國立臺灣大學醫學院臨床醫學研究所 碩士論文
Graduate Institute of Clinical Medicine
College of Medicine
National Taiwan University Master Thesis
The role of Hepatitis B virus X gene on the natural history of hepatitis B virus infection
B 型肝炎 X 基因對於兒童 B 型肝炎感染自然病史所扮演 的角色
張凱琪 Kai-Chi Chang
指導教授: 張美惠 教授
Advisor: Mei-Hwei Chang, Professor
中華民國 105 年 01 月
Jan 2016
目 錄
口試委員審定書………. ……… i
致謝 ………. ………. ………… ii
中文摘要………. ………. …….. iii
英文摘要………. ………. …….. v
碩士論文內容 一、 緒論 1.1 研究背景 ……… 1
1.1.1 慢性 B 型肝炎感染與免疫耐受 ………. 1
1.1.2 HB x 基因 ……… 1
1.1.3 抗病毒先天性免疫系統 ……… 2
1.1.3.1 MAVS 蛋白質在宿主的先天性免疫系統扮演一個重要的角色 2
1.1.3.2 B 型肝炎病毒利用多種策略逃脫先天性免疫系統的攻擊 …. 3
1.1.3.3 HBx 基因與宿主先天性免疫系統之間的關聯性 ………….. 3
1.2 未解決的問題及假說 ………. 4
1.3 研究目的 ………. 4
二、研究方法及材料 2.1 研究對象 ……… 5
2.2 研究方法 ……… 6
2.2.1 B型肝炎病毒標記, 血清學檢測, 病毒定量, 基因型 ……… 6
2.2.2 B型肝炎X基因序列比對分析 ……… 6
2.2.3 免疫組織化學染色法分析HBx蛋白質與MAVS蛋白質的表現… 7
2.3 統計方法 ……… 8
三、結果 3.1 免疫耐受期的結束 ……… 8
3.2 HBx 基因定序 ………. 9
3.3 影響免疫耐受期的因素 ………. 9
3.4 比較不同時間點所測得之HBx蛋白突變 ……….. 10
3.5 使用免疫組織化學染色法研究肝臟組織切片內HBx蛋白質以及 MAVS蛋白質的表現 ……… 10
四、討論 ………. 11
五、結論及展望 ………. 13
六、論文英文簡述 ………. 15
七、參考文獻 ……… .. 33
八、圖表 ………. 41
ii
致謝
感謝指導老師張美惠教授的指導,老師指引我研究方向並且不厭其煩、鉅細靡遺 地與我討論,提供我許多寶貴的意見。並且感謝許宏遠教授、倪衍玄教授、陳慧 玲教授、吳嘉峯副教授、鄭永銘教授、蔡慧慧博士、以及小兒腸胃科實驗室同仁 的指導與協助。
iii
中文摘要 研究背景及目的
慢性 B 型肝炎感染是一個全球的健康議題。持續性的 B 型肝炎感染常常會演變 成為慢性肝臟發炎或是肝癌。B 型肝炎感染,流行地區例如--台灣,通常開始於嬰 幼兒期,並主要藉由母嬰垂直感染而使兒童成為帶原者。兒童的慢性 B 型肝炎感 染在年幼時期通常沒有症狀,這段免疫耐受期通常會持續 15 年甚至更久。 B 型肝 炎病毒如何躲匿並對抗宿主固有的先天性免疫,並使患童不產生免疫反應的機制 目前還是未明。近來發現 B 肝病毒功能性基因之一的 HBx 基因可能在 B 型肝炎自 然病史中扮演了某種重要的角色。HBx 基因的變異也曾被報導與肝臟腫瘤發生的 原因相關。然而,對慢性 B 型肝炎兒童來說, HBx 基因的變異是否跟免疫耐受期 (immune tolerance phase)的維持或結束有關係,目前仍然不清楚。HBx 基因的突變 對於兒童慢性 B 型肝炎的長程自然病史之影響仍然有待研究。我們希望探討 HBx 基因變異與兒童慢性 B 型肝炎感染長程病史之間的交互作用。
研究方法
臺大醫院兒童肝膽腸胃科自兒童期開始對一群慢性 B 型肝炎感染的患者做成長 程追蹤。自幼年開始,他們在門診規律性定期接受病史詢問及理學檢查,並且也 接受抽血檢驗,其中包括肝轉胺酶、B 型肝炎病毒標記(HBsAg, HBeAg, Anti-HBe, Anti-HBs, Anti-HBc)、胎兒蛋白、B 型肝炎病毒去氧核醣核酸的定量、與 B 肝病毒 基因型測定等。在這些慢性 B 型肝炎感染的個案中,我們追蹤未曾接受 B 型肝炎 抗病毒藥物治療、未合併有其他 C 型肝炎或 D 型肝炎…等可能導致肝臟發炎因素 的病患,取其兒童期第一次接受抽血檢查之血清,萃取其中的 B 型肝炎病毒去氧 核醣核酸,並進行 HBx 基因的定序。接著與已知資訊中的 HBx 基因序列進行比對 並分析,以找出突變點,並且與臨床資料整合做分析, 我們希望能探討 B 肝病童 之 HBx 基因突變與免疫耐受期長度以及與自然病程之間的關連性。
研究結果
在這個研究中,我們追蹤了155位同意加入本研究之病患至少5年以上,其中有 105位是男性。他們平均起始追蹤年紀為8.9± 4.5歲,平均追蹤時間為23.4± 6.7年。
有110位病患在追蹤期間經歷了免疫耐受期的結束,並且有81位患者經過B型肝炎e
iv
抗原抗體轉換的過程。根據155位患者聚合酶連鎖反應附帶分析比對,基因型B型 占所有病患的77.4%。另外, HBx基因出現機率最高的突變位點有五個,包括 V44G/L, A66T, R87G, N118T, I127V。利用Kaplan-Meier存活分析,我們發現基因 型B型、HBx基因的R87V、 N118T、以及I127V突變,相較於基因型C型及沒有發
生突變者,有較高的比例容易結束免疫耐受期而進入到免疫廓清期(發炎期),並且
達到統計學上的顯著意義(p<0.001, p=0.017, 0.044, 0.001)﹔在多變項分析,僅有 HBx基因的I127V突變發現到有較高的機會較早突破免疫耐受期而進入到免疫廓 清期,其HR為2.07; p值為0.022,達統計學上顯著意義。
結論
在慢性 B 型肝炎感染自然病史中, 我們發現 HBx 基因的 I127V 突變和較早突 破免疫耐受期有顯著相關。
關鍵詞
慢性 B 型肝炎; B 型肝炎 x 基因; 免疫耐受期; 第一型干擾素; MAVS 蛋白
v
Abstract
Backgrounds and Aims
Chronic hepatitis B virus (HBV) infection is a world-wide health problem. Persistent HBV infection is often associated with chronic liver disease and may lead to liver cirrhosis and hepatocellular carcinoma (HCC). HBV infections were mostly acquired in the infancy and early childhood through the route of mother-to-infant transmission, especially in endemic areas such as Taiwan. Children with HBV infection often
remained asymptomatic at their young age, and their immune tolerance phases may last 2 decades or longer. The mechanism of HBV to evade or overwhelm the human innate immune system, and to keep a state of relative hypo-responsiveness to HBV in children still remained largely unclear. Recently, HBx gene, which was one of the four HBV functional genes, had been reported to be involved in the host immune abrogation.
However, the relationship between HBx mutation and either the persistence or the ending of immune tolerance was unknown. The impacts of HBx mutation on long term natural course of chronic HBV infection remained to be elucidated, particularly in children. We aim to investigate prognostic value of mutant HBx gene on the natural history of HBV infection from childhood to adult life.
Materials and Methods
Patients were recruited from long term prospective chronic HBV carrier cohort,
vi
which was conducted in Pediatric department of National Taiwan University Hospital.
They received evaluations periodically including history taking and physical
examinations. Liver enzymes, HBV markers (HBsAg, HBeAg, Anti-HBe, Anti-HBs, Anti-HBc), AFP, DNA quantitation, and HBV genotyping were also assessed. The blood samplings collected at initial time of enrollment were examined for HBx gene.
We analyze the sequences of HBx genes and tried to find the mutations.
We attempt to elucidate the correlations between HBx gene mutations and the lasting or ending of immune tolerance phase.
Results
A total of 155 children (105 males) were followed for 23.4± 6.7 years with initial age of 8.9± 4.5 years. Genotype B accounted for 77.4% of all patients according to sequence findings. One hundred and ten patients had breakthrough immune tolerance phase and 81 patients achieved HBe seroconversion. HBx gene mutations of V44G/L, A66T, R87G, N118T, I127V had highest frequencies. In Kaplan-Meier survival analysis, genotype B and mutants of R87V, N118T, and I127V in HBx gene had significantly
higher cumulative proportion of breakthrough of immune tolerance phase (log rank test,
p<0.001, p=0.017, 0.044, and 0.001, respectively). In multivariate analysis, we found
that mutation of I127V in HBx gene was an independent favorable factor to predict
vii
earlier breakthrough of immune tolerance phase, with a statistically significant value
(hazard ratio 2.07, 95% CI 1.11-3.85, p=0.022)
Conclusions
In chronic HBV infection, patients with I127V mutation in HBx gene had higher potential to breakthrough the immune tolerance phase.
