國立臺灣大學生物資源暨農學院 森林環境暨資源學研究所
碩士論文
School of Forestry and Resource Conservation
College of Bio-Resources & Agriculture
National Taiwan University Master Thesis
淡黃木層孔菌在臺灣中部及北部與環境暨生物因子之研究 Study on the Relationship between Phellinus gilvus
and the Environmental and Biotic Factors in Central and Northern Taiwan
陳雅得 Ya-te Chen
指導教授:李靜峯 助理教授 Advisor: Ching-Feng Li, Ph.D.
共同指導:張東柱 博士(林業試驗所) Co-advisor: Tun-Tfschu Chang, Ph.D. (TFRI)
中華民國一 O 八年二月
誌謝
很幸運的能遇到五木老師,如明燈一樣讓我逐漸看清研究之路。還有 David 與 VegLab 的大家對植群調查的執著、嚴謹的態度,以及開放的討論風氣,也都深 深折服我。在學業後期,我覺得最難的不是研究本身,而是記得自己的初心。每 當被工作淹沒,我就會想想你們,重新校正方向。
這份研究源自於林業試驗所的張東柱與傅春旭老師,你們帶我入門,而你們 對樹病與真菌持續的熱情始終激勵著我。謝謝 Dora、阿暉,讓我下定決心讀研究 所,跟你們總能聊出許多靈感。謝謝芳孜,陪我上山做 ARBOTOM 探測老樹腐朽,
雖然最後沒放入論文,卻是兩年中最有趣的經驗之一。謝謝林試所實驗室的大家,
以及中研院的朱宇敏老師與師母,在你們那兒做真菌鑑定總是讓我沉迷忘食,彷 彿在迷宮中時日飛逝,直到好像終於能說自己對真菌開始有一點點了解。
感謝邱春火老師,不僅供應我經費,也不吝與我長篇討論,讓我更了解各種 分析方法的得與失。謝謝在植物標本鑑定上大力幫忙的建帆學長跟信彥,以及幫 我畫地圖的政倫。感謝 DA,不僅是我的 coding 外掛,也一如往常地老是批評到讓 我想放棄,然後不得不更深入審視自以為已經搞定的東西。
當初追著樹世無常,來到了這裡,想深入一窺大樹森林的奧秘。沒想到在最 後找到的卻在意料之外。跟大樹們一樣,我們也面臨許多無常與逆境,然後繼續 堅定而溫柔的站立。但願你們也如此,永遠青翠。
2
摘要
心材腐朽菌在森林動態中扮演重要的角色,但在臺灣相關的研究稀少。本論 文的目的為釐清影響臺灣中海拔常見的心材腐朽菌 -- 淡黃木層孔菌(Phellinus gilvus)生態分布的重要因子:藉由設計快速的廣泛取樣調查,來取得淡黃木層孔菌 在臺灣北部與中部天然林中分布地點或未發現地點的生物變數(寄主樹木與森林植 群資料)與環境變數(氣候資料),並量化分析,找出影響它們分布的最關鍵變數。
快速調查方法,係以「臺灣現生天然植群圖集」中所定義之植群群系為主要 分層取樣依據,著重於取得現地的森林與樹木資料,並加上「區域」這一變數,
以分析各項變數是否具跨區域的影響力。經實踐後,確認本調查方法確實可達到 快速取樣、建立特定物種分布資料庫的目的。所獲得之資料為兼顧「發現」與「未 發現」之高度不平衡物種分布數據。後續分析則顯示,加入生物因子建立的分布 模型,比僅使用環境因子的模型更為準確、簡潔、合理且易詮釋,可見生物因子 在決定物種生態分布上之重要性,建議物種分布研究應將生物因子納入。
分析後顯示影響淡黃木層孔菌分布最重要的變數為:平均樹木胸徑大、植群 中殼斗科所佔比例高、以及氣溫季節性變化相對小的天然林中。符合此環境條件 之植群群系為山地常綠闊葉林,其次為空間上相鄰的下部山地次生常綠闊葉林、
山地針闊葉混淆林、山地常綠闊葉矮林、山地針葉林與山地針闊葉次生混淆林。
淡黃木層孔菌應為常見於雲霧盛行地區老熟殼斗科天然林的物種。
本研究的結果,為後續研究心材腐朽菌如何影響森林生態不可或缺的一步,
所發展之快速調查方法亦可調整應用於其他物種分布之研究中。
關鍵字:淡黃木層孔菌,心材腐朽菌,快速調查,物種分布,植群
3
Abstract
Heartwood decay fungi play important roles in forest dynamics. However, the study on their biology, physiology, and ecological roles in forest regeneration is very limited in Taiwan. The objective of this study is to identify the key factors that determine the ecological distribution of a heartwood decay fungus, Phellinus gilvus, often found in mid-altitudinal natural forest in Taiwan. With the design and
employment of a quick stratified sampling method based on Actual Vegetation Maps of Taiwan, data of the occurrences and absence of P. gilvus across natural forest in
northern and central Taiwan was obtained, including biological variables concerning the host trees and host forest, as well as environmental variables concerning the habitat and climate. The quick survey method was proven to be efficient in acquiring samples to build a dataset of a certain species for the analysis of its distribution. The analysis of the dataset shows that P. gilvus generally occurs in locations with relatively little
temperature fluctuation and in forest with a greater proportion of Fagaceae trees. All of the fruitbodies occur on trees of the family Fagaceae with large diameters of breast height. Montane evergreen broadleaf forest is the most suitable vegetation type for P.
gilvus, while occurrences were also recorded scattering in other vegetation types with similar climatic conditions and large dominant Fagaceae trees. The results of this study provide important base for future studies on the ecological roles of heartwood decay fungi in forest. Moreover, the quick survey method may be applied to the study of distribution of other species.
Keywords: Phellinus gilvus, heartwood decay fungi, quick survey, species distribution, vegetation types
4
目錄
誌謝……….……….………....1
摘要……….……….………....2
Abstract ………..…..3
目錄 ..……….………….………...4
圖目錄 ……….………...…………..…...7
