國立臺灣大學生物資源暨農學院農業化學系 碩士論文
Department of Agricultural Chemistry College of Bioresources and Agriculture
National Taiwan University Master Thesis
利用單核苷酸多型性資料庫及基因編輯技術 建立 OsNramp5 基因突變水稻
Generation of OsNramp5 Dysfunction Rice Mutants Using Single Nucleotide Polymorphism Database and Genome Editing
Technology
林煜翔 Yu-hsiang Lin
指導教授:洪傳揚 博士 Advisor: Chwan-Yang Hong, Ph.D.
中華民國 108 年 7 月
July, 2019
誌謝
碩士生涯過得特別快,感覺前一刻才剛進實驗室還在學怎麼種水稻,這一刻 已經在寫論文致謝了。轉眼間就在農化舊館的 309 室度過了兩年,能在這一刻邊 寫著致謝邊回憶起這兩年的點點滴滴,除了感謝生我養我的父母外,我要特別感 謝我的指導教授洪傳揚老師,謝謝老師包容我這樣一個奇怪的學生,讓我能在實 驗室中學習成長。另外也要感謝實驗室的每一位成員對我的支持與鼓勵,感謝建 志學長、民宇學長、晉宇學長、Harsha 平日對我的指導、幫我莫名其妙的實驗結 果 debug,還有追到女朋友。也謝謝教我這個菜鳥怎麼做 PCR 的佳晴、半夜幫我 種水稻的柏宥、Andrew、為我講解如何做土壤消化的亮心、一起上試驗設計課的 健一、迷路好夥伴的凱彥、抓寶好朋友的上俊、辛苦打 ICP-MS 的致勳、珮雯、崩 潰好戰友的立成、糖心蛋專精的翰霖、尬聊達人的仲凱、衰神胖寶、噪音製造機 亮喻,願意做我在上課、做實驗時的夥伴,陪伴我度過了開心有趣的實驗室生活。
最後還要謝謝楊芥,在我經歷困難時總是陪伴在我身邊,並鼓勵我往目標前進。
謝謝大家,我就讀碩士班的這兩年因為你們而充滿色彩。
中文摘要
鎘 (Cd) 非植物的必須元素,卻容易經由必須元素通道累積在水稻穀粒中,鎘 米會造成人體肝、腎的慢性毒害,因此控制穀粒的鎘含量對於糧食安全極為重要。
在 水 稻 吸 收 鎘 的 機 制 中 , Natural resistance-associated macrophage protein 5 (OsNramp5) 運 輸 蛋 白 是 水 稻 根 部 吸 收 鎘 最 主 要 的 通 道 , 過 去 的 研 究 已 知 , OsNramp5 基因被離子束誘變法剔除時,可大幅降低水稻穀粒鎘含量,而利用基因 編輯技術剔除 OsNramp5 的水稻植株也同樣可獲得含鎘量極低的穀粒,這些結果顯 示 OsNramp5 是影響水稻穀粒鎘累積最關鍵的基因。本研究目的在使台灣本土水稻 品種也具有低鎘累積特性,研究從兩個方向進行:(1)利用單核苷酸多型性資料庫 篩選 OsNramp5 核苷酸具有變異的水稻品系,以作為育成低鎘水稻的種源;(2)應 用基因編輯技術剔除台灣水稻品種的 OsNramp5 基因。從水稻單核苷酸多態性資 料庫中,篩選出 50 個 OsNramp5 核苷酸具有變異的水稻品系,分析這些品系 OsNramp5 胺基酸序列,發現有十個胺基酸序列改變的品系,具有穀粒低鎘累積潛 力,其中 NSF-20、NSF-21 及 Nona Bokra 三個水稻都出現第 505 個胺基酸由丙胺 酸 (Alanine, A, 基因編碼為 GCC) 變為蘇胺酸 (Threonine, T, 基因編碼為 ACC)之 突變,分析鎘處理後 50 個品系幼苗地上部、根部的鎘累積量,發現 8 種鎘含量較 低的品系。基因編輯試驗中,選擇台灣目前栽培面積最廣的台南 11 號、及研究資 源最多的台農 67 號進行基因編輯,針對 OsNramp5 基因第十個外顯子進行編輯,
再利用基因定序篩選編輯成功的植株,未來可將進一步種植這些具有低鎘累積潛 力的水稻於含鎘土壤,以評估具有低鎘累積潛力之水稻品系。
關鍵字: 鎘、水稻、OsNramp5 基因、單核苷酸多態性、CRISPR 基因編輯
ABSTRACT
Cd is a toxic heavy metal and can lead to Cd-related diseases such as renal tubular dysfunction and bone disease. Cadmium (Cd) is not an essential element for plants. However, rice uptakes and accumulates Cd in grains by essential element’s transporters. The accumulation of Cd is a serious threat to human being since it can be concentrated in body through the food chain. Therefore, reducing the Cd content in grains is very important for food safety. Natural resistance-associated macrophage protein 5 (OsNramp5) is the main transporter which uptake Cd in roots. Knocking out of OsNramp5 has been reported to dramatically reduced Cd accumulation in rice grains.
To generate low Cd accumulating Taiwanese cultivars, this research ainmed to carry out in two ways: 1. Screening for osnramp5 dysfunction mutants from single nucleotide polymorphism (SNP) database. 2. Genome-editing the OsNramp5 to Taiwanese cultivars. After screening SNP database from the rice 3000 genomes, 50accessions showed polymorphic genotypes on OsNramp5 genes. Analysis of amino acid sequences from 50 selected accessions showed that 10 accessions contained a change of amino acid. . In addition, three of ten accessions showed the same change at 505th amino acid, where Alanine (Ala, GCC) was substituted by Threonine (Thr, ACC). Examined of Cd concentrations on roots and shoots of rice seedlings showed that 8 lines have lower Cd content in roots or shoots. To generate genome-edited rice plants, two domestic rice cultivars, Tainan 11 (TN11) and Tainung 67 (TNG67), were used to precisely mutagenized the OsNramp5. Tainan 11 (TN11) is the most growing rice cultivars, while Tainung 67 (TNG67) has the most available research resources. Both cultivars’ 9th exon of OsNramp5 was chose for CRISPR/Cas9–based mutagenesis. Mutation was determined by DNA sequencing. Identification of OsNramp5 mutated accessions can be
used as a parent for low cadmium rice breeding. On the other hand, genome-edited rice plants can further screen low cadmium and transgene-free progenies.
Key word:Cadmium、Rice、Nramp5、SNP、CRISPR
目錄
口試委員會審定書 ... #
誌謝 ...i
中文摘要 ... ii
ABSTRACT ... iii
目錄 ... v
圖目錄 ... vii
表目錄 ... viii 縮寫對照表 ...ix
第壹章、緒論 ... 1
一、鎘對人造成的危害 ... 1
二、鎘對植物的影響 ... 2
三、鎘與水稻的關係 ... 3
四、現行鎘污染場址整治 ... 3
五、藉改變栽培方法降低水稻穀粒鎘的累積 ... 4
六、低鎘水稻的育成 ... 5
七、水稻的核苷酸多態性 ... 6
八、基因編輯技術 ... 7
第貳章、研究目的 ... 10
第參章、材料與方法 ... 11
一、植物材料與生長條件 ... 11
二、DNA 定序 ... 11
三、基因表現分析材料準備與處理 ... 13
四、基因表現分析 ... 14
五、水耕試驗及幼苗鎘含量分析 ... 15
第肆章、結果 ... 17
一、OsNramp5 的基因結構與胺基酸突變位點 ... 17
二、50 株栽培種的相關資訊 ... 17
三、50 株栽培種的基因定序及胺基酸變化 ... 17