Keywords
Chronic hepatitis B infection; hepatitis B X gene; immune tolerance phase; type I interferon; MAVS protein
1
一、緒論
1.1 研究背景:
1.1.1 慢性B型肝炎感染與免疫耐受期
慢性B型肝炎感染會造成一連串的嚴重肝臟疾病包括慢性肝炎、肝硬化、或 肝細胞癌。在B型肝炎盛行的地區,例如台灣,慢性B型肝炎的感染主要是在嬰 幼兒期發生,並且主要的途徑為透過母嬰感染而造成。此種傳播途徑在B型肝炎 疫苗廣泛施打地區,是疫苗失敗的感染(breakthrough infection)最主要原因(Chu et al., 1989; Chang, 2008)。
在慢性B型肝炎感染的早期,由於宿主(兒童)與B型肝炎病毒之間多處於相互 不對抗的狀態,達成一個巧妙的平衡,因而建立起一段免疫耐受期(immune tolerance phase)。由於此時患童的免疫系統並不積極的將病毒清除體外,於是導 致B型肝炎病毒得以持續存在人體當中(Chisari et al., 1995)。我們推測免疫耐受期 的成因是因為短時間大量的B型肝炎病毒透過母體傳給當時免疫系統尚未成熟 的嬰幼兒,由於嬰幼兒免疫系統辨識外來抗原的能力還不足,無法產生有效的 抗體-抗原作用,也就無法發揮將病毒清出體外的能力,導致免疫耐受期的產生。
一般認為B型肝炎病毒透過抑制第一型干擾素(type I interferon)來躲避先天性 免疫系統(innate immunity)的攻擊。B型肝炎病毒基因體四段開放讀碼區中的其 中一段--HBx 基因,被報導具有重要的能力可以去抑止干擾素的生成途徑 (Busca et al., 2014)。不只如此,HBx gene 基因還可以調控病毒本身及宿主細胞 的許多功能,它還可能使B型肝炎病毒成功躲匿宿主免疫的攻擊。
1.1.2 HB x基因
HBx gene是B型肝炎病毒基因體四段開放讀碼區中最小的一段,它跟另外的 聚合酶基因(polymerase gene)以及前核心區基因(precore gene)都有部分的重疊片 段。HBx 基因的基因產物為HBx 蛋白,長度約154個胺基酸,分子量的大小約 17.2kDa。根據前人的研究,HBx具有多種細胞調控能力,不但牽涉蛋白質與蛋
2
白質之間的交互作用,並且也發現具有轉活化(transactivator)的功能,能活化許 多基因,包括病毒的啟動子(promoters)與加強子(enahncers)。它也被認為可以影 響許許多多細胞內的運作,包括訊息傳遞( signal transduction )、轉錄調控 ( modulation of transcription )、細胞凋亡( apoptosis )、細胞週期調控( cell-cycle regulation )與癌症的形成( carcinogenesis )…等等(Bouchard et al., 2004; Benhenda et al., 2009; Bouchard et al., 2004 )。
由其廣泛調節能力顯示HBx 基因可以藉由調控病毒本身啟動子(promoters) 的作用回過頭來促進B型肝炎病毒基因下游的其他表現,HBx 基因也可以藉由 和宿主細胞核內的各式基礎複合物(basal machinery)和轉錄因子(transcription factors)產生交互作用,改變細胞內正常訊號分子的傳遞,藉此來調節周遭環境 給予B型肝炎病毒的影響,轉變成為有利於病毒複製的條件(Wei et al., 2010)。
除了具有轉活化的功能以外,HBx 基因 也具有活化許多胞內訊息傳遞路徑 (signal transduction pathways)的功能,前中牽涉到包括: Ras-Raf-mitogen-activated protein kinase (Ras-Raf-MAPK), extracellular signal-regulated kinase (ERK),
stress-activated protein kinases/ NH2 -terminal-Jun kinase (SAPK/ JNK), protein kinase B (PKB/Akt), and the Janus kinase /STAT (JAK/STAT) ..等的重要訊息傳遞 分子(Benn et al., 1996; Guidotti et al., 2006)。
1.1.3 抗病毒先天性免疫系統 (Antiviral innate immunity)
1.1.3.1 MAVS蛋白在宿主的先天性免疫系統扮演一個重要的角色
第一型干擾素(Type 1 interferon, IFN) 的製造和自然殺手細胞(natural killer cell, NK cell)在第一線將病原體清除於人體外之功能上扮演重要的角色。除了類 鐸 接 受 體 (Toll-like receptor, TLRs)和 Nod 樣 接 受 體 (Nod-like receptor, NLRs)可 以 辨 認 出 外 來 入 侵 的 抗 原 以 啟 動 信 號 傳 遞 以 外,還 有 另 一 個 重 要 的 傳 遞 路 徑 是 透 過粒線體抗病毒信號蛋白(mitochondrial antiviral signaling protein, MAVS protein) (Takeuchi et al., 2009; Hoffmann et al., 2015)。 MAVS 蛋白 會利用與第 I 型類維他命 A 酸可誘導基因(retinoic acid-inducible gene I , RIG-I)以
3
及黑色素瘤分化相關基因(melanoma differentiation-associated gene 5, MDA5)結合 形成穩定的信號小體(RIG-I-MDA5-MAVS),進一步往下誘導干擾素的產生 (Bowie et al., 2008)。過去的研究曾發現,A 型肝炎病毒和 C 型肝炎病毒都會透 過破壞 MAVS 蛋白質,使得信號傳遞途徑的下游無法生成活化性的 nuclear factor–keppaB (NF-kB)和干擾素調控因子 3 (IFN regulatory factor-3, IRF3),於是 使後續的第一型干擾素無法生成(Bowie et al., 2008)。
1.1.3.2 B型肝炎病毒利用多種策略逃脫先天性免疫系統的攻擊
對於 B 型肝炎病毒來說,過去已被報導它會利用數種不同的策略來逃避宿主 的免疫系統攻擊。B 型肝炎核心抗原(HBV core antigen)被發現會抑制 β 型干擾 素(IFN-β)的轉錄(Thomas et al., 1991)。另外,B 型肝炎病毒微粒(HBV virons)、
表面抗原(surface antigen)、e 抗原(e antigen)、和前核心區抗原(precore antigen) 都曾被發現具有阻斷類鐸受體(Toll-like receptor, TLR)所誘發之抗病毒作用的能 力,因此導致β 型干擾素(IFN-β)及干擾素往後所誘發之基因跟著無法被表現(Jun et al,. 2007; Chen et al., 2004; Reignat et al., 2002; Wu et al., 2009)。B 型肝炎病毒 陸續被發現會利用多種方式去干擾拮抗第一型干擾素和細胞激素(cytokines)的 誘發,進一步阻斷先天性免疫系統中抗病毒功能的發揮。
1.1.3.3 HBx基因與宿主先天性免疫系統之間的關聯性
既然A型肝炎病毒和C型肝炎病毒皆被發現會利用阻斷MAVS 蛋白來干擾先 天性免疫系統的作用,則MAVS 蛋白在B型肝炎病毒的感染途徑中所扮演的角 色也為人所好奇、想進一步探討。近年來報導,B型肝炎病毒可能利用其附加蛋 白質-HBx 蛋白,來辨認MAVS蛋白並造成該蛋白質的分解(Hoffmann, et al., 2015)。
當HBx 蛋白靠近粒線體(mitochondria)上的MAVS 蛋白時,可能主要利用其C 端(C-terminus)的胺基酸片段來附著,因此這段C 片段是不可或缺的。若HBx基 因發生突變而造成胺基酸序列的改變,HBx蛋白將失去附著上MAVS蛋白的能力 (Li et al., 2008; Wei et al., 2010)。
4
既然MAVS蛋白在傳遞訊息、往下製造生成β型干擾素的路徑上扮演一個重責 大任,並且β型干擾素又與先天性免疫系統功能的發揮、甚至誘導後天性免疫反 應的出現息息相關,我們不禁想問: HBx基因有無產生變異、以及它所生成的蛋 白質產物是不是也在抗病毒的防禦面上,甚至關於慢性B型肝炎感染的免疫耐受 期佔有一定的影響?