表目錄 ……….………...8
第一章 前言 ……….……….…….…..9
第二章 文獻回顧 ………..……….…………10
2.1 物種分布之理論基礎………...………...…10
2.2 影響心材腐朽菌分布的因子………...……….….….11
2.2.1 植群類型……….…………...….….12
2.2.2 氣候條件………...…..….12
2.2.3 森林結構多樣性、棲地連續性…...………....…….12
2.2.4 真菌生存策略與寄主範圍…...………..………….13
2.3 淡黃木層孔菌的生理與生態………….……….………...…………...….14
2.3.1 生態定位:弱病原性之初級心材腐朽菌………….………...…….14
2.3.2 巨觀與微觀型態……….……….…….….……….…..15
2.3.3 營養模式與生長環境…………...……….………..….15
2.3.4 繁殖與傳播………….……….………...…….……….…16
2.3.5 生命史………….………. …….………...………....17
2.3.6 淡黃木層孔菌在臺灣之研究現況……….……….…...17
2.3.7 小結……….……….……….…...18
5
2.4 研究尺度……….……….……….…………..19
2.5 調查方法論:生物分布取樣方法……….……….……19
第三章 研究方法……….22
3.1 快速調查方法……….………...….22
3.1.1 調查範圍……….………...…...22
3.1.2 調查分層單位……….………...…...22
3.1.3 變數選擇……….………...………...24
3.1.4 取樣方式……….………...………...25
3.1.5 調查結果………...……….……….…..26
3.2 資料分析方法……….……….………...……28
3.2.1 分類樹法……….……….………...…. 29
3.2.2 以隨機森林分析自變數重要值……….………. .31
3.2.3 分類樹模型之評價………….……….………..32
第四章 資料分析結果………...……….………..…34
4.1 各項變數之初步分析……….……….…….……..34
4.1.1 生物變數……….…………...……….…….….34
4.1.2 環境變數……….………..40
4.1.3 變數間的相關性………...………...….42
4.2 單一分類樹模型分析.……….……….……….….…44
4.2.1 自變數篩選……….……….…………...………. 45
4.2.2 「生物與環境自變數」組………...….………46
4.2.3 「環境自變數」組……….……….………..……48
4.3 以重複隨機子取樣驗證法比較兩組分類樹之效用……….………....49 4.3.1 跨區(全部資料)與分區(中臺灣)分析中兩組模型之準確率比較.…50
6
4.3.2 影響淡黃木層孔菌分布的關鍵自變數分析….………..……52
4.3.3 模型樹之簡潔度之比較….……….………...…..54
4.4 小結….……….……….……….….………55
第五章 總結與建議 ……….……….….……56
參考文獻 ……….……….……….60
附錄一 蘇鴻傑所劃分之全台地理氣候區。………..………...…...68
附錄二「臺灣現生天然植群圖集」各植群群系基本資料。………..69
附錄三 調查地點與時間。………...………....72
附錄四 樹種名錄。………..………...73
附錄五 重複隨機子取樣驗證法之 R 語言 code。……..………..…....78
附錄六 淡黃木層孔菌之標本採集、鑑定照片範例。…..………...82
7
圖目錄
圖 1 淡黃木層孔菌在臺灣已知「發現」與「未發現」地點總覽. ………..28
圖 2 淡黃木層孔菌在各植群群系中發現之頻度. ……….………..35
圖 3 淡黃木層孔菌寄主樹種圓餅圖. ……….………..37
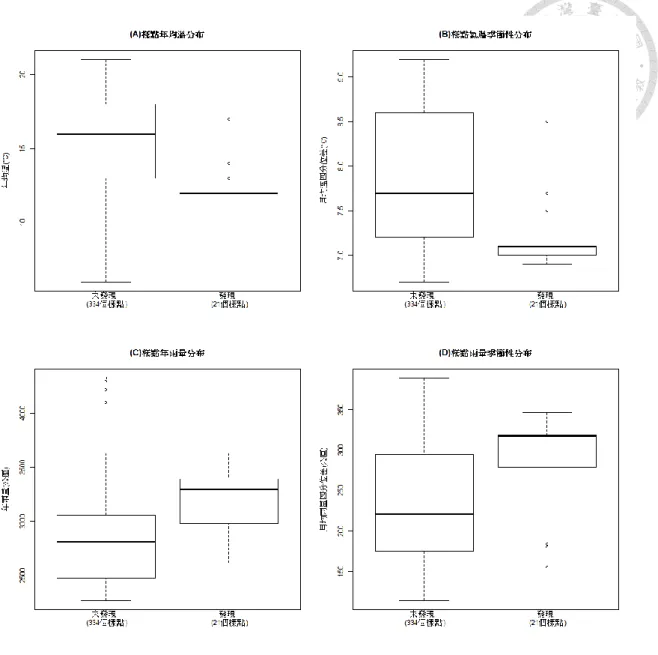
圖 4 樣點樹均胸徑分布盒鬚圖. ………..………….38
圖 5 樣點森林冠層高度分布盒鬚圖. ………..……….39
圖 6 所有樣點的海拔分布. ………...…………...…..41
圖 7 淡黃木層孔菌之有無與氣候因子的關係盒鬚圖. ………...……..42
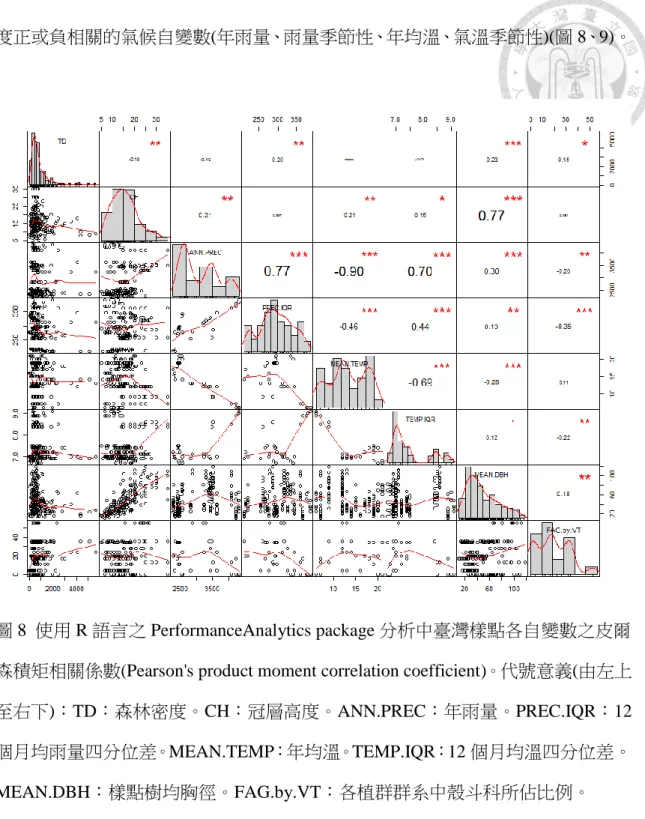
圖 8 中台灣樣點各自變數間之相關性. ……….……….….43
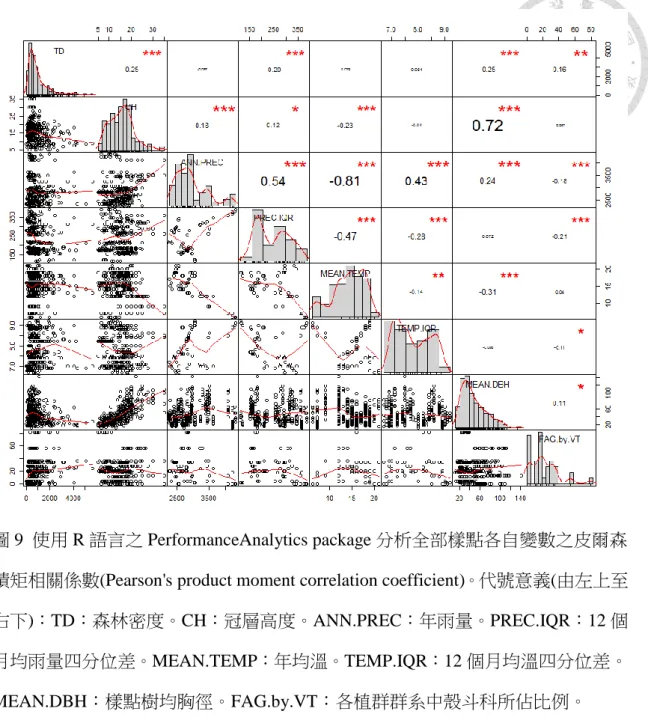
圖 9 全部樣點各自變數間之相關性. ……….…………..…44
圖 10 以隨機森林分析各自變數的平均準確率降低值. ……….………45
圖 11 以完整調查資料庫建立的分類樹, 使用生物與環境自變數. ……….….48
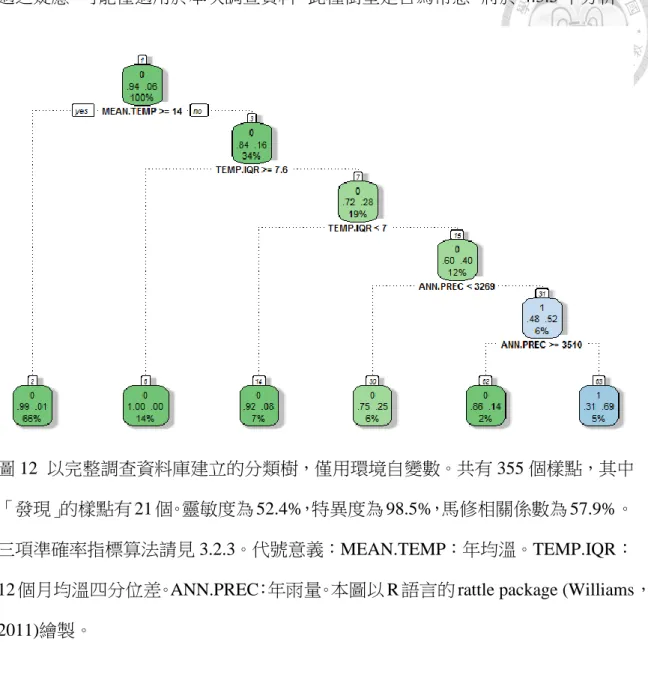
圖 12 以完整調查資料庫建立的分類樹, 僅用環境自變數. ………..49
圖 13 全部資料的 1,000 組訓練子集與測試子集之準確率分布. ……….…..50
圖 14 中台灣資料的 1,000 組訓練子集與測試子集之準確率分布. …………...……51
8
表目錄
表 1 生物變數. ………..….23
表 2 環境變數. ………..….…24
表 3 取樣總數. ………...27
表 4 混淆矩陣(confusion matrix). ……….………...….33
表 5 用隨機森林計算各自變數相對於依變數(植群群系)的重要值. ………36
表 6 重要自變數之獨立雙樣本 T 檢定結果. ………..…….39
表 7 跨區與分區模型準確率對照表. ………..…….52
表 8 在總共 4,000 棵分類樹中, 各自變數用於模型建立的次數統計. ………….…54
表 9 在總共 4,000 棵分類樹中, 各自變數用於分割點的總次數統計. ………..55
9
第一章 前言
心材腐朽菌在森林動態中扮演重要的角色(Boddy & Heilmann-Clausen,2008)。
它們為具初級木材分解能力的真菌,能先驅腐生於莖幹或根部的心材。其中有些 為純粹的分解者,可與樹長久並存,也可促成樹木傾倒或逐步分解,時程與規模 不一。也有些兼具病原性,可能使樹木致病死亡。數種心材腐朽菌已證實為森林 更新的推手,而實際影響為何,不僅受其生物學影響,也與分布模式有極大關係 (Hansen & Goheen,2000;Hawkins & Henkel,2011;Heilmann-Clausen & Boddy,
2008;Hennon,1995;Holah,1992;Holah,1997)。然而,這類真菌的分布情況,
在臺灣目前僅有各地菌相普查與零星觀察報告,並無系統性的調查,導致任何進 一步的生態學研究皆無法進行。
本研究的目的即在釐清影響臺灣中海拔森林常見的心材腐朽菌 -- 淡黃木層 孔菌(Phellinus gilvus)生態分布的重要因子。淡黃木層孔菌在臺灣分布廣泛,但從 未有過系統性的調查,影響其分布的因子亦僅止於推測,尚無量化分析。本研究 設計快速、廣泛的取樣調查方法,來取得淡黃木層孔菌在北臺灣與中臺灣天然林 中的分布地點與棲地資料,用以分析影響它分布的關鍵因子,同時評估此調查方 法的效用。此為後續研究腐朽菌如何影響森林生態不可或缺的一步,而所發展的 研究方法亦可應用於其他生理與生態特性類似的物種具時效性的調查中。
具體的目標有三:
(1) 設計一可快速取得物種分布位置資料的調查方法。
(2) 分析調查資料,找出影響淡黃木層孔菌生態分布的關鍵因子。
(3) 建立分類樹模型並驗證之,以評估調查方法的效用。
10
第二章 文獻回顧
2.1 物種分布之理論基礎
物種的「分布」,為一具多層意義的概念。除了指地理上的分布,也指生態 上的空間:該物種在生態系中的角色與地位,由複雜的種間關係,例如競爭、共 生等關係所塑造而成。此外,由於物種群落與環境條件皆具消長動態,因此許多 分布理論會加上時間因子,例如族群數量動態、族群對所處生態系持續的回饋作 用等等。每一種生物或環境條件都是一項限制因子(limiting factors),一方面反應出 環境提供該物種利用的資源幅度,另一方面也對應到該物種本身生理與生態學上 的耐性範圍(tolerant range)。多個條件維度集合的結果,使每個物種都佔有獨特的 空間(Hutchinson,1957;Guisan,2005;Franklin,2009;Holt,2009)。