四、50 株栽培種的 OsNramp5 表現量 ... 18
五、鎘處理下水稻幼苗的植體鎘含量 ... 18
六、CRISPR 轉植株的轉基因檢測 ... 18
七、CRISPR 轉植株的基因編輯成果 ... 18
第伍章、討論 ... 19
第陸章、參考文獻 ... 21
第柒章、附錄 ... 24
圖目錄
圖 1. OsNramp5 的基因結構 ... 25
圖 2. SNP 資料庫中胺基酸的改變位置 ... 26
圖 3. 50 株栽培種的胺基酸序列比對 ... 27
圖 4. 50 株栽培種的 OsNramp5 表現量比較 ... 28
圖 5. 50 株水稻幼苗的地上部、根部鎘濃度 ... 29
圖 6. CRISPR/Cas9 系統載體構築及目標序列 ... 30
圖 7. CRISPR 轉殖株的轉基因分析 ... 31
圖 8. CRISPR 轉殖株的基因編輯成效 ... 32
表目錄
表 1. 50 株栽培種的資料 ... 33 表 2. 50 株栽培種的 SNP 位置 ... 35 表 3. 論文中使用引子表 ... 36
縮寫對照表
Cas CRISPR-associated proteins
Cd Cadmium
CdS Cadmium sulfide
CRISPR Clustered Regularly Interspaced Short Palindromic Repeats HMA heavy metal ATPase
ICP-MS Inductively coupled plasma mass spectrometry IRRI International Rice Research Institute
IRT iron-regulated metal transporter LCR low-molecular-weight cysteine-rich lcd-kmt low-Cd Koshihikari mutants
MT metallothionein
NHEJ non-homologous end-joining
NSFTV National Science Foundation-"Exploring the Genetic Basis of Transgressive Variation in Rice”
Nramp Natural resistance-associated macrophage proteins PCR Polymerase chain reaction
ROS reactive oxygen species
SNP single nucleotide polymorphisms TAE buffer Tris-Acetate-EDTA buffer
TN11 Tainan 11
TNG67 Tainung 67
第壹章、 緒論
一、 鎘對人造成的危害
鎘 (Cadmium, Cd) 是外觀為銀灰色的過度金屬。由於密度大於 5 g/cm3 ,鎘 與砷 (As) 、鉻 (Cr) 、銅 (Cu) 、汞 (Hg) 、鎳 (Ni) 、鉛 (Pb) 及鋅 (Zn) 等元 素一同被歸類為重金屬。上述八種重金屬由於其危害性,被我國環保署訂定為土 壤管制的檢驗對象,其中鎘運用於製作電池、金屬電鍍、塑膠製造及電子組件生 產,倘若產業的廢水沒有妥善處理,鎘將隨著污水進入環境中。由於鎘並非會自 行分解的污染物,污染場址不經由環境整治很難恢復正常。前人文獻指出鎘具有 致癌能力(Takao et al., 1996),長期暴露於含鎘環境會造成人體的胃、腎臟、肝臟等 器官的病變(Cancer, 1993; Waalkes and Berthan, 1995; Waalkes et al., 1996)。隨食物 鏈進入人體的鎘會與金屬硫蛋白 (metallothionein, MT) 結合並累積於肝臟、腎臟 (Klaassen, 1981; Klaassen et al., 1999)。雖然藉金屬硫蛋白螯合可以減少鎘的危害,
但這也造成鎘難以自人體移除,延長鎘的生物半衰期(Takao et al., 1996)。
鎘毒害最早的紀錄可回溯至 1950 年代。日本九州富山縣為取得原物料進行大 規模採礦活動,並將採礦及冶礦的廢水排放入神通川及其支流,造成該流域受到 礦業廢水的污染。由於神通川是當地的飲用水及灌溉水源,該地的民眾因長期攝 取受鎘污染的食物及水,身體出現骨質疏鬆、腎功能衰竭等問題,其中嚴重者甚 至會因關節和脊椎骨極度疼痛而行動不便。此病徵由當地醫生診斷為慢性鎘中 毒,並以患病者疼痛的喊聲命名為痛痛病。此為鎘損害人體健康的首例(Inaba et al., 2005)。
1960 年代臺灣推行「客廳即工廠」的政策,鼓勵家庭代工發展,因此小型工 廠零星散佈於農地間。由於當時排水系統及灌溉系統並未分流,工業廢水就近排 放並污染灌溉系統。農田長期以污染水灌溉,日積月累下造成農作物的重金屬含 量超標。於 1980 年代,台灣的彰化縣、台中縣、雲林縣及桃園縣陸續檢出鎘米。
衍生出的食安問題嚴重影響我國人民健康。此後臺灣開始重視重金屬污染防治議 題。
二、 鎘對植物的影響
植物主要依靠根部來吸收必須元素,其中金屬元素多以離子的形式進入植 體。植體中的鎘元素大多源於被工業廢水污染的土壤,並以正二價鎘離子的形式 被植物吸收。雖然鎘並非植物的必須元素,甚至會對植物造成毒害,然而鎘離子 的化學性質與鋅、鐵、錳離子等必須元素相似 (Nakanishi et al., 2006; Pedas et al., 2008; Sasaki et al., 2012),植物的運輸蛋白無法區分何者為必須元素,而錯將鎘吸 收。進入植體後鎘危害植物的機制可分為 (1) 降低植物養分吸收 (2) 降低植物水 分吸收 (3) 破壞植物氧化還原平衡。
2.1、降低植物養分吸收
如同前文所述,鎘並非植物的必須元素,植體中並沒有專門負責吸收鎘的轉運 蛋白,鎘與鋅、鐵、錳等必須元素競爭進入植體的途徑,造成植物對該類養分的 吸收減少。舉一些常見作物為例,鎘處理會降低豌豆的鉀、錳吸收量(Hernandez et al., 1996; Pandey and Sharma, 2002)、降低高麗菜的鐵吸收量(Pandey and Sharma, 2002)、降低水稻錳的吸收量(Ishimaru et al., 2012)。這些元素雖然並非植物的主要 成分,但會作為酵素或輔酶參與植物多種生理反應。如鐵、錳協助光合作用,鉀 協助植物細胞的勢能平衡。缺乏必須營養時會造成植物生理反應混亂,進而抑制 植物生長。
2.2、干擾植物蒸散速率
水分由根部至葉片的運輸動力主要依靠蒸散作用,當水分被根部吸收後,會經 由木質部運輸至葉片,最後從氣孔回歸大氣中。此過程在維管束中形成向上的拉 力,是植物運輸水分及水溶性養分的動力來源。鎘會改變葉片的水分含量及擾亂 葉片氣孔的開闔,進而影響蒸散速率。舉例來說:在菜豆 (Phaseolus vulgaris L. cv Contender) , 鎘 會 降 低 葉 片 的 水 含 量 , 提 升 氣 孔 阻 力 使 蒸 散 速 率 降 低 (Poschenrieder et al., 1989)。在蠶豆 (Vicia faba L.) ,鎘會與鈣競爭鈣通道蛋白,干 擾保衛細胞的滲透調節 (Perfus‐Barbeoch et al., 2002)。當植物蒸散拉力遭到擾亂,
會使養分及水分缺乏輸送動力,造成植物黃化或凋萎。
2.3、生成活性氧化物
當鎘進入植物細胞的葉綠體或粒腺體,會干擾胞器中電子傳遞鍊的反應,導致 超氧陰離子自由基 (superoxide radical, O2.–)、氫氧自由基 (hydroxyl radical, . OH) 、過氧化氫(hydrogen peroxide, H2O2) 等活性氧化物 (reactive oxygen species, ROS) 大量生成 (Møller, 2001)。由於 ROS 的反應性很高,在細胞內會作為氧化反 應的電子接收者攻擊周遭,令細胞膜、蛋白質、核酸受損 (Foyer and Noctor, 2005)。
隨著 ROS 濃度提高,植物會陷入氧化逆境而無法進行生長所需的生理反應。
上述鎘造成的生理危害會反映於植物的外表型態,若和無鎘毒害的植株比 較,可觀測到鎘會抑制種子的萌芽、分化、根部的延長、葉綠素的生成以及導致 新葉黃化(Chmielowska-Bąk et al., 2014; Iqbal et al., 1991)。長期的生理干擾與活性 氧化物累積會導致植株矮小、生質量下降甚至死亡。即使不考慮人體健康危害,
單就作物的生長與產量而言,鎘毒害也是相當重要的議題。
三、 鎘與水稻的關係
鎘被水稻根部吸收後,經過維管束運輸至地上部,最後累積於穀粒中。由於 鎘毒性不會立即導致植物死亡,即使是鎘污染環境中的水稻,也有可能成長至足 以結出成熟穀粒,也就是所謂的鎘米。雖然穀粒所含的鎘含量不高,但鎘的生物 半衰期極長,一旦攝取入體內很難藉由新陳代謝排出。再加上米飯為亞洲地區的 主食,即使是低鎘含量的穀粒,長期的攝取鎘米仍能累積到足以危害人體的劑量。
考量到國人飲食習慣及鎘的毒性,我國的食米鎘管制標準為 0.4 ppm。
四、 現行鎘污染場址整治 4.1、土壤污染管制標準
我國鎘污染場址的管制標準為 20 mg/kg soil,監測標準是 10 mg/kg soil,如果是 栽種食用作物的農地則會更加嚴格,管制標準及監測標準分別為則是 5 mg/kg soil 和 2.5 mg/kg soil。當土壤鎘濃度超越管制標準時,該處會被公告為污染控制場址,
於管制改善期間除了農地必須停止耕作外,受到污染的作物則需銷毀以防止流入 市面。
4.2、污染場址整治方式
現今污染場址的整治方法可依處理位置分為現地整治與離地整治,又可依處理 的反應機制細分為 (1) 固化法:注入固化劑或穩定劑,使污染物被包覆吸附並降 低生物活性。 (2) 玻璃化法:在土壤中插入電極後通高壓電產生高溫,造成有機 污染物無氧分解,金屬類污染物被玻璃狀物質固定。 (3) 淋洗法:依照污染物性 質添加介面活性劑、螯合劑或是酸,將污染物從土壤中淋洗出來,如果是現地整 治的話需要針對混入洗出液的地下水收集處理。 (4) 熱脫附法:針對易揮發的有 機物或是汞加熱土壤,收集脫附的污染物後再以其他系統處理。 (5) 排土/客土 法:排土法是將污染土壤挖除,客土法則是混入從他處取得無污染土,物理性減 少污染物濃度。 (6) 植生復育:利用植物或其根系的微生物,對土壤中污染物吸 收、固化、轉劃分解。
