行政院國家科學委員會專題研究計畫 成果報告
臺灣地區醫學中心與區域醫院大腸桿菌與克雷氏肺炎桿菌 產生廣效性頭孢子黴素水解酵素分析與分子流行病學研究
(2/2)
計畫類別: 個別型計畫
計畫編號: NSC93-2320-B-006-016-
執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立成功大學醫學系病理學科
計畫主持人: 顏經洲 計畫參與人員: 蔡淑慧
報告類型: 完整報告
處理方式: 本計畫可公開查詢
中 華 民 國 94 年 9 月 29 日
行政院國家科學委員會補助專題研究計畫 █ 成 果 報 告
□期中進度報告
臺灣地區醫學中心與區域醫院大腸桿菌與克雷氏肺炎桿菌產生廣效性 頭孢子黴素水解酵素分析與分子流行病學研究
計畫類別:█ 個別型計畫 □ 整合型計畫
計畫編號:NSC 93 - 2320 - B - 006 - 016 - 執行期間: 92 年 8 月 1 日至 94 年 7 月 31 日
計畫主持人: 顏 經 洲 共同主持人:
計畫參與人員: 蔡 淑 慧
成果報告類型(依經費核定清單規定繳交):□精簡報告 █完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列 管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:國立成功大學醫學院病理學科
中 華 民 國 94 年 9 月 27 日
中文摘要
關鍵詞:廣效性頭孢子黴素水解酵素;廣效性頭孢子黴素;大腸桿菌;克雷氏肺炎桿 菌
具有廣效性頭孢子黴素水解酵素之格蘭氏陰性菌的散佈已是全球性的問題。頭孢子黴素水解
酵素主要分成A、B、C、D 四類。要預防這些酵素擴散,首要工作,必須了解其流行情形,
然而,目前臺灣地區關於廣效性頭孢子黴素水解酵素之流行病學調查結果,多屬單一醫院的 資料,缺乏全國性的資料。因此,本研究計畫目的為:(1)建立一全國性的廣效性頭孢子黴素 水解酵素之分子流行病學資料;(2)了解臺灣地區各類水解酵素之盛行情形;(3)了解不同區域
抗藥機轉差異與抗藥菌株之關聯性。本研究分析收集來自全國 7 家醫學中心對廣效性頭孢子
黴素有抗藥性之291 株大腸桿菌與 282 株克雷氏肺炎桿菌。利用瓊膠稀釋法分析其藥物敏感
度;使用isoelectric focusing、菌落雜交法、聚合脢連鎖反應與核酸定序決定其各類頭孢子黴
素水解酵素基因型、運用接合實驗與質體分析了解抗藥基因散佈情形,最後以菌株分型進行
分子流行病學研究。在這些菌株中,60.5%大腸桿菌與 94.0%克雷氏肺炎桿菌產生 A 類廣效性
頭孢子黴素水解酵素,43.6%大腸桿菌與 14.5%克雷氏肺炎桿菌產生 C 類廣效性頭孢子黴素水
解酵素。在大腸桿菌,CTX-M (54.3%)、SHV 廣效性頭孢子黴素水解酵素(9.3%)與 CMY-2 AmpC 水解酵素(43.6%) 被偵測到。在克雷氏肺炎桿菌,CTX-M (55.0%), SHV 廣效性頭孢子黴素水 解酵素 (47.9%), CMY-2 AmpC 水解酵素 (3.5%)與 DHA-1 AmpC 水解酵素(11.0%)被偵測到。
核酸定序分析 blaCTX-M 基因發現 7 種亞型,blaCTX-M-3、blaCTX-M-9、blaCTX-M-14、blaCTX-M-15、 blaCTX-M-17、blaCTX-M-19, 與一新型的 blaCTX-M-9相關的基因。在這7 種 blaCTX-M亞型中,blaCTX-M-3
與 blaCTX-M-14 分別在克雷氏肺炎桿菌與大腸桿菌佔大多數,而 blaCTX-M-17、blaCTX-M-19與新型
的 blaCTX-M-9相關的基因為在臺灣首度被發現 blaCTX-M亞型。分子流行病學研究發現抗藥菌株
具有基因的多型性。接合實驗與質體分析顯示抗藥基因散佈主要是以平行性的轉移為主而非 clone 的散佈。本研究結果顯示臺灣具有廣效性頭孢子黴素抗藥性之大腸桿菌與克雷氏肺炎桿 菌其抗藥基因具有複雜性與多樣性,必須監控這些抗藥基因進一步的散佈。
英文摘要
Key Words: extended-spectrum β-lactamases;extended-spectrum β-lactams;Klebsiella pneumoniae;
Escherichia coli
Widespread of gram-negative bacilli producing extended-spectrum β-lactamases is a global problem. β-Lactamases involved in resistance to extended-spectrum β-lactams are grouped into four major classes, classes A, B, C, and D. The purposes of the present project were: (1) to provide an overview of various β-lactamases in Klebsiella pneumonia and Escherichia coli in Taiwan; (2) to investigate the possible mechanisms of the spread of the resistance genes; (3) to establish the relationships of the resistance strains among different healthcare settings. A total of 291 Escherichia coli and 282 Klebsiella pneumoniae isolates that showed decreased susceptibilities to extended-spectrum cephalosporins were collected between March and August 2003 from seven medical centers. Overall, ESBL production was detected in 60.5% of the E. coli isolates and 94.0% of the K. pneumoniae isolates, and 43.6% of the E. coli isolates and 14.5% of the K.
pneumoniae isolates exhibited plasmid-mediated AmpC enzymes. In E. coli, CTX-M (54.3%), SHV ESBLs (9.3%), and CMY-2-related AmpC enzymes (43.6%) were detected; in K. pneumoniae, CTX-M (55.0%), SHV ESBLs (47.9%), CMY-2-related enzymes (3.5%), and DHA-1-related enzymes (11.0%) were detected. Thirty-five (12.0%) of the 291 E. coli isolates and 46 (16.3%) of the 282 K. pneumoniae isolates harbored two or three β-lactamases involved in resistance to extended-spectrum cephalosporins. Nucleotide sequencing of blaCTX-M genes revealed the presence of 7 subtypes, which were blaCTX-M-3, blaCTX-M-9, blaCTX-M-14, blaCTX-M-15, blaCTX-M-17, blaCTX-M-19, and a novel blaCTX-M-9-related gene. Among the seven blaCTX-M subtypes, blaCTX-M-3
and blaCTX-M-14 were predominant in K. penumoniae and E. coli, respectively, and blaCTX-M-17, blaCTX-M-19, and the novel blaCTX-M gene were firstly identified in Taiwan. Randomly amplified polymorphic DNA analysis revealed genetic diversity among the isolates with blaCMY-2, blaCTX-M-3, or blaCTX-M-14 randomly selected from different hospitals, and conjugation experiments and plasmid analysis suggest the interhospital spread of similar resistance plasmids.