1.2 未解決的問題及假說
慢性B型肝炎感染的兒童大部分的時間都是處於免疫耐受期。因此,探討免 疫耐受期的維持跟結束對於了解B型肝炎的自然病史很重要。但為什麼免疫耐受 期可以從嬰幼兒時期持續到青少年及青年,目前大家仍無法提出合理的解釋,
另外它的機制也仍然不明。
因此在此提出我們的假設:
1. 造成免疫耐受期結束並開始免疫廓清期(發炎期)的原因仍為未知,既然HBx基 因過去被報導可以透過其產物HBx蛋白來破壞MAVS蛋白,進而阻斷下游的訊 息傳導來間接避免病毒遭到宿主的先天性免疫系統的攻擊,我們推測HBx 基因 跟免疫耐受期的維持及結束之間可能具有相關性。但是目前關於HBx 基因的長 期性研究仍然十分欠缺,因此若我們對HBx 基因進行深入的探討跟研究,或許 可以找出免疫耐受期維持與結束的原因,並且也進一步對B型肝炎造成的慢性 肝臟疾病增加認識與了解。
2. B型肝炎病毒在進入肝細胞時,會透過抑制TLR所誘發的抗病毒能力以及干擾
素的製造來躲避第一線先天性免疫系統的攻擊,而透過破壞MAVS 蛋白同時 也抑制β型干擾素生成的另外這一項路徑,到目前為止,報導較少。而我們目 前仍缺乏有效的方式來阻擋B型肝炎病毒進入宿主細胞,往積極面著想,透過 增加對HBx 基因的了解,我們期待可以找出更好的策略對抗B型肝炎病毒的入 侵,或許在自然病史的早期就可以阻斷免疫耐受期的生成以及維持。
1.3 研究目的
到目前為止,沒有任何一種有效的辦法可以促使先天性免疫系統成功對抗B
5
型肝炎病毒的進入,來避免免疫耐受期的產生與建立。我們進行HBx 基因的相 關研究,目的是要探討是否可以利用對於HBx 基因多一些研究,來知道它在慢 性B型肝炎的自然病史上所扮演的角色,並且可能在不造成宿主肝細胞的傷害 下,建立新的策略來扭轉免疫耐受期。
二、
研究方法及材料
2.1 研究對象
病患血清中B型肝炎表面抗原(HBsAg)陽性持續長達6個月以上定義為慢性B 型肝炎感染。在臺大醫院兒童肝膽腸胃科有一群慢性B型肝炎患者自幼年時期就 持續進行長期追蹤,他們固定至少每6個月回診一次,接受抽血檢驗(包括各項B 型肝炎標記和肝功能)以及腹部超音波的檢查,我們在每次回診皆仔細進行身體 評估和臨床症狀的問診,並且對於家族病史是否有B型肝炎感染或罹患肝細胞癌 的成員,以及個案過去是否接種B型肝炎疫苗、和免疫球蛋白都詳細的紀錄。基 於我們團隊過去的研究,我們定義肝功能異常為丙氨酸轉胺酶 (Alanine
aminotransferase, ALT )數值超過30 IU/L (Wu et al., 2012)。當他們的臨床症狀或 血液檢驗指標有所變化或異常時,則依據其臨床病情而安排較近的門診返診時 距。
對於慢性B型肝炎患者的自然病史,我們界定免疫耐受期為B型肝炎e抗原陽 性、B型肝炎e抗體陰性、並且ALT數值在正常範圍之內。並且我們定義ALT > 30 U/L持續超過6個月為結束免疫耐受期,進入到免疫廓清期。另外,若患者被偵 測到B型肝炎e抗原消失、B型肝炎e抗體出現,穩定持續超過6個月則被認為是肝 炎e抗原血清轉換(HBeAg seroconversion)。
在這群接受長期門診追蹤的慢性B型肝炎帶原者中,若自幼年時期開始病童 舉凡因為B型肝炎病毒感染而在本科追蹤達五年以上,經過家長或法定代理人同 意並簽署同意書,曾保留凍存其加入長程追蹤後第一次抽血、以及突破免疫耐 受期前最後一次抽血之檢體者,則納入為本試驗的研究對象。假設患者有合併
6
感染C型、D型、E型肝炎,或者是先天性人類免疫不全病毒感染(HIV)的病患,
以及曾經接受過抗B型肝炎病毒治療藥物,則會被排除在收案之外。
我們詳細研究他們B型肝炎感染的病史,並辨別他們是否歷經免疫耐受期的 結束與進入免疫廓清期,或者仍然處於免疫耐受期之中。另外,我們取出這些 病患在幼年第一次接受追蹤時所留存的血液檢體,以及突破免疫耐受期前最後 一支血液檢體,萃取並檢驗其感染病毒株之HBx基因,以找尋B型肝炎自然病史 和HBx 基因之間的關係。本計畫研究通過台灣大學醫學院附設醫院研究審查委 員會批准通過。
2.2 研究方法
2.2.1 B型肝炎病毒標記, 血清學檢測, 病毒定量, 基因型
在每次追蹤所採取的血清檢體皆進行肝功能檢測及各項B型肝炎病毒標記,包 含B型肝炎表面抗原,表面抗體(Anti-HBs),e抗原,e抗體,B型肝炎核心抗體 (anti-HBc),檢測方式是利用已經商業化的放射免疫分析法檢測模組(Abbot Laboratories, North Chicago, IL)。第一次追蹤時所留存的血液檢體藉由聚合酶連 鎖反應分析(polymerase chain reaction, PCR)以及後續的熔解曲線分析(melt curve assay)檢驗出B型肝炎感染之病毒量和其病毒株的基因型(Yeh et al., 2004)。血清 中肝臟肝功能生化指標ALT的濃度以自動分析儀 (7450; Hitachi, Tokyo, Japan) 測定。
2.2.2 B型肝炎X基因序列比對分析
患者所感染之B型肝炎病毒的去氧核醣核酸(deoxyribonucleic acid, DNA)取自 長程追蹤之第一次抽血、以及患者突破免疫耐受期前最後一次抽血所留存的血 液檢體,我們利用DNA minikit (Qiagen, Hilden, Germany)將去氧核醣核酸萃取出 來。接著運用巢式聚合酶連鎖反應(nested polymerase chain reaction ,nested PCR) 的技術,將HBx gene作放大倍數以利作分析(HBV nucleotides 1374-1839)。 聚合 酶連鎖反應的產物將使用瓊脂糖凝膠電泳(agarose gel electrophoresis)做分離。
接著其產物將被純化並且作定序。數據將會依照目前公認的HBV DNA sequence
7
做一系列的序列分析(strain ayw, NCBI Reference sequence: NC_003977.2)。
2.2.3 免疫組織化學染色法分析HBx蛋白質與MAVS蛋白質的表現
在我們收案的慢性B型肝炎的患者中,有部分患者過去曾因為肝功能異常而 接受過肝臟組織切片。我們找出他們保存在台大醫院病理部的肝臟組織並裁取 分出部分切片。由於這些肝臟組織檢體,主要是經由福馬林固定,再以石蠟包 埋處理。因此在進行免疫組織化學法之前,須將檢體浸泡於Trilogy™ reagent (Cell Marque, Rocklin, CA),並微波加熱十分鐘,以便同時去除福馬林
(deparaffinization),再水化(rehydration),及修復抗原(antigen unmasking)。為了 更全面修復因石蠟包埋而造成檢體中的抗原被破壞,Trilogy處理後的檢體需再 浸泡於AR-10 antigen retrieval solution, pH 10 (BioGenix, Fremont, CA),微波十分 鐘,放在室溫下冷卻三十分鐘,使用無離子水清洗兩次,最後用PBS緩衝液浸 泡。
免疫組織化學染色法的操作主要是採用UltraVision Quanto Detection System HRP (Thermo Scientific , Fremont, CA)試劑套組,並依據廠商所提供的建議執行:
首先,在室溫中用百分之三的雙氧水浸泡十分鐘,以便消除細胞內生性過氧小 體的活性。之後用PBS緩衝液徹底沖洗三次,每次五分鐘,再利用試劑套組中 所提供的ultra V block於室溫下處理五分鐘以阻斷組織中的雜訊,不須沖洗,直 接加入本實驗室自製的抗HBV X抗原的單株抗體(anti-HBx monoclonal
antibody),或是購買之抗MAVS的多株抗體(anti-MAVS polyclonal antibody)。抗 體先以PBS緩衝液稀釋(若是進行HBV X抗原的單株抗體染色,比例為一比五﹔
