國立台灣大學醫學院生物化學暨分子生物學研究所 碩士論文
Graduate Institute of Biochemistry and Molecular Biology College of Medicine
National Taiwan University Master Thesis
棕櫚酸促進陰電性低密度脂蛋白在巨噬細胞中引起 IL-1產 生透過增加鉀離子流出
Palmitic acid enhances electronegative LDL-induced production of IL-1 by increasing potassium efflux in macrophages
張庭瑜 Ting-Yu Chang
指導教授:呂紹俊 博士 Advisor: Shao-Chun Lu, Ph.D.
中華民國 一百零七年七月
July, 2018
口試委員審定書
摘要
近年來,由於飲食習慣改變,肥胖率越來越高,肥胖者為代謝症候群的高危險 族群,容易罹患第二型糖尿病、心血管疾病,如動脈粥狀硬化與心肌梗塞等。這些 病人血液中,陰電性低密度脂蛋白(LDL(-))、游離脂肪酸及介白激素 1 (IL-1) 都 明顯提高,巨噬細胞分泌的 IL-1在這些疾病扮演重要的角色,因此在這個研究我 們探討LDL(-) 與游離脂肪酸在巨噬細胞引起之 IL-1產生。
IL-1的產生需要兩條訊號:第一,啟動訊號為 NF-κB 活化,接著 NF-κB 作 為轉錄因子促進 pro-IL-1β 和 NLRP3 的表現;第二,激活訊號為 NLRP3 發炎小 體活化,導致 Caspase-1 被切割且活化後,將 pro-IL-1β 切割轉變為成熟型 IL-1β。
先前實驗室研究指出,LDL(-) 在巨噬細胞能誘導 NF-κB 及 Caspase-1 活化,但 Caspase-1 活化程度並不高。另外,游離脂肪酸被認為可以促進 NLRP3 發炎小體 活化。因此,我們想探討 LDL(-)、游離脂肪酸和 IL-1β 在發炎反應之間的交互作 用,且是否游離脂肪酸能提高 LDL(-) 在巨噬細胞誘導之 IL-1β 產生。
我們從餵食高膽固醇脂飲食之兔子的血漿分離到 LDL(-),將其與飽和或不飽 和游離脂肪酸一起處理人類單核球細胞 (THP-1) 分化之巨噬細胞,分析分泌到細 胞培養液中的 IL-1β 含量。結果顯示,棕櫚酸單獨不能造成 IL-1β 產生,且其沒 有誘導 NF-B 活化之能力;但能促進 LDL(-) 誘導之 IL-1β 產生。而 LDL(-) 會 誘導清道夫受體 LOX-1 表現增加,LOX-1 也被指出和發炎反應有密切關係,然 而棕櫚酸並不會進一步透過促進 LOX-1 表現,而增加 IL-1β 產生。將細胞培養 在無鉀離子之培養基下可以使 LDL(-) 誘導之 IL-1β 達到 LDL(-) 與棕櫚酸一起 處理的程度。相對的,幾種非專一性的鉀離子通道抑制劑會抑制 LDL(-) 與棕櫚酸 一起所誘導之 IL-1β。這些結果顯示,棕櫚酸可能是透過促進鉀離子流出細胞之作 用,造成 NLRP3 發炎小體與 caspase-1 進一步活化,而使 IL-1β 產生增加。
總結,在我們的研究發現LDL(-) 和棕櫚酸對於 IL-1β 產生有協同效應,並且
發現棕櫚酸會提高 LDL(-) 誘導 IL-1β 產生之機制可能是增加鉀離子排出。因此 我們認為病人血液中若是 LDL(-) 和飽和游離脂肪酸升高,可能會引起 IL-1β 大量 產生,是引起發炎的關鍵因素。但我們並不清楚棕櫚酸是影響那個鉀離子通道,其 機制有待進一步探討。
關鍵詞:第二型糖尿病、巨噬細胞、發炎反應、陰電性低密度脂蛋白、游離脂肪 酸、介白激素 1、NF-κB、NLRP3 發炎小體、棕櫚酸
Abstract
Recently, prevalence of obesity is increasing worldwide because of changing of diet preference. And obesity is a major risk factor for metabolic diseases, which usually progresses to type 2 diabetes mellitus and cardiovascular diseases; such as atherosclerosis, myocardial infarction. Inflammation is associated with these diseases, and macrophages are the predominant contributor of inflammation. Moreover, electronegative LDL (LDL(-)), free fatty acids and interleukin-1IL-1) are increased significantly in the
blood of these patients.
Production of IL-1 requires two signals. Firstly, the priming signal is to activate NF-κB signaling. NF-κB as a transcription factor that promotes pro-IL-1and NLRP3
expression. Secondly, the activating signal is to activate NLRP3 inflammasome and caspase-1. Then activated caspase-1 proteolytically cleaves pro-IL-1into mature IL- 1Our previous study showed that LDL(-) could induce NF-κB activation in
macrophage, and weakly promote caspase-1 activation. Evidence has shown that palmitic acid could promote NLRP3 inflammasome/caspase-1 activation. Thus, in this study we investigated whether there is an interaction between LDL(-) and palmitic acid in inducing IL-1production in macrophages.
LDL(-) was isolated from the plasma of the rabbits fed with high-fat/cholesterol diet.
THP-1 macrophages were treated with LDL(-) and BSA bound saturated or unsaturated free fatty acids, then the levels of IL-1in culture medium were analyzed by ELISA. The
results show that palmitate, a saturated fatty acid, alone is unable to induce NF-κB activation and IL-1production in macrophages. However, palmitate enhances LDL(-)- induced IL-1production. Palmitate did not further activate NF-κB or increase the levels of pro-IL-1 mRNA and protein when treated with LDL(-). Our data also show that
palmitate did not further increase LDL(-)-induced increase of LOX-1, a scavenger receptor for LDL(-). These results suggest that palmitate enhances LDL(-)-induced IL- 1production was not through activation of signal one (NF-κB signaling) or increase
levels of LOX-1.
Cells treated with LDL(-) under a potassium free medium, and the level of IL-1in
the medium is close to that of LDL(-) and palmitate co-treated cells. Treated potassium channel blockers significantly lower LDL(-) and palmitate-induced levels of IL-1. The results suggest that palmitate enhances LDL(-)-induced IL-1βproduction is likely through increase potassium efflux.
In conclusion, our study show that palmitate enhances LDL(-)-induced IL- 1production, the results is physiological relevant since the concentrations of LDL(-) and palmitate used in this study are under physiological ranges in plasma of patients with AMI and diabetes. However, we do not know which potassium channel is activated by
palmitate. The mechanism underlie deserved further investigation.
Keywords: type 2 diabetes, macrophages, inflammation, electronegative LDL, free fatty acids, Interleukin-1NF-κB, NLRP3 inflammation, palmitate
目錄
口試委員審定書 ... i
摘要 ... ii
Abstract ... iv
第一章 緒論 ... 1
第一節 文獻回顧 ... 2
(一) IL-1 和發炎反應的關係 ... 2
(二) IL-1 的產生 ... 3
(三) Oxidized LDL/陰電性低密度脂蛋白 (LDL(-)) 和巨噬細胞的關係 ... 5
(四) LDL(-) 的特性... 7
(五) 游離脂肪酸在發炎反應扮演的角色 ... 8
(六) 循環中的游離脂肪酸 ... 9
(七) LOX-1 對於 LDL(-) 誘導發炎反應的重要性 ... 10
第二節 研究動機與實驗目的 ... 11
(一) 研究動機 ... 11
(二) 實驗目的 ... 12
第二章 材料與方法 ... 13
第一節 實驗材料 ... 14
第二節 細胞培養 ... 17
第三節 分離脂蛋白 ... 17
第四節 游離脂肪酸牛血清蛋白複合物製備 ... 18
第五節 酵素結合免疫吸附法 (Enzyme-Linked ImmunoSorbent Assay, ELISA) .... 18
第六節 細胞存活率分析 ... 19
第七節 細胞 mRNA 表現分析 ... 19
(一) 細胞 RNA 抽取 ... 20
(二) 合成 First strand cDNA ... 20
(三) 即時定量聚合酶連鎖反應 (Real-time quantitative PCR) ... 20
(四) 引子資訊 ... 21
第八節 西方點墨分析法 ... 21
(一) 細胞蛋白抽取 ... 21
(二) 樣品前處理 ... 22
(三) 膠體配方 ... 22
(四) SDS-PAGE ... 22
(五) 半濕式蛋白質轉印 ... 23
(六) 抗體結合免疫分析 ... 23
(七) Enhanced Chemiluminescence (ECL) 系統偵測冷光訊號 ... 23
第九節 Knockdown 細胞中標靶基因透過 RNA 干擾 ... 24
(一) 慢病毒 Clone 資訊 ... 24
(二) 慢病毒感染 ... 25
第十節 Potassium-free 溶液配製 ... 25
(一) 溶液配方 ... 25
第十一節 統計分析 ... 26
第三章 結果 ... 27
第一節 人類 LDL(-) 會誘導巨噬細胞產生 IL-1,而 native LDL 不會... 28
第二節 兔子 LDL(-) 單獨在巨噬細胞引起小幅度 IL-1 產生 ... 28
第三節 Palmitic acid 在巨噬細胞對 IL-1 的產生沒有影響 ... 29
第四節 兔子 LDL(-) 和 palmitic acid、stearic acid 在巨噬細胞對 IL-1 的產生 之影響有協同作用,其他測試的脂肪酸則無影響 ... 29
第五節 LDL(-) 在巨噬細胞引起 IL-1 產生經由 LOX-1 ... 30
第六節 Palmitic acid 不會促進 LDL(-) 進一步引起 NF-B 活化 ... 31
第七節 Palmitic acid 不會促進 NF-B 下游基因 IL-1、TNF- 和 IL-6 的 mRNA 和蛋白質表現 ... 32
第八節 Palmitic acid 促進 LDL(-) 誘導 IL-1 產生不是經由 phagocytosis 和 lysosomal degradation ... 33
第九節 Palmitic acid 促進 LDL(-) 誘導 IL-1 產生可能經由 potassium efflux 34 第四章 討論 ... 36
第一節 Palmitic acid 促進 LDL(-) 誘導之 IL-1 產生增加 ... 37
第二節 LDL(-) 透過 LOX-1 可能不是經由胞吞作用引起下游訊號 ... 38
第三節 PA 不是藉由 LOX-1 促進 LDL(-) 在巨噬細胞誘導 IL-1 產生增加 39 第四節 游離脂肪酸和 IL-1 產生的關係 ... 40
第五節 PA 在巨噬細胞不活化 NF-B 路徑 ... 41
第六節 PA 在巨噬細胞透過 potassium efflux 促進 NLRP3 inflammasome /caspase-1 活化……….. ... 42
第七節 總結 ... 43
第五章 圖表 ... 45
第六章 參考文獻 ... 62
圖表目錄
Figure 1. Effects of human nLDL, LDL(-) and PA on the production of IL-1, TNF-, and IL-6 in THP-1 macrophages. ... 46 Figure 2. Effects of rabbit LDL(-) on the production of IL-1 in THP-1 macrophages. 48 Figure 3. Effects of PA on the production of IL-1 in THP-1 macrophages. ... 49 Figure 4. Effects of rabbit LDL(-) plus PA and rabbit LDL(-) plus various FFAs on the production of IL-1 in THP-1 macrophages. ... 51 Figure 5. LDL(-) and PA induce IL-1 production through a LOX-1 but not CD36, dependent pathway. ... 54 Figure 6. Effects of rabbit LDL(-) and PA on NF-B activation in THP-1 macrophages.
