨機研磨至耳 石 表 面 完 全 露 出 。 接 著 以 電 腦 自 動 化 控 制 微 量 刮 取 器(micromill, ESI;
www.esi.com)沿著較明顯的成長紋路設定取樣路線,再內插數條取樣線條在取樣 路線之間增加時間序列解析度(圖 9),內插區域的取樣數量會以質譜儀可接受最 低限制約碳酸鈣體積0.016 mm3為限。
在蒐集粉末子樣本前先在耳石邊緣外刮出兩到三條取樣線寬度的溝槽,以利 之後蒐集子樣本,刮取器在耳石表面刮出粉末後,以解剖刀刀尖依序收集子樣本 的粉末到玻璃樣品瓶,並以石臘膜封住瓶口和編號,每條取樣線在刮取後都拍照 記錄,用在對照特定時序的同位素值所對應的成長紋路。
盛裝在玻璃樣品瓶的子樣本置入 Kiel Carbonate Device IV (Thermo Electron Corporation; www.thermoscientific.com)自動進樣系統,儀器會維持在 70˚C,並自 動將純磷酸滴入玻璃樣品瓶和耳石粉末反應,接著反覆升降溫去除反應產生的水 分子與其他的氣體,再將二氧化碳送進質譜儀(Finnigan MAT 253; Thermo Electron Corporation)進行碳氧同位素比值分析,分析的子樣本之間會穿插國際標準品 NBS19 (International Atomic Energy Agency, Vienna),用於校正樣品數值。同位素 數值是以VPDB (Vienna Pee Dee belemnite)為參考值表示,計算公式如下:
(1) δ =(Rsample - Rstandard) × Rstandard-1 × 1000(‰)
上式中R 為18O/16O 在樣本(sample)或標準品(standard)中的比值。標準品 NBS19 的穩定性氧同位素值為-2.2 ‰,標準品重複測定間精度約為 0.09 ‰。在分析完成
後,根據採樣區域的間格距離可以繪出耳石從核心到邊緣的穩定性氧同位素的時 序變化圖,藉穩定性氧同位素值變化趨勢推估土魠魚類棲息海域。
2.3 年齡與成長 2.3.1 年齡估計
Grandcourt et al. (2005)對波斯灣土魠魚的年齡估計,是以平均出生日期到第 一暗帶的時間,加上第一暗帶之後的暗帶以及最後暗帶到捕獲的時間,由於邊緣 暗帶的判別在暗帶逐漸出現和結束的季節較為不易判別,同一樣本有可能在不同 判讀中造成相差一輪暗帶的結果,進而讓年齡估計相差一歲,因此本研究將同月 份捕獲但邊緣沉積情況不同的讀輪結果,以不同的公式估計年齡,藉此消除邊緣 明暗帶判讀差異造成的誤差。本研究的年齡估計方式如下,土魠魚產卵期約為3- 6 月,5 月最多個體的卵巢達到性成熟,且 6 月開始母魚生殖腺出現退化卵細胞 (翁等, 2016),故本研究假設土魠魚的孵化日期是 5 月 1 日,所使用的年齡計算 公式詳細說明如下:
(2) age =5+12𝑛+(𝑚−9) 12
(3) age =5+12(𝑛−1)+(𝑚−9) 12
(4) age =5+12(𝑛−1)+(3+𝑚) 12
(5) age =5+12(𝑛−2)+(3+𝑚) 12
其中,n 為耳石年輪數量,m 為捕獲月份,由本研究的各月份邊緣明暗帶變 化可知明帶主要在4–9 月形成,10 月開始形成暗帶,持續至隔年 3 月。10-12 月 捕獲的土魠魚,若其耳石邊緣為暗帶者(圖 10),其年齡計算方式為:假定孵化日 為5 月 1 日到暗帶開始出現日期 10 月 1 日共 5 個月,加上第一暗帶之後的年輪 估計月份,為12 (n – 1)個月,再加上邊緣暗帶開始到捕獲的月份,為(m – 9)個 月,最後除以一年12 個月,此為第三公式。1 - 3 月捕獲的土魠魚,若其耳石邊
緣為暗帶者(圖 11),則年齡的估計方式為:孵化日 5 月 1 日到暗帶開始出現日期 10 月 1 日,共 5 個月,加上第一暗帶之後的年輪估計月份,為 12 (n – 1) 個月,
加上邊緣暗帶開始到該年結束的 3 個月,和隔年到捕獲月份的 m 個月,最後除 以一年 12 個月,此為第四公式。4–6 月捕獲的土魠魚,若其耳石邊緣為暗帶者 視為暗帶延後結束(圖 12),同樣以第四公式計算。而 7–9 月捕獲的土魠魚,若其 耳石邊緣為暗帶者(圖 13),視為暗帶提早出現,最後一組明暗週期未滿 1 年,故 以年輪估計的月份需再減一組明暗週期,由第四公式中12 (n – 1) 改為 12 (n – 2) 成為第五公式。
捕獲月份在4–9 月的土魠魚,若其耳石邊緣為明帶者(圖 14)以第四公式估計 年齡。捕獲月份在1–3 月的土魠魚,若其耳石邊緣為明帶(圖 15)者視為暗帶提早 結束,同樣以第四公式估計年齡。而10–12 月捕獲的土魠魚,若其耳石邊緣為明 帶者(圖 16)視為暗帶延後出現,應補算一輪暗帶,故同樣以核心到第一暗帶的 5 個月,加上明暗週期12 n (補算一輪暗帶,第三式的 12 (n - 1)改成 12 n),再加上 應該要出現暗帶的10 月到捕獲月份的時間(m - 9),最後總月分量除以一年 12 個 月,成為第二公式。第二公式的分子在移項之後等同第四公式,因此10–12 月捕 獲且邊緣為明帶的個體可直接由第四公式估計年齡而不需由第二公式估計年齡。
第五公式也在移項之後等同第三公式,因此7–9 月捕獲,耳石邊緣為暗帶者,可 直接以第三公式計算。所有情況可以第三或第四公式估計年齡。而有五尾小於50 cm 的樣本,以及用在季節性分布估計的 12 尾樣本,是直接以日輪除以一年 365 天作為年齡估計。