4.3、整治方式的實施問題
我國以土壤中污染物的全量為規範標準,因此不將污染物移除,僅是降低生物 活性的固化法、玻璃化法等整治方式不適用於台灣農地,且固定的污染物有可能 因環境變動再次釋出。淋洗法則需考慮土壤的透水性及當地的水文條件,否則可 能造成淋洗不足污染物殘留,或是污染物隨地下水擴散形成二次污染等問題。植 生復育法則會受限於生物因素:整治深度依植體根系長度決定、整治濃度上限依 植株耐受程度決定、整治時間依植物生長速度及生質量決定等,是在廣泛應用前 仍需研發的技術。考量現有技術及成本,排土/客土法為最常使用的整治方式,但 排土法的廢土處置及客土法場址的污染物仍是有待解決的問題。
五、 藉改變栽培方法降低水稻穀粒鎘的累積 5.1、環境因素對鎘移動性的影響
前述整治方法是從環境層面減少作物的鎘吸收,但考量到成本及可行性,很難 有令人滿意的結果。為了有效降低穀粒鎘含量,科學家改從離子的移動性機制著 手:改變環境因素以降低鎘的移動性。舉例來說,鎘的溶解度受土壤的氧化還原 電位 (Eh 值) 影響 (Iimura, 1978),當土壤浸水處於還原態時會生成硫化氫,解離 的硫化氫會與鎘形成難溶的硫化鎘 (CdS) 沉澱 (Ksp = 5.0 *10-28)。反之,經過排
水的土壤由於成氧化態,其中的硫多以硫酸根的形式存在,無法將鎘固定不被水 稻吸收 (Ito and Iimura, 1976)。因此可藉由稻田的水分管理調控土壤 Eh 值,進而 降低水稻的鎘吸收量。日本曾利用此機制,制定了抽穗前後三周浸水灌溉稻田的 對策,藉此成功降低稻米的鎘含量,目前約有 40000 公頃的稻田受到此規範管制 (Ishikawa et al., 2016)。
5.2、管理方式的實施問題
由於土壤中的反應機制牽涉許多層面,且土壤基質中含有許多可能受反應影響 的成分,就單一機制來擬訂污染對策可能會造成預估外的風險。舉例來說:雖然 水分管理可以確實降低穀粒鎘含量,但假如鎘污染的土壤中也含有砷,淹水環境 會令砷偏向氧化價數較低的三價態,造成砷在水稻的移動性增加並導致穀粒中的 砷含量上升 (Ishikawa et al., 2016)。
六、 低鎘水稻的育成
因為環境整治與栽培管理法各自有其限制,科學家仍在尋找有效減少穀粒鎘含 量的方法。近年由於分子技術的進步,有許多科學家改從作物細胞的運輸蛋白著 手,希望藉此途徑降低作物的鎘吸收。
6.1、參與鎘運輸的運輸蛋白介紹
為從養分吸收機制控管鎘的吸收,理解吸收機制的原理可以達到事半功倍的效 果。先前已有提到植物藉由運輸蛋白吸收鎘,水稻植體內鎘的運輸可詳細分為根 部的吸收、根部至地上部的轉運以及地上部的再移動 (remobilization) ,各個過程 有不同運輸蛋白參與。在根部,OsIRT1 (iron-regulated metal transporter 1),OsNramp1 及 OsNramp5 皆會將土壤溶液中的鎘運輸至根細胞中,即參與根部的鎘吸收。而 在木質部,OsHMA2 (heavy metal ATPase 2) 會將根細胞中的鎘裝載入木質部,也 就 是 參 與 根 部 至 地 上 部 的 轉 運 。 在 地 上 部 , OsLCR1 (low-molecular-weight cysteine-rich1) 會參與鎘的再移動,包括生殖生長期將鎘輸送至穀粒。其中影響根 部鎘吸收的 OsNramp5 ,已被證實缺失該蛋白功能的突變株穀粒鎘累積量會顯著 減少(Sasaki et al., 2012)。
6.2、Nramp5 家族及功用
Nramp 家族 (Natural resistance-associated macrophage proteins) ,自然免疫相關 巨噬細胞蛋白家族。第一個發現的 Nramp 蛋白位於哺乳類細胞膜並影響細胞對於 抗原的敏感性 (Vidal et al., 1993)。後續研究中,科學家發現 Nramp 家族可作為轉 運蛋白,運輸 Mn2+ ,Zn2+ ,Cu2+ ,Fe2+ ,Cd2+ ,Ni2+ ,Co2+ 及 Al3+ 等金屬離子(Colangelo and Guerinot, 2006; Nevo and Nelson, 2006; Xia et al., 2010),顯示這些蛋白參與了金 屬離子的吸收、轉運、細胞間運輸、排毒 (Nevo and Nelson, 2006)。以模式生物阿 拉伯芥 (Arabidopsis thaliana) 為例,阿拉伯芥的基因組有六種不同的 Nramp 蛋 白,酵母菌的蛋白表現實驗顯示 AtNramp1, 3, 4 具有運輸 Mn2+,Fe2+ 及 Cd2+ 等金 屬離子的能力(CURIE et al., 2000; Lanquar et al., 2005; Thomine et al., 2003;
Thomine et al., 2000)。這些蛋白在阿拉伯芥的生長過程中參與根部的錳吸收和細胞 中鐵的平衡。
6.3、nramp5 突變株與鎘的關係
日本學者以越光米 (Koshihikari) 為背景,利用碳離子束照射誘發水稻突變,篩 選 出 三 株 鎘 累 積 量 低 的 突 變 株 後 命 名 為 lcd-kmt1, 2, 3 (low-Cd Koshihikari mutants) 。倘若將篩選出的突變株與正常株種植於鎘污染土中 (1.8 mg/kg) ,突變 株的穀粒鎘含量皆小於 0.05 mg/kg,小於食米鎘濃度標準 0.4 mg/kg,而正常株平 均穀粒鎘含量為 1.73 mg/kg,此外突變株與正常株不同處僅有穀粒鎘含量低的性 狀,其他諸如植株型態、稻稈重、食米品質等農藝性狀皆與正常株相同。經過基 因鑑定,發現在突變株的 OsNramp5 基因有缺失 (deletion) 或插入 (insertion) 突 變:lcd-kmt1 在 exon X 有插入突變,lcd-kmt2 在 exon IX 有缺失突變,lcd-kmt3 則 為整段 Nramp5 缺失 (Ishikawa et al., 2012),這項結果顯示 OsNramp5 基因功能的 有無會大幅影響水稻的鎘吸收量,若能從基因層面著手令 Nramp5 失去作用,即 可獲得低鎘累積的水稻。
七、 水稻的核苷酸多態性 7.1、水稻相異性狀的來源
自然界族群中的多種性狀源自於等位基因的不同,等位基因的變化又可分為簡 單 重 複 序 列 (simple sequence repeats, SSRs) 、 節 段 插 入 / 刪 除 (segmental insertions/deletions) 及單一核苷酸多型性 (single nucleotide polymorphisms, SNPs) (Mammadov et al., 2012),倘若這些變化改變轉譯出蛋白質的表現量或結構,便會 使基因持有者產生不同的性狀。水稻的族群中有許多不同性狀的栽培種,其中可 能存在 OsNramp5 基因序列不同的水稻品系,倘若有低鎘累積性狀的野生型水 稻,可作為親本株提供育種使用。藉由雜交、回交分離出缺陷的 OsNramp5 基因,
導入欲改良的品種中,即可育成同時具有良質米性狀且穀粒鎘累積量低的水稻。
7.2、單一核苷酸多態性資料庫
在 2005 年,國際水稻基因體定序計劃 (International Rice Genome Sequencing Project, IRGSP) 完成了針對日本粳稻品種日本晴 (Nipponbare) 全基因體的 DNA 序列定序,為水稻的分子育種技術奠定了基石。學者接著投入比較稻屬物種基因 體計畫 (The Oryza Map Alignment Project, OMAP) ,針對諸多栽培種、野生種水 稻進行全基因體定序,統合多種水稻的基因體序列後完成 SNP 資料庫。在 SNP 資 料庫中育種家可以針對目標基因搜索 SNP 的品種、SNP 位點、核苷酸變化等資 訊,用以挑選合適的 SNP 品種做為育種種源。相較於傳統育種需要在大量的親本 株中尋找適合的種源,SNP 資料庫可以先就基因層面做篩選,減少選擇親本株的 成本與時間。
八、 基因編輯技術
除了在自然中尋找不同基因型的種源,新興的基因編輯技術也可以改良現有 栽培種,培育出具有目標性狀的植株。相較於尋找自然的野生種導入基因,基因 編輯可以直接破壞或導入基因功能,節省尋找適合種源的時間。
8.1、常見基因編輯技術
現今常見的基因編輯技術有鋅指核酸酶 (zinc-finger nucleases) 、類轉錄活化 因子核酸酶 (Transcription Activator-Like Effector Nucleases, TALEN) 、常間回文重 複 序 列 叢 集 /CRISPR 關 聯 蛋 白 系 統 (Clustered Regularly Interspaced Short
Palindromic Repeats/CRISPR-associated proteins, CRISPR/Cas system) 等,上述基因 編輯的原理為引導核酸酶至目標基因,剪切雙股 DNA 後啟動細胞修復機制,修復 機制可分為非同源性末端接合 (non-homologous end-joining, NHEJ) 及同源重組 (homologous recombination, HR)。其中非同源性末端接合由於沒有參照模板,修復 後的基因容易出現插入或缺失突變,即可達成敲除基因的目的。
8.2、CRISPR 系統的由來
CRISPR 為細菌對嗜菌體的後天免疫系統,由多種噬菌體的基因片段和核酸酶 的基因組成,當細菌遭受噬菌體攻擊且存活下來,便會將病毒的基因片段納入 CRISPR 系統,此後細菌即可藉由該基因組辨識外來的 DNA 或質體。當同種噬菌 體再次將 DNA 注入細菌,便會啟動防衛機制轉錄出對應的 single guide RNA (sgRNA),sgRNA 與核酸酶形成複合體後,會辨識外來基因並引導核酸酶剪切目 標,藉此防禦嗜菌體攻擊。
