國立宜蘭大學動物科技 國立宜蘭大學動物科技 國立宜蘭大學動物科技
國立宜蘭大學動物科技學系 學系 學系 學系(研究所 研究所 研究所 研究所) 碩士論文 碩士論文 碩士論文
碩士論文
Department of Animal Science National Ilan University
Master Thesis
飼料飼料飼料
飼料添加丁酸添加丁酸添加丁酸鈉添加丁酸鈉鈉鈉及羥丁胺酸對離乳仔豬生長及羥丁胺酸對離乳仔豬生長及羥丁胺酸對離乳仔豬生長性能及羥丁胺酸對離乳仔豬生長性能性能性能、、、、腸道發育腸道發育腸道發育及腸道發育及及免及免免免 疫反應疫反應疫反應
疫反應之影響之影響之影響 之影響
Effects of diets supplemented with sodium butyrate and threonine on the growth performance, intestinal development,
and immune responses of weaned pigs
指導教授指導教授指導教授
指導教授:::李德南:李德南李德南 博士李德南 博士博士 博士
Der-Nan Lee, Ph. D.
研究生研究生
研究生研究生:::余秉樺:余秉樺余秉樺 余秉樺
Bing-Hua Yu
本授權書所授權之論文為授權人在國立宜蘭大學 動物科技研究所 系所
96 學年度第 2 學期取得 博士 ▓ 碩士 學位之論文。
論文名稱: 飼料添加丁酸鈉及羥丁胺酸對離乳仔豬生長性能、腸道發育及免疫反應之 影響
指導教授: 李德南 教授
茲同意將授權人擁有著作權之上列論文全文資料,非專屬、無償授權國立宜蘭大學及國 家圖書館,不限地域、時間與次數,以光碟或其他各種數位化方式將上列論文重製散布、
發行或上載網路,提供讀者基於個人非營利性質之線上檢索、閱覽、下載或列印。
授權選項:
■ 校內、校外公開
□ 校內公開,校外因上列論文為授權人向經濟部智慧財產局申請專利之 附件或相關文件之一 (申請專案號: ),請 於 年 月 日後再將上列論文公開或上載網路。
□ 校內公開,校外因上列論文尚未正式對外發表,請於 年 月 日 後再將上列論文公開或上載網路。
□ 校內公開,校外不予公開
數位化公開方式若未勾選本人同意視同授權校內、校外公開。
授權人: 余秉樺 身分證字號:
E-mail: [email protected]
簽名或蓋章: 中華民國 97 年 8 月 6 日
誌謝 誌謝 誌謝 誌謝
本論文承蒙恩師李德南博士耐心指導與激勵,並自大學進入實驗室後,
給予秉樺生活與實驗上的經驗傳授,以及在百忙中指導秉樺的論文撰寫與 修改論文,在此表示由衷感激。
研究期間,感謝東華大學翁慶豐老師與台灣大學朱有田老師給予我參與 他們實驗室訓練之機會,並提供許多寶貴建議,獲益匪淺。同時,非常感 激諸位評審老師百忙中撥空參加秉樺論文口試,並給予本論文多方面的教 導與指正,使本論文更加完整。
本試驗承農委會 95 農科-5.1.1-牧-U1 與 96 農科-2.1.1-牧-U1(13)計畫經 費補助,僅此申謝。衷心感謝比利時 INVE-Nutri-Ad 公司提供丁酸鹽與德 國 Degussa 公司慨贈羥丁胺酸,由衷感激。
近修期間,承蒙眾多師長、同學、朋友及家人,給予秉樺無限鼓勵與支 持。此外,感謝本實驗室莊雨軒學長、邱后妍學姊、洪玉祥同學、呂効儒、
陳彥廷、黃伯驥、林皇志學弟以及王若綺、陳乙潁學妹之協助,銘記在心,
特此致謝。因感謝的人數眾多,故無法一一列出,特此表示由衷的感謝。
最後感謝我的父母親與我的哥哥,感謝他們在經濟、生活及精神上的鼓 勵與支持,讓我在碩士班求學期間能無後顧之憂的全心投入於研究當中,
秉樺在此獻上我最真誠的感謝。
余秉樺 民國九十七年六月
中文摘要 中文摘要 中文摘要 中文摘要
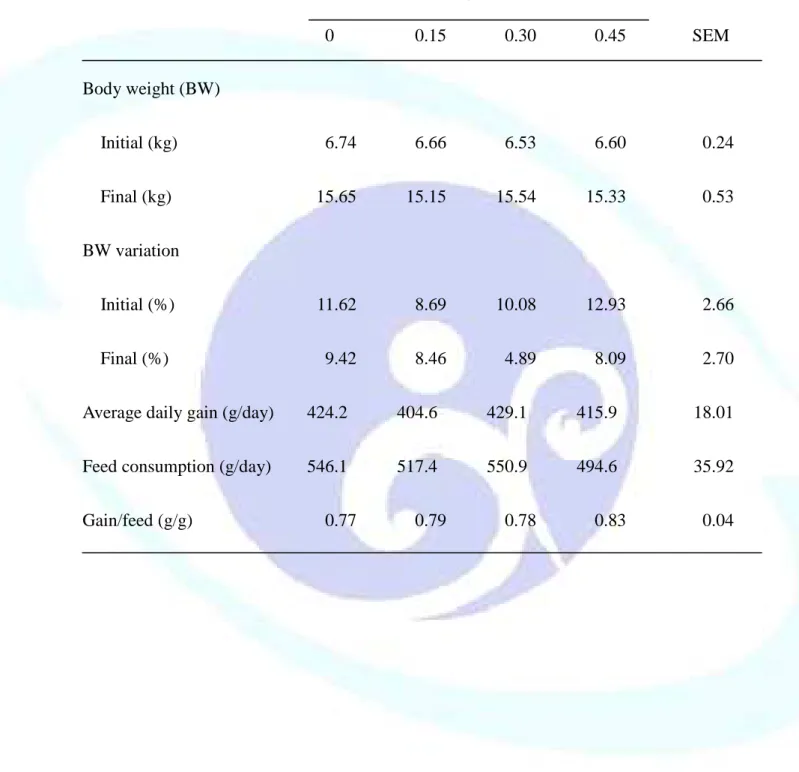
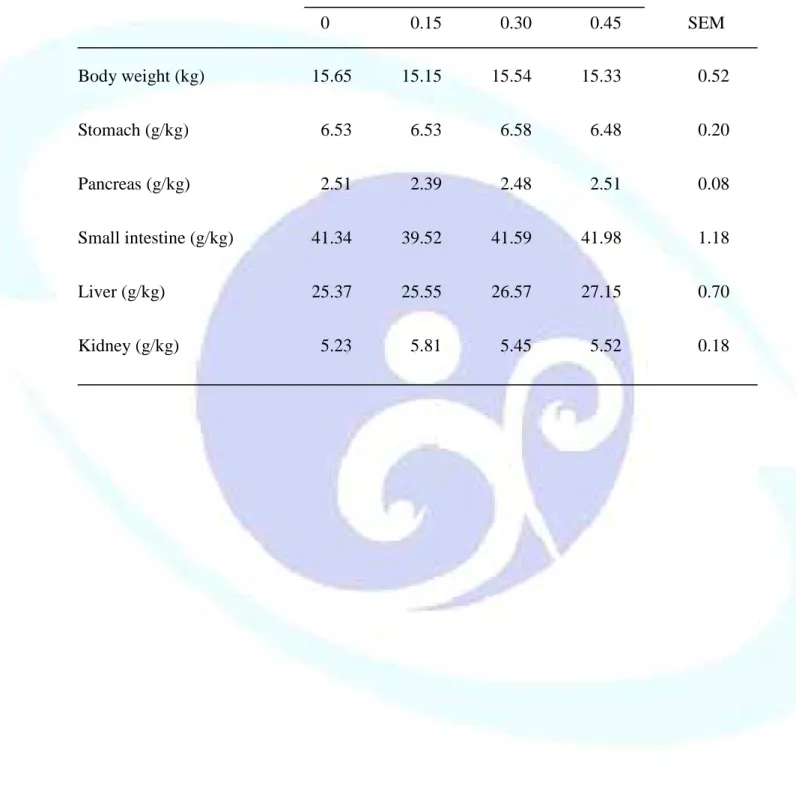
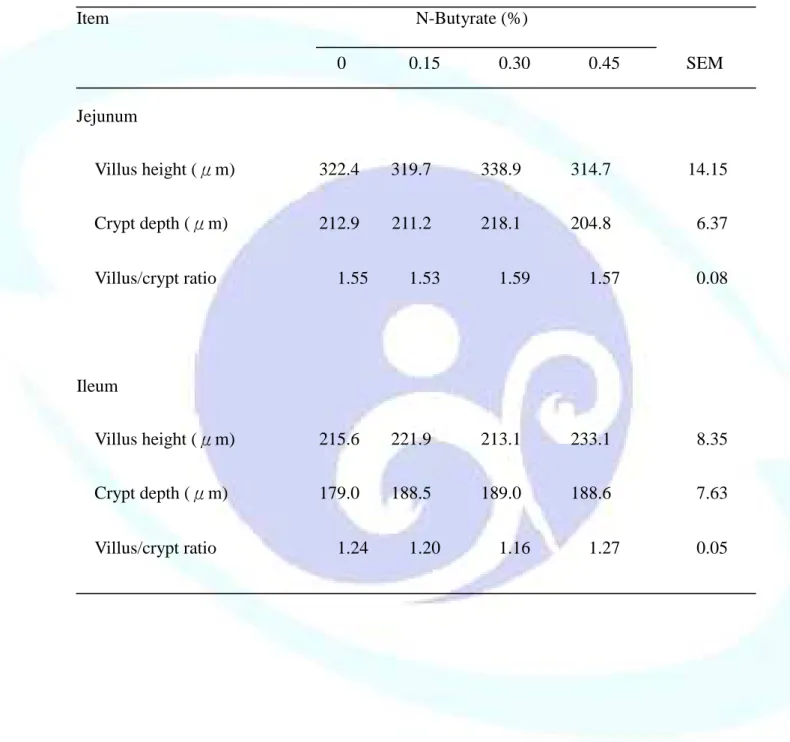
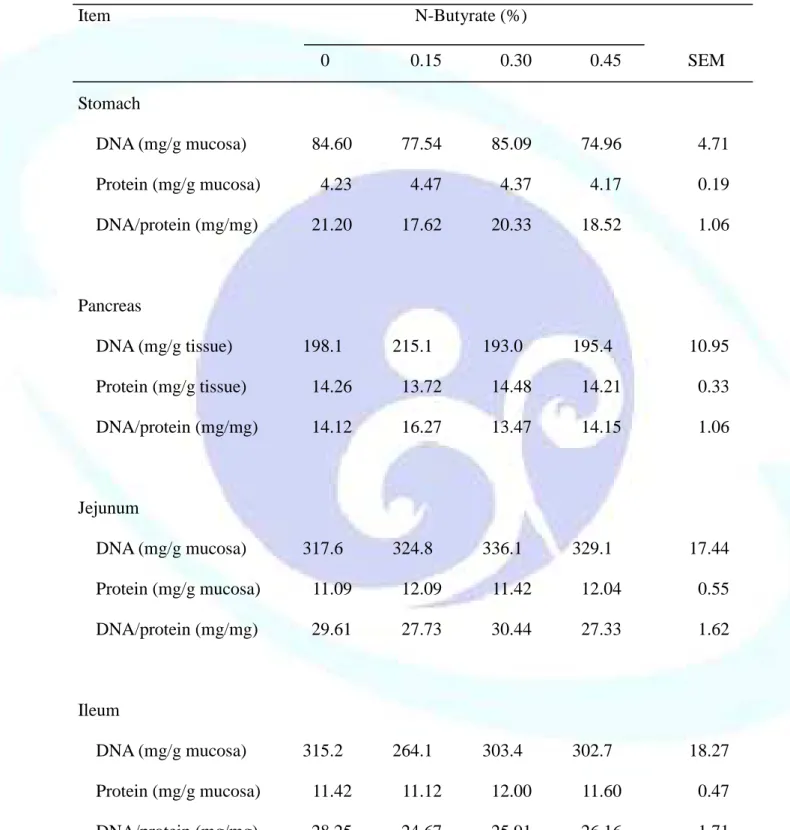
仔豬於離乳階段消化道尚未發育完整,飼料供應丁酸鹽(Sodium butyrate, NaB)可滋養腸細胞並促進其分化成熟,羥丁胺酸(Threonine, Thr)則可提供蛋 白質合成之胺基酸骨架材料來源,故本試驗目的探討飼料添加 NaB 與 Thr 對離乳仔豬生長性能、消化道發育及免疫反應之影響。試驗一利用 48 頭三 週齡雜交離乳仔豬隨機餵飼 0、0.15、0.30 及 0.45%未包覆 NaB 之四種飼料 處理,每欄 2 頭,每處理 6 欄,餵食 21 天後全部犧牲以分析離乳仔豬腸道 發育與消化道相關淋巴組織免疫反應。結果發現,飼料添加 NaB 未能顯著 影響離乳仔豬生長性能、消化器官重、小腸黏膜形態、黏膜 DNA 濃度及消 化酵素活性、小腸食糜大腸桿菌或乳酸菌數,及血球組成或體液 IgA 濃度。
其中發現飼料添加 0.45% NaB 可較對照組增加空腸食糜黏液蛋白濃度達 21%,但未達顯著差異水準(P > 0.05)。飼料添加 0.15% NaB 有最高之派亞 氏結淋巴細胞經植物血凝素(Phytohemagglutinin, PHA)刺激之增生反應,而 添加 0.45% NaB 可增加周邊血液單核細胞(Peripheral blood monocyte, PBMC) 分泌干擾素-γ(interferon-γ, IFN-γ)之能力,但卻顯著降低脾臟細胞經 PHA 刺激之增生反應(P < 0.05)。
試驗二選取 64 頭三週齡雜交離乳仔豬,利用 2×2 複因子設計隨機餵食 添加脂肪包覆型式之 NaB 組(0-14 天:添加 0.09%;15-42 天:添加 0.06%)、
添加 Thr 組(0-6 週:添加 0.1%)、同時添加 NaB 與 Thr 組及皆未添加當作對
照組,每處理 4 欄,每欄 4 頭,試驗第 21 天時每欄犧牲 2 頭豬隻以評估腸 道生理與免疫反應,其餘豬隻持續餵飼飼料至 42 天結束。結果發現,飼料 處理未影響仔豬全期之生長性能,但飼料添加 Thr 可減少試驗結束時體重 變異程度。飼料中添加 NaB 或 Thr 皆可顯著提升仔豬離乳後第 21 天血漿中 Thr 濃度(P < 0.02),且 Thr 處理組可延續提高至第 42 天(P < 0.01),但未影 響血清尿素態氮濃度。飼料添加 NaB 和 Thr 分別具有提高試驗 21 天迴腸胺 基胜肽酶與鹼性磷酸酶活性(P < 0.05),飼料添加 NaB 可增加第 21 天空腸食 糜黏液蛋白濃度,而飼料添加 Thr 則會增加空腸絨毛部位酸性杯狀細胞大 小,而 NaB 與 Thr 同時影響空腸絨毛部位中性杯狀細胞之數目與其面積。
飼料處理未能顯著影響試驗 21 與 42 天之免疫球蛋白濃度,試驗處理亦未 影響試驗 21 天之膽汁、小腸黏膜及其食糜之 IgA 濃度。但試驗 21 天時,
飼料添加 NaB 會增加脾臟細胞經由刺激之淋巴細胞增生反應(P < 0.05)並同 時降低脾臟細胞 CD4+含量,而飼料添加 Thr 具有增加 PBMC 經伴刀豆球蛋 白 A (concanavalin A, con A)刺激之淋巴細胞增生反應(P < 0.05)。
飼料添加不同形式 NaB 皆未影響小腸結構或功能,添加 0.45%未保護之 NaB 具有增加 PBMC 分泌 IFN-γ之能力,而添加脂肪包覆之 0.15-0.2% NaB (保育 I 期-保育 II 期)易會抑制脾臟細胞之 CD4+比例。飼料添加 Thr 可增 加離乳仔豬空腸酸性杯狀細胞數目與迴腸鹼性磷酸酶活性,亦可提高 PBMC 之增生反應,但未與 NaB 交互影響離乳仔豬腸道發育或免疫反應。
Abstract
Sodium butyrate (NaB) is one of energy source for epithelial cells of intestinal, and plays a central role in maintaining the mucosal barrier in the gut.