物種的生態分布研究,一般指藉量化分析已知發現或為發現該物種之地點的 資料,定義出生態分布空間,亦即找出決定該物種分布的因子及其梯度(Guisan,
2005;Franklin,2009)。物種理論上可生存的空間,與現實中物種生存的空間,兩 者有所落差,因前者代表物種潛在的分布範圍,但還需通過多重濾網後(filters)才 形成今日所見的實際分布狀態,這些濾網包括歷史因子,如因傳播限制而沒有機 會到達適合但隔絕的環境中,或因擾動而局部滅絕;也包括生物因子,如捕食或 病原的傳播,使族群的發展受限(Hutchinson,1957;Franklin,2009;Lambers,2008)。
不考慮時間因子的物種分布研究,需建立在一重要假設上:調查中所發現之 本物種分布情形,皆已與其生態環境達成一定平衡,是物種特性、適應力與該地
11
資源、環境條件互相符合的結果,可在其中生存並繁殖(Guisan,2005)。因為若考 慮時間因子1,則有例外:例如,今日所見的族群並非存在於適合的環境中,有可 能正在遷徙或逐漸滅絕;阿利效應(allee effect)可能使密度夠大,或傳播力夠強的 族群,在適合環境之外生存下來;物種繁殖單位的源匯動態(source-sink dynamics) 也會跨越適合分布之區塊界線。這些狀況需以更多假設或模型來涵蓋與解釋 (Guisan,2005;Franklin,2009)。
本文研究淡黃木層孔菌的分布,採用生態上的分布概念與上述之平衡假設,
僅探討與目標物種分布相關的環境與生物因子,不探討地理上的分布,且不考慮 時間維度與動態變化。
2.2 影響心材腐朽菌分布的因子
心材腐朽菌屬木棲性擔子菌(wood-inhabiting basidiomycetes),它們為具初級木 材分解能力的真菌,能先驅腐生於莖幹或根部的心材。若干研究分析了區域尺度 下多項因子對真菌種類分布的影響,發現最顯著相關者為植群類型、氣候條件、
棲地結構與連續性,以及真菌物種本身的生存策略與寄主範圍。而目前仍無適當 解釋的分布現象,則可能必須追溯到過往歷史或尚未深入研究的環境濾網
(Heilmann-Clausen & Boddy,2008;Schwarze et al,2000;Heilmann-Clausen et al, 2014;Abrego, Bassler, Christensen, & Heilmann-Clausen,2015;Abrego, Bassler, Christensen, Ainsworth, & Heilmann-Clausen,2016)。
1另一個加入時間觀點的重要領域為演化與系統發生學中的物種分布研究,涉及物種的定義、族群 遺傳學等,在本論文中不加以討論。
12
2.2.1 植群類型
由於植物為其能量來源,森林植群類型的分布與木棲性真菌的分布緊密相關 (Heilmann-Clausen & Boddy,2008; Yamashita, Hattori, & Abe,2010; Stokland,2012),
此現象在或大或小的研究尺度上皆明顯。例如,橫跨北美與歐亞大陸的北方針葉 林(boreal forest)中,有超過 80%相同的木材腐朽菌物種(Gilbertson,1980);歐美與 東亞樹種組成相近的溫帶落葉林,地中海、東非與北美南部的亞熱帶森林,皆有 相對應的木棲性真菌,隨之跨洲分布(Heilmann-Clausen & Boddy,2008)。木棲性 真菌與森林類型高度相關,一方面反映出所適應之氣候與環境因子的相似性,另 一方面也代表著共同演化的歷史,例如北方針葉林中的樹種相對較少,在過去百 萬年以來的變化也較少,多數木棲性真菌已適應該地長而寒冷的冬天、大陸性氣 候與降水特性,已無法在較溫暖的區域生存(Heilmann-Clausen & Boddy,2008)。
2.2.2 氣候條件
氣候對植群分布有極大的影響,但對真菌的分布是否同等重要,則尚未有詳 細的研究。有些真菌分布的現象受地域性氣候條件,尤其是降水與氣溫的影響相 當顯著(Gilbertson,1980; Heilmann-Clausen,2005;Heilmann-Clausen,2014;Abrego et al,2016)。一些真菌在較易對寄主造成逆境的氣候帶分布較廣、數量也較多。
氣候變遷與暖化的現象,也已反映在真菌分布範圍與物候的改變上 (Heilmann-Clausen & Boddy,2008)。
2.2.3 森林結構多樣性、棲地連續性
13
森林的組成因子中,對木棲性真菌分布有重大影響者,除了植群組成,其林 齡結構與棲地連續性也相當重要,而這些與該地的人類活動常有密切關係。已有 許多研究比較了人為干擾程度與真菌多樣性的關係(Pentilla, Lindgren, Miettinen, Rita, & Hanski,2006)。在天然林中,由於樹種較多、樹齡不一,棲地多樣性較高,
因此真菌組成也較多樣(Schmit, Mueller, Leacock, Mata, Wu, & Huang,2005);而人 為干擾多的森林,通常平均樹齡較小,樹圍較小,砍伐傷口與中小型落枝吸引較 多先驅菌種,但寄生或腐生於老樹心材與巨大倒木的真菌種類,幾乎只能在足夠 老齡的天然林中才找得到(Heilmann-Clausen & Boddy,2008)。
棲地的限縮與破碎化,是人為造成的另一個重要影響,對孢子不適合長途傳 播的真菌種類尤其有害。儘管孢子傳播距離與棲地破碎化的關係尚待研究,但一 般認為,族群數目較少、生長於不易受擾動影響之棲地、孢子傳播力弱或孢子數 目變化大、對棲地類型或寄主高度專一者,棲地的不連續性對他們的影響甚巨;
而相對的,原本就僅需小塊(例如小斷枝)、易受破壞(例如易燃樹種)的棲地之木棲 性真菌,孢子的傳播力較強,棲地的破碎化可能反而提高其競爭力(Stokland,2001;
Abrego,2015)。棲地破碎化若與森林結構單一化同時發生,則可對高度專一的真 菌物種產生加乘的傷害(Pentilla et al,2006;Heilmann-Clausen & Boddy,2008)。
2.2.4 真菌生存策略與寄主範圍
在上述環境背景下,不同的木棲性真菌有不同的生存策略與寄主範圍,這決 定了他們的分布型態,同時反映了其生態地位。
不同的生存策略,可從他們在樹木或木材上佔據不同的演替階段、持續時程
14
與傳播方式看出。專精心材腐朽之先驅菌種通常具備分解木質素、纖維素的能力,
此為相當緩慢的過程,故多長期棲於心材材積量大之老樹或大型枯木,對環境因 子的接受範圍較大,對逆境也有更高的耐受性。它們的傳播方式常為孢子與菌絲 透過根部接觸並進。部分真菌物種具病原性,能夠侵犯樹木的活細胞,包括邊材 與維管束,此種生存模式稱為「兼性寄生」;此類菌種對材積需求較小,可感染 年輕樹木,因此,林齡結構與棲地限縮等因子對其影響也較小。褐根病菌(Phellinus noxius)即為具強悍寄生能力腐朽菌之例子,其寄主範圍寬廣,因而成為佔臺灣樹 木病害通報案件最大宗的樹木病原。在演替階段中較晚出現的真菌物種,由於需 面對更多他者的競爭,會較傾向 R 策略,也相對更依賴孢子來傳播(Boddy &
Heilmann-Clausen,2008; Schwarze et al,2000)。
寄主廣泛的物種,受限於森林類型、棲地狀況的程度相對較低,而寄主專一 的物種,在競爭上可能有其優勢,但分布受限,也較易受環境改變的影響。寄主 廣泛者較不受植群分布的限制。其中寄主的偏好,以及不同樹種對同種真菌抵抗 力之差異,是促進森林更新與結構改變的重要因子。
2.3 淡黃木層孔菌的生理與生態
2.3.1 生態定位:弱病原性之初級心材腐朽菌
木層孔菌屬(Phellinus spp.)為世界泛佈的木材腐朽菌與樹木病原,其中包括了 各級木材分解者,本文所探討的淡黃木層孔菌,為心材腐朽菌,會削弱莖幹或根 系的機械支撐力,而導致樹木風折或傾倒;目前對淡黃木層孔菌的生存策略並未 有深入的研究,但由於其以莖腐與莖基部腐朽(stem and butt rot)為主,根據田野觀
15
察,應為腐朽速度緩慢、病原性不強的兼性寄生者 (Davidson, Campbell, & Vaughn,
1942;張東柱,2004,2005)。
2.3.2 巨觀與微觀型態