8.3、CRISPR 的應用
目前 CRISPR 技術在水稻、阿拉伯芥、菸草及蕃茄等植物中皆有應用,科學 家將 CRISPR 基因組內的基因片段改成想要編輯的目標基因,將設計好的 CRISPR 質體送入農桿菌中,再藉由農桿菌感染目標將 CRISPR 系統的基因嵌入植物 DNA 中,此時 CRISPR 系統對目標基因的精準性為科學家所用,令 sgRNA 引導核酸 酶至目標基因剪切,引發 NHEJ 的機制使 DNA 在修補過程中產生突變。由於相較 於其他 zinc-finger nucleases、TALEN 基因編輯技術,CRISPR 系統具有操作容易、
設計簡單等優點,目前被廣泛應用於遺傳工程中,提供給使用者準確且有效的基 因編輯工具。
8.4、CRISPR 編輯水稻
在日本成功製造出低鎘累積稉稻後,中國的學者參照成功突變位置設計 CRISPR 載體,針對國內較常食用的秈稻敲除 OsNramp5 基因,並成功獲得具有低 鎘累積性狀的秈稻品種,即使將這些 CRISPR 編輯株種植於鎘污染的田地,穀粒 的鎘累積量仍小於 0.05 mg/kg,遠低於中國食米管制標準 0.02 mg/kg。這項試驗也
證實無論在稉稻還是秈稻品系中,OsNramp5 是影響鎘吸收的關鍵基因(Tang et al., 2017)。
第貳章、 研究目的
鎘為有毒性的重金屬之一,具有極長的生物半衰期。食物中的鎘會進入人體 並造成慢性毒害,因此控制農作物的鎘含量對於食品安全極為重要。在作物吸收 鎘的機制中,運輸蛋白扮演重要的角色。以水稻為例:OsNramp5 的蛋白位於水稻 根部,負責將土壤中的鎘運輸至根部細胞中。前人研究指出含鎘環境中,osnramp5 水稻突變株的穀粒鎘含量顯著低於正常株,這證實從基因層面抑制鎘累積是可行 的方法。
本研究目的為獲得穀粒鎘累積量低的台灣水稻品系,分別使用兩種方法:(1) 利用單核苷酸多型性資料庫篩選 OsNramp5 核苷酸具有變異的水稻品系,以作為育 成低鎘水稻的種源;(2)應用基因編輯技術剔除台灣水稻品種台農 67 號及台南 11 號的 OsNramp5 基因,以育成無轉基因的本土低鎘水稻品種。
第參章、 材料與方法
一、 植物材料與生長條件 1.1、植物材料來源
本試驗使用水稻根據國際稻米研究所 (International Rice Research Institute, IRRI) 的單核苷酸多態性資料庫 (Rice SNP-seek database) 篩選,以日本晴 (Oryza sativa L. cv. Nipponbare) 的基因序列作為比較基準,選取在 Nramp5 的序列中有鹼基變 化的栽培種。各栽培種的編號採用 National Science Foundation-"Exploring the Genetic Basis of Transgressive Variation in Rice" (NSFTV,其後以 NSF 簡稱) 之種子 編號,種子來自農藝系董老師實驗室。CRISPR 植株來自同實驗室黃建誌學長製 做植株。
1.2、植株栽培條件
挑取無蟲咬、無發芽之水稻種子,依編號放入發芽盤中,浸泡於 2% 漂白水表 面消毒 30 分鐘,剔除空心浮於水面的種子,以自來水沖洗 5 次至無消毒水的氣 味殘留,添加適量水分 (不淹過水稻表面) 後放入 37℃ 培養箱中發芽兩天。將發 芽種子放置鐵網架上,移入 30 L 塑膠方盆中,並添加木村氏水耕液供給成長所需 養分,水稻幼苗置於人工氣候室 (日 30℃/夜 25℃) 照射自然光,每隔兩天更換 水耕液。待所有幼苗第三葉完全展開後進行後續處理。
二、 DNA 定序 2.1、DNA 萃取
取 1~2 片水稻葉片放入 2 mL 微量離心管,加入小鋼珠並以均質機 1200 rpm 研 磨 1 分鐘,樣品粉末加入 0.7 mL Urea extraction buffer 及 0.7 mL phenol: chloroform (1:1) 靜置 15 分鐘,使用離心機以 8000 rpm 離心 10 分鐘後,將上清液加入含 70 μL NaOAc 及 0.7 mL Isopropanol (IPA) 的微量離心管混勻,以微量移液管勾取 DNA 放入含 1 mL 70% 酒精的微量離心管清洗雜質,洗淨後倒掉酒精風乾 DNA,加入 50 μL ddH2O 回溶。取 2μL DNA 與 loading dye 混合,加入 1% 瓊脂 膠體 (含 ethidium bromide) 於 TAE buffer 中以 100 伏特電泳 20 分鐘,確認有無
DNA 條帶。
2.2、DNA 片段製備
利用 NCBI 的 primer designing tool 設計定序用引子 (Nramp5-GeFw/Rv),定序位 置由第 13 個外顯子 (exon) 至 3'非編碼區 (3'UTR) (附圖)。以前述 DNA 為模板,
將下列反應物加入微量離心管中,進行聚合酶連鎖反應 (Polymerase chain reaction, PCR) 。
per sample
DNA 0.5 μL
2 mM dNTP 2.5 μL
25 mM MgSO4 1μL
10 μM Fw primer 0.75 μL 10 μM Rv primer 0.75 μL 10 X taq buffer 2.5 μL
ddH2O 16.5 μL
KOD taq polymerase 0.5 μL
Total 25 μL
Step 反應條件如下:
1 94℃ 加熱 2 min
2 94℃ 加熱 15 sec
3 60℃ 加熱 30 sec
4 68℃ 加熱 30 sec (重複 step 2→4,循環 39 次)
5 68℃ 加熱 7 min
6 16℃ 暫停反應
取 20 μL PCR 產物與 loading dye 混合,加入 1% 瓊脂膠體 (含 ethidium bromide, EtBr) 於 TAE buffer 中以 100 伏特電泳 20 分鐘,確認有無 552 bp 之 PCR 產物 條帶。
2.3、PCR 產物回收
將電泳膠體置於 UV 燈下,以刀片切下含有 PCR 產物的膠體,放入 1.7 mL 微 量離心管中,利用 GeneDireX PCR Clean-Up & Gel Extraction kit 自膠體中回收 PCR 產物。加入 500 μL Binding solution 後放置於 60℃ 加熱槽 10 分鐘,加熱期 間將 spin column 裝於收集管上,膠體溶解後倒入 spin column , 13000g 離心一 分鐘後丟棄廢液,加入400 μL wash solution 1,13000g 離心一分鐘後丟棄廢液,
再加入600 μL wash solution 2,13000g 離心一分鐘後丟棄廢液,再 13000g 離心兩 分鐘去除殘留的 wash solution 2 ,將 spin column 裝於 1.7 mL 微量離心管上,加 入30 μL ddH2O 回溶 PCR 產物兩分鐘,再 13000g 離心兩分鐘得到 PCR 產物。
2.4、DNA 定序
PCR 產物交託明欣生物科技有限公司 (Mission Biotech) 定序。
三、 基因表現分析材料準備與處理 3.1、RNA 萃取
剪取三周大水稻根部擦乾,以鋁箔紙包覆並保存於液態氮中,研缽經酒精擦拭,
以液態氮預冷後開始研磨植體,研磨過程中適時添加液態氮防止回潮,植體粉末 使用藥杓刮入微量離心管,加入 1 mL TRIzol 後靜置 5 分鐘,再加 0.2 mL chloroform 靜置 2~3 分鐘,使用離心機在 4℃ 下 12000g 離心 15 分鐘,取上層 水相溶液移入新的微量離心管,加入 0.5 mL IPA 後靜置 10 分鐘,再次使用離心機 於4℃ 下 12000g 離心 10 分鐘並丟棄懸浮液,加入 1 mL 75% 酒精並輕微震盪,
使片狀沉澱物不黏著於管壁,倒掉酒精風乾 5 至 10 分鐘,加入 30 μL RNase-free H2O 回溶 RNA。取 2 μL RNA 與 loading dye 混合,加入 1% 瓊脂膠體 (含 ethidium bromide) 於 TAE buffer 中以 100 伏特電泳 20 分鐘,將膠體置於 UV 箱中觀察 RNA 品質。
3.2、合成第一股 cDNA
以 MMLV Reverse transcriptase 1st-Strand cDNA Synthesis Kit 進行。定量 RNA
per sample RNA (1000 μg) X μL 100 μM oligo-dT 1 μL
ddH2O 9-X μL
Total 10 μL
RNA 接合引子後靜置於冰上 1 分鐘,加入 cDNA 合成試劑混合均勻,於 37℃
反應 60 分鐘。
per sample Annealed RNA 10 μL 10X RT-script buffer 2 μL
10 μM dNTPs 1 μL
RT-script enzyme 1 μL RNase-free H2O 6 μL
Total 20 μL
反應完成後,於85℃ 加熱 5 分鐘終止反應,保存於 -80℃ 冰箱用於後續實 驗。
四、 基因表現分析
4.1、半定量反轉錄聚合酶連鎖反應 (semi-quantitative RT-PCR)
以前述 cDNA 為模板,並設計 cDNA RT-PCR 的引子 (產物 572 bp) ,增強位 置由第 12 個外顯子至 3'非編碼區 (附圖)。,將下列反應物加入微量離心管中進行 PCR,另外設計 Actin 的引子 (產物 444 bp) ,以常態表現的 OsActin1 為比較基 因表現量的基準。
per sample
cDNA 2 μL
10 mM dNTP 0.4 μL 10 μM Fw primer 0.4 μL
10 μM Rv primer 0.4 μL 10 X taq buffer 1 μL
ddH2O 5.6 μL
YEA taq polymerase 0.2 μL