Threonine (Thr), an indispensable amino acid for pig nutrition, is also main role for the maintenance of mucosal proteins because some of these proteins contain a high amount of Thr in their peptide backbone. The disadvantage of immature in gut causes poor growth during weanling of pigs. The aim of this experiment was to test whether the effects of diets supplemented with NaB and Thr on growth performance, intestinal development, and immune responses of weaned pigs.
In Exp. 1, forty-eight pigs (21 day weaned) were randomly allotted into four treatments including 0, 0.15, 0.30, and 0.45% non-protected NaB. Pigs were fed 2 pigs per pen, and each treatment had 6 pens for 21 days. At the end of the Exp., all of the pigs were sacrificed to investigate the effects of dietary treatment on intestinal structure and function as well as gut-associated lymphocyte tissues immune responses. The results showed that dietary supplemented with NaB did not affect the growth performance, organ weights, small intestine mucosal morphology, mucosal DNA concentration, digestive enzymes, the intestinal microbial ecosystem, blood composition, and humoral IgA concentration of
weaned pigs. Interestingly, dietary supplemented with 0.15% NaB had the highest proliferation by phytohemagglutinin (PHA) stimulation on lymphocytes of Peyer’s patch. Dietary supplemented with 0.45% NaB increased jejunal digesta mucin concentration about 21% compared with the control group (P >
0.05), while significantly increased IFN- γ secretion of peripheral blood monocytes (PBMC) and decreased in proliferation of splenocytes by PHA stimulation were observed.
In Exp. 2, sixty-four pigs (21 day weaned) randomly allotted to four treatments including fat-coated NaB (days 0 to 14: supplemented with 0.09%;
days 15 to 42: supplemented with 0.06%), Thr (days 0 to 42 supplemented with 0.10%), NaB plus Thr and control groups. Pigs were fed 4 pigs per pen, and each treatment had 4 pens. And on day 21, 2 pigs per pen were randomly sacrificed to investigate the effects on gut physiology and immune responses.
The remain pigs were fed to the end of the Exp. The data revealed that dietary NaB and Thr supplement failed to affect the growth performance, serum urea nitrogen and immunoglobulin concentrations, bile, small intestinal mucosa, and digesta IgA concentration during 42 days period of weaned pigs, but dietary supplemented with Thr decreased the variation of body weight (P < 0.01). On day 21, dietary supplemented with NaB and Thr were significantly increased
plasma Thr concentration (P < 0.02) and ileal amino-peptidase N and alkaline phosphatase (P < 0.05), while dietary supplemented with Thr was sustained enhance plasma Thr concentration on day 42. On day 21, dietary supplemented with Thr increased the villus acidic goblet cell size of jejunum, and NaB plus Thr had a synergistic effect on the villus neutral goblet cell numbers and area of jejunum. Dietary supplemented with NaB had significantly increased the splenocyte proliferation by PHA stimulation (P < 0.05) and decreased in CD4%
of spleen, while dietary supplemented with Thr was significantly increased PBMC proliferation by concanavalin A stimulation (P < 0.05) on day 21.
Conclusively, dietary supplemented with unprotected NaB has no effects on small intestinal structure and functions, and dietary supplemented with 0.45%
NaB has significantly increased IFN- γ concentration, but dietary supplemented with fat-coated 0.09-0.06% NaB during phase 1 and phase 2 has improved immune response of weaned pigs. Dietary supplemented with Thr is increased jejunal acidic goblet cell numbers, ileal alkaline phosphatase, and PBMC proliferation of weaned pigs, but no effects of interaction with NaB on intestinal development and immune responses of weaned pigs.
