國立臺灣大學生物資源暨農學院農藝學系 碩士論文
Department of Agronomy
College of Bioresources and Agriculture National Taiwan University
Master Thesis
高溫下水稻穀粒粒型對米質形成之生理影響 The physiological effects of rice grain shape on grain
quality formation under high temperature
張芳瑜 Fang-Yu Chang
指導教授:盧虎生 博士 Advisor:Huu-Sheng Lur, Ph.D
中華民國 102 年 7 月
July,2013
致謝
這篇論文的完成,一路上跌跌撞撞走了將近 5 年的時間,讓我深深地感受到 自己的不足及需要加強的地方,並感謝一路上支持我的師長、前輩、不棄不離的 好朋友、情義相挺的學弟妹們以及我的家人,沒有你們,我走不到今天這步。
首先感謝盧虎生老師的指導與包容,當初沒有想到自己會在就學中途先出社 會工作,心中總是懸掛著一件未完成的事。然而老師既是個嚴師亦像個慈父般,
無論是在工作或是學業上總是給予最大的鼓勵與方向的指正。也感謝張孟基老師 及朱鈞老師,每當看我回到學校總是不忘關心我學業的進度並給予鼓勵。
再來感謝雲林縣政府農業處的許永瑜副處長(當時是我的科長),若當時沒 有她的支持與包容,同意讓我轉調到研究單位,如今也無法再回來完成這份學 業。還有感謝之前在縣府的同事們對於剛出社會的我總是給與許多協助。
接下來感謝高雄改良場的黃德昌場長大力支持研究人員進修,感謝吳志文 課長、周國隆主持人與張芯瑜學姊對於要邊處理學業的我,在工作上不成熟表現 的種種包容。
對於實驗室的大家,感謝佳諭學姊、雪玉學姊、芹如學姊、佳玲學姊、學 謙學長、凡萱、景雯、書華、子怡、雅婷、清倫、豐鳴、筱均、昌烜。謝謝你們 大家的幫忙。尤其對於佳諭姊姊致上最深的謝意,感謝妳總是不厭其煩地教導功 力不足的學妹,甚至還給與人生上的鼓勵,若沒有妳的那些激勵,我可能都沒有 勇氣撐到這一刻。再來就是,不離不棄的好朋友們,順惠、泰佑及遠在美國的若 芸。一路上感謝你們陪我度過無數的低潮時期,但你們總是能夠做出最適當的反 應讓我迅速脫離低潮的漩渦。很感謝這輩子有你們這樣的朋友。
接著感謝我的家人,謝謝阿爸阿母無限的愛,謝謝弟弟的體貼,離家工作後 才感受到家的好及發現自己原來那麼愛你們。還有感謝許多幫助過我的人。
最後感謝老天,這一路上雖然走得不順利,但感謝您給的一切考驗。
將此論文獻給我愛的每個人。
I
中文摘要
前人研究指出種子在發育時間其內部是處於低氧之環境,越靠近種子中心氧 分子濃度逐漸下降,能量生成越少。而種子內部氧氣濃度亦會隨溫度之提升而下 降。水稻在高溫下容易使產量及品質下降,尤其在開花後 20 天以內的影響最甚。
一般認為,在稉稻與秈稻兩個亞種中,秈稻的耐熱性相對較佳;因此,本研究期 望探討不同之水稻粒型在高溫下對米質形成之生理過程之影響差異,期望作為耐 熱育種改良之參考。本試驗以農業試驗所提供之臺農 67 號 (TNG67,短粒型)及 經由疊氮化鈉誘變之品系 SA1739 (長粒型)為材料,於開花後進行 15 天之日夜溫 25/20 oC 及 35/30 oC 處理,分別取開花後 6、9、15、20、25 DAF (Days after flowering) 及成熟之種子進行各項品質及生理試驗分析。
經米粒外觀分析結果顯示 SA1739 較 TNG67 心腹白比例較少,且其成熟穀 粒重量受高溫處理影響少,屬較耐熱之品系。
在常溫環境下,SA1739 的米質外觀無 TNG67 常見之腹白現象,且在能量代 謝之基因表現上與穎果內部 ATP 濃度果可知。常溫下 SA1739 在 6~15DAF 尚可 由有氧呼吸途徑提供 ATP,約自 15DAF 才開始走向無氧呼吸。然而 TNG67 穎果 在常溫下其有氧呼吸相關途徑隨發育減少其表現,取而代之逐漸提升無氧呼吸之 表現。由此顯示 SA1739 粒型改變後,已對穎果內能量合成造成影響。
在高溫環境下,TNG67 之穎果澱粉合成基因(SSIIa、GBSS)抑制較多,為高 溫下穀粒重量下降之原因。SA1739 之穎果在高溫下其 Glycolysis、TCA cycle 相 關基因(IDH、SUSB、MDH)及 ATPSB 皆受抑制,但 PPDKA 大量表現,推測 SA1739 穎果於高溫下其 PPDKA 可扮演 PK(pyruvate kinase)之角色,將 PEP 轉 pyruvate,
提供無氧呼吸之原料,以生產 ATP 供應其內部之代謝反應。反觀 TNG67 高溫下 參與有氧呼吸途徑之基因亦受到抑制但 ADH 提升之幅度較少,可能無法供應生 理代謝所需之足夠能量。能量方面,高溫下 SA1739 在早期其 ATP 仍可維持常溫 之狀態,且常溫與高溫下 ATP 最大濃度差異小,可能有利於減少白堊質粒產生。
II
另 SA1739 榖粒充實早期有較高之 SOD 表現,推測可將 superoxide 轉換為毒性 較弱之 H2O2,降低脂肪過氧化程度(MDA),此外其多元胺合成基因(OsSAMDCI、
SPDSYI)表現提升,可能有較多含量多元胺而使得其在高溫下細胞死亡指標(PBZ)
基因表現相較 TNG67 低。反觀 TNG67 高溫下 ACO1 早期表現提升幅度大,可 能有較多乙烯之產生,另多元胺合成基因提升幅度相對較少,加上有多元胺降解 基因(PAO)大量表現之現象,而導致其細胞死亡指標(PBZ)相較嚴重。
依本試驗 TNG67 與 SA1739 初步之結果推斷,外觀具細長粒型之 SA1739 具有較好之能量利用效率,可能減緩高溫對穀粒充實之傷害,但是否代表其穎果 內部具有較高之含氧含量還需待實測該穎果內部氧含量才能驗證。本試驗之水稻 長粒型與短粒型品系對高溫之品質與生理反應之差異性的研究成果,將可提供因 應氣候暖化之耐熱性育種的理論基礎。
關鍵字:高溫、粒型、白堊質、能量
III
英文摘要
It is known that the seed usually develops under hypoxia state with the oxygen concentration decreased from outer to inner layers of the seed. Reports also found that increasing temperature would further decrease oxygen concentration in the seed.
Many researches reveal that high temperature stress may lead to a poor yield and grain quality, especially before 20 days after flowering. Furthermore, indica type rice, which usually has long grain shape, usually had better heat tolerance than does the japonica type. In the present study, we want to understand the effects of grain shape on grain quality formation under high temperature stress. After flowering, TNG67 and SA1739 (a mutant line fromTNG67, with slender/long grain shape) were treated with 25/20oC (day/night temperature) and 35/30oC for 15 days . The current results
revealed that grains of mutant line had a lower chalkiness than that of TNG67 under high temperature.
From the results of energy metabolism related gene expression and ATP concentration under normal temperature (25/20oC, day/night), we found that
SA1739 could go through oxidative phosphorylation during 6~15 DAF to supply ATP.
However, TNG67 only could produce ATP from fermentation even under 25/20oC.
Mutation of the grain shape of SA1739 from TNG67 might changed their energy metabolism during grain filling.
In gene expression, starch synthesis genes (SSIIa, GBSS) decreased seriously in TNG67 under high temperature. The grains of SA1739 had higher ATP concentration at early developmental stage. SA1739 showed up-regulation in expressions of
PPDKA, ADH and down regulation of TCA cycle genes under high temperature,
suggesting a role of PPDKA to change PEP to pyruvate, so that alcohol fermentation in the long grain mutant line could supply enough ATP under high temperature. In addition, SA1739 had higher expression levels on SOD expression to decrease lipid
IV
peroxidation and high expression level of OsSAMCDI, SPDSYI , but less expression on PAO, that might implied a higher content of polyamines under high temperature.
SA1739 also showed lower expression of cell death related gene PBZ under high temperature. On the other hand, the short grain cultivar TNG67 had higher expression rate of ACO1 at early stage and less expression on OsSAMCDI and SPDSYI under high temperature, suggesting that it might release higher amount of ethylene and lower amount of polyamines under high temperature, and in turn enhanced a higher expression of cell death related gene PBZ.
From the present results, it is suggested that SA1739 may have higher energy metabolizing efficiecly under high temperature, so that it may have better dry matter accumulation and less chalk grains. However, the relationship between the internal oxygen status in caryopsis with grain shape needs to be clarified in the future .
