國立臺灣大學生物資源暨農學院園藝學系 碩士論文
Department of Horticulture College of Bioresources and Agriculture
National Taiwan University Master Thesis
秋季土壤水分變化對臺灣亞熱帶低海拔地區 低需冷性梨樹地上部與根系生長之影響
Effects of soil water status on late season shoot and root growth of low chill Asian pears in lowland
subtropical Taiwan
蕭元彰
Yuan-Chang Hsiao
指導教授:李國譚 博士 Advisor:Dr. Kuo-Tan Li
中華民國 100 年 1 月
January, 2011
目錄
目錄... i
圖目錄... ii
摘要... iii
Abstract...v
第一章 總論-前人研究及試驗假說 ...1
一、 前言 ... 1
二、 前人研究 ... 3
(一) 臺灣地區之降雨與氣候趨勢 ... 3
(二) 土壤缺水對果樹作物生長之影響 ... 4
(三) 浸水對土壤環境之影響 ... 9
(四) 土壤淹水對果樹作物生長之影響 ... 12
(五) 梨樹根系之生長特性 ... 23
(六) 秋根對薔薇科落葉果樹生長之影響 ... 24
(七) 溫帶地區落葉果樹季節性根部與地上部生長之關係 ... 26
(八) 亞熱帶地區常綠果樹季節性根部與地上部生長之關係 ... 27
(九) 果樹作物根系與地上部生長之關係 ... 27
(十) 果樹作物根系之觀測方法 ... 28
三、 試驗假說 ... 31
四、 結論 ... 31
參考文獻... 32
第二章 秋季土壤水分變化對臺灣亞熱帶低海拔地區低需冷性梨樹地上部與根系 生長之影響... 48
摘要... 48
一、前言... 50
二、材料與方法... 52
三、結果... 60
四、討論... 78
參考文獻... 107
第三章 結論與未來研究方向 ...113
一、 結論 ...113
二、 未來研究方向 ...114
參考文獻...117
附錄...119
圖目錄
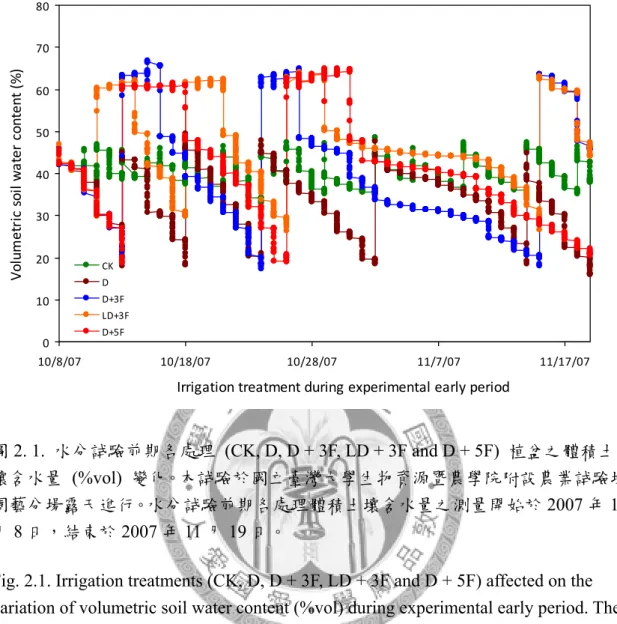
圖2. 1. 水分試驗前期各處理植盆之體積土壤含水量變化。 ... 85
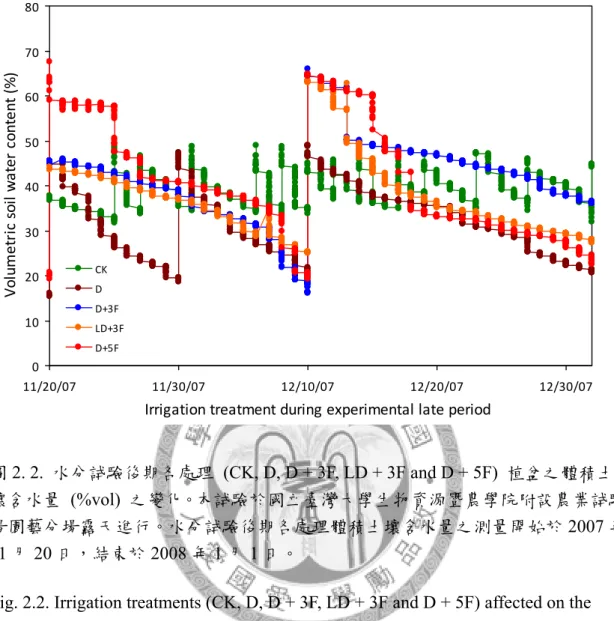
圖2. 2. 水分試驗後期各處理植盆之體積土壤含水量之變化。 ... 86
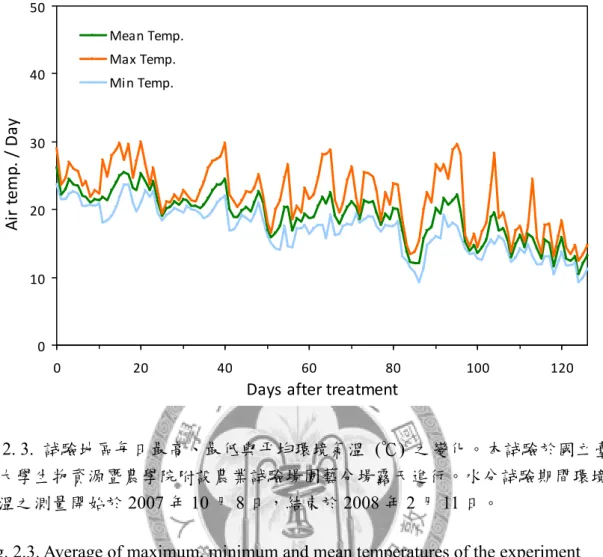
圖2. 3. 試驗地區每日最高、最低與平均環境氣溫之變化。 ... 87
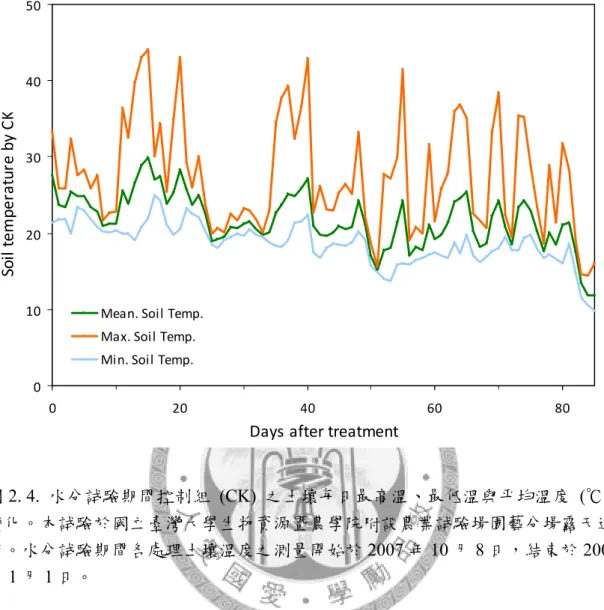
圖2. 4. 水分試驗期間控制組之土壤每日最高溫、最低溫與平均溫度變化。 ... 88
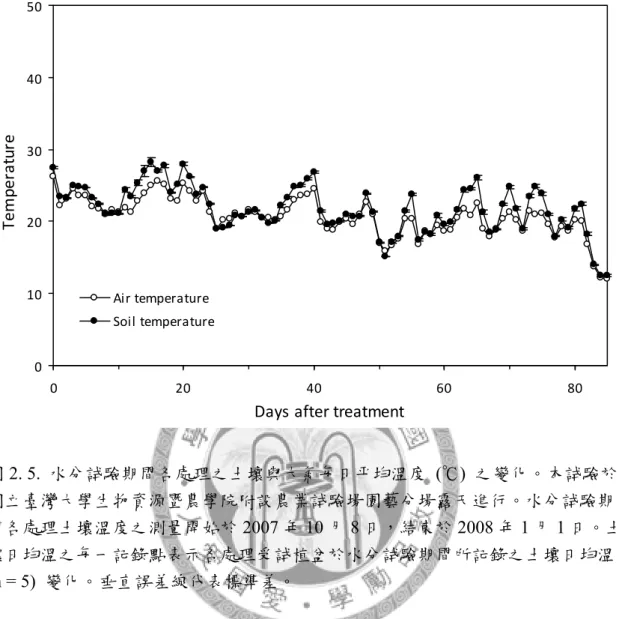
圖2. 5. 水分試驗期間各處理之土壤與大氣每日平均溫度之變化。 ... 89
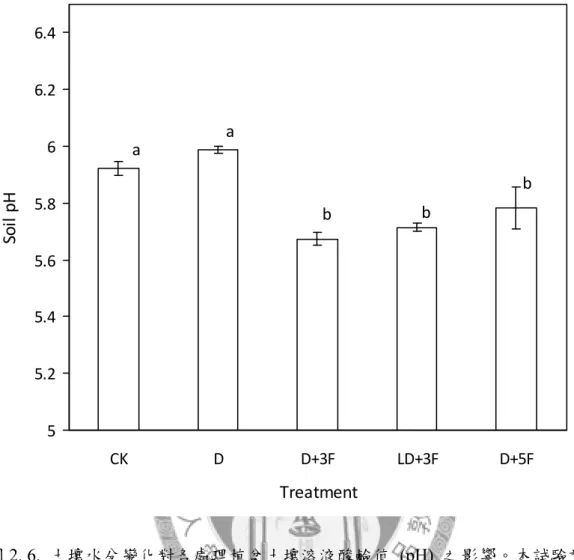
圖2. 6. 土壤水分變化對各處理植盆土壤溶液酸鹼值之影響。 ... 90
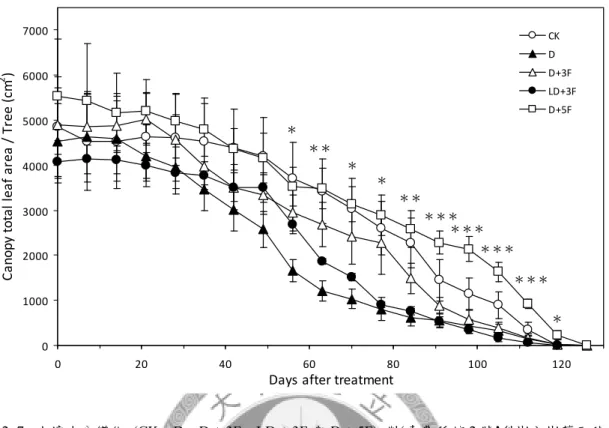
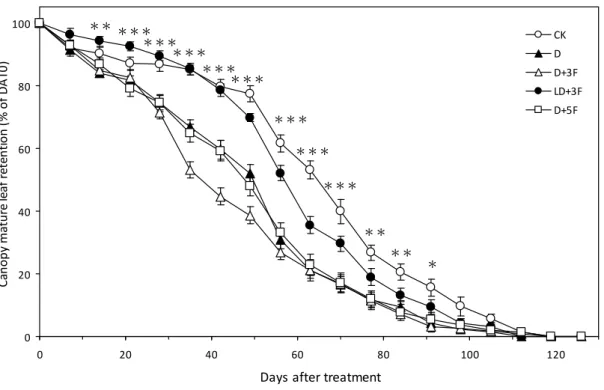
圖2. 7. 土壤水分變化對‘臺農種苗 2 號’梨樹全樹葉面積生長之影響。... 91
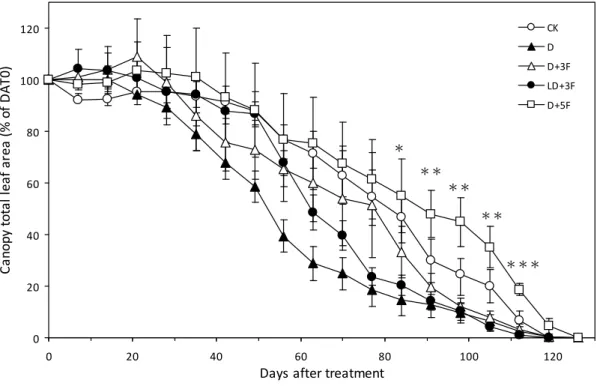
圖2. 8. 土壤水分變化對‘臺農種苗 2 號’梨樹全樹葉面積比例之影響。... 92
圖2. 9. 土壤水分變化對‘臺農種苗 2 號’梨樹宿存葉比例之影響。... 93
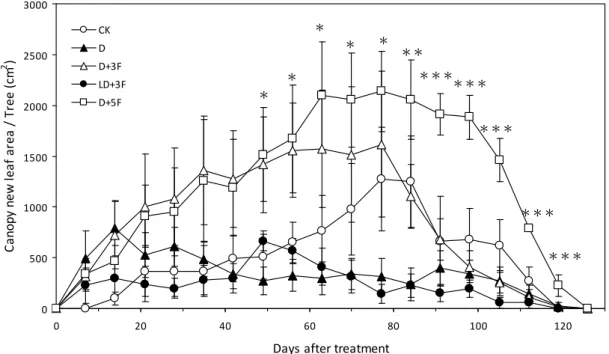
圖2. 10. 土壤水分變化對‘臺農種苗 2 號’梨樹新葉生長之影響。... 94
圖2. 11. 土壤水分變化對‘臺農種苗 2 號’梨樹有效根系分佈比例之影響。... 95
圖2. 12. 土壤水分變化對水分試驗前期 ‘臺農種苗 2 號’ 梨樹新生根生長模式之影 響。... 96
圖2. 13. 土壤水分變化對水分試驗前期 ‘臺農種苗 2 號’ 梨樹新生根存活率之影 響。... 97
圖2. 14. 土壤水分變化對水分試驗後期 ‘臺農種苗 2 號’ 梨樹新生根生長模式之影 響。... 98
圖2. 15. 土壤水分變化對水分試驗前期 ‘臺農種苗 2 號’ 梨樹標定新生根維持白色 比率之影響。... 99
圖2. 16. 土壤水分變化對水分試驗前期 ‘臺農種苗 2 號’ 梨樹標定新生根存活率之 影響。... 100
圖2. 17. 土壤水分變化對水分試驗後期 ‘臺農種苗 2 號’ 梨樹標定新生根維持白色 比率之影響。... 101
圖2. 18. 土壤水分變化對水分試驗後期 ‘臺農種苗 2 號’ 梨樹標定新生根存活率之 影響。... 102