目錄
頁次
誌謝---i
摘要--- ii
英文摘要--- iv
目錄--- vii
表次--- x
圖次--- xiii
I、前言--- 1
II、文獻檢討--- 3
一、仔豬腸道與免疫發育過程--- 3
(一)仔豬腸道黏膜與免疫組織之結構組成--- 3
(二)仔豬出生前後之腸道與免疫系統發育--- 5
(三)仔豬離乳階段之腸道與免疫系統發育--- 8
二、影響仔豬離乳期間腸道與免疫發育之飼養因素--- 12
(一)離乳年齡--- 12
(二)飼料型態--- 13
(三)飼養環境--- 14
三、飼料抗原與營養成分對離乳仔豬腸道與免疫之影響--- 15
(一)飼料抗原--- 16
(二)功能性脂肪酸--- 17
1.多元不飽和脂肪酸--- 17
2.中鏈脂肪酸--- 18
3.有機酸與短鏈脂肪酸--- 19
4.丁酸--- 21
(三)功能性胺基酸---27
1.麩醯胺 ---28
2.精胺酸 ---29
3.羥丁胺酸 ---30
III、試驗部份---36
一、丁酸鹽添加量對離乳仔豬生長性能、腸道發育及免疫反應之影響- 36 摘要--- 36
前言--- 37
材料與方法 ---39
結果 ---44
討論 ---59
結論 ---63
二、飼料添加丁酸鹽與羥丁胺酸對離乳仔豬生長性能、腸道發育及免疫
反應之影響--- 64
摘要--- 64
前言--- 65
材料與方法---68
結果 ---74
討論 ---90
結論 ---95
IV、總結論 ---96
V、參考文獻--- 97
表次
頁次
文獻檢討--- 3
表一. 仔豬黏膜免疫系統發育階段---5
表 1-1. 基礎飼料配方和營養分組成--- 41
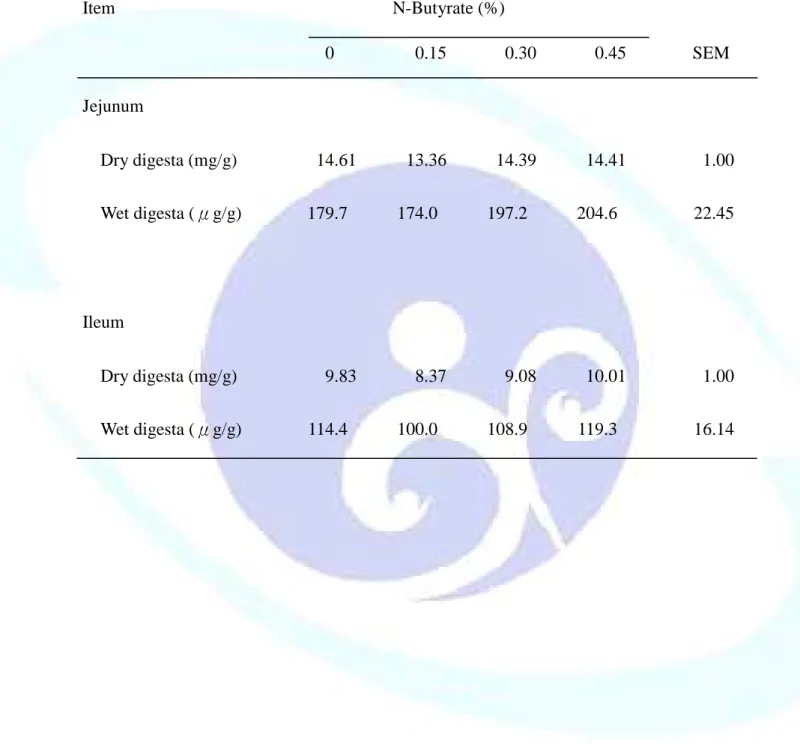
表 1-2. 飼料添加丁酸鹽對離乳仔豬生長性能之影響--- 45
表 1-3. 飼料添加丁酸鹽對離乳仔豬器官重量之影響--- 47
表 1-4. 飼料添加丁酸鹽對離乳仔豬小腸絨毛形態之影響--- 48
表 1-5. 飼料添加丁酸鹽對離乳仔豬胃黏膜、胰臟及小腸黏膜 DNA 和蛋 白質濃度之影響--- 49
表 1-6. 飼料添加丁酸鹽對離乳仔豬小腸食糜黏液蛋白濃度之影響--- 50
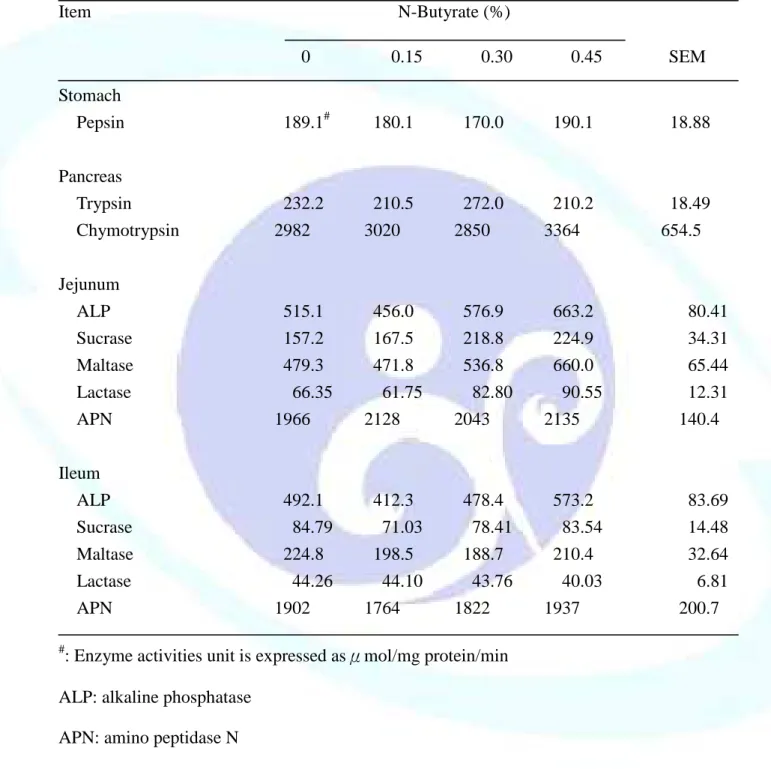
表 1-7. 飼料添加丁酸鹽對離乳仔豬胃黏膜、胰臟及小腸黏膜酵素活性 之影響--- 51
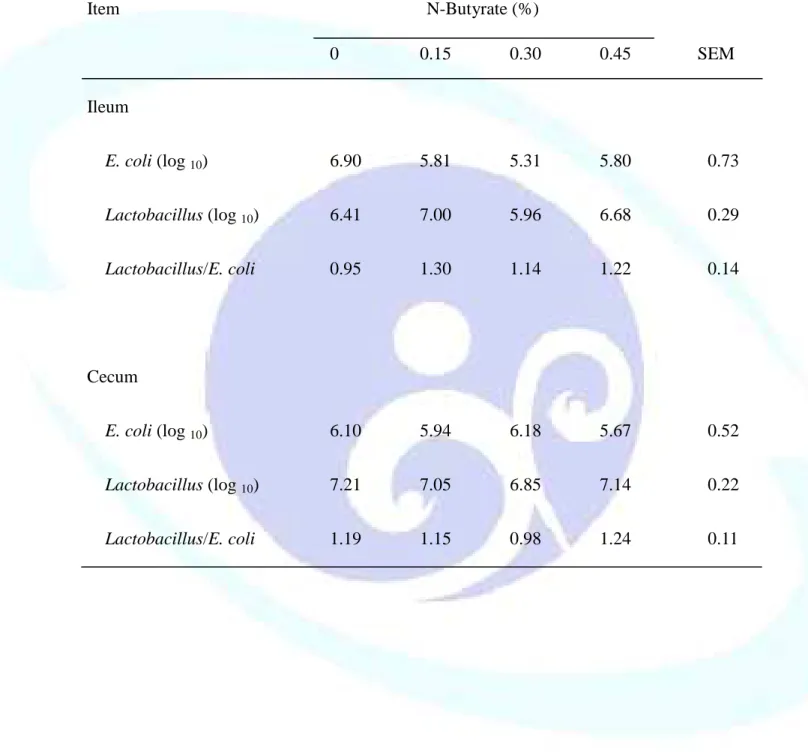
表 1-8. 飼料添加丁酸鹽對離乳仔豬腸道微生物菌相之影響--- 52
表 1-9. 飼料添加丁酸鹽對離乳仔豬血球數之影響--- 54
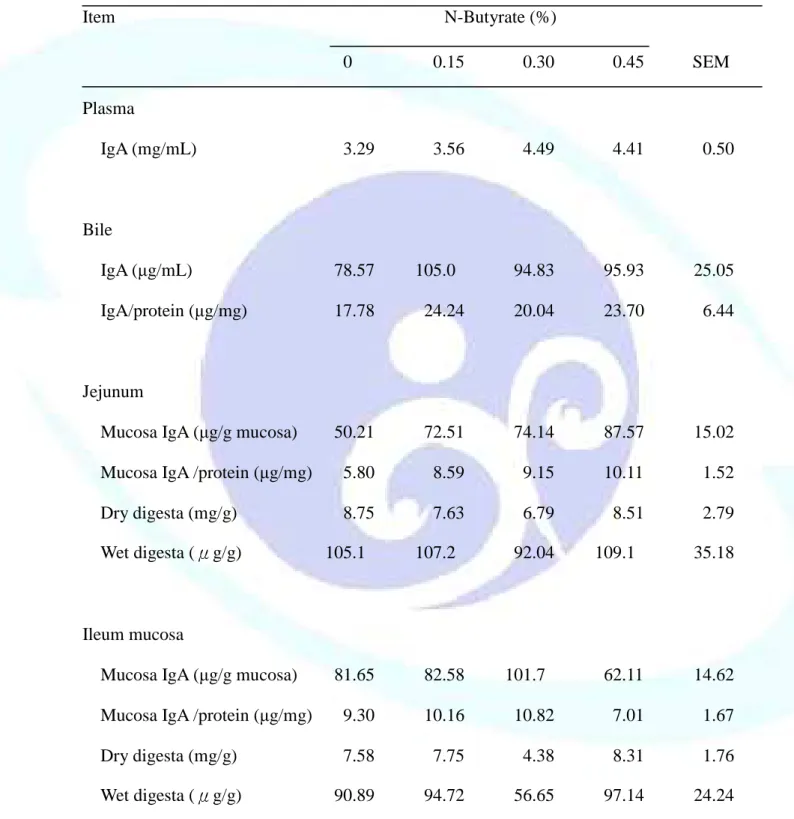
表 1-10. 飼料添加丁酸鹽對離乳仔豬血漿、膽汁、小腸黏膜及食糜 IgA 濃度之影響--- 55
表 1-11. 飼料添加丁酸鹽對離乳仔豬周邊血液與消化道相關淋巴組織淋 巴細胞增生反應之影響--- 56
表 1-12. 飼料添加丁酸鹽對離乳仔豬周邊血液與消化道相關淋巴組織淋 巴細胞 IFN-γ與 IL-10 濃度之影響--- 57 表 2-1. 基礎飼料配方和營養分組成--- 69 表 2-2. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬生長性能之影響--- 75 表 2-3. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬血漿胺基酸與血清尿素
態氮濃度之影響--- 77 表 2-4. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬小腸黏膜形態之影響--- 78 表 2-5. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬小腸中性杯狀細胞之影
響--- 79 表 2-6. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬小腸酸性杯狀細胞之影
響--- 80 表 2-7. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬小腸硫化杯狀細胞之影
響--- 82 表 2-8. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬小腸黏膜消化酵素活性
之影響--- 83 表 2-9. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬小腸食糜黏液蛋白濃度
之影響--- 84 表 2-10. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬血清免疫球蛋白濃度之
影響--- 85
表 2-11. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬膽汁、小腸黏膜及 IgA 濃度之影響--- 87 表 2-12. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬周邊血液與消化道相關
淋巴組織淋巴細胞增生反應之影響--- 88 表 2-13. 飼料添加丁酸鹽及羥丁胺酸對離乳仔豬周邊血液與消化道相關
淋巴組織淋巴細胞表面抗原之影響--- 89
圖次
頁次 圖 1-1. 飼料添加丁酸鹽對離乳仔豬腸繫膜淋巴結淋巴細胞 IFN-γ
濃度之影響--- 58
I、 、 、 、前言 前言 前言 前言
仔豬離乳時畜群、營養及環境的改變,會誘導劇烈的自我持恆反應而形 成緊迫,造成仔豬生長、腸道功能及免疫低下,而易提高疾病感染風險性 與死亡率。研究發現 28 天離乳仔豬飼養於環境衛生不佳之豬舍,會降低豬 隻生長性能與血漿羥丁胺酸濃度(Le Floch et al., 2006),顯示環境的變化會 改變仔豬體內營養分利用。新生仔豬於離乳前可藉由吸吮母豬乳汁獲得母 體抗體與養分進而提高保護力,然而離乳後仔豬食物來源由乳汁轉換成乾 性飼料,此時因腸道黏膜尚未發育完成,並易受飼料及微生物抗原刺激而 導致腸道受損。雖然延後仔豬離乳日齡可提高日增重與降低死亡率(Main et al., 2004),但早期離乳具有增加母豬生產潛能,若能供應促進離乳仔豬免疫 促進效果及腸道黏膜滋養性之物質,將可改善離乳仔豬生產效率。
丁酸屬於短鏈脂肪酸其為經由腸道微生物發酵飼料纖維的代謝產物,具 有降低飼料酸度、抑制微生物增生、且於吸收同時可伴隨促進鈉離子與水 分子之吸收,可直接供作腸細胞能量來源,對細胞也具有抑制組蛋白去乙 醯酶的效果,使染色體結構鬆散並提高基因轉錄作用,故能促進腸細胞分 化成熟(Smith et al., 1998),而有助於預防離乳仔豬下痢。因為腸腔中丁酸濃 度在仔豬離乳後降低(Mathew et al., 1996),因此,推論仔豬離乳時於飼料 中,額外添加丁酸應能改善離乳仔豬腸道健康狀態,但丁酸在常溫下為液 態不穩定,因而市面上常以丁酸鹽(Sodium butyrate, NaB)形式販售,便於乾
性飼料中混合添加。
羥丁胺酸(Threonine, Thr)是動物體之必需胺基酸之ㄧ,可防止肝臟脂肪 堆積與合成黏膜黏液蛋白。在一般以穀類為主之豬隻飼料原料中,Thr 常是 第二或第三限制胺基酸。依據 NRC (1998)之營養需要量標準顯示 5-10 kg 和 10-20 kg 仔豬總 Thr 需要量分別為 0.86 與 0.74%,而真可消化 Thr 需要 量分別為 0.74 與 0.63%。豬隻必需胺基酸在小腸組織部位的首渡代謝 (First-pass metabolism)約為 70% (Schaart et al., 2005),而生長中豬隻消化道 胺基酸耗用量大於總飼料胺基酸 50%以上,其中小腸 Thr 耗用率即佔總飼 料胺基酸之 60%,是所有胺基酸的最高者。腸道黏膜表面廣布由杯狀細胞 所分泌之黏液蛋白,具有隔絕腸腔與腸道組織之功能,黏液蛋白之胺基中 Thr 含量超過 40% (Carlstedt et al., 1993),由於豬隻飼料中添加抗營養消化 原料會增加黏液蛋白分泌,而且仔豬離乳期間供應超過生長所需之 Thr,可 以增加離乳仔豬之免疫反應(Wang et al., 2006a),但飼料添加超過生長所需 Thr 是否會影響腸道發育仍不明瞭。
綜合上述 NaB 與 Thr 之營養功能,NaB 具有促使腸細胞趨向分化以增 加腸細胞分泌消化酵素、IgA 與黏液蛋白等能力,而 Thr 為提供蛋白質結構 材料來源,故 NaB 是否會與 Thr 同時影響離乳仔豬腸道仍未明瞭。因此本 試驗設計二試驗,分別探討飼料添加不同劑量 NaB 及同時探討飼料添加 NaB 與 Thr 對離乳仔豬生長性能、腸道發育及免疫反應之影響。
II、 、 、 、文獻檢討 文獻檢討 文獻檢討 文獻檢討
一 一 一
一、 、 、 、仔豬腸道與免疫發育過程 仔豬腸道與免疫發育過程 仔豬腸道與免疫發育過程 仔豬腸道與免疫發育過程
(一
一一一)仔豬仔豬仔豬仔豬腸道黏膜與免疫組織之結構組成腸道黏膜與免疫組織之結構組成腸道黏膜與免疫組織之結構組成腸道黏膜與免疫組織之結構組成仔豬腸道不僅具有消化吸收之功能,也是一種天然的物理障壁,可防 止有毒化合物與病源從腸道黏膜進入血液循環系統。小腸組織係由單層上 皮細胞與固有層細胞所構成,而腸細胞依功能性可區分為(1)刷狀緣細胞 (Brush border cells):其具有分泌雙糖酶與胜肽酶之能力,位於黏膜表皮主 司消化與吸收之功能;(2)杯狀細胞(Goblet cells):位於黏膜表皮主司分泌黏 液,以保護腸道;(3)腸內分泌細胞:主司分泌腸性荷爾蒙與神經荷爾蒙,