目錄
頁數 封面
中文摘要 ……..………...I
英文摘要 ……..………...II
目錄 ……..………...III
前言 ……..………... 1
研究目的 …..……….. 2
文獻探討 …..………..3-4
研究方法 …..………..5-7
結果 …..………..8-10
討論 …..………..11-12
參考文獻 …..………..13-15
圖 …..………..16-18
表 …..……….. 19-21
前言
由於抗生素的廣泛使用,細菌中產生抗藥性的比率越來越高,人類往往必須發展 更強更廣效性的抗生素來對抗這些細菌。但卻往往跟不上細菌產生抗藥性的速度。抗 藥菌株的發生與散佈,不僅危及生命,亦造成醫療支出的增加。針對這個問題,目前 最佳的對策還是預防它們的發生與散佈。
在所有的抗生素中,頭孢子黴素 (cephalosporins) 為使用最多的藥物之一。特別 是針對格蘭氏陰性菌。而格蘭氏陰性菌產生對頭孢子黴素的抗藥性,最常見的機轉便 是細菌自己製造會破壞頭孢子黴素結構的水解酵素(1)。這些水解酵素,依照其分子結 構,主要可分為A、B、C、D 四大類 (2)。
醫院中的格蘭氏陰性菌,由於常常具有多重抗藥性,當其造成感染時,往往必須 使用廣效性抗生素,尤其是第三代頭孢子黴素,也因此醫院內對第三代頭孢子黴素有 抗藥性的菌株也逐漸增加,而其抗藥機轉主要是由於細菌產生廣效性頭孢子黴素水解 酵素。廣效性頭孢子黴素水解酵素多屬A 類頭孢子黴素水解酵素,主要為 SHV 與 TEM 兩大型(3)。在過去幾年內,這兩型酵素,不管是流行病學或臨床研究方面,被研究得 最為清楚。由於具有這型酵素菌株往往具有多重抗藥性,特別是 fluoroquinolone 類抗 生素(4),如 ciprofloxacin,當被其感染時,carbapenem 類抗生素,如 imipenem 或 meropenem,往往成為最後一線用藥(5)。不幸的是,目前亦有可以水解這最後一線用 藥的酵素出現,主要是IMP 與 VIM 兩型酵素 (6-9)。這類酵素,主要屬於 B 類頭孢子 黴素水解酵素。另外,C 類頭孢子黴素水解酵素也可以使細菌對第三代頭孢子黴素有 抗藥性,主要以CMY-1 與 CMY-2 兩型酵素為代表(10,11)。C 類與 A 類頭孢子黴素水 解酵素主要不同處在於 A 類可被β-lactamase 抑制劑抑制,且較不易水解 cephamycin 類抗生素,C 類則皆不被這兩類藥物抑制。
目前 A 類廣效性頭孢子黴素水解酵素在大腸桿菌與克雷氏菌的偵測,已有標準方 法可讓一般臨床實驗室使用 (12)。B 類與 C 類頭孢子黴素水解酵素則否,一般臨床實 驗室很難確定它們的存在與否。由於它們不易被偵測出來,目前全世界關於它們的流 行病學與臨床研究僅限少數的幾個實驗室,此類流行病學與臨床研究資料也很有限;
也正因為目前大家還不甚了解,它們容易散佈開來,造成病患與醫療照護系統的問題。
研究目的
目前臺灣地區關於廣效性頭孢子黴素水解酵素之流行病學調查結果,多屬單一醫院的資料,
缺乏全國性的資料,而這些資料正是訂定預防廣效性頭孢子黴素水解酵素散佈之策略或政策 的重要根據之一。因此,本研究計畫之目的為:(1)建立一全國性的廣效性頭孢子黴素水解酵 素之分子流行病學資料;(2)了解臺灣地區各類頭孢子黴素水解酵素之盛行情形;(3)了解不同 區域抗藥機轉差異與抗藥菌株之關聯性。本計畫之研究結果將有助於了解臺灣地區廣效性頭 孢子黴素水解酵素之流行情形,便於及早採取適當對策,避免更多患者的感染及酵素之擴散。
本研究計畫將以克雷氏肺炎桿菌與大腸桿菌為研究標的。
文獻探討
過去幾年,臺灣關於廣效性頭孢子黴素水解酵素的研究,主要屬於A 類頭孢子黴素水解酵素 (13-15)。盛行率方面,以克雷氏肺炎桿菌而言,一般是在 5 到 10%之間,最高者為中部某醫 院,達30%左右(13),而成大在 1998 年的數據約在 8%左右 (14)。種類以 SHV 型為主,大多 為SHV-5 與 SHV-12 兩種亞型。而大腸桿菌方面,成大約在 2-3%之間 (15)。比較特別的是其 種類我們發現是以CTX-M-3 這一類為主。CTX-M-3 一開始是被報告出現在波蘭(16),我們是 首度發現其出現在臺灣。A 類廣效性頭孢子黴素水解酵素目前在一般臨床微生物實驗室皆可 用標準方法去篩選確認,因此,它們的散佈比較容易被追蹤與預防。
C 類頭孢子黴素水解酵素由於沒有標準方法去偵測,因此到目前沒有,也很難有整體的 盛行率數據。我們成大在1998 年收集的克雷氏肺炎桿菌菌株中,首度發現三株具有一新型的 C 類頭孢子黴素水解酵素。此酵素其分子結構與在韓國被發現的 CMY-1 相近(14)。在大腸桿 菌方面,我們在1999 年一至九月收集的菌株中,首度發現五株具有 CMY-2 型 C 類頭孢子黴 素水解酵素出現在臺灣(15),而此酵素首次是發現在希臘。這一型酵素最近在成大分離出來 的大腸桿菌中,似有明顯增加的趨勢,由於CMY-2 不易以一般臨床微生物實驗室偵測,加上 它可在不同菌種、不同環境中廣為分佈,使得防止它進一步漫延,變得難上加難。另外我們 成大在1999 與 2000 年收集的克雷氏肺炎桿菌菌株中,首度發現產生具誘導性表現的 DHA-1 酵素(17),此類菌株主要特性與產生內源性 C 類頭孢子黴素水解酵素的格蘭氏陰性桿菌,如 Enterobacter 與 Citrobacter 類似,在β-lactamase 抑制劑或 cephamycin 類抗生素存在下,容易 被誘發出來,使菌株對廣效性頭孢子黴素有明顯抗藥性。
九 0 年代初期,在日本首度發現可在不同細菌間傳播的水解 carbapenem (院內感染常用的最 後一線抗生素) 的 B 類頭孢子黴素水解酵素,主要為 IMP-1 型(6)。此酵素的出現。在日本引 起相當重視,而後進行一連串的研究,發現此酵素在日本主要存在於Serratia 與綠膿桿菌。其 基因可在染色體上,亦可在質體上(18-21)。最近兩年,相繼在法國與義大利發現此酵素之亞 型,主要為IMP-2、VIM-1 與 VIM-2 (7-9)。在臺灣,我們亦發現 B 類頭孢子黴素水解酵素的 存在(22, 23)。在假性單胞菌、包括綠膿桿菌,主要是 VIM-2 與一我們所新發現的 VIM-3 型 酵素,另外也有少部分的IMP-1 酵素。更重要的發現是,我們在一 1998 年克雷氏肺炎桿菌菌 株中發現一 IMP-2 亞型,我們命名為 IMP-8。由於克雷氏肺炎桿菌具有儲存各種抗藥基因的 特性,使得此類酵素容易被擴散開。在1999 與 2000 年的研究中,我們發現有產生 IMP-8 的 克雷氏肺炎桿菌與 Enterobacter cloacae 在成大醫院爆發院內感染的情形 (24, 25),同時也首 度發現產生 VIM-2 的 Citrobacter freundii 菌株 (25)。B 類頭孢子黴素水解酵素雖可水解 carbapenem,然在藥物敏感試驗中卻常呈現對 carbapenem 敏感的現象,因此不易要用一般藥 物敏感試驗去篩選出來。此類酵素一旦擴散開來,將對住院病患造成極大威脅,醫師可能面 臨無藥可用的窘境。
最近幾年,廣效性頭孢子黴素水解酵素流行趨勢似乎有逐漸改變的情況出現。根據最近 我們剛完成的91 年度疾病管制局之科技計畫結果顯示,從 1999 年至 2002 年收集至成大醫院
的菌株中,在大腸桿菌與克雷氏肺炎桿菌,產生A 群廣效性頭孢子黴素水解酵素比例,分別 約在1.5%至 3.5%與 4.8%與 13%之間,盛行率略有波動但增加情形並不明顯。C 類酵素則有 快速增加趨勢,在大腸桿菌與克雷氏肺炎桿菌之盛行率,分別從 0.4 及 1.5%增加到 5.5 與 8.8%;其中變化最大者為 CMY-2 型,分別從 0.4 及 0%增加到 5.5 與 6.7%;其次為 DHA-1 型,
在克雷氏肺炎桿菌,從 0.6%逐年增加到 2%;CMY-8 型則有減少的情形,且從 2000 年後便 不復出現。B 類酵素在 Enterobacter cloacae 與克雷氏肺炎桿菌發現的是 IMP-8,在假性單胞 菌是VIM-3 與 IMP-1。這些酵素盛行率皆出現減少的情形。分子流行病學分析方面,CMY-2 菌株之增加,主要是由於抗藥基因在不同菌株間散佈。在引起血行性感染之產生CMY-2 大腸 桿菌菌株中,37%可能為社區獲得,其中又有一半可能來自安養中心,表示 CMY-2 菌株可能 在社區環境廣為散佈,特別是安養中心。產生IMP-8 之克雷氏肺炎桿菌之出現,主要是由於 發生院內感染爆發所致。產生DHA-1 之克雷氏肺炎桿菌與 IMP-8 之 E. cloacae 之流行,與院 內感染爆發及抗藥基因在不同菌株間散佈有關,少部份菌株可能其他醫院或安養中心。這些 結果可歸納成幾點結論:(1) 台南地區廣效性頭孢子黴素水解酵素之種類具複雜與多樣性;(2) CMY-2 C 類酵素有快速增加趨勢,且已成為一重要且常見的廣效性頭孢子黴素;(3) 抗藥基 因已在不同菌種間相互散佈;(4) 一些抗藥基因已在醫院外環境散佈;(5) 具抗藥基因之菌株 具基因多樣性。