total 10 μL
Step 反應條件如下:
1 94℃ 加熱 5 min
2 94℃ 加熱 30 sec
3 55℃ 加熱 30 sec
4 72℃ 加熱 30 sec (重複 step 2→4,循環 39 次)
5 72℃ 加熱 10 min
6 16℃ 暫停反應
取 10 μL PCR 產物與 loading dye 混合,加入 1% 瓊脂膠體 (含 ethidium bromide, EtBr) 於 TAE buffer 中以 100 伏特電泳 20 分鐘,將膠體置於 UV 箱中 比較 444 bp (Actin) 及 572 bp (PCR 產物) 條帶的明暗程度。
五、 水耕試驗及幼苗鎘含量分析 5.1、水耕液鎘處理試驗
取三周大之水稻作試驗材料,水耕液中添加5 μM CdCl2 (每隔三天更換含鎘水 耕液) 處理一周,處理結束後以 5mM CaCl2 溶液浸泡根部 5 分鐘,將水稻植株抽 離鐵網並以 ddH2O 清洗根部,以擦手紙吸收殘留水分後分為地上部及根部,放入 70℃ 烘箱烘乾。
5.2、水稻植體消化
取烘乾的植體稱重並記錄,樣品剪碎倒入分解管中,於抽氣櫃中加入 70% 濃 硝酸 8 mL 使樣品浸於硝酸中,使用 Anton Paar 的微波消化儀 (Multiwave PRO) 消化植體,操作條件及步驟如下。
Max. Pressure Increase Rate 0.5 bar/s IR Temperature Limit 180℃
Max. Pressure 40.0 bar Internal Temperature Limit 210℃
Max. Microwave Power 1500 W
Step 反應條件如下:
Step Power Time
1 Power ramp 600 W 5 min
2 Power hold 600 W 10 min
3 Power ramp 800 W 10 min
4 Power hold 800 W 10 min
5 Cooling 0 W
待消化液冷卻後倒入 50 mL 定量瓶中,以 ddH2O 潤洗分解管併入消化液,再 以 ddH2O 定量至 50 mL 標線,以 Advantec 1 號濾紙重力過濾至樣品瓶中。
5.3、檢量線製備
精確量取鎘標準品 (1000 mg/L) 5 mL,以 0.5 N 硝酸溶液定容至 50 mL (100mg/L) ,重複上述過程序列稀釋,製成 1 mg/L 及 0.1 mg/L 鎘濃度的檢量線標 準溶液,另精確取 10 mg/L 之溶液 4 mL、2.5 mL、1 mL,以 0.5 N 硝酸溶液定容 至 50 mL,製成 0.8 mg/L、0.5 mg/L、0.2 mg/L 等鎘濃度檢量線標準溶液。
5.4、鎘含量分析
使用 Perkin Elmer 的 AAnalyst 200 火焰原子吸收光譜儀,以 228.80 nm 波長的 鎘元素燈管作為光源,檢測標準溶液的吸收度並建立檢量線,將樣品的吸收度代 入檢量線以求取鎘濃度。每個樣品檢測三次取平均值。
第肆章、 結果
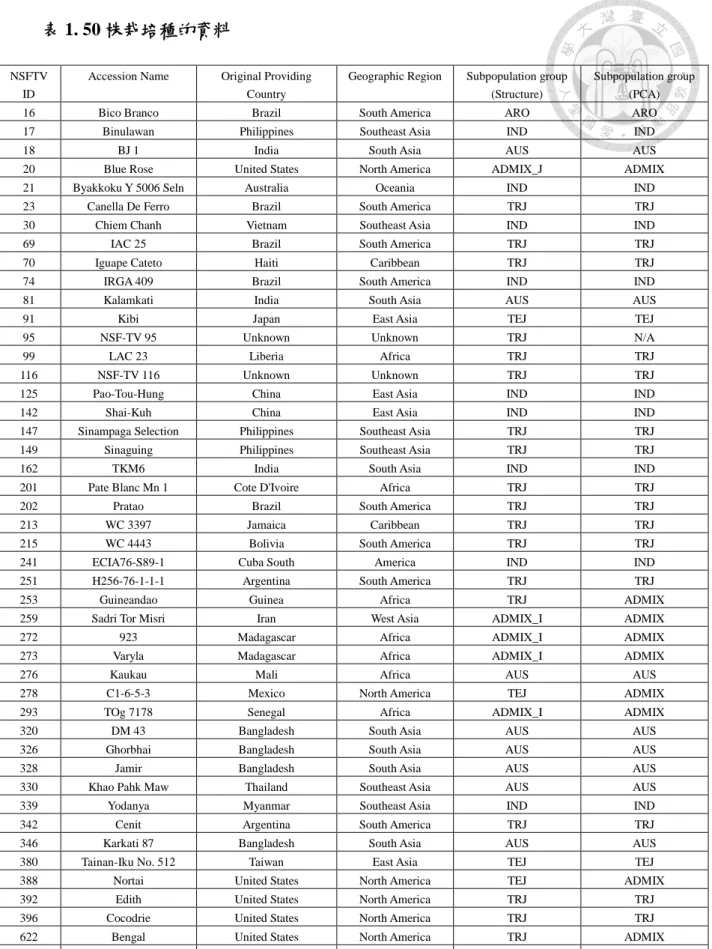
一、 50 株栽培種的相關資訊
利用 SNP 資料庫篩選出 50 株栽培種,參照 NSFTV 的編號整理於 (表 1)。
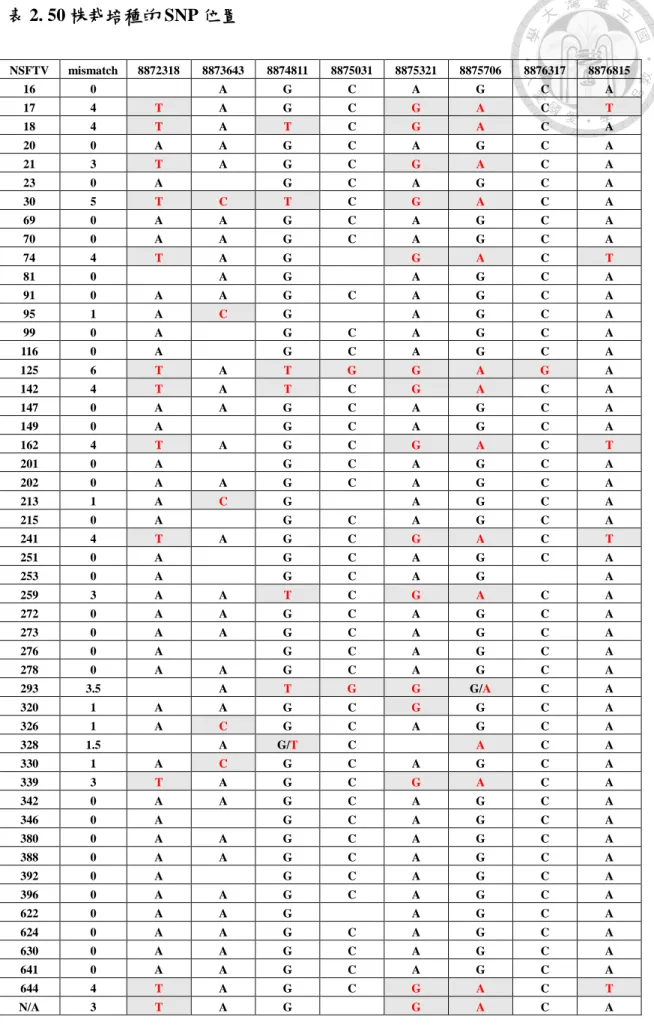
二、 OsNramp5 的基因結構與胺基酸突變位點
在水稻註解計畫資料庫(The Rice Annotation Project database, RAP-DB) 中檢索 OsNramp5 的 資 料 , 將 獲 得 的 資 訊 整 理 如 下 所 述 : OsNramp5 基 因 編 號 為 Os07g0257200,基因結構如 (圖 1) ,一共具有 13 個外顯子及 12 個內隱子,全基 因長度為 7263 bp,編碼序列長度為 1617 bp。在 International Rice Informatics Consortium (IRIC) 的 SNP 資料庫中搜索 OsNramp5 有 SNP 的水稻品系,於 HDRA 品種集 (Variety set) 中 SNP 位點皆位於內隱子上 (表 2) ,由於內隱子的基因序列 不會轉錄成 mRNA,即使變化了也不會改變 Nramp5 的蛋白結構。而在另外一個 3k 品種集中有部分 SNP 位點位於外顯子上,但由於密碼子簡併性,多種密碼子 可能轉譯出同一個胺基酸,即使有鹼基變化也不一定會造成胺基酸改變。在外顯 子 SNP 點位中,僅有位於第十三個外顯子的 SNP 會造成胺基酸改變 (圖 2) ,即 紅 色 標 示 的 丙 胺 酸 變 成 蘇 胺 酸 。 在 資 料 庫 中 符 合 此 胺 基 酸 序 列 的 栽 培 種 有 NSF-20、NSF-21、Nona Bokra 共三種。
三、 50 株栽培種的基因定序及胺基酸變化
將 50 株水稻 DNA 定序片段轉換為胺基酸序列並排序 (圖 3) 。可見 NSF-20、
NSF-21 及 Nona Bokra 有前述預期的胺基酸變化,另外有七株植株也有測得未預 期的胺基酸變化,分別為 NSF-16 及 74 的第 523 個胺基酸由穀胺酸 (Glutamate, E) 變為賴胺酸 (Lysine, K) ,NSF-30 及 388 的終止密碼子各變為亮胺酸 (Leucine, L) 及酪胺酸 (Tyrosine, Y),NSF-201 的第 537 個胺基酸由脯胺酸 (Proline, P) 變為精 胺酸 (Arginine, R) ,NSF-392 的第 519 個胺基酸由天冬胺酸 (Aspartate, D) 變為酪 胺酸 (Tyrosine, Y) ,NSF-396 則有多處胺基酸變化。這十種栽培種可能因此具有 活性較低甚至是失去作用的 Nramp5。
四、 50 株栽培種的 OsNramp5 表現量
取 50 株栽培種水稻的 RNA ,反轉錄第一股 cDNA 後以 OsActin1 作為參照 基因,針對 OsNramp5 進行半定量 RT-PCR (圖 4) 。電泳圖的條帶明暗顯示基因表 現量多寡。在 50 個栽培種中並沒有顯著的表現量差異,可以排除 SNP 位點影響 啟動子調控區域等可能性,在本試驗使用的栽培種中,Nramp5 蛋白的功能有無才 是影響穀粒鎘含量的關鍵因素。
五、 鎘處理下水稻幼苗的植體鎘含量
以 5 μM CdCl2 處理水稻幼苗一周,並將植體分為地上部及根部分析鎘含量 (圖 5) ,NSF-251 及 Nona Bokra 無論在地上部、根部皆有較低的鎘含量 (小於 15 mg/kg dw 及 400mg/kg dw) 。其他栽培種諸如 NSF-125、NSF-278、NSF-388 以及 NSF-74、NSF-241、NSF-644 各具有較低的地上部鎘含量及根部鎘含量。具有作為 低鎘水稻育種親本株的潛力。
六、 CRISPR 轉植株的轉基因檢測
CRISPR 系統的載體構築如 (圖 6) 使用 pRGEB32 載體,由 Rice snoRNA U3 promoter 表現設計好的 sgRNA , UBI promoter 表現 Cas9 核酸酶。基因編輯位 置為 Exon IX 上的 PS1 及 PS2,構築成兩種目標序列不同的載體,藉由農桿菌轉 殖入 TN11 及 TNG67 水稻植株。CRISPR 轉殖株可藉由對應的引子 (針對 HPT 基 因序列設計) 進行 PCR 確認是否成功導入轉基因。試驗 PCR 中以正常株 DNA 作 為陰性對照組、質體為陽性對照組、無菌水為空白對照組。結果可見 (圖 7) 中轉 殖株皆帶有與質體相同的轉基因,正常株與無菌水則沒有,顯示所有轉殖株皆成 功導入轉基因。
七、 CRISPR 轉植株的基因編輯成果
於 (圖 6) 中顯示 CRISPR 編輯的位點,轉殖株基因定序結果整理如 (圖 8) , 左側是針對 ps1 序列的編輯結果,右側兩個是針對 ps2 序列的編輯結果。
第伍章、 討論
水稻為亞洲人口的主要糧食,因此保護消費者遠離鎘米危害十分重要,倘若能 育成低鎘累積量的水稻,便可以讓消費者放心食用。為達成此目的本研究以 OsNramp5 為目標基因,得到 osnramp5 失去功能而無法吸收鎘的突變株。本研究 從兩個方向進行:一為利用單核苷酸多態性資料庫篩選 OsNramp5 核苷酸具有變 異的水稻品系,以作為育成低鎘水稻的種源;二為應用基因編輯技術剔除台灣水 稻品種的 OsNramp5 基因,以育成無轉基因的本土低鎘水稻品種。
在前人的研究中,錳是植物進行光合作用的重要成分,同時也是原本 OsNramp5 基因負責運輸的對象,可以預期失去活性的 osnrmap5 突變株除了鎘吸收量減少,
錳吸收量也會降低。日本及中國的學者有針對 osnramp5 突變株檢測植株的錳含 量,無論是離子束照射或 CRISPR 誘發突變,突變株的錳含量皆顯著低於正常株 (Ishikawa et al., 2012; Tang et al., 2017) 。在後續田間試驗中發現產量有些微下降,
可能是缺錳導致的現象。經過多次試驗發現倘若供給足量的錳,水稻的產量只會 減少 5%,在學者評估安全性與產量的取捨後,認定為可以接受的損失(Kobayashi et al., 2018; YANG et al., 2019)。
除了植體錳、鎘含量顯著降低,與正常株不同的性狀不僅無益甚至可能有害,
因此確認突變株是否有異常性狀非常重要。前述日本學者以離子束誘發突變獲得 了 lcd-kmt1, 2, 3,其中 lcd-kmt1, 2 在植株及穀粒的表型、食米品質、稻稈重等農 藝性狀不受誘發突變影響。在 lcd-kmt3 則有提早抽穗、植株矮小等未預期性狀,
但學者推測原因為該突變株缺失大片段的基因 (大於 200k bp) ,除 OsNramp5 以 外其他基因也失去功能,方導致突變株與正常株出現差異。由於 lcd-kmt1, 2 沒有 出現負面的性狀,日本學者認定可以做為育種的親本株,並在對子代的產量、基 因固定度檢驗後選擇 lcd-kmt 2 品系命名為越光環 1 號 (Ishikawa et al., 2019) 。另 一方面,中國學者檢測 CRISPR 編輯試驗得到的秈稻品系,同樣認定突變株的農 藝性狀與正常株並無差異,失去 OsNramp5 功能不會對植株造成負面影響。由這些 前例可知取 osnramp5 突變株作為低鎘水稻育種的親本株沒有問題。