... 56 Figure 7. Effects of rabbit LDL(-) and PA on expressions of NF-B downstream genes, IL-1, TNF- and IL-6, and their protein levels in THP-1 macrophages. ... 58 Figure 8. PA enhances LDL(-)-induced IL-1 production is not through phagocytosis and lysosomal degradation. ... 59 Figure 9. PA enhances LDL(-)-induced IL-1 production is possibly through potassium efflux. ... 60
第一章 緒論
第一節 文獻回顧
(一) IL-1和發炎反應的關係
促發炎細胞因子 Interleukin 1IL-1參與第二型糖尿病(type 2 diabetes mellitus, T2DM)、痛風性關節炎、粥狀動脈硬化、心血管疾病(cardiovascular diseases, CVD)和肥胖相關併發症等的發展。文獻指出,以IL-1抗體治療,能有效改善第
二型糖尿病;也能減少心血管疾病的發生;且能改善胰島素的分泌。除了阻
止IL-1的作用,也抑制了IL-1在胰島細胞所促進的發炎反應,像是 IL-8 和 IL-6 等細胞因子釋出 [3]。研究發現,ST 時段上升心肌梗塞STEMI病人血漿
(plasma)中,IL-1濃度比正常人高 [4]。臨床證據顯示患有 T2DM 之病人很容易
走向 CVD,如心肌梗塞 (myocardial infarction, MI)、粥狀動脈硬化 (atherosclerosis) 等。主要成因是高血糖、胰島素抗性 (insulin resistance),造成血脂、血糖代謝異常。
臨床發現在急性心肌梗塞 (acute myocardial infarction, AMI) 病人與 T2DM 患者 血液中游離脂肪酸 (free fatty acids, FFAs)都偏高 [5-7]。因此,循環中游離脂肪酸 對於 insulin resistance 和造成 AMI 發生的機制可能隱含重要的問題。胰島素能抑 制脂質分解,促進脂質儲存在脂肪細胞;抑制肝臟糖質新生、刺激葡萄糖透過 GLUT-4 進入到脂肪及肌肉細胞被儲存或利用。而外源性的 FFAs 增加,像飲食中 攝取的油脂,若造成 FFAs 過多,則會減少食物的攝取和抑制肝臟糖質新生等 [6];
使循環中的葡萄糖減少,進而減弱細胞對胰島素的敏感性,導致 insulin resistance,
且發現常引起心血管功能障礙 [6]。Insulin resistance 和飲食誘導之發炎反應有關,
藉由細胞發炎反應干擾 IRS 的磷酸化 (Insulin signaling pathway) [8]。AMI、T2DM 病人等也被發現血液中的促發炎細胞因子,如 IL-1、TNF-等,較正常人高[6, 8-10]。很多人在研究,脂肪酸除了誘導 insulin resistance 發生,其在發炎反應中扮 演什麼重要角色,而學者經常透餵食高膽固醇脂飲食 (high-fat diet, HFD) 誘導 insulin resistance 或糖尿病發生之動物模式進行研究。
也有許多文獻提到,inflammasome 活化促進 IL-1的產生,在肥胖、insulin
resistance 和 T2DM 的發生扮演重要的角色 [11, 12]。 IL-1常在這些病人血液中 被發現,除了作為一個發炎反應的指標,會刺激下游產生更廣的發炎進展。IL-1
只在特定的細胞中產生,像是血液中單核球、組織中的巨噬細胞跟樹突細胞。IL- 1產生會刺激許多細胞因子和趨化因子的產生和分泌、誘導內皮細胞表現細胞黏 附分子,促單核球分化移動到血管內膜,分化成巨噬細胞等 [13]。而 oxidized LDL 以往被認為是造成 atherosclerosis 的主要因子,也有人在探討其和 T2DM、發炎反 應和 IL-1產生之關係 [14]。非酒精脂肪肝病 (NAFLD) 嚴重會發展到肝硬化、
肝癌,也常伴隨著 T2DM 和 MI 的發生 [15-17],且 oxidized LDL 也被認為是造 成非酒精性脂肪肝炎 (NASH) 的危險因子之一 [18]。以上這些疾病都可能和 oxidized LDL、FFAs 跟 IL-1 有密切的關係。
在臨床上,當人體血液循環 pH 值下降(正常生理 pH = 7.4),生理機能會弱
化。實驗也證明,胞外酸化 (acidification)增加細胞外的氫離子濃度,和發炎反應有 關,可能伴隨著使鉀離子流出細胞,而活化 inflammasome,造成 IL-1的產 生。另外,IL-1被釋放到細胞外,是一個重要的過程,進而影響下游許多細胞因 子產生和免疫反應。IL-1如何被分泌到細胞外,有幾個機制被提出,IL-1由
secretory lysosomes 攜帶、形成exosomes,被胞吐至細胞外(exocytosis)、透過 specific transporters 運送,或從和細胞膜融合 vesicles 釋出 [20]。而其中細胞膜上 的受體等可能影響 IL-1的釋出,如P2X7 離子通道在其中扮演重要角色,P2X7 之 拮抗劑在許多發炎反應和自體免疫相關疾病有治療的效用 [21]。
(二) IL-1的產生
典型的 IL-1產生需要兩條訊號傳遞路徑,第一條為啟動訊號 (priming signal),
核因子活化 NF-κB 作為轉錄因子,調控 IL-1 的基因表現。當上游 I-Bα 磷酸 化,和NF-κB 次單元 p50 和 p65 分開後,p65 磷酸化,活化的 NF-κB 進到細胞 核中,增強IL-1的基因轉錄,再轉譯成pro-IL-1;亦促進 inactive NLRP3 的表 現增加。Pro-IL-1和NLRP3 主要在免疫相關細胞表現,像單核球細胞 (monocyte)、
樹突細胞 (dendritic cell) 和巨噬細胞 (macrophage) 等。但平時在這些細胞保持低 度表現,在有損傷或病原 (Damage/ Pathogen -associated molecular patterns) 刺激,
造成 priming signal,進而促進他們的轉錄 [22]。成熟態 IL-1的產生需要進行轉 譯後修飾,藉由第二條訊號 (activating signal) 刺激、誘導 NLRP3 inflammasome 的 組裝和 caspase-1 的活化,切割 pro-IL-1使成為成熟態 IL-1。
發炎小體 (inflammasome) 是一個很大的複合物,能促細胞內的 Caspase-1 活 化進而促進成熟態的 IL-1和IL-18 產生分泌至細胞外。其中 NLRP3 為最常見的 inflammasome 。NLRP3 在許多肥胖 (obesity) 相關疾病,特別是 T2DM 中扮演 重要角色。已有文獻指出,肥胖 (HFD) 誘導小鼠產生胰島素抗性、粥狀動脈硬化,
而 NLRP3-/- 缺失的小鼠受到保護 [23, 24]。而典型 IL-1 的產生是透過 Caspase- 1/ASC/NLRP3 路徑,NLRP3 (nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin domain containing 3)、ASC (apoptosis-associated speck-like protein containing a caspase recruitment domain) 和 pro-Caspase-1 (cysteine protease) 經過彼 此連接且摺疊,組成大型複合物,稱為 NLRP3 inflammasome。被活化的 NLRP3 inflammasome 其中 pro-Caspase-1 進行自我切割 (autocleavage), 產生活化態 Caspase- 1 ( p10/p20 tetramer),再將 pro-IL-1進行切割,形成成熟態 IL-1,進而 分泌到細胞外 [25]。故常以分泌至胞外的 IL-1和 Caspase-1 增加,表示 NLRP3 inflammasome 的活化。不過在肥胖相關疾病和 NLRP3 所介導的 IL-1 產生之間 的分子連結機制目前尚不完全清楚。
活化 NLRP3 inflammasome 上游有幾個常見的機制,包括粒線體 ROS (reactive oxygen species) 的產生、溶酶體功能失調 (lysosome dysfunction) [26]、鉀 離子排出到細胞外 (potassium efflux) 和胞外高濃度 ATP (ATP release) 等 [27]。而 aluminum hydroxide (alum)、oxidized LDL、膽固醇結晶體 (cholesterol crystals)、單 鈉尿酸鹽晶體 (monosodium urate, MSU crystals) 和游離脂肪酸 (free fatty acids) 等 被認為是 NLRP3 inflammasome 的活化子 (activator) [28]。有人甚至提出,alum 和
MSU 被細胞吞噬,造成 lysosome dysfunction 和 ATP release,形成的濃度梯度,
使ATP 和 P2X7 (purinergic receptor) 離子通道結合,伴隨 potassium efflux 發生,
觸發 NLRP3 inflammasome 活化 [29]。近年來,越來越多人指出,飽和脂肪酸 (saturated fatty acid) 能誘導 NLRP3 inflammasome 活化 [26, 30]。
(三) Oxidized LDL/陰電性低密度脂蛋白 (LDL(-)) 和巨噬細胞的關係
粥狀動脈硬化 (atherosclerosis) 是心血管疾病發生主要原因。現今知道成因是 oxidized LDL 刺激動脈內皮細胞產生趨化因子,如 MCP-1 和 M-CSF 等,這些因 子跟細胞黏附分子,促使單核球進入到血管內膜空腔 (intima),分化成巨噬細胞,
並且表現清道夫受體 (scavenger receptors) 將 intima 中的 oxidized LDL 攝入。過 多的三酸甘油酯跟膽固醇堆積在巨噬細胞,形成泡沫細胞 (form cells)。加上平滑 肌細胞聚集於血管內膜壁,逐漸形成纖維性斑塊,使血管腔變窄,阻礙血流通過;
嚴重時斑塊破裂,造成血栓。家族性高膽固醇血症(Familial hypercholesterolemia)患 者,其血液中膽固醇 (LDL-cholesterol) 濃度很高,且發生心血管疾病的風險高。