淡黃木層孔菌屬刺革菌科、木層孔菌屬。菌絲二次元,具隔,生殖菌絲無扣 子體(simple-septate)。子實體一至多年生,表面暗黃褐色,遇氫氧化鉀(KOH)溶液 變黑,平伏、反捲至有菌蓋,木栓質至木質,單生或成群覆瓦狀生長,半圓形,
多皺摺,寬可達數十公分,菌孔近圓形,每毫米 6-8 個,黃褐色至紫褐色。剛毛豐 富,厚壁,源自子實層(hymenial setae),大小 12-42*4-6 微米。不具剛毛菌絲(setal hyphae)。擔孢子無色,光滑,單層壁,橢圓形至卵形,大小 4-5*3-5 微米。培養之 菌落前緣菌絲氣生至平伏,棉狀,白色、鮭色至褐色 (Davidson et al,1942;Gilbertson,
1979;Larsen & Cobb-Poulle,1990;張東柱、周文能、王也珍、朱宇敏,2001;
史香芸,2012)。本研究中對淡黃木層孔菌的鑑定以子實體與培養菌落之巨觀特徵 為主,子實層與孢子之微觀型態特徵為輔。
2.3.3 營養模式與生長環境
心材腐朽菌在自然界中(亦即木材內)的生長過程不易觀察,而僅能以木材顯微 切片來觀察其腐朽模式、在木材細胞內之生長狀況,以實驗室培養來測量其最適 生長環境與生長速度。不論早期或近年的實驗室研究,都顯示 30°C 左右為淡黃木 層孔菌的生長最適溫度 (Davidson et al,1942;Rew et al,2000;Jo et al,2006;
史香芸,2012)。在自然環境中,木材內部的溫度難以測量,並且除了受氣溫影響,
濕度、是否為活樹、木材量、暴露程度、腐朽階段等也是決定溫度的極重要因子。
16
淡黃木層孔菌為均質白色腐朽菌(uniform / simultaneous white rot fungus。
Gilbertson,1979。本詞為自行中譯),其營養菌絲伸入死亡中空的木材細胞內,分 泌 laccase、tyrosinase 等酵素(史香芸,2012;鑑定為 Fuscoporia gilva),將鄰近木 材細胞壁的主要成分,包括木質素、纖維素與半纖維素,由內往外以差不多的速 度降解為自己的碳源,同時不斷產生菌絲分枝,分解並穿透木材細胞壁與中膠層 (middle lamella),形成許多孔洞。與另一種白色腐朽 -- 選擇性木質素降解 (selective delignification)比起來,均質白腐導致木材更快速且全面的鬆散脆化 (Schwarze et al,2000)。
2.3.4 繁殖與傳播
淡黃木層孔菌的有性生殖由擔孢子完成,為四極異宗交配(tetrapolar bifactoral heterothallism),兩個交配型因子(mating type factors)皆為複等位基因
(multiallelic)(Rizzo et al,1995)。子實體在自然界中相當常見,應為主要的傳播方 式。目前尚無發現無性孢子之報告。
擔孢子的產生季節、數量、傳播方式與距離目前皆無研究,但通常此類真菌 多依靠氣流與風,傳播距離可至數米、數公里;厚壁、存活力強的擔孢子,若在 適當的季節飛散,則有跨越大洲傳播的潛力(Heilmann-Clausen & Boddy,2008)。
此外,有些根腐菌在森林中靠營養菌絲從樹木根接(root graft)處傳播,形成聚 集分布(clumped),亦即鄰近的樹木會先遭傳染,甚至形成不斷往外擴大之疾病中 心(disease center)。淡黃木層孔菌並非典型根腐菌,但在野外仍可觀察到小群感染 樹木形成集落,因此,根接傳播法可能仍佔一定的比重。
17
2.3.5 生命史
淡黃木層孔菌為腐朽菌,木材為其能量來源,故其生命史與樹木密不可分。
本文採用樹病週期的觀念,將其生命史分為幾個環節:感染期、生長期、產孢期 與傳播期。
感染,或稱接種(inoculation),指找到或建立進入基質的入口;對活樹的心腐 菌而言,通常這指樹木的傷口、枯枝或小枝條,使孢子或菌絲可以避開樹皮的防 護與活細胞的免疫抵抗,進入已死亡的心材空細胞中(Shigo,1979;Schwarze,2000)。
成功感染後,可能進入生長期,擴展群落(colonization);若環境不適,也可能進入 休眠,潛伏至逆境過去再開始生長;由於本菌較不侵犯活組織,大部分僅利用樹 心木材,因此能與樹並存很長一段時間,直到樹幹因強度弱化而折斷、或因其他 原因死亡。一旦累積足夠能量、或當基質養分即將用盡時,菌落開始產生子實體,
進入有性生殖之產孢期,準備往外尋找出口,此時的感染源具有最強的傳染力。
孢子成熟且環境符合釋放條件時便會散播至大氣中,隨機落至樹木傷口或適合的 基質(例如倒木)上,開始下一輪的週期。然而這僅為簡化之模型,對淡黃木層孔菌 的基本生理,及其與樹木的互動方式,仍有諸多空白環節有待研究。
2.3.6 淡黃木層孔菌在臺灣之研究現況
淡黃木層孔菌為世界泛布的木材白腐菌。從熱帶、亞熱帶到溫帶皆有分布,
記錄過的寄主樹種相當廣泛,至少有 75 個樹種,包括針葉及闊葉樹(Fungal Databases,2017;Encyclopedia of Life,2017)。然而在臺灣並無系統性的全面調查。
集結各地菌相調查報告來看,其子實體幾乎皆發現於海拔 1,000~2,500 公尺之間的
18
天然闊葉或針闊葉混合老熟林中,大部分長在殼斗科、尤其是長尾栲(Castanopsis cuspidate var. carlesii)與杏葉石櫟(Lithocarpus amygdalifolius)大樹或死樹、倒木上 (張東柱,2004,2005;周文能,2000;黃靜芬,2014;林偉峻,2013;陳昇明,
1999;王也珍,2004),有些學者更推測其為櫧櫟林更新演替的重要推手(張東柱,
2004,2005;王也珍,2004)。
目前並無足夠資料可直接驗證此推論,因為世界各地的資料繁雜且準確度未 知,臺灣現有的資料則稀少零散,且皆缺乏長期記錄。筆者曾在南投北東眼山兩 個森林永久樣區調查大量長有淡黃木層孔菌之殼斗科大樹以及大量倒木,並比對 兩個樣區間隔 12 年的兩次每木普查資料,得知兩次普查之間這些大樹的變化並不 大:立者恆立,倒木則幾乎皆於 12 年前已倒,可見 12 年的時間並不足以記錄腐 朽菌可能造成的森林動態(宋國彰,1996;Song et al,2010)。因此,本論文將不探 討時間上的動態,而專注於分析生態上的分布。
2.3.7 小結
雖然淡黃木層孔菌造成心材的腐朽速度,以及從接種到形成子實體所需時間,
目前皆無法得知,但可從以上對心材腐朽菌的研究結論中,推論其為生長與擴張 速度緩慢、耐受性高、與樹木或倒木長久並存的真菌物種。
本節所列舉文獻,以及本研究之野外調查,絕大部分是建立在真菌子實體野 外調查的基礎上,因此對真菌實際的分布範圍皆有相當程度的低估,因為即使沒 有子實體,孢子或菌絲仍可能在寄主內部存活或休眠,此為本研究的一大限制。
然而,子實體的出現,代表該真菌在當地的群落已有一定規模與歷史,並且能夠
19
行有性生殖,有完整的生命史,符合 2.1 提及的基本假設:調查中所發現之本物種 分布情形,皆已與其生態環境達成一定平衡,是物種特性、適應力與該地資源、
環境條件互相符合的結果。
2.4 研究尺度
地理上的研究尺度會決定各項因子的影響力,因為在不同尺度會觀察到不同 的分布特性與現象,目標物種的移動範圍也。尺度越大、解析度越粗,例如跨洲 的物種分布研究,則巨觀氣候條件、地理隔離等因子就越重要。而若採小至幾百 公頃的保護區、大至數個縣市的區域尺度,則區域性的植群、土地利用、地形與 氣候等因子的影響力就往往較為突顯。適當的資料解析度才能鑑別出影響物種分 布的因子(Guisan,2005)。本研究採區域尺度,所有資料如植群與氣候的解析度皆 與此相符。
2.5 調查方法論:生物分布取樣方法
對特定物種生態分布資料之取得方法,大致可分為兩種。一為大規模的調查:
設計統一的隨機取樣方法,根據物種特性設計調查變數(variables),可明確記錄該 物種之「發現」(presence / occurrences)與「未發現」(absence),以及現地生物與環 境因子,資料完整,但往往曠日廢時。許多生物已有這種普查方法的歷年資料庫,
例如各國繁殖鳥類大調查(BBS)或全國性的植群普查,可用於研究;但對於單一物 種的調查則不多,研究者往往需自行取得,成本較高之外,也較難涵蓋廣大範圍。
第二種方法較為常見:集結以往「發現」地點之記錄,包括學者或業餘人士的調 查報告、或標本館藏,再從環境資料庫中取得這些地點的環境數據;例如 A. K.