2.3.2 年輪判讀和年齡估計精準度分析
為了估計耳石判讀年輪以及估計年齡的精準度,以平均百分誤差(APE, average percent error) (Beamish and Founier, 1981)和變異係數(CV, Coefficient of variation) (Campana, 2001)對兩次讀輪的結果進行分析,APE 與 CV 的公式如下:
(6) APE = 100% ×1
𝑅∑ |𝑋𝑖𝑗−𝑋𝑗|
𝑋𝑗 𝑅𝑖=1
(7) CV = 100% ×
√∑ (𝑋𝑖𝑗−𝑋𝑗)2 𝑅−1 𝑅 𝑖=1
𝑋𝑗
上式中,R 為重複讀輪的次數,Xij為第i 次判讀時,第 j 個樣本的讀輪結果 或估計年齡,Xj是第j 個樣本讀輪結果或估計年齡的平均值。
2.3.3 成長方程式
本研究用 von Bertalanffy growth model (von Bertalanffy, 1938)以非線性最小 平方法套適所有耳石樣本的體長與年齡資料,以及公、母魚的體長與年齡資料,
其中有8 尾小於 1 歲的個體尚無法分辨性別,因此將這 8 尾年齡體長資料同時加 入公及母魚的年齡體長資料,並將所得之參數和其他研究比較,由於本研究的樣 本平均年齡為 1.36 歲,樣本體長頻度比漁獲體長頻度偏小,因此也和其他研究 比較由成長方程式所估計的土魠魚初期體長,其中von Bertalanffy growth model 公式如下:
(8) 𝐿𝑡= 𝐿∞(1 − 𝑒−𝐾(𝑡−𝑡0))
其中Lt為t 歲時的體長,L∞為理論極限體長,K 為成長係數,t0為體長為0 的年 齡。
2.3.4 年齡與長度對應表
根據耳石切片的讀輪結果和對應的尾叉長資料,建立以每10 cm 尾叉長為 間距的土魠魚Age-Length Key,並利用此 Age-Length Key 與馬公第三魚市場所 量測的土魠魚尾叉長資料(表 2、圖 2、圖 3),推測臺灣海峽內各月份的土魠魚 年齡組成。
2.4 季節性分布估計
2.4.1 穩定性氧同位素值對應日期估計
將內插取樣線區域所包含日輪,除以內插取樣線的數量,作為每個取樣線子 樣本包含之平均日輪,再以耳石最邊緣日輪對應捕獲日期,回推每個子樣本對應 日期,由於土魠魚耳石在核心區有再礦化(remineralization)現象,可能取樣時無法 完全避開,使一特定期間樣本受到其他時期的穩定同位素值干擾,故季節性分布 估計將排除發生耳石再礦化區域所對應之期間(圖 17)。
2.4.2 臺灣周邊海域之耳石穩定性氧同位素分布估計
根據子樣本對應日期,將每日海面下 10-80 m 之平均溫度和平均鹽度(取自 Copernicus:http://marine.copernicus.eu/)以下列公式估計康氏馬加鰆在臺灣周邊海 域每日耳石氧同位素分布,首先將鹽度換算成海水所含的穩定性氧同位素比值,
再以海水穩定性氧同位素值和溫度估計土魠魚耳石所沉積的穩定性同位素比值,
並移除水深超過500 m 之區域的估計(取自 NOAA:http://www.noaa.gov/):
(9) δ18Oseawater (VSMOW) = 0.385S(‰) − 13.05;R = 0.87 (張, 2000) (10) δ18Ootolith (VPDB) = δ18Oseawater (VSMOW) − 0.27T (℃) + 5.19;𝑟2 =
0.81 (Kiwataga et al., 2013)
上式中 S 為鹽度,T 為溫度,海水氧同位素以 Vienna Standard Mean Ocean Water (VSMOW)參考值表示,耳石氧同位素以 Vienna Pee Dee Belemnite (VPDB) 參考值表示。
再根據子樣本之δ18O 分析值,對照上式所估計的臺灣周邊耳石氧同位素分 布,藉此重建土魠魚可能的季節性分布,由於子樣本的氧同位素分析值為一段期 間內,耳石氧同位素的平均值,在該期間內每日的氧同位素值,應在該平均值上 下波動,因此該平均值等高線在此期間內的分布,可以做為土魠魚分布的參考,
故將臺灣周邊海域以長寬皆20 公里之網格劃區,並呈現對應期間每個網格有子 樣本分析值以等高線經過的頻率,作為等高線頻率圖。由於部分區域可能只在該 期間短暫出現平均值,因此另外嘗試直接藉由各區域每日的耳石估計值和該子樣 本的分析值做方均根差(root-mean-square deviation, RMSD)比較,了解哪些區域的
氧同位素估計值比較接近子樣本氧同位素分析值。不過目前對土魠魚所處環境的 溫度及鹽度變化梯度尚不了解,方均根差的計算會擴大差異,可能會排除溫度及 鹽度變化較大的區域,因此本研究也估計每個區域的耳石同位素估計值和該子樣 本的平均差分析(mean deviation, MD),作為平均差圖。方均根差估計及平均差估 計是以臺灣周邊海域之估計耳石δ18O 在對應期間內和子樣本 δ18O 分析值之方均 根誤差和平均差,公式分別如第十一式和第十二式:
(11) RMSD = √Σstart dateend date (δ18Oest oto−δ18Oanal oto)2 t
(12) MD =Σstart dateend date (δ18Oest oto−δ18Oanal oto) t
上式中start date 為子樣本對應起始日期,end date 為子樣本對應結束日期,