圖2. 19. 水分試驗前期 ‘臺農種苗 2 號’ 梨樹植株宿存葉下降比例與標定新生根維 持白色比率之相關性。... 103
圖2. 20. 水分試驗前期 ‘臺農種苗 2 號’ 梨樹植株宿存葉下降比例與標定新生根存 活率之相關性。... 104
圖2. 21. 水分試驗後期 ‘臺農種苗 2 號’ 梨樹植株宿存葉下降比例與標定新生根維 持白色比率之相關性。... 105
圖2. 22. 水分試驗後期 ‘臺農種苗 2 號’梨樹植株宿存葉下降比例與標定新生根存 活率之相關性。... 106
摘要
本研究於國立臺灣大學生物資源暨農學院附設農業試驗場園藝分場戶外進 行,處理始於2007 年 10 月 8 日,結束於 2008 年 1 月 3 日,水分試驗期間調查測 量植株地上部葉片與根系之生長,探討秋季土壤水分變化對臺灣亞熱帶低海拔地 區低需冷性梨樹地上部與根系生長之影響。本試驗以2 年生,嫁接於鳥梨 (Pyrus lindleyi Rehder) 實生苗之蜜雪梨 (Pyrus pyrifolia Nakai cv. ‘Tainung No.2’) 裸根苗 為試驗材料,分別進行5 種灌溉處理,處理一為控制組 (CK),該處理之土壤體積 含水量 (θv) 維持於 30%-45%。處理二 (D) 之植株於灌溉後停止供水至 θv < 20%
再給水至 θv = 45%。處理三 (D+3F) 之植株於灌溉後停止供水至 θv < 20%時行淹 水處理3 日。處理四 (LD+3F) 之植株於灌溉後停止供水至 θv = 25-30%時行淹水 處理3 日。處理五 (D+5F) 之植株於灌溉後停止供水至 θv < 20%時行淹水處理 5 日。試驗結果如下,比較水分試驗期間各受試植株地上部葉片之生長可知,乾旱 (D) 與土壤水分劇烈變化 (D + 3F, D + 5F) 促進植株成熟葉提早脫落。而由水分試驗期 間梨樹根系之生長可知,水分試驗前期各受試植株具2 至 3 次之生長高峰,於冬 季水分試驗後期根系生長量降低,1 月後新生根大量死亡,各受試植株僅保留少數 白色與黃色根越冬。比較水分試驗前期梨樹根系生長模式可知,劇烈土壤水分變 化降低新生根存活率;CK、D、D + 3F、LD + 3F 與 D + 5F 處理於水分試驗前期 之新生根總存活率分別為89.26±1.21%、82.95±2.43%、74.14±6.14%、92.28±2.10%
與68.08±4.30%,各處理間比較達顯著差異水準。比較水分試驗前期與後期各受試 植株標定新生根維持白色比率與新生根存活力下降至50%所需之日數可知,CK、
D 與 LD + 3F 處理之標定新生根存活力高於 D + 3F 與 D + 5F 處理。分析水分試驗 前期植株宿存葉下降比例與標定新生根存活力兩者間之相關性可知,植株宿存葉 下降比例與與新生根存活力呈顯著負相關;新生根存活力隨宿存葉比例下降而減 少,水分試驗前期新生根維持白色比率、新生根存活率與植株宿存葉下降比例之 決定係數分別為0.5562 與 0.7371。
關鍵字:秋根、新生根、宿存葉、新葉、體積土壤含水量
Abstract
This research was carried out in Horticulture Research and Experimental Farm, College of Bio-Resources and Agriculture of National Taiwan University from October 8, 2007 to January 3, 2008. We measured shoot and root growth of low chill Asian pears in lowland subtropical Taiwan to document the effect of soil water status on late season shoot and root growth of potted two-year-old ‘Tainung No.2’/Lindley’s pear trees. Trees were irrigated with five irrigation treatments: Control treatment (CK), plants were maintained at 30-45% soil water content; Drought treatment (D), plants were not watered until soil water content below 20% and then irrigated back to 45%; Drought and 3 three-day Flooding (D+3F), plants were not watered until soil water content below 20% and then flooded for 3 days; Moderate Drought and 3 three-day Flooding (LD+3F), plants were not watered until soil