而其 LDL receptor (LDLR)基因發生缺陷,LDL 卻會被巨噬細胞攝入,在 1970 年 代,許多學者提出 oxidized LDL 和 atherosclerosis 之間的關係,還有對巨噬細胞 造成的影響,並在一些 in vitro 實驗驗證 [31-33]。首先有學者將從人類血漿分離 得到的 LDL 和兔子主動脈 (aorta) 的內皮細胞 (endothelial cell) 一起培養一段時 間後,會對內皮細胞造成細胞毒性 [32, 34]。並且將這些 LDL 分離出來,發現和 原本的(native) LDL 相比,密度增加,由於其中的膽固醇酯和磷脂質減少了,蛋白 質等組成改變;且發現在 apolipoprotein B (ApoB)上 lysine 殘基的εamino groups 進行了修飾而提高負電荷。將培養後含有經過自然修飾之 LDL 的培養基,轉移至
巨噬細胞進行培養,這些修飾的LDL 會被巨噬細胞攝入並促進形成泡沫細胞。而
經化學修飾的 acetylated LDL (acetyl-LDL) 亦會被巨噬細胞攝入,並且形成泡沫細 胞 [33]。而這種自然發生的 LDL 修飾,被發現是一種氧化性修飾 (oxidative modification),故稱這種 LDL 為 oxidized LDL (oxLDL) [35, 36]。之後常有人以二
價金屬離子氧化 LDL 進行相關實驗,如 Cu2+-oxLDL。不過也證實了巨噬細胞上 的一些受體對於acetyl-LDL 和 oxLDL 的專一性不一樣 [37-39]。推測有數個因子 會造成LDL 氧化,而其中一個重要的氧化因子為骨髓過氧化酶 (Myeloperoxidase, MPO),被活化的嗜中性球和單核球中會大量表現 MPO,其會使 LDL 上的脂質和 蛋白質被氧化,進而產生活性氧化物質 (Reactive oxygen species, ROS),促進更多 的 LDL 被氧化[40]。
2003 年 Yang 等人,以陰離子交換層析法從血漿 LDL 分離到多種和 native LDL 相比,帶負電的 LDL,(依帶負電性低至高分離出 L1 (native LDL)到 L5)
[41],我們稱其為陰電性低密度脂蛋白 (Electronegative LDL, LDL(-))。過去認為,
oxLDL 會被巨噬細胞攝入,進而形成 form cell,並且認為是在 intima 被氧化。而 LDL(-) 是氧化程度較輕微的 oxLDL (依 malondialdehyde, MDA 過氧化產物和 thiobarbituric acid, TBA 結合的產物 (TBARS) 生成量比較),且可由循環中的LDL 分離到。因此我們認為,LDL 可能在循環中就已經發生氧化作用,而非一定在 intima 被氧化。LDL(-) 為現今一個新穎的心血管疾病指標,在 STEMI、抽菸、高 膽固醇症和糖尿病患者的血液中能被分離到 [42],其中以 STEMI 患者含的比例 最高,約佔全部 LDL 的 3~10%。而糖尿病患者,血液裡以氧化過程中出現的一 種氧化型態,為small dense LDL (sdLDL),可能是 LDL 經 glycation 修飾產生的 終產物,引起發炎反應 [43]。循環中的 LDL (-) 能活化發炎和免疫反應,進而造成 atherosclerosis 的發展,但 LDL (-) 在發炎反應中扮演的角色,目前尚未有很詳細 的了解。不過,我們認為 LDL (-) 可能在其中扮演一個訊息分子,透過訊息傳傳的 引起發炎反應。先前實驗室研究發現,高脂飲食誘導倉鼠肝門靜脈周圍巨噬細胞 (Kupffer cell) 堆積的位置,同時也有 LDL(-) 聚集 [44],認為 LDL(-) 可能透過被 巨噬細胞攝入而引起發炎反應。學長在 in vitro 實驗中,證實 LDL(-) 在巨噬細胞 中誘導 IL-1、TNF-和 IL-6 促發炎細胞因子產生,並且可能是透過活化 NF-B 路徑[4, 44]。而細胞因子 G-CSF 和 GM-CSF 的表現和 AMI 嚴重程度呈正相關,
我們也發現 LDL(-) 會誘導 G-CSF 和 GM-CSF 的產生 [45]。在這先研究也比較 了LDL(-) 和 oxLDL (commercial Cu2+-oxLDL) 對在巨噬細胞引起促發炎因子表現 的影響,發現兩者差異很大。在生理濃度下 LDL(-) 引起明顯的發炎反應,而 oxLDL 與 native LDL 相似並沒有明顯的作用 [4]。表示 LDL(-) 在發炎反應中是 重要因子之一,且比起和過去大家認知的 oxLDL 對巨噬細胞的影響還大,LDL(-) 似乎扮演不一樣的角色。LDL(-) 也確實存在人類血液循環中,然而其中引起發炎 反應的機制是我們目前正在探討的方向。
(四) LDL(-) 的特性
native LDL/LDL(-) 和 oxLDL 抗體進行免疫競爭的方式證明,倉鼠的肝門靜 脈周圍有 LDL(-) 的堆積,而非 native LDL,並證明 LDL(-) 能夠從倉鼠血液中分 離出來[44]。有許多文獻指出,minimally oxLDL (MM-LDL) 常用來描述開始進入 氧化過程輕微氧化的 LDL,其生物活性就已經和未經修飾的 LDL 不同,能在內 皮細胞或巨噬細胞誘導促發炎因子和趨化因子產生 [46]。而其生理修飾通過多種 方式進行,包括酵素反應和非酵素反應,酶反應和非酶反應,LDL 最外層主要是 由磷脂質所組成,可能由此先被氧化[46]。自由基、ROS 的攻擊或 lipoxygenase 的 催 化 造 成 脂 質 過 氧 化 (peroxidation) [46] ; 而 LDL 脂 質 上 的 修 飾 常 見 有 , phospholipases A2 (PLA2) 水解 phosphatidylcholine,產生 lysophosphatidylcholine,
而 游 離 脂 肪 酸 的 含 量 增 加[46] 。 Sphingomyelinase (SMase) 將 LDL 上 的 sphingomyelin 水解成 ceramide [46],而過程中,構型發生了改變,增加了 SMase 對 ApoB 的活性。 膽固醇酯 (cholesteryl ester) 則被 水解,不 過由於其不像 phospholipid 在血漿發生水解作用,故其產生的過氧化物,為 atherosclerosis lesions 中找到最主要的脂質氧化產物[47]。而 LDL(-) 被認為是 mildly 或是 moderately oxidized LDL,氧化程度介於 MM-LDL 和 extensively oxLDL [46]。亦有學者發現,
LDL(-) 和 native LDL 相比,其中 ApoB 的二級結構減少,整個構型嚴重改變 [38],
且因為ApoB 發生錯誤摺疊 (misfolding),進而造成 LDL 聚合 (aggregation),而
LDL 發生 aggregation 和 atherosclerosis 發生的成因有密切的關係 [48, 49]。此外 , ApoB 上的 tyrosine 殘基也可能透過 MPO 被修飾 [46]。而 LDL 除了可能因為 ApoB 上的 lysine 殘基被修飾;發生 glycation 和 desialylation 亦會增加負電性 [46]。LDL(-) 和 native LDL 相比,主要的蛋白質組成為 ApoB (pI = 6.62),而 LDL(-) 則增加了Apo(a)、 ApoA-I、ApoC-II、ApoC-III 和 ApoE (pI < 6.62) 等 [50],也 有學者認為因此增加了 LDL 的負電性。LDL(-)的 vitamin E 和多不飽和脂肪酸含 量減少;總膽固醇 (total cholesterol)、三酸甘油酯 (triglyceride)、游離脂肪酸 (non- esterified fatty acids)、sialic acid 和 TBARS 含量增加;和 LDLR 之親和力下降 [47]。
(五) 游離脂肪酸在發炎反應扮演的角色
在 T2DM 病人血液中 FFAs 的濃度增加;在以 high-fat diet (HFD) 誘導糖尿 病或胰島素抗性 (insulin resistance)之動物模式中也有 FFAs 提高的現象。FFAs 可 能是造成 insulin resistance 的主要因素之一。Insulin resistance 在臨床上很重要,
和 T2DM、高血糖和血脂異常等密切相關,為心血管疾病 (CVD) 的危險因子 [51]。
心肌梗塞 (MI) 病人,FFAs 濃度也提升,故血液中 FFAs 增加和 CVD 有極大的 關聯性 [5]。在臨床上,FFAs 被視為潛在的生物標記 (biomarker) [5]。而 IL-1為 CVD 相關發炎反應的重要介質,也發現 AMI 和 T2DM 病人等,血液中 IL-1
濃度偏高 [6, 8-10]。因此有人探討 insulin resistance 和 FFAs 跟發炎反應之間的關 係。FFAs 在脂肪組織 (adipose tissue) 透過 toll-like receptor 4 (TLR4) 引起發炎反 應,且促進巨噬細胞產生細胞因子 (cytokines);而 TLR4 缺失的骨骼肌細胞能避 免脂質所誘導的 insulin resistance [52]。也有報導指出,肥胖或 T2DM 因為 insulin resistance,使血液中 FFAs 濃度提高,促進 adipose tissue 中巨噬細胞浸潤 [53]。而 FFAs 作為訊號經由巨噬細胞上表現的 TLRs 活化 MAPK (mitogen- activated protein kinase) pathway [53, 54]、促發炎 signaling pathways [51, 55, 56]。也 就是說,FFAs 可能對於 IL-1的產生可能是經由 TLRs,活化 priming signal,但 都尚未有直接的證明。
多篇報導指出,FFAs 中的飽和脂肪酸 (saturated fatty acids, SFAs),特別指血 液中含量最多的兩個 FFAs,棕櫚酸 (palmitic acid) 和硬脂酸 (stearic acid) 在巨噬 細胞能活化NLRP3 inflammasome (activating signal),進而促進 IL-1 的產生 [26, 30]。SFAs 活化 NLRP3 inflammasome,且其中有AMPK-ROS signaling 參與[56]。