20
Wollan 等(2008)就用標本館藏預測了九種大型真菌的分布範圍。此類方法相對容易 取得廣大範圍的資料,唯無統一取樣方法,多為歷年來之逢機發現,或靠專業知 識進行的刻意搜尋,且多缺乏「未發現」記錄以及現地資料。以下就取樣方式與 變數選擇兩方面,討論這兩種方式的不同。
在取樣方面,取樣的目的是為了推論母群體的樣貌,由於第二種方法不符合 隨機取樣原則,亦即並非建立在「樣本可代表母群體」以及「在取樣期間,發現 機率會隨該物種真實分布的機率而改變」之假設上,因此必須另行建立假設。取 樣的方式也影響結果。T. C. Edwards (2006)發現,隨機取樣與跟據專家知識的刻意 搜索,兩種方式會導致模型不同,生態詮釋也不同,需採用不同的基礎假設與分 析工具;後者較易產生偏差。
僅記錄「發現」的資料(presence-only data),也需另建假設來處理「未發現」
的部分,不然會因選樣偏差(sample selection bias)或未發現偏差(non-detection bias) 而影響結果的準確性;一些研究建議設計合適的資料分析方法、或盡可能取得當 初取樣過程資訊,以減少偏差(Yackulic,2013;Hefley, Tyre, Baasch, & Blankenship,
2013),例如 L. Mair 等(2016)就在一種木層孔菌(Phellinus ferrugineofuscus)分布模 擬中,使用僅記錄「發現」的調查資料,但以訪問調查者的方式來輔助推論整體 分布機率。相對的,若調查方式符合隨機取樣,且「發現」與「未發現」並重 (presence-absence data),則需要的偏差校正較少,發生機率的各項條件也較易定義,
結果通常(但不一定)較為準確(Franklin,2009)。
不同調查方法也導致變數的選擇不同。第二種方法依賴既有資料庫,例如 WorldClim.org 網站提供氣候數據(bioclimatic data),衛星與地圖系統提供地形、坡
21
向資料,而有些地區可取得生物數據,例如林齡、立木材積、棲地破碎度等。變 數選擇基於對目標物種的了解與假設,但在此也受限於資料庫的可得性。
變數選擇的不同,會影響分析結論。例如 L. Mair 等(2016)發現,Phellinus ferrugineofuscus 的分布與特定樹種的材積以及與老熟林的距離最為相關,氣候因 子次之。Copot 等(2017)則使用歷年發現記錄研究靈芝(Ganoderma lucidum),發現 林型(forest type)的影響超過氣候。H. S. Yuan 等(2015)也使用歷年發現記錄對中國 大陸三種木層孔菌(P. baumii, P. igniarius, P. vaninii)的分布進行模擬,僅使用氣候 資料,最後依據影響力最大的幾項氣候因子推估潛在分布範圍。N. B. Klopfenstein 等(2016)以類似方法模擬重要樹木病原菌 -- 有害木層孔菌(Phellinus noxius,褐根 病菌)的分布。這四份研究的對象皆為腐朽菌(除了 P. noxius 的病原性較高),但僅 第一份有相應的國家普查成果可供驗證,結果呈現高度準確;其餘 3 份則僅以自 己的資料交叉驗證,預測與實際狀況的差距無法得知。這些研究顯示僅用氣候資 料為此領域的研究主流之一,但其準確度亟需驗證。
22
第三章 研究方法
跟據目前對目標物種 -- 淡黃木層孔菌的生物學了解,結合於北臺灣、中臺灣 數次先行勘查的經驗,設計介於 2.4 中所述兩種調查法之間的折衷方法:首先選擇 可能具有重要性的變數,再以快速、大範圍的分層隨機取樣(stratified random sampling),採沿多條穿越線進行的多樣點法,現地記錄與淡黃木層孔菌的出現與 否,並測量各項變數數值。接著,使用調查資料中「生物與環境自變數」與僅用
「環境自變數」兩個組合,分別建立分布模型並驗證之。最後,除了篩選影響淡 黃木層孔菌分布的關鍵變數外,也探討在分析心材腐朽菌物種分布時,生物變數 是否必要、若僅使用環境變數,結果是否相同。
3.1 快速調查方法
3.1.1 調查範圍
本研究的調查範圍限於蘇鴻傑所劃分之全台地理氣候區中之北臺灣內陸區(代 號 NWI)、中臺灣內陸區(代號 CWI)(蘇鴻傑,1985。附錄一)。以北橫公路、中橫 公路沿線為主,其他地區為輔,選定若干登山步道、地域性林業、管線維護步道 或獵徑,沿步道或於步道周邊穿越之各植群群系帶設立樣點調查。選址以易達、
安全、人為干擾儘可能小為原則。
3.1.2 調查分層單位
23
淡黃木層孔菌之生存與樹木密不可分,本研究假設淡黃木層孔菌的寄主範圍 不大,森林類型為影響其分布的重要因子之一,因此調查單位以「臺灣現生天然 植群圖集」之植群群系(邱祈榮,2009)為基本劃分依據。臺灣現生天然植群圖集對 全臺灣各地森林的分類,首先依據植相區分出群系綱(class),再依葉子與物候特徵 區分群系亞綱(subclass),最後依優勢植物物種與特定環境地貌形成之森林形相,
來定義植群群系(formation),此分類反映了植物物種組成,也間接反映出該地區之 氣候條件。其它可能影響淡黃木層孔菌分布的變數還有:寄主樹種、樹木大小與 優勢度、森林密度、年均溫、年雨量、氣溫季節性及雨量季節性(是否具乾雨季差 異)。其中,每個樣點的氣候資料皆由網路資料庫取得,生物變數則需現地測量。
本調查根據以下三個維度區劃分出分層取樣之單位小區:
(1)「臺灣現生天然植群圖集」之植群群系。但刪除與木本植物無關、過於稀 少、或人為干擾嚴重之群系(附錄二)。共選擇了 16 個群系。
(2) 當同一植群群系跨越不同海拔時,包括亞高山、上部山地、山地、下部山 地、低地,則於不同海拔分別取樣。海拔之區分大致為:低地 400 < m,下部山地 400 ~ 1500m,山地 1500 ~ 2500m,上部山地、亞高山均 > 2500m,但個別群系之 區分略有不同,在臺灣北部、中部也不同。
(3) 根據蘇鴻傑(1985)所劃分之全台地理氣候區,個別植群群系之分布若跨北 臺灣內陸區(NWI)、中臺灣內陸區(CWI)的群系,則分開取樣。但由於北部海拔 2500m 以上的山地較少,故上部山地與亞高山群系大多僅於中部取樣。
24
3.1.3 變數選擇
本調查一共記錄 7 個生物變數與 6 個環境變數。
(1) 生物變數:本調查記錄之生物變數如表 1。除植群群系外,皆於現地測得。
表 1 生物變數
變數(variable) 資料類型 備註
植群群系(vegetation type) 類別(categorical) 共 16 種 有無淡黃木層孔菌(P. gilvus presence) 二元(binary) 0 (無) / 1 (有) 樹種(tree species) 類別(categorical) 共 131 種 樹木胸徑(DBH) 連續(continuous) 單位:公分 樣點樹均胸徑(mean DBH)2 連續(continuous) 單位:公分 該地森林密度(tree density)3 連續(continuous) 單位:棵/公頃 該地冠層高度(canopy height) 連續(continuous) 單位:公尺
(2) 環境變數:本調查記錄之環境變數中(表 2),海拔由 GPS 現地測得,雨量 及氣溫資料則從 WorldClim 網站4下載 1 平方公里解析度之全球氣候資料後,將所 有樣點 GPS 點位位置的各月氣候資料抽出計算而得(Fick & Hijmans,2017)。
2 為每個樣點五棵樣樹的胸徑平均值。
3 森林密度計算公式:40000/(A2+B2+C2+D2)。A~D 為四象限中,距樣點最近的四棵樹與樣點 之距離。
4 WorldClim 網站:www.worldclim.org,雨量與氣溫資料乃根據 1970-2000 年記錄所計算。雖然 WorldClim 的資料有所誤差,特別是在如臺灣這種小範圍內地形變化較大處,但仍為準確度可接受 之氣候資料。
25
表 2 環境變數
變數(variable) 資料類型 備註
區域(region) 類別(categorical) 北部 / 中部 海拔(altitude) 連續(continuous) 單位:公尺 年雨量(annual precipitation) 連續(continuous) 單位:公厘
雨量季節性(precipitation IQR) 連續(continuous) 年雨量四分位距(IQR) 年均溫(mean temperature) 連續(continuous) 單位:°C
氣溫季節性(temperature IQR) 連續(continuous) 年均溫四分位距(IQR)
(3) 描述性的輔助資料:寄主樹木健康狀況、傾倒方式、腐朽程度,若發現淡 黃木層孔菌則記錄其在樹上之生長位置、數量,棲地狀況及人為干擾程度等。
3.1.4 取樣方式
根據 3.1.2 的分層單位,在每個單位小區選擇穿越線,沿線取多個樣點調查。
本方法力求快速與效率,並兼顧調查者安全,因此多選擇位於主要道路或產道附 近的登山步道、管線修繕道或林業道路,較易行且人為干擾小。步驟如下:
(1) 調查準備:準備 GPS 植群群系、道路、等高線圖與登山步道等圖層, 以 QGIS version 3.0.0-Girona 疊圖顯示道路途經之植群群系,以篩選合適的穿越線。
空間資料來源:底圖圖資取自 NGIS 國土資訊圖台網站、臺灣現生天然植群圖集 GIS 圖層5。等高線地圖與登山步道航跡檔取自地圖產生器資料庫6。
(2) 野外調查程序:
5 NGIS 國土資訊圖台網站,取自 http://tmap.geospatial.org.tw/index.html。
6 地圖產生器,取自 https://map.happyman.idv.tw/~mountain/twmap3/。
26
a. 進入各植群群系範圍後即可開始設樣點。每個樣點相距 25-50 米(水平距 離)。若經不適合調查的區塊,例如為新近崩塌裸露地,地形危險,或植 被與該植群群系明顯不同,則續行至植群群系相符、安全且適合調查的 點為止。
b. 每個單位小區共擇 10 個樣點7。於每個樣點處,採以雷射測距儀測量森 林密度、冠層高度、GPS 點位與海拔高度。森林密度之測量採用中心點 四分法(point-centered quarter method),或稱四象限法(Cottam,1953,
1956)。
c. 於每個樣點處,檢查距離樣點 15 米以內胸徑最大的 5 棵樣樹,記錄樹 木資料,以及是否出現淡黃木層孔菌。樣樹之胸徑需 10 公分以上,可 為活樹或死樹,死樹不限倒木、立枯、樹頭或斷落之樹幹木段,亦不限 腐朽程度,只需符合胸徑 10 公分以上之條件即可。若大樹位置危險難 達,則找其他樹取代之。若無法湊足 5 棵樣樹,則另行註記。
(3) 所有長有淡黃木層孔菌的樣樹皆須鑑定樹種,必要時採回標本、鑑定保存。
所有淡黃木層孔菌皆須採樣,並以巨觀與微觀方法鑑定之。鑑定標準詳見 2.3.2.。
保存玻片標本以供查驗。微觀鑑定範例詳見附錄六。
3.1.5 調查結果
取樣總數如表 3。總共調查了 364 個樣點,1952 棵樣樹,58 筆淡黃木層孔菌 出現記錄。其中有效樣本為 345 個樣點,1775 棵樣樹,27 筆淡黃木層孔菌,出現
7 分層取樣的比例問題,為本調查所面臨的限制之一。本調查以植群群系為分層取樣之畫分單位,
但各植群群系所佔面積並不相同。未來若欲進行類似的調查,宜盡量取得植群群系所佔面積之可靠 資料,依比例取樣,以準確對應各植群群系的代表性,消除這對資料分析可能造成的影響。
27
於 7 個植群群系中,佔全體的 1.5%。其他 19 個樣點、177 棵樣樹、31 筆淡黃木層 孔菌出現記錄,則是在同樣地點、以同樣方式進行的調查,但非取樣時記錄而得,
或因有些單位小區取樣超過 10 個點而刪去。這些多餘的點不計入統計分析中,但 重要的發現將另行補述。此外,中部的 FB31(上部山地-山地-下部山地崩塌地次 生落葉闊葉林)之「山地」部分,因找不到合適地點而僅做 5 個點。調查地點與時 間詳見附錄三。調查前後既有與新增的淡黃木層孔菌分布點,標示於圖 1。
表 3 取樣總數
總數 北部 中部
樣點數目 355 150 205
有菌樣點 21 (5.9%) 4 (2.7%) 17 (8.3%)
樣樹數目 1775 750 1025
有菌樣樹 27 (1.5%) 5 (0.7%) 22 (2.1%)
28
圖 1 淡黃木層孔菌在臺灣已知「發現」與「未發現」地點總覽。圖示:紅圈為本 次調查中發現淡黃木層孔菌的樣點;藍圈為本次調查未發現菌的樣點。黃圈為前 人文獻或調查中曾記錄過淡黃木層孔菌的地點;綠圈為前人地方菌相調查中未記 錄過發現此菌的地點。前人記錄的參考價值有限,且大部分為模糊定位,因此僅 以圖示,未放入分析中。本次調查點位則皆為精確定位。本圖以 ARCGIS version 10.6 繪製。
3.2 資料分析方法
29
調查所得之資料,除了就各項變數以描述統計進行初步分析外(4.1),亦將採用 決策樹(decision tree)中之分類樹法(classification tree)進行多變數整合分析(4.2)。依 此方法篩選出來的最重要變數建立之分類樹模型之效用,將以集成(ensembles of classifiers)的方式來驗證(4.3)。
為了評估加入「生物自變數」與僅用「環境自變數」建立之模型的差異,多 變數之整合分析均分為「生物與環境自變數」與「環境自變數」兩組分別進行。
本次調查所獲得資料之整理、標準化、視覺化、統計分析(獨立雙樣本 T 檢定、
獨立雙樣本 Wilcoxon 等級和檢定、皮爾森積矩相關係數)、隨機森林與分類樹,皆 由 R 3.5.1 執行(R Core Team,2018)。集成驗證的 R 語言 code 詳見附錄五。
3.2.1 分類樹法
分類樹為機器學習(machine learning)的一個分枝,適合依變數為類別性資料 (categorical data)時使用。在分類樹中的每個節點,母資料會根據所選擇的演算法,
依其中一個自變數的某個數值或某些類別分類。如此重複若干次的分類後,資料 會形成樹狀的多層結構,而當每類資料不再符合分割的演算條件時,就不再分割,
而形成最終子集合,或稱「終結點」或「葉節點」(end nodes / leaves)。這些最終 子集合內,依變數各類別中比例最大者,即為該子集合預測的依變數類別,此比 例也可視為該終結點預測正確的機率。
分類樹法的準確度雖常不如傳統方法如線性或非線性迴歸分析,但若邊界條 件與演算法設定得宜,對於物種發現與否能有良好的鑑別力(Johnson, Chawla, &
30
Hellmann,2012;James et al,2013)。本研究採用分類樹法,另一考量為其長處符 合研究目標:呈現各項自變數的影響力大小,且結果容易詮釋。而最主要的缺點:
當缺乏與依變數高度相關之自變數、或多個自變數高度互相影響時,分類樹的結 構較為不穩,甚至即使小範圍的樣本改變就會改變整個模型,也連帶影響預測力 (James et al,2013)。此缺點可用集成的方式克服。
分類樹的樣貌,除了取決於輸入的自變數之外,也受所選擇做為分裂標準的 演算法影響。在本研究所使用之 R 語言 rpart package 中(Therneau & Atkinson,2018),
內建兩種最常見的演算法供選擇:吉尼係數(Gini index)與資訊獲利(information gain),用以選擇「能使分割後資料中的不純度(impurity)降低值最大」的自變數數 值或類別做為分割點。前者計算的依據為吉尼係數。
吉尼係數公式(James et al,2013):
其中 指第 m 個資料子集合中,共 k 個類別的依變數中各類別所佔的比例。
0 ≤ ≤ 1,因此 0 ≤ − log 。吉尼係數越小,代表資料純度越高,亦即 代表較佳的分割選項。
資訊獲利(information gain)則用「熵(entropy)」來度量資料的歧異度,
熵值公式(James et al,2013):
由公式可看出,吉尼係數傾向放大所佔比例大的依變數類別、忽略所佔比例
31
小的類別。相較之下,依變數類別所佔比例無論大小,資訊獲利演算法皆會給予 一定權重,因此,小比例類別的訊號較不會被忽略,而消失在著重大比例類別純 度的分類樹中。
在 4.2 的分類樹分析中,依變數為二元類別:「淡黃木層孔菌是否發現」,其 中「發現」的比例相當小(5.9%)。因此,為了兼顧「發現」與「未發現」的資料分 析,選擇資訊獲利演算法。
設定 D 值可控制分割標準與分類樹的大小,避免過度配適(overfitting),然而 本研究採取集成方式來平均單棵分類樹潛在因過度配適而拉高的錯誤率(4.3),故略 過此步驟。
3.2.2 以隨機森林分析自變數重要值
使用分類樹分析資料時,選擇輸入哪些自變數,不僅影響分割節點,也影響 最終的預測效果與詮釋複雜度。選擇自變數這個步驟的目標,在於使用儘量少的 自變數來達到足夠良好的預測效果(Genuer,2010)。因此在進行分類數分析之前,
先以隨機森林(random forest)計算各自變數的重要值,刪除重要性低的自變數。
隨機森林之演算法同分類樹,但程序相反:先隨機從 m 個自變數中抽出其中 一個做為分割點,再計算該分割點的重要值,最後從 n 棵樹的眾多節點中統計各 自變數的總重要值。此方法的特點為「隨機選擇分割點」,因此不致偏重選擇重 要值突顯的自變數,而忽略、不評估其他自變數。每個自變數的重要值分為兩種:
根據吉尼係數算出的平均不純度降低值(mean decrease of Gini),與經過隨機置換
32
(permutation)的同一自變數放回同一節點後,整棵樹預測準確率的降低值,該值越 高(準確率降低越多)表示該自變數越重要(mean decrease in accuracy)。前者僅顯示 一自變數在每個分割點的重要值,後者則評估每個自變數對全樹的影響。在 4.2.1 中將根據平均準確率降低值來篩選自變數。
3.2.3 分類樹模型之評價