以調節腸細胞生長與分化作用;(4)腸間隙淋巴細胞(Paneth cells):主司分泌 抗菌蛋白,屬於天生之免疫細胞。前三類腸細胞散佈於腸組織中,其由位 於腺窩基部之幹細胞,歷經數次細胞分裂後向絨毛頂端堆積,然後自絨毛 頂端經細胞凋亡而脫落至腸腔中,此過程週期約 3 - 5 天。腸道上皮細胞表 面具有多種受體,故可辨識腸道中不同抗原,其中類鐸受體(Toll-like receptor, TLR)可分為病原或共生相關分子形式,具有辨認抗原與免疫調節功能,並 可活化訊號傳導路徑(NFκB 與 MAPK)以啟動基因轉錄(Stokes et al., 2004)。經抗原刺激後會分泌細胞激素等物質,當上皮內淋巴細胞、固有層 細胞及固有層淋巴細胞接受訊息後,得以調節淋巴細胞之分化並啟動腸道 免疫反應。
豬隻免疫系統可區分為細胞性免疫系統(Cellular immune system)與體 液性免疫系統(Humoral immune system)二大類,細胞性免疫系統之免疫功能 是由淋巴細胞所提供,而體液免疫系統之免疫功能則是由血液與淋巴液中 之抗體所提供。此二大類系統相互配合下可順利活化免疫反應,其主要步 驟包括:抗原抓取、抗原呈現、激活抗原敏感細胞及激起細胞媒介性免疫 反應或體液免疫反應(產生抗體與維持細胞對特定抗原之記憶性) (Tizard, 1992)。腸道免疫組織可分為:組織性淋巴組織(主要為派亞氏與腸繫膜淋巴 結)與散漫性淋巴組織(散佈在黏膜層中之淋巴細胞) (Stokes et al., 2004)。一 般腸組織分為絨毛部份依序可分為上皮層與固有層,在成熟的腸絨毛上皮 層具有大量 CD8+ T 淋巴細胞,而固有層則具有 CD4+ T 淋巴細胞以及存在 於腺窩中具有分泌性的漿細胞。表 1 為不同年齡仔豬黏膜免疫系統發育狀 態,仔豬出生後 2 週黏膜免疫淋巴細胞尚未分化成熟,直至第 2-6 週間才出 現分化完成之淋巴細胞與抗體分泌性細胞,因此,仔豬黏膜免疫系統至少 需 2 個月左右才漸趨發育成熟。
病原微生物如果黏附至上皮細胞表面,就有機會穿過上皮層而導致疾 病之發生(Stokes et al., 2004)。而腸道具有不同免疫組織,可啟動免疫活化 反應,以對抗病原與啟動耐受性,並可適應共生微生物與食物之無害抗原,
但此 2 種功能皆需消耗養分以啟動反應(Bailey et al., 2005),若腸道與免疫 組織對有害病原形成不適當的作用反應,將會造成過敏反應或提高疾病感
表 1. 仔豬黏膜免疫系統發育階段
Table 1. Stages of the mucosal immune system development of piglets
階段一 新生仔豬 少量抗原呈現細胞、幾乎沒有 T 淋巴細胞及沒有漿 細胞
階段二 1 天-2 週齡 1.上皮與固有層中出現不正常 CD2+CD4-CD8-與 CD2+CD4-CD8αα T 淋巴細胞
2.固有層中出現一些活化態 CD4+ T 淋巴細胞
3. MHCII+細胞出現於固有層中
階段三 2-4 週齡 1.成熟記憶性 CD4+ T 淋巴細胞出現於固有層中
2. IgM+ B 淋巴細胞出現於腺窩區域
階段四 4-6 週齡 1.上皮與固有層中出現記憶性 CD8+ T 淋巴細胞
2. IgA+ B 淋巴細胞出現於腺窩區域
(Bailey and Haverson, 2006)
受性。因此,發育良好之腸道首要需要腸道黏膜免疫系統之平衡發育,其 中牽涉腸細胞與黏膜內淋巴細胞間之交互作用與成熟度,次而則仰賴腸道 共生微生物與有益微生物的定殖。然而腸細胞、淋巴細胞及微生物間交互 關係複雜,故在飼料中補充營養成分,以期待能夠促進仔豬腸道發育與免 疫反應。
(二
二二二)仔豬仔豬仔豬仔豬出生前後出生前後出生前後出生前後之之之之腸道與免疫腸道與免疫腸道與免疫腸道與免疫系統系統系統系統發育發育發育發育懷孕母豬之生理狀態與胎兒發育息息相關,任何品系的母豬飼養於溼 熱環境下,皆會因抑制飼料採食而降低體重,並減輕仔豬之出生重量 (Gourdine et al., 2006),且初產之母豬在限食狀態下不僅會影響黃體激素
(Luteinizing Hormone, LH) , 也 會 影 響 促 濾 泡 激 素 (Follicle stimulating hormone, FSH)之分泌量,而影響泌乳期間濾泡發育狀態(Kauffold et al., 2008)。另外,限制懷孕母豬蛋白質攝取(0.7%蛋白質),也會影響懷孕中期 (63 天)胎兒體重、器官重、器官蛋白質、RNA 及 DNA 濃度(Pond et al., 1991),
因為此時期是胎兒發育最快速時期,故母豬蛋白質攝取劇烈影響仔豬發 育。荷爾蒙分泌量降低會影響懷孕母豬子宮內生長限制作用(Intrauterine growth restriction)而導致胞內訊號傳導不足、氧化還原不平衡、抑制蛋白質 合成作用及提高蛋白質降解作用,最終導致新生仔豬營養代謝不正常而抑 制其生長發育(Wang et al., 2008),因此,母豬飼養環境條件不佳而降低飼料 採食量與荷爾蒙分泌量,最後抑制子宮、濾泡及胎兒發育。
泌乳期母豬攝取能量不足,會有降低乳量之現象,此時若短暫給予高 脂飼料,雖可提供高能量以維持母豬體重,但卻不易維持母豬體重,而且 會增加血液中代謝物含量(非酯化脂肪酸、β-羥基丁酸及尿素)、降低胰島 素及類胰島素生長因子-1(Insulin-like growth factor-1)之分泌量而間接影響 黃體激素與雌素二醇之分泌量與濾泡發育(van den Brand and Kemp, 2006)。
因此,懷孕母豬飼料營養成分與能量攝取量,會調控母豬內泌素而影響母 豬子宮與胎兒發育,甚至也會影響爾後之濾泡發育與生產性能。因此,於 母豬懷孕至泌乳期時需控制採食量與荷爾蒙分泌量,以預防母豬生產性能 降低進而影響胎兒發育。
仔豬出生至離乳階段之消化道發育,依賴腸腔性(Luminal)與系統性 (Systemic)因子之調控。其中腸腔性因子包括腸道內之皮質醇與類升糖素胜 肽-2 等荷爾蒙調節因子之共同影響,如皮質醇會影響胃酸與胃泌素之分泌 及消化道水解酶活性(胃蛋白酶、乳糖酶及胺基胜肽酶);而糖皮質激素影響 消化道形態、細胞學及功能性分化;系統性因子如出生後攝取母乳中之生 長因子、荷爾蒙及養分而影響消化道發育。因此腸腔性與系統性因子,分 別調控仔豬出生前後消化道之發育。文獻指出,仔豬出生時之小腸黏膜,
即具有乳糖酶與麥芽糖酶,但蔗糖酶則是在仔豬出生後第 1 週出現,同時 也是乳糖酶分泌高峰期,隨後開始降低,緊接著第 2 週為蔗糖酶與麥芽糖 酶的分泌高峰期,隨後開始降低(Aumaitre and Corring, 1978),由上述可知 仔豬正由依賴非消化道性營養來源之胚胎轉換成依賴消化道性營養來源 (Trahair and Sangild, 1997),這也說明新生仔豬消化道正處於適應期。
仔豬出生後因先天免疫系統尚未發育完全,因此可藉由母豬乳汁中免 疫球蛋白所提供之被動免疫保護,並可與消化道上皮細胞及黏液構築一個 保護功能完整之腸道黏膜障壁。雖然母豬初乳中免疫球蛋白濃度高,但分 娩 2 天後乳汁免疫球蛋白抗體含量開始急速下降,故新生仔豬被動免疫保 護能力會隨日齡之增長而減弱。
雖然新生仔豬被動免疫保護能力隨年齡增長而減弱,但新生仔豬先天 免疫系統,會受環境與飼料抗原刺激而逐步發育。文獻顯示,新生仔豬腸
道 CD2+ T 細胞數與單核細胞/顆粒細胞比雖少,但介白素-2R (Interleukin-2R, IL-2R) 細胞數量眾多(Vega-López et al., 1995),且 14 日齡仔豬仍具有來自 於母豬乳汁的移行抗體,而能維持穩定的 IgG 濃度(Niekamp et al., 2007),
顯示此時仔豬仍受到被動免疫反應之保護。此外,仔豬出生後一週腸道固 有層即出現免疫球蛋白生產細胞(Bourne, 1976),且 CD4+細胞數量急遽增 加,但 CD8+細胞數量直至 5-7 週時仍然不足(Vega-López et al., 1995),由此 可知仔豬出生後約一個月其非特異性免疫能力正逐漸成熟。
雖然總淋巴細胞數目與增生反,應會隨仔豬年齡增加而提高(Blecha et al., 1983; McCauley and Hartmann, 1984),但嗜中性球數目、總 IgG 濃度及 經裂質原刺激之 B 細胞增生反應,則會隨仔豬年齡增加而下降(Blecha et al., 1983; McCauley and Hartmann, 1984; Hoskinson et al., 1990; Niekamp et al., 2007)。因此,仔豬出生後之健康,雖逐漸受到非特異免疫之保護,但主要 仍是藉由細胞性免疫保護能力持續發揮其影響力。
(三
三三三)仔豬仔豬仔豬仔豬離乳離乳離乳離乳階段階段階段階段之腸道與免疫之腸道與免疫之腸道與免疫之腸道與免疫系統系統系統系統發育發育發育發育仔豬腸道與免疫組織發育完成前會經歷離乳階段,此時因飼料轉換造 成仔豬採食下降而抑制生長,至今已證實離乳的確會改變仔豬腸道黏膜形 態、消化道水解酵素活性與分泌能力及免疫反應。其中無論仔豬是否經過 離乳過程皆會依年齡之增長而增加腺窩深度,但離乳會更加深腺窩深度
(Hampson, 1986a; Hampson, 1986b)。且離乳後因食物之轉換造成仔豬食慾與 採食量降低,導致消化酵素劇烈下降,強烈影響仔豬腸道黏膜形態(Gu et al., 2002)與降低絨毛高度/腺窩深度比,而提高下痢風險(Pluske et al., 1996);另 外,三週齡仔豬在離乳後 2-3 週之腸道黏膜形態仍未恢復至離乳前水準 (Brown et al., 2006a)。 但也有文獻顯示,離乳後 3 天腸道絨毛寬度增加,