但若是進行抗MAVS的多株抗體染色,比例為一比五十),靜置於四度C的冰箱 內靜置二十四小時(supporting table 1)。接著,以PBS緩衝液沖洗五分鐘,重複三 次,加入“Primary Antibody Amplifier Quanto ”放置在室溫下十分鐘。再用PBS緩 衝液以上述方式沖洗。再者,加入“HRP Polymer Quanto”繼續在室溫下作用一小 時。在三次PBS緩衝液沖洗後,加入“DAB Quanto Chromogen”和“DAB Quanto
8
Substrate”之混和液。最後用hematoxylin染色,再用micromount(Surgipath, Richmond, IL)封片。
2.3 統計方法:
我們利用卡方檢定(chi-square test)來做計算及校正,分析HBx 基因突變以及 其他變項分析。Student t 檢定 (student t test) 或是Mann–Whitney U test 被使用 來計算兩組的連續變項其分布之平均值(mean)或中位數(median)之間的差異,並 且檢測95%信賴區間(95% confidence interval)。若為多組的分析,則使用
Bonferroni correction來進行multiple comparisons。我們利用存活分析 (Kaplan –meier survival analysis) 以及Cox 比例風險模型(Cox proportional hazards model) 來檢測突破免疫耐受期的預測因子。
統計上的軟體分析則使用Small STATA 12.0 ® (Stata Corp. Texas, USA)和 SPSS Windows 版本 19 (IBM SPSS Inc., Chicago, IL, USA)。P值<0.05被定義為 統計上達到顯著意義。
三、
結果:
3.1 免疫耐受期的結束
我們總共收集了 155 位慢性 B 型肝炎患者,其中有 105 位男性,50 位女性。
他們從幼年時期即開始接受慢性 B 型肝炎的長期追蹤,他們平均的起始追蹤年 紀為 8.9± 4.5 歲,追蹤時間為 23.4± 6.7 年;其中有 110 位患者在追蹤過程中,曾 經經歷免疫耐受期的結束,當時的平均年紀為 18.3 ±5.2 歲(表 1)﹔另外有 45 位 患者則是在最近一次追蹤檢查,仍然發現處於免疫耐受期之中。
在此 110 位結束免疫耐受期的患者,他們在免疫廓清期期間,ALT 上升之最 高值取 log10 的平均值為 2.31±0.41 U/L。並且此 110 位中有 81 位患者(73.6%) 在追蹤期間,達成 B 型肝炎 e 抗原的消失以及 B 型肝炎 e 抗體的出現(HBeAg loss and Anti-HBe emergence)﹔他們達成 B 型肝炎 e 抗原抗體轉換(HBeAg
9
seroconversion)的平均年齡為 21.87 ±6.34 歲,約是在結束免疫耐受期的 4.72 ± 3.30 年後。
3.2 HBx 基因定序
將 B 型肝炎病毒的去氧核醣核酸自這 155 位患者的血清分離出來之後作分 析。發現這群病人感染 B 型肝炎病毒的基因型以 B 型較多,共 120 位、佔我們 研究群體的 77.4%﹔另外有基因型 C 型 35 位(22.7%)。 另外,B 型肝炎病毒量 在這群患者取 log10 的平均值為 7.48± 0.68 IU/ml。
接著,在經過巢式聚合酶連鎖反應將患者 HBx 基因放大倍數並跑膠送定序、
定序結果根據 NCBI website 所公布之感染人類之 B 型肝炎病毒做比對後發現:
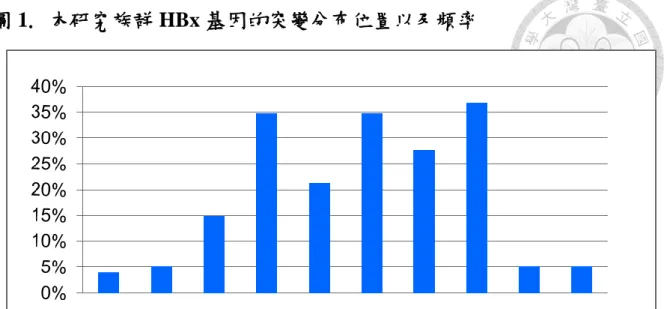
此 155 位患者感染之 B 型肝炎病毒的 HBx 基因,在長程追蹤之第一次抽血中共 有 74 個胺基酸的點位變異。若以 HBx 蛋白的 154 個胺基酸來闡述,我們發現 有五個胺基酸突變位置: V44G/L, A66T, R87G, N118T, I127V 出現的機率最高,
分別偵測到出現頻率為 34.8%, 21.3%, 34.8%, 27.7%, 和 36.8%,並且這些突變大 部分都是出現在基因型 B 型的患者(圖 1)。
3.3 影響免疫耐受期的因素
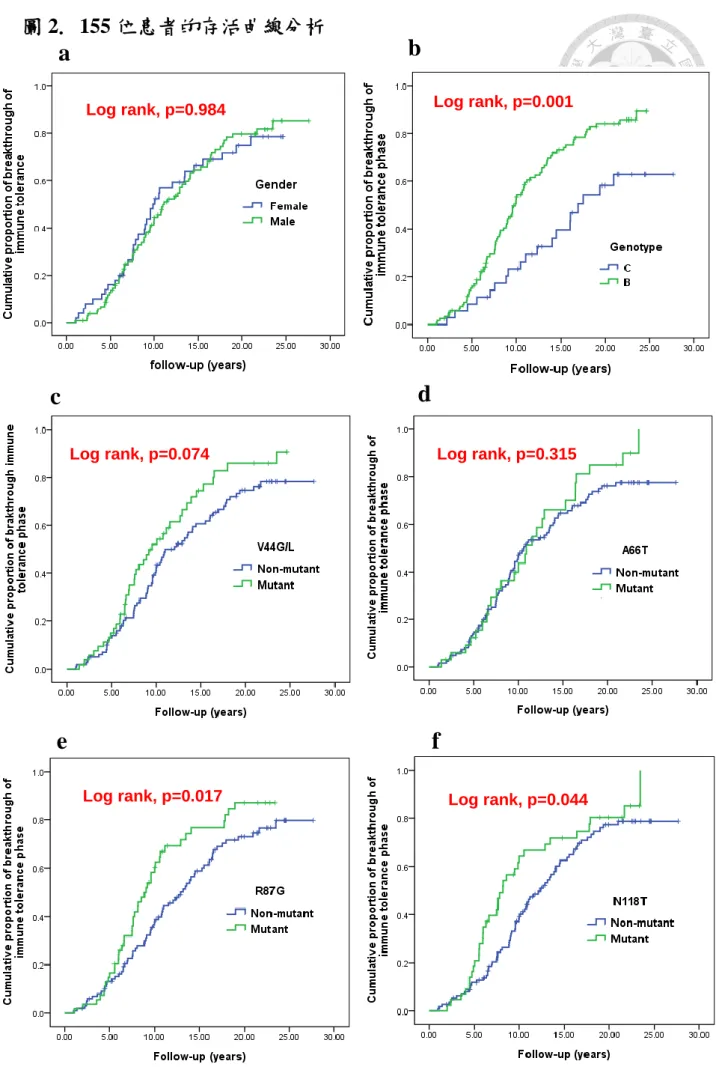
我們利用存活分析(survival curve with log rank test)和Cox 比例風險模型(Cox proportional hazard model) 探討病患的基本特性、HBx 基因和免疫耐受期長短 之間的關聯性。我們發現以Kaplan-Meier圖探討發現: 基因型B型相對於基因型 C型有較高的累積機率會較早突破免疫耐受期(log rank, p=0.001)。另外,在長程 追蹤之第一次抽血,其HBx 基因發生R87V, N118T,或I127V突變的患者,突破 免疫耐受期的機率也比沒有發生突變者的機率高,p值達到統計學上的顯著意義 (log rank, p=0.017, 0.044, 和0.001)。除此之外,HBx基因發生V44G/L的突變之 患者,有較容易結束免疫耐受期、進入到免疫廓清期的趨勢,但p值未達顯著意 義(p=0.074) (圖 2)。在多變項分析,當我們相互校正性別、病毒基因型、病毒 量以及此五項HBx基因突變的因素後,我們發現:在控制其他預測因子後,唯 獨第一次抽血發現HBx基因有I127V突變者,相較於沒有發生突變者,較容易突
10
破免疫耐受期而進入到免疫廓清期,其風險值為2.07,95%信賴區間為1.11- 3.85,達統計上的顯著差異(p=0.022)﹔另外,基因型B型相較於基因型C型,在 預測早期突破免疫耐受期方面也出現一個較高的趨勢,風險值為1.74,95%信賴 區間為0.96- 3.16,但尚未達統計上的顯著差異(p=0.068) (表2)。
3.4 比較不同時間點所測得之 HBx 蛋白突變
我們發現 51 位病患在長程追蹤的第一支血液、以及突破免疫耐受期前最後一 支血皆持續有 HBx 基因 I127V 突變。另外,有 3 位患者在起初追蹤的第一支血 液並未偵測到 HBx 基因 I127V 突變,但卻在突破免疫耐受期前最後一支血發現 到 I127V 突變的存在;相反的,6 位個案在長程追蹤的第一支血液檢體中,原 本偵測到 HBx 基因 I127V 突變,但此現象在突破免疫耐受期前最後一支血液檢 體卻消失發現。透過分析,我們發現 HBx 基因 I127V 突變持續存在的個案群明 顯比 HBx 基因 I127V 從未被偵測到的個案群,會較早突破免疫耐受期進入到免 疫廓清期(log rank, p<0.001)。
3.5 使用免疫組織化學染色法研究肝臟組織切片內 HBx 蛋白以及 MAVS 蛋白的表
現
共有 23 位患者(15 位男性和 8 位女性)曾接受過肝臟切片檢查,這些患者接受 切片檢查的目的,是希望透過組織病理學方式評估肝臟細胞在免疫廓清期階段
因發炎而受到的損害。這些患者接受肝臟切片檢查的平均年齡為18.37±4.09
歲,並且是在進入到免疫廓清期後大約 2.84±2.51 年後接受此項檢查。我們發現 HBx 蛋白和 MAVS 蛋白在這些肝臟組織內的表現具有三種不同的情況: (1)兩種 蛋白都沒有表現( HBx(-)MAVS(-); 13.0%, 3/23 個案) (2) HBx 蛋白染色為陽 性、MAVS 蛋白染色為陰性( HBx(+)MAVS(-) ; 30.4 %, 7/23 個案) (3) HBx 蛋白 染色為陰性、MAVS 蛋白染色為陽性( HBx(-)MAVS(+); 56.5 %, 13/23 個案)。扣 除掉兩位患者之可能因為肝組織檢體年代較為久遠,受保存狀況影響而染色受 限以外,我們發現在這些患者的肝組織檢體中,其 HBx 蛋白和 MAVS 蛋白呈現 一個負相關(圖 3)。但是,比對這些患者感染之的 HBx 基因定序結果,發現是
11
否有 I127V 的突變和肝臟的 MAVS 蛋白表現並沒有明顯相關性(p=0.414)。另 外,對照他們的病程,也發現肝組織染色有 MAVS 蛋白表現者,亦並無較早結 束免疫耐受期的趨勢。
四、討論:
慢性B型肝炎感染的患童通常處在免疫耐受期當中,並且為高病毒複製量,