water content to 20-30% and then flooded for 3 days; Drought and five-day Flooding (D+5F), plants were not watered until soil water content below 20% and then flooded for 5 days. Mature leaves after D, D+3F and D+5F treatments defoliated sooner than those subjected to CK and LD+3F. During the early period of the experiment, there were two to three peaks of root growth. In January, the vast majority of the new roots turned dark brown or was invisible, leaving on a small portion of survival white and yellow roots. By comparing the five irrigation treatments during the early period of the experiment, soil moisture fluctuation
significantly decreased new root survival. New root survival (%) for the five irrigation treatments during the early period were: 89.26±1.21% (CK), 82.95±2.43% (D),
74.14±6.14% (D+3F), 92.28±2.10% (LD+3F), and 68.08±4.30% (D+5F). Overall by analyzing the longevity at when 50% of the new white roots turned yellow and at when 50% of the new roots turned dark brown, black or invisible, we found that plants subjected to CK, D and LD+3F had a higher new root survivorship compared to those subjected to D+3F and D+5F. By analyzing the correlation of canopy mature leaf retention (% of DAT0) versus cohort new roots remaining white (%) in DAFR40, we found that there was a high correlation between these two events. The coefficient of determination, r2, between new roots remaining white and canopy mature leaf retention (% of DAT0), and between new root survival and canopy mature leaf retention (% of DAT0) were 0.5562 and 0.7371, respectively.
Keywords: Autumn roots, New roots, Volunetric soil water content, Mature leaves, New leaves.
第一章 總論-前人研究及試驗假說
一、前言
梨 (Pyrus spp.) 為薔薇科 (Rosaceae),梨亞科 (Pomoideae),梨屬 (Pyrus)多 年生落葉喬木,全世界約有35 個種,野生於亞洲、歐洲及非洲等地,主要集中於 地中海、高加索、中亞及中國,為世界主要經濟果樹,栽培分類上可概分成兩大 類:東方梨 (oriental pear) 與西洋梨 (common pear; European pear; Pyrus communis L.) (廖,1995; Challice, 1974)。西洋梨原產於歐洲,主要種植於歐美地區。東方梨 原產於中國,主要種植於亞洲地區,可分為秋子梨 (Ussurian pear; Pyrus ussuriensis Max.)、白梨 (White pear; Ya pear; Pyrus brestchneideri Rehder) 及砂梨 (Sand pear;
Japanese pear; Pyrus pyrifolia Nakai; Pyrus serotina Rehd.) 三大品系 (康,1997; 櫻 井,1944)。其中白梨與秋子梨為中國北方品系,多分佈於黃河以北至東北等冷凉 乾燥地區;砂梨為中國南方品系,分佈於淮河以南等溫暖潮濕地區 (康,1997),