t 為子樣本對應天數,δ18Oest oto 是以每天平均溫度、平均鹽度估計之耳石氧同位
素,δ18Oanal oto是子樣本之氧穩定性同位素分析值。
再由Kitagawa et al. (2013)的報告所提供的數據,估計該溫度與同位素平均 值關係式的標準誤約為 0.14‰,因此從方均根差 0-0.1‰呈現紅到黃色,並在差 異大於0.11‰後由黃轉變成和陸地相同的白色,差異在約 0.2‰之後不再呈現顏 色,而平均差則從 0 到±0.1‰呈現紅到黃色,差異超過±0.11‰後由黃轉變成和 陸地相同的白色,差異在約±0.2‰之後不再呈現顏色。
叁、結果
3.1 年輪週期驗證
耳石邊緣呈現暗帶的情況在每個月的樣本中都有出現(圖 18),暗帶在 10 月 到隔年3 月出現比例均大於 60%,6、8 和 9 月的比例也高過 50%,其中 8 月更 超過70%,而耳石邊緣呈現明帶則在 3-11 月中出現,明帶高峰則出現在 4-7 月,
其中只有6 月明帶比例低於 60%。儘管在 12 月到隔年 3 月暗帶比例均高於 80%,
甚至 12 月到隔年 2 月暗帶比例高達 100%,在春夏之際的明暗帶並無較明顯的 消長趨勢。
從本研究的耳石橫切樣本可以觀察到,第二暗帶時常和第一或第三暗帶重疊,
像是從第一或第三暗帶分岔形成,出現第二、三暗帶間距比第一、二暗帶間距長 或是第三、四暗帶間距比第二、三暗帶長的不合理現象,非常類似 Lewis and Mackie (2002)所描述情形(第一和第二年輪之間的連續案帶沉積)。在經過各暗帶 與第一暗帶的距離測量後發現,第二暗帶距離第一暗帶最短為83 µm,最長為 160 µm,其中有超過 70%分布在 71-130 µm 之間(圖 19)。第三暗帶距離第一暗帶最 短為167 µm,最長為 246 µm,其中有超過 90%分布在 191-250 µm 之間。第四 暗帶與第一暗帶之距離最短為253 µm,最長為 384 µm,其中有超過 80%分布在 291-400 µm 之間。由於第二暗帶出現在本研究 5–10 月捕獲的樣本邊緣,距離第 一暗帶出現高峰的 11–3 月未滿一年,故將和第一暗帶距離 160 µm 以下的第二 暗帶判讀為偽年輪,並參照第三暗帶與第四暗帶的分布判讀第二年輪與第三年輪。
在將疑似偽年輪的第二暗帶判成明帶之後,結果如圖 20,耳石邊緣暗帶從 9 月開始出現,到隔年5 月結束,高峰期依舊在 12 月至隔年 3 月,均超過 80%而 12 月至隔年 2 月高達 100%。明帶高峰期為 5-9 月,百分比均高於 80%,其中 6- 8 月高達 100%,一年之中在 3-5 月以及 9-11 月同時有明帶及暗帶出現,且在 3-
5 月有明顯暗帶減少而明帶增加的趨勢,而在 9-11 月也有明顯的明帶減少同時 暗帶增加的趨勢,均勻地呈現一年由一暗帶及一明帶之輪紋所組成。
3.2 年齡與成長
3.2.1 耳石年輪判讀精準度
將經過相隔兩個星期的兩次讀輪結果互相比較,324 個年輪判讀標本中有 30 個樣本判讀結果不同,皆為相差1 歲。而以年齡估計公式估計年齡後,有 11 個 樣本有不同的年齡估計結果,兩次估計年齡的平均 APE 與平均 CV 分別為 0.83
%、1.18 %,由於兩次判讀結果差異相當小,本研究直接將第一次判讀結果用於 估計成長方程式,年齡範圍為0.24-5.08 years,平均年齡為 1.36 ± 0.62 year。
3.2.2 成長方程式
使用 von Bertalanffy growth model 對所有體長與年齡資料所套適得到的方程 式為 𝐿𝑡= 117.41 × (1 − 𝑒−0.50(𝑡−(−1.04))),L∞、K 和 t0分別為117.41 cm 、0.50 year-1、-1.04 year,從成長方程式可以估計出 1 歲之前成長快速,1 歲時可達 74.8 cm,隨後成長速率逐漸減緩,2 歲時可達 91.4 cm。雄魚的成長方程式為𝐿𝑡 = 118.84 × (1 − 𝑒−0.44(𝑡−(−1.23)))。L∞、K 和 t0分別為118.84 cm 、0.44 year-1、-1.23 year,而雌魚的成長方程式為𝐿𝑡= 109.55 × (1 − 𝑒−0.78(𝑡−(−0.48))),L∞、K 和 t0分 別為109.55 cm 、0.78 year-1、-0.48 year,成長曲線如圖 21。以成長方程式,雄 魚和雌魚在1 歲時尾叉長差異不大,之後逐漸變大,2 歲時差異可達約 4 公分,
之後差異逐漸變小,4 歲時尾叉長差異不到 1 cm,隨後差異再次拉開,5 歲時差 異可達3 公分。
3.2.3 年齡與長度對應表
根據2016 年 1 月到 2017 年 4 月採樣的 324 尾土魠魚之尾叉長,以 10 cm 為 間距,和每筆體長資料對應的年齡估計結果建立Age-Length Key (表 4),此 Age- Length Key 涵蓋長度範圍 30-130 cm。 2016 年和 2017 年間分別量測了 2859 尾
及3762 尾土魠魚之尾叉長。其中 2016 年的 11 月及 2017 年的 4 和 8 月的體長資 料不足100 尾,其餘月份均有 100 尾以上。