研究方法
第一年
產生A 類廣效性頭孢子黴素水解酵素菌株確認:
本年度主要研究A 類廣效性頭孢子黴素水解酵素。A 類廣效性頭孢子黴素水解酵素菌株
之確認乃根據National Committee for Clinical Laboratory Standards (NCCLS) 方法 (12),比較 ceftazidime 與 cefotaxime 加了 clavulanic acid 後抑制環變化大小決定。
藥物敏感性試驗:
使 用 瓊 膠 稀 釋 法 來 決 定 抗 藥 菌 株 之 藥 物 敏 感 性(12) 。 使 用 藥 物 包 括 cefotaxime 、 ceftazidime、ceftriaxone、cefoxitin、cefmetazole、imipenem、meropenem、ampicillinn、amoxicillin 與clavulanic acid、piperacillin、piperacillin 與 tazobactam、ciprofloxacine、gentamicin、amikacin 與tobramycin。
等電點聚焦法(Isoelectric focusing)分析
利用等電點聚焦法來決定細菌產生之頭孢子黴素水解酵素特性(26, 27)。水解酵素之粗萃 取,乃利用超音波震破細胞而得(26)。將細胞粗萃取液點於 pH3.0 至 9.5 之膠 (Amersham Pharmacia Biotech,Hong Kong)上進行電泳。電泳完畢後用 nitrocefin 呈色。
攜帶頭孢子黴素水解酵素基因菌株之偵測:
利用聚合脢連鎖反應法偵測攜帶頭孢子黴素水解酵素基因菌株。主要標的基因為SHV、
TEM、三群 CTX-M 型基因 (14, 15, 22, 23)。利用非放射性菌落雜交法,使用以上聚合脢連 鎖反應法產物來製造探針,來排除聚合脢連鎖反應法偽陰性的可能性。利用直接核酸定序法 來決定各個基因亞型。
接合實驗 (Conjugation experiments) 與質體分析(Plasmid analysis):
利用接合實驗來了解抗藥基因之存在位置(28)。將菌株與對 streptomycin 有抗藥性的 C600 大腸桿菌混合生長,分別用含10 µg/ml ceftazidime 或 cefotaxime 或 64µg/ml cefoxitin 與 64µg/ml streptomycin 的培養基培養篩選。接合實驗成功的 C600 大腸桿菌分別用聚合脢連鎖 反應法、非放射性南方墨漬雜交法(29)與等電點聚焦法來確認。從接合實驗成功的 C600 大腸 桿菌抽取質體後,用限制脢 EcoRI 切取,電泳後比對,決定質體大小與彼此間關係。
菌株分型 (30)
利用Arbitrarily primed PCR (AP-PCR)及脈衝電場瓊膠電泳法(pulsed-field gel
electrophoresis) 來決定菌株間之關係,以了解菌株散佈情形,探討抗藥基因之散佈是否為同 型菌株散佈所致或平行散佈 (horizontal transfer)有關。
第二年
本年度主要研究 B 與 C 類廣效性頭孢子黴素水解酵素。
藥物敏感性試驗:
使用瓊膠稀釋法來決定 NCCLS 廣效性頭孢子黴素水解酵素確定方法陰性者之抗藥菌株
藥物敏感性(12)。使用藥物包括 cefotaxime、ceftazidime、ceftriaxone、cefoxitin、cefmetazole、
imipenem、meropenem、ampicillinn、amoxicillin 與 clavulanic acid、piperacillin、piperacillin 與tazobactam、ciprofloxacine、gentamicin、amikacin 與 tobramycin。
頭孢子黴素水解酵素特性分析:
1. 等電點聚焦法(Isoelectric focusing)分析:
利用等電點聚焦法來決定細菌產生之頭孢子黴素水解酵素特性(26, 27)。水解酵素之粗萃 取,乃利用超音波震破細胞而得(26)。將細胞粗萃取液點於 pH3.0 至 9.5 之膠 (Amersham Pharmacia Biotech,Hong Kong)上進行電泳。電泳完畢後用 nitrocefin 呈色。
2. 頭孢子黴素水解酵素水解試驗:
頭孢子黴素水解酵素水解試驗主要針對產生 B 類頭孢子黴素水解酵素之菌株,因其往往
對carbapenem 藥物呈現敏感性,必須用水解試驗來確定其作用。方法乃是將細胞粗萃取液與
等量(150 µM 之)各種頭孢子黴素混合,利用紫外線分析儀測定對應於該頭孢子黴素 OD 值之
變化,來決定其水解速度(7)。
攜帶頭孢子黴素水解酵素基因菌株之偵測:
利用聚合脢連鎖反應法偵測攜帶頭孢子黴素水解酵素基因菌株。主要標的基因為SHV、
TEM、CTX-M-3、CMY-1、CMY-2、DHA、IMP 與 VIM 型基因(14, 15, 22, 23)。利用菌落雜 交法,使用以上聚合脢連鎖反應法產物來製造探針,來排除聚合脢連鎖反應法偽陰性的可能 性。利用直接核酸定序法來決定各個基因亞型。
接合實驗 (Conjugation experiments) 與質體分析(Plasmid analysis):
利用接合實驗來了解抗藥基因之存在位置(28)。將菌株與對 streptomycin 有抗藥性的 C600
大腸桿菌混合生長,分別用含10 µg/ml ceftazidime 或 cefotaxime 或 64µg/ml cefoxitin 與 64µg/ml streptomycin 的培養基培養篩選。接合實驗成功的 C600 大腸桿菌分別用聚合脢連鎖 反應法、非放射性南方墨漬雜交法(29)與等電點聚焦法來確認。從接合實驗成功的 C600 大腸 桿菌抽取質體後,用限制脢 EcoRI 切取,電泳後比對,決定質體大小與彼此間關係。
菌株分型 (30)
利用Arbitrarily primed PCR (AP-PCR),ribotyping,及脈衝電場瓊膠電泳法(pulsed-field gel electrophoresis) 來決定菌株間之關係,以了解菌株散佈情形,探討抗藥基因之散佈是否為同 型菌株散佈所致或平行散佈 (horizontal transfer)有關。
結果
Detection of ESBL producers. Overall, 176 (60.5%) of 291 E. coli isolates and 265 (94.0%) of 282 K. pneumoniae isolates were considered ESBL producers with two phenotypic tests for ESBL detection. ESBL production was inferred by the NCCLS confirmatory tests in 171 E. coli isolates and 260 K. pneumoniae isolates; among the remainder, 5 E. coli isolates and 5 K.
pneumoniae isolates gave positive results in the double-disk synergy tests.
β-Lactamase identification. IEF analysis revealed the presence of β-lactamases with pIs of 9.0 (n = 127), 8.6 (n = 2), 8.4 (n = 39), 8.2 (n = 41), 8.0 (n = 1), 7.9 (n = 116), and 5.4 (n = 238) alone or in combination in 288 E. coli isolates and no β-lactamase in 3 E. coli isolates. In 282 K.
pneumoniae isolates, β-lactamases with pIs of 9.0 (n = 10), 8.4 (n = 98), 8.2 (n = 126), 8.0 (n = 2), 7.9 (n = 55), 7.8 (n = 31), 7.6 (n = 282), 6.1 (n = 5), and 5.4 (n = 194) were detected alone or in combination. The β-lactamases with pIs of 8.6, 8.4, 8.2, 8.0, 7.9, and 6.1 were only detected in putative ESBL producers, and the β-lactamases with pIs of 9.0, 7.8, 7.6, and 5.4 were detected in both putative ESBL and non-ESBL producers.