而砂梨也為臺灣主要梨之栽培品系 (廖,1995)。
砂梨為臺灣主要經濟果樹之一,根據調查資料顯示,民國 96 年梨收穫面積達 8098 公頃,主要種植區域依序為臺中縣、苗栗縣與新竹縣,合計約佔全臺梨種植 面積的89% (行政院農業委員會,2007)。臺灣地區目前梨主要栽培模式可分成三 大類:一是高海拔地區栽培的日本梨、二是平地的橫山梨、第三則是高接梨 (吳,
1994)。近年來由於消費水準提高,橫山梨因品質較差而售價偏低,造成平地梨園 多改以生產高接梨為主;橫山梨為副之栽培模式 (廖,1995),佔全臺種植面積約 69% (林和高,2005)。
然而近年來由於高接梨栽培成本過高,加上氣候變遷與國土復育等限制,梨 產業競爭力逐漸降低,故選拔品質優良,適合於低海拔栽培生產之低需冷性梨品 種,可降低生產成本,提升梨產業競爭力。故行政院農業委員會農業試驗所與臺
中區農業改良場自1976 年開始,分別以高品質之高需冷性梨與低需冷性之橫山梨 進行雜交,以此自後裔中選育出數個適合於低海拔地區栽培之低需冷性梨新品 種,例如:臺農一號 (明福梨)、臺種苗農二號 (蜜雪梨)、臺農三號 (玉金香梨)、
臺中一號 (福來梨)、臺中二號 (晶圓梨)、臺中三號 (晶翠梨),目前已陸續技術轉 移推廣中 (行政院農業委員會農業試驗所,2007; 臺中區農業改良場,2009)。
臺灣位處東亞溫帶及熱帶季風氣候區,主要水資源為雨水,主要降水期為 2-4 月的春雨、5-6 月的梅雨與夏秋的颱風雨,夏末至初春為臺灣中南部的旱季,年平 均雨量約2500mm (中央氣象局,2003; 陳,1993)。雖臺灣氣候具高溫多雨之特徵,
但因中央山脈南北貫穿全島,造成臺灣各河流坡陡且短、水資源涵養能力差,且 中、小尺度地區性天氣系統常受氣候帶與複雜地形影響,降雨時間與空間上分佈 不均勻,山區常因地形抬升作用並配合部分天氣系統變化,帶來短時間的極端降 水 (陳,1993),相對易造成作物淹水與排水後缺水等現象。
臺灣地區平均年降雨量為全球平均值之 3 倍,但因降水之時間與空間分佈變 化大,加上環境破壞、都市化與人口密度上升之影響,不同地區水資源利用極不 均勻,一般而言,臺灣北部及東部地區年降水量大於蒸發量,缺水機率較低,中 南部因降水量多集中於梅雨季及夏季,乾季長達半年之久,具有潛在性乾旱的特 性,若當年梅雨不顯著或侵臺颱風不足,隔年則易缺水 (吳,1996; 臺灣大學全球 變遷研究中心,1997; Yu et al., 2002)。目前臺灣地區因氣候變遷與暖化等影響,未 來氣候型態將逐漸走向極端化,即溫度上升、乾旱期間變長,降雨日數減少或雨 季極端降雨,也易產生暴雨型態,氣象預測難度也升高 (經濟部水利署,2007; 廖 等,2009; Liu et al., 2009; Yu et al., 2006)。
目前臺灣地區梨樹生產多以高接梨栽培系統生產果實 (廖,1995)。許多例子 顯示,砧木之生長與生理會直接影響接穗的生長、產量與品質 (Lockard and Scineider, 1981; Twiorkoski and Miller, 2007)。水分為作物生長的重要環境因子,土
壤水分狀況會直接影響果樹生長、開花及結實等過程 (Jones et al., 1985; Schaffer et al., 1992)。臺灣氣候環境雖高溫多濕、年降雨量高,但降雨過度集中、土壤水分變 化劇烈;低窪排水不良與水田轉作地區於大量降水後常造成土壤淹水現象,果樹 為長期作物,栽培時間較長,無法臨時轉作,故探討臺灣低海拔地區栽培梨樹之 水分與根部生理有助於了解環境土壤水分變化下梨樹之生長模式。
對大多數果樹而言,淹水與乾旱逆境會影響植物體地上部生長,產生葉片缺 水、低氣孔導度、低淨二氧化碳同化率等抑制地上部生長之作用 (Jones et al., 1985;
Schaffer et al., 1992),相關研究大多僅探討單一逆境變化對作物之影響,然而作物 於淹水與乾旱交互作用下之生長變化卻尚未明瞭。一般植物體可分成地上部與地 下部,過去對作物生理之研究方向大多偏重於地上部之調查,然而植物根系對固 定植株與養、水分吸收具重要作用,但因土壤不可觀測性之限制,給根系生態學 之研究帶來一定的困難,故本研究將設計根窗,探討植物體根部於土壤水分變化 下之生長,進一步釐清梨樹根部與水分生理之關聯性。
為了解秋季土壤水分變化對臺灣亞熱帶低海拔地區低需冷性梨樹生長之影 響,本研究擬利用植盆控制水分,模擬不同程度之土壤水分變化,調查梨樹盆苗 地上部與地下部之營養生長,探討並比較不同土壤水分變化下梨樹之生長與生理。
二、前人研究
(一) 臺灣地區之降雨與氣候趨勢
臺灣位處東亞溫帶及熱帶季風氣候區,同時位於大陸與海洋之緩衝區,氣候 受大尺度環流之影響。臺灣主要水資源來源為雨水;主要降水期為2-4 月的春雨、
5-6 月的梅雨與夏秋的颱風雨,夏末至初春為臺灣中南部的旱季,年平均雨量約 2500mm (中央氣象局,2003; 陳,1993)。一般而言,臺灣地區降水之時間與空間 分佈變化大,北部及東部地區年降水量大於蒸發量,缺水機率較低,中南部因降