V
目錄
致謝……….Ⅰ 中文摘要………..………Ⅱ 英文摘要………Ⅳ 目錄………Ⅵ 圖與副表目錄………Ⅶ 縮寫字對照表………Ⅸ
壹、 前言………1
一、 高溫導致水稻產量、外觀及食味品質下降………1
二、 高溫與穎果發育之內生低氧環境與氧化逆境 ………6
三、 植物荷爾蒙對水稻穎果發育與品質影響………7
四、 穀粒粒型與水稻產量品質………8
五、 試驗推論………..10
貳、 材料與方法.……….11
參、 結果與討論 ………18
一、 TNG67 與 SA1739 穎果發育之差異……….18
二、 TNG67 與 SA1739 之生理反應差異及相關基因表現……….22
肆、 結論……….37
伍、 參考文獻……….39
陸、 圖與表格……….46
VI
圖與附表目錄
圖 1. TNG67 與 SA1739 於常溫及高溫下不同階段穎果發育之外觀。…………49
圖 2. TNG67 與 SA1739 於常溫及高溫處理下穎果發育之生長曲線(A)、含水率與 水分含量(B)與成熟粒重(C)。………...………50
圖 3.TNG67 與 SA1739 開花後 30 天內於常溫及高溫下之葉綠素值反應。 …...…52
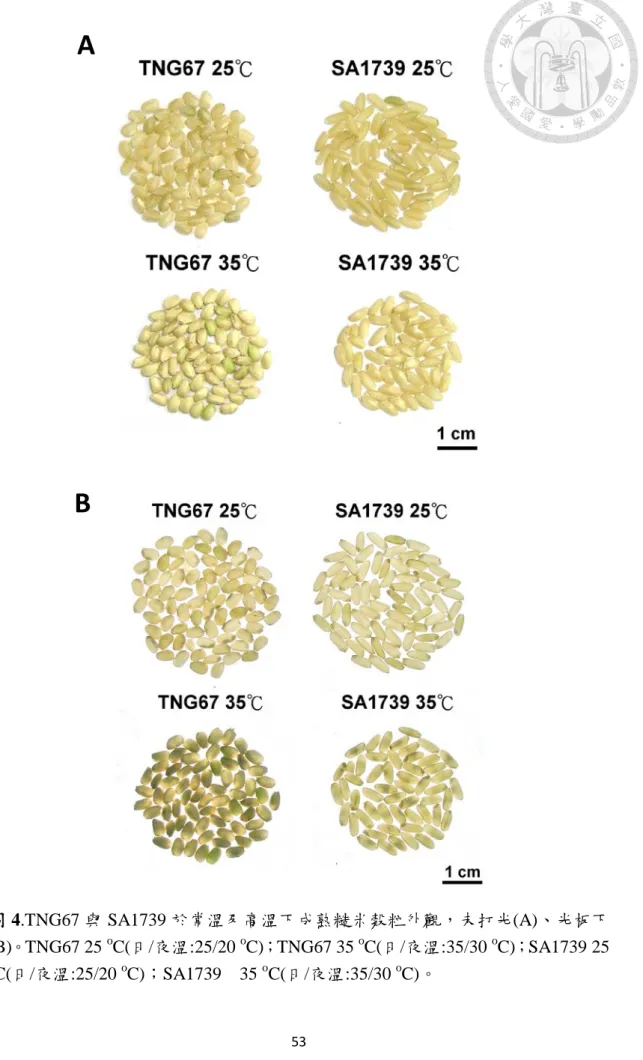
圖 4.TNG67 與 SA1739 於常溫及高溫下成熟糙米穀粒外觀,未打光(A)、光板下 (B)。……….………53
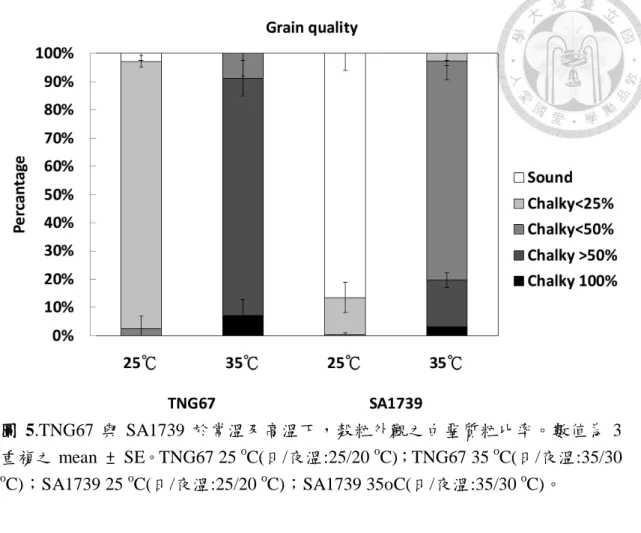
圖 5.TNG67 與 SA1739 於常溫及高溫下,穀粒外觀之白堊質粒比率。 ………..…54
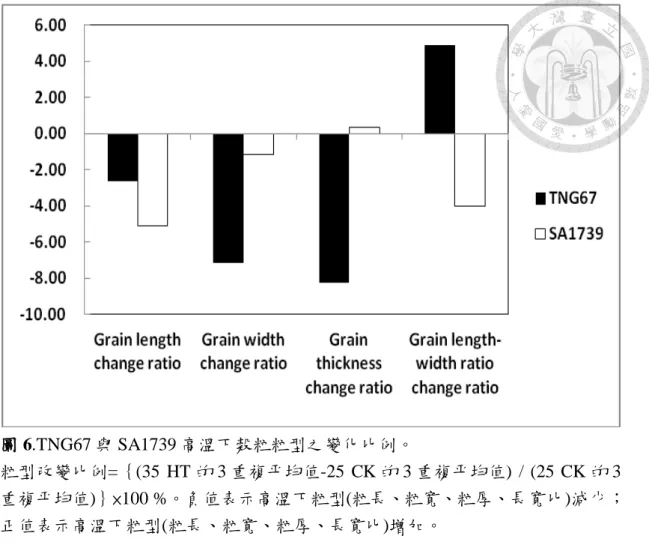
圖 6.TNG67 與 SA1739 高溫下穀粒粒型之變化比例。………..…………55
圖 7.TNG67 與 SA1739 穎果 pH 變化。………57
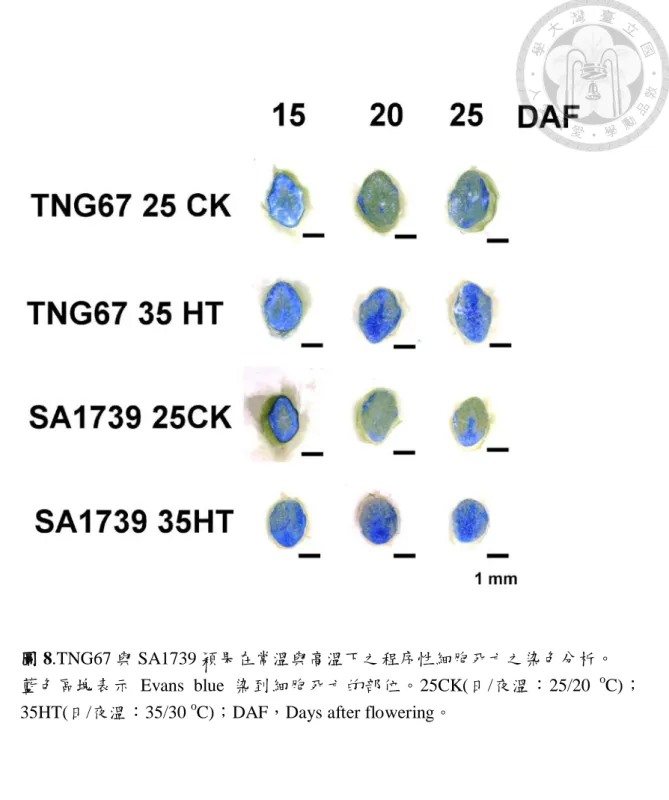
圖 8.TNG67 與 SA1739 常溫與高溫下程序性細胞死亡過程。………..………58
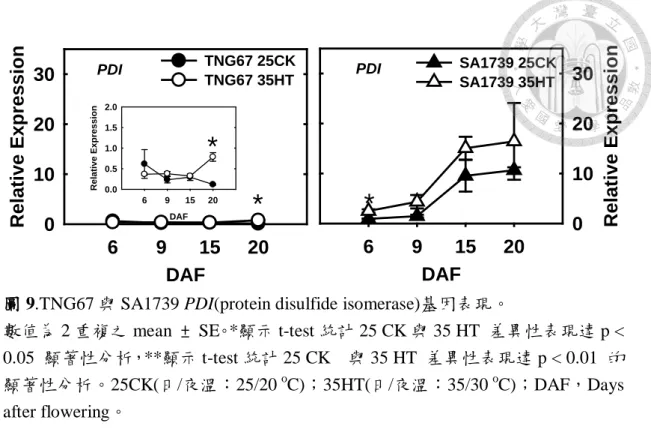
圖 9.TNG67 與 SA1739 PDI(protein disulfide isomerase)基因表現。………....……59
圖 10.TNG67 與 SA1739 細胞死亡基因表現。………60
圖 11.TNG67 與 SA1739 在常溫及高溫下 ATP 濃度之變化。………61
圖 12.TNG67 與 SA1739 參與澱粉合成基因表現。………62
圖 13.TNG67 與 SA1739 儲藏性蛋白相關基因表現。………63
圖 14.TNG67 與 SA1739 糖解作用相關基因表現。………64
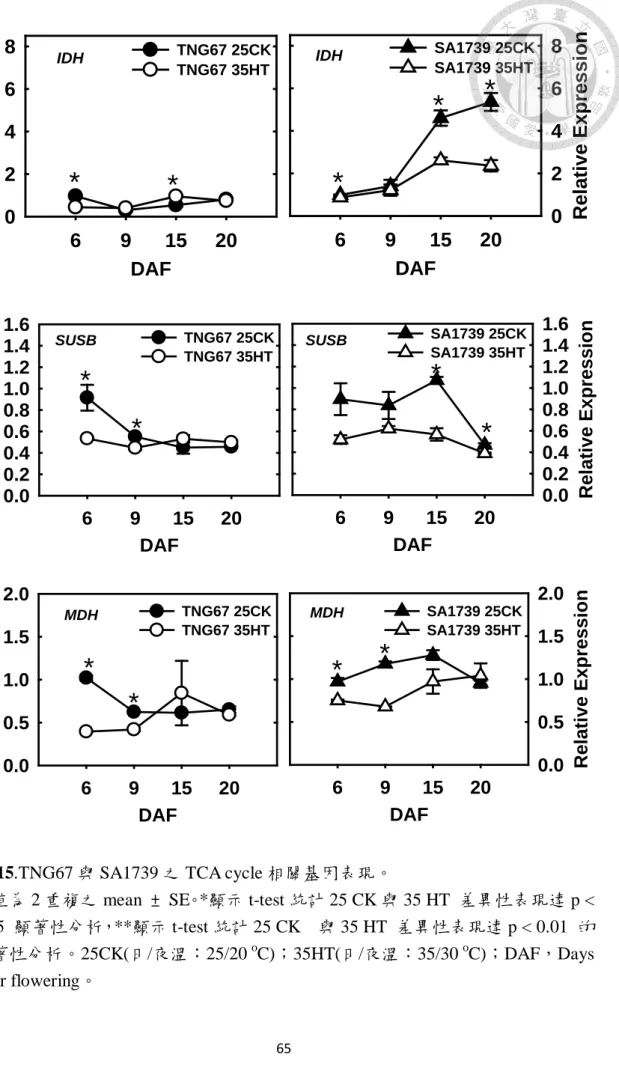
圖 15.TNG67 與 SA1739 之 TCA cycle 相關基因表現。……….………65
圖 16.TNG67 與 SA1739 有氧呼吸與無氧呼吸相關基因表現。………66
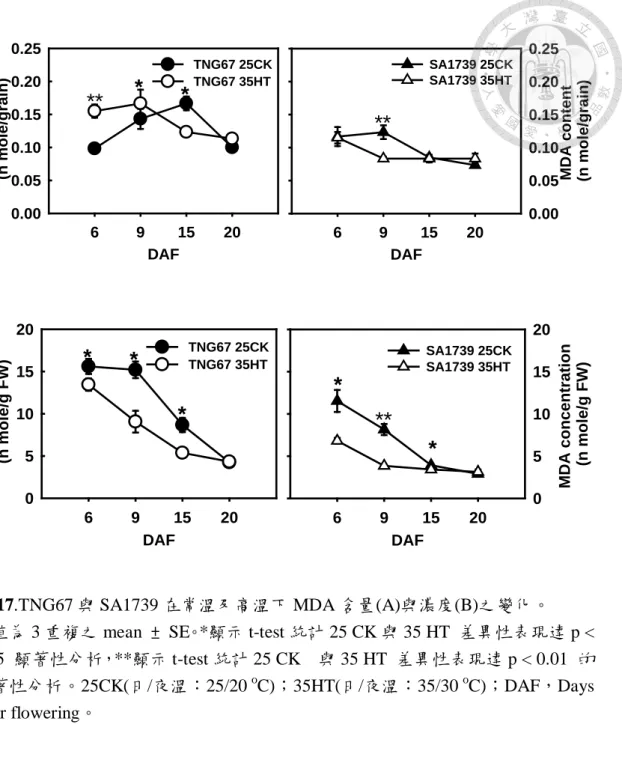
圖 17.TNG67 與 SA1739 在常溫及高溫下 MDA 含量(A)與濃度(B)之變化。……..67
圖 18.TNG67 與 SA1739 之 SOD 基因表現。………...……68
圖 19. TNG67 與 SA1739 消除 H2O2相關基因表現。 ………...……69
圖 20.TNG67 與 SA1739 在常溫及高溫下 H2O2含量(A)與濃度(B)之變化。……...70
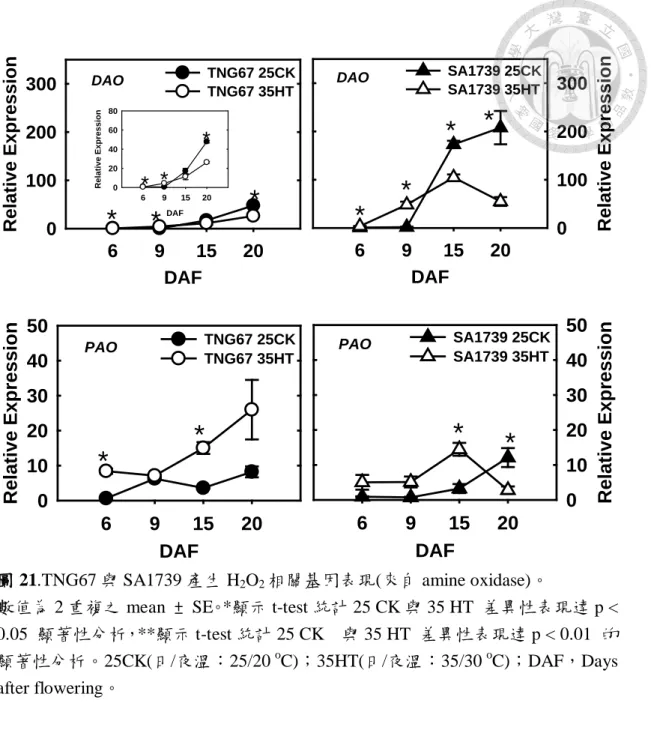
圖 21.TNG67 與 SA1739 產生 H2O2相關基因表現(來自 amine oxidase)。…………71
圖 22.TNG67 與 SA1739 參與乙烯相關基因表現。………72
VII
圖 23.TNG67 與 SA1739 參與多元胺合成相關基因表現。………74
圖 24.TNG67 與 SA1739 逆境相關基因表現。………76
圖 25.高溫下澱粉合成相關基因表現。………..…………78
圖 26.SA1739 耐熱機制。………...79
表 1、水稻生長發育生理基因及逆境相關基因列表,及期表現分析使用之引子擴 增之片段長度。………46
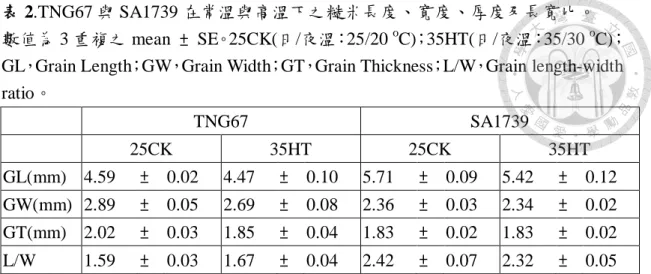
表 2.TNG67 與 SA1739 在常溫與高溫下糙米長度、寬度、厚度及長寬比。……..…56
VIII
縮寫字對照表
ACO ACC oxidase
ADC Arginine decarboxylase ADH Alchohol dehydrogenase
AGPase ADP-glucose-1-phosphate uridylyltransferase AlaAT Alanine amynotransferase
ASC ACC synthase
ATP Adenosinetriphosphate BE Starch branching enzyme CAT Catalase
DAF Days after flowering DHAR Dehydoascorbate reductase DNAase Ca2+ dependant-endonuclease DW Dry weight
FBPase Fructose-bisphosphatase FW Fresh weight
GBSS Granule-bound starch synthase GLO1 Glyoxylase Ⅰ
HSP Heat shock protein IDH Isocitrate dehydrogenase MDA Malondialdehyde
MDH malate dehydrogenase MG Methylglyoxal
ODC Ornithine decarboxylase PAO Polyamine oxidase PB Protein body
PBZ Probenazole-induced protein PCD Program cell death
PDI Protein disulfide isomerase PFK Phosofructokinase
PK Pyruvate kinase
PPDK Pyruvate phosphate dikinase Pro7 Prolamine 7
SAM S –Adenosylmethionine
SAMDC S -Adenosylmethionine decarboxylase SAMS S –Adenosylmethionine synthetase
IX
SMS Spermine synthase SOD Superoxide dismutase SPDSY spermidine synthase SUSB succinyl-CoA synthetase B
UGPase UDP-glucose-1-phosphate uridylyltransferase
X
壹、前言
根據世界糧食組織(FAO)於之統計年報顯示,2009 年每人平均可耕地面積約 0.2 公頃,約為過去 50 年之一半。水稻為全球生產量前 3 名之糧食作物,在全球 人口不斷上升,而可耕地面積逐年下降之情況下,據 FAO 2012 年之統計年報估 計至 2050 年全球所需之水稻量尚需提升一倍。另地球自 1906 年至 2005 年,平 均氣溫約提升了 0.75 oC(IPCC 2007)。全球暖化之現象已造成日本九州當地生產 一等米比例逐年下降,影響當地農民之收入。臺灣位處之緯度尚比九州低,臺灣 良質米生產之溫度臨界值為:日高溫小於 28 oC、日均溫小於 26 oC 及日低溫小 於 22 oC(盧 等,2006)。在台灣一期作除臺東地區,個地區水稻充實期容易遭受超 過 26 oC 之環境,二期作以高屏地區較易超過 26 oC(Lin et al., 2005)。臺灣自 2001 年至 2008 年白堊質粒和位熟粒比率有逐年上升現象,而一、二期稻作穀粒充實 溫度與白堊質粒及未熟粒呈現正相關(Lur et al.,2009),全球暖化對臺灣稻米生長 及品質之衝擊已是不可忽視之課題。此外,氣候變遷也使全球水稻生產面積最大 之國家-印度在 2012 年遭受豪雨威脅,使該國水稻生長遭受衝擊進而影響全球 之稻米供應。以上顯示在氣候變遷下維持世界糧食安全是現今農藝學家所需面對 之挑戰。
一、高溫導致水稻產量、外觀及食味品質下降
水稻(Oryza sativa)常見之兩亞種為秈稻 indica 及稉稻 japonica,一般稉型稻 較適合生長於溫帶地區,而秈稻較適應熱帶地區。臺灣所處之緯度為最適稉稻栽 培區之最南端,而一般臺灣穀粒充實期平均溫度多高於 26 oC,日低溫容易超過 22 oC,日高溫容易超過 30 oC,均容易產生白堊質粒(盧 等,2006)。高溫對穀粒 充實期造成之損害有:(1)白堊質粒產生、(2)穀粒容積變小、(3)粒重下降、(4)食 味品質下降之現象。水稻之充實平均溫度若超過 25 oC,易導致米粒外觀及食味 品質下降(松江 et al., 2003; 森田, 2009)。水稻抽穗後 20 天內為對高溫最敏感之
1
時期 (Tashiro and Wardlaw, 1991a),其平均溫若高於 24oC 容易導致心白粒,若 高於 27oC 則背白粒及乳白粒之比例提升,超過 36oC 容易導致死米之發生。以 遭受高溫逆境之時期探究,於充實初期至中期 (抽穗後 4 至 20 天)遭受高溫,容 易產生乳白粒;而在充實中後期 (抽穗後 16 至 24 天)遭受高溫則容易產生背白 粒(Tashiro and Wardlaw, 1991a)。高溫使穀粒充實不良、穀粒變薄及米糠層增厚,
增加未熟粒之比率,進階造成粒重下降之現象。穀粒充實期平均氣溫若超過 24oC,容易導致糙米粒重下降(Yoshida and Hara, 1977),其關鍵期約在開花後 6 至 16 天(佐藤 et al., 1973);而 Tashiro and Wardlaw(1991b)之研究約在開花後 12 至 20 天。另在秈稻及稉稻兩個亞種間影響粒重下降之溫度,秈稻所能適應之溫 度較稉稻高(Yoshida and Hara, 1977)。高溫導致白堊質粒產生及粒重變輕,使米 質及產量下降,對市場價格產生影響。