其他報導甚至認為,細胞內SFAs 增加、lysosomal dysfunction 和 SFAs 衍生的晶 體能夠在巨噬細胞活化 NLRP3 inflammasome [26]。在我們先前的研究發現,LDL(-) 在巨噬細胞可能透過 ROS 誘導 IL-1產生,且非經吞噬作用(phagocytosis) 和 lysosomal dysfunction [4]。而我們好奇,若是 LDL(-) 能活化 IL-1產生之priming signal,那 SFAs 是否扮演活化NLRP3 inflammasome 的角色,促進 IL-1大量產 生,並且是否透過上述AMPK-ROS signaling、potassium efflux、lysosomal dysfunction 或 phagocytosis 等機制去活化。
(六) 循環中的游離脂肪酸
在飲食中,豬油含有約24% 棕櫚酸palmitic acid)、約硬脂酸stearic acid)、 約油酸oleic acid) 和約亞油酸(linoleic acid);椰子油含有約月桂 酸 (lauric acid)。在台灣人的飲食中,常用的大豆沙拉油,含有 52~65% linoleic acid。
許多文獻也探討攝取的油脂,可能對發炎反應產生的影響 [57-60]。在循環中FFAs 正常的濃度平均為約 400~500 M,中長鏈的FFAs 通常由白蛋白 (albumin) 攜帶。
而palmitic acid (C16:0) 和 stearic acid (C18:0) 這兩種飽和脂肪酸 (SFAs) 含量最 高,palmitic acid 佔其中的二到三成,約為 100~150 M,且為脂肪酸合成過程中 產生的第一產物,是其它長鏈脂肪酸和不飽和脂肪酸的前驅物。lauric acid (C12:0) 和myristic acid (C14:0) 在循環中含量較少,為短中鏈脂肪酸。而 Oleic acid (C18:1 n-9) 和 linoleic acid (C18:2 n-6) 為循環中較多的不飽和脂肪酸 (unsaturated fatty acids, UFAs);-亞麻酸 (-linolenic acid) (C18:3 n-3)、花生四烯酸 (arachidonic acid) (C20:4 n-6) 和 DHA (docosahexaenoic acid) (C22:6 n-3) 等含量較少 [61]。 SFAs 和發炎反應之間的關係,目前還不太清楚。不過,許多文獻指出 SFAs 能誘導
NLRP3 inflammasome 活化,甚至造成其他和肥胖、代謝異常相關的發炎過程, 不過看起來似乎需要 priming signal 才能造成 IL-1產生。並且目前也不知道是否 全部的SFAs 在巨噬細胞上皆有相同的影響,甚至不清楚何種濃度或是碳鏈長短和 構造,是否會影響調控機制。UFAs,像 oleic acid 和 linoleic acid,在巨噬細胞抑 制 NLRP3 inflammasome 活化,能抑制 SFAs 引起的 IL-1 產生 [26, 30]。-3 脂 肪酸也被指出能透過抑制 NLRP3 inflammasome 活化避免發炎反應和代謝失調 [62]。但其他 UFAs 對於 NLRP3 inflammasome 活化和發炎反應所扮演角色,目 前並不清楚。
(七) LOX-1 對於 LDL(-) 誘導發炎反應的重要性
我們知道,native LDL 會被細胞膜上的 LDL 受體 (LDL receptor, LDLR)辨認,
進而將膽固醇回收至肝臟或運送到肝外組織再利用。LDLR 辨認的是 LDL 上的 ApoB-100,而 LDL(-) 和 LDLR 的親和力低 [50, 63]。 Acetyl-LDLR 在 1980 年 代被發現且被選殖出來,稱為 Scavenger receptor A1 (SR-AⅠ),不過 oxLDL 並不會 和 acetyl-LDL 進行競爭,故認為在巨噬細胞上有不只一種 modified-LDL 之清道 夫受體 (scavenger receptor)。在 1993 年的研究,發現 CD36 為 oxLDL 的 scavenger receptor,在 oxLDL 能被巨噬細胞上的 CD36 結合降解 [39]。巨噬細胞上最主要 的 scavenger receptor 有 SR-AⅠ/Ⅱ 和 CD36;而有學者認為,不同的 scavenger receptor 辨認 oxLDL 上不同的組成。像 SR-A1 辨認 ApoB 的修飾;CD36 辨認 被氧化的 phospholipid;Toll like receptor 4 (TLR4) 則辨認被氧化的 cholesteryl ester,
這些 scavenger receptor 也表現在其他細胞,包括平滑肌細胞和血小板等 [46]。
而 Lectin-like oxidized LDL receptor-1 (LOX-1) 過去被認為是內皮細胞上 oxLDL 的主要受體,最早從牛的主動脈內皮細胞之 cDNA library 篩選出。LOX-1 對於 oxLDL 的結合和降解能力強,而非 native LDL 和 acetyl-LDL [46]。一開始 發現 LOX-1 和 oxLDL 作用會造成內皮細胞失去功能或是細胞凋亡 (apoptosis),
和 atherosclerosis 的發生有相當的關係 [42],且在人類早期 atherosclerosis 發生
時大量表現 LOX-1 [64]。不過 LOX-1 的表現在多數病變中發現,也在堆積於 intima 的巨噬細胞和平滑肌細胞有高度表現 [64, 65]。有學者甚至指出這些不同受 體和 oxLDL 之間的關係,還會依據 oxLDL 不同的氧化程度,如 CD36 和 extensively oxidized LDL;LOX-1 和 mildly oxLDL [46];LOX-1 和 LDL(-) [50] 之 間有較強的交互作用。後來出現許多報導指出,LDL(-) 和 LOX-1 引起發炎反應 之間的重要關係 [42, 66]。 LDL(-) 經由LOX-1 路徑,活化 protein kinase C (PKC)、
p38 MAPK 和 NF-B;甚至自己扮演黏附因子(adhesion molecule) 的角色,吸引單 核球等,引起發炎反應 [42, 65]。看起來 LOX-1 和 LDL(-) 之間的關係,和其他 scavenger receptors 接受 oxLDL 為配體和其結合並且進行內吞作用 (endocytosis) 不完全相同。在實驗室先前的研究發現, LDL(-)會在巨噬細胞誘導 LOX-1 的表 現,並且指出 LDL(-) 是經由 LOX-1 誘導促發炎細胞因子 IL-1 產生 [4]。並且 我們認為 LOX-1 的調控是在訊息路徑的最上游,LOX-1 和 LDL(-) 之間的交互 作用可能是造成發炎反應的關鍵第一步。另一方面,LDL(-) 透過 LOX-1 調節肌 纖維膜離子通道,改變所藉導的心臟復極化,則是減少 potassium 往細胞外流動 [66]。也有文獻指出,palmitic acid 造成 LOX-1 在巨噬細胞的表現增加 [64]。因 此,我們想探討 SFAs 是否對於 LDL(-) 誘導 IL-1產生有增加效應之外,並且 LDL(-) 和 SFAs 是否透過 LOX-1 產生作用。
第二節 研究動機與實驗目的 (一) 研究動機
先前實驗室研究指出,LDL(-) 在巨噬細胞會引起 IL-1的產生,且LDL(-) 能 增加 Caspase-1 活性[4]。但是據我們判斷,Caspase-1 活化程度並不高,LDL(-) 被 認為主要是啟動 priming signal,活化 NF-B 路經,促進轉錄、轉譯,表現大量 pro-IL-1和 NLRP3。而 NLRP3 inflammasome 與 Caspase-1 的活化主要應該是透 過其他因子。心肌梗塞或是糖尿病病人共通的是,血液中除了發現 LDL(-) 和 IL- 1的存在, FFAs 的濃度也較正常人的高,這些都和心血管疾病 (CAD) 的發生
有相當的關聯性 [5]。2011 年,Wen 等人提出,在 lipopolysaccharide (LPS)-priming 的情況下, FFAs 在巨噬細胞誘導 NLRP3 inflammasome 活化,進而促進 IL-1
的產生,並且探討了多個NLRP3 inflammasome 活化相關的機制 [56]。因此我們 想要知道,是否LDL(-) 在人類巨噬細胞扮演 priming 的角色,而透過 FFAs 的刺 激,造成 NLRP3 inflammasome 活化,以提高 IL-1的產生,達到加乘作用。
(二) 實驗目的
1. 第一步先探討,游離脂肪酸是否會〝促進〞LDL(-) 在巨噬細胞所誘導之 IL- 1 的產生。
2. 探討不同游離脂肪酸,包括飽和脂肪酸和不飽和脂肪酸,對於 LDL(-) 誘導 IL-1 的產生,是否有不同的影響。
3. 最後,探討游離脂肪酸是否在巨噬細胞中誘導NLRP3 inflammasome 活化,並 且探討參與在其中的路徑。
第二章 材料與方法
第一節 實驗材料
1. THP-1 (Human monocytic leukemia cell line) 購 自 American Type Culture Collection (Manassas, VA, USA)。
2. Hyclone RPMI 1640 (SH30017.02) 及 fetal bovine serum (FBS) 購自 Thermo Scientific (Logan, UT, USA)。
3. Laboratory rabbit diet 購自 LabDiet (St. Louis, MO, USA)。
4. Cholesterol powder 購自 Wako (Ginza, Tokyo, Japan)。
5. Dialysis membrane (12-14 KDa) 購 自 Spectrum Laboratories Inc. (Rancho Dominguez, CA, USA)。
6. UNO Q6 及 Q12 Column 購自 Bio-Rad (Hercules, CA, USA)。
7. Syringe filter (0.22 μm) 購自 Sartorius Stedim Biotech (Göttingen, Germany)。
8. Centrifuge tubes 購自 Beckman (Brea, California, USA)。
9. Centrifuge filter Units (10K) 購自 Millipore (Taipei, Taiwan)。
10. Quantikine Human IL-1、TNF- 及 IL-6 ELISA kits 購 自 R&D Systems (Minneapolis, MN, USA)。