水量多集中於梅雨季及夏季,乾季長達半年之久,具潛在性乾旱的特性 (吳,1996;
臺灣大學全球變遷研究中心,1997; Yu et al., 2002)。
近年來臺灣地區因氣候變遷與暖化等影響,未來氣候型態將逐漸走向極端 化,即溫度上升、乾旱期間變長,降雨日數減少或雨季極端降雨,也易產生暴雨 型態,氣象預測難度也升高 (經濟部水利署,2007; 廖等,2009; Yu et al., 2006)。
Liu 等(2009) 利用量化方式以全球降雨氣候計畫 (Global Precipitation Climatology Project; GPCP) 於 1979-2007 年之觀測資料分析全球 17 個氣候區與臺灣地區極端 降雨與全球溫度之關係發現,全球溫度每上升1 度 K,全球前 10%之強降雨增加 約110%,而 30%-60%降雨等級之中、小強度降雨則減少約 20%。而全球溫度每增 加1 度 K,臺灣地區前 10 %之強降雨則增加約 140 %,而前 10 %小雨則減少約 70
%。強降雨的增加可能導致淹水,臺灣中、南部春冬兩季之中、小雨為保持土壤濕 潤及地下水之關鍵水源,其持續性減少將導致乾旱之威脅。
(二) 土壤缺水對果樹作物生長之影響
1. 土壤缺水影響果樹作物生長之反應
水為植物體的主要組成分,含量佔組織鮮重的 70-90%。水分一般由土壤或栽 培介質供應,為根部所吸收,供植物體利用。植物體藉水分進行光合作用、蒸散 作用、礦物元素吸收及各種生理之代謝;水不僅是光合作用的原料,植物體內各 種代謝作用也多在水溶液中進行。水分同時也是養分吸收和運輸的媒介,植物體 也藉由蒸散作用進行土壤養水分之吸收、運輸及體內溫度之調節 (Jones et al., 1985;
Kramer, 1983)。
土壤水分依其存在狀態可分為毛細管水 (capillary water)、吸著水 (hygroscopic water) 與重力水 (gravitational water) 三種,惟毛細管水可供根部所吸收利用。土 壤缺水 (water deficit) 導致作物乾旱逆境 (drought stress),然而作物依失水量不同
可分為暫時性凋萎 (temporary wilting) 及永久性凋萎 (permanent wilting) 兩種。土 壤於降水後,大量重力水經逕流排出後,土壤於飽和狀態所含之水量稱田間容水 量 (field capacity)。土壤於不供應水分的狀態下,土壤水分因植物體根部吸收與蒸 發散 (evaportranspiration; E.T.) 作用逐漸減少,當植株蒸散作用耗損之水分大於根 部所吸收之水分時,植株會造成暫時性萎凋之現象,暫時性萎凋常見於夏季中午,
然而午後根部吸收水量與植物體蒸散作用達到平衡時,植株將逐漸恢復正常之膨 壓,土壤水分含量於暫時萎凋點 (temporary wilting point) 之範圍內,只要再供應 水分,作物即可恢復正常。但土壤若不予供水,土壤水分持續下降,土壤內供根 部吸收之毛管水 (有效水) 耗盡,使得植株根部吸水不足,影響植株水分吸收,葉 片開始萎凋,此時若再加水,植株之萎凋狀態仍無法復原,此時之土壤含水量稱 永久萎凋點 (permanent wilting point) (Kramer, 1983; Kramer and Boyer, 1995a)。
木本植物對缺水之反應較草本植物敏感,主要因木本植物具樹幹,故具較高 的液壓阻力 (hydraulic resistance) (Lakso, 2003)。作物於不同之生長階段,需水量 皆不同,然而作物缺水導致乾旱逆境,作物於土壤缺水下會產生許多生理及型態 上之調適以抵抗乾旱逆境。作物於缺水狀態下,早期之反應為單葉面積的減少,
其次為成熟葉脫落與根系生長量增加,最後氣孔關閉以減少水分消耗 (Hasegawa, 2002)。
2. 土壤缺水對果樹作物水分狀態與光合作用生理之影響
對絕大多數果樹作物而言,乾旱逆境造成植物膨壓、氣孔導度與光合作用率 下降為其典型之生理反應 (Lasko, 2003)。然而乾旱對果樹作物水分狀態與光合作 用生理特別敏感,果樹作物對乾旱逆境最先出現的生理反應便是氣孔導度的降低 與中午莖部水勢下降,主要因失水與減低蒸散作用所造成 (Francisco et al., 2007;
Hasegawa, 2002)。
乾旱逆境造成作物失水,土壤缺水同時也會降低植物體水分狀態。植株中午
莖部水勢 (midday stem water potential) 可代表中午缺水逆境下作物整體的水分狀 態 (Scholander, 1965),短期土壤缺水明顯降低植物體中午莖部水勢與水分利用率 (water use efficiency),然而乾旱逆境下植株水分利用率下降之敏感度亦可作為植株 對抗乾旱逆境之指標 (Behboudian et al., 1994; Francisco et al., 2007; Lakso, 2003)。