(一) 高溫與水稻穎果發育
在 Morita 等人 (2004) 之研究中顯示,穗遭受到高溫逆境下之傷害比葉高溫 之傷害大,其穀粒千粒重明顯下降,顯示水稻穎果之生長發育受到影響。一般環 境下,水稻穎果在開花後 2 至 8 天為穀粒大小增加時期,而胚乳之細胞分裂與分 化都在開花後 8 天前進行,胚之發育在開花後 10 天左右成熟,而在開花後 11 至 15 天穎果已達成熟穀粒大小,開花後 5~20 天開始為澱粉累積主要時期,蛋白質 累積自開花後 7 天開始,水稻穎果中澱粉約佔了 80%,蛋白質約 7%到 8%,是 穎果重量主要來源,因此該階段為穀粒充實期。約在開花後 18 天,穎果之乾鮮 重皆不再持續增加,進入脫水階段而達穀粒成熟(Ishimaru et al., 2003; Xu et al., 2008)。
在 Yamakawa 等人 (2007) 之研究中,高溫下(日/夜溫為 33/28oC)加速穀粒 充實速率,縮短穀粒充實期,並使穀粒乾重下降,並且呈現完整米粒比率下降及 白堊質粒增加之情況。夜高溫對產量之影響較日高溫嚴重,日低溫每上升 1oC
2
將使產量減少 10%(Peng et al., 2004)。夜高溫和日高溫均會使穎果之胚乳細胞數 增加,但夜高溫下每個胚乳細胞平均面積減少最為嚴重,而使得胚乳之總面積減 少,使產量下降(Morita et al., 2005)。
Westgate 及 Boyer (1986)在玉米之研究中發現,當種子內之乾物種累積至最 高峰時,有最大之水分含量,之後種子內的水勢(Ψw)急速下降,水分含量跟著 下降,表示種子的最大含水量可能 sink 的容積有關。水稲方面,夜高溫下產生 之粒重下降現象外,亦觀察到夜高溫會使水稻穎果內之水分含量及含水率下降嚴 重,推測可能和胚乳細胞發育受到抑制有關(森田, 2009)。另在 Ishimaru (2009) 研究中發現,高溫下造成白堊質粒主要為乳白粒及心白粒,其白堊質發生在穀粒 中心部位。透過核磁共振影像觀察發現,高溫處理下之發育前期穎果之胚乳中心 有較低之含水量,伴隨著在電子顯微鏡下觀察到之澱粉粒堆疊不整齊現象。然而 在充實前期及中期並無偵測到澱粉水解酵素α-amylase 之基因表現,顯示乳白 粒及心白粒並非由α-amylase 導致而成,可能和水分之調控有關。
從 sink(葉)-source(穎果)角度探討高溫對產量之影響,可發現穀粒充實期高 溫下葉片光合作用下降(Chowdhury and Wardlaw, 1978),加上穀粒基部之通導輸 送組織可因脫水酵素之作用而提早退化,造成葉片裡之碳水化合物及水分無法有 效從穗軸供給至穎果;另一方面穎果因呼吸作用提升降低了碳水化合物之合成作 用,且珠心表皮也在高溫下發生充實初期到中期之提早退化現象,致使最後之粒 重下降。(中山, 1969; 岩澤 et al., 2003; 森田, 2009)
(二)高溫與水稻穎果內醣類與澱粉之合成代謝
白堊質粒之產生主要是因為澱粉粒與蛋白質粒堆疊不整齊,空隙中之空氣反 射產生之白濁現象。此外,高溫下使穎果內碳水化合物同化作用抑制也影響了澱 粉之合成代謝。因此,穎果內澱粉合成之調控成為關鍵之因素。
在 Yamakawa 等人 (2007) 與 Yamakawa and Hakata (2010)之研究探討高溫
3
下導致澱粉合成下降的原因可能有:(1)sucrose transporter (SUT1,SUT2)基因表現 下降,使 sucrose 運輸進入胚乳細胞能力下降,合成澱粉之能力因而受影響;(2) 澱粉合成之前驅物 ADP-glucose 之合成酵素 UGPase 及 AGPase 基因表現量下 降,但 ADP-glucose 之含量卻在高溫下提升,顯示 ADP-glucose 產生後之生合成 步驟可能受影響。也發現 ADP-glucose transporter 之 BT1 基因表現降低,導致 ADP-glucose 無法運送進入 amyloplast,降低澱粉合成能力;(3)澱粉合成基因 GBSSⅠ,SSⅡa,SSⅢa 及 BEⅡb 皆受到抑制;(4) α-amylase 之基因表現及酵素活
力提升使澱粉降解轉換為能量利用;(5)參與糖解作用的酵素(phosphofructokinase 及 pyruvate kinase)基因表現超過醣質新生之酵素(FBPase 及 PPDK),使 sugar phosphate 不易走向澱粉合成途徑;(6)呼吸傳遞鏈上的 cytochrome c oxidase, cytochrome c reductase 及 F-type H+-transproting ATPase 皆受到抑制,降低 ATP 之 合成。其中,抑制 BEⅡb 之表現可影響支鏈澱粉結構並導致長臂鏈支鏈澱粉增 加,導致白堊質粒產生且影響食味值(Nishi et al., 2001; Yamakawa et al., 2007;
Yamakawa and Hakata, 2010)。另利用 RNAi 抑制α-amylase 之轉殖株在高溫下表 現,白堊質粒比率有下降之現象,尤其抑制乳白粒之發生,顯示α-amylase 可能 廣泛地在胚乳細胞中影響白堊質粒產生(Hakata et al., 2012)。
(三)高溫影響穎果胺基酸代謝與蛋白質合成
水稻穎果內蛋白質含量,雖僅占 7~8%,對產量之影響有限,但於品質及營 養價值影響甚大。水稻儲藏性蛋白質可分為酸鹼溶性蛋白 glutelins、醇溶性蛋白 prolamins、鹽溶性蛋白 globulins 及水溶性蛋白 albumins。一般穀類以 prolamins 含 量 最多,而水稻穎 果內 儲藏蛋白以 glutelins 為主,約 60~80% ,其次為 prolamins,約 20%。Glutelins 約在開花後 5 天開始累積於胚乳細胞,prolamins 約在開花後 10 天開始合成。此外 protein body Ⅱ(PBⅡ,glutelins 及 globulins 主 要累積之部位)合成較 protein body Ⅰ(PBⅠ,prolamins 累積之部位)早於胚乳細
4
胞 中 (Tanaka et al., 1980; Yamagata et al., 1982; Shewry and Halford, 2002;
Nishimura et al., 2006)。PBⅠ較 PBⅡ難消化,且 prolamines 與食味品質成負相關 (Ogawa et al., 1987; Lin et al., 2005)。
高溫下穎果內 amino acids 發生累積之現象,其中的因素(1) amino acid transporter 在高溫下不受影響,穩定運送 glutamine、asparagine 及 serine;(2)胺 基酸由碳代謝途徑轉換而來,尤其從 pyruvate 與 oxaloacetate 這種介於碳氮代謝 的轉換點。其中 AlaAT 和 ALS 酵素在高溫下表現量提升,將 pyruvate 轉換為 analine、valine 及 leucine。而 AspAT 的表現量提升,使 oxalateacetate 轉換為 aspartate,觀察到高溫下參與糖解作用與 TCA cycle 的 sugar phosphates 與有機酸 有降低之現象;(3)aminoacyl t-RNA synthrtase 表現量下降,tRNA 攜帶 amino acid 能力降低,進而影響蛋白質的合成;(4)轉錄與轉譯作用受到抑制使蛋白質合成 能力之降低,且蛋白質水解作用在高溫下不受影響,使得蛋白質降解產生之胺基 酸 無 法 有 效 進 入 蛋 白 質 合 成 途 徑 , 導 致 穎 果 內 部 胺 基 酸 含 量 累 積 之 現 象 (Yamakawa and Hakata, 2010)。雖然 amino acid 對產量影響不大,但對於外觀品 質與營養價值有較大之影響。
另高溫下穎果內 13-kD prolamin 含量下降及 protein disulfide isomerase(PDI) 表現量下降之現象(Yamakawa et al., 2007)。由於 prolamines 主要累積於內質網衍 生之蛋白質粒(PB-ER),並以 protein body I(PB-I)之形式存在。glutelin 之前驅物在 cisternal ER(C-ER)合成,並由高基氏體運送至蛋白質儲存液胞 (protein storage vascuole,PSV,也稱為 PBⅡ)進行後轉譯作用,並藉由雙硫鍵將酸鹼亞基結合 (Müntz, 1998)。PDI 含有 ER resident signal,主要在內質網幫助多肽之雙硫鍵形 成,此外亦在 C-ER 影響 glutelin 前驅物之成熟(Wilkinson and Gilbert, 2004;
Satoh-Cruz et al., 2010),因此 PDI 之缺失將使蛋白質之組成及含量改變(Yasuda et al., 2009)。而在 PDI 缺失的水稻突變體中,穀粒外觀呈現白堊質之現象,胚乳有 不規則的澱粉粒及 PB-I 累積(Kim et al., 2012),顯示 PDI 參與胚乳內部蛋白質合
5
成之調控。
二、高溫與穎果發育之內生低氧環境與氧化逆境
種子發育過程是在低氧環境進行(Rolletschek et al., 2004),氧濃度由種子外部 向內部遞減,而其 ATP 含量於種子分布之位置與氧濃度一致,澱粉、蛋白質與 脂質的分布與 ATP 及氧濃度有關。高溫下使發育中向日葵種子降低種子內部氧 氣溶度(Rolletschek et al., 2007)。低氧環境容易導致氧化逆境之產生。活化氧族 (ROS)產生是低氧環境之特性,亦是為了刺激細胞產生氧氣(Blokhina et al., 2003)。
在大麥穎果內發現,隨著發育過程胚乳細胞逐漸酸化,此現象始於乾物質累 積達最大值、細胞開始進行脫水及種皮葉綠素降解時期,而酸化之來源自種皮或 糊粉層釋放之大量 malate 及少量 citrate 及 lactate。而種皮上 PEP carboxylase 及 Alcohol dehydrogenase(ADH)之活性也隨著提升。種皮在酸化過程仍保有較高的 呼吸速率,而胚乳細胞呼吸作用則逐漸降低(Macnicol and Jacobsen, 1992)。