當他們面臨長期的肝細胞慢性發炎,可能會使他們遭遇肝纖維化或肝硬化的風 險。當血中ALT數值上升,間接代表免疫耐受期結束。目前,我們沒有其他非 侵犯性的工具可以來預測B型肝炎病程從免疫耐受期進入到免疫廓清期。若在免 疫廓清期之中較早經歷到B型肝炎e抗原-抗體陰轉,一般多被認為可以預測較好 的預後。免疫耐受期的早點結束通常暗示病毒量開始下降、並且可視為病程中 一個好的開端,值得我們好好研究(Yang et al., 2002)。
從宿主免疫防禦的觀點來看,無論是先天性免疫系統或後天性免疫系統都牽 涉在對抗 B 型肝炎病毒的過程中。先天免疫系統負責的通常是初始的關鍵步 驟,要及時遏止病毒侵入,並且有效誘導後續特異性的後天性免疫反應發生 (Fisicaro et al., 2009)。但是,B 型肝炎病毒利用策略抑制先天性免疫刺激發生,
進而達到感染宿主肝臟細胞、不被清除掉的目的。這被認為是成功建立起免疫 耐受期的原因(Dunn et al., 2009; Luangsay et al., 2015)。
一些臨床和實驗證據證明,對抗 B 型肝炎病毒特異性的 T 細胞免疫力(T cell immunity)並非不存在,部分研究發現新生兒具有對特定的病毒產生特定 T 細胞 免疫(virus-specific T cell response)的能力(Marchant et al.,2003; Koumbi et al., 2010; Vermijlen et al., 2010)。既然先天性免疫系統中信號的增強可以幫助引發下 游的 T 細胞反應,並且誘導細胞激素所調控之非細胞毒殺力(cytokine-mediated noncytopathic effects),一旦上游的傳遞訊號上升,就可以產生大量的後天性免 疫系統攻擊反應 (Adkins et al., 2004; Levy, 2007)。在兩種免疫系統的交互作用 中,第一型干擾素( IFN-α/β)在先天性免疫系統中扮演重要角色,不但直接抑制
12
病毒的複製、並且間接刺激人類樹突細胞(dendritic cell)的分化,隨後引發一連 串動態效應(Kadowaki et al., 2000; Trinchieri, 2003)。此外,基因組分析和細胞實
驗也注意到β 型干擾素與會誘發一系列對抗 B 型肝炎病毒作用之基因群之間有
密切相關(Wieland et al., 2003; Wieland et al., 2004)。
我們的研究發現當 HBx 基因發生突變,尤其是 1I127V 變異,可以預測 B 肝 感染病史中免疫耐受期的突破。雖然在分組各別分析中,一度發現 B 型肝炎基 因型 B 型、HBx 基因發生 R87V、N118T、或 I127V 突變的患者都會較早進入 到免疫廓清期,但在相互校正各個因子後,僅有 HBx 基因 I127V 突變在多變項 Cox 比例風險模組(Cox proportional hazard model)分析中,具有獨立預測較早突 破免疫耐受期的能力(HR 2.07,95%CI 1.11- 3.85,p= 0.022)。HBx 蛋白已被報 導會降解破壞 MAVS 蛋白,轉而抑制下游的干擾素生成所必須的干擾素調節轉 錄因子(interferon regulatory transcription factor 3, IRF3) (Hoffmann, et al., 2015)。
當 HBx 基因發生突變,可能會使 HBx 蛋白失去附著並破壞 MAVS 蛋白的能力,
進而重新啟動訊號傳遞路徑並且復甦β 型干擾素的製造。以前的研究曾經報導:
牽涉在 HBx 蛋白辨識、並共同黏著至粒線體上 MAVS 蛋白的關鍵是位在其 C 末端(C terminus)的第 111-116 的胺基酸序列。並且學者特別強調 HBx 蛋白中第 115 個位置上的 cysteine (Cys 115)的重要性,一旦 Cysteine 因為核苷酸突變而轉 為生成 Alanine (HBx C115A),則 HBx 蛋白附著上粒線體的能力會完全消失,也 就無法靠近粒線體上的 MAVS 蛋白而破壞它 ( Li et al., 2009; Wei et al., 2010)。
我們的研究結果發現,I127V位在HBx蛋白的C末端,因此對於HBx蛋白附著 並破壞MAVS蛋白的過程也可能有影響。我們推定既然β型干擾素被認為可以有 效的增強宿主的免疫誘發反應,並且有助於後續的後天性免疫系統產生專一的 機制消滅病毒,我們的此項研究或許可以部份解釋T細胞活性復甦恢復,以及接 下來使患者ALT數值因發炎而上升、B型肝炎病毒量下降等數項結束免疫耐受期 的特徵。
另外,我們的研究發現,這155位參與研究的慢性B型肝炎患者,所感染之
13
病毒的基因型以B型及C型為主,此發現符合過去在慢性B型肝炎的研究上,關 於全球地域性及種族上的分布及表現 (Liu et al., 2013)。在調整過各項因素後,
患者感染B型肝炎病毒基因型B型者比起基因型C型者,雖然未達統計意義,但 似乎也有較高趨勢可以較早突破免疫耐受期(HR 1.74,95%CI 0.96- 3.85,p=
0.022)。過去我們的團隊已經發現基因型B型的患者比基因型C型的患者,會較 早經歷e抗原的血清轉換(HBeAg seroconversion) (Ni et al., 2004) ; 而肝功能ALT 上升、突破免疫耐受期常常是e抗原血清轉換的前奏曲,因此我們的發現再次暗 示B肝病毒基因型的重要性,並且它的影響力可能牽涉在整個免疫廓清期之中。
在肝臟免疫組織化學染色檢驗中,除了兩個檢體無法被成功染色外,我們發 現在其它的肝組織切片染色中,HBx蛋白和MAVS蛋白的表現為一個負相關。這 再次印證HBx蛋白的存在會干擾MAVS蛋白的表現。然而,我們無法進一步證明 B型肝炎患者感染病毒之HBx基因序列若有I127V突變,與其肝組織是否可染出 MAVS蛋白表現之間有顯著關連性。由於肝組織切片樣本數太少,即使兩者之 間有所相關,亦不容易看出其顯著性,這一點是本研究主要的侷限性。未來我 們需要更多的病人數量、以及他們所保存的血液及肝臟組織檢體,再進一步做 相關研究跟探討。
根據以上的研究結果,利用此一長期追蹤的慢性B型肝炎感染族群,我們發 現HBx基因對於B型肝炎感染的自然病史,可能扮演一個重要的角色。積極面來 說,HBx基因有I127V突變,或許可以被視為一個保護因子,用來預測免疫耐受 期將會比其他患者提早結束。
五、結論及展望
過去文獻回顧, HBx基因的突變被發現和肝細胞癌的發生與肝細胞癌患者的 預後息息相關(Yeh et al., 2010)。但是有關HBx基因在免疫耐受期所扮演角色的 研究卻很少。另外,慢性B型肝炎感染的自然病史中,關於免疫耐受期是怎麼形 成又怎麼持續的,許多專家學者曾提出不同的見解,卻始終眾說紛紜。我們透
14
過本研究,發現當HBx基因發生突變,或許是我們人類宿主獲得啟動免疫反應、
再次對抗並清除B型肝炎病毒的一個契機。接著,我們預計透過實驗來檢測B肝 病毒野生株、以及I127V的突變株,其HBx蛋白產物對於MAVS蛋白的拮抗力量 是否有所不同,來進一步驗證我們的發現(利用Cycloheximide –chase assay檢驗 HBx蛋白不同的表現,是否使MAVS蛋白的半衰期有所不同)。
未來,這一個研究主題仍有許多未知的面向,需要我們繼續努力探討。我們 希望藉由持續的發掘,能對慢性B型肝炎患者的照護上有所助益。
15
六、論文英文簡述
1. Introduction
1.1 Background
1.1.1 Chronic hepatitis B virus infection and immune tolerance phase
Chronic hepatitis B virus (HBV) infection causes a spectrum of serious diseases including chronic liver disease, liver cirrhosis, and hepatocellular carcinoma (HCC).