基改食品的疑慮之一為作物中的轉基因,民眾對於轉基因除了毒性、過敏性等 安全考量外,甚至在宗教因素也有疑慮。CRISPR 技術雖然目的為誘發基因突變 使其失去功能,不會在目標基因中留下轉基因,但仍需要將 CRISPR 系統的基因 植入作物中,反基改食品的民眾可能因此產生不必要的恐慌,因此 CRISPR 轉殖 株還需要將編輯成功的基因與轉基因分離。分離轉基因的方式可以參考傳統育 種,藉由轉殖株的自交分離保留失去功能的目標基因,並獲得不含轉基因的作物。
依照此步驟產生的基因編輯作物不同於基改作物,在安全性上沒有轉基因的疑 慮。目前各國對於基因編輯食品的態度各有不同,在 2018 年 3 月 18 日美國農業 部 (U.S. Department of Agriculture) 宣布不會對 CRISPR 等基因編輯作物採取管制 措施,而歐盟則是採取保守態度,認定基因編輯技術屬於基因改造生物法的管理 範疇。日本厚生勞動省於 2019 年 3 月 27 日,宣布未含有外源基因的基因編輯產 品不受基因改造相關法規之管制,只需準備相關資料供厚生勞動省審查。在我國
依「食品安全衛生管理法」第三條之定義,「基因改造」意指使用基因工程或分子
生物技術將遺傳物質轉移或轉殖入活細胞或生物體,產生基因重組現象,使表現 具外源基因特性或使自身特定基因無法表現之相關技術。但不包括傳統育種、同 科物種之細胞及原生質體融合、雜交、誘變、體外受精、體細胞變異及染色體倍 增等技術。但由於基因編輯為新興技術,是否適用這條法規尚未有定論。
第陸章、 參考文獻
Cancer, I. A. f. R. o. (1993). Beryllium, cadmium, mercury, and exposures in the glass.
Apresentado em: IARC Working Group on the Evaluation of Carcinogenic Risks to Humans: Beryllium, Lyon, 1993.
Chmielowska-Bąk, J., Gzyl, J., Rucińska-Sobkowiak, R., Arasimowicz-Jelonek, M., and Deckert, J. (2014). The new insights into cadmium sensing. Frontiers in plant science 5, 245.
Colangelo, E. P., and Guerinot, M. L. (2006). Put the metal to the petal: metal uptake and transport throughout plants. Current opinion in plant biology 9, 322-330.
CURIE, C., ALONSO, J. M., Marie, L., ECKER, J. R., and BRIAT, J.-F. (2000).
Involvement of NRAMP1 from Arabidopsis thaliana in iron transport.
Biochemical Journal 347, 749-755.
Foyer, C. H., and Noctor, G. (2005). Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses. The Plant Cell 17, 1866-1875.
Hernandez, L., Carpena‐Ruiz, R., and Garate, A. (1996). Alterations in the mineral nutrition of pea seedlings exposed to cadmium. Journal of Plant Nutrition 19, 1581-1598.
Iimura, K. (1978). Behavior and balance of contaminant heavy metals in paddy soils-studies on heavy metal pollution of soils (part 2). Bull Hokuriku Natl Agric Exp Stn 21, 95-145.
Inaba, T., Kobayashi, E., Suwazono, Y., Uetani, M., Oishi, M., Nakagawa, H., and Nogawa, K. (2005). Estimation of cumulative cadmium intake causing Itai-itai disease. Toxicol Lett 159, 192-201.
Iqbal, M., Mahmood, M. T., and Ahmed, F. (1991). Influence of cadmium toxicity on germination and growth of some common trees. Pakistan Journal of Scientific and Industrial Research (Pakistan).
Ishikawa, S., Abe, T., Kuramata, M., and Hayashi, S. (2019). Development of Low-Cadmium-Accumulating Rice. In "Cadmium Toxicity", pp. 139-150.
Springer.
Ishikawa, S., Ishimaru, Y., Igura, M., Kuramata, M., Abe, T., Senoura, T., Hase, Y., Arao, T., Nishizawa, N. K., and Nakanishi, H. (2012). Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice. Proceedings of the National Academy of Sciences 109, 19166-19171.
Ishikawa, S., Makino, T., Ito, M., Harada, K., Nakada, H., Nishida, I., Nishimura, M., Tokunaga, T., Shirao, K., and Yoshizawa, C. (2016). Low-cadmium rice (Oryza sativa L.) cultivar can simultaneously reduce arsenic and cadmium concentrations in rice grains. Soil science and plant nutrition 62, 327-339.
Ishimaru, Y., Takahashi, R., Bashir, K., Shimo, H., Senoura, T., Sugimoto, K., Ono, K., Yano, M., Ishikawa, S., and Arao, T. (2012). Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport. Scientific reports 2, 286.
Ito, H., and Iimura, K. (1976). The absorption and translocation of cadmium in rice plants and its influence on their growth, in comparison with zinc: Studies on heavy metal pollution of soils (Part 1). Bull Hokuriku Natl Agric Exp Stn 19, 71-139.
Klaassen, C. D. (1981). Pharmacokinetics in metal toxicity. Fundamental and Applied
Klaassen, C. D., Liu, J., and Choudhuri, S. (1999). Metallothionein: an intracellular protein to protect against cadmium toxicity. Annual review of pharmacology and toxicology 39, 267-294.
Kobayashi, A., Hori, K., Yamamoto, T., and Yano, M. (2018). Koshihikari: a premium short-grain rice cultivar–its expansion and breeding in Japan. Rice 11, 15.
Lanquar, V., Lelièvre, F., Bolte, S., Hamès, C., Alcon, C., Neumann, D., Vansuyt, G., Curie, C., Schröder, A., and Krämer, U. (2005). Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. The EMBO journal 24, 4041-4051.
Møller, I. M. (2001). Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annual review of plant biology 52, 561-591.