將各月份的尾叉長頻度分布經 Age-Length Key 轉換成年齡組成,各月份捕 獲的土魠魚年齡組成如圖22、圖 23 顯示,各月份漁獲並無明顯季節性變化,主 要由4 歲以下的個體所組成,除了 2017 年 6 月,1 歲年級群穩定在 40%以上,2 歲年級群大多在20%-30%之間,3 歲年級群幾乎都在 15%以下,而 4 歲以上個體 在各月分居未超過10%。2016 和 2017 整年漁獲年齡組成中(圖 24),1 歲個體約 佔50%,2 歲個體約 25%,3 歲個體 2016 年較少約 8%,2017 年則佔快 15%,4 歲以上個體均不超過10%。
3.3 氧同位素變化趨勢
在12 個耳石氧同位素分析中(圖 25),有 8 個耳石樣本(編號:3、218、274、
278、304、384、385、386)的氧同位素比例呈現相同的季節性趨勢,在夏季(7–9 月)達到最低值,δ18O 約在-2‰和-3‰之間,只有 218 號和 386 號樣本低於-3‰,
隨後逐漸升高,約在冬季(1–3 月)達到最高,δ18O 約在-0.5‰和-1.5‰之間,只有 218 號和 274 號樣本略低於-1.5 ‰。另有 4 個樣本的同位素比值變化較為特殊,
其中 200 號和 219 號同位素最低值出現在春季(4–6 月),δ18O 分別為-2.44‰和- 2.57‰,並在夏季(7–9 月)達到最高,δ18O 分別為-1.55 ‰和-1.68 ‰。11 號樣本在 秋季中期(11 月)才達到最低,δ18O 為-1.91 ‰,不過隨後也在冬季(1–3 月)達到最 高,δ18O 為-0.69 ‰。而 221 號樣本則是從春季到秋季幾乎都在-2‰上下些微波 動。
3.4 季節性分布
由於耳石在第一暗帶之後成長速率驟降,使得子樣本的時間解析度也跟著驟 降,故本研究接著以上述未滿一歲或一歲左右共 12 尾樣本的耳石同位素,進行 季節性分布估計,其中有2016 年 3 月捕獲的 3 號及 11 號樣本、2016 年 10 月捕
獲的200、218、219、221 號樣本,2017 年 2 月捕獲的 274 號及 304 號樣本,2107 年 3 月捕獲的 278 號樣本,以及 2017 年 5 月捕獲的 384、385、386 號樣本(表 5)。
以下季節性分布敘述從 2016 年春天開始,到 2017 年春天結束,可藉由 219 號(圖 30、圖 42、圖 54)接續 384 號(圖 35、圖 47、圖 59)的分布了解此段時間內 的連續性變化。其餘樣本可作為參考:2016 年春天請參考 200、219、221 號樣本 前期分布;2016 年夏天請參考 3 號初期,200、218、219、221 號中到後期,274、
304、385 初期;2016 年秋天請參考 3、274、385 號中期,11、278 號初到中期,
304 號中到後期,384 號初期;2016 年冬天請參考 3、11、274、304、385 號後 期,278、384 號中到後期;2017 年春天請參考 384、385、386 號最後分布。
在等高線頻率分布以及 MD 分布中,中國沿岸分布以及離岸的主要分布帶 有不同趨勢,中國沿岸在各季節常有分布,且該分布在冬季時向北迴縮,與曾等 (1971)觀察到的漁獲分布變化相反,由於溫度和耳石氧同位素呈現反比關係,而 鹽度和耳石氧同位素呈現正比關係,推測中國沿岸分布是受到較低鹽度影響而出 現的雜訊,因此下文中並不將中國沿岸分布變化加入敘述。
等高線頻率分布(圖 26-37)顯示稚魚在春末時主要分布於臺灣西部沿岸以及 澎湖經海峽中線到南海北部一帶,分布區域在夏季時可能有些個體往中國沿岸、
南海以及東海擴張,接著分布區域會在臺灣海峽中部分裂後分別繼續往南及往北 移動,秋季時分布區域又聚集在臺灣海峽中部形成連續分布,在冬季時分布區域 往南移動至南海,在春季時沿臺灣淺灘外側經澎湖水道延伸至臺灣西部沿岸。
MD 分布(圖 38-49)趨勢同等高線頻率變化趨勢,春末夏初之時,主要分布從 臺灣西部沿岸擴張到整個臺灣海峽,接著可能往中國沿岸、南海及東海繼續擴張,
在東海及南海均有大範圍分布之後,分布區域在臺灣海峽中部一分為二,秋季時 分布區域又從南海及東海往臺灣海峽內會合,冬季時分布區域再往南海移動,冬 末春初沿臺灣淺灘外側及澎湖水道延伸進入臺灣海峽。和等高線頻率分析相比,
分布面積在不同季節和區域都更為細窄,等高線頻率在 10%以上的區域幾乎都 有 MD 分布,不過在 MD 分布較大的區域不一定對應在等高線頻率 10%以上的 區域。
RMSD 分布(圖 50-61)情況無論是季節或是區域都較為零散,各季節之間的 分布連續性比等高線頻率的變化較不明確,不過基本上和 MD 分布中面積較大 或估計值較小的區域重疊,春末只在臺灣淺灘以及臺灣西部沿岸有絲狀分布,夏 季在東海及臺灣海峽出現零散的絲狀分布,臺灣淺灘的西南方出現稍大的片狀分 布,接著對應等高線頻率分布在東海面積最廣時,RMSD 在比沖繩海槽更西北側 的海域出現大面積分布,並且海峽內的中國沿岸出現絲狀分布,隨後沖繩海槽西 北側片狀分布消失,東海及臺灣海峽呈現零散絲狀分布,臺灣淺灘西南方有往澎 湖水道西南側延伸的漏斗狀分布,秋季時東海的絲狀分布進入臺灣海峽,在秋末 持續往西南移動進入南海。春季時再由南海經臺灣淺灘延伸至臺灣南部沿岸,
RMSD 分布面積比等高線頻率分布少了更多,RMSD 分布大致對應在 MD 分布 中面積較大和MD 值較接近 0 的區域,和以上兩中分析相比,最明顯的區域差異 是在東海的中國沿岸於四季幾乎沒有出現RMSD 分布。
肆、討論
4.1 年輪週期驗證