11. Lauric acid、Myristic acid、Stearic acid、Docosahexaenoic acid、Dimethyl sulfoxide (DMSO)、bromophenol blue、NaCl、CaCl2、NaOH、KOH、D-glucose、phorbol 12-myristate 13-acatate (PMA)、polymyxin B (PMB)、MTT、bovine serum albumin (BSA)、ammonium persulfate (APS) 及 β-mercaptoethanol 購自 Sigma-Aldrich (St. Louis,MO, USA)。
12. L-glutamine 、 penicillin/streptomycin 、 TRizol 及 HEPES 購 自 Gibco/Life Technologies (Rockville, MD, USA)。
13. Isopropanol、methanol、ethanol、chloroform、glycerol、Na2HPO4、KH2PO4 及 Tris- base 購自 J.T.Baker (Phillipsburg, NJ, U.S.A.)。
14. dNTP mixture 、 Protein and DNA ladder 及 Oligo dT18 primer 購自 Bioman Scientific (Taipei, Taiwan)。
15. M-MLV-RT 5X buffer 及 M-MLV-reverse transcriptase 購自 Promega (Madison, WI, USA)。
16. Primer 由源資或每得公司合成。
17. 2x SYBR mix 購自 Bioline (Trento, Italy)。
18. KCl、MgCl2、Isoamyl alcohol (IAA)、sodium dodecyl sulfate (SDS)、N, N, N’, N’- tetramethyl ethylenediamine (TEMED)、ethylenediaminetetraacetic acid (EDTA) 及 glycine 購自 Merck (Whitehouse Station, NJ, USA)。
19. Phenyl methane sulfonyl fluoride (PMSF) 購 自 Boehringer Mannheim GmbH (Ingelheim, Germany)。
20. Protease inhibitor cocktail tablets 購自 Roche (Basel, Schweiz)。
21. DC Protein Assay Kit 及 30% acrylamide/bis solution 購自 Bio-Rad (Hercules, CA, USA)。
22. PVDF membrane 購自 Millipore (Billerica, MA, USA)。
23. Monoclonal antibodies against Phospho-p65 (ser536) 、 phospho-ERK1/2 及 cleaved- IL-1β (Asp116) 購自 Cell Signaling Technology (Beverly, MA, USA)。
24. Monoclonal antibody against phospho-I-B、TEA、4-AP 及 A 438079 購自
bcam (Cambridge, UK)。
25. Polyclonal antibodies against p65 、 ERK1/2 及 I-B購 自 Santa Cruz Biotechnology (Santa Cruz, CA, USA)。
26. Monoclonal antibody against LOX-1 購自 BioLegend (San Diego, CA, USA)。
27. Goat anti-mouse IgG HRP、Goat anti-rabbit IgG HRP 及 Monoclonal antibody against -actin 購自 Chemicon (Temecula, USA)。
28. Enhanced-chemiluminesence (ECL) 購自 PerkinElmer Inc. (Waltham, MA, USA)。
29. BAY117082、U0126、SB203580、SP600125 及 L-JNKi 1 trifluoroacetate 購自 Calbiochem (San Diego, CA, USA)。
30. Lentiviruses 購自 National RNAi Core Facility Platform (Taipei, Taiwan)。
第二節 細胞培養
THP-1 (人類單核球細胞株) 以 RPMI 1640 添加 10 % 胎牛血清 (FBS)、1%
L-glutamine、1% penicillin/streptomycin 和 0.001% β-mercaptoethanol 的培養液於 37˚C 和 5% 二氧化碳的培養箱進行培養。此細胞為懸浮型細胞,細胞種植時,先 將細胞以 3500 rpm 離心 2~3 分鐘後,移除原有培養液,留下沉澱的細胞,加入 新的培養液重新沖散細胞,加入 phorbol 12-myristate 13-acetate (PMA)至終濃度 162 nM 然後分盤,使其分化三天後成為巨噬細胞。在進行細胞處理之前,用不含 FBS 的培養液潤洗 2 次,之後換成不含 FBS 培養液培養 2 小時後,進行後續實驗。
第三節 分離脂蛋白
由高脂高膽固醇飼料 (5% lard, 2% soybean oil and 0.15% cholesterol in chow diet) 餵食的紐西蘭大白兔、ST elevation myocardial infarction (STEMI) 或是 Diabetes mellitus (DM) (+) 病人取的之血液,以 3500 rpm 於 4˚C 離心 20 分鐘,取上層 之血漿。將 6 mL 血漿置於超高速離心管中,加入 1 mL 密度為 1.097 g/mL 的 NaBr 溶液,進行超高速離心 (Beckman L-100K, Rotor 70.1 Ti, 4˚C, 41000 rpm) 16 小時,取出最上層 2 mL 之 Chylomicrons 和 VLDL (0.95 < d < 1.006 g/mL)。剩餘 溶液再加入 1mL,密度為 1.283 g/mL 的 NaBr 溶液,經超高速離心 18 小時,取 出最上層液體 2 mL 之 LDL (1.019 < d < 1.063 g/mL)。剩餘溶液再加入 3 mL 密 度為 1.406 g/mL 的 NaBr 溶液,以超高速離心 20 小時,取出最上層 2 mL 之 HDL (d > 1.063 g/mL)。脂蛋白皆保存於 4˚C,並於一週內使用 。
上述分離之 LDL 在分離 native LDL 與 LDL(-) 前,需先進行透析,透析膜 事先浸泡在 50% ethanol/1x PBS 並保存於 4˚C,以透析夾密封後,浸泡在 2000 倍 體積的 1x PBS (含 1 mM EDTA) 中,於 4˚C 透析約 16 小時。取出之後,利用 蛋白質純化分析系統儀 (AKTA Explorer 10S FPLC System),以 Buffer A (0.02M Tris HCl, pH 8.0, 1 mM EDTA) 流洗陰離子交換樹脂層析管柱 (Q6 or Q12 Column),再 隨著Buffer B (含 1 M NaCl in Buffer A)的濃度逐漸增加,產生鹽離子梯度,依據
LDL 帶電性的不同,分離 native LDL 與 LDL(-)。在我們的條件下,Buffer B 濃 度在 11.5~21.5 分鐘,從 0% 提升至 15% ;Buffer B 濃度在 62~71.5 分鐘,從 20% 快速提升至 100%。在 26~30 分鐘出現第一個波峰,為 native LDL 被洗提 出來的位置;而在 66~70 分鐘出現另一個波峰,為 LDL(-)。
分別將 native LDL 和 LDL(-) 利用濃縮管以 4000 rpm 於 4˚C 離心,進行濃 縮,並在無菌操作台內以 0.22 μM Syringe Filter 過濾,保存於 4˚C。利用 DC Protein Assay Kit (Bio-Rad) 進行蛋白質濃度測定,以波長 655 nm 測量吸光值,樣 品以BSA (A6003) 為標準品得到的標準曲線換算出濃度,用於後續實驗。
第四節 游離脂肪酸牛血清蛋白複合物製備
若在室溫下為固態的脂肪酸,如棕櫚酸 (palmitic acid),其熔點為 62.9˚C ,需 先隔水加熱使其為液態,且配製時盡量避光。將純的非脂化脂肪酸 (NEFA) 與 NaOH 以莫耳數 1:1 混合均勻進行皂化反應,加入 HEPES 進行緩衝,搖晃均勻 後,將脂肪酸鈉鹽與 BSA (A9647) 以莫耳數 3:1 的比例結合,接著填充氮氣,置 於定溫培養箱以 150 rpm 於 40˚C 震盪數小時至溶液呈清澈後,在無菌操作台內 以 0.22 μM Syringe Filter 過濾,保存於 -20˚C。欲配製 5mL 的 10 mM 脂肪酸溶 液:0.05 mmole 純的非脂化脂肪酸加入 250 L 0.2 N NaOH、5 mL 25 mM HEPES 和 1.1 g BSA。
第五節 酵素結合免疫吸附法 (Enzyme-Linked ImmunoSorbent Assay, ELISA) 使用 Quantikine Human IL-1、TNF-和 IL-6 ELISA kits 偵測 2 x 105 cells in 24-well plate 之 500 L 培養液中的分泌蛋白質濃度,藉此分析分泌性蛋白質 IL- 1、TNF- 和 IL-6 的表現量。培養液保存於 -80˚C。
原理是,先將樣品中的蛋白質和 Microplate 上的 Capture 抗體結合,接著加 入和 HRP (horseradish peroxidase) 共軛鍵結的專一性抗體,鍵結在樣品蛋白質上,
形成三明治排列,再加入 HRP 的 substrate(Hydrogen peroxide 和 Chromogen),
呈色後,用 Stop Solution 終止反應,溶液呈黃色。最終,以波長 450 nm 和 540 nm 測量吸光值,扣掉背景值,並利用標準品濃度製作標準曲線,計算出樣品濃度。