但腺窩深度直到 25 天後仍未恢復至離乳前程度,而離乳後第 11 天可得到 較高之絨毛高度/腺窩深度比值(Brown et al., 2006b),雖未影響絨毛高度,但 卻抑制腺窩細胞之增生能力。上述離乳過程對仔豬腸道黏膜形態之變化,
推測可能是仔豬離乳早期腸道正處於食物來源與飼養環境改變之適應期而 造成腸道發育變化。
離乳過程對仔豬消化道功能之影響,除脂解酶外,其他胰臟酵素活性 與基因表現皆在短期內恢復至離乳前水準,且仔豬離乳後 3 天,胰臟外分 泌功能似乎已能適應乾性飼料(Marion et al., 2003)。此外,離乳早期胰液中 雖具有較高含量之蛋白質、胰蛋白酶、解脂酶及澱粉酶,但解脂酶活性卻 處於不活化狀態(Huguet et al., 2006)。離乳過程並不會直接影響胰臟分泌消 化酵素能力,但四週齡仔豬食糜胃蛋白酶活性離乳後卻會迅速下降,離乳 後第 5 天恢復至離乳前程度,而小腸食糜胰蛋白酶、羧基胜肽酶 A 、澱粉 酶及解脂酶在離乳後 2-7 天皆處於活性低下狀態,以及離乳後第 7 天胰蛋白 酶活性恢復至離乳前程度,但其他酵素仍未達到離乳前程度,胰凝乳蛋白
酶、羧基胜肽酶 B 及羧基脂解酶活性至離乳後 9 天仍未恢復(Hedemann and Jensen, 2004),因此離乳會抑制仔豬小腸中胰臟消化酵素活性及功能特性。
另外,仔豬三週齡離乳會降低腸道乳糖酶活性,而蔗糖酶活性亦會急速下 降,其離乳後 4-5 天為活性最低時期,之後可回復到離乳前程度(Hampson and Kidder, 1986)。上述結果顯示離乳對仔豬胃腸道消化功能影響甚巨,尤 其是腸道杯狀細胞也扮演著一個保護腸道的角色(Gu et al., 2002),研究發 現,三週齡離乳仔豬在離乳 3 天後,腸道中性與酸性杯狀細胞數目顯著降 低,於 2 週後才會逐漸增加腸道絨毛杯狀細胞數目(Dunsford et al., 1991)。
另外之試驗亦發現,三週齡離乳仔豬離乳後 3 天十二指腸與空腸中性杯狀 細胞數會降低,離乳後 25 天則發現會降低迴腸中性杯狀細胞數(Brown et al., 2006b),此外也發現離乳後 3 週十二指腸硫化杯狀細胞數顯著較離乳後 1 週增加(Brown et al., 2006a)。由上述結果顯示,雖然杯狀細胞具有分泌黏液 之功能以保護腸道,但也因離乳過程使其數量降低。
腸道為仔豬養分吸收與代謝的主要器官,並具有物理障壁特性以抵抗 外來抗原(Eckmann et al., 1995; Pitman and Blumberg, 2000),故腸道發育狀 態,會影響仔豬生長性能與腸道物理防禦能力。離乳階段同時也是仔豬生 長發育最快速時期,新生仔豬之免疫系統,易受離乳影響而抑制免疫能力 發育。此外,離乳後因飼料由乳汁轉換成乾性飼料,而產生飼料抗原誘發 腸道免疫系統短暫性過敏反應,導致腸細胞轉化與絨毛萎縮,並增加仔豬
對疾病的感受性(Stoke et al., 1987)。
研究顯示,三週齡離乳仔豬腸道絨毛、腺窩 CD2+ T 細胞數及巨噬細胞 /顆粒細胞數量比顯著提高(Vega-López et al., 1995),而離乳後 2 天小腸組織 便會出現 CD4+與 CD8+ T 淋巴細胞(McCracken et al., 1999),其反映出仔豬 正處於先天性免疫反應發育期(Niekamp et al., 2007)。此外,研究顯示腸道 黏膜也可以產生某些重要的細胞激素與趨化激素,以活化嗜中性球、巨噬 細胞、T 細胞、B 細胞及樹突細胞,提高腸道黏膜中淋巴細胞數量,以增加 先天與後天免疫反應之能力,因此腸道黏膜就像是免疫系統中的”看門狗”
(Eckmann et al., 1995; Jiang et al., 2000)。腸道黏膜淋巴細胞也具調節上皮更 新、生長及分化的重要功能(Matsumoto et al., 1999),且上皮內淋巴細胞與 杯狀細胞數量多寡,可反應出上皮形態與微生物狀況(Deplancke and Gaskins, 2001; McCracken and Lorenz, 2001),可能是腸道上皮內淋巴細胞,經飼料抗 原、養分及微生物之共同刺激並分泌細胞激素,藉細胞激素相關訊息傳導 物質與固有層細胞或黏膜中淋巴細胞交換訊息,而調節腸道黏膜中淋巴細 胞之分布(Stokes et al., 2004)。三週齡離乳仔豬周邊血液 T 淋巴細胞、CD2+ T 細胞與γ/δ T 淋巴細胞接受器百分比,隨年齡增加而增加,且離乳後 2 週 可提高周邊血液單核細胞,經植物血凝素刺激之增生指數,但卻會降低 CD4+ T 淋巴細胞百分比而提高 CD4/CD8 淋巴細胞百分比(Brown et al., 2006a),
周邊血液單核細胞數目可隨離乳年齡增加,且離乳後第 2 週血液 T 淋巴細
胞對抗原具有較高的反應能力,但離乳後 11 或 25 天卻會降低空腸與迴腸 CD2+ T 淋巴細胞數,且離乳後 3-11 天會降低空腸 MHC II+數目(Brown et al., 2006b)。離乳後 3 週仔豬腸道免疫能力尚未完全恢復,雖然離乳過程會抑制 仔豬腸道免疫發育,但此時周邊已具有某些免疫保護能力,因此推估仔豬 出生約 2 個月或三週齡仔豬離乳後 3 週以上,其腸道免疫系統才會逐漸發 育完全。
二 二 二
二、 、 、 、影響仔豬 影響仔豬 影響仔豬 影響仔豬離乳期間 離乳期間 離乳期間 離乳期間腸道與免疫發育之 腸道與免疫發育之 腸道與免疫發育之 腸道與免疫發育之飼養 飼養 飼養因素 飼養 因素 因素 因素
腸道是一種敏感且複雜的器官,除具有消化吸收功能外並具有免疫組 織,以預防動物疾病發生,仔豬腸道約在 2 月齡左右才漸趨發育完全,期 間仔豬會面臨離乳過程,且有多種因素會影響仔豬腸道生理狀態與免疫反 應,這些因素包括:飼養與營養因素,其中飼養因素包括仔豬離乳年齡、
飼料型態及飼養環境。
(一
一一一)離乳離乳離乳離乳年齡年齡年齡年齡仔豬不同日齡(17、21、28 及 35 日齡)離乳對小腸黏膜發育也具有不同 之影響,21 與 35 日齡離乳之仔豬具有較高之小腸絨毛高度與腺窩深度,但 17 日齡離乳之仔豬小腸絨毛高度與腺窩深度卻是最低,28 日齡離乳仔豬具 有較多之絨毛上皮內淋巴細胞,21 日齡離乳仔豬絨毛上皮內淋巴細胞含量
卻是最少,杯狀細胞密度會隨離乳日齡增加而降低(Gu et al., 2002),另外 7 日齡仔豬在離乳後 3 天雖可提高小腸蔗糖酶、胜肽酶及麥芽糖酶活性以及 麥芽糖酶 mRNA 表現量,但卻會降低乳糖酶活性與絨毛高度(Marion et al., 2005),而仔豬於 21、28 及 35 日齡離乳後 7 天分別會降低十二指腸乳鐵蛋 白 mRNA 表現 77%、53%及 59%,且只有在 28 天離乳後第 2 週十二指腸乳 鐵蛋白 mRNA 表現量會顯著降低(Wang et al., 2006b)。上述結果綜合顯示,
離乳會劇烈影響小腸生理狀態,但 21 日齡離乳可減輕腸道受損。
(二
二二二)飼料飼料飼料飼料型型型型態態態態離乳後飼料形式的轉換會造成仔豬食慾降低,但於乳汁與固體飼料中 添加風味劑不會影響仔豬離乳後 7 小時內採食量,而哺乳與固體飼料在不 添加風味劑情況下交替餵食,哺乳會降低固體飼料採食量(Millet et al., 2008),且研究發現仔豬離乳後 2 天內,幾乎未有採食現象,直至第 7 天才 有明顯進食行為,因此導致腸道絨毛高度與腺窩深度降低,卻也發現黏膜 CD4+與 CD8+ T 淋巴細胞逐漸增加,以保護腸道與預防疾病感染(McCracken et al., 1999),給予新生仔豬教槽料可降低一週齡至五週齡之飼料採食量,但 在離乳後前 4 週時會具有較高之體重(Kuller et al., 2007),離乳後因飼料轉 換成固體形式,而降低採食與改變腸道黏膜形態,但仔豬經過約一週之腸 道適應期而,攝取到均衡營養分並改善生長性能。此外,飼料物理特性會
影響豬隻消化道微生物菌相之組成,餵食粗糙非粒狀飼料會增加胃食糜乾 物質含量、厭氧菌數及有機酸濃度,並可抑制體外沙門氏菌增生速率,此 外,小腸後段、盲腸及中結腸也具有較低量之大腸菌數(Mikkelsen et al., 2004),飼料纖維的粗細也會影響腸道微生物菌相的發展,飼料纖維較粗之 飼料可提高盲腸丁酸濃度,並降低小腸後段、盲腸及中結腸大腸菌數 (Mikkelsen et al., 2004),由此可知飼料纖維含量高可促進腸道有益微生物之 發展,這可能也與腸道黏膜發育狀態有關。
(三
三三三)飼養環境飼養環境飼養環境飼養環境離乳仔豬飼養於獨立空間會提高飲水頻率,且需誘導採食並導致體重 低於群體飼養之離乳仔豬,因此,群體飼養環境下可刺激離乳仔豬採食與 生長性能(Sorrells et al., 2006);光照時間長短則會影響離乳仔豬生長與免疫 反應,光照時間短(8 小時/每天)可提升 2-4 週齡離乳仔豬之免疫反應,卻會 抑制生長性能(Niekamp et al., 2007)。另外,四週齡離乳仔豬飼養於環境衛 生不佳之豬舍會降低日增重、飼料利用效率及血漿羥丁胺酸濃度,但卻會 提高血漿離胺酸濃度(Le Floch et al., 2006)。且不同飼養條件(商業一貫肉豬 場與早期離乳隔離豬舍)也會影響三週齡離乳仔豬腸道發育與免疫反應,商 業一貫肉豬場會提高離乳後第 1 天空腸硫化杯狀細胞數和 3-25 天迴腸酸性 與硫化杯狀細胞數,也會提高離乳後第 11 天空腸 CD8+與 CD25+ T 淋巴細
胞,但卻會降低三週齡離乳仔豬空腸絨毛高度/腺窩深度比值、離乳後第 3 天空腸 CD8+與 CD25+ T 淋巴細胞數及第 25 天空腸硫化杯狀細胞數(Brown et al., 2006b)。另外,社會地位緊迫也會增加早期離乳仔豬對沙門氏菌的感 受性與糞便中的濃度(Callaway et al., 2006)。群飼與延長光照時間可提高採 食量而有利於生長性能,但卻提高社會地位緊迫與疾病感受性,且飼養環 境衛生條件不佳也會降低腸黏膜保護作用與活化免疫反應。