By PCR, the detection of blaTEM, blaOXA-10-related, blaDHA-1-related, andblaCMY-2-related genes corresponded with the detection of β-lactamases with pIs of 5.4, 6.1, 7.8, and 9.0, respectively, by IEF. The blaCTX-M-1–related genes were detected in all isolates with the pI 8.6 or 8.4 β-lactamase, and the blaCTX-M-9–related genes were detected in all isolates with the pI 8.0 or 7.9 β-lactamase. A combination of IEF and the PCR-NheI method was used to identify multiple SHV-type
β-lactamases per isolate. All 41 E. coli isolates with a pI 8.2 β-lactamase showed two bands of 770, and 247 bp in the PCR-NheI analysis, suggesting the expression of SHV-5-like ESBLs.
Among the 282 K. pneumoniae isolates, all 126 isolates with a pI 8.2 β-lactamase showed three bands of 1,017, 770, and 247 bp in the PCR-NheI analysis, suggesting coproduction of an
SHV-5-like ESBL and an SHV-1-like non-ESBL by each of these isolates. Among the remaining 156 K. pneumoniae isolates, 9 isolates showed two bands of 770 and 247 bp, suggesting the expression of SHV-2-like ESBLs; 147 isolates yielded blaSHV products with no NheI site, suggesting that each of these isolates had an SHV-1-like non-ESBL. All 432 blaTEM-positive isolates of E. coli and K. pneumoniae expressed a pI 5.4 β-lactamase, suggesting that they had TEM-1-like narrow-spectrum β-lactamases. The blaCMY-1-related genes were not detected.
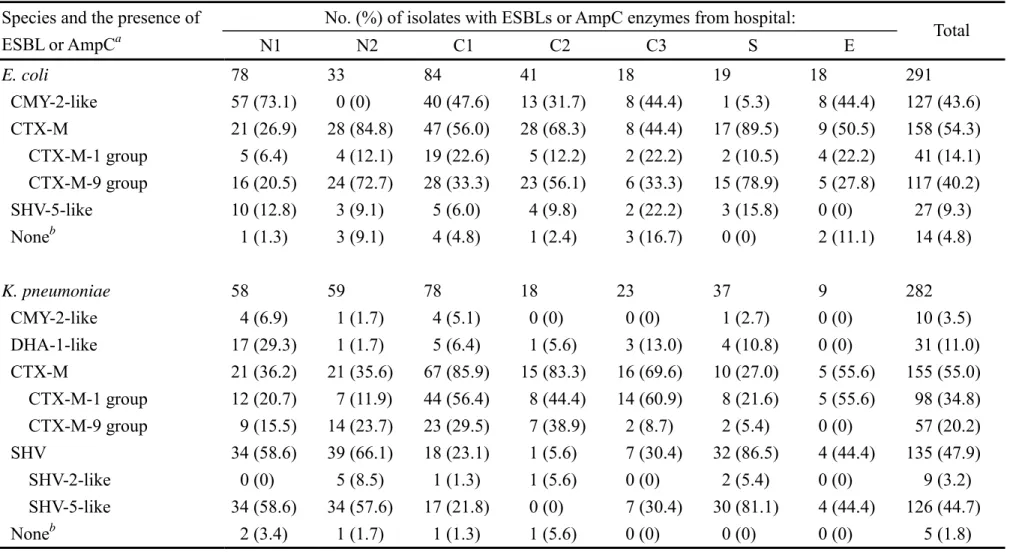
Overall, ESBL genes were detected in 176 (60.5%) of the 291 E. coli isolates and 265 (94.0%) of the 282 K. pneumoniae isolates, and AmpC genes were detected in 127 (43.6%) E. coli isolates and 41 (14.5%) K. pneumoniae isolates. The prevalence rates of the major types of ESBLs and AmpC enzymes among the extended-spectrum cephalosporin-resistant isolates were estimated based on the results of IEF and PCR, and the data are shown in Table 1. In E. coli, CTX-M ESBLs and
CMY-2-related AmpC enzymes were the most common types of β-lactamase involved in resistance to extended-spectrum cephalosporins and were found in 54.3% and 43.6%, respectively, of the 291 isolates. In K. pneumoniae, the most prevalent types were SHV and CTX-M ESBLs, which were
found in 55.0% and 47.9%, respectively, of the 282 isolates. ESBLs of CTX-M-1 group were more prevalent than those of CTX-M-9 group in K. pneumoniae, while ESBLs of CTX-M-9 group were more prevalent than those of CTX-M-1 group in E. coli.
Thirty-five (12.0%) of the 291 E. coli isolates and 46 (16.3%) of the 282 K. pneumoniae isolates harbored two or three β-lactamases involved in resistance to extended-spectrum cephalosporins.
Five ESBL-producing E. coli isolates that were not detected by the NCCLS ESBL confirmatory tests were found to produce a CMY-2-related AmpC enzyme and an SHV-5-related ESBL. Five ESBL-producing K. pneumoniae isolates that were not detected by the NCCLS confirmatory tests were found to produce a DHA-1-related AmpC enzyme and an SHV-5-related ESBL.
PCR products of blaCTX-M-1-, blaCTX-M-9-, blaDHA-1-, blaCMY-2-, and blaOXA-10-related genes obtained from selected isolates were sequenced. Nucleotide sequencing of the amplified blaCTX-M-9-related genes obtained from 134 isolates revealed the presence of blaCTX-M-14 (n = 88), blaCTX-M-9 (n = 4), and blaCTX-M-19 (n = 1) in E. coli and the presence of blaCTX-M-14 (n = 33), blaCTX-M-19 (n = 2), blaCTX-M-17 (n = 5), and an unpublished blaCTX-M-9–related gene (n = 1), temporarily named blaCTX-M-38 (GenBank accession no. AY822595) in K. pneumoniae. The unpublished blaCTX-M gene differs from blaCTX-M-14 by a single nucleotide, which leads to a
Ser220Arg substitution. Nucleotide sequencing of the amplified blaCTX-M-1-related genes obtained from 117 isolates revealed the presence of blaCTX-M-3 (n = 33) and blaCTX-M-15 (n = 2) in E. coli and the presence of blaCTX-M-3 in all K. pneumoniae isolates (n = 82) that were sequenced. All PCR products of β-lactamase genes related to blaDHA-1, blaCMY-2, and blaOXA-10 from selected isolates were identified to be blaDHA-1 (n = 23), blaCMY-2 (n = 16), and blaOXA-10 (n = 5), respectively, by nucleotide sequencing. blaOXA-10 encodes a narrow-spectrum Ambler’s class D β-lactamase (1, 7).