在水稻穎果發育過程中,開花後 15 天澱粉粒開始於胚乳內部累積至開花後 18 天遍部整個胚乳,程序性細胞死亡(Program cell death, PCD)發生於澱粉粒堆疊 後。而約在開花後 12 天於內部之胚乳可偵測到 H2O2,於開花後 15 天有 H2O2 大量累積之現象,顯示 H2O2發生於 PCD 前,H2O2可能扮演著調控胚乳細胞內 部澱粉累積、細胞死亡及養分運輸之角色。隨著穎果發育,穎果內 ATP 濃度遞 減且無氧呼吸作用上升,表示水稻穎果發育是低氧逆境(Xu et al., 2008; Xu et al., 2010)。
水稻根部在低氧環境下會誘導 alanine 及 GABA 的產生(Reggiani et al., 1988)。水稻穎果在高溫下呼吸傳遞鏈相關酵素之基因表現量受到抑制且有 alanine 及 GABA 累積之現象,顯示高溫下穎果亦是低氧之環境,而與其合成之 酵素 Ala AT 及 glutamate decarboxylase 之基因表現亦有所提升,但參與 GABA
6
的代謝酵素 GABA aminotransferase 卻受到抑制(Yamakawa and Hakata, 2010)。相 關研究指出 GABA shunt 與氧化逆境耐性有關(Smirnoff and Cumbes, 1989;
Bouché et al., 2003)。此外,水稻高溫穎果內有 glutathione 累積之現象(Yamakawa and Hakata, 2010),而 glutathione 是重要的氧化還原劑,藉由將電子轉給 DHAR 以維持 ascorbate 的含量,去除 H2O2。在(Liao et al., 2012)等人的研究也指出,高 溫下 ROS 於植物細胞內扮演調控 TCA cycle、呼吸傳遞鏈、核酸複製、細胞分化 等角色。
三、植物荷爾蒙與水稻穎果發育對高溫之生理反應
水稻穎果隨發育過程有 H2O2 累積現象。植物體內 H2O2 主要產生來源:
(1)superoxide dismutase、(2)apoplastic oxalate oxidase、(3)amine oxidase、(4)class Ⅲ peroxidase、(5)fatty acid β-oxidation、(6)glycolate oxidase (Mittler, 2002)。Amine oxidase 參與多元胺之降解,其 polyamine oxidase(PAO)及 diamine oxidase (DAO) 分解 spermidine、spermine 及 putrecine 後產生 H2O2。多元胺參與植物體內之細 胞分裂、型態發生、胚胎發育、果實發育及逆境反應。
水稻上穗位的穎果主要分布於在 primary branch,相較於下穗位之穎果有較 高之細胞分裂、穀粒充實速率及粒重。上穗位之穎果除有較高之 spermidine 及 spermine 含量,且 arginine decarboxylase(ADC)、S-adenosylmethionine
decarboxylase(SAMDC) 及 spermidine synthase 之 活 性 亦 較 高 。 另 額 外 供 給 spermidine 及 spermine 與下穗位,可提升其細胞分裂及穀粒充實速率,並伴隨著 澱粉合成相關酵素活性之提升,表示多元胺參與著穎果之發育及澱粉代謝(Yang et al., 2008) 。此 外 ,下穗 位 較上 穗 位含有 較多乙烯及 1-aminocylopropane- 1-carboxylic acid (ACC) 含 量 , 且 乙 烯 合 成 相 關 基 因 1-aminocyclopropane-1- carboxylate oxidase 1 (ACO1) 及 ACO3 表向量亦較高,乙烯釋放量與細胞分裂級 穀粒充實速率成負相關且會抑制澱粉合成基因之表現(Yang et al., 2006; Zhu et al.,
7
2011)。
在玉米敗育的穀粒中,多元胺含量降低,其現象與低胚乳細胞核數及 DNA 含量相關(Liang and Lur, 2002)。穀類作物胚乳細胞的 PCD 過程,伴隨著 nuclease 活性提升及 DNA 降解現象,並受到乙烯及 abscisic acid (ABA)之調控(Young and Gallie, 2000),顯示多元胺亦可能參與著 PCD 之過程。
水稻穎果內乙烯釋放速率及 ACC 濃度與白堊質形成有關,高濃度 ACC 容 易導致澱粉粒堆疊鬆散,澱粉粒之間有較大之空隙,增加白堊質比率(Yang et al., 2007)。由於高溫下白堊質粒發生比率提高,水稻穎果內α-amylase 表現量及活 性上升 。由 於在 種子 發芽過 程中,會 誘導α-amylase 大量表現,主要受到 gibberellin(GA)之正向調控,受到 abscisic acid (ABA)之負向調控。但在高溫下水 稻穎果之 ABA、jasmonic acid(JA)及 salicylic acid (SA)含量下降,而 cytokinin(特 別是 isopentenyl adenine)、amino acid conjugated auxin 含量有提升之現象,顯示 若干植物荷爾蒙可能與白堊質粒形成相關,但它們的生理機制目前瞭解相當有限 (Hakata et al., 2012)。
四、穀粒粒型與水稻產量品質
在生理上,高溫下穀粒粒厚減少使產量構成要素中之充實粒比例下降,而導 致產量下降(佐藤 et al., 1973; Tashiro and Wardlaw, 1991)。高溫亦會造成粒寬及粒 厚減少之現象,使胚乳容積變小進而影響粒重,尤其以高夜溫之影響較為嚴重 (Morita et al., 2005)。而水稻耐熱品種 Kimmaze 在高溫下減少粒長卻粒寬與粒 厚,而高溫下其穎果內部之 ATP 濃度高峰可以維持常溫之水平,而其他不耐之 品種日本晴與台中 65 號之粒厚與粒寬在高溫下有縮減之現象,穎果內部之 ATP 濃度高峰皆會下降(She et al., 2010)。
秈稻與稉稻在耐熱性上的研究,高溫皆會影響稔實率,但高溫下秈稻的花粉 的萌發率及活性下降比率普遍較稉稻低(Prasad et al., 2006)。在秈稉稻的演化研
8
究,利用與野生稻 O. rufipogon 內的酵素,atpB-rbcL、p-VATPase 及 SAM 的 haplotype 分析,可推測秈稻演化源於西瑪拉雅山南端,如東印度、緬甸及泰國;
而稉稻演化自中國南方(Londo et al., 2006)。利用同功酶 (Isozyme)分類亞洲水稻 品種的研究裡,起源於孟加拉、泰國的品種其酵素分群屬第Ⅰ群;起源於韓國及 日本的品種其酵素分群屬第Ⅵ群。而針對先稉稻分析發現,秈稻的酵素分群多屬 第Ⅰ群,稉稻的分群多屬第Ⅵ群(Glaszmann, 1987)。在影響粒型寬度基因 qSW5 的研究裡,將 Kasalath 的 qSW5 利用染色體至換將或利用回交育種方式導入日本 晴中,導致粒型寬度減少。另外 qSW5 在地裡分布上,帶有 qSW5 的品種(粒寬較 窄)較帶有 qsw5 者(粒寬較寬)多分於較低之緯度(Shomura et al., 2008)。以上顯示 秈稻在演化的起源及長粒型在地理位置分布上皆在熱帶地區,可能因此使秈稻普 遍較能適應高溫環境。
9
五、試驗推論
由上述前人研究,一般具長粒型之秈稻皆較短粒型之稉稻適應高溫環境,但 多著墨於在產量層面,生理層面之機制探討甚少。推測秈稉稻可能因粒型之不 同,而使穎果發育過程內部之氧濃度較易擴散到秈稻之內部,可使穎果避免缺氧 現象,進而擁有較高之能量以因應高溫下澱粉與蛋白質等物質合成。所以本試驗 利用臺農 67 號(短粒形)及臺農 67 號誘變品系-SA1739(長粒型),探討不同粒型 在高溫下穎果內能量代謝、氧化逆境、植物荷爾蒙對米質之影響,瞭解造成秈稉 型稻對高溫不同耐受性之原因,以作為因應氣候暖化之水稻耐熱育種之理論依 據。如下試驗推論圖所示。(藍色件頭表示影響的方向;紅色箭頭表示兩者為對 立關係)
10
貳、材料與方法 一、試驗材料栽培與取樣
本試驗材料由台灣農業試驗所(Taiwan Agriculture Research Institute)所提供 之稉型水稻(Oryza sativa sp. Joponica)-臺農 67 號(TNG67)及以疊氮化鈉(Sodium azide, NaN3)誘變處理之誘變品系 SA1739。TNG67 及 SA1739 未去穎之榖粒長 度、寬度及長寬比為:0.69 cm/0.37 cm/1.88 ; 0.83 cm/0.33 cm/2.57。外觀如下圖 所示。
選取健康之榖粒於燒杯內以 1 %次氯酸鈉(NaOCl)消毒種子 20 分鐘後,於 燒杯內加入少許水量,置於 37 oC 生長箱中催芽。待種子發芽後,於室溫及自然 光照下以木村氏水耕液栽培生長。水耕液每隔兩天換一次。待幼苗長至 2~3 葉齡 即可移植至面積為 1/200000 (22×22 cm),深度 28 cm 的塑膠桶中,單本植,每盆 種 9 株。
於插秧當天施基肥,待植株長至 6~7 葉齡施予追肥,11 葉齡時施予穗肥。每 次每盆肥料施用量之氮磷鉀為:0.5 g 尿素、3.5 g 過磷酸鈣及 1.0 g 氯化鉀。植 株從插秧到劍葉抽出時期皆生長於台灣大學校總區農業試驗田之玻璃溫室中。待 植株長出劍葉後移至台灣大學人工氣候室日夜溫 25/20 oC 自然光照玻璃溫室進 行馴化。植株開花期(有 50 %開花)將一半之材料移入 35/30 oC 自然光照玻璃室中 進行 15 天的溫度處理後再移回 25/20 oC。各處理於榖粒充實期中取 6、9、15 及 20 DAF (Days after flowering)及成熟之穎果做為試驗分析材料。脫粒後之穎果丟
11
入液態氮,並存放於-85 oC,待分析。
本試驗共進行四種處理:
1. TNG67 25/20 oC(日/夜溫)常溫處理,標示為 TNG67 25CK。
2. SA1739 25/20 oC(日/夜溫)常溫處理,標示為 SA1739 25CK。
3. TNG67 35/30 oC(日/夜溫)高溫處理 15 天,標示為,TNG67 35HT。
4. SA1739 35/30 oC(日/夜溫)高溫處理 15 天,標示為 SA1739 35HT。
二、葉片葉綠素值測定