步驟為以下,先以 Calibrator Diluent 適當稀釋樣品,並序列稀釋已知濃度的標準 品之後,適當加入 Assay Diluent 和樣品至 Microplate (coated with monoclonal antibody) 樣品槽中,蓋上乾淨封膜並在室溫於震盪器上搖晃 2 小時。之後,移除 樣品槽中溶液,以 1x Wash Buffer 清洗後,將殘留液體徹底去除。加入 Conjugate 抗體 (Polyclonal antibody conjugated to HRP),蓋上乾淨封膜並在室溫於震盪器上 搖晃 1 小時。接著,移除樣品槽中溶液,以 1x Wash Buffer 清洗後,將殘留液體 徹底去除。在 15 分鐘內、避光環境下,加入事先以 1:1 混和均勻的 Color Reagent A (Hydrogen peroxide) 和 Color Reagent B (Chromogen) 溶液,蓋上乾淨封膜並在 室溫於震盪器上搖晃 20 分鐘,之後加入 Stop Solution ,溶液完全混和均勻,顏 色呈黃色,在 30 分鐘內用酵素連結免疫吸附分析儀 (Thermo Scientific Multiskan EX) 以波長 450 nm 和 540 nm 測量吸光值。
針對不同抗體之 ELISA 套組,步驟上有些許差異,應參考商提供測定程序。
自 行 塗 層 的 ELISA 套 組 , 需 將 Capture 抗 體 以 PBS 稀 釋 達 建 議 working concentration,並加入 100 μL 至 Microplate 樣品槽中,室溫下放置約 16 小時,
後續步驟參考廠商提供測定程序。
第六節 細胞存活率分析
將 4 x 104 cells in 96-well plate 之 100 L 細胞培養液體移除,以 37℃ 1x PBS 潤洗後,在避光條件下,加入 40 L/well MTT (5 mg/mL PBS),於 37˚C 和 5% 二 氧化碳培養箱培養 2 小時之後,移除 MTT ,加入 50L/well DMSO,在室溫於 震盪器上搖晃約 10 分鐘,將細胞代謝產生的 Formazan 紫色結晶完全溶出。之後 用酵素連結免疫吸附分析儀以波長 540 nm 測量吸光值。結晶的生成量與活細胞數 目成正相關,以此比較對照組和實驗組之間的細胞存活率。
第七節 細胞 mRNA 表現分析
(一) 細胞 RNA 抽取
將 1 x 106 cells in 6-well plate 之細胞培養液體移除,加入 1 mL 的 TRizol 並 置於震盪器搖晃數分鐘,待細胞脫落後,收集細胞懸浮液約 1mL 加入 200 μL CIAA (Chloroform:Isoamyl alcohol = 24:1) 混合均勻,以 12000 rpm 離心 3~5 分 鐘,取出 500 μL 上清液至新試管(切勿取到中間層),加入等體積 500 μL 冰的 100% Isopropanol 混合均勻,靜置 -20˚C 約 16 小時,進行沉澱。以 12000 rpm 離 心 10~15 分鐘,去除上清液,以 200 L 冰的 70% ethanol 潤洗沉澱物(total RNA)
2 次,完全去除殘留液體,加入適當體積(10~40 μL)的 DEPC (Diethyl pyrocarbonate) -ddH2O,置於 65℃ 水浴槽 5 分鐘,使 RNA 完全回溶。之後立即置於冰上。
(二) 合成 First strand cDNA
用 NanoDrop™ 2000 Spectrophotometers 以波長 260 nm 測量吸光值定量 RNA 濃度,260/280 值落在 1.8–2.0 之間; 260/230 值落在 2.0–2.2 之間。取 5 μg RNA 並以 DEPC-ddH2O 補足體積至 13 μL,再加入 1 μL Oligo dT primer ( 50 ng/μL ) 混合均勻,置於 PCR 反應器在 70˚C 反應 10 分鐘後立即置於冰上,防 止 RNA 復性。接著,加入 4 μL 5x M-MLV-RT buffer、1 μL M-MLV-Reverse transcriptase (200 U/μL ) 和 1 μL dNTP mix (10 mM) 混合均勻,並置於 PCR 反應 器 (GeneAmp PCR system 9700) 進行反轉錄反應:先在 25˚C 反應 10 分鐘,接 著在 42˚C 反應 50 分鐘以形成 cDNA ,最後在 90˚C 作用 5 分鐘去除酵素活 性,反應完立即置於冰上,以 ddH2O 補至總體積為 100 μL,使 cDNA 濃度為 5
g/100 L 並保存於 -20˚C。
(三) 即時定量聚合酶連鎖反應 (Real-time quantitative PCR)
配置反應溶液包含: 0.4 μL Forward Primer (10 μM)、 0.4 μL Reverse Primer (10 μM) 、10 μL 2x SYBR Green mix (SensiFAST SYBR Hi ROX Kit)、18 μL ddH2O 和 2 μL cDNA,使 cDNA 濃度為 5mg/mL,於定量聚合酶連鎖反應器 (Applied Biosystems StepOnePlus™ Real-Time PCR Systems) 進行反應。反應條件:50˚C,2
分鐘;95˚C,10 秒;95˚C, 15 秒,60˚C,1 分鐘,40 個循環,測量 cycle threshold (CT);最後 95˚C,15 秒;60˚C,30 秒;9 ˚C、15 秒偵測溶點溫度 (Tm)。
(四) 引子資訊
Targets Forward sequence Reverse sequence
36B4 5’-CAACCCAGCTCTGG
AGAAAC-3’
5’-TGCCCCTGGAGATT TTAGTG-3’
hIL-1 5’-GCTGAGGAAGATGC TGGTTC-3’
5’-CGTGCACATAAGCC TCGTTA-3’
hTNF- 5’-GCTGCACTTTGGAG TGATCG-3’
5’-GTTTGCTACAACAT GGGCTACAG-3’
hIL-6 5’-TGTGAAAGCAGCAA
AGAGGCACTG-3’
5’-ACAGCTCTGGCTTG TTCCTCACTA-3’
hTGF- 5’-CAAGGATCTGGGCT GAAGTGGA-3’
5’-CCAGGACCTTGCTG TACTGCGTGT-3’
hIL-10 5’-TGCAAAACCAAACC ACAAGA-3’
5’-TCTCGGAGATCTCG AAGCAT-3’
hLOX-1 5’-GTGCTTGTCTTTGG ATGCCA-3’
5’-AGTGGGGCATCAAA GGAGAA-3’
hCD36 5’-GAGAACTGTTATGG
GGCTAT-3’
5’-TTCAACTGGAGAGG CAAAGG-3’
第八節 西方點墨分析法 (一) 細胞蛋白抽取
將 1 x 106 cells in 6-well plate 之 1 mL 細胞培養液體移除,以冰的 1x PBS 潤 洗 2 次後,加入 100 μL RIPA Buffer (150 mM NaCl, 50 mM Tris-HCl, pH 7.4 , 5
mM EDTA, pH 8.0, 0.5% NP-40, 0.5% Triton X-100, 0.1% SDS, 1 mM NaF, 1 mM PMSF, 1 mM Na3VO4, 1x Protease inhibitor cocktail),並在冰上以細胞刮勺將細胞刮 下,收集細胞懸浮液,若測量膜蛋白的表現,蛋白進行超音波震盪處理 5 秒共 3 次,之後靜置冰上 30 分鐘,接著以 12000 rpm 於 4˚C 離心 10 分鐘,取上清液 即為蛋白質,保存於 -80˚C。
(二) 樣品前處理
利用 DC Protein Assay Kit (Bio-Rad) 進行蛋白質濃度測定,需加入 Reagent S (Reagent S : Reagent A = 1 : 50),避免 detergent 干擾;以分光光度計以波長 755 nm 測量吸光值,樣品以 BSA 為標準品得到的標準曲線換算出樣品蛋白質濃度。
取 20~40μg 的蛋白質並以 ddH2O 補足體積至 14 μL,加入 6 μL 事先以體積比 2:1 混和均勻的 5x Sample Buffer (250 mM Tris-HCl, pH 6.8, 10 % SDS, 0.1 % Bromophenol blue, 50% (v/v) Glycerol) 及 10 % β-Mercaptoethanol 溶液,置於 PCR 反應器以 99.9˚C 加熱 10 分鐘後立即置於冰上,保存於 -20˚C 冰箱。
(三) 膠體配方
10 mL Running Gel (10%) (40 ~ 60 kDa)
5 mL Stacking Gel (5%)
ddH2O 4 mL ddH2O 3.4 mL
30% Acrylamide Mix 3.3 mL 30% Acrylamide Mix 0.83 mL 1.5 M Tris-HCl (pH 8.8) 2.5 mL 1 M Tris-HCl (pH 6.8) 0.63 mL
10% SDS 0.1 mL 10% SDS 0.05 mL
10% Ammonium persulfate 0.1 mL 10% Ammonium persulfate 0.05 mL
TEMED 0.004 mL TEMED 0.005 mL
(四) SDS-PAGE
膠體可以提前製備,使 Acrylamide 單體間鍵結更完全。將製作好的膠體架在
電泳槽中,加入 200 mL 1x SDS-PAGE Running Buffer (25 mM Tris-base, 192 mM Glycine, 0.1 % SDS),再將樣品及蛋白質標準品 (protein ladder) 注入樣品槽中,連 接好電極及電源,先以 70 V 進行電泳半小時,待樣品堆疊整齊後,再以 100 V 進 行 1~1.5 小時電泳。
(五) 半濕式蛋白質轉印
將膠體從電泳槽取出,並去除上膠後,浸泡於 1x Transfer Buffer (48 mM Tris- base, 39 mM Glycine, 0.037 % SDS, 20 % (v/v) Methanol, pH 8.3);PVDF 膜先浸泡 於 Methanol 1 分鐘後,接著浸泡於 1x Transfer Buffer 約 5 分鐘;濾紙亦先浸泡 於 1x Transfer Buffer。
以適量 1x Transfer Buffer 潤濕轉印槽 (Hoefer TE 70 Semiphor Transphor),依序堆 疊三張濾紙、 PVDF 膜、膠體和三張濾紙,去除所有隔層的氣泡後,蓋上上層電 極板,並放置重物於上方,插上電源,以 60 mA/8.5 x 5.5 cm2 膠體的電流進行轉 印,測量分子量 < 60 kDa 蛋白質表現,轉印 1~1.5 小時;分子量 > 60 kDa 蛋白 質,轉印 2 小時。
(六) 抗體結合免疫分析