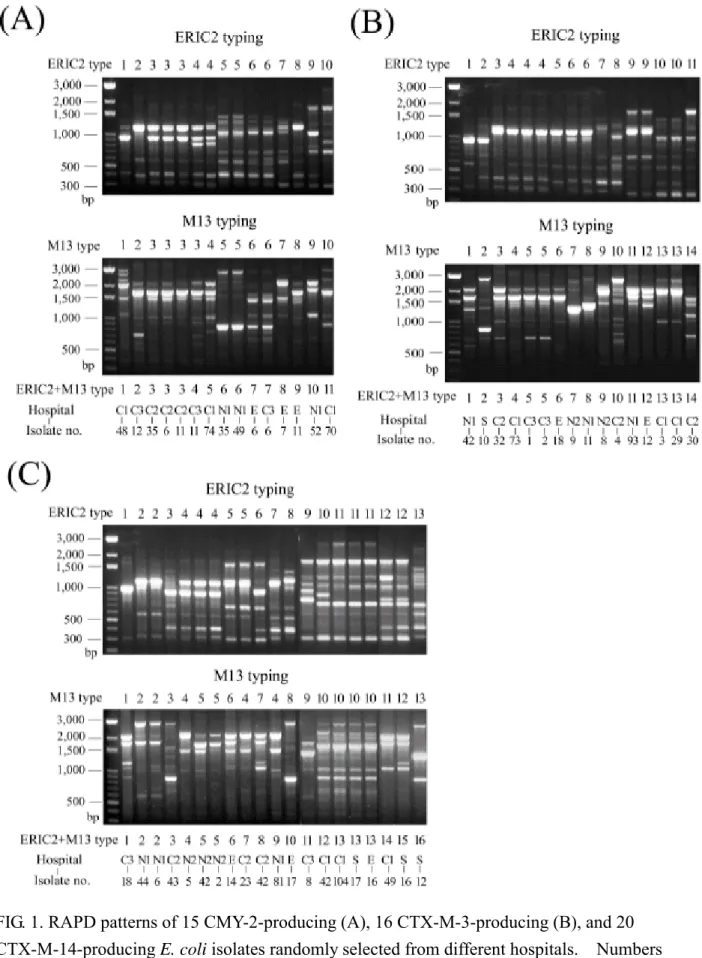
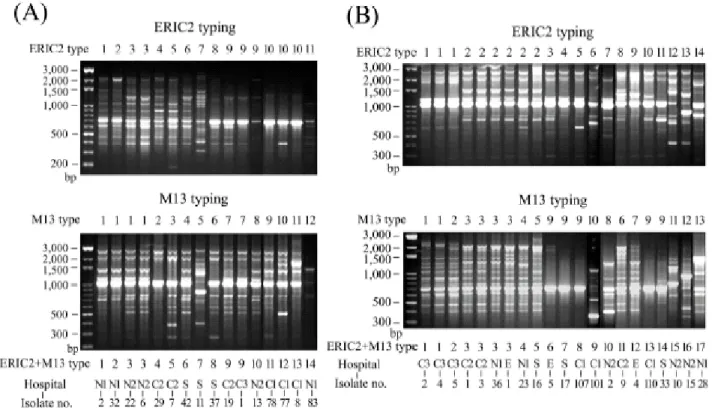
RAPD analysis. Isolates with blaCTX-M-14, blaCTX-M-3, or blaCMY-2 were randomly selected for RAPD typing. By combination of the results obtained with primers M13 and ERIC2, 16, 14, and 11 types were observed among 20 CTX-M-14-producing isolates, 16 CTX-M-3-producing isolates, and 15 CMY-2-producing isolates, respectively, in E. coli, and 14 and 17 types were observed among 16 CTX-M-14-producing isolates and 21 CTX-M-3-producing isolates, respectively, in K.
pneumoniae (Fig. 1 and 2).
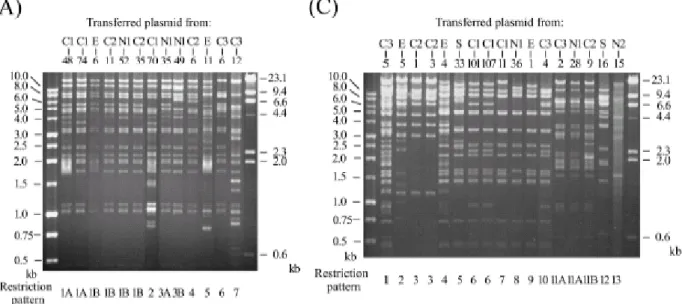
Conjugation experiments and plasmid analysis. Selected isolates that had been typed by RAPD were subject to conjugation experiments and plasmid analysis. Thirteen, 18, and 17
transconjugants were obtained from 15 CMY-2-producing E. coli isolates, 20 CTX-M-14-producing E. coli isolates, and CTX-M-3-producing K. pneumoniae isolates, respectively. The presence and expression of the resistance genes were confirmed by PCR and IEF, respectively. The results of fingerprinting analysis of the transferred plasmids are shown in Fig. 3. Although the transferred plasmids showed genetic diversity, the isolates from different hospitals were found to have blaCMY-2-positive, blaCTX-M-14-positive, or blaCTX-M-3-positive plasmids with identical or similar restriction patterns.
Susceptibility testing. Among the 291 E. coli isolates, 178 (61.2%), 127 (43.6%), 138 (47.4%),
87 (29.9%), 31 (10.7%), and 2 (0.7%) isolates were resistant to cefoxitin (MIC, 2 - >256 µg/ml;
MIC90, >256 µg/ml), cefotaxime (MIC, 0.06 - >128 µg/ml; MIC90, >128 µg/ml), ceftazidime (MIC, 0.13 - >128 µg/ml; MIC90, 128 µg/ml), aztreonam (MIC, 0.06 - >128 µg/ml; MIC90, 64 µg/ml), cefepime (MIC, 0.03 - >64 µg/ml; MIC90, 32 µg/ml), and imipenem (MIC, 0.06 - >32 µg/ml; MIC90, 0.25 µg/ml), respectively, based on the NCCLS criteria (24). Among the 282 K. pneumoniae isolates, 113 (40.1%), 134 (47.5%), 173 (61.3%), 205 (72.7%), 86 (30.5%), and 5 (1.8%) isolates were resistant to cefoxitin (MIC, 2->256 µg/ml; MIC90, 256 µg/ml), cefotaxime (MIC, 0.06->128 µg/ml; MIC90, >128 µg/ml), ceftazidime (MIC, 0.13->128 µg/ml; MIC90, >128 µg/ml), aztreonam (MIC, 0.06->128 µg/ml; MIC90, >128 µg/ml), cefepime (MIC, 0.03->64 µg/ml; MIC90, 64 µg/ml), and imipenem (MIC, 0.06->32 µg/ml; MIC90, 0.5 µg/ml), respectively. MICs of various β-lactam agents for representative isolates producing ESBLs and/or AmpC enzymes are shown in Table 2.
討論
This report describes the distribution of ESBLs and acquired AmpC enzymes among clinical isolates and E. coli and K. penumoniae that exhibited decreased susceptibilities to
extended-spectrum cephalosporins in Taiwan. Two surveys performed before 2001 at a university hospital in southern Taiwan showed the predominance of SHV and CTX-M ESBLs in K.
pneumoniae and E. coli, respectively (39, 40). In this study, the SHV-type enzymes remained the most prevalent ESBLs in K. pneumoniae at three hospitals in northern and southern Taiwan (Table 1); however, the CTX-M-type β-lactamases have become predominant at all three hospitals in central Taiwan. In E. coli, CTX-M-type enzymes remained the most prevalent ESBLs as described previously (38, 39).
The members of the CTX-M family have been increasing, among which CTX-M-14, CTX-M-3, and CTX-M-2 have been reported most commonly (4). In Taiwan, the predominance of CTX-M-3 among ESBL-producing E. coli isolates has been observed (39), and CTX-M-3 followed by
CTX-M-14 has been reported to be the most commonly identified CTX-M β-lactamase (39, 43).
Moreover, CTX-M-9 and CTX-M-15 (W. L. Yu, K. C. Cheng, L. T. Wu, M. A. Pfaller, P. L.
Winokur PL, and R. N. Jones, Letter, Antimicrob. Agents Chemother. 48:362-363, 2004) have been identified from E. coli and K. pneumoniae isolates, respectively, from two hospitals in southern Taiwan (38). In the present study, CTX-M-3 remained the most prevalent CTX-M-type enzyme in K. pneumoniae; however, CTX-M-14 has become the most commonly identified CTX-M enzyme in E. coli throughout Taiwan. Moreover, this report describes the countrywide spread of blaCTX-M-9
and the first appearance of CTX-M-15 in E. coli in Taiwan. CTX-M-15 has been noted for its increased activity against ceftazidime and has been described in many countries recently (4, 11, 15, 20, 21, 28).
Three CTX-M variants, CTX-M-17, CTX-M-19, and an unpublished enzyme temporarily named CTX-M-38, were identified for the first time in Taiwan in this study. blaCTX-M-17 differs from blaCTX-M-14 by two nucleotides, which lead to a Glu289Lys substitution (8). CTX-M-19 derives from CTX-M-15 by a Pro167Ser substitution and is also known to confer a higher level of
resistance to ceftazidime than cefotaxime (16, 29). Compatible with this finding, our isolates with CTX-M-19 were all found to have a high level of resistance to ceftazidime (≥ 64 µg/ml) (Table 2).