以葉綠素計 SPAD-502 Chlorophyll Meter(Minota,Japan),於開花起各處理取 6 植株,每天測量劍葉之葉綠素值(SPAD value)至開花後 30 DAF。以 student t-test 進行顯著性分析。
三、建立穎果生長曲線及發育期外觀標準
取發育中種子 6、9、15、20、25 DAF 及成熟種子,於液態氮下去除內外穎,
拍照建立發育模式圖。並將各發育階段種子稱重紀錄,建立生長曲線。進行 3 重複試驗,並以 student t-test 進行顯著性分析。
四、糙米外觀調查
取各處理之成熟稻穗 3 穗,風乾後脫粒,於光板下觀察糙米外觀及白堊質面 積比例。糙米外觀等級分為完整粒(sound)、白堊質小於 25%(chalky<25%)、白 堊 質 25~49%(chalky < 50%) 、 白 堊 質 50~99%(chalky > 50%) 及 白 堊 質 100%(chlky100%),共 5 等級。
五、穀粒粒型測定
取各處理之糙米穀粒 30 粒,以穀粒判別機(Satake RGQI20,Japan)測量單粒之
12
粒長、粒寬、粒厚及穀粒長寬比,並比較常溫及高溫下之改變比粒。
粒型改變比例={(35oC-25oC)/25oC}×100%
六、穎果 pH 測定
依 Macnicol and Jacobsen (1992)之方法進行,取各試驗處理之糙米樣品 2 粒,放入 2 ml 微量離心管(QSP, USA)並加入一顆直徑 5 mm 鎢鋼珠,以均質機 SH-100 (KURABO,Japan)將樣品磨碎後加入 200μl 蒸餾水,混合後成米漿狀,迅 速以 pH meter(XP-701,SUNTEX,Taiwan)測量。進行 3 重複試驗,並以 student t-test 進行顯著性分析。
七、ATP extraction
依 Napolitano and Shain (2005)之方法修正,取發各發育階段之穎果 3 粒,稱 重後裝於 2 ml 微量離心管並加入一顆直徑 5 mm 鎢鋼珠,於液態氮低溫下以均 質機 SH-100 (KURABO,Japan)將樣品磨碎後加入 500μL 7% perchloric acid (ice cold),混合均勻後至冰上放置 10 分鐘,在 0 oC 下以 13,500rpm 離心 10 分鐘後取 上清液至新管,pellet 再加入 500μL 7% perchloric acid (ice cold)萃取第二次。兩 次萃取之上清液合併後,以 3 M KOH、0.4 M Tris 及 3 M KCl 配成之混合液將 pH 調製 7.75。中和後在 0 oC 下以 13,500 rpm 離心 10 分鐘,取出上清液稀釋 1000 倍放冰上待測定。以 ENLITEN®ATP Assay System 配製 ATP standard 及 rL/L Reagent,取 10μL 稀釋萃取液加入 100μl rL/L Reagent,混和後以 GLOMAX 20/20 LUMINOMETER(Promega,USA)進行測定。空白組以 7% perchloric acid 代 替萃取液。進行 3 重複試驗,以 student t-test 進行顯著性分析。
八、H2O2 extraction
依 Jana and Choudhuri (1982)之方法修正,取發各發育階段之穎果 10 粒,稱
13
重後裝於 2 ml 微量離心管並加入一顆直徑 5 mm 鎢鋼珠,於液態氮低溫下以均 質機 SH-100 (KURABO,Japan)將樣品磨碎後加入 600μl 之 sodium phosphate buffer(50 mM,pH 6.8 內含 1 mM hygroxylamine),混合均勻後在 4 oC 下以 12,000 xg 離 心 20 分 鐘後取上 清液 500 μ l 至新 管 ,加入 200 μ L TiCl4(Titanium Chloride(0.1 %,v/v)溶於 20 %(v/v)H2SO4),混合後以室溫 12,000 xg 離心 15 分鐘,
取上清液以 Specrtophotometer U-200(HITACHI,Japan)測定 A410吸光值。空白組以 sodium phosphate buffer(50 mM,pH 6.8 內含 1 mM hygroxylamine)代替上清液,
以 0.4,0.2,0.1,0.05,0.025 μmole H2O2建立標準曲線。進行 3 重複試驗,以 student t-test 進行顯著性分析。
九、MAD extraction
依 Heath and Packer (1968)之方法修正,取發各發育階段之穎果 5 粒,稱重後 裝於 2 ml 微量離心管並加入一顆直徑 5 mm 鎢鋼珠,於液態氮低溫下以均質機 SH-100 (KURABO,Japan)將樣品磨碎後加入 1.2 mL 之 TCA〔Trichloroacetic acid(5%,v/v)〕混合均勻後在 20 oC 下以 10,000 xg 離心 5 分鐘後取上清液 300μl 至新管,加入 1.2 mL TBA〔Thiobarbituic acid,0.5%(w/v)溶於 TCA(20%,w/v)〕,
混合後以 95 oC 水浴 30 分鐘後立刻插入冰上終止反應,以超音波震盪器去除氣 泡 5 分鐘,常溫下 10,000 xg 離心 10 分鐘後取上清液以 Specrtophotometer U-200(HITACHI,Japan)測定 A532及 A600吸光值。空白組以 300μL TCA(5%,w/v) 代替上清液,以 0.25,0.5,1,1.5,2,2.5,3 μM TEP(1,3,3-Tetraethoxypropane,Sigma) 建立標準曲線。進行 3 重複試驗,以 student t-test 進行顯著性分析。
十、細胞死亡染色
依 Young and Gallie (1999)之方法修正,取個處理試驗中 15、20、25 DAF 之 穎果,於低溫下以解剖刀將穎果以橫向切斷,至入 0.1 % Evans Blue 染色 2 分鐘,
14
再以蒸餾水退染 40 分鐘。以 Nicon D3200 拍照紀錄。
十一、基因表現 (一)RNA Extraction
依 Chomczynski and Sacchi (1987)的方法進行修正。穎果放入 2 ml 微量離心 管(QSP, USA)並加入一顆直徑 5 mm 鎢鋼珠丟入液態氮,以均質機 SH-100 (KURABO,Japan) 1200 rpm 之速度震碎穎果(10 s/次,共 5 次)。加入 1.3 ml TRIZOL Reagent® (Sigma,USA) 萃取 穎果 RNA, 於 Thermomixer comfort (Eppendorf, USA)25 oC,1350 rpm 下反應 30 分鐘。以離心機 Z300k (HERMLE,Germany) 4 oC, 13500 xg 離心 20 分鐘後,吸取上清液。加入 260μl Chloroform (MERCK,Germany) vortex 靜置 10 分鐘後,4 oC,13500 xg 離心 20 分鐘。取上清液 500μl,加入 300 μl high salt solution (1.4 M NaCl & 0.8 M sodium citrate)和 600 μl isopropanol (MERCK,Germany)混勻後置於 4 oC 沉澱 1 小時。4 oC,13500 xg 離心 20 分鐘後去 除上清液,加入 1 ml 70% EtOH 清洗 pellet 2 次。乾燥 pellet 約 10 分鐘後加入 60 μl TE buffer(10 mM Tris-HCl,pH 8.0; 1 mM EDTA,pH 8.0)和直徑 3 mm 小鋼珠,
於均質機下以 1600 rpm 振 2 分鐘,打碎 pellet。再於 Thermomixer comfort 下 25
oC, 1350 rpm 反應 30 分鐘。4 oC, 12000 xg 離心 20 分鐘,取上清。加入 1/6 體積 之 95% EtOH,於冰上反應 20 分鐘。4 oC, 12000 xg 離心 10 分鐘後取上清,加入 DEPC QH2O 使體積達 200μl,再加入 20μl 3 M NaOAC(pH5.0)和 520μl 95%
EtOH,混勻後於-20 oC 沉澱 overnight。4oC,13500 xg 離心 20 分鐘,去上清,以 1 ml 75%EtOH 清洗 pellet。乾燥 pellet 約 10 分鐘後加入 10 mM Tris-HCl(pH 8.0) 回溶 RNA,於冰上反應 5 分鐘再離心 10 分鐘,取出上清液。RNA 經分光光度 計 BioPhotometer(Eppendorf,USA)定量後,取 1μg RNA 於 1X BPET agarose gel 確定 RNA 品質。
15
(二)First-strand cDNA synthesis
依照 Invitrogen SuperScript III Frist Strand Synthesis System for RT-PCR 之 protocol。取 1μg RNA 加入 0.25μl 50μM oligo(dT)20VN(Bioman,Taiwan)、1μl 之 10 mM dNTP mix 補 DEPC QH2O 至 3.25μl。於 65 oC 下反應 5 分鐘後,立刻 將 PCR tube 插在冰上。再加入 1μl 之 5X RT buffer、0.25μl 之 0.1M DTT、0.25 μl 之 RNase OUT(40U/μl, Invitrogen,USA)及 0.25μl 之 SuperScriptIII(200 U/μ l,Invitrogen,USA)混勻,spindown。於 PCR 機器 Veriti 96 well Thermocycler (Applied Biosystems,USA)以 55 oC,60 分鐘合成第一股 cDNA 再以 70 oC,15 分鐘 終止反轉錄酶之作用。
(三)Real Time PCR