Mammadov, J., Aggarwal, R., Buyyarapu, R., and Kumpatla, S. (2012). SNP markers and their impact on plant breeding. International journal of plant genomics 2012.
Nakanishi, H., Ogawa, I., Ishimaru, Y., Mori, S., and Nishizawa, N. K. (2006). Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+
transporters OsIRT1 and OsIRT2 in rice. Soil Science & Plant Nutrition 52, 464-469.
Nevo, Y., and Nelson, N. (2006). The NRAMP family of metal-ion transporters.
Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1763, 609-620.
Pandey, N., and Sharma, C. P. (2002). Effect of heavy metals Co2+, Ni2+ and Cd2+ on growth and metabolism of cabbage. Plant Science 163, 753-758.
Pedas, P., Ytting, C. K., Fuglsang, A. T., Jahn, T. P., Schjoerring, J. K., and Husted, S.
(2008). Manganese efficiency in barley: identification and characterization of the metal ion transporter HvIRT1. Plant physiology 148, 455-466.
Perfus‐Barbeoch, L., Leonhardt, N., Vavasseur, A., and Forestier, C. (2002). Heavy metal toxicity: cadmium permeates through calcium channels and disturbs the plant water status. The Plant Journal 32, 539-548.
Poschenrieder, C., Gunse, B., and Barcelo, J. (1989). Influence of cadmium on water relations, stomatal resistance, and abscisic acid content in expanding bean leaves.
Plant Physiology 90, 1365-1371.
Sasaki, A., Yamaji, N., Yokosho, K., and Ma, J. F. (2012). Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. The Plant Cell 24, 2155-2167.
Takao, W., Shimbob, S., Moon, C., S., Zhang, Z., W., and Ikeda, M. (1996). Cadmium contents in rice samples from various areas in the world. The Science of the Total Environment 184, 191-196.
Tang, L., Mao, B., Li, Y., Lv, Q., Zhang, L., Chen, C., He, H., Wang, W., Zeng, X., and Shao, Y. (2017). Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield.
Scientific reports 7, 14438.
Thomine, S., Lelièvre, F., Debarbieux, E., Schroeder, J. I., and Barbier‐Brygoo, H.
(2003). AtNRAMP3, a multispecific vacuolar metal transporter involved in plant responses to iron deficiency. The Plant Journal 34, 685-695.
Thomine, S., Wang, R., Ward, J. M., Crawford, N. M., and Schroeder, J. I. (2000).
Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis with homology to Nramp genes. Proceedings of the National Academy of Sciences 97, 4991-4996.
Vidal, S. M., Malo, D., Vogan, K., Skamene, E., and Gros, P. (1993). Natural resistance to infection with intracellular parasites: isolation of a candidate for Bcg. Cell 73, 469-485.
Waalkes, M., and Berthan, G. (1995). Handbook on metal-ligand interactions of biological fluids. Marcel Dekker, New York.
Waalkes, M., Misra, R., and Chang, L. (1996). Toxicology of metals. Boca Raton, FL:
CRC.
Xia, J., Yamaji, N., Kasai, T., and Ma, J. F. (2010). Plasma membrane-localized transporter for aluminum in rice. Proceedings of the National Academy of Sciences 107, 18381-18385.
YANG, C.-h., ZHANG, Y., and HUANG, C.-f. (2019). Reduction in cadmium accumulation in japonica rice grains by CRISPR/Cas9-mediated editing of OsNRAMP5. Journal of integrative agriculture 18, 688-697.
第柒章、 附錄
5’UTR 3’UTR
非轉錄區 外顯子 內隱子
8872040
OsNramp5
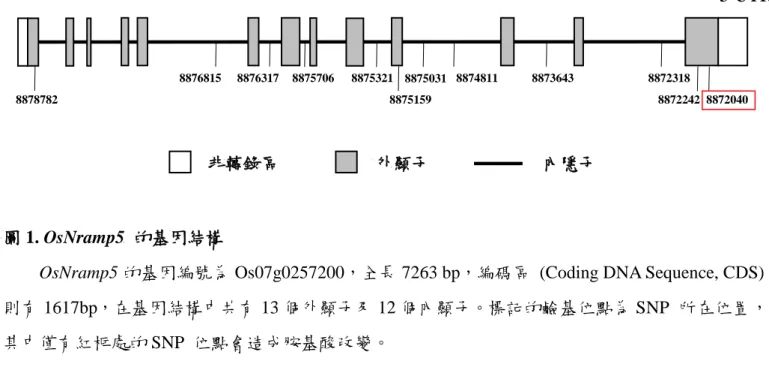
圖 1. OsNramp5 的基因結構
OsNramp5 的基因編號為 Os07g0257200,全長 7263 bp,編碼區 (Coding DNA Sequence, CDS) 則有 1617bp,在基因結構中共有 13 個外顯子及 12 個內顯子。標記的鹼基位點為 SNP 所在位置,
其中僅有紅框處的 SNP 位點會造成胺基酸改變。
8872318 8873643
8874811 8875031
8875321 8875706
8876317 8876815
8872242 8875159
8878782
Amino acid sequence of OsNramp5
>MEIERESSER GSISWRASAA HDQDAKKLDA DDQLLMKEPA WKRFLAHVGP GFMVSLAYLD PGNLETDLQA GANHRYELLW VILIGLIFAL IIQSLAANLG VVTGRHLAEI CKSEYPKFVK IFLWLLAELA VIAADIPEVI GTAFAFNILF HIPVWVGVLI TGTSTLLLLG LQKYGVRKLE FLISMLVFVM AACFFGELSI VKPPAKEVMK GLFIPRLNGD GATADAIALL GALVMPHNLF LHSALVLSRK TPASVRGIKD GCRFFLYESG FALFVALLIN IAVVSVSGTA CSSANLSQED ADKCANLSLD TSSFLLKNVL GKSSAIVYGV ALLASGQSST ITGTYAGQYI MQGFLDIRMR KWLRNLMTRT IAIAPSLIVS IIGGSRGAGR LIIIASMILS FELPFALIPL LKFSSSKSKM GPHKNSIYII VFSWFLGLLI IGINMYFLST SFVGWLIHND LPKYANVLVG AAVFPFMLVY IVAVVYLTIR KDSVVTFVAD SSLAA VVDAE KADAGDLAVD DDEPLPYRDD LADIPLPR*
GCC (A) → ACC (T)
圖 2. SNP 資料庫中胺基酸的改變位置
SNP 資料庫中顯示 NSF-20、NSF-22、Nona Bokra 等植株在 OsNramp5 蛋白的第 505 個胺基 酸由丙胺酸 (Alanine, A) 變為蘇胺酸 (Threonine, T),可能會因此改變 OsNramp5 的性能。
圖3. 50 株栽培種的胺基酸序列比對
圖中第一排為參照水稻日本晴,與日本晴相同的胺基酸以.省略表示,僅在相異處標示出胺基酸的改變。可見NSF-20、NSF