前人利用土魠魚耳石切片樣本的研究中,可由圖片看出第一年輪的判定略有 出入。有將反射光下,遠離sulcus 的 ventral side 上的暗帶判定年輪(Grandcourt et al., 2005),且此第一年輪寬度遠超過其他暗帶;也有在穿透光下沿著 ventral side 判讀年輪的方式(Newman et al.,2012),此第一年輪也明顯較其他年輪更細窄;還 有在穿透光下沿sulcus 一側判讀年輪的方式(Lewis and Mackie, 2002),此研究清 楚標示出偽年輪的型態,不過並無量化偽年輪和年輪之間的差異,但多數圖片仍 清楚的呈現年輪的間距將隨年齡越大而越窄(Lewis and Mackie, 2002;Grandcourt et al., 2005)。本研究的樣本在前三暗帶的型態大多無明顯差異,在年輪週期驗證 和年齡判讀上容易誤判,故先觀察年齡較大且輪紋清楚的樣本,各輪紋的分布情 形。在耳石暗帶間距觀察中,疑似偽年輪的第二暗帶約在第一暗帶後100 μm 左 右,而第三暗帶距離第一暗帶約在230 μm 前後。若將第二暗帶認定為年輪,則 第一、第二年輪的間距約100 μm,將比第二、三年輪的間距約 130 μm 來的窄。
如果不將第二暗帶讀作年輪,則第一、二年輪間距約為230 μm,第二、三年輪間 距約為 110 μm,此結果更符合絕大部分魚類耳石隨成長減緩而耳石年輪的間距 逐漸變窄的情形,且第二暗帶出現在5–10 月,而其他暗帶出現高峰在 12–3 月。
故本研究以19 片暗帶分布明確,且至少出現三暗帶的樣本中,最大之一、二暗 帶間距 160 μm 為界,將距離小於等於 160 μm 的第二暗帶判判定為偽年輪,當 邊緣暗帶與第一暗帶間距小於160 μm 則不以認定為代表年輪的暗帶,而是在第 一暗帶與第三暗帶之間的偽年輪。在過去的研究中也發現土魠魚的耳石一年可能 形成兩輪的紀錄,例如於澳洲昆士蘭海域1–5 月、南非海域 5 月的土魠魚形成之 暗帶比主要暗帶更為細窄(McPherson, 1992; Govender, 1994),因此在 McPherson (1992)的研究中被認定為偽年輪,而本研究所判定之偽年輪出現在 5-10 月。根據
Lewis and Mackie (2002)的研究,在寬的暗帶之間有較為細窄的暗帶,且隨年齡 增長,寬和窄的暗帶合併在一起,形成寬暗帶和窄明帶所組成的年輪周期。如果 後續研究包含更廣的年齡組成,將不同年級群之個體分開分析,或許可以看出從 年輕到年長樣本群,由兩個暗帶形成高峰逐漸併合成跨越更長時間的暗帶形成高 峰。
儘管在1、2、4 和 11 月所檢視的耳石樣本數小於 20 個,且多數樣本年齡不 超過2 歲,將偽年輪排除後,明暗帶同時出現的月份從 9 個月便縮短為 6 個月,
且原本明暗帶共存的9 個月中並無明顯的明暗消長趨勢,而在校正後,明暗共存 的6 個月分並非連續出現,而是在明帶與暗帶的高峰期之間逐漸改變比例,在暗 帶形成高峰前3 個月中,暗帶比例逐漸升高,暗帶形成高峰後 3 個月則暗帶比例 逐漸降低。除了呈現更準確的年輪周期,此校正也避免高估本研究中佔多數的年 輕個體的年齡,讓成長曲線更接近現實。
過去研究通常認為耳石形成的暗帶和溫度、光線條件、生殖或攝食有關 (Beckman and Wilson, 1995),而 Grandcourt et al. (2005)在南波斯灣觀察到土魠魚 耳石暗帶形成比環境升溫慢 2 個月,但趨勢和溫度變化似乎有關連,而在鄰近的 阿曼海域的土魠耳石暗帶主要在冬季形成(McIlwain et al., 2005),昆士蘭海域觀 察到土魠暗帶的形成則與繁殖季重疊(McPherson, 1992; McPherson, 1993)。於年 輪判讀時,穩定的周期能幫助更準確的年齡估計,因此本研究在年輪判讀時並不 包括偽年輪。然而藉由年輪形成的季節討論影響年輪形成的因子時,或許同時考 慮偽年輪形成的季節,能更準確的推測暗帶形成原因,未校正的各月邊緣明暗帶 比例顯示,明帶高峰期在春季,而暗帶從 8 月到隔年 3 月均占 50%以上,根據前 人研究,攝食量和溫度提升均會使鱈魚耳石產生不透明度較高的沉積(Høie et al., 2008),在校正前的結果顯示春季有較高的明帶沉積比例,此時在臺灣周邊葉綠 素 a 濃度大於 2 mg/m3的區域也較其他時期狹窄(圖 62,資料來源為 APDRC:
http://apdrc.soest.hawaii.edu/)。而秋季和冬季在臺灣周邊葉綠素 a 濃度大於 2
mg/m3的區域分布較廣、夏季海溫較高,或許暗示 8 月到隔年 3 月多數土魠魚有 較高的新陳代謝率,因此反應在耳石的輪紋結構上。
4.2 年齡與成長
在 前 人 的 研究 中 , 波 斯 灣 土魠 魚 耳 石 讀 輪 結 果 的平 均 APE 為 14.4%
(Grandcourt et al., 2005);在澳洲西部為 3.7 % (Newman et al., 2012);阿曼沿海為 2.8 % (McIlwain et al., 2005),澳洲東岸各年級群的平均 APE 有顯著差異,1–5 歲 各年級群的平均APE 約在 0.6–1.6 之間(Ballagh et al., 2006)。本研究中平均 APE 較小,可能是因為本研究多屬年輕個體,年輪數目較少,判讀差異的幅度被限制,
且年輕個體的輪紋間距較寬,不易因輪紋重疊而產生判讀誤差。本研究也透過定 義偽年輪的方式,避免高估年齡,同時透過年齡估計公式大幅降低邊緣判讀差異 造成的年齡差異。
本研究所採集的樣本中,最年輕的個體約為0.24 歲,體長為 37.5 cm,年紀