將與水稻穎果發育相關之重大生理反應之關鍵基因於 NCBI(National Center for Biotechnology Information)搜尋其 cDNA 序列,並於 KOME(Knowledge-based Oryza Molecular biological)確認其 cDNA 來源。利用 Primer Express 3.0 Software (Applied Biosystem,USA)設計引子(引子資訊如表一)。將設計出的引子由 Bioneer (USA)合成,合成產量 2 O.D.,OPC 純化。依照 KAPA™ SYBR® FAST qPCR Master Mix(2x) Roche LightCycler® 480 protocol 加以修改,以 LightCycler® 480 Multiwell Plate 384(Roche,USA)進行 PCR。7.5ng cDNA、100 nM primer、1x KAPA™ SYBR®
FAST qPCR Master Mix(2x) Roche LightCycler® 480 最後補滅菌二次水至 5μl 後 上 Roche LightCycler® 480 進行 real time PCR。PCR 反應條件為 Pre-incubation(1 cycle):95 oC,3min、Amplification(40 cycles):95oC,10s;60oC,1min(signal)、Melting Curve(1 cycle):95 oC,5s;65oC,1 min;95 oC、Cooling(1 cycle):40 oC,10s。以 eEF1A 作為 internal contril,利用 2-△△Ct計算相對表現量,每個材料的基因均以各自 25 oC 6 DAF 表現量為基準。進行 3 重複試驗,取 2 相近值以 student t-test 進行顯著性 分析。
16
(四)TNG67 與 SA1739 發育穎果的 DNA microarray 分析
萃取 TNG67 與 SA1739 25 CK 與 35 HT 處理過程之 6、9、12 DAF 三重複的 RNA 樣品送華聯公司進行後續分析。透過 Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA)與 Nano Drop spectrophotometer (Thermo Scientific, Waltham, MA, USA) 進行 RNA 品質分析,之後取 1μg RNA 進行 antisense RNA 合成 (OneArray® Amino Allyl aRNA Amplification Kit),Cy5 螢光 標 示 (Amersham Pharmacia, Piscataway, NJ, USA) 與 DNA microarray 雜 合 (RiceOneArray, Phalanx Biotech Group, Taiwan) 。由 AXON4000B 進行雜合訊號 掃描,將掃描圖檔以 GenePix™ 4 軟體輸出成數據檔。Array 的資料使用商業套 裝軟體 Rosetta Resolver System® (Rosetta Biosoftware) 進行 normalization 與顯 著性分析後,取 log2 (倍率) ≥ 1 且 p < 0.05 的顯著性水準的差異結果進行後續的 基因功能分析。針對參與澱粉合成代謝途徑相關基因表現,以 VANTED 軟體進 行繪圖。
17
参、結果與討論 一、 TNG67 與 SA1739 穎果發育之差異
(一)、常溫下 SA1739 發育速率較 TNG67 快速,高溫下更加速穎果之充實速 率,縮短充實時期
常溫下(25 CK),TNG67 穎果發育速率較 SA1739 慢,如圖 1 所示。TNG67 至 15 DAF 才長到成熟穀粒之體積大小,而 SA1739 在 9 DAF~15 DAF 已發 育至成熟穀粒體積大小。TNG67 至 25 DAF 穎果外觀開始呈現透明度,
SA1739 至 20 DAF 穎果已呈現透明外觀。TNG67 至 25 DAF 穎果葉綠素還 存在整顆穎果,SA1739 至 25 DAF 穎果大部分葉綠素已降解,只剩穎果背 部保留些許葉綠素。高溫下(35 HT),加速 TNG67 及 SA1739 穎果發育速率,
仍以 SA1739 生長較快。高溫下 TNG67 至 9~15 DAF 已長到成熟穀粒體積 大小,至 20 DAF 穎果外觀呈現透明度,25 DAF 只剩穎果背部保留些許葉 綠素。SA1739 至 9~15 DAF 已長到成熟穀粒體積大小,15 DAF 穎果已只剩 背部保留些許葉綠素,20 DFA 穎果葉綠素已完全將解。以上顯示經疊氮化 鈉誘變之長粒型 SA1739 在改變穀粒形狀後其穎果發育速率較 TNG67 快,
而兩試驗材料在高溫處理下均促進穎果之發育,縮短充實期間。
(二)TNG67 及 SA1739 穀粒充實期穎果鮮重、乾重、水分含量與劍葉葉綠素 含量變化
圖 2A 的穎果鮮重變化顯示,TNG67 常溫下鮮重累積自 6 DAF 至 25 DAF 持續上升,其中 9~15 DAF 為穎果膨大最快時期。於高溫下其鮮重自 6D AF 到 20 DAF 快速累積,9~15 DAF 鮮重均高於常溫下,20 DAF 鮮重與常溫同,
自 25 DAF 鮮重下降且比常溫低。而常溫下乾重累積與鮮重同步進行,9~15 DAF 有最大物質累積速率,高溫下乾物質累積亦是在 9~15 DAF 高於常溫,
20 DAF 與常溫同,自 25 DAF 下降且比常溫低。高溫下乾物重累積無法與
18
鮮重同步進行,而 9~15 DAF 亦有物質累積最大之速率,至 20 DAF 有最大 乾物重,達生理成熟階段,隨後乾物重下降。高溫下成熟糙米百粒重較常溫 下少 5 mg(常溫下平均百粒重 20 mg,高溫下為 15 mg),成熟種子體積減少(圖 2C & 圖 1)。
對 SA1739 而言,常溫下其鮮重自 6~25 DAF 呈穩定上升趨勢,高溫下之 鮮重自 6~20 DAF 持續上升且高於常溫之表現,在 6~9 DAF 有最大穎果膨大 速率,自 25 DAF 鮮重下降且比常溫低。乾物重於常溫下隨發育持續累積,
15~20 DAF 有最大之物質累積速率,而高溫下亦是 6~20 DAF 持續上升且高 於常溫之表現,且乾物質累積與鮮重同步進行,在 6~9 DAF 有最大物質累 積速率,至 15 DAF 有最大乾物重,達生理成熟階段,自 25 DAF 下降且比 常溫低(圖 2A)。高溫下成熟之糙米鮮重減少 1 mg(常溫平均鮮重 19 mg,高 溫下為 18 mg)(圖 2C)。
穎果水分含量隨穎果發育逐漸下降,高溫下穎果含水率(%)皆較常溫 少,且含水率下降最大時期皆發生於乾物質累積最大速率階段(圖 2B &
2C)。比較本試驗材料之表現,常溫下 TNG67 含水率下降較 SA1739 早,反
觀高溫下 TNG67 含水率下降速率較 SA1739 慢。TNG67 高溫與常溫 6 DAF 含水率相差 11.72 %,而 SA1739 高溫與常溫含水率差異則是 8.1%。另 SA1739 高溫下 20、25 DAF 含水率略有上升趨勢。
若就水分含量(mg/grain)而言,TNG67 常溫下在 15 DAF 前水分含量逐 漸增加,至 15 DAF 有最高水分含量 10.03 mg/grain 隨後水分含量開始下降 至 8.59 mg/grain,但在高溫下 6~9 DAF 水分含量皆高於常溫,高溫下 9 DAF 有最高含水量 13.6 mg/grain 且高於常溫最大含水量,之後快速下降至 7.33 mg/grain。SA1739 常溫下 6~9 DAF 水分含量快速增加,至 15 DAF 有最大 含水量 13.35 mg/grain,之後快速下降至 7.75 mg/grain,而高溫下 6 DAF 有 最大含水量 12.43 mg/grain,之後水分含量逐漸下降至 8 mg/grain,且高溫
19
20 DAF 含水量高於常溫 20 DAF。
前人研究發現穎果發育過程中水分分部由中心向外流失,高溫下穎果中 心早期有水分下降現象但到中晚期穎果中心的水分略較外圍高,此現象是因 心 白 形成 導致 穎 果 中心 結 構 鬆 散使 水 分 分布 於 空隙 間(Ishimaru et al., 2009)。在本試驗中,兩試驗材料於常溫下之表現顯示,TNG67 含水率下降 快且水分含量較少,而 SA1739 在穎果發育中含水率下降較緩慢及有較高水 分含量,另高溫下 6 DAF 含水率下降程度 TNG67 較 SA1739 嚴重。在前人 研究發現,在高日溫及高夜溫在水稻穎果發育早期水分含量有上升之現象,
其中高日溫處理下之最大含水量高於常溫含量,但高夜溫之最大含水量明顯 降低。最大含水量與胚乳容積及粒重增加速度相關,高夜溫最高含水量下降 導致胚乳容積減少與粒重增加速率下降(森田, 2009)。而在玉米子實中發 現,子實最大含水量與子實最終重量呈現正相關(Borrás et al., 2003)。雖然高 溫下 TNG67 早期水分含量增加,表是此時穎果快速膨大,在 9 DAF 之含水 量最高,但穎果粒重增加速率卻較常溫低,表示高溫已對 TNG67 穎果之物 質合成累積造成傷害,導致其最終粒重下降。反觀 SA1739 高溫下 6 DAF 有最高含水量,此時提升粒重增加速度,可能因此有利減緩粒重之損失。
上述顯示 TNG67 與 SA1739 在高溫下皆提升穀粒充實速率,並提早達 到生理成熟階段,但比較常溫與高溫下成熟粒重之差異亦顯示 SA1739 較 TNG67 具耐熱特性。SA1739 先天上具較好之保水能力,且高溫早期含水率 下降幅度較小,可能因此減緩高溫之傷害導致白堊質之形成。此外,最高含 水量與粒重累積速度之同步化對穎果發育具重要性。
由圖 3 的葉綠素值測定結果,TNG67 在穀粒充實期間之葉綠素值在常 溫下自 41 降至 33,高溫下則自 41 降至 30,其中以高溫 20DAF 以後下降幅 度較常溫明顯。SA1739 常溫下,葉綠素值自 37 降至 20,高溫下自 37 降自 22。顯示高溫下會加速 TNG67 劍葉後期老化之速率,而 SA1739 於高溫下
20
相較穩定。但在圖 2A 中可知,高溫下 TNG67 與 SA1739 分別在 20 DAF 及 15 DAF 乾物重累積達最高,但這段期間葉綠素值並無下降,推測高溫造成 兩試驗材料粒重減少可能不是來自劍葉光合作用能力下降。
(三)TNG67 與 SA1739 穎果外觀品質差異