對多數果樹作物而言,植物體光合作用與氣體交換對缺水逆境特別敏感。一 般而言,缺水逆境影響植物體光合作用之途徑有兩個:氣孔的關閉與膜結構的破 壞。土壤缺水造成氣孔關閉對植物體光合作用只有輕微的影響,乾旱逆境對光合 作用主要之影響為光系統Ⅱ效率的降低 (Hasegawa, 2002)。果樹作物於短期乾旱逆 境下植物體會先關閉氣孔,降低蒸散速率,以保持植物體水分狀態,雖植物體受 逆境影響水勢下降,但葉片膜結構不受影響,故光系統Ⅱ效率 (PS efficiency) Ⅱ 暫 不受輕度缺水所影響。但植物體於中度、嚴重或長期缺水逆境時葉綠體內類囊體 膜 (thylakoid membranes) 結構因缺水與高溫受破壞,進而影響光系統 (PS ; Ⅱ Ⅱ P680) 與電子傳遞鍊,此時光系統Ⅱ效率才會受影響降低;造成葉綠素螢光值 (chlorophyll fluorescence; Fv/Fm) 下降 (Flore and Lakso, 1989; Hasegawa, 2002;
Lakso, 2003) ,故乾旱逆境下植株葉綠素螢光值下降之敏感度可作為植株對抗乾旱 逆境之指標。
土壤早期缺水導致植物體光合作用速率降低主要是因為氣孔關閉所造成;土 壤缺水使根部合成離層酸 (abscisic acid; ABA) 能力增加,離層酸向地上部傳導,
葉片離層酸上升與細胞分裂素 (cytokinins) 降低造成氣孔關閉 (高,1994)。
3. 乾旱逆境對薔薇科果樹滲透勢調節之影響
植物體對乾旱逆境的另一個反應,即為溶質累積造成滲透勢的降低,此過程 稱滲透勢調節 (osmotic adjustment);植物體細胞內溶質濃度增加造成溶質濃度淨 增,滲透勢調節產生一個更為負值的葉片水分潛勢 (negative leaf water potential),
使得水分流向葉片,維持葉片膨壓,即:(Ψw (-) = Ψs (-) + Ψp (+),Ψw: total water
potential, Ψs: osmotic potential or solute potential, Ψp: Turgor or pressure potential)。滲 透勢調節可幫助葉片維持膨壓,使氣孔開張,於輕微的水分逆境下仍可持續進行 光合作用。山梨糖醇 (Sorbitol) 及甜菜鹼 (betaine) 為薔薇科果樹中 2 種常見的相 容性溶質(compatible solutes),為植物體細胞質中重要的滲透調節劑,作物受水分 逆境時,甜菜鹼與山梨糖醇會在細胞內累積而提高溶質濃度,使得水分仍能流向 葉片,進行滲透勢調節 (Gao et al., 2004; Hasegawa, 2002; Lakso, 2003; Wang et al., 1995)。
乾旱逆境會誘導酥梨 (Pyrus bretschneideri Rehd.‘Suly’) 成熟葉片甜菜鹼累 積,且於輕微的水分逆境下葉片甜菜鹼累積量即明顯上升。然而當水分逆境持續 發展時,甜菜鹼的累積不會因逆境強度的增加而持續增加累積的量,推測可能與 梨本身耐旱性質有關。樹體於乾旱逆境下外施甜菜鹼能促進枝條生長,且外施甜 菜鹼對枝條生長之促進於重度缺水逆境時更加明顯 (Gao et al., 2004)。
4. 土壤缺水對果樹作物地上部營養生長之影響
植物體地上部營養生長對缺水特別敏感;植物體地上部生長速率隨中午莖部 水勢下降而減少,中午莖部水勢與地上部生長速率呈線性負相關。植物體於缺水 初期的反應便是減緩地上部營養生長 (vegetative growth),尤其是葉片。在乾旱逆 境下,減少葉面積的擴大,使樹冠面積縮小以及降低蒸散速率,對植物而言是有 利的。乾旱逆境亦可能誘導植株老葉落葉,此過程稱為葉面積調節 (leaf area adjustment) 作用,主要為植物體水分來源受限時,降低全樹葉面積和蒸散作用之 機制 (Behboudian and Mills, 1997; Hasegawa, 2002; Lakso, 2003)。
5. 果樹水分狀態與果實發育及品質之關係
缺水易影響果實生長發育與品質;於新世紀梨盆栽試驗中發現,果實發育早 期缺水導致成熟果實鈣含量降低、可溶性醣類濃度增加、增加FSD (flesh spot decay) 發生比率,早期缺水後缺鈣與FSD 發生之原因,可能為缺水導致蒸散流降低、鈣
無法運移至果實,以及細胞分裂期產生細胞壁較薄弱所致 (Behboudian and Lawes, 1994)。新世紀梨果實於細胞分裂期缺水逆境試驗中發現,早期缺水逆境後若再度 灌溉,果實直徑仍會增加,恢復至原來完整灌溉的果徑 (Behboudian et al., 1994)。
6. 土壤缺水對果樹作物根系生長之影響
作物於乾旱環境中汲取水分主要依賴根系之長度與密度 (Buwalda, 1993)。根 系為土壤水分的直接吸收利用者,當土壤缺水時,作物根系首先感應並發出信號,
使植株對土壤缺水進行反應,同時根系之形態、结構、植物荷爾蒙也產生相對應 之變化,影響地上部的生長。多年生果樹對土壤缺水 (water deficit) 格外敏感,主 要因缺水導致植物組織內水力傳導係數 (hydraulic conductivity) 降低所致
(Buwalda, 1993; Jones et al., 1985; Kramer and Boyer, 1995b)。Waring 和