In endemic areas like Taiwan, chronic HBV infection initiates mainly during infancy or early childhood through mother-to-infant transmission. This transmission route is particularly dominant in those HBV vaccinated birth cohorts who have breakthrough HBV infection (Chu et al., 1989; Chang, 2008).
During the early phase of chronic HBV infection, immune tolerance status is established in infants and children through interaction of HBV and the young host.
Neonatal tolerance to HBV is probably responsible for viral persistence (Chisari et al., 1995).We postulate that due to the huge amount of HBV transmitted from the mothers, and the immature immune system of the small infants, immune tolerance status is thus established.
It is widely accepted that HBV escapes from innate immunity by inhibiting type I interferon (IFN) production. HBx gene, the smallest of four HBV functional genes, was reported to have a cardinal role in repressing this pathway (Busca et al., 2014).
HBx seemed to modulate a wide range of viral and cellular functions, and its mutant
16
variants were considered to result in a viral escape strategy. In addition, mutant HBx
may also have the oncogenic potential for chronic HBV carriers to develop HCC.
1.1.2 Hepatitis B X (HBx) Gene
HBx gene is the smallest of open reading frames which were partially overlapping with precore and polymerase genes in HBV virus. It had 154-amino acid polypeptide with a molecular weight of 17.5 kDa. HBx appears to have regulatory function with protein-protein interactions, and had been suspected to be a promiscuous
transactivator of cellular and viral promoters /enhancers. It had been suggested to affect viral replication, host cell functions, by modulating a wide of cellular processes including gene transcription, cell cycle progression, genotoxic stress response, DNA damage repair and apoptosis (Bouchard et al., 2004; Benhenda et al., 2009; Bouchard et al., 2004 ).
Its ability to regulate a wide range of genes suggests that HBx could up-regulate the expression of HBV genes by transactivating its own promoters, as well as interact with basal machinary and several transcription factors in the nucleus to mimick the cellular coactivation function, modifying the environment to facilitate viral replication (Wei et al., 2010).
Aside from its transactivating capabilities, another proposed mechanism for HBx had been shown to be activation of several different signal transduction pathways in the cytoplasm, including Ras-Raf-mitogen-activated protein kinase (Ras-Raf-MAPK),
17
extracellular signal-regulated kinase (ERK), the stress-activated protein kinases/
NH2 -terminal-Jun kinase (SAPK/ JNK), protein kinase B (PKB/Akt), and the Janus
kinase /STAT (JAK/STAT) (Benn et al., 1996; Guidotti et al., 2006).
1.1.3 Antiviral innate immunity
1.1.3.1 MAVS system plays a cardinal role in host innate immunity.
Type 1 interferon (IFN) cytokine production and NK cell activation are essential for elimination of invading pathogens from innate immunity. Besides of Toll-like receptors (TLRs) and NOD-like receptors (NLRs) system which can recognize invading pathogens to initiate signaling, another critical arm is launched by the mitochondrial antiviral signaling (MAVS) system (Takeuchi et al., 2009; Hoffmann, et al., 2015). This protein may interact with retinoic acid reducible gene-I (RIG-I) and melanoma differentiation-associated gene (MDA) helicase, and transmit signaling cascade downstream (Bowie et al., 2008). Previous studies showed that several kinds of viruses could block host innate immunity through destroying MAVS protein. For instance, hepatitis A virus (HAV) and hepatitis C virus (HCV) both cleave MAVS, and result in impaired signaling pathway which can activate NF-kB and IFN regulatory factor-3 (IRF3). They use this strategy to repress type 1 IFN production and then evaded host innate immunity (Bowie et al., 2008).
1.1.3.2 Hepatitis B virus evades innate immunity through several strategies
18
For HBV, several mechanisms have been reported to be adopted by HBV to escape the host immunity. HBV core antigen is indicated to repress IFN-β transcription (Thomas et al., 1991). HBV virions, surface antigen, e antigen, and precore antigen have also been shown to abrogate TLR-elicited antiviral process, which subsequently block the expression of IFN-β and IFN stimulated genes (Jun et al,. 2007; Chen et al., 2004; Reignat et al., 2002; Wu et al., 2009). Hepatitis B virus has been shown to use several methods to antagonize first-line interferons and
cytokines induction, in order to block the trigger of antiviral innate immunity.
1.1.3.3 Association between HBx gene and host innate immunity
Since both HAV and HCV can interfere with innate signaling pathway by
targeting MAVS, the role of this protein in HBV infection needs to be clarified. It is recently reported that HBV utilizes its “accessory” protein, HBx protein, to target
MAVS for ubiquitination and proteasomal degradation (Hoffmann, et al., 2015).
The C-terminus of HBx is indispensable for its specific localization on the mitochondria .The key amino acids for MAVS targeting are also mapped to be in C terminal fragment. Substitution mutants may result in disturbed HBx protein, and this may let HBx protein lose the MAVS binding ability (Li et al., 2008; Wei et al., 2010).
Since MAVS protein displays a myriad role initiating a signaling cascade to promote the induction of type 1 IFN, which impacts on innate immunity. We wonder
19
if HBx gene and its subsequent wild or mutant products have unique roles in antiviral
defense ability of innate immunity and HBV immune tolerance phase.
1.2 Unsolved problems
Chronic HBV infected children are considered as mostly in immune tolerant phase. The immune tolerance phase during the natural history of HBV infection from early childhood to young adults is important, yet still remained largely unclear.
1. Factors being involved in the transition of immune tolerance phase to
inflammatory phase remained unknown. Since HBx gene could be involved in HBV virions hiding from innate immunity, we speculated that HBx gene might have some correlations with the sustaining or ending of immune tolerance phase.
Yet the longitudinal follow-up information of HBx gene mutational changes in chronic HBV infected children is lacking. Understanding the role of HBx may enhance our understanding of the immune tolerance and related liver diseases in HBV persistent infection.
2. It is accepted that HBV escapes from host innate immunity by avoiding trigger TLR pathway and IFN production at initial entry. The reports regarding interfering with the other pathway of MAVS protein working in cooperation with
RIG-I-MDA5 were seldom described. Yet efficient intervention to reverse the immune tolerance to HBV is still not applicable. The research in this aspect exploring these signaling transmission routes may be helpful to the future
20
development of appropriate strategies to reverse immune tolerance at early stage,
and may even combat the HBV virus.
1.3 Aim
To investigate the prognostic values of HBx gene and HBx protein on the natural history of HBV infection from childhood to adult life.
2. Subjects and Methods
2.1 Subjects
Children who have been HBsAg-positive for more than 6 months were defined as chronic HBV carriers. A group of chronic HBV carriers who have been diagnosed with HBV infection since early childhood have received long-term follow-up at Pediatrics Department of National Taiwan University Hospital. Series of evaluations including physical examinations, abdominal sonography, HBV markers, liver
function profiles were performed at each 6-month interval visit. If elevated serum ALT levels are detected, the follow-up interval was shorted to 1-3 months, and we defined upper normal limit (UNL) for ALT in children as 30IU/L based on our previous report (Wu et al., 2012).
We also recorded their HBV immunization history, clinical course and outcome, and relevant family histories in detail. The immune-tolerant phase of chronic HBV infection was defined as HBeAg-positive, anti-HBe-negative, and ALT level below
21
UNL. Persisted serum ALT elevation detected twice for at least 6 months apart was defined as breakthrough of immune-tolerance phase and entry into immune-clearance
phase. In addition, clearance of serum HBeAg and emergence of anti-HBe exceeding 6 months in duration was the surrogate markers of HBeAg seroconversion.
In this longitudinal HBV carrier follow-up cohort, we recruited chronic HBV carriers who had been followed for more than 5 years, and who have kept their first available specimen in immune tolerant phase and the last one before ending of immune tolerance phase at our laboratory. Patients who were co-infected with hepatitis C, D, E, or human deficiency virus (HIV) and who used antiviral therapy or immunosuppressant agents were excluded.
In order to assess the association between natural history of chronic HBV infection and HBx gene, the blood specimens collected at initial time at entry to receive regular follow-up and the last time before they breakthrough immune tolerance phase were examined for HBx gene. The study protocol was approved by the institutional review board of the National Taiwan University Hospital, and the
patients or their guardians provided signed informed consent to collect blood samples
and clinical data.