綜合上述節仔豬離乳年齡、抗原、飼料形態及飼養環境會增加仔豬離 乳時的緊迫,假使能在此時期飼料中添加某些營養成分,以減少離乳緊迫,
應能改善離乳仔豬腸道黏膜結構、功能及免疫反應,提高其生長性能與健 康狀態。
三 三 三
三、 、 、 、飼料抗原與 飼料抗原與 飼料抗原與 飼料抗原與營養成分對離乳仔豬腸道與免疫之影響 營養成分對離乳仔豬腸道與免疫之影響 營養成分對離乳仔豬腸道與免疫之影響 營養成分對離乳仔豬腸道與免疫之影響
飼料營養成分,舉凡蛋白質、脂肪、維生素及礦物質等,皆為動物生長 與維持正常機能所必需,但特殊脂肪酸與胺基酸,除供應體內一般體組成 之目的外,尚具有提高動物免疫之生理功能,諸如丁酸、羥丁胺酸及精胺 酸等。飼料組成除營養因子外尚包含飼料抗原,其亦可能干擾豬隻腸道發 育與免疫能力亦一併討論。
(一
一一一)飼料抗原飼料抗原飼料抗原飼料抗原仔豬暴露於抗原下可分成二階段,第一階段:出生後仔豬腸道初次接 觸自然環境中之大量抗原,其包括母乳與環境微生物,直至離乳前仔豬腸 道已具有大量之共生微生物,此時腸道黏膜與免疫仍處於發育階段,因此 易受抗原刺激影響仔豬生長。第二階段:仔豬離乳後面臨飼養環境、食物 及社會地位之改變,不僅提高仔豬緊迫與降低腸道黏膜障壁功能,使腸道 黏膜容易受外來抗原之刺激(Bailey et al., 2005)。
仔豬離乳後育成期飼料常使用大豆原料,對三週齡離乳仔豬易造成生長 遲緩與下痢之情況(Armstrong and Cline, 1977),而其原因為大豆製品含有 Glycinin 與 β -conglycinin 等 抗 原 , 會 引 起 仔 豬 區 域 性 過 敏 (Local hypersensitivity response)與系統性免疫反應(Systemic immune response) (Li et al., 1990)。區域性過敏反應會使小腸絨毛受損與萎縮而影響養分之吸收 (Cera et al., 1988; Hankins et al., 1992),且三週齡離乳仔豬以大豆原料之飼料 餵食 4 天後,其近端小腸絨毛與腺窩中具有較多之 CD2+淋巴細胞與單核細 胞/顆粒細胞比(Vega-Lopez et al., 1995),或者經大豆蛋白抗原刺激會提高 13 倍十二指腸黏膜嗜酸性細胞密度(Dréau et al., 1994),除飼料抗原外,自然環 境中也具有多種刺激性抗原(腸道共生及病原微生物抗原),正常情況下腸道 黏膜會對抗原產生適當反應,但腸道黏膜免疫系統仍會對這些抗原產生短 暫性過敏反應或提高腸細胞轉化作用,而導致絨毛萎縮並影響仔豬腸道發
育(Stokes et al., 1987)。
因此,在飼料中添加促進腸道之營養成分,應可減緩飼料抗原對離乳仔 豬腸道影響程度、縮短腸道過敏反應時間及免疫活化程度。
(二
二二二)功能性脂肪酸功能性脂肪酸功能性脂肪酸功能性脂肪酸脂肪由不同形式之脂肪酸組成,因其物理、化學及生理特性之差異而 對動物而言是一種重要的營養因子。一般在自然環境下,脂肪可溶於有機 溶劑之中,卻不溶於水,並受脂肪酸長度與雙鍵數目而影響吸收效率。自 然環境具有多種脂肪酸對仔豬也具有不同之作用:
1.多元不飽和脂肪酸(Polyunsaturated fatty acid)
穀物與植物油脂中含有 Omega-3 (ω-3)與 Omega-6 (ω-6)之多元不飽和 脂肪酸,其在生理與營養上皆具重要角色,但哺乳類動物無法自體合成,
故為哺乳類動物必需脂肪酸。α-亞麻油酸(α-linolenic acid, ALA, 18:3 n-3) 與亞麻油酸(Linoleic acid, LA, 18:2 n-6),分別為 ω-3 與 ω-6 之代表性脂肪 酸,其在動物體內可分別轉化成不同長度之脂肪酸,例如:ALA 在動物體 內可轉化成二十碳五烯酸(Eicosapentaenoic acid, EPA, 20:5 n-3)與二十二碳 六烯酸(Docosahexaenoic acid, DHA, 22:6 n-3),而 LA 進入體內則可轉化成 花生四烯酸(Arachidonic acid, ArA, 20:4 n-6) (Kim et al., 2007)。研究顯示,
穀物油脂含有高比例 ω-6 與低比例 ω-3,如果在母豬飼料中添加高量 ω-6 則 對仔豬之存活率與免疫功能具負面影響。而母豬攝取含有多元不飽和脂肪 酸之飼料,雖可增加多元不飽和脂肪酸進入胎兒體內並提高其利用率,但 不影響胎兒體重與對疾病之感受性,但提高母豬飼料中 ω-3 濃度卻可提高 約 0.6 頭窩仔數並改善發炎狀態以提高母豬生產性能,這可能與 ArA 與 EPA 在發炎與免疫反應中之角色有關,由於 ω-3 會抑制促發炎細胞激素之分泌 且 EPA 為 ω-3 之ㄧ(Grimble, 2005),故 EPA 會競爭抑制 ArA 轉化成二十碳 烯酸而達到抗發炎之效果。綜合上述文獻,多元不飽和脂肪酸具有改善母 豬生產性能、母豬及仔豬免疫能力之功能。
2.中鏈脂肪酸(Medium-chain fatty acid, MCFA)
MCFA 對於新生仔豬是一種穩定的能量來源,於仔豬出生後一天內即可 分 泌 胰 臟 脂 解 酶 而 分 解 MCFA 並 形 成 中 鏈 三 酸 甘 油 脂 (Medium-chain triacylglycerol, MCT)進而改善 MCT 之利用效率(Odle, 1997)。因肝細胞對 MCT 具有較高之氧化代謝效率,故 MCFA 經小腸吸收後直接進入肝臟進行 氧化代謝,藉此 MCFA 因其代謝特性而不具有提高體脂肪之功能(Odle, 1997;
St-Onge and Jones, 2002)。
3.有機酸(Organic acid)與短鏈脂肪酸(Short-chain fatty acid, SCFA)
有機酸可降低胃酸鹼度而維持消化道微生物菌相平衡,並促進食糜蛋 白質分解作用,但三週齡離乳仔豬經餵食不同比例之有機酸 7 天後,未能 改變總消化道消化率,而餵食含有甲酸/延胡索酸之飼料後,會提高胃酸鹼 度、丙酸鹽及丁酸鹽百分比,卻不會影響微生物菌相百分比,而飼料添加 甲酸/乳酸混合酸雖未能改變小腸短鏈脂肪酸百分比,卻能降低大腸桿菌屬 微生物百分比(Franco et al., 2005),飼料共同添加微生物植酸酶與有機酸,
可 改 善 保 育 期 仔 豬 飼 料 有 效 磷 之 利 用 率 , 而 共 同 抑 制 有 效 磷 之 流 失 (Omogbenigum et al., 2003),飼料添加微膠囊化有機酸會降低結腸短鏈脂肪 酸比例、空腸及盲腸大腸桿菌屬細菌數目(Piva et al., 2007),飼料添加有機 酸可維持豬隻體內磷含量,並降低消化道短鏈脂肪酸百分比與腸道大腸桿 菌屬細菌數目,故有機酸可提高腸道有益菌/有害菌百分比以促進腸道健康。
短鏈脂肪酸(Short-chain fatty acid, SCFA)為含 1-6 個碳之有機脂肪酸,經 腸道微生物利用飼料碳水化合物(多醣、寡醣及醣蛋白)進行發酵所產生之代 謝產物。主要為醋酸、丙酸及丁酸所組成,其在腸道中之莫爾比率約為 60:
20: 20 或 3: 1: 1,其比率會依發酵位置、腸道微生物組成、飼料形態、養分 組成及纖維含量差異而不同。大腸黏膜細胞可快速吸收 SCFA,其吸收主要 機制為質子化 SCFA 簡單擴散與陰離子交換蛋白(Cook and Sellin, 1998),僅 有 5-10% SCFA 會殘留於糞便中而排出體外,而 SCFA 排出過程與鈉離子吸
收和重碳酸離子分泌有關。 此外,SCFA 也可提高腸細胞中葡萄糖運送蛋 白與刷狀緣鈉/葡萄糖共同運送蛋白 mRNA 含量,並直接參與腸道適應反應 與刺激腸內分泌 L 細胞分泌類升糖素胜肽-2 (Glucagon-like peptide-2, GLP-2),SCFA 中之丁酸除具有提高血漿 GLP-2 含量之功能外,也具有腸 黏膜滋養效果(Tappenden et al., 2003)。
雖說 SCFA 可由大腸黏膜細胞快速吸收,但不同 SCFA 其代謝作用位置 卻不盡相同,周邊肌肉組織可氧化醋酸殘基產生能量,肝臟可代謝少許丁 酸與丙酸作為糖質新生之受質,並吸收 50-70%醋酸,腸上皮細胞主要利用 丁酸作為能量產生受質(Hijova and Chmelarova, 2007),由上述可知丁酸不僅 可提供腸細胞作為能量來源(Roediger, 1980),並可促進鈉離子與水分子之吸 收、抑制組蛋白去乙醯酶活性而促進特異基因轉錄(Mathew et al., 1996;
Jobin and Sartor, 2000),故推測若在飼料中添加丁酸應可改善仔豬離乳後腸 道發育。
總結上述各種脂肪酸因其物理、化學及生理特性,對動物而言皆是重 要的營養因子,但由於消化道消化吸收程度不一,且因其在體內代謝位置 不同而對仔豬生理具有不同影響,推測腸細胞能有效代謝利用結構簡單之 丁酸,因而較能有效改善仔豬離乳時之腸道發育。
4.丁酸(Butyric acid) (a)丁酸鹽之化學特性
丁酸(Butyric acid)是 4 個碳的短鏈脂肪酸,是大腸表皮細胞主要能量來
源(Roediger, 1980)。丁酸可提高動物飽足感、強化黏膜障壁防禦功能、抑制 大腸癌症生成、發炎及氧化壓力(Hamer et al., 2008),此外也具有組蛋白去 乙醯酶抑制劑的效果,以基因外調節功能而高度乙醯化染色體組蛋白,使 染色體結構鬆散,並提高轉錄因子對核糖體 DNA 之感受性,而加強基因轉 錄作用,故能調節腸細胞增生或分化作用(Smith et al., 1998)。另外,也發現 丁酸可抑制核因子 kappa B (Nuclear factor kappa B, NFκB)活化,而 NFκB 具有調控促發炎細胞激素、趨化素及發炎誘導酵素(Jobin and Sartor, 2000)。