CTX-M-17 and CTX-M-19 are very rare and have been reported in clinical isolates of K.
pneumoniae from Vietnam and France, respectively (8, 9, 29). To our knowledge, this is the first report to document the presence of these two enzymes outside Vietnam and France. The sequence of blaCTX-M-38 was recently deposited in the GenBank database and the β-lactamase gene was obtained from a K. pneumoniae strain from China. This gene differs from blaCTX-M-14 by a single nucleotide, which leads to a Ser220Arg substitution. The characteristics of the novel enzyme have not yet reported. However, like the isolates with CTX-M-14, the isolate with CTX-M-38 had a much higher level of resistance to cefotaxime than ceftazidime, suggesting that the two enzymes have similar biochemical characteristics.
Two plasmid-mediated AmpC enzymes, DHA-1 and CMY-2, were detected in this study. We have reported previously the occurrence of DHA-1 in K. pneumoniae isolates from two hospitals in southern Taiwan and the rapid spread of blaCMY-2 in a university hospital in southern Taiwan (37, 41, 42). The present study demonstrated the countrywide spread of blaDHA-1 and revealed that the enzyme was the most commonly identified AmpC enzyme in K. pneumoniae in Taiwan. CMY-2 was the only plasmid-mediated AmpC enzyme identified in E. coli and blaCMY-2-like genes were detected in 37.1% to 73.1% of extended-spectrum cephalosporin-resistant E. coli isolates from four hospitals, indicating that that the widespread of CMY-2-producing E. coli has become a serous problem in Taiwan.
RAPD analysis showed genetic diversity among isolates with blaCMY-2, blaCTX-M-14, or blaCTX-M-3
from different hospitals. Although a few isolates with blaCMY-2, blaCTX-M-14, or blaCTX-M-3 from different hospitals showed the same RAPD patterns (Fig. 1 and 2), further pulsed-fielded gel electrophoresis analysis revealed that they all belonged to different clones (data not shown).
Similar plasmids containing blaCMY-2, blaCTX-M-14, or blaCTX-M-3 were observed among different strains from different hospitals. These findings together suggest that horizontal transfer and interhospital spread of similar resistance plasmids played a role in the dissemination of these resistance genes in Taiwan.
In this study, 12.0% of the 291 E. coli isolates and 16.3% of the 282 K. pneumoniae isolates were found to contain two or three β-lactamases involved in resistance to extended-spectrum
cephalosporins. ESBL production was not detected in 5 (2.8%) of 176 ESBL-producing E. coli and 5 (1.9%) of 265 ESBL-producing K. pneumoniae by the NCCLS confirmatory method due to the coproduction of ESBLs and AmpC, and the ten false-negative isolates were identified by the double-disk synergy method. This data indicate the complexity of the extended-spectrum
cephalosporin-resistant K. pneumoniae and E. coli isolates in Taiwan. The complexity may result from the widespread and increasing prevalence of various β-lactamases in Taiwan. Our result also suggests that isolates resistant to cephamycins and extended-spectrum cephalosporins should be tested by adjunctive methods such as the double-disk synergy method for ESBL detection at institutions where plasmid-mediated AmpC enzymes are prevalent.
參考文獻
1. Pitout, J. D. D., C. C. Sanders, and E. Sanders. 1997. Antimicrobial resistance with focus on β-lactam resistance in gram-negative bacilli. Am. J. Med. 103; 51-59.
2. Bush, K., G. A. Jacoby, and A. A. Medeiros. 1995. A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob. Agents Chemother.
39:1211-1233.
3. Nordmann, P. 1998. Trends in β-lactam resistance among Enterobacteriaceae. Clin. Infect.
Dis. 27 (Suppl. 1):S100-S106.
4. Paterson, D. L., L. Mulazimoglu, J. M. Casellas, W. C. Ko, H. Goossens, A. Von Gottberg, S.
Mohapatra, G. M. Trenholme, K. P. Klugman, J. G. McCormack, and V. L. Yu.2000.
Epidemiology of ciprofloxacin resistance and its relationship to extended-spectrum
beta-lactamase production in Klebsiella pneumoniae isolates causing bacteremia. Clin Infect Dis. 30:473-478.
5. Schiappa, D. A., M. K. Hayden, M. G. Matushek, F. N. Hashemi, J. Sullivan, K. Y. Smith, D.
Miyashiro, J. P. Quinn, R. A. Weinstein, and G. M. Trenholme. 1996. Ceftazidime-resistant Klebsiella pneumoniae and Escherichia coli bloodstream infection: a case-control and molecular epidemiologic investigation. J. Infect. Dis. 174:529-536.
6. Osano, E., Y. Arakawa, R. Wacharotayankun, M. Ohta, T. Horii, H. Ito, F. Yoshimura, and N.
Kato. 1994. Molecular characterization of an enterobacterial metallo-β-lactamase found in a clinical isolates of Serratia marcescens that shows imipenem resistance. Antimicrob. Agents Chemother. 38:71-78.
7. Riccio, M. L., N. Franceschini, L. Boschi, B. Caravelli, G. Cornaglia, R. Fontana, G.
Amicosante, and G. M. Rossolini. Characterization of the metallo-β-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of blaIMP allelic variants carried by gene cassettes of different phylogeny. Antimicrob. Agents Chemother. 44:1229-1235.
8. Lauretti, L., M. L. Riccio, A. Mazzariol, G. Cornaglia, G. Amicosante, R. Fontana, and G. M.
Rossolini. 1999. Cloning and characterization of blaVIM, a new integron-borne
metallo-β-lactamase gene from a Pseudomonas aeruginosa clinical isolate. Antimicrob.
Agents Chemother. 43:1584-1590.
9. Poirel, L., T. Naas, D. Nicolas, L. Collet, S. Bellais, J. D. Cavallo, and P. Nordmann. 2000.
Characterization of VIM-2, a carbapenem-hydrolyzing metallo-β-lactamase and its plasmid- and integron-borne gene from a Pseudomonas aeruginosa clinical isolate in France.
Antimicrob. Agents Chemother. 44:891-897.
10. Bauernfeind, A., I. Stemplinger, R. Jungwirth, and H. Giamarellou. 1996. Characterization of the plasmidic β-lactamase CMY-2, which is responsible for cephamycin resistance.
Antimicrob. Agents Chemother. 40:221-224.
11. Bauernfeind, A., I. Stemplinger, R. Jungwirth, R. Wilhelm, and Y. Chong. 1996.
Comparative characterization of the cephamycinase blaCMY-1 gene and its relationship with
other β-lactamase genes. Antimicrob. Agents Chemother. 40: 1926-1930.
12. National Committee for Clinical Laboratory Standards. 1999. Performance standards for antimicrobial susceptibility testing; ninth informational supplement. M100-S9. National Committee for Clinical Laboratory Standards. Wayne, Pa.
13. Liu, P. Y. F., J. C. Tung, S. C. Ke, and S. L. Chen. 1998. Molecular epidemiology of
extended-spectrum β-lactamase-producing Klebsiella pneumoniae isolates in a district hospital in Taiwan. J. Clin. Microbiol. 36: 2759-2762.
14. Yan, J. J., S. M. Wu, S. H. Tsai, J. J. Wu, and I. J. Su. 2000. Prevalence of SHV-12 among clinical isolates of Klebsiella pneumoniae producing extended-spectrum β-lactamases and identification of a novel AmpC enzyme (CMY-8) in southern Taiwan. Antimicrob Agents Chemother 44: 1438-1442
15. Yan, J. J., W. C. Ko, S. H. Tsai, H. M. Wu, Y. T. Jin, J. J. Wu JJ. 2001. Dissemination of CTX-M-3 and CMY-2 beta-lactamases among clinical isolates of Escherichia coli in southern Taiwan. J Clin Microbiol. 2000 Dec;38(12):4320-5.