最大的個體約為5.08 歲,體長為 128 cm,其中只有 6 尾樣本年紀低於 0.5 歲。
套適von Bertalanffy growth model 所得到的參數:L∞、K、t0,分別為117.41 cm、
0.50 year-1、-1.04 year,將此結果與前人在其他海域的研究結果比較,澳洲東岸 的Queensland (Ballagh et al., 2006)、西岸的 Pilbara (Newman et al., 2012)、阿曼 (McIlwain et al., 2005)、波斯灣(Grandcourt et al., 2005)所估計的土魠魚 von Bertalanffy growth model 的參數如表 6。其中土魠魚在阿曼和波斯灣的理論極限 體長均在140 cm 上下,而在澳洲東岸的 Queensland (Ballagh et al., 2006)、西岸 的Pilbara (Newman et al., 2012)和本研究的理論極限體長均在 120 cm 上下。除了 理論極限體長在各地區有差異之外,過去研究曾指出雌魚成長速率較慢,但有比 雄魚還高的理論極限體長,推測是由不同性別投資在繁殖的能量比例不同所造成 (McPherson, 1992; McIlwain et al., 2005),而本研究的結果正好相反,雄魚成長比 雌魚緩慢,但雄魚有更高的理論極限體長,或許是本研究的高齡個體不足所致,
由本研究樣本的體長頻度和漁獲體長頻度相比,樣本包含的體長偏小(圖 4),最
大體長為128 cm,用於 Age-Length key 的體長量測資料中,最大體長為 152 cm,
因此理論極限體長和成長參數有待未來加入更多高齡個體後再評估。不過在以上 研究中均充足的年輕樣本中,由成長方程式所估計的土魠魚初期成長,在第一年 已經可以看出差異(表 7)。由前人研究可以知道土魠魚在澳洲西岸的 Pilbara、澳 洲東岸的Queensland、南波斯灣、阿曼以及臺灣的繁殖期高峰分別在十月、十一 月、六月、五月以及五月(Grandcourt et al., 2005; McIlwain et al., 2005; Ballagh et al., 2006; Newman et al., 2012)。由以上月份的近海海表溫度和葉綠素 a 濃度(表 7、
圖63、圖 64、圖 65,資料來源為 APDRC:http://apdrc.soest.hawaii.edu/),可以 看出除了南波斯灣外,越溫暖以及葉綠素a 濃度越高的海域,土魠魚在 1 歲可以 有越大的尾叉長,如阿曼沿岸海域的繁殖季高峰海表溫度約可達 30˚C,且葉綠 素a 濃度可超過 4 mg/m3,其土魠魚一歲時估計體長可超過75 cm;而在澳洲東 岸Queensland 的繁殖季海表溫度較低約在 26-29˚C,葉綠素 a 濃度約在 1 mg/m3 以下,其土魠魚一歲估計體長則低於65 cm。Neuheimer et al. (2011)的研究中指 出Cheilodactylus spectabilis 在超過適合成長的水溫後,成長會減緩。從圖 61 可 以看出比起阿曼海域,南波斯灣在土魠魚繁殖季之後的海表溫度持續上升,可達 33˚C 上下,或許此水溫已經超過最適合土魠魚仔稚魚成長的溫度範圍,進而反 應在初期較慢的成長上。
4.3 漁獲年齡組成
臺灣在 2016 年到 2017 年的土魠魚漁獲年齡組成較無明顯的季節性變化,且 目前對於此土魠魚族群有多少比例會進行遷徙尚不明確,較難推測臺灣的土魠魚 漁獲年齡組成變化受到哪些不同因子的影響,不過仍可以看出土魠魚在1 歲左右 加入流刺網漁業,主要漁獲年齡為 4 歲以下之個體。從本研究的耳石樣本觀察 中,秋季捕獲體長60-70 cm,且耳石尚未出現第一暗帶的個體,尚未經歷第一個 冬季,而土魠魚繁殖高峰在隔年3-5 月(翁等, 2016),代表秋季捕獲 60-70 cm 之 個體尚未經歷第一次繁殖就被捕獲,這暗示臺灣土魠魚捕獲量逐年減少或許跟網
目過小有關,不過本研究的樣本少有大於4 歲的個體,因此建構出的 Age-Length Key 可能低估高齡魚的比例,有待未來加入更多高齡魚的年齡體長資料構築 Age- Length Key,並加入更多年份的體長頻度資料估計臺灣捕獲土魠魚的年齡組成。
4.4 等高線頻率分析、平均差分析與方均根誤差分析之差異
本研究所使用的三種分布估計方式在原理上的差異也呈現在估計結果上,
以下將從原理出發討論估計結果的解讀。
等高線頻率分析呈現所有分布機率大於零的區域,而實測的耳石氧同位素 涵蓋日期從幾天到一、兩個月不等,只要耳石分析值於該期間均在估計同位素 變化範圍內,等高線頻率圖即會顯示在該海域有出現此魚體,估計分布結果將 隨時間越長而有越大的分布面積,雖然這個性質讓單一子樣本所反映的分布較 難解讀,卻可以在連續的子樣本對應分布之間有更好的連續性,這讓子樣本對 應時間較長或在游泳能力越好的物種的分析中更易了解遷徙的動向。