2.2 Methods
2.2.1 HBV markers, serological tests, HBV viral loads, and genotyping
22
Serum samples were obtained at each visit to examine liver function profiles and HBV markers. HBV markers, including HBsAg, antibody to HBsAg (anti-HBs), HBeAg, and anti-HBe were assessed by immunoassay by commercial kits (Abbot Laboratories, North Chicago, IL). HBV viral load and genotype from the first available specimens at the immune tolerant phase were determined using real-time
polymerase chain reaction (PCR) and melt curve assays. Serum alanine
aminotransferase (ALT) levels were determined by autoanalyzer (7450; Hitachi,
Tokyo, Japan).
2.2.2 HBx Gene Sequencing
HBV DNA will be extracted from serum samples from patients with chronic HBV infection, by using a DNA minikit (Qiagen, Hilden, Germany). Nested PCR will be performed to amplify HBV X gene (HBV nucleotides 1374-1839).
The PCR product will be identified in 2% agarose gel electrophoresis stained with ethidium bromide. The nested PCR products will be purified and then sequenced. All sequencing data will be analyzed based on the consensus HBV sequence (strain ayw, NCBI Reference sequence: NC_003977.2).
2.2.3 Liver tissue samples, immunohistochemistry staining for HBx protein and
MAVS protein
Liver tissues from some of our enrolled HBV subjects were obtained. These
23
patients received liver biopsy procedures mostly due to abnormal liver functions during follow up. Paraffin-embedded tissue sections were collected from the
Department of Pathology at National Taiwan University Hospital. Deparaffinization, rehydration and unmasking were performed simultaneously using the Trilogy™
reagent (Cell Marque, Rocklin, CA) with microwave heating for 10 min. To
completely retrieve nuclear antigens, sections were successively incubated in AR-10 antigen retrieval solution, pH 10 (BioGenix, Fremont, CA) and microwave heated for another 10 min. After cooling to room temperature, immunostaining were performed using an UltraVision Quanto Detection System HRP (Thermo Scientific, Fremont, CA) kit. All the primary antibodies used (Supporting Table 1) were incubated with
tissue sections at 4°C overnight, and the staining procedures were performed according to the manufacturer’s instructions.
2.3 Statistics
The X2 test with Yates’ correction will be used to analyze the variables and HBx gene mutants. HBV-DNA levels will be transformed to Log 10 vlues. Student’s t test or the Mann–Whitney U test will be used to test differences in 95% confidence interval/mean or distribution/median between two groups for continuous variables, and Bonferroni correction will be used to adjust for multiple comparisons.
Kaplan-meier survival analysis will be conducted and Cox’s proportional hazard
24
model will be used to predict the factors of breakthrough of immune tolerance phase.
Statistical analyses were conducted using commercially available software Small Stata version 12.0 (Stata Corp., Texas, USA) and SPSS Window version 19 (IBM SPSS Inc., Chicago, IL, USA). All tests were two-sided with significance set at P- value <0.05.
3.
Results
3.1 Breakthrough of Immune tolerance phase
A total of 155 chronic HBV infected patients (105 males and 50 females) were enrolled into our study. The mean age of enrollment was 8.9± 4.5 years-old. They were followed for 23.4± 6.7 years. One hundred and ten subjects had breakthrough of immune tolerance phase at the mean age of 18.3 ±5.2 years-old (table 1). During the inflammatory phase, the mean value of log10 peak ALT level was 2.31±0.41 IU/L. In addition, 81 of these 110 patients achieved HBeAg loss and Anti-HBe emergence, and the mean age was 21.87 ±6.34 years-old, which was about 4.72 ± 3.30 years after ending of immune tolerance phase (Table1).
3.2 HBx gene amplification and sequencing
DNA was successfully extracted from the serum of these 155 patients. Genotype distributions showed a prevailing percentage of genotype B, 120 (77.4%); compared with genotype C, 35 (22.7%). HBV DNA quantification of these 155 subjects revealed mean log10 level of 7.48± 0.68 IU/ml. In addition, the DNA sequences
25
flanking the HBx region were sequenced. Mapping with consensus HBV sequence, point mutations with amino acid changes were identified in 74 sites. Mutations of V44G/L, A66T, R87G, N118T, I127V had highest frequencies, which had been identified in 34.8%, 21.3%, 34.8%, 27.7%, and 36.8% of all 155 patients (Figure 1).
The distribution of these 5 HBx gene mutations was more prominent in genotype B than genotype C.
3.3 Factors associated with duration of immune tolerance phase
We analyzed the relationship between baseline characteristics, HBx gene
mutations of initial blood test, and the duration of immune tolerance phase with
log-rank test and cox proportional hazards model. Kaplan-Meier survival curve
method showed genotype B had significantly higher cumulative proportion of
breakthrough of immune tolerance phase rate than genotype C (log rank, p=0.001).
In addition, the ratios of ending of immune tolerance phase for mutants of R87V,
N118T, and I127V in HBx gene of initial blood samplings were also significantly
higher than those of non-mutants (long rank p values =0.017, 0.044, and 0.001,
respectively). One mutational site, V44G/L, was associated with shorter immune
tolerance phase at a borderline significance level (log rank p=0.074) (Figure 2). After
considering univariate prognostic factors as potential covariates in developing the
multivariate Cox proportional hazard models, we uncovered that ending of immune
26
tolerance phase was independently associated with mutation of I127V in HBx gene
of initial blood sampling with statistically significant level (Hazards ratio (HR) 2.07,
95 % confidence interval (CI) 1.11- 3.85, p=0.022). Genotype B, compared with
genotype C, also contributed to a higher trend to predict earlier breakthrough of
immune tolerance phase, but the p value was insignificant (HR 1.74, 95 % CI
0.96-3.16, p=0.068) (Table 2).
3.4 HBx gene mutations of blood samplings at two different time-point
Fifty-one patients were consistently found to have I127V mutations both at initial blood sampling in immune tolerance phase, and the last blood sampling before ending of immune tolerance phase. However, three cases who did not be examined with HBx I127V mutation at early time were found to have I127V mutation before breakthrough immune tolerance phase. On the contrary, there were six cases found with HBx I127V mutations at initial blood sampling lose such presentation at the time before breakthrough immune tolerance phase. The subgroup of cases who consistently preserved HBx I127V mutations seemed to have higher potential to breakthrough immune tolerance phase at early age than the subgroup that did not be detected with I127V mutations at these two time-points (long rank p values <0.001).
3.5 Liver immunohistochemical staining for HBx protein and MAVS protein
Twenty-three patients (15 males and 8 females) in our enrolled groups have
27
received liver biopsies. The indication for histopathology examination was evaluating the degree of hepatic injury during the immune clearance phase. The mean age when they received liver biopsy was 18.37± 4.09 years-old, and the mean interval between breakthrough of immune tolerance phase and receiving biopsy procedure was 2.84± 2.51 years. We explored their HBx and MAVS proteins expression, and IHC staining revealed three distribution patterns: (1) both negative ( HBx(-)MAVS(-);13.0%, 3/23 cases); (2) HBx protein positive and MAVS protein negative ( HBx(+)MAVS(-); 30.4 %, 7/23 cases); and (2) HBx protein negative and MAVS protein positive ( HBx(-)MAVS(+) ; 56.5 %, 13/23 cases). Except for those liver tissues without being detected with any of two proteins, we found a negative correlation between HBx protein and MAVS expression (Figure 3). However, compared with HBx gene sequencing results, there was no significant association between I127V mutation and hepatic MAVS protein expression (p= 0.414). In addition, the patients with liver biopsies detected as positive MAVS staining did not have higher potentials to end their immune tolerance phases at early stages.
4. Discussion
High viral replication rate is usually found in immune tolerance phase in children with chronic HBV infection, and persistently high viral load with long-term
inflammation may cause liver fibrotic and cirrhotic changes. Until ALT flaring up,
28
which was a surrogate marker of ending of immune tolerance phase, we have no tool to predict the time of entrance of clearance phase. Since HBeAg serocoversion following peak ALT level often indicate better outcome, ending of immune tolerance phase is a noteworthy event (Yang et al., 2002).
From the point of views of host defense, both innate and adaptive immune systems are usually involved in the response to viral infection. Innate responses usually play the initial crucial role for early viral containment and timely efficient induction of specific adaptive response (Fisicaro et al., 2009). However, HBV evolves strategies to inactivate innate signaling and as such is able to establish persistent infection without combating by following adaptive system. This is
believed to be the reason of establishment of HBV immune tolerance phase (Dunn et al., 2009; Luangsay et al., 2015).
Some clinical and experimental evidences showed that HBV-specific T cell immunity are not completely absent in the initial phase of HBV infection, and newborns have been shown to mount virus-specific T cell responses in viral infections (Marchant et al., 2003; Koumbi et al., 2010; Vermijlen et al., 2010). As enhanced activation of components of the innate immunity can help to elicit T cell responses and trigger cytokine-mediated noncytopathic effects, once upstream innate signaling strengthens, it may generate distinctly rapid-rise patterns of adaptive
29
immune responses (Adkins et al., 2004; Levy, 2007). Among the interplay between the two immune systems, type 1 IFN (IFN-α/β) in innate immunity play a cardinal role by not only directly inhibiting viral replications but also indirectly stimulating the differentiation of dendritic cells, subsequently trigger a series of dynamic
responses (Kadowaki et al., 2000; Trinchieri, 2003). In addition, the close association between IFN-β and the induction of hepatocellular genes that may mediate strong anti-HBV effects were also demonstrated by genomic analysis of the livers and hepatocyte cell lines in previous studies (Wieland et al., 2003; Wieland et al., 2004).