將轉印完成之 PVDF 膜浸泡於以注射用水配製的 1x TBST Buffer (140 mM NaCl, 25 mM Tris-base, 0.05% Tween-20, pH 7.4),並在室溫於震盪器上搖晃 5 分 鐘,接著浸泡於 Blocking Buffer (5% Skim Milk Powder or BSA (A9647) in 1x TBST),
並在室溫於震盪器上搖晃 1 小時,再接著以 1x TBST Buffer 在室溫於震盪器上搖 晃,潤洗 5 分鐘共 3 次後,置換於以 Blocking Buffer 適當比例稀釋的一級抗體,
置於 4˚C 冰箱的震盪器搖晃 16 小時。隔天,將一級抗體回收,再以 1x TBST Buffer 潤洗 PVDF 膜 10 分鐘共 3 次,之後將 PVDF 膜置換於以 Blocking Buffer 適當稀釋的二級抗體 (conjugated to horseradish peroxidase) 中,並在室溫於 震盪器上搖晃 1 小時,最後以 1x TBST Buffer 潤洗 PVDF 膜 10 分鐘共 3 次。
(七) Enhanced Chemiluminescence (ECL) 系統偵測冷光訊號
將 PVDF 膜 浸 泡 於 事 先 以 體 積 比 1 : 1 混 合 均 勻 的 Enhanced Luminol Reagent 和 Oxidizing Reagent 溶液,反應 1 分鐘後,將膜放置於透明墊板上,接 著蓋上另一張透明墊板,將氣泡完全趕出,即可用 UVP 系統 (ImageQuant™ LAS 4000) 進行冷光偵測。
第九節 Knockdown 細胞中標靶基因透過 RNA 干擾
從中研院國家型核醣核酸干擾設施平台 (National RNAi Core Facility Platform) 訂購慢病毒,其具有專一性抑制人類 LOX-1 基因表現的 shOLR1,用此感染細胞。
(一) 慢病毒 Clone 資訊 shLuciferase
Clone ID promegaLuc_221s1c1 NCBI Gene ID 0
Gene Symbol LUCIFERASE Gene Description n/a
Target Taxonomy
Target Sequence 5’-CAAATCACAGAATCGTCGTAT-3’
Oligo Sequence 5’-CCGGCAAATCACAGAATCGTCGTATCTCGAGATAC GACGATTCTGTGATTTGTTTTTG-3’
Region 3UTR
shOLR1 (LOX-1)
Clone ID TRCN0000060518 NM ID NM_002543 NCBI Gene ID 4973
Gene Symbol OLR1
Gene Description Oxidized low density lipoprotein (lectin-like) receptor 1 Target Taxonomy Human
Target Sequence 5’-GCTCGGAAGCTGAATGAGAAA-3’
Oligo Sequence 5’-CCGGGCTCGGAAGCTGAATGAGAAACTCGAGTTTC TCATTCAGCTTCCGAGCTTTTTG-3’
Region CDS
(二) 慢病毒感染
將 1 x 106 之細胞回溶於 8 μg/mL polybrene 培養液,幫助慢病毒感染,將其 種至 6 cm dish,培養液總體積為 5 mL。依據 RNAi 核心以 A549 細胞株定相對 病毒量:Titer (R.I.U/L),計算 2 < M.O.I (R.I.U/cell number) < 3,滴入適當體積的 慢病毒,並放置於 37˚C 和 5% 二氧化碳的培養箱進行培養。當感染 24 小時後,
移除含有病毒的培養液,並以 10 μg/mL puromycin 的培養液進行篩選,7 ~30 天 後可進行細胞 mRNA 或蛋白質表現分析,以確認 Knockdown 效率。
第十節 Potassium-free 溶液配製
細胞做處理前,去除不含 FBS 的培養液,用 Potassium-free 溶液潤洗細胞 2 次,並換成 Potassium-free 溶液。
(一) 溶液配方 K+-free buffer
20 mM HEPES 0.95 g/200 mL
100 mM NaCl 1.17 g/200 mL
5 mM Na2HPO4 0.12 g/200 mL
1 mM CaCl2 0.028 g/200 mL
1 mM MgCl2 0.028 g/200 mL
2 mg/mL D-glucose 0.4 g/200 mL
第十一節 統計分析
實驗結果以 mean ± SD 表示,各組獨立樣本組間差異用 Student’s t-test 進行 分析,若用 two-way ANOVA 進行分析,各組間顯著差異再進行 Bonferroni t-test 多重比較分析檢定。
第三章 結果
第一節 人類 LDL(-) 會誘導巨噬細胞產生 IL-1,而 native LDL 不會
THP-1 細胞株,人類單核球細胞,以 PMA 處理三天使其分化成為巨噬細胞。
有文獻指出,serum 會影響 oxLDL 和巨噬細胞上受體的交互作用 [39, 67],為了 避免這影響,我們將含有胎牛血清 (FBS) 之細胞培養液置換成 serum-free 培養液,
並且培養 2 小時後再進行細胞處理。
STEMI 病人血液中 LDL(-) 的生理濃度約為 10~40 g/mL,實驗以 10~40
g/mL 病人 LDL(-) 刺激細胞 6 小時,之後以 ELISA測量巨噬細胞分泌到培養 液中的IL-1,結果顯示病人 LDL(-) 顯著提升 IL-1 的產生。
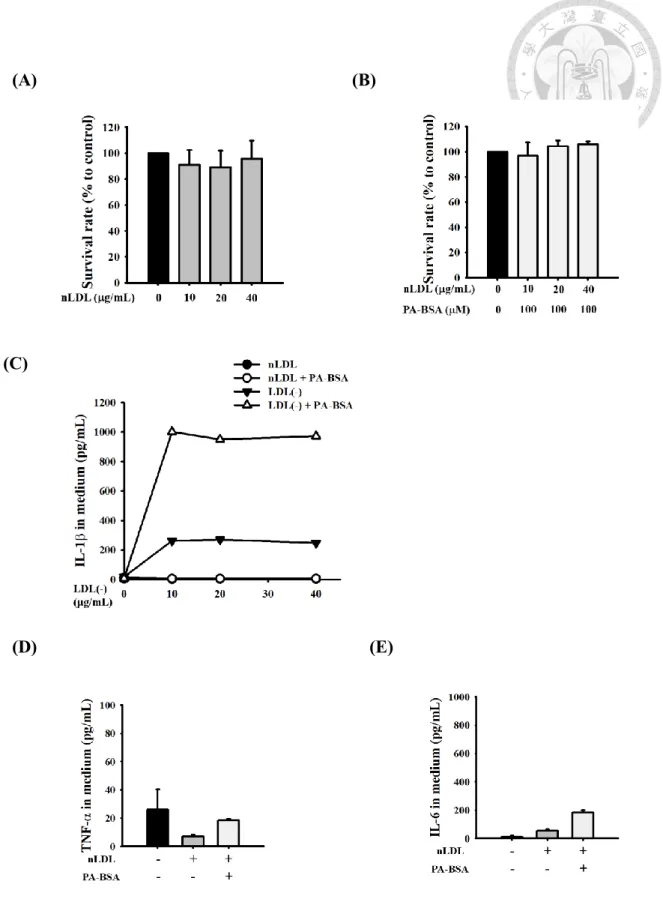
游離脂肪酸 (FFAs) 在血液中會和血清白蛋白 (serum albumin)結合,在實驗中 我們以牛血清白蛋白 (BSA)和 FFAs 做結合,形成複合物。以 10~40 g/mL 正常 人之 native LDL 和 PA 共同處理巨噬細胞,以 ELISA測量巨噬細胞分泌到培養 液中的促發炎細胞因子,IL-1、TNF-α 和 IL-6。結果顯示,而在 native LDL 處 理巨噬細胞並不會造成細胞死亡的條件下 Figure 1A & 1B,各處理組和對照組培 養液中 IL-1、TNF-α 和 IL-6 之濃度皆無顯著差異 Figure 1C, 1D & 1E。病人 LDL(-) 加上 palmitic acid-BSA (PA-BSA) 共同處理細胞後,培養液中 IL-1濃度 大幅增加Figure 1C。表示native LDL 加 PA-BSA 不會更造成巨噬細胞產生發炎 反應,而 LDL(-) 會顯著提升 IL-1 的產生,LDL(-) 加上 PA-BSA 會更增加 IL- 1 的產生。
第二節 兔子 LDL(-) 單獨在巨噬細胞引起小幅度 IL-1 產生
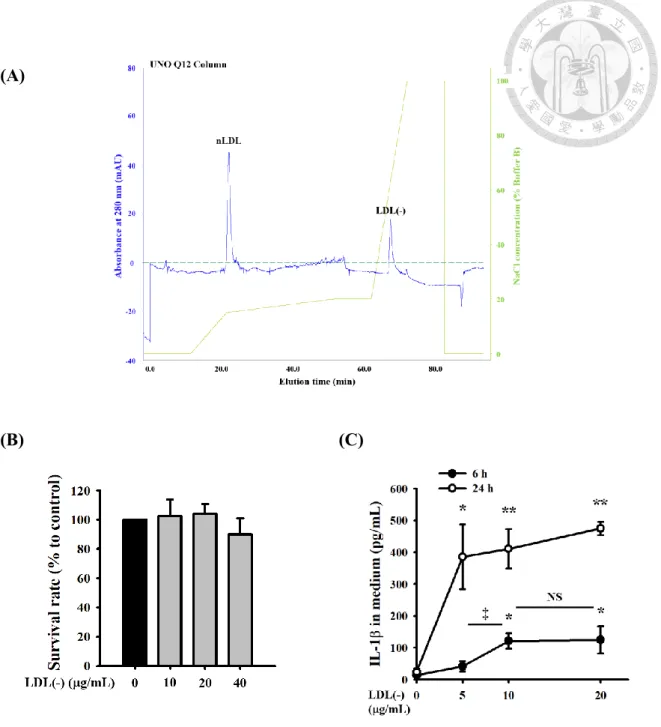
實驗使用的 LDL(-) 為從餵食高膽固醇酯飲食之兔子循環中分離出來之
LDL(-),由於高血脂兔子為易引發 atherosclerosis 之動物模型 [39],且由於病人的 LDL(-) 取得不易,加上兩種來源的 LDL(-) 引起 IL-1 產生的狀況相似,所以用 兔子的 LDL(-) 來做後續的實驗。
我們從兔子血漿分離之 LDL,依帶電性不同,透過陰離子交換樹脂管柱分離 出比較帶負電之 LDL(-) Figure 2A。在 LDL(-) 處理巨噬細胞並不會造成細胞死
亡的條件下 Figure 2B,分別以 5~20 g/mL LDL(-) 處理巨噬細胞 6 小時和 24 小時,透過 ELISA測量巨噬細胞分泌到培養液中的IL-1。結果顯示,在 6 小時,
10 g/mL 以上之 LDL(-) 比起對照組,培養液中 IL-1濃度顯著提升;在 24 小 時,5 g/mL 以上之 LDL(-) 比起對照組,培養液中 IL-1濃度顯著提升Figure 2C。且隨著LDL(-) 處理的時間從增加至小時,培養液中IL-1的濃度顯著增 加Figure 2C。綜合以上,10 g/mL 兔子 LDL(-) 在巨噬細胞對於 IL-1產生已 有顯著之效應。
第三節 Palmitic acid 在巨噬細胞對 IL-1 的產生沒有影響
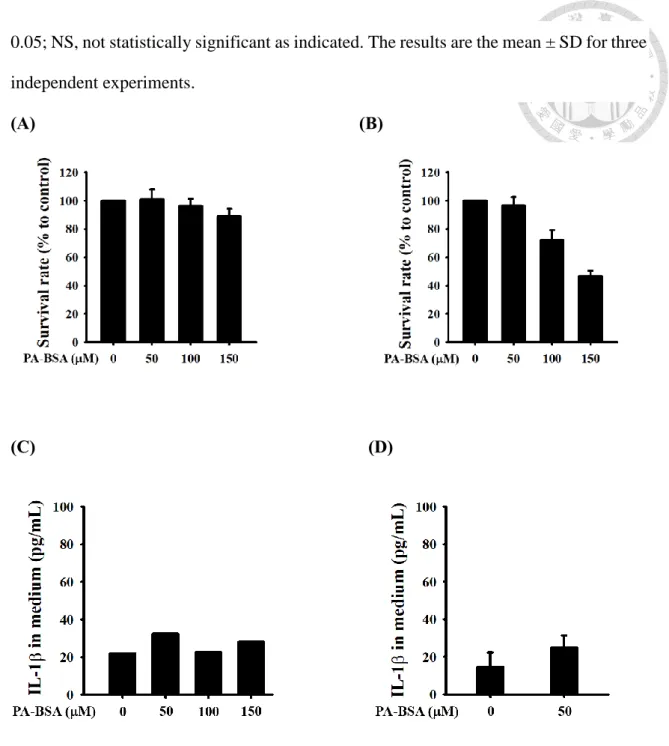
Palmitic acid (PA) 為十六個碳的飽和脂肪酸,生理濃度在上下。我們以
到不同濃度之PA 處理細胞 6 小時,細胞存活率皆在八成以上Figure 3A;但是處理時間增長為 24 小時,100 PA 會使細胞存活率掉至八成以下,
150 PA 存活率更不到六成,其隨著濃度增加,存活率下降 Figure 3B;可能 是細胞在 serum-free 培養液中長時間培養。不過,在不會造成細胞死亡之 PA 濃 度處理細胞,皆不會造成巨噬細胞引起 IL-1產生,培養液中IL-1濃度比起對 照組,無顯著差異Figure 3C & 3D。
第四節 兔子 LDL(-) 和 palmitic acid、stearic acid 在巨噬細胞對 IL-1 的產生 之影響有協同作用,其他測試的脂肪酸則無影響
已知以PA 單獨處理巨噬細胞 6 小時,並不會誘導 IL-1分泌;LDL(-) 單獨 處理細胞則會引起 IL-1產生。接著我們以LDL(-) 和 PA共同處理巨噬細胞。結 果發現,在細胞存活率達九成以上的情況下Figure 4A & 4B),IL-1分泌至培養 液的量,約達到 LDL(-) 單獨處理細胞的五倍量 Figure 4C。且 PA 與 10 g/mL LDL(-) 即可以達到接近 PA 與 20 g/mL LDL(-) 引起的 IL-1的產量 Figure 4C。 接著以10 g/mL LDL(-) 和不同濃度()的PA 共同處理巨噬細胞,PA 濃度在 150 時,IL-1分泌的量達到高峰Figure 4D。因此綜合以上,10 g/mL LDL(-) 和 100 PA 共同處理 THP-1 巨噬細胞,對於 IL-1的產生可以有協
同的效應且達到高峰。
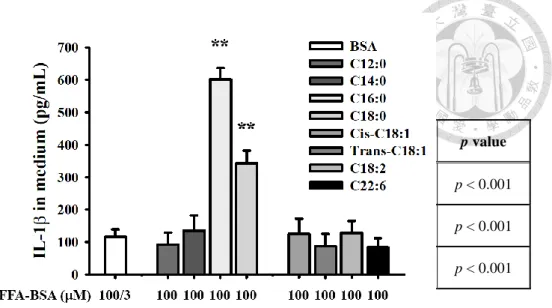