但丁酸在常溫下為液態不穩定,故一般以 NaB 形式便於使用,NaB 攝取進 入消化道中,可被解離形成丁酸與金屬鹽類離子,而具有滋養黏膜與降低 腸道酸鹼值之特性,故推測 NaB 具有滋養腸細胞與促進細胞分化成熟之功 能。
(b)丁酸之吸收代謝途徑與生產
約有 85-90%丁酸為哺乳動物消化道微生物利用碳水化合物進行發酵所 產生,其具有降低飼料酸鹼值及抑菌功能(Knudsen et al., 2003; Fay and Farias, 1975)。丁酸可透過簡單擴散與陰離子交換蛋白之作用,直接進入腸
細胞達到快速吸收的功效(Wächtershäuser and Stein, 2000),同時也促進鈉離 子與水分之吸收,減少腸道中水分含量而有助於預防離乳仔豬下痢,但其 量在仔豬離乳後降低(Mathew et al., 1996)。研究顯示,消化道上皮細胞所吸 收之 80%丁酸先經腸細胞β-氧化代謝轉化成酮體而進入血液(Roediger, 1980; Bugaut, 1987; Hijova and Chmelarova, 2007),但仍有部分丁酸會進入血 液當中。腸上皮細胞主要利用丁酸作為能量來源,肝臟可代謝少許丁酸與 丙 酸 作 為 糖 質 新 生 之 受 質 並 吸 收 50-70%醋 酸 (Hijova and Chmelarova, 2007)。
動物本身能藉由外源非澱粉類多醣(抗性澱粉、非消化性寡醣及醣醇) 與內源物質(醣蛋白、剝落之上皮細胞及胰臟分泌物)經大腸微生物發酵產生 丁酸(Knudsen et al., 2003);此外,給予豬隻馬鈴薯飼料可得到最高比例之 丁酸,而同種穀物飼料原料如具有高纖維含量,也可提高總短鏈脂肪酸與 丁酸之比例(Knudsen et al., 2003)。因此,腸道與血液中丁酸含量會隨纖維 攝取量高低而改變,無論如何,腸道中丁酸濃度對細胞適當營養效果並不 能相互做為對照。
豬隻餵食 45 g 大麥粉中之非澱粉類多醣後 7 小時,可在門脈血液中監 測到約 50 μM NaB,但豬隻餵食 116 g 裸麥麵包之非澱粉類多醣體後 7 小 時卻只測得約 40 μM NaB,然而在第 12 小時則可測得約有 60 μM NaB (Knudsen et al., 2003),由此可知不同穀物會影響血液 NaB 濃度,且高量非
澱粉類多醣能進一步延長腸道微生物發酵時間,並可維持血液 NaB 濃度。
此外,當小鼠口服不同比例之三丁酸甘油酯(Tributyrin, TB; 3.1、5.2、7.8 及 10.3 g/kg 小鼠體重)後發現,口服 5 分鐘後即可在血漿中測得 NaB,而 1 小 時內血漿中 NaB 濃度分別為 0.5、0.9、1 及 1.75 mM NaB 濃度,意外的是 口服 10.3 g/kg TB 會有 10%小鼠猝死之現象發生(Egorin et al., 1999)。
(c)丁酸鹽對豬生長之影響
近年來陸續有研究探討 NaB 對於離乳仔豬生長性能之影響,但結果並 不一致。研究發現,餵食 7 kg 離乳仔豬 0.17% NaB 至 102 kg 肥育豬,可提 高隻日增重 23.5%及飼料利用效率 11.8%,進而降低飼料成本 9%與增加豬 隻售價 13% (Galfi and Bokori, 1990),而飼料添加 0.30% NaB 則可提高三週 齡離乳仔豬試驗 4 週期間之飼料利用效率(Manzanilla et al., 2006)。但其他研 究顯示,飼料添加 0.10-0.40% NaB 皆不會影響仔豬生長性能(Biagi et al., 2007),另外,飼料添加 0.05-0.40%脂肪包覆形式 NaB 也不會影響三週齡離 乳仔豬試驗 4 週之生長性能(Weber and Kerr, 2008),由以上試驗結果推測 NaB 改善離乳仔豬飼料利用效率之效果大於體增重,故離乳仔豬飼料添加 NaB 應具有改善離乳仔豬生長性能之作用。
(d)丁酸鹽對腸道發育之影響
前人研究發現丁酸之生理功能,可能包括降低飼料酸鹼值及殺菌之能力
(Fay and Farias, 1975),此等酸化劑可減少腸道微生物對腸黏膜的負面影響 並促進動物生長(Partanen and Mroz, 1999),且可透過簡單擴散直接進入腸細 胞達到快速吸收之功效(Wächtershäuser and Stein, 2000),同時也會伴隨吸收 鈉離子及水分子進入腸細胞,減少腸道中水分含量而有助於預防離乳仔豬 下痢。腸細胞直接利用簡單擴散進入細胞中之丁酸作為能量來源,進而促 進腸細胞分化成熟(Smith et al., 1998),因為丁酸是腸道黏膜上皮細胞重要的 營養成分之ㄧ(Scheppach et al., 1996),其可保護腸道不受微生物侵害 (Gardiner et al., 1995)。動物試驗結果顯示,飼料添加 0.17% NaB 可提高豬 隻絨毛高度與降低胃潰瘍發生(Galfi and Bokori, 1990),而人工乳中添加 0.30% NaB 餵食新生仔豬可藉由增加血中胰臟多肽(Pancreatic polypeptide) 及膽囊收縮素(cholecystokinin)濃度,進而提高仔豬之隻日增重及小腸絨毛 發育(Kotunia et al., 2004),另外,餵食 0.10% NaB 可提高離乳仔豬腸道絨毛 高度(Wang et al., 2005)。由以上結果顯示,飼料添加 NaB 可滋養豬隻腸道 絨毛上皮細胞,而改善豬隻腸道形態與吸收能力。其他試驗則顯示,餵食 0.30% NaB 可提高結腸腺窩杯狀細胞數及增加空腸腺窩深度(Manzanilla et al., 2006),更進一步證實,飼料添加 NaB 可促進保護腸道的杯狀細胞的增 生作用,這些試驗結果推論可能是提升腸道丁酸濃度而產生之增生效果
(Tsukahara et al., 2003),但丁酸在腸道很快速被吸收代謝,因此消化道丁酸 濃度並不會隨 NaB 添加量提高而增加(Biagi et al., 2007)。
NaB 除對腸細胞具有滋養功能,動物試驗也發現添加 0.17% NaB 可提 高豬隻迴腸乳酸菌/大腸桿菌比之比例(Galfi and Bokori, 1990),以及飼料添 加 0.30% NaB 雖只具有改善三週齡離乳仔豬腸道乳酸菌/腸桿菌比例之趨 勢,但微生物嘌呤鹼基與揮發性脂肪酸(Volatile fatty acid, VFA)濃度較穩 定,雖然迴腸澱粉含量較多而導致前段結腸 VFA 及後段結腸支鏈 VFA 含量 較低,但有趣的是試驗發現 NaB 可調整微生物菌相,使微生物種類多樣化,
進而防止某些菌種或單一菌種過度增生(Castillo et al., 2006)。因此,NaB 改 變微生物複雜度與種類組成比降低消化道微生物數更具相關性,並增加腸 道有益菌比例進而改善離乳仔豬腸道微生物菌相與腸道發育。
(e)丁酸鹽對離乳仔豬之免疫調節功能
體外試驗添加 2 mM NaB 可顯著抑制豬周邊血液單核細胞增生反應,
但卻又可增加干擾素-γ之濃度(Weber and Kerr, 2006);另外之試驗在體外培 養小鼠脾臟細胞添加 0.2-0.6 mM NaB 則可提高其抗綿羊紅血球抗體力價及 IL-2 分泌量(Kishiro et al., 1994)。體外培養大鼠腸繫膜淋巴節淋巴細胞添加 1.5 mM NaB 經 con A 刺激後會抑制淋巴細胞增生反應與 IL-2 分泌量 (Cavaglieri et al., 2003),且飼料中添加 NaB 也可提高大鼠腸細胞存活率與提
高電阻,並會降低黏膜通透性與固有層嗜中性球細胞數(Venkatraman et al., 2003) 。 體 外 培 養 人 類 巨 噬 細 胞 並 添 加 5-10 mM NaB 在 脂 多 醣 (Lipopolysaccharide, LPS)刺激下,皆可增加腫瘤壞死因子α分泌量(van Nuenen et al., 2005)。綜合上述,當 NaB 濃度超過 1 mM 以上可趨使淋巴細 胞走向 Th1(T helper type 1)路徑而加強發炎反應。
飼料添加 NaB 對豬隻免疫能力影響之研究還不多,小鼠經口灌注 10 mM NaB 在結腸上皮細胞表面可發現 IL-18 之表現(Kalina et al., 2002),但天 竺鼠結腸缺乏 NaB 將誘導結腸細胞大量凋亡,並促使過量巨噬細胞進入固 有層與啟動血液中單核細胞進入腸組織中,而促進發炎反應,當灌注 10 mM NaB 後則不活化此腸道固有層發炎反應(Luciano et al., 2002)。此外,餵食四 週齡離乳仔豬發酵碳水化合物飼料,可提高迴腸 IL-1β與結腸 IL-6 mRNA 表現量並與支鏈脂肪酸具有正相關性(Pié et al., 2007);而大鼠給予發酵纖維 飼料可提高盲腸腺窩 CD8+與 CD161+密度,且與 SCFA 濃度具有相關性,
除此之外也發現 CD8+會出現於分化中腸細胞部位(Ishizuka et al., 2004),因 此給予發酵纖維飼料可改變大腸腺窩層免疫細胞之分布,並調節腸道上皮 內平衡作用,故推測 NaB 具有調節離乳仔豬免疫系統走向 Th1 路徑。
總而言之,飼料添加 NaB 可改善豬隻飼料利用效率,並滋養腸細胞而 改善腸道黏膜形態,當提供 1 mM 以上 NaB 也可驅動免疫反應走向 Th1 路 徑,以提高免疫反應能力。
(三