方均根誤差(RMSD)分析是同時要求精度與準度的分析,只有在分析日期內 的每日估計氧同位素都接近耳石氧同位素分析值的區域才會呈現魚體出現在該 海域中,因此當分析時間越長或環境中的溫鹽變化梯度越大,估計魚隻的分布面 積將越小。在分析涵蓋時間越長,或環境溫鹽變化梯度大的情況,此性質將降低 分布呈現的準確性。而在分析時間越短或適溫性洄游物種以及游泳能力較低的物 種分析中,能更準確地呈現可能的分布區域。如304 號樣本的夏季子樣本對應日 期較秋季短,因此 RMSD 分布與等高線頻率分布的差異在夏季差異較小,而秋 季差異較大,9/11 – 9/15,304 號樣本在這四天的 RMSD 分布與等高線頻率分布 一致性高,均在東海有大面積分布而在臺灣海峽及南海有零星分布,而在 10/29 – 11/29 一整個月中,在沖繩海槽兩側、中國沿岸經臺灣海峽到南海北端有大面 積等高線頻率分布,而RMSD 分布只零星出現在臺灣周邊和沖繩海槽兩側(圖 34、
圖58)。另外編號 274 的樣本在夏季與冬季的子樣本對應的期間長度差異不大,
因此 RMSD 分布與等高線頻率分布在夏季與冬季都有明顯差異。274 號樣本在
8/2 - 8/30 之間,等高線頻率分布從中國沿岸延伸入臺灣海峽,在同一區域 RMSD 僅有零星分布。在隔年1/3 – 2/3 之間,等高線頻率分布出現在中國沿岸而 RMSD 並無在臺灣周邊出現(圖 32、圖 56)。
平均差分析(MD)是要求準度而不要求精度的分析,因此溫鹽變化梯度大和 分析時間越長對平均差分析的影響比方均根誤差分析小。而由於氧同位素值的變 化和鹽度成正比且和溫度成反比,因此往溫暖海域的移動和往鹽度較低海域移動 將無法區分,互相成為雜訊,如編號384 的樣本在冬季時,在臺灣海峽內的分布 隨著溫暖水團逐漸往南移動,而中國沿岸分布則逐漸往北移動至更低鹽度的區域 (圖 47)。因此在已經確定鹽度或溫度範圍而能剔除雜訊的情況下,這些性質讓平 均差分析較適用於生活在溫度或鹽度變化梯度大的物種上。如大西洋鮭魚在夏季 往低鹽度的河川上游移動(Hanson et al., 2010),此時的分析就可以剔除往溫暖海 域移動的分布,進而得到更準確的估計。
由於土魠魚耳石中的氧同位素含量是在空間及時間都有變化的情況下所得 到的平均值,而MD 分布和 RMSD 分布是以每個定點的溫度及鹽度隨時間變化 所做估計,因此 MD 分布應該是核心區域,而實際分布可能比 MD 分布還大。
等高線頻率估計則能包含時間和空間的變化,這也是等高線分布和實際捕獲位置 重疊率較高的原因。未來期望能將樣本分析所得的氧同位素平均值之標準偏差和 更多篩選因子納入分析中。
在等高線分布估計中,共有 7 尾樣本的實際捕獲位置與捕獲前估計分布相重 疊,4 尾樣本的實際捕獲位置在捕獲前估計分布的鄰近區域;在 MD 分布中,有 4 尾樣本的實際捕獲位置與捕獲前估計分布相重疊,7 尾樣本的實際捕獲位置在 捕獲前估計分布的鄰近區域;在 RMSD 分布中有 3 尾樣本的實際捕獲位置與捕 獲前估計分布相重疊,8 尾樣本的實際捕獲位置在捕獲前估計分布的鄰近區域。
編號11、304、384、385 樣本的實際捕獲位置與等高線頻率分布相重疊,不過是 在頻率20%以下的區域,這幾尾樣本的實際捕獲位置則在估計範圍更縮窄的 MD
以及RMSD 分布的鄰近海域。在 12 尾樣本的三種分析估計中,有 11 尾樣本的 實際捕獲位置,與捕獲前估計分布相重疊或在鄰近區域,顯示此估計方式有相當 程度的可靠性,只有274 號樣本較為例外,實際捕獲位置並不在三種估計分布的 鄰近海域,估計分布應在更為溫暖的南海海域,這暗示土魠魚可能在短期內進行 長距離遷徙,從南海移動到臺灣海峽被捕獲。
4.5 康氏馬加鰆之季節性分布與生活史
本研究以鹽度及溫度資料推算不同時空下土魠魚耳石氧同位素比值,藉此比 對實測的耳石氧同位素分析值在對應日期內所代表的魚隻分布海域,以下透過溫 度(取自 Copernicus:http://marine.copernicus.eu/)的季節變化討論土魠魚的棲息海 域變化。
80 m 以淺的平均溫度 23-25˚C 分布的四季變化和等高線頻率分布變化一致 性相當高(圖 67 中淺綠色分布),春季時此溫度帶分布從臺灣淺灘延伸經澎湖水 道至臺灣西部沿岸,春末時往中國沿岸及東海擴張,夏季時在臺灣海峽內分裂為 不連續分布並分別繼續往南和往北擴張。秋季時此溫度帶在東海的分布逐漸退回 臺灣海峽,秋末時從臺灣西部沿岸經澎湖延伸到臺灣淺灘。冬季時分布在澎湖周 遭延伸往臺灣淺灘。春季再度從臺灣西部沿岸向北延伸。
從溫度分布搭配土魠魚估計分布之間的關係,以及前人所描述的臺灣海峽流 況變化(Jan et al., 2002),推測土魠魚可能會隨著黑潮支流和中國沿岸流的鋒面在 東海到南海之間遷徙,中國沿岸流的鹽度及溫度均較低,但受到河水注入影響而 有較高的營養鹽含量(Liu et al., 2000)。黑潮支流的溫度及鹽度均較高、營養鹽含 量較低(Chung et al., 2001),土魠魚跟隨鋒面遷徙則可在溫暖且生產力較高的環境 快速成長,由此推論土魠魚的早期生活史從臺灣淺灘孵化開始,春季由臺灣暖流 送往東海,夏季的東海因為黑潮及長江沖淡水注入成為溫暖又且生產力高的育幼 地(Gong et al., 2011),稚魚在此得以快速成長,秋季東海開始降溫,喜好暖水域 的土魠魚則沿著黑潮支流與中國沿岸流的鋒面進入臺灣海峽,冬季時臺灣海峽繼
續降溫,部分個體進入南海避冬,隔年春季回到臺灣淺堆,春季正是土魠魚的繁 殖季(翁等, 2016)。過去研究中,秋季臺灣西北沿岸開始有土魠魚漁獲,捕獲分布 最北處到淡水沿岸,隨著溫度繼續降低,漁獲分布逐漸南移集中到澎湖周邊海域,
冬季時漁獲稍減,春季時澎湖周邊海域漁獲又再增加,此時在新竹沿岸也出現土 魠魚漁獲(曾等,1971),此紀錄相當符合本研究所估計的季節性分布變化。前人也 有發現澳洲的康氏馬加鰆族群中有不進行遷徙的個體(Collette, 2001),而澎湖周 遭海域整年均可捕獲土魠魚,推測原因為澎湖周遭有生產力高的大面積珊瑚礁,