16. Gniadkowski, M., I. Schneider, A. Palucha, R. Jungwirth, B. Mikiewicz, and A. Bauernfeind.
1998. Cefotaxime-resistant Enterobacteriaceae isolates from a hospital in Warsaw, Poland:
Identification of a new CTX-M-3 cefotaxime-hydrolyzing β-lactamases that is closely related to the CTX-M-1/MEN-1 enzyme. Antimicrob. Agents Chemother. 42:827-832.
17. Yan JJ, Ko WC, Hung YC, Chuang CL, Wu JJ. 2002 Emergence of Klebsiella pneumoniae isolates producing inducible DHA-1 β-lactamase in a university hospital in Taiwan. Journal of Clinical Microbiology 40(9):3121-26.
18. Senda, K., Y. Arakawa, K. Nakashima, H. Ito, S. Ichiyama, K. Shimokata, N. Kato, and M.
Ohta. 1996. Multifocal outbreaks of metallo-β-lactamase-producing Pseudomonas aeruginosa resistant to broad-spectrum-β-lactams, including carbapenems. Antimicrob.
Agents Chemother. 40:349-353.
19. Senda, K., Y. Arakawa, S. Ichiyama, K. Nakashima, H. Ito, S. Ohsuka, K., Shimokata, N. Kato, and M. Ohta. 1996. PCR detection of metallo-β-lactamase gene (blaIMP) in gram-negative rods resistant to broad-spectrum β-lactams. J. Clin. Microbiol. 34:2909-2913.
20. Hirakata, Y., K. Izumikawa, T. Yamaguchi, H. Takemura, H. Tanaka, R. Yoshida, J. Matsuda, M.
Nakano, K. Tomono, S. Maesaki, M. Kaku, Y. Yamada, S. Kamihira, and S. Kohno. 1998.
Rapid detection and evaluation of clinical characteristics of emerging multiple-drug-resistant gram-negative rods carrying the metallo-β-lactamase gene blaIMP. Antimicrob. Agents Chemother. 42:2006-2011.
21. Ito, H., Y. Arakawa, S. Ohsuka, R. Wacharotayankun, N. Kato, and M. Ohta. 1995.
Plasmid-mediated dissemination of the metallo-β-lactamase gene blaIMP among clinical isolated strains of Serratia marcescens. Antimicrob. Agents Chemother. 39:824-829.
22. Yan, J.J., W. C. Ko, J. J. Wu. 2001. Identification of a plasmid Encoding SHV-12, TEM-1, and a variant of IMP-2 metallo-beta-Lactamase, IMP-8, from a clinical isolate of Klebsiella
pneumoniae. Antimicrob Agents Chemother. 45:2368-71.
23. Yan, J.J., P. R. Hsueh, W. C. Ko, K. T. Luh, S. H. Tsai, H. M. Wu, and J. J. Wu. 2001.
Metallo-beta-lactamases in clinical pseudomonas isolates in Taiwan and identification of vim-3, a novel variant of the vim-2 enzyme. Antimicrob Agents Chemother. 45:2224-8.
24. Yan JJ, Ko WC, Tsai SH, Wu HM, Wu JJ. 2001 Outbreak of infection with
multidrug-resistant Klebsiella pneumoniae carrying blaIMP-8 in a university medical center in Taiwan. Journal of Clinical Microbiology 39:4433-4439.
25. Yan JJ, Ko WC, Chuang CL, Wu JJ. 2002. Metallo-β-lactamase-producing Enterobacteriaceae isolates in a university hospital in Taiwan: prevalence of IMP-8 in Enterobacter cloacae and first identification of VIM-2 in Citrobacter freundii. Journal of Antimicrobial Chemotherapy 50:503-11.
26. Bauernfeind, A., H. Grimm, and S. Schweighart. 1990. A new plasmidic cefotaximase in a clinical isolate of Escherichia coli. Infection 18: 294-298.
27. Matthew, M., M. Harris, M. J. Marshall, and G. W. Rose. 1975. The use of analytical isoelectric focusing for detection and identification of β-lactamases. J. Gen. Microbiol.
88:169-178.
28. Provence, D. L., and R. Curtiss III. 1994. Gene transfer in gram-negative bacteria, p.
319-347. In P. Gerhardt, R. G. E. Murray, W. A. Wood, and N. R. Krieg (ed.), Methods for general and molecular bacteriology. American Society for Microbiology, Washington, D. C.
29. Southern, E. M. 1975. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
30. Piggot, P. J., M. Amjad, J. J. Wu, H. Sandoval, and J. Castro. 1990. Genetic and physical maps of Bacillus subtilis 168, p. 493-543. In C. R. Harwood and S. M. Cutting (ed.), Molecular biology methods for bacillus. John Wiley & Sons Ltd., West Sussex, England.
FIG. 1. RAPD patterns of 15 CMY-2-producing (A), 16 CTX-M-3-producing (B), and 20 CTX-M-14-producing E. coli isolates randomly selected from different hospitals. Numbers designating RAPD types generated with either primer ERIC2 or M13 are shown above the gels.
Numbers designating isolates and numbers designating RAPD types obtained by combination of the results with the two primers are shown below the gels.
FIG. 2. RAPD patterns of 16 CTX-M-14-producing (A) and 21 CTX-M-3-producing K.
pneumoniae isolates randomly selected from different hospitals. Numbers designating RAPD types generated with either primer ERIC2 or M13 are shown above the gels. Numbers designating isolates and numbers designating RAPD types obtained by combination of the results with the two primers are shown below the gels.
FIG. 3. EcoRI restriction patterns of conjugative plasmids from 13 CMY-2-producing E. coli isolates (A), 17 CTX-M-14-producing E. coli isolates (B), and 18 CTX-M-3-producing K.
pneumoniae isolates (C). The numbers designating the donor isolates are shown above the gels, and numbers designating restriction patterns are shown below the gels.
TABLE 1. Distribution of ESBLs and AmpC enzymes among extended-spectrum β-lactam-resistant E. coli and K. pneumoniae isolates collected from seven medical centers in Taiwan
No. (%) of isolates with ESBLs or AmpC enzymes from hospital:
Species and the presence of
ESBL or AmpCa N1 N2 C1 C2 C3 S E Total
E. coli 78 33 84 41 18 19 18 291
CMY-2-like 57 (73.1) 0 (0) 40 (47.6) 13 (31.7) 8 (44.4) 1 (5.3) 8 (44.4) 127 (43.6) CTX-M 21 (26.9) 28 (84.8) 47 (56.0) 28 (68.3) 8 (44.4) 17 (89.5) 9 (50.5) 158 (54.3) CTX-M-1 group 5 (6.4) 4 (12.1) 19 (22.6) 5 (12.2) 2 (22.2) 2 (10.5) 4 (22.2) 41 (14.1) CTX-M-9 group 16 (20.5) 24 (72.7) 28 (33.3) 23 (56.1) 6 (33.3) 15 (78.9) 5 (27.8) 117 (40.2) SHV-5-like 10 (12.8) 3 (9.1) 5 (6.0) 4 (9.8) 2 (22.2) 3 (15.8) 0 (0) 27 (9.3) Noneb 1 (1.3) 3 (9.1) 4 (4.8) 1 (2.4) 3 (16.7) 0 (0) 2 (11.1) 14 (4.8)
K. pneumoniae 58 59 78 18 23 37 9 282
CMY-2-like 4 (6.9) 1 (1.7) 4 (5.1) 0 (0) 0 (0) 1 (2.7) 0 (0) 10 (3.5) DHA-1-like 17 (29.3) 1 (1.7) 5 (6.4) 1 (5.6) 3 (13.0) 4 (10.8) 0 (0) 31 (11.0) CTX-M 21 (36.2) 21 (35.6) 67 (85.9) 15 (83.3) 16 (69.6) 10 (27.0) 5 (55.6) 155 (55.0) CTX-M-1 group 12 (20.7) 7 (11.9) 44 (56.4) 8 (44.4) 14 (60.9) 8 (21.6) 5 (55.6) 98 (34.8) CTX-M-9 group 9 (15.5) 14 (23.7) 23 (29.5) 7 (38.9) 2 (8.7) 2 (5.4) 0 (0) 57 (20.2) SHV 34 (58.6) 39 (66.1) 18 (23.1) 1 (5.6) 7 (30.4) 32 (86.5) 4 (44.4) 135 (47.9) SHV-2-like 0 (0) 5 (8.5) 1 (1.3) 1 (5.6) 0 (0) 2 (5.4) 0 (0) 9 (3.2) SHV-5-like 34 (58.6) 34 (57.6) 17 (21.8) 0 (0) 7 (30.4) 30 (81.1) 4 (44.4) 126 (44.7) Noneb 2 (3.4) 1 (1.7) 1 (1.3) 1 (5.6) 0 (0) 0 (0) 0 (0) 5 (1.8)
aBased on the results of PCR and IEF.
bNo ESBLs and AmpC were detected.