Schlesinger(1985) 則認為一般木本植物之根系無法生長於土壤水勢低於-0.7MPa 之 環境。
對絕大多數果樹作物而言,地上部生長對缺水較地下部敏感;於低土壤水勢 條件下,作物地上部停止生長,但根系卻繼續伸長,土壤缺水下根系的伸長被認 為有利於作物從土壤中尋找、吸收水分,維持作物生長與水分狀態,所以地上部 與地下部對乾旱逆境之反應通常認為是作物對乾旱條件之適應性機制 (Buwalda, 1993; Garnier et al., 1986; Hasegawa, 2000; Kramer and Boyer, 1995b)。於乾旱逆境 下,作物根系也會產生向水性 (hydrotropism) 以增加水分之搜索與吸收 (Kramer and Boyer, 1995b)。
即使如此,持續乾旱逆境會降低根系產量,於 1 年生蘋果‘Mutsu’盆苗乾旱試 驗可知,土壤水勢下降,植株根乾物重也會逐漸下降,兩者間呈正相關,且根部 之呼吸速率也隨植株根系乾物重下降而降低 (Psarras and Merwin, 2000)。
由上可知,適度土壤缺水增加作物根系之生長量,多數作物根系於土壤水勢
低於-50kPa 時,根的伸長速率才會下降 (Buwalda and Lenz, 1992; Richards, 1983)。
比較不同供水量 (100%灌水、50%灌水、25%灌水) 對‘Golden Declicious’ / M.9 蘋 果樹地上部與地下部生長可知,植株地上部之生物產量 (biomass) 隨土壤水分減 少而降低,然而比較根系之生物產量可知,正常灌溉蘋果樹之全樹根部乾物重為 50%灌溉的 95%,為 25%灌溉的 140%,顯示適度缺水會增加根系之生長 (Buwalda and Lenz, 1992)。一般認為土壤缺水誘導根系分佈範圍增加主要是因為根部要尋找 水分,而不是增加根的密度 (root length density);Buwalda (1993) 以 5 年生奇異果 植株為樣本,於試驗期間進行土壤灌溉與不灌溉處理,觀測半徑範圍2.75 公尺,
深1 公尺植株根系之發展,結果顯示正常灌溉植株之根系最遠僅發展至樹幹半徑 2 公尺之範圍,且根系總長度多分佈於半徑0.5-1.25 公尺處,然而未灌溉植株之根系 最遠延長至距植株2.75 公尺遠,但根系分佈深度仍維持於 1 公尺之範圍,然而缺 水下奇異果根系向外延伸而非向下延伸也是藤本植物特有的根系發展狀態。葡萄 植株於栽培時減少水分供應,地下部對地上部重量之比值增加,細根與白色吸收 根增加且也產生較多也較長之側根 (lateral root) (Richards, 1983)。
地下部與地上部之比例 (root : shoot ratio) 通常會反映出地上部枝條對養水份 的需求與供應,因此缺水會增加地下部與地上部之比例。地下部與地上部之比例 的增加與缺水有關,顯示光同化物分配往根系的比例增加 (Buwalda, 1993; Kramer and Boyer, 1995),植物體根系生長大約會消耗 1/3 地上部製造的光合產物,推測植 物體遭遇缺水逆境下地上部乾物重與根系分佈之原因,主要可能因輕度缺水會刺 激植物體根系伸長,因而消耗樹體之光合產物 (Buwalda, 1993)。
(三) 浸水對土壤環境之影響
土壤是由土體、液相與氣相所組成之三相系,三者之比例影響土壤之物理與 化學性質。土壤物理性質決定作物根系生長與發育之基本因子、影響作物生產力。
優良的土壤物理性質,至少須具備良好排水、通氣之條件,土壤植物才能有好的
生長與生產 (張,1980; Lyon and Buckman, 1948)。土壤為一個多孔體,孔隙裏主 要貯存水分與空氣,土壤中各相所佔之容積比率為動態而非固定的;液相與氣相 彼此有顯著反比之關係。降水時,水分進入土壤將除去同量體積之氣體;反之水 分則因排水、蒸發或植物生長吸收水分等因素被除去。正常通氣良好之土壤應具 10-30%之孔隙,而於浸水狀況下,土壤孔隙常降至 0%,土壤孔隙為水所填滿 (Grable, 1966)。
大氣中約含 79%氮 (N2)、20.4%氧 (O2) 及 0.03%的二氧化碳 (CO2),而土壤 中由於根及其他生物體的呼吸作用消耗氧氣、釋放二氧化碳,加之二氧化碳於土 壤中之擴散不若空氣中迅速,因此土壤空氣之二氧化碳濃度約為大氣的10-100 倍,氮氣濃度仍維持79%,氧氣濃度稍低。一般乾燥土壤中,1 立方公尺即有 5-24 克氧氣。土壤孔隙為土壤氣體與大氣交換之場所,由於氧氣與二氧化碳於大氣及 土壤分壓的不同,故引起大氣之氧氣向土壤擴散,而土壤之二氧化碳則向大氣中 擴散,產生動態平衡 (Kozlowski, 1984)。
1. 淹水對土壤氣體含量變化之影響
淹水逆境直接導致土壤缺氧,主要發生於瞬間大量降雨、土壤過度黏重、排 水不良、天然濕地、地下水位過高、河谷淹水與過量灌溉。氣體之交換主要藉由 擴散作用,土壤浸水使土壤孔隙為水分所填滿,氣體擴散空間或通路呈不連續性,