三三三)功能性胺基酸功能性胺基酸功能性胺基酸功能性胺基酸動物體內每天會有大量的蛋白質合成與降解之轉化作用,因此動物必 須攝取飼料蛋白質,以維持每天轉化作用。單胃動物之胺基酸種類,可分 為必需與非必需胺基酸,而動物能自體合成得到非必需胺基酸,但動物卻 無法合成必需胺基酸之碳骨架結構,因此必須由採食獲得必需胺基酸。動 物腸外組織並無法有效利用飼料中 30-50%胺基酸,需透過小腸黏膜之胺基 酸分解作用方可利用,而小腸黏膜可降解精胺酸、脯胺酸、甲硫胺酸、離 胺酸、苯丙胺酸、羥丁胺酸、甘胺酸及絲胺酸(Wu, 1998)。Kim 等人(2007) 將近年來功能性胺基酸相關研究已做初步討論,可分成下列幾點:
第一、細胞內轉化作用:例如白胺酸、麩醯胺及精胺酸對胚胎發育扮 演著重要的角色,白胺酸可提高年輕動物肌肉蛋白合成作用,且白胺酸、
異白胺酸及纈草胺酸為動物組織麩醯胺合成作用中之受質,故麩醯胺也參 與支鏈胺基酸之合成代謝。
第二、促進荷爾蒙分泌與調節新陳代謝:酪胺酸是腦中神經傳導物之 一,可促進甲狀腺、腎上腺及腦下垂體之功能;白胺酸是身體許多重要生 化成份的原料,包括與能量代謝有關的物質,以及腦中神經傳導物;丙胺 酸是肌肉組織及腦部中樞神經能量來源之一,可幫助產生抗體,協助糖類 及有機酸的代謝;苯丙胺酸是腦部及神經細胞合成神輕傳導物「新腎上腺 素」(Norepinephrine)的原料;甲硫胺酸可降低膽固醇與降低肝脂肪含量,
並協助腎臟排泄氨;甘胺酸可協助血液氧氣釋放到組織細胞,幫助荷爾蒙 的製造,加強免疫功能;絲胺酸幫助肌肉及肝臟儲存肝糖,協助製造抗體,
合成神經纖維之外鞘。
第三、免疫功能:包括麩醯胺可提高淋巴細胞增生、巨噬細胞吞噬能力、
T 淋巴細胞細胞激素分泌量及 B 淋巴細胞抗體生產量;精胺酸可誘導產生 一氧化氮;半胱胺酸在動物感染疾病狀態下可高效率地增強免疫功能;色 胺酸可增強先天免疫與抑制發炎;脯胺酸可降低腸道氧化作用。
各種重要胺基酸之特殊功能茲分述如下:
1.麩醯胺(Glutamine, Gln)
Gln 是目前研究較多之功能性胺基酸,因 Gln 是消化道與其他免疫細胞 主要能量來源之ㄧ,並且在多種異化代謝下是一個必需因子,腸道黏膜對 Gln 具有高度之耐受性,在高劑量下並不會造成副作用。研究顯示,Gln 也 是腸細胞增生、腸液/電解質吸收之影響因子,同時也是生長因子之競爭性 因子,但上皮生長因子與 Gln 共同存在下,才能提高增生效率(Rhoads, 1999);此外,肌肉內 Gln 濃度與肌肉蛋白質合成作用具有正相關 ,除骨骼 肌外,Gln 也會刺激小腸黏膜蛋白合成作用與抑制蛋白質降解作用(Kim et al., 2007),以上結果顯示 Gln 的確具有滋養腸道與促進蛋白質合成之功能。
在感染情況下,Gln 仍具有修復腸道黏膜、提高鈉離子與水分子吸收之功能
factor-I, IGF-1)刺激下,可提高仔豬腸細胞鈉離子依賴性 Gln 吸收作用,但 腸道黏膜質量多寡並不會影響 Gln 吸收作用(Alexander and Carey, 2002)。麩 醯胺除具有腸道黏膜滋養功能外,尚具有免疫調節功能,文獻指出餵食三 週齡離乳仔豬 0.5% Gln,可提高試驗第 7 天與第 14 天膽汁 IgA 與蛋白質含 量,餵食 1.0% Gln 可提高周邊血液單核細胞第 14 天之增生反應,而餵食 1.5% Gln 則可提高第 14 天之脾臟與腸繫膜淋巴結淋巴細胞增生反應(Lee et al., 2003a)。此外,飼料添加 Gln 具有提高十二指腸絨毛高度與絨毛高度/腺 窩深度比值之效應,並會降低第 14 天小腸/屠體重量比(Lee et al., 2003b)。
2.精胺酸(Arginine, Arg)
Arg 近年來也逐漸受到研究者重視,因 Arg 為必需胺基酸之ㄧ,在疾病 感染與快速生長期時會增加 Arg 之需求量,其可經由鈉離子-依賴性運送機 轉被小腸廣泛地吸收代謝而增強免疫功能與黏膜修復。若在體外試驗,腸 細胞培養液中缺乏 Arg 狀態下,會降低細胞增生與熱緊迫蛋白表現量,並 提高腸細胞對細胞凋亡之感受性(Lenaerts et al., 2007)與一氧化氮濃度並在 感染與發炎狀態下扮演著關鍵性角色;但卻必須考慮到一氧化氮毒性之提 高與抗低血壓動力學之不穩定(Luiking et al., 2005)。動物試驗結果顯示,雞 隻給予 Arg 飼料並接種傳染性華氏囊病毒,會增進腸上皮內淋巴細胞毒殺 能力、淋巴細胞增生能力及特異抗體濃度(Tayade et al., 2006),顯示 Arg 具 有促進腸道與系統性免疫反應。另外,3 日齡新生仔豬同時給予含有 Arg
與隱孢子蟲卵母細胞之飼料,會啟動依賴前列腺素之分泌性下痢,但並未 啟動腸上皮防禦機制或障壁功能(Gookin et al., 2008),而餵飼含 2% Arg 飼 料後,再誘導小鼠腸源性膿毒症(gut origin sepsis),會提高周邊血液黏附分 子、IL-4 及器官發炎細胞激素表現,但卻降低周邊血液干擾素-γ分泌量(Yeh et al., 2006),攝取含有 2% Arg 可調節 Th1/Th2 反應,並可抑制腸源性膿毒 症期間所誘導之細胞性免疫反應。由上述顯示,Arg 雖可產生一氧化氮以保 護腸道並抑制發炎作用,且具有調節發炎與抗發炎反應,但不具有促進腸 道障壁與滋養功能。
3.羥丁胺酸(Threonine, Thr)
Thr 為豬隻重要之必需胺基酸(Guzik et al., 2005; Thong and Liebert, 2004a; Thong and Liebert, 2004b),依據 NRC (1998)之營養需要量標準顯示,
5-10 kg 和 10-20 kg 體重豬隻之總 Thr 需要量分別為 0.86 和 0.74%,而真可 消化 Thr 需要量分別為 0.74 和 0.63%,但添加超過 NRC 推薦量標準之真可 消化或總 Thr 需要量 0.10%並無法進一步改善離乳仔豬生長性能與血清尿 素態氮濃度(Li et al., 1999; Wang et al., 2006a)。另外,Thr 對大鼠與新生仔 豬腸道黏液蛋白合成作用也具有重大影響,在缺乏 Thr 狀態下會降低腸道 黏液蛋白濃度,但可依 Thr 添加量之提高而增加腸道黏液蛋白濃度(Faure et al., 2005; Law et al., 2007)與血清 IgG 濃度(Li et al., 1999; Wang et al., 2006a)。
白質合成而降低蛋白質降解程度,且無論是何種功能性胺基酸皆具有調節 動物生理之功能,但若在飼料中額外添加 Thr 較能提升腸道黏液蛋白分泌 能力以保護腸道,此外,又能提升體液免疫能力以改善離乳仔豬腸道發育 與免疫反應。
茲將 Thr 之消化、吸收及功能分述如下:
(a)羥丁胺酸之消化與吸收代謝途徑
Thr 是一般穀物飼料中第二或第三限制胺基酸,且不同飼料原料之 Thr 含量也有所不同,如玉米粉與乳清粉雖易消化吸收,但 Thr 含量皆不多,
且 與 玉 米 澱 粉 混 合 時 , 易 造 成 飼 料 型 態 過 細 而 抑 制 採 食 (Wang et al., 2006a)。因此,有研究者提出不同飼料原料具有不同之 Thr 消化率,而建立 迴腸表面可消化係數(Coefficients of ileal apparent degestability, CIAD)與迴 腸真可消化係數(Coefficients of ileal true degestability, CITD),以評估飼料原 料 Thr 在動物體內的消化程度。Wang 等人(2006a)先評估其試驗主要飼料原 料 Thr 之消化率,其中玉米、花生粉及乳清粉中 Thr 之 CIAD 分別為 0.70、
0.78 及 0.88,而其 CITD 分別為 0.89、0.92 及 0.92。因此,在配製飼料前先 評估飼料原料之 Thr 消化率,以防止飼料 Thr 不足或過量而對動物個體產生 營養缺乏或毒性。
動物體養分吸收主要器官為小腸,故所有養分會經過腸細胞首渡代謝 (first-pass metabolism)轉化後,才有機會進入門脈循環中,且腸外器官無法
有效利用飼料 Thr 約 30-50%,故必需經過腸道組織異化代謝後,才能有效 利用(Wu, 1998),因此腸道扮演著一個代謝 Thr 之重要角色。飼料 Thr 皆由 小腸黏膜吸收而進入門脈循環以供給其他器官,這些器官分別是胃、腸、
胰臟及脾臟並稱為「門靜脈排流器官」(portal-drained viscera, PDV),而 PDV 會消耗飼料 Thr 總採食量一半以上(van Goudoever et al., 2006)。因此,Thr 對動物體而言是一種重要的營養成分。
(b)羥丁胺酸對仔豬生長性能之影響
Thr 為豬隻重要之必需胺基酸(Guzik et al., 2005; Thong and Liebert, 2004a; Thong and Liebert, 2004b),依據 NRC (1998)之營養需要量標準顯示,
5-10 kg 和 10-20 kg 體重豬隻之總 Thr 需要量分別為 0.86 和 0.74%,而真可 消化 Thr 需要量分別為 0.74 和 0.63%。近年來陸續有不同試驗以驗證仔豬 Thr 需要量是否需要改變,若在添加高量乳蛋白情況下,不論有無共同添加 抗生素情況下,飼料中含有 0.75%真可消化 Thr 可得到最佳之日增重與飼料 換肉率,而添加超過 NRC 推薦量標準之 0.85%真可消化 Thr 並無法進一步 改善離乳仔豬生長性能,並可依真可消化 Thr 濃度之提高而降低血清尿素 態氮濃度(Li et al., 1999; Wang et al., 2006a)。採食行為試驗顯示,豬隻能依 體內營養需求自行採食不同 Thr 濃度飼料,結果顯示採食高量 0.67%真可消 化 Thr 之飼料組,具有最佳之生長性能與最低之血漿尿素態氮(Ettle and Roth,