19
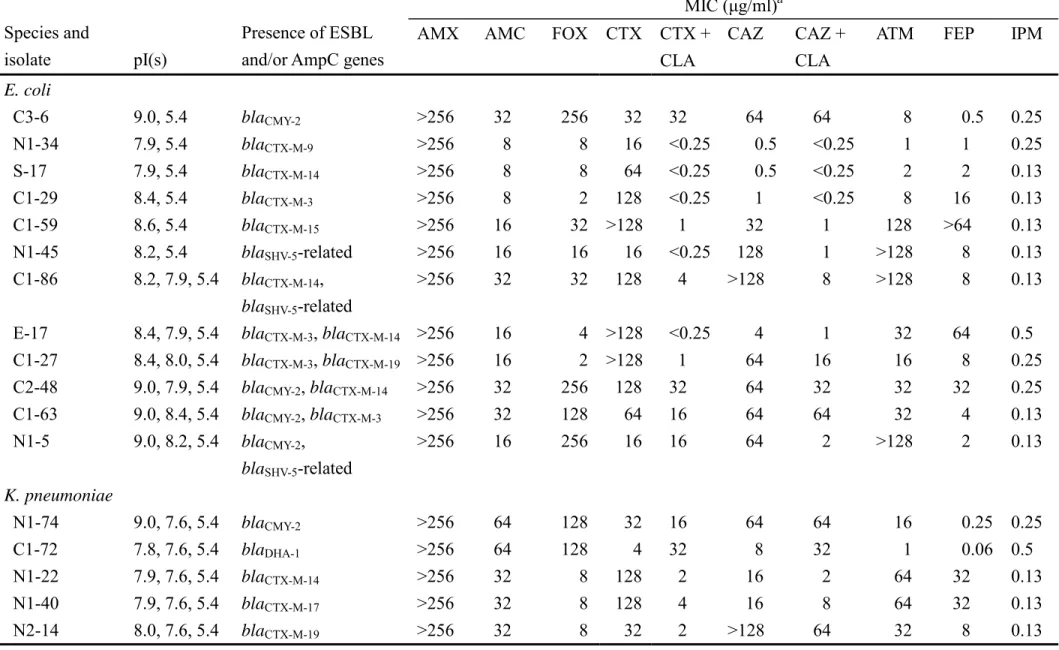
TABLE 2. MICs of selected β-lactam agents for representative clinical isolates of K. pneumoniae and E. coli that produced ESBLs and/or acquired AmpC enzymes
MIC (µg/ml)a Species and
isolate pI(s)
Presence of ESBL and/or AmpC genes
AMX AMC FOX CTX CTX +
CLA
CAZ CAZ +
CLA
ATM FEP IPM
E. coli
C3-6 9.0, 5.4 blaCMY-2 >256 32 256 32 32 64 64 8 0.5 0.25 N1-34 7.9, 5.4 blaCTX-M-9 >256 8 8 16 <0.25 0.5 <0.25 1 1 0.25 S-17 7.9, 5.4 blaCTX-M-14 >256 8 8 64 <0.25 0.5 <0.25 2 2 0.13 C1-29 8.4, 5.4 blaCTX-M-3 >256 8 2 128 <0.25 1 <0.25 8 16 0.13 C1-59 8.6, 5.4 blaCTX-M-15 >256 16 32 >128 1 32 1 128 >64 0.13 N1-45 8.2, 5.4 blaSHV-5-related >256 16 16 16 <0.25 128 1 >128 8 0.13 C1-86 8.2, 7.9, 5.4 blaCTX-M-14,
blaSHV-5-related
>256 32 32 128 4 >128 8 >128 8 0.13
E-17 8.4, 7.9, 5.4 blaCTX-M-3, blaCTX-M-14 >256 16 4 >128 <0.25 4 1 32 64 0.5 C1-27 8.4, 8.0, 5.4 blaCTX-M-3, blaCTX-M-19 >256 16 2 >128 1 64 16 16 8 0.25 C2-48 9.0, 7.9, 5.4 blaCMY-2, blaCTX-M-14 >256 32 256 128 32 64 32 32 32 0.25 C1-63 9.0, 8.4, 5.4 blaCMY-2, blaCTX-M-3 >256 32 128 64 16 64 64 32 4 0.13 N1-5 9.0, 8.2, 5.4 blaCMY-2,
blaSHV-5-related
>256 16 256 16 16 64 2 >128 2 0.13
K. pneumoniae
N1-74 9.0, 7.6, 5.4 blaCMY-2 >256 64 128 32 16 64 64 16 0.25 0.25 C1-72 7.8, 7.6, 5.4 blaDHA-1 >256 64 128 4 32 8 32 1 0.06 0.5 N1-22 7.9, 7.6, 5.4 blaCTX-M-14 >256 32 8 128 2 16 2 64 32 0.13 N1-40 7.9, 7.6, 5.4 blaCTX-M-17 >256 32 8 128 4 16 8 64 32 0.13 N2-14 8.0, 7.6, 5.4 blaCTX-M-19 >256 32 8 32 2 >128 64 32 8 0.13
C2-20 7.9, 7.6, 5.4 blaCTX-M-38 >256 32 16 >128 8 8 1 32 32 1 S-33 8.4, 7.6, 5.4 blaCTX-M-3 >256 16 2 64 0.13 2 1 4 16 0.25 N2-4 7.6 blaSHV-2-related >256 16 4 16 <0.25 8 1 4 4 0.13 N2-17 8.2, 7.6, 5.4 blaSHV-5-related >256 16 32 16 0.5 128 2 >128 2 0.13 C1-61 7.9, 7.6 blaCTX-M-14,
blaSHV-2-related
>256 16 4 >128 1 8 1 16 16 0.25
C1-88 8.4, 8.2, 7.6, 5.4
blaCTX-M-3, blaSHV-5-related
>256 16 4 64 4 64 1 64 16 0.25
C1-31 8.2, 7.9, 7.6, 5.4
blaCTX-M-9, blaSHV-5 >256 32 64 64 4 >128 8 >128 16 0.5
S-12 9.0, 8.4, 7.6, 5.4
blaCMY-2, blaCTX-M-3 >256 64 256 64 16 >128 64 >128 >64 0.25
N1-92 9.0, 8.2, 7.6 blaCMY-2, blaSHV-5-related
>256 32 256 64 32 >128 128 >128 8 0.5
N1-53 7.9, 7.8, 7.6, 5.4
blaDHA-1, blaCTX-M-9 >256 64 >256 64 4 32 >128 32 4 0.25
N1-17 8.2, 7.8, 7.6, 5.4
blaDHA-1, blaSHV-5-related
>256 64 >256 32 32 >128 >128 >128 2 0.25
C1-89 8.4, 8.2, 7.8, 7.6, 5.4
blaDHA-1, blaCTX-M-3, blaSHV-5-related
>256 32 >256 >128 64 >128 128 >128 64 0.5
C1-112 8.2, 7.9, 7.8, 7.6
blaDHA-1, blaCTX-M-14, blaSHV-5-related
>256 32 128 >128 64 >128 8 >128 64 0.5
aAMX, amoxicillin; AMC, amoxicillin-clavulanic acid; FOX, cefoxitin; CTX, cefotaxime; CTX+CLA, cefotaxime and clavulanic acid (2 µg/ml);
CAZ, ceftazidime; CAZ+CLA, ceftazidime and clavulanic acid (2 µg/ml); ATM, aztreonam; FEP, cefepime; IPM, imipenem.