圖 4 及圖 5 顯示,常溫下 TNG67 在穀粒腹部有些許白堊質,完整米粒 僅佔 2.8 %,有 94.7 %穀粒為白堊質面積小於 25 %,白堊質面積大於 25 % 但小於 50 %者佔 2.5 %。高溫下,TNG67 穀粒白堊質嚴重,84 %穀粒為白 堊質面積大於 50 %,且有 7.2 %者為白堊質面積 100 %。另一方面,SA1739 在常溫下,86.4 %之穀粒為完整米粒,13.1 %之榖粒為白堊質面積小於 25
%;高溫下有 77.6 %者為白堊質面積大於 25 %但小於 50 %,且幾乎出現於 穀粒腹部,另有 3 %者為白堊質面積 100 %。由榖粒外觀之比較顯示高溫下 長粒型之 SA1739 亦具有較好之外觀品質。
(四)高溫處理下 TNG67 與 SA1739 穀粒粒型之變化
圖 6 顯示 TNG67 高溫下糙米長度、寬度及厚度分別較常溫減少 2.6 %、
7.1 %及 8.2 %,由於寬度下降比例高而導致高溫下之長寬比增加 4.8 %。反 觀 SA1739 高溫下糙米長度及寬度分別減少 5 %及 1.1 %,而厚度則增加 0.3
%,由於長度下降比例較高導致長寬比減少 4 %。在 Morita 等人 ( 2005)的 研究中,高溫容易導致穀粒寬度及厚度減少,在本試驗結果顯示 TNG67 受 高溫影響較嚴重,整體穀粒體積減少,而 SA1739 之變化相對較穩定。由於 種子內部的氧溶度可由擴散作用及種皮上之葉綠素行光合作用而來。前人之 研究顯示(Rolletschek et al., 2004; Rolletschek et al., 2007),由於種子內部氧溶 度由外部向內部遞減,高溫環境更減少使氧溶度減少,又本研究中 TNG67 穀粒寬度及厚度均較 SA1739 寬厚,如表 2 所示,推測 TNG67 之寬(短)型穀
21
粒內部氧溶度較 SA1739 低,高溫下穀粒內部缺氧程度亦可能較嚴重,因此 高溫下 TNG67 藉由減少粒寬及粒厚,以因應高溫逆境之缺氧衝擊。
二、 高溫下 TNG67 與 SA1739 之生理反應差異及相關基因表現 (一)TNG67 與 SA1739 穀粒酸化程度與酸化相關基因表現
由圖 7 可知 TNG67 在高溫下穎果 pH 均較常溫低,以 9 DAF 時期之 pH 差異最大,常溫下為 6.8、而高溫下為 6.64;至成熟穀粒時其 pH 在常溫為 7.18,高溫下為 7.06。反觀 SA1739 高溫下其穀粒之 pH 變動相對較小,至 成熟時穀粒 pH 常溫為 7.06,高溫為 7.01。此結果顯示 TNG67 在高溫下穎 果內部酸化程度較 SA1739 嚴重。但與 Macnicol and Jacobsen (1992)不同的 是,該研究中大麥隨發育胚乳之 pH 逐漸下降,而本實驗則以整個穎果進行 試驗,並未將胚乳與種皮分離,因此在本實驗結果發現隨發育過程 pH 有上 升之趨勢。
在本實驗室之另一研究發現,處理乙烯釋放劑會增加胚乳酸化程度,高 溫下處理乙烯抑制劑則可減緩穎果酸化(李,2013)。於本研究之乙烯合成基因 表現對照可發現(圖 22),TNG67 高溫下參與乙烯合成 ACC oxidase ,ACO1 於 6 DAF 及 9 DAF 之表現量提升,ACO2 則無;反觀 SA1739 之 ACO1 與 ACO2 在發育前期皆有提升現象,但以 ACO2 較明顯。在水稻穎果穎果中發
現,下位穗 ACO1 與 ACO3 較上位穗表現量高,且隨穎果發育而下降(Zhu et al., 2011)。推測乙烯影響穎果發育與品質,以 ACO1 影響之層面較大,並參 與高溫下穎果早期之酸化作用,使 TNG67 在高溫下之 9DAF 的穎果酸化程 度較 SA1739 嚴重。
穎果酸化之原因,亦可來自於有機酸 malate、citrate 和 lactate 之累積 (Macnicol and Jacobsen, 1992),而有機酸的累積,可能與 TCA cycle 活性有 性。在 MDH(malate dehydrogenase)基因表現結果發現(圖 15),TNG67 其表
22
現常溫下前中期隨發育而下降,15 DAF 後上升,而高溫下之趨勢則是隨發 育期表現量上升,在 6 DAF 表現量較常溫低,9 DAF 其表現量與常溫類似,
15 DAF 開始高過常溫之表現量。將此現象與圖 7 對應可發現,TNG67 高溫 下穎果發育中期(15 DAF)之酸化可能與 MDH 表現逐漸上升有關。反觀 SA1739 高溫下 MDH 表現量受抑制,推測高溫下其 MDH 參與穎果中後期之 酸化反應。
(二)TNG67 與 SA1739 之穎果細胞死亡現象及儲藏性蛋白差異及影響米質形 成之相關基因表現
由圖 8 所示 TNG67 與 SA1739 常溫下之穎果細胞死亡於 25 DAF 才開始 自胚乳細胞發生,而高溫下 TNG67 在 20 DAF 及 25 DAF 穎果胚乳中心均有 明顯細胞死亡及向外擴散現象。SA1739 細胞死亡亦在 20 DAF 及 25 DAF 有明顯現象,不同的是發生部位在穀粒腹部。結果顯示高溫會加速細胞死亡 過程,以 TNG67 危害程度較重,而兩個材料之細胞死亡主要部位與圖 4B 光板下所見白堊質區塊符合。由於白堊質為澱粉粒與蛋白質體堆疊不整齊而 產生空隙,空氣折色後導致穀粒外觀呈現不透明。在(星川, 1968)研究中澱 粉粒的累積方向,水稻澱粉粒自胚乳中心往周圍、由背部往腹部、由頂端往 基部累積,顯示高溫下 TNG67 可能在澱粉累積早期出問題,而在胚乳中心 產生明顯細胞死亡現象,反觀 SA1739 則可能為充實後期由於穀粒背部為維 管束已退化(圖 1)養分無法持續供應,而在腹部容易有細胞死亡。
PBⅠ主要累積 prolamines,PBⅡ主要累積 glutelins,而 PDI 可幫助 prolamines 及 glutelins 之運輸。因此該基因表現量在高溫下受到抑制可能影 響穀粒外觀品質(Takemoto et al., 2002; Lin et al., 2010)。在 PDI 缺失的水稻突 變體中,觀察到穀粒外觀呈現白堊質之現象,且胚乳有不規則的澱粉粒及 PB-I 累積(Kim et al., 2012)。由圖 9 結果可知 PDI 在 TNG67 常溫下隨發育階
23
段表現量下降,而 SA1739 之 PDI 於常溫下則是隨發育階段表現象逐漸上 升。TNG67 常溫下大部分的穀粒外觀就有些許的白堊質(chalky<25 %),而 SA1739 常溫下則以完整粒比例居多,PDI 可能為其外觀差異之原因。TNG67 在高溫下 PDI 之表現於 9 DAF 些微提升,15 DAF 又些微下降,至 20 DFA 期表現量提升。而 SA1739 高溫下其 PDI 之表現量較常溫高且隨發育階段逐 漸提升。但兩試驗材料 Pro 7、Glu 與 19kD Glb 常溫下之表現量皆隨發育而 上升。TNG67 高溫下之表現量亦是隨發育而上升,但自 15 DAF 起表現量受 皆較常溫低。高溫下 SA1739 則是 6~9 DAF 表現量提升,但自 15 DAF 起表 現量皆受抑制。儲藏性蛋白質若無法正確地修飾與摺疊,大量之不成熟蛋白 質累積將會傷害內質網產生 ER stress (Oono et al., 2010)。耐熱之 SA1739 高 溫下提升 PDI 之表現,但 Pro7 之表現量在高溫後期受抑制(圖 13),還需由 測定穎果儲藏性蛋白含量來證實。由以上可推測 PDI 之差異表現為導致 TNG67 與 SA1739 外觀品質差異之因子之一。
PBZ(Probenazole-induced protein)被認為是水稻自體防禦的分子標誌,
PBZ1 參與水稻生物逆境反應,如稻熱病及白葉枯病;非生物逆境,如低溫,
並受植物荷爾蒙的調控,GA 及 ABA 皆會誘導 PBZ1 之表現。病變突變體 (lesion mimic mutant, LMMs)即使在無病原菌感染情況下,植株體內依然有 高度敏感反應(hypersensetive response, HR),常被做為研究 PCD 的材料 (Lorrain et al., 2003)。在水稻病變突變體,spotted leaf 1 (spl1)裡發現細胞死 亡指標 PBZ1 與 PCD 有高度相關性,PBZ1 高度存在於 PCD 之組織內。水 稻根經淹水逆境缺氧後,容易產生通氣組織(aerenchyma),並且誘導 PBZ1 表現,因此 PBZ1 可以做為細胞死亡之指標。而其作用機制可能藉由細胞內 之 RNase 活性導致細胞死亡(Kim et al., 2008; Kim et al., 2011)。在 TNG67 穎 果在高溫下 PBZ 之表現量在發育中後期大量誘導且表現程度較 SA1739 高,
對照其高溫下米質外觀與細胞死亡結果,可更加確認 TNG67 在高溫下穎果
24
發育之細胞死亡程度較嚴重,另亦暗示 TNG67 在高溫下缺氧之程度較 SA1739 嚴重(圖 10)。
另外做為細胞死亡指標之 DNAase(Ca2+ dependant-endonuclease),常溫 下 TNG67 隨發育而上升,SA1739 則是 6~9 DAF 上升,隨後下降。高溫下 TNG67 僅在 9 DAF 表現量略較常溫高,其餘皆受到抑;SA1739 則是在高溫 6~9 DAF 表現提升,15~20 DAF 則受到抑制(圖 10)。本研究中 DNAase 在高 溫逆境下表現的角色,可能還需要由 DNA fragmentation 之試驗進一步證明。
(三)TNG67 與 SA1739 穎果 ATP 濃度差異與能量調控相關基因表現
由圖 11 所示常溫下 TNG67 在 15 DAF 其 ATP 濃度達最高,之後隨發育 而下降;高溫下 6 DAF 含量最高,之後隨發育階段而下降。反觀 SA1739 在常溫下 6~9 DAF 有最大 ATP 濃度,之後隨發育階段而下降;高溫下則在 6 DAF 有最高 ATP 濃度,9 DAF 時還可以維持與常溫的水平,至 15 DAF 濃度急速下降。兩試驗材料比較下,SA1739 較 TNG67 有較高之 ATP 濃度。
She 等人 (2010) 研究發現耐高溫之品種 Kinmaze,高溫下粒重減少程度 低,且穀粒大小及粒型變化小,其發育階段穎果內部 ATP 含量常溫下 10 DAF 達最高峰,而在高溫 10 DAF 亦可維持常溫下之表現量,反觀在 flo2 mutant EM37 在高溫下粒重減少嚴重,其高溫下雖 ATP 下降緩慢,但於高溫下 7DAF 的 ATP 含量無法維持常溫下之狀態,可見 ATP 含量之維持與耐高溫有關。