Our study emphasized the predictive value of HBx gene mutations, especially I127V variant, for the breakthrough of tolerance phase. Although in the subgroup analysis, patients with genotype B, mutations of R87V, N118T, or I127V in HBx gene had higher chances to enter immune clearance phase at younger age, only
I127V mutation in HBx gene served as an independent prognostic factor in
multivariate Cox proportional hazard model(HR 2.07, 95 % CI 1.11- 3.85, p=0.022).
HBx gene was reported to be capable to degrade MAVS protein, and then in turn
inhibit the activation of interferon regulatory transcription factor 3(IRF3) which was
necessary for IFN-β production (Hoffmann, et al., 2015). Mutant HBx stain may
thwart the binding ability of HBx protein to MAVS protein and thus resume signal
propagation and finally induce IFN-β production. Previous studies once reported that
30
the key amino acid for mitochondria targeting was mapped to HBx C terminus 111-116 amino acids, and they particularly mentioned the importance of the115 amino acid. When HBx 115 Cysteine was mutated to alanine (HBx C115A), the binding property of HBx was impaired (Li et al., 2008; Wei et al., 2010). Our finding was inconsistent with previous finding, but HBx 127 was also located at C terminus, and I127V mutation may also involve in the MAVS targeting process. Since IFN-β has been shown to potently enhance immune response and contribute to subsequent adaptive effector mechanism eliminating viruses, our finding may partially explain the possible T cell restoration in late period of immune tolerance phase.
In our study cohort, genotype B was the prevailing genotype, accounting for 77.4% of the subjects, followed by genotype C (22.7%). This finding was consistent with the previous report regarding geographic and ethnic distributions of HBV genotypes (Liu et al., 2013). After adjustment for other factors, carriers of genotype B also seemed to have a higher trend to enter clearance phase earlier than genotype C
(HR 1.74, 95 % CI 0.96-3.16, p=0.068). Our previous data revealed genotype B had
an advance of HBeAg seroconversion compared with genotype C (Ni et al., 2004).
Elevation of ALT was likely to be the prelude of seroconversion, and our finding
displayed that HBV genotype may also have impacts on this sentinel event.
In immunohistochemical staining of liver biopsy tissues, except for two samples
31
without being stained with any of them, there was a reverse relationship of MAVS protein and HBx protein expressions. This may reconfirm that MAVS protein exhibition would be interfered by HBx protein. However, no significant association between HBx I127V mutation of infected HBV stain and patients’ hepatic MAVS protein expression could be demonstrated in our study, and positive correlation was initially expected to be found. Small sample size of liver biopsy tissues was our limitation, which may obscure the potential relevance. More HBV infected cases and their biopsy tissues should be enrolled and further investigation will be needed.
In conclusion, with the aid of a prospective long term follow up cohort, we found
HBx gene mutations affected the natural course of HBV infection. I127V mutation in HBx gene was an independent factor to predict earlier ending of immune tolerance
phase to enter clearance phase, and it may work by host immune function restoration
through increment of IFN-β.
5. Conclusions and future perspectives
Previously, HBX gene had been identified to be related to hepatocarcinogenesis.
Several HBx gene mutations at specific locations were reported to have prognostic value in HCC-HBV patients (Yeh et al., 2010). However, the information about the role of HBx gene in immune tolerance phase is limited. With the aid of a prospective long term follow-up cohort, we found HBx gene mutations may affect the duration of
32
immune tolerance phase. HBx mutations may re-activate the diminished host innate and adaptive immune responses, and restore T-cell functions to eliminate HBV particles. We are going to perform a cycloheximide -chase assay to examine that whether HBx I127V mutant protein could resume the half-life of MAVS protein, which is downregulated by HBx wild type protein. We expect to verify our findings by further and more examinations.
The reason of persistence or ending of immune tolerance phase in chronic HBV carriers is still unknown, and more and more alternative interpretations regarding the immunopathological events that take place during this early phase have been
proposed. Future studies are required to decipher the mechanisms that underlie to regulate these host-virus interactions. This is an essential step to provide a better care to HBV-infected patients.
33
七、參考文獻
Adkins B, Leclerc C, Marshall-Clarke S. Neonatal adaptive immunity comes of age.
Nat Rev Immunol. 2004;4:553-64.
Benhenda S, Cougot D, Buendia MA, Neuveut C. Hepatitis B virus X protein molecular functions and its role in virus life cycle and pathogenesis. Adv Cancer Res.
2009;103:75-109.
Benn J, Su F, Doria M, Schneider RJ. Hepatitis B virus HBx protein induces
transcription factor AP-1 by activation of extracellular signal-regulated and c-Jun N-terminal mitogen-activated protein kinases. J Virol. 1996;70:4978-85.
Birrer RB, Birrer D, Klavins JV. Hepatocellular carcinoma and hepatitis virus. Ann Clin Lab Sci. 2003;33:39-54.
Bouchard MJ, Schneider RJ. The enigmatic X gene of hepatitis B virus.
J Virol. 2004;78:12725-34.
Bowie AG, Unterholzner L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat Rev Immunol. 2008;8:911-22
Busca A, Kumar A.Innate immune response in hepatitis B virus (HBV) infection. Virol J.
2014;11:22.
Chang MH. Natural history and clinical management of the chronic hepatitis B virus infection in children. Hepatol Int. 2008;2:28-36.
34
Chen MT, Billaud JN, Sallberg M, Guidotti LG, Chisari FV, Jones J, Hughes J, Milich DR. A function of the hepatitis B virus precore protein is to regulate the immune response to the core antigen. Proc Natl Acad Sci U S A. 2004;101:14913-8.
Chen WN, Oon CJ, Leong AL, Koh S, Teng SW. Expression of integrated hepatitis B virus X variants in human hepatocellular carcinomas and its significance. Biochem Biophys Res Commun. 2000;276:885-92.
Chisari FV, Ferrari C. Hepatitis B virus immunopathogenesis. Annu Rev Immunol.
1995;13:29-60.
Chu CM, Karayiannis P, Fowler MJ, Monjardino J, Liaw YF, Thomas HC. Natural history of chronic hepatitis B virus infection in Taiwan: studies of hepatitis B virus DNA in serum. Hepatology. 1985;5:431-4.
Dunn C, Peppa D, Khanna P, Nebbia G, Jones M, Brendish N, Lascar RM, Brown D, Gilson RJ, Tedder RJ, Dusheiko GM, Jacobs M, Klenerman P, Maini MK. Temporal analysis of early immune responses in patients with acute hepatitis B virus infection.
Gastroenterology. 2009;137:1289-300.
Fisicaro P, Valdatta C, Boni C, Massari M, Mori C, Zerbini A, Orlandini A, Sacchelli L,
Missale G, Ferrari C. Early kinetics of innate and adaptive immune responses during
hepatitis B virus infection. Gut. 2009;58:974-82.
Guidotti LG, Chisari FV. Immunobiology and pathogenesis of viral hepatitis. Annu Rev
35
Pathol. 2006;1:23-61.
Hoffmann HH, Schneider WM, Rice CM. Interferon and viruses: an evolutionay arms race of molecular interactions. Trends Immunol. 2015;36:124-38.
Iavarone M, Trabut JB, Delpuech O, Carnot F, Colombo M, Kremsdorf D, Bréchot C, Thiers V. Characterisation of hepatitis B virus X protein mutants in tumour and non-tumour liver cells using laser capture microdissection. J Hepatol.
2003;39:253-61.
Kadowaki N, Antonenko S, Lau JY, Liu YJ. Natural interferon alpha/beta- producing cells link innate and adaptive immunity. J Exp Med. 2000;192:219-26.
Kew MC. Hepatitis B virus x protein in the pathogenesis of hepatitis B virus-induced hepatocellular carcinoma. J Gastroenterol Hepatol. 2011;26:144-52.
Koike K, Moriya K, Iino S, Yotsuyanagi H, Endo Y, Miyamura T, Kurokawa K.
High-level expression of hepatitis B virus HBx gene and hepatocarcinogenesis in transgenic mice. Hepatology. 1994;19:810-9.
Koumbi L, Bertoletti A, Anastasiadou V, Machaira M, Goh W, Papadopoulos NG, Kafetzis DA, Papaevangelou V. Hepatitis B-specific T helper cell responses in uninfected infants born to HBsAg+/HBeAg- mothers. Cell Mol Immunol.
2010;7:454-8.
Kremsdorf D, Soussan P, Paterlini-Brechot P, Brechot C. Hepatitis B virus-related