各組分類樹模型之評價(evaluation)方法,採用重複隨機子取樣驗證(repeated random sub-sampling validation),隨機將所有資料分為 70%的訓練資料與 30%的 測試資料。然而為了避免「發現」樣點不足而無法建立有效模型,設定訓練資料 中需至少有 5 個「發現」的樣點(在 4.3.1 的中臺灣資料集裡,依比例改為至少有 4 個),測試資料中則至少 1 個,並且刪除根資料集分類失敗、無法成樹的抽樣。將 每份訓練資料,用「生物與環境」及「環境」兩組自變數分別建立 1,000 棵分類樹 後,再以對應之測試資料驗證之。
每棵樹模型之預測準確率,乃根據訓練與測試資料的混淆矩陣(confusion matrix,表 4)分別計算的特異度、靈敏度與馬修相關係數三項指標來評估。最後觀 察 1,000 個模型中這三項指標之分布情況與平均數,以比較「生物與環境自變數」
及「環境自變數」兩組模型的差異。此種集成方式能克服單棵樹模型結構不穩定、
或因過度配適(overfitting)造成準確率下降的問題。
混淆矩陣與準確率之計算使用 R 語言的 caret package (Kuhn,2018),為比對 原資料與預測結果之樣點總數計算而成。三項指標的公式如下:
33
表 4
混淆矩陣(confusion matrix)
預測「未發現」 預測「發現」
實際「未發現」 真陰性(TN) 偽陽性(FP) 實際「發現」 偽陰性(FN) 真陽性(TP)
「未發現」之預測準確率:
特異度(specificity,SPE) = TN / (TN + FP)
「發現」之預測準確率:
靈敏度(sensitivity,SEN) = TP / (TP + FN)
馬修相關係數(Matthew's Correlation Coefficient,MCC):
若分母為 0,則自動將分母改為 1(以避免產生闕漏值 NA),此時 MCC 值為 0。
(Tichy & Chytry,2006)
本研究之資料「未發現」與「發現」的比例懸殊,導致準確度(accuracy,ACC
= (TP + TN) / (TP + FP + TN + FN))會被大量「未發現」的正確預測數目大幅拉高,
參考價值有限,因而改以馬修相關係數做為評估標準。馬修相關係數同時考慮「未 發現」與「發現」的數目,其值介於-1 到 1 之間,1 表示預測結果完全準確,-1 表示預測結果與實際狀況相反,越接近 0 則表示預測結果為隨機;接近 1 與-1 都 代表模型有一定效用,而接近 0 則代表模型的效用不大。
34
第四章 資料分析結果
在快速調查所取得的資料中,淡黃木層孔菌的樣點佔全體的 5.9%;若以單棵 樣樹為單位來看,則僅佔 1.5%;此為兼顧「發現」與「未發現」之高度不平衡物 種分布數據(imbalanced data)。這顯示,淡黃木層孔菌的分布並不如原先以為的廣 泛。由於以單棵樣樹為單位的資料,「發現」所佔比例太小,故後續分析將以「樣 點」為單位(表 3 中粗體字)。而僅能以單棵樣樹為單位分析的資料,包括樣樹胸徑、
樣數樹種,與發現淡黃木層孔菌處之描述性資料,則在 4.1 中分析。
4.1 各項變數之初步分析
4.1.1 生物變數
(1) 植群群系
在七個發現淡黃木層孔菌的植群群系中,僅 FB41(山地常綠闊葉林)在北部、
中部皆出現,其餘群系北部、中部皆不同。北部有 FB52(下部山地-低地次生常綠 闊葉林)及 FM41(山地針闊葉混淆林),這兩個群系在中部也有調查採樣,但未發現 淡黃木層孔菌。中部則尚有 FB42(山地常綠闊葉矮林)、FC41(山地針葉林)與 FM32(上部山地-山地-下部山地針闊葉次生混淆林),後兩者在北部亦採樣但未 發現(圖 2)。
35
圖 2 淡黃木層孔菌在各植群群系中發現頻度。紅色為北部樣點,藍色為中部樣點,
未發現淡黃木層孔菌之植群群系不予列入。代號意義:FB41:山地常綠闊葉林、
FB42:山地常綠闊葉矮林、FB52:下部山地次生常綠闊葉林、FC41:山地針葉林、
FM32:山地針闊葉次生混淆林、FM41 山地針闊葉混淆林。
由於發現淡黃木層孔菌的樣點多位於北部或中部獨有的植群群系,因此,植 群群系無法用於跨區分析,須以其它高度相關的變數代替之。代替變數的挑選,
使用隨機森林計算各變數在以植群群系為依變數時的重要值。其中由於淡黃木層 孔菌皆於殼斗科樹木上發現(死樹若無法鑑定則不計入),故設一個新變數:「各植 群群系中殼斗科所佔比例」,加上其他以樣點為單位的自變數,年均溫與冠層高 度則因可能影響分析結果而刪除。結果顯示「殼斗科比例」的重要值遠超其它變 數,其次為各項氣候變數(表 5)。因此後續分析中將以殼斗科比例取代植群群系。
36
表 5
用隨機森林(random forest)計算各自變數相對於依變數(植群群系)的重要值 自變數 / 代號 平均準確率降低值(MeanDecreaseAccuracy) 殼斗科比例 / FAG.by.VT 0.58
樣點樹均胸徑 / MEAN.DBH 0.13
森林密度 / TD 0.02
氣溫季節性 / TEMP.IQR 0.20
雨量季節性 / PREC.IQR 0.20
年雨量 / ANN.PREC 0.29
註:「各植群群系中殼斗科所佔比例」(FAG.by.VT)之重要值遠超過其他自變數,
故在後續分析中將以其取代植群群系。本表使用 R 語言的 randomForest package (Liaw & Wiener,2002)計算而得。
(2) 樹種
本調查共記錄 127 個樹種、其中有 4 種僅鑑定到屬;無法鑑定者另外歸入「未 知(unknown)」,共 401 棵,佔 22.6%,但長有淡黃木層孔菌的所有活樹皆鑑定到 種。有 3 筆資料因現地樹木不足而未取樣。樹種名錄詳見附錄四。
淡黃木層孔菌的寄主偏好十分明顯:除了樹種不明的死樹以外,全都發現於 殼斗科樹上。除了圖 3(B)中的長尾栲(Castanopsis cuspidate var. carlesii)、青剛櫟 (Quercus glauca)、森氏櫟(Quercus morii)、毽子櫟(Quercus sessilifolia)以外,圖 3(A) 加入額外取樣但未算入有效樣本的資料,多了杏葉石櫟(Lithocarpus amygdalifolius) 與南投石櫟(Lithocarpus nantoensis)兩種寄主。
37
圖 3 淡黃木層孔菌寄主樹種圓餅圖。(A)為包含無效樣點的所有資料中之記錄,共 發現 58 點。(B)為 1775 筆有效取樣資料中的 27 筆發現淡黃木層孔菌記錄。括號中 數字為樣樹數量。
(3) 樹木胸徑
單棵樣樹的胸徑方面,資料差異極大,從 10 cm ~ 200 cm 不等。經 log 標準化 之後大致成常態分布,北部、中部的資料分開觀察,亦皆如此。獨立雙樣本 T 檢 定結果顯示,不論以單棵樣樹胸徑或樣點樹均胸徑來看,北部與中部的分布皆無 顯著差異,「無淡黃木層孔菌的樹木 DBH 值分布」亦然;而長有淡黃木層孔菌的 樣樹胸徑分布,經由獨立雙樣本 Wilcoxon 等級和檢定(Wilcoxon rank sum test),也 顯示北部(5 筆)與中部(22 筆)之間並無顯著差別(W = 66, p = 0.51)。
淡黃木層孔菌對寄主樹木的胸徑有明顯偏好。圖 4 顯示長有淡黃木層孔菌的 樹木胸徑平均較沒有淡黃木層孔菌的樹木大,而用此數據做獨立雙樣本 T 檢定的 結果,p 值遠小於 0.001(表 6),支持此項觀察。改以樣點樹均胸徑來分析的結果亦
38
同。胸徑越大的樹,死亡的心材也越多,推測這是最適合心材腐朽菌生長並行有 性繁殖的環境。
圖 4 樣點樹均胸徑分布盒鬚圖。(A)「發現」與「未發現」淡黃木層孔菌生長的樣 點樹均胸徑分布。「未發現」的部分中,DBH > 100 的離群值大部分為針葉樹。(B) 中臺灣與北臺灣的樣點樹均胸徑分布。
(4) 冠層高度與森林密度
森林密度的跨距極大,每公頃 28 ~ 5644 棵樹,即使在同一片森林中,也可能 因區域狀況不同,而在相鄰地區測出相差甚大的數字。無論樣點處有無淡黃木層 孔菌,森林密度皆無顯著差異(表 6)。
森林冠層高度從 4.9 ~ 35 米不等(圖 5)。冠層高度在各種植群群系中差異很大,
若僅比較相同的植群群系,則有淡黃木層孔菌的樣點冠層較高(表 6),若北部、中 部分開分析,結果亦同。
39
圖 5 樣點森林冠層高度分布盒鬚圖。
表 6
重要自變數之獨立雙樣本 T 檢定結果
檢定項目 df t p
單棵樣樹胸徑,北台灣/中臺灣比較 1674.33 -0.03 0.98 樣點樹均胸徑,北台灣/中臺灣比較 353.00 1.12 0.27 單棵樣樹胸徑,發現 > 未發現 28.92 14.61 < .001 樣點樹均胸徑,發現 > 未發現 26.09 8.52 < .001 樣點森林密度,發現 > 未發現 24.63 -0.65 0.52 樣點冠層高度,北台灣/中臺灣比較 325.30 1.71 0.09 樣點冠層高度,發現 > 未發現 32.98 8.54 < .001 註:本表中各項數值皆先經對數轉換處理(logarithmic transformation)。