及Nona Bokra 有前述預期的胺基酸改變,另外有七株植株也有測得未預期的胺基酸變化,分別為NSF-16及74的第523個胺基酸由
穀胺酸(Glutamate, E) 變為賴胺酸(Lysine, K) ,NSF-30 及388的終止密碼子各變為亮胺酸(Leucine, L) 及酪胺酸(Tyrosine, Y)
NSF-201的第537個胺基酸由脯胺酸(Proline, P) 變為精胺酸(Arginine, R) ,NSF-392的第519個胺基酸由天冬胺酸(Aspartate, D)
為酪胺酸(Tyrosine, Y) ,NSF-396則有多處胺基酸變化。這十種栽培種可能因此具有活性較低甚至是失去作用的Nramp5。
圖4. 50 株栽培種的OsNramp5 表現量比較
半定量RT-PCR 電泳圖的條帶明暗顯示基因表現量多寡,以OsActin1 基因為比較基準,可見50個栽培種中並沒有顯著的表現
量差異,可以排除SNP 位點影響啟動子調控區域等可能性,在本試驗使用的栽培種中,Nramp5 蛋白的功能差異才是影響穀粒鎘含
量的關鍵因素。
Cd content (mg/kg)(dw)
Cultivars have different aa seq
sho o t
root
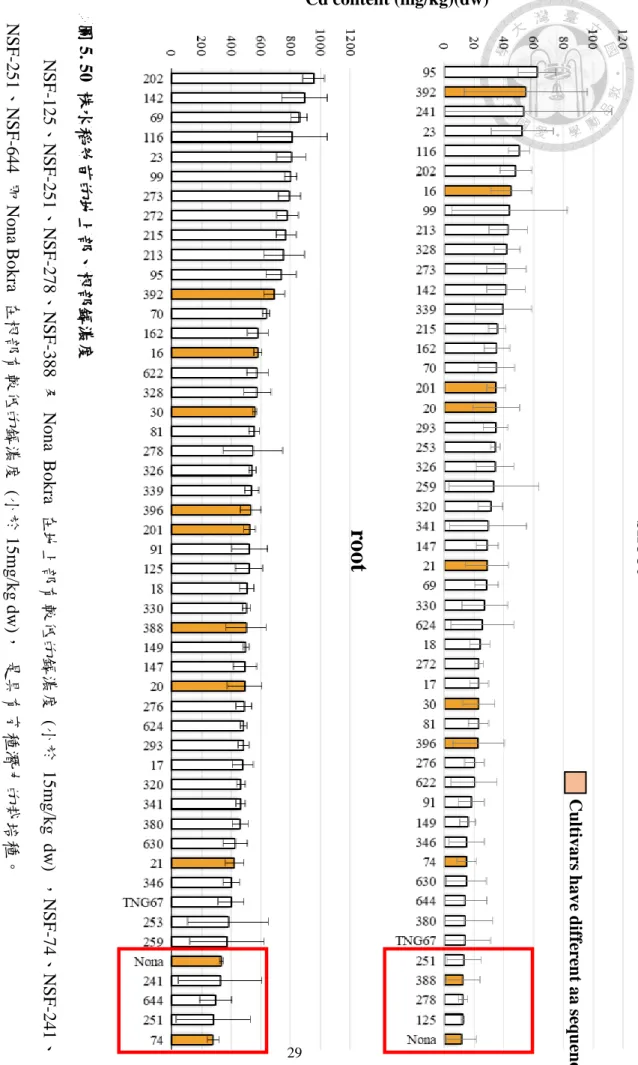
圖5. 50 株水稻幼苗的地上部、根部鎘濃度
NSF-125、NSF-251、NSF-278、NSF-388及NonaBokra 在地上部有較低的鎘濃度(小於15mg/kg dw) ,NSF-74、NSF-241
NSF-251、NSF-644 即Nona Bokra 在根部有較低的鎘濃度(小於15mg/kg dw), 是具有育種潛力的栽培種。
OsNramp5
圖 6. CRISPR/Cas9 系統載體構築及目標序列
使用 pRGEB32 載體,由 Rice snoRNA U3 promoter 表現設計好的 sgRNA , UBI promoter 表 現 Cas9 核酸酶。基因編輯位置為 Exon IX 上的 PS1 及 PS2,構築成兩種目標序列不同的載體,
並藉由農桿菌轉殖入 TN11 及 TNG67 水稻植株。
pRGEB32 15.9 kb
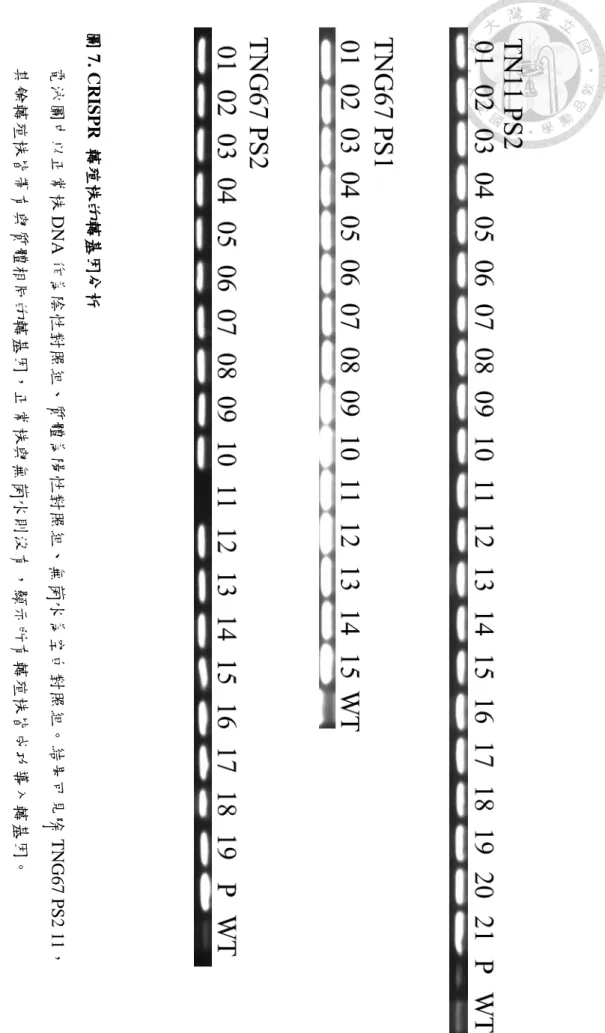
圖7. CRISPR 轉殖株的轉基因分析
電泳圖中以正常株DNA作為陰性對照組、質體為陽性對照組、無菌水為空白對照組。結果可見除TNG67 PS2 11,
其餘轉殖株皆帶有與質體相同的轉基因,正常株與無菌水則沒有,顯示所有轉殖株皆成功導入轉基因。
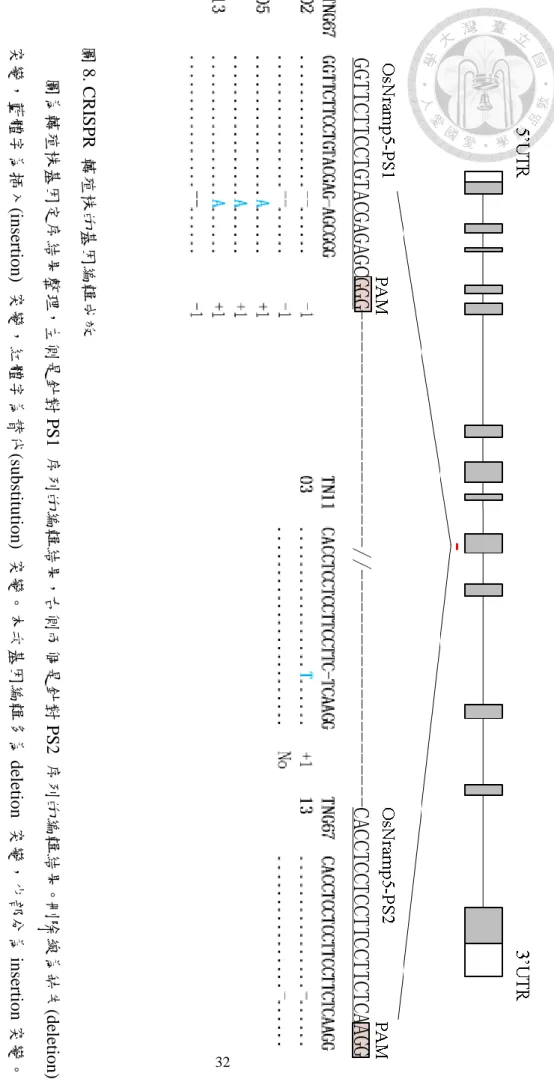
圖8. CRISPR 轉殖株的基因編輯成效
圖為轉殖株基因定序結果整理,左側是針對PS1 序列的編輯結果,右側兩個是針對PS2 序列的編輯結果。刪除線為缺失(deletion)
突變,藍體字為插入(insertion) 突變,紅體字為替代(substitution) 突變。本次基因編輯多為deletion 突變,少部分為insertion突變。
表 1. 50 株栽培種的資料
NSFTV ID
Accession Name Original Providing Country
Geographic Region Subpopulation group (Structure)
Subpopulation group (PCA)
16 Bico Branco Brazil South America ARO ARO
17 Binulawan Philippines Southeast Asia IND IND
18 BJ 1 India South Asia AUS AUS
20 Blue Rose United States North America ADMIX_J ADMIX
21 Byakkoku Y 5006 Seln Australia Oceania IND IND
23 Canella De Ferro Brazil South America TRJ TRJ
30 Chiem Chanh Vietnam Southeast Asia IND IND
69 IAC 25 Brazil South America TRJ TRJ
70 Iguape Cateto Haiti Caribbean TRJ TRJ
74 IRGA 409 Brazil South America IND IND
81 Kalamkati India South Asia AUS AUS
91 Kibi Japan East Asia TEJ TEJ
95 NSF-TV 95 Unknown Unknown TRJ N/A
99 LAC 23 Liberia Africa TRJ TRJ
116 NSF-TV 116 Unknown Unknown TRJ TRJ
125 Pao-Tou-Hung China East Asia IND IND
142 Shai-Kuh China East Asia IND IND
147 Sinampaga Selection Philippines Southeast Asia TRJ TRJ
149 Sinaguing Philippines Southeast Asia TRJ TRJ
162 TKM6 India South Asia IND IND
201 Pate Blanc Mn 1 Cote D'Ivoire Africa TRJ TRJ
202 Pratao Brazil South America TRJ TRJ
213 WC 3397 Jamaica Caribbean TRJ TRJ
215 WC 4443 Bolivia South America TRJ TRJ
241 ECIA76-S89-1 Cuba South America IND IND
251 H256-76-1-1-1 Argentina South America TRJ TRJ
253 Guineandao Guinea Africa TRJ ADMIX
259 Sadri Tor Misri Iran West Asia ADMIX_I ADMIX
272 923 Madagascar Africa ADMIX_I ADMIX
273 Varyla Madagascar Africa ADMIX_I ADMIX
276 Kaukau Mali Africa AUS AUS
278 C1-6-5-3 Mexico North America TEJ ADMIX
293 TOg 7178 Senegal Africa ADMIX_I ADMIX
320 DM 43 Bangladesh South Asia AUS AUS
326 Ghorbhai Bangladesh South Asia AUS AUS
328 Jamir Bangladesh South Asia AUS AUS
330 Khao Pahk Maw Thailand Southeast Asia AUS AUS
339 Yodanya Myanmar Southeast Asia IND IND
342 Cenit Argentina South America TRJ TRJ
346 Karkati 87 Bangladesh South Asia AUS AUS
380 Tainan-Iku No. 512 Taiwan East Asia TEJ TEJ
388 Nortai United States North America TEJ ADMIX
392 Edith United States North America TRJ TRJ
396 Cocodrie United States North America TRJ TRJ
622 Bengal United States North America TRJ ADMIX
624 Kaybonnet United States North America TRJ TRJ
630 Saber United States North America TRJ TRJ
641 Tainung 67 Taiwan East Asia TEJ TEJ
644 IR64-21 Philippines Southeast Asia IND IND
Subpopulation identified by STRUCTURE analysis based on 36 SSRs (Ali et al.
2011). Aus is coded with AUS, indica with IND, aromatic (GroupV) with ARO, temperate japonica with TEJ, tropical japonica with TRJ. Admix_I contains a greater proportion of ancestry from ether indica or aus, and ADMIX_J is admixture of mostly temperate or tropical japonica.
Subpopulation identified by PCA analysis using 36,901 high quality SNPs (Zhao et al. 2011). Aus is coded with AUS, indica with IND, temperate japonica with TEJ, tropical japonica with TRJ, aromatic (GroupV) with ARO, and ADMIX is a mixture of more than one subpopulation.