加上整年均有23-27˚C 之間的溫暖海水。
編號 386 的樣本其季節性分布較為不同,在春季繁殖期之前,在受中國沿岸 流影響而鹽度較低的區域,以及在超出本研究估計範圍之外的更南方有分布,春 季繁殖期開始才出現在臺灣周邊海域,從南海北端延伸入臺灣海峽。由於一尾魚 只能在同一時間出現在同一海域,中國沿岸之分布是因鹽度較低,而形成如同分 布在溫暖海域的雜訊,排除雜訊後,這樣的分布變化暗示在更南方有其他土魠魚 族群與本研究的土魠魚族群有基因交流。
4.6 耳石穩定性氧同位素與魚隻分布動態
前人利用耳石穩定性氧同位素估計魚類於海洋中棲息海域的方法與本研究 有些微差異(Hanson et al., 2010; Sakamoto et al., unpublished),以下針對耳石處理、
同位素分析和日期對應進行討論。
在耳石處理上,Hanson et al. (2010)和 Sakamoto et al. (unpublished)均使用平 磨(saggital plane)的方式,使核心露出表面。而本研究是以橫切(transverse plane) 再研磨使耳石核心露出。其中以平磨處理,將有更足夠的面積使用 micromill 進 行採樣,進而提升時間解析度,不過土魠魚的耳石結構較為特殊且立體,較難使 用平磨同時讓核心與邊緣在最大面積的研磨面上出現,並且同時保有足夠厚度進 行 micromill 採樣。另外,土魠魚耳石從橫切面可以清楚觀察到再礦化區域,而 無法從平磨樣本中輕易觀察到,這可能造成核心區域的氧同位素值遭受魚隻初期
生活史以外的氧同位素訊號干擾,耳石的平磨與橫切各有利弊,端看目標物種的 耳石結構而定。
在同位素分析時,Hanson et al. (2010)使用二次離子質譜儀(Secondary ion mass spectrometer, SIMS),可以分析極小範圍內(約 15 μm)的同位素組成,並觀察 到大西洋鮭魚從淡水進到海水時,明顯的氧同位素比值躍昇。而在本研究與 Sakamoto et al. (unpublished)的研究中是使用 micromill 採樣,再以同位素比質譜 儀(Isotope Ratio Mass Spectrometry, IRMS)進行分析。Hanson et al. (2010)的研究中 指出IRMS 所得到的同位素訊號精確度比 SIMS 高,但 IRMS 需要的樣本質量也 比 SIMS 高,而 SIMS 的技術也在提升當中。以 micromill 採樣雖然時間解析度 會降低,但仍足夠看出基本的季節性分布。由於SIMS 分析的樣本只會在表面留 下點狀的軌跡,仍可以由分析後的樣本判讀日輪,因此未來如能以橫切耳石樣本 搭配更精準的SIMS,將能避開再礦化區域,並更準確的對應每個子樣本所代表 的時間。
在子樣本對應日期上,Hanson et al. (2010)是以第一個同位素比值躍昇的訊 號作為進入海水的訊號並對應上多數幼鮭進入海水的五月,再以最大的氧同位素 比值對應上最冷的二月,然而在冬季大西洋鮭魚實際分布在更北方,Hanson et al.
(2010)則認為或許是近似物種的同位素分化公式和所研究的大西洋鮭魚不同。不 過在本研究以及Sakamoto et al. (unpublished)的研究中可以看到同位素比值上升,
不一定代表越接近冬季,可能是個體往低溫或是往高鹽度區域移動。因此以日輪 估計子樣本代表的時間或許是更可靠的做法。Sakamoto et al. (unpublished)對應子 樣本代表日期的方式是以子樣本所跨越的日期的中位數作為子樣本的日期,在時 間解析度可接受的情況,這是更方便的處理,然而本研究是以時間解析度較低的 橫切耳石樣本進行採樣。加上土魠魚游泳能力好、臺灣周遭海域受溫鹽性質差異 極大的黑潮及中國沿岸流影響,每個子樣本代表日期內的實際同位素變化難以估 計,或許不適合此對應日期的方式。
Hanson et al. (2010)最後採納的同位素分化公式所估計的魚隻分布,緯度均 跨越10 度以上,其餘公式所估計的魚隻分布緯度超過 10 度則占少數。由於估計 魚隻分布的窄或廣同時受同位素分化公式的標準差、同位素分化公式的斜率及估 計的耳石同位素分布梯度所影響,且該研究並無明確描述分布估計方式,因此難 以藉由該研究討論更加精準的估計方式。不過該研究嘗試不同同位素分化公式的 結果顯示,同一分化公式所估計的魚隻分布在不同時間有不同的跨越緯度,可以 推測估計的耳石同位素分布梯度對魚隻的估計分布有重要的影響。
在 Hanson et al. (2010)及 Sakamoto et al. (unpublished)的研究中,都期待能將 估計出的分布範圍再縮小,尤其在經度變化的解析度,能符合更實際的情況。在 本研究中,透過臺灣捕獲土魠魚的分布和fishbase 所呈現的土魠魚分布範圍,推 測土魠魚生活在陸棚區域,因此遮蔽了臺灣灣周遭深度超過500 m 的地形,同時 在解讀上避開渡冬移動卻出現往低鹽度移動的雜訊,大幅減少了較為不合理的分 布,如能對目標物種的習性更為了解,將能提升此分布估計方式的準確性。期待 未來在分析上,可以藉由更高時間解析度的同位素分析,搭配同位素比值在子樣 本涵蓋時間內的標準差作為估計分布的篩選條件,或許能增加經度上的解析度,
提升此具有潛力的方法的實用性。