(5) 發現淡黃木層孔菌處之描述性資料
綜觀發現淡黃木層孔菌的 27 棵樣樹,可歸納出幾項共通點。淡黃木層孔菌大 部分長於離地高度兩米內的莖基部,特別常見於樹幹與地際連接處與隆起的根領 部分;僅 1/10 樣本長於離地兩米以上的高處樹幹。大部分子實體長於板根或樹幹
40
皺摺的凹陷處;若樹幹傾斜,則傾向出現於向地面(朝下的一側),推測是因此面較 不受降雨影響,微氣候較穩定、利於孢子飄散的緣故。寄主樹木的外觀大部分有 中空、開裂或明顯腐朽等缺陷,但仍有 1/3 樣本長於外觀健壯、無明顯缺陷的活樹 上。最特別的一例樣本長在一棵主幹死亡、僅剩萌蘗的長尾栲基部,數量極多,
連直徑不到 10 公分的活萌蘗枝也長上去;這代表淡黃木層孔菌在勢力旺盛、寄主 樹木衰弱時,有攻擊樹木活細胞的能力。
4.1.2 環境變數
(1) 海拔
樣點的海拔分布,從 500 ~ 3100 米不等,跨越低、中、高海拔,中部樣點的 平均海拔比北部高(圖 6(B))。其中淡黃木層孔菌出現海拔幾乎僅限於 2200 ~ 2300 米之間(圖 6(A))。然而,海拔高度僅為樣點「地點」的參考資料,對生物或生態的 直接影響,主要透過其對氣溫、雨量、植群的影響而來,但同樣海拔在臺灣各地 所代表的生物或環境意義皆不相同。因此,本研究將海拔當做描述性資料,排除 在量化分析之外。
41
圖 6 所有樣點的海拔分布。(A),「發現」與「未發現」淡黃木層孔菌之樣點的海 拔分布盒鬚圖,「發現」的樣點海拔明顯集中於 2200 ~ 2300 米之間。(B),北區與 中區的海拔分布盒鬚圖。
(2) 氣候
氣候變數有四項:年均溫、年雨量、12 個月均溫與雨量的四分位差,後兩者 代表氣溫與雨量的季節性。所有樣點的年均溫在 6 ~ 21 之間,年雨量在 2266 ~ 4326 公厘之間。其中淡黃木層孔菌發現地點的年均溫為 12 ~ 17°C,平均 12.5°C;年雨 量 2600 ~ 3600 公厘,平均 3194 公厘;有菌的樣點各月溫差在 6.9 ~ 8.5°C 之間,
平均 7.2°C;各月雨量變化則在 155.8 ~ 345.5 公厘間,平均為 286.1 公厘。
跟未發現淡黃木層孔菌的樣點比起來,有菌地點的年雨量較多,雨量季節變 化較大;年均溫則較低,氣溫季節變化較小(圖 7)。此趨勢與中部樣點的氣候趨勢 一致。由於各項氣候數值皆無法調整為常態分布,故不做獨立雙樣本 T 檢測。
42
圖 7 有無淡黃木層孔菌樣點之氣候條件範圍盒鬚圖。(A),有無淡黃木層孔菌樣點 之年均溫。(B),有無淡黃木層孔菌樣點之氣溫季節性。(C),有無淡黃木層孔菌樣 點之年雨量。(D),有無淡黃木層孔菌樣點之降雨季節性。
4.1.3 變數間的相關性
使用 R 中的 PerformanceAnalytics package (Peterson & Carl,2018)分析各自變 數之皮爾森積矩相關係數(Pearson's product moment correlation coefficient)發現,若 以 70%定義高相關性,呈現高相關的自變數有:樣點樹均胸徑與冠層高度,及高
43
度正或負相關的氣候自變數(年雨量、雨量季節性、年均溫、氣溫季節性)(圖 8、9)。
圖 8 使用 R 語言之 PerformanceAnalytics package 分析中臺灣樣點各自變數之皮爾 森積矩相關係數(Pearson's product moment correlation coefficient)。代號意義(由左上 至右下):TD:森林密度。CH:冠層高度。ANN.PREC:年雨量。PREC.IQR:12 個月均雨量四分位差。MEAN.TEMP:年均溫。TEMP.IQR:12 個月均溫四分位差。
MEAN.DBH:樣點樹均胸徑。FAG.by.VT:各植群群系中殼斗科所佔比例。
44
圖 9 使用 R 語言之 PerformanceAnalytics package 分析全部樣點各自變數之皮爾森 積矩相關係數(Pearson's product moment correlation coefficient)。代號意義(由左上至 右下):TD:森林密度。CH:冠層高度。ANN.PREC:年雨量。PREC.IQR:12 個 月均雨量四分位差。MEAN.TEMP:年均溫。TEMP.IQR:12 個月均溫四分位差。
MEAN.DBH:樣點樹均胸徑。FAG.by.VT:各植群群系中殼斗科所佔比例。
各項變數的初步分析顯示,淡黃木層孔菌偏好大棵殼斗科,以及冠層較高、
年均溫較低,氣溫季節變化較小的森林中,年雨量及雨量季節性在中臺灣有影響,
但在北臺灣並不明顯。
4.2 單一分類樹模型分析
45
4.2.1 自變數篩選
進入分類樹分析前,先以隨機森林計算各自變數的重要值並刪去不重要者,
以降低雜訊干擾。自變數之間若存在高度相關性(correlation),會影響重要值評估 結果,導致其一被低估。因此須先檢查各自變數之間的相關性(見圖 8、9)。呈現高 相關的自變數有:樣點樹均胸徑與冠層高度,及高度正或負相關的氣候自變數。
移除冠層高度與年雨量後,進行隨機森林分析。
圖 10 以隨機森林分析各自變數的平均準確率降低值。(A),生物與環境自變數合 併分析時,各自變數相對於「是否發現淡黃木層孔菌」的平均準確率降低值。森 林密度(TD)重要值極小,予以刪除。(B),同前,但僅考慮環境自變數;各項重要 值差距不大,皆予保留。代號意義:MeanDecreaseAccuracy:平均準確率降低值。
FAG.by.VT:各植群群系中殼斗科所佔比例。MEAN.DBH:樣點樹均胸徑。
TEMP.IQR:12 個月均溫四分位差。PREC.IQR:12 個月均雨量四分位差。
MEAN.TEMP:年均溫。TD:森林密度。本表使用 R 語言的 randomForest package (Liaw & Wiener,2002)計算而得。
46
結果如圖 10。(A)為生物與環境變數合併分析時,各自變數平均準確率降低值 的高低。(B)相同但僅考慮環境變數。本研究自變數的篩選僅參考平均準確率降低 值。平均準確率降低值最低者為森林密度,不到 1%,因此刪除。其餘各項的值差 距不大;由於兩個以上解釋力相當的自變數會壓低且逼近彼此的準確率降低值,
因為隨機置換其中之一並不會大幅影響結果。因此不刪除剩下的自變數。
經過篩選後,確定用於分類樹分析的自變數為:
「生物與環境自變數」組:樣點樹均胸徑、植群群系之殼斗棵比例、年均溫、
年雨量、月均溫與月均雨量之四分位差。
「環境自變數」組:年均溫、年雨量、月均溫與月均雨量之四分位差。
以下分類樹分析以「淡黃木層孔菌發現與否」做為依變數,依據資訊獲利演 算法,分析此兩組自變數對結果模型的影響。
4.2.2 「生物與環境自變數」組
以圖 11 為例,分類樹由上而下分割,最上方為根資料(root),每個節點皆為二 分點,顯示分割根資料集所使用的自變數及其數值,符合的資料走左邊,反之走 右邊。圖中第一分割點為 MEAN.DBH(樣點樹均胸徑(公分)),第二分割點為 FAG.by.VT(殼斗棵所佔比例(%)),第三分割點為 TEMP.IQR(12 個月均溫之四分位 差(°C))。綠色框表示該資料子集合中「未發現淡黃木層孔菌(以 0 表示)」的樣點佔 多數,藍色則表示「發現(以 1 表示)」的樣點佔多數。各框第二行呈現「未發現(左)」
與「發現(右)」的比例。第三行則為該子集合佔所有資料的比例。
47
圖 11 中以樣點樹均胸徑 < 70 公分左右做為第一分割點,意指樣點周邊最大 的樹均胸徑 < 70 公分的地方(圖中紅字 A)幾乎不會發現淡黃木層孔菌。此處的例 外(紅字 B)為雜有大型殼斗科的山地常綠闊葉矮林(植群群系代號 FB42),這種林型 多位於山間小稜線,多為胸徑 30~40 公分左右的杜鵑純林,但附近若有發現淡黃 木層孔菌之常綠闊葉林,則杜鵑林中也可能混雜有長菌的殼斗科大樹。
第二分割點為該地植群群系殼斗科所佔比例 < 35%。符合者為紅字 C,多為 高海拔之紅檜-鐵杉林、冷杉林,或低海拔的肖楠林,殼斗科比例低,樹木雖普遍 大棵,但幾乎無法發現淡黃木層孔菌。此處的例外為紅字 D,多為紅檜、鐵杉為 優勢樹,但仍有零星大型殼斗科的針闊葉混淆林。
前二個分割點皆與植群群系相關。最後分割點為 12 個月均溫之四分位差 > = 7.4°C。換言之,即使森林組成適合淡黃木層孔菌,但溫度季節變化太過劇烈,則 也不容易發現它。紅字 E 與 F 多為海拔較高的針闊混淆林,或低地常綠闊葉林的 上緣。此處可謂淡黃木層孔菌分布的最高與最低邊界,其間恰好大致為雲霧林的 分布區。在所有樣點中,氣溫變化最劇烈者為高海拔、陽明山、以及低地,這可 能解釋了為何淡黃木曾孔菌偏好中海拔,且幾乎無法在林型適合的東北部如福山 樣區、或中部低海拔闊葉林如蓮華池發現。
整體而言,若一地樹均胸徑 > 70 公分、森林中殼斗棵樹木佔整體 35%以上,
且氣溫季節變化 < 7.4°C,則發現淡黃木層孔菌的機會最大。反之,若樹木胸徑不 大,或樹大但殼斗科不多、季節性溫差大,則要發現淡黃木層孔菌相當困難。