接著探討其他脂肪酸和 LDL(-) 是否也對於在巨噬細胞誘導 IL-1有類似的 作用。將 10 g/mL LDL(-) 分別和不同的飽和游離脂肪酸,包括月桂酸 (lauric acid, C12:0)、肉豆蔻酸 (myristic acid, C14:0)、棕櫚酸 (palmitic acid, C16:0) 和硬脂酸 (stearic acid, C18:0),以及幾種不飽和脂肪酸,包括油酸 (oleic acid, cis-C18:1)、反 式油酸 (elaidic acid, trans-C18:1)、亞麻油酸 (linoleic acid, C18:2) 和二十二碳六烯 酸 (docosahexaenoic acid, C22:6),其共同處理巨噬細胞 6 小時。(Figure 4E) 的結 果顯示,跟單獨處理 LDL(-) 之處理組比,只有以 palmitic acid 或 stearic acid 處 理,會顯著提高 IL-1的產生,且palmitic acid (PA) 比 stearic acid (SA) 處理組 產生的量較高;其餘脂肪酸皆沒有顯著作用。結果顯示,只有飽和脂肪酸 PA、SA 對於 LDL(-) 誘導 IL-1產生有協同作用。
上述實驗的結果顯示,PA 能夠更提高 LDL(-) 在巨噬細胞誘導 IL-1的產生。
Figure 4F 以 LDL(-) 單獨或是加上PA一同處理細胞,分為四個處理組;分別為 (1) 無LDL(-)/無 PA (-)、(2) 有 LDL(-)/無 PA、(3) 無 LDL(-)/有 PA 和(4) 有 LDL(-)/有 PA。透過 two-way ANOVA 統計分析結果為,LDL(-) 對於 IL-1的產生顯著影響
p < 0.001);PA 對於 IL-1的產生也顯著影響p < 0.001);且在 LDL(-) 和 PA 雙 因子之間有顯著交互作用 (p < 0.001)。除了無LDL(-)/無 PA 和 (3) 無 LDL(-)/
有PA 對於 IL-1的產生無顯著差異,其他四組之間皆有顯著差異。這結果顯示,
PA 需要和 LDL(-) 一同處理細胞才會對 IL-1的產生有顯著影響。
第五節 LDL(-) 在巨噬細胞引起 IL-1 產生經由 LOX-1
實驗室先前的研究已經指出, LDL(-) 在巨噬細胞誘導 IL-1產生經由LOX- 1 [4],且也有文獻指出 PA 在巨噬細胞會促進 LOX-1 表現 [64],所以進一步探 討PA 是否也透過 LOX-1 作用使 LDL(-) 誘導 IL-1 產生增加。
我們利用帶有抑制人類 LOX-1 基因表現的 shOLR1 之慢病毒感染巨噬細胞,
將 LOX-1 基因 knockdown 後,在此細胞株處理 LDL(-) 和 PA 6 小時,LDL(-)
單獨處理組別和 LDL(-) 加 PA 組別,培養液中 IL-1之濃度皆比knockdown 控 制組顯著的低 Figure 5A。另外,LDL(-) 加 PA 處理,和 LDL(-) 單獨處理之 LOX-1 mRNA 表現並無顯著差異,且 PA 對於 LOX-1 之 mRNA 表現沒有顯著 影響p = 0.515);LDL(-) 和 PA 雙因子之間也無顯著交互作用 p = 0.678)Figure 5B。另外,LDL(-) 和 PA 在巨噬細胞上皆不造成清道夫受體,CD36 基因表現的 影響 Figure 5B。而從LOX-1 蛋白質表現來看,無論 LDL(-) 加 PA 處理 6 小 時或 24 小時,皆不會造成比單獨處理 LDL(-)更增加 LOX-1 表現 Figure 5C。
這些結果顯示,LDL(-) 獨立經由 LOX-1 誘導在巨噬細胞 IL-1的產生;而 PA 不是經由 提高 LOX-1 基因或蛋白質表現而促進 IL-1的產生。
第六節 Palmitic acid 不會促進 LDL(-) 進一步引起 NF-B 活化
IL-1 產生需要兩條獨立的訊號傳遞,其中第一條路徑為 NF-B 活化。首先,
NF-B 的 inhibitor,I-B 被磷酸化後,會離開 NF-B 且被降解,接著 NF-B 被 磷酸化後,進入細胞核中,作為轉錄因子,調控下游基因表現,經過轉錄、轉譯提 高 pro-IL-1表現。
巨噬細胞在處理 LDL(-) 6 小時之下,I-B的磷酸化程度顯著增加Figure 6A;且pro-IL-1的蛋白表現量大量增加,約為控制組的倍Figure 6B。實驗 結果顯示,LDL(-) 在巨噬細胞會誘導 NF-B 活化,並且造成 pro-IL-1大量表現。
再分析PA是否也透過活化NF-B 進一步使 LDL(-) 單獨引起 pro-IL-1表現更 增加。實驗設計分為以下四個處理組,分別為 (1) 無 LDL(-)/無 PA、(2) 有 LDL(-) /無 PA、(3) 無 LDL(-)/有 PA 和(4) 有 LDL(-)/有 PA;結果發現,(2) 有 LDL(-)/無 PA 和 (4) 有 LDL(-)/有 PA 處理組在 I-B的磷酸化程度和pro-IL-1蛋白表現 皆沒有顯著變化Figure 6A & 6B。並且透過 two-way ANOVA 統計分析結果為,
LDL(-) 對於 I-B磷酸化程度顯著影響,但PA 對於 I-B磷酸化程度則無顯著 影響p = 0.699),雙因子之間也無顯著交互作用 (p = 0.398);LDL(-) 對於 pro-IL- 1蛋白表現顯著影響,但PA 對於 pro-IL-1蛋白表現沒有顯著影響p = 0.558),
且雙因子之間也沒有顯著交互作用 (p = 0.427) Figure 6C)。上述結果表示,PA 沒 有使 LDL(-) 誘導 NF-B 活化的程度更增加。結果顯示,PA 不是透過促進 pro- IL-1表現更增加而促進mature IL-1產生更增加。另一方面也顯示,PA 可能是透 過活化第二條路徑,也就是透過 NLRP3 inflammasome/Caspase-1 的活化,促進 LDL(-) 誘導的 IL-1產生。
我們也探討了,是否 NF-B 路徑或其他 MAPK (mitogen-activated protein kinase) 路徑也參與在 LDL(-) 和飽和脂肪酸在巨噬細胞誘導 IL-1 產生,我們透 過 NF-B 抑制劑和幾個 MAPK 抑制劑進行分析。在巨噬細胞處理 10 g/mL LDL(-) 和50 M stearic acid (SA) 24 小時,產生的 IL-1為±pg/mL。
細胞分別前處理多種抑制劑 1 小時,其中 Bay11-7082 為 NF-B 抑制劑;U0126 為 MEK1/2 抑制劑,而 ERK1/2 受上游 MEK1/2 調控;SB203580 為 p38 抑制 劑;SP600125 和 L-JNKi 為 JNK 抑制劑,接著再同時處理 LDL(-) 和 SA 24 小 時。結果發現,只有在前處理 Bay11-7082 和 U0126 的組別,比起前處理 DMSO 的細胞,培養液中 IL-1 的濃度顯著下降 Figure 6D。這結果顯示LDL(-) 和 SA 在巨噬細胞引起 IL-1 產生的作用,會經過 NF-B 和 ERK1/2 活化路徑。
第七節 Palmitic acid 不會促進 NF-B 下游基因 IL-1、TNF- 和 IL- 的 mRNA 和蛋白質表現
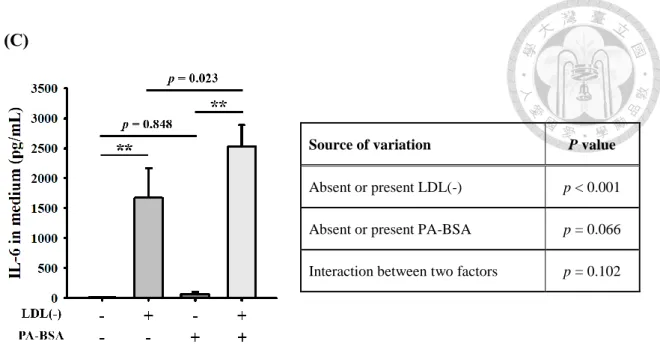
NF-B 活化,除了調控 IL-1表現,還會調控其他促發炎因子TNF-和IL-6 等基因表現。實驗結果顯示,巨噬細胞在處理 LDL(-) 6 小時之下,IL-1、TNF-
和IL-6 的 mRNA 表現顯著增加 Figure 7A。但在加上PA 這個因子後發現,單 獨處理 LDL(-) 和處理 LDL(-)與 PA 的情況下 IL-1、TNF-和IL-6 mRNA 表 現相似 Figure 7A。two-way ANOVA 統計分析結果為,LDL(-) 對於 IL-1、TNF-
和IL-6 mRNA 表現顯著影響,但PA 對於 IL-1、TNF-和IL-6 mRNA 表現則 沒有影響p = 0.320/0.440/0.543),且在 LDL(-) 和 PA 雙因子之間沒有顯著交互作 用 (p = 0.188/0.471/0.410)Figure 7A bottom。TNF-和IL-6 跟 IL-1一樣會被分
泌至細胞外,而LDL(-) 對於 TNF-的蛋白產生有顯著影響p < 0.001);PA 對於 TNF-的蛋白質產生也顯著增加p = 0.020),雙因子之間也有顯著交互作用 (p = 0.024)Figure 7B,但比起對於 IL-1的產生,p value 較大,影響較小Figure 7B。 最後,LDL(-) 對於 IL-6的蛋白產生有顯著影響p < 0.001);但 PA 對於 IL-6的 蛋 白 產 生 沒 有 顯 著影 響p = 0.066) ; 雙 因 子 之 間 也 沒 有 顯 著 交 互 作 用 (p = 0.102)Figure 7C,表示 LDL(-) 對於 IL-6的產生之影響為獨立的,不依賴PA 的 存在。綜合以上結論,PA 對於 LDL(-) 誘導IL-1的產生,影響較大,且可以促 進培養液中IL-1的量增加至約五倍Figure 4F),顯示PA 在其中扮演重要角色。
而先前也證實 native LDL 和 PA 一同處理巨噬細胞,亦不會誘導 TNF- 和 IL-6 的產生。
第八節 Palmitic acid 促進 LDL(-) 誘導 IL-1 產生不是經由 phagocytosis 和 lysosomal degradation
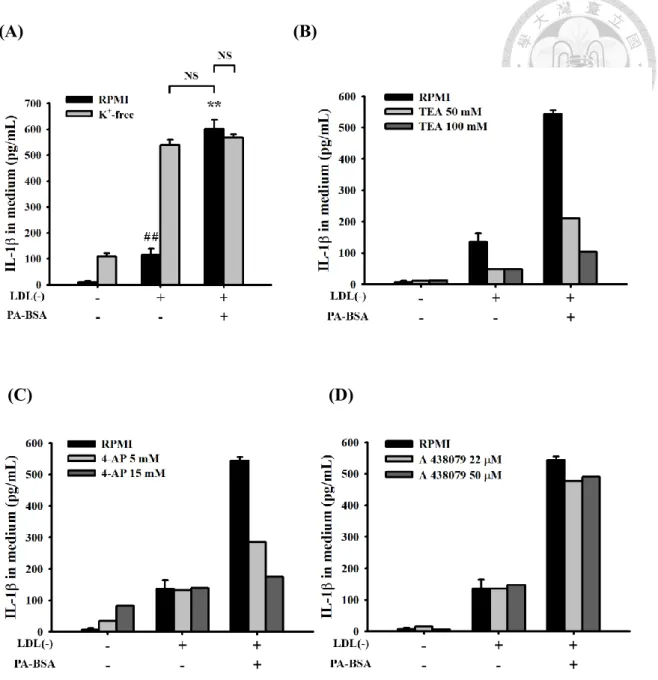
我們推測 palmitic acid 可能參與在 IL-1 產生所需之第二條路徑,NLRP3 inflammasome/caspase-1 路徑,而 NLRP3 inflammasome/caspase-1 的活化,已知可 能透過 phagocytosis [29, 68]、lysosomal degradation [26, 29] 和 potassium efflux [27, 28, 68] 等。因此我們探討 palmitic acid 是否透過上述三種途徑活化 NLRP3 inflammasome/caspase-1,進而促進 IL-1 產生增加。
Cytochalasin D 能夠限制肌動蛋白的 polymerization 和 elongation,藉此抑制 細胞進行 phagocytosis [69],且文獻指出其能抑制 monosodium urate (MSU) 之 phagocytosis [70]。我們將巨噬細胞前處理 cytochalasin D 約 1 小時,接著同時處 理,LDL(-) 和 PA 6 小時。結果發現,和控制組(無前處理 cytochalasin D 之細 胞)相比, LDL(-) 誘導之 IL-1 產生並不受影響;也不會影響 PA 促進 LDL(-) 誘導之 IL-1 產生增加的作用 Figure 8A。這結果顯示, LDL(-) 可能不是經由 phagocytosis 在巨噬細胞誘導 IL-1 產生;PA 也可能不是透過 phagocytosis 活化 NLRP3 inflammasome/caspase-1 進而造成 LDL(-) 誘導之 IL-1 產生增加。而