而在本試驗材料的 SA1739 粒重變化與 ATP 表現模式,可符合 She 等人 (2010)的結果。
另在 She 等人 (2012)的試驗材料其大部分發育中水稻穎果 ATP 濃度有 2 個高峰期,而高溫下會使第 2 個高峰提前表現像皆較常溫低,且高溫與常 溫第 2 高峰之 ATP 濃度比與高溫下完整米粒有正相關。在本試驗材料中,
皆只有一個高峰期,且高溫下亦有提前表現之現象。比較高溫下與常溫高峰
25
期 ATP 濃度之比例,TNG67 僅 79 %而 SA1739 則為 107 %,對應高溫下完 整米粒比率,TNG67 高溫下無完整米粒,而 SA1739 尚有 2.6 %之完整米粒。
若以完整米粒加 Chalky < 25 %之比例,高溫下 TNG67 為 0 %,而 SA1739 有 80%。
另由圖 2C 已得知 TNG67 高溫下 6~9 DAF 為乾物重累積速率最快時 期,但此時其穎果內 ATP 濃度已下降,至 20 DAF 有最大乾物重,此時 ATP 已達最低濃度。SA1739 高溫下 6~9 DAF 為乾重累積最快時期,穎果內部有 最高之 ATP 濃度,至 15DAF 有最大乾物重,ATP 濃度也已迅速下降。
由以上結果顯示,SA1739 不僅在高溫下可以維持 ATP 濃度高峰期之時 間點亦可減少與常溫 ATP 高峰之濃度差異,並且能在穎果有最大 ATP 濃度 時大量累積乾物質,顯示 SA1739 可以有效地利用能量,避免能量之浪費。
穀粒內大分子物質之合成皆需消耗能量,澱粉合成相關基因 SSⅡa 及 GBSS,在 TNG67 及 SA1739 兩個試驗材料中,常溫下隨發育階段表現量逐
漸增加,於 15 DAF 大量表現,但 SA1739 表現量在 15 DAF 達高峰後下降(圖 12)。由先前兩試驗材料之 ATP 含量(圖 9)及乾物重累積(圖 2A)可發現,澱
粉合成大量表現與乾物重持續累積,但此時穎果內部 ATP 濃度皆處於較低 之狀態,表示常溫下此生育時期之澱粉合成所需之能量消耗較低。
觀察兩試驗材料高溫下澱粉合成與儲藏性蛋白質基因表現(圖 13),在高 溫下皆受到抑制,由其以 TNG67 抑制較嚴重。TNG67 及 SA1739 在高溫下 Porlamine 7、Glutaline、19kD Globulin 在穀粒充實早期表現量提升,但在 15 DAF 之後表現量下降,其中以 SA1739 高溫下抑制量較多。水稻穎果有 80%以上組成分為澱粉,故澱粉為影響粒重主要之因素,對照基因表現與成 熟穀粒重(圖 2C)之結果,推測 TNG67 高溫下粒重下降之主要原因來自於澱 粉合成基因抑制所導致。
在 Micorarray 的結果裡(圖 25),針對澱粉合成途徑中,高溫下誘導
26
SA1739 的 sucrose synthase(Os04g0309600 及 Os030g401300)、ADP-glucose transporter(Os05g0171300)、ADPase(Os05g0580000 及 Os07g0243200)、starch branching enzyme(Os06g0726400)、soluble starch synthase(Os06g0160700)。
在澱粉降解層面,前人研究裡發現高溫逆境誘導水稻穎果內 Amy1A, Amy1C, Amy3A, Amy3D 及 Amy3E 的表現(Hakata et al., 2012)。而在本試驗結果裡高
溫下也誘導了兩試驗材料的 Amy3D 及 Amy3E,但 TNG67 在 9 DAF 誘導 Amy3D;SA1739 則到 12 DAF 才誘導 Amy3D 表現。兩試驗材料的 Amy3E
在高溫下 6、9 DAF 皆被誘導,但 SA1739 到 12 DAF 其表現量受到抑制。
綜觀澱粉之合成代謝表現,SA1739 在高溫下較 TNG67 有利於進行澱粉合 成。
生物的能量代可由(1)有氧呼吸:糖解作用進入檸檬酸循環再由粒線體 之 oxidative phosporylation 產生 ATP;(2)無氧呼吸:由糖解作用最後之產物 pyruvate 走向酒精發酵途徑產生較少量之 ATP。
TNG67 常溫下參與糖解作用之 PFK 在 6~15 DAF 表現量上升隨後下 降,而 G3PDH 則是隨發育天數持續上升,PK 在 6~9 DAF 下降隨後又上升 (圖 14)。但在檸檬酸循環之 IDH 在 6~9 DAF 下降隨後上升,而 SUSB 與 MDH 則隨發育天數表現量下降(圖 15),參與 oxidative phosphorylation 之 ATPSB 在 15 DAF 有些微上升,但整體趨勢是隨發育天數下降,而參與無氧呼吸之 ADH 則是自 9 DAF~15 DAF 表現量上升(圖 16)。由以上現象可發現,雖然
糖解作用在發育中後期還持續表現,但由 PK 所產生之 pyruvate 在穎果發育 中並無法走進檸檬循環進而走向 oxidative phosphorylation,取而代之是走入 無氧呼吸之途徑,表示 TNG67 常溫下穎果發育過程是走向低氧之過程,與 前人研究結果是相同(Xu et al., 2008)。
SA1739 常溫下參與糖解作用之 PFK 在 6~9 DAF 表現量些微下降,自 15DAF 表現量達最高峰,隨後下降,而 G3PDH 與 PK 則是 6~15 DAF 持續
27
上升隨後下降(圖 14)。在檸檬酸循環之 IDH 隨發育天數持續上升,而 SUSB 與 MDH 則在 15DAF 表現量下降(圖 15),參與 oxidative phosphorylation 之 ATPSB 在 15 DAF 後下降,而參與無氧呼吸之 ADH 則是自 9 DAF~15 DAF
表現量上升(圖 16)。由以上現象可知 SA1739 常溫下自 15 DAF 以前可藉由 糖解作用、檸檬酸循環與 oxidative phosphorylayion 產生能量,15 DAF 以後 則是藉由無氧呼吸維持後續之能量。
由於在大麥、玉米種子內發現氧濃度越靠近種子內部,氧濃度逐漸下降 (Rolletschek et al., 2004; Rolletschek et al., 2005)。常溫下兩試驗材料能量之走 向可推測,SA1739 比起 TNG67 可維持穎果內發育前 15 天有較多之氧含量,
可能因 SA1739 有較細長之粒型以利氧氣之擴散有關,但還需進一步的驗證 使得知。另對照先前澱粉合成基因表現、ATP 濃度與乾物重累積結果,澱粉 合成除可需較少之能量也能適應低氧之環境。
TNG67 高溫下參與糖解作用之 PFK 趨勢與常溫同,但在發育早期表現 量較常溫提升,到 15、20 DAF 表現量受抑制。而高溫下 G3PDH 表現量都 受到抑制,PK 則只在 9 DAF 表現量較常溫高,但趨勢是 6~15 DAF 隨發育 而提升,到 20 DAF 開始下降。而參與檸檬酸循環之 IDH、SUSB 及 MDH 高溫下的表現量接受到抑制,而 ATPSB 亦是受到抑制,反觀在參與無氧呼 吸的 ADH 高溫下表現量提升,且隨發育階段上昇,這表示糖解作用最後的 產物 pyruvate 在高溫下趨向走入無氧呼吸,表示 TNG67 高溫下穎果內部有 缺氧之現象。
SA1739 高溫下參與糖解作用之 PFK、G3PDH 及 PK 接受受到抑制,而 檸檬酸循環中的 IDH、SUSB 及 MDH 亦受到抑制,而 ATPSB 亦受到抑制,
但 ADH 卻大量表現,表示高溫下 SA1739 穎果內部亦是缺氧之現象,且依 靠無氧呼吸維持穎果內部所需之能量。
值得注意的是,PK 是將 PEP 轉為 pyruvate,pyruvate 經由 pyruvate
28
decarboxylase 轉為 acetoaldehyde 再經由 ADH 轉為 ethenol。但 SA1739 高溫 下的 PK 相較常溫下之表現受到抑制,進入無氧呼吸所需 pyruvate 由此推定 應不是透過 PK 所提供。PPDK (pyruvate phosphate dikinase)是可以扮演 PK 之功能將 PEP 轉為 pyruvate,產生 ATP;亦可將 pyruvate 轉為 PEP,消耗 ATP 和 Pi 產生 AMP 及 PPi。隨著水稻穎果發育,PPDK 表現量下降(Chastain et al., 2006; Xu et al., 2008),在穎果發育早期的調控是傾向將 PEP 轉為 PK(Chastain et al., 2006)。水稻有 3 種類型之 PPDK 基因,分別為在細胞質 內(cytosolic type)的 OsPPDKA 和 cyOsPPDKB,在葉綠體內之 chOsPPDKB。
在 T-DNA 插入造成 OsPPDKB 缺陷之水稻突變株,穀粒外觀呈現心白之現 象(Kang et al., 2005),表示 PPDK 扮演著胚乳內物質累積之調控角色。細胞 質內之 PPDK 調控代謝合成途徑:(1)將 pyruvate 轉為 PEP,在細胞質中產 生之 PPi 為誘導澱粉合成中之 sucrose sythase 之訊號,有利澱粉之合成;(2) 由於 pyruvate 為碳、氮代謝與脂肪合成之轉折點,PPDK 若將 PEP 轉為 pyruvate 增加細胞質內之 pyruvte 含量,有利於使之轉為有機酸之合成,如 Ala 及 Glu,或是走向於脂肪酸之合成。(3)PPDK 會由低氧所誘導(Moons et al., 1998),種子發育過程中是低氧之環境(Rolletschek et al., 2004),PPDK 若走 向將 PEP 轉為 pyruvate 之途徑可以產生 ATP,將使缺氧之組織有利產生能 量。
在 DNA microarray 的結果發現,高溫下誘導 SA1739 在 6 DAF 的 OsPPDKB 表現(圖 25)。在水稻 OsPPDKB 突變體裡,沒有 OsPPDKB 轉譯
及 OsPPDKB 蛋白質,但伴隨有外觀呈現心白、粒重下降、蛋白質及脂肪含 量提高之現象,表示 OsPPDKB 的缺失使 pruvate 走向脂肪的合成(Kang et al., 2005)。在不耐熱的水稻品種裡,高溫下抑制 OsPPDKB 的表現,外觀有白 堊質 現象(Yamakawa et al., 2007)。而本試驗中高溫下誘導 SA1739 的 OsPPDKB 表現,推測此現象將有利於 pruvate 走向 PEP,產生 PPi 做為誘導
29