以致無法進行正常的氣體擴散。空氣中氧氣的擴散系數為0.205 cm2 / sec,於水中 僅為1×10-5 cm2 / sec;氣體於水中之擴散速率為空氣中的萬分之一,土壤中剩餘之 氧氣則因植株根系及微生物進行呼吸作用而逐漸消耗完,且土壤微生物行呼吸作 用釋放出之二氧化碳亦無法有效擴散至大氣中,造成土壤中氧氣缺乏,二氧化碳 濃度增加 (Drew and Sisworo, 1977; Kozlowski, 1982; Rowe and Beardsell, 1973)。
當土壤氧氣濃度低於正常值時,稱低氧 (hypoxia) 狀態,此情形於短期淹水、
過量灌溉、天然濕地環境或地下水位過高、排水不佳時即可能發生。若土壤經過 長時間淹水,低於淹水面土壤深層則完全缺氧,稱無氧 (anoxia) 狀態 (Kozlowski, 1984; Smith et al., 1990)。
於排水不良或淹水時,嫌氣性微生物會取代好氣性微生物進而分解有機質產 生甲烷、乙烷、丙烯與乙烯等氣體,植物體根部也會因缺氧產生乙烯 (Kozlowski, 1997)。
2. 淹水對土壤氧化還原電位與植物體土壤養分利用之影響
土壤浸水降低土壤氧化還原電位,易產生土壤還原反應與解離現象,排水良 好的土壤其氧化還原電位 (redox potentials) 為 +300mV 以上或更高,於浸水狀況 下,土壤氧化還原電位降低至 +300mV 以下或更低 (Kozlowski, 1997)。土壤氧化 還原電位降低使其易產生還原反應;例如:硝酸鹽 (NO3-; nitrate)、氧化錳 (MnO;
manganic oxide)、氧化鐵 (FeO; Ferric oxide)、硫酸鹽 (SO42-; sulfate) 等物質進行 還原反應,造成土壤離子型態改變,如Fe3+→Fe2+或NO3-→NO2-等解離現象,土壤 離子含量上升,因此於浸水之土壤常有錳、鐵離子量上升,過多之情形,高濃度 下常造成植物的離子毒害現象 (Patrick and Mahapatra, 1968)。
土壤浸水造成土壤中營養成分型態之改變而影響其利用性,其中磷酸鐵易解 離為磷酸亞鐵而流失,但也造成磷有效性提高,而於短時間內提供植物體利用 (Patrick and Mahapatra, 1968)。短期浸水也會提高鉀 (K)、硫 (S)、鉬 (Mo)、鈉 (Na) 等礦物元素的可利用性 (Rowe and Beardsell, 1973)。
正常情況下土壤所進行之硝化作用 (nitrification) 由 NH4+→NO2-→NO3-,再經 由植株根部吸收。於浸水狀況下由於氧化電位變化,使硝酸根離子經由去硝化作 用 (denitrification) 由 NO3-→NO2-→N2逸散,造成損失,且於去硝化過程中亞硝酸 根之生成亦有害植物,於浸水土壤中每日經由去硝酸化作用所逸散之硝酸根可達
20-50ppm (Grable, 1966)。
(四) 土壤淹水對果樹作物生長之影響
1. 淹水影響果樹作物生長之過程與反應
土壤淹水後對作物與環境最大之影響與變化即為缺氧所造成的缺氧逆境 (oxygen deficiency),但土壤淹水對作物發育所造成之影響並非單純僅由缺氧所造 成,尚受其他因子所影響。例如:土壤淹水間接造成土壤產生還原態的有毒物質、
土壤中無機離子的流失以及根代謝作用產生有毒物質等逆境與傷害 (Hasegawa, 2002; Schaffer, 1998)。土壤淹水後,植物體於缺氧下會產生許多生理及形態上的調 適反應,觀察果樹遭遇淹水逆境之反應,最先受到影響的是光合作用,主要是因 為氣孔關閉造成。一般而言,大多數作物於土壤淹水下最早出現之反應便是氣孔 導度下降 (Kozlowski, 1982; Kozlowski and Pallardy, 1984),之後光合作用率與蒸散 作用降低、根部與地上部之生長受抑制、葉片萎凋、根吸收離子效率降低,最終 造成植株死亡 (Kawase, 1981; Schaffer, 1998; Schaffer, et al., 1992)。Crane 和 Davies(1989) 以兔眼種藍苺 (Vaccinium ashei Reade) 植株為例,詳細整合連續淹水 處理對植株生長之反應,將植株連續淹水造成之生理現象分成3 種階段,階段 1 (phase1) 需時 2-5 日,主要造成氣孔導度下降、光合作用效率與量子產率 (quantum yield)降低、根部水力傳導係數(root hydraulic conductance) 降低與夏季花芽數減 少,階段2 (phase2) 需時 6-14 日,主要加重階段 1 之症狀並增加落葉率與營養生 長終止,階段3 (phase3) 需時 16-120 日,主要造成莖部水勢下降與果實皺縮,然 而連續淹水造成植株死亡需時46-120 日。
淹水逆境對作物影響程度依物種、樹齡與遭遇之環境因子而有不同
(Kozlowski, 1984),一般而言,淹水時間越長,對作物損害越大,植株排水後之恢 復力越低 (Andersen et al., 1985; Ismail and Noor, 1996; Schaffer, 1998; Schaffer et al.,
1992; Smith et al., 1990)。以 1-2 年生杏桃 (Prunus americana) 植株而言,土壤淹水 會降低植株存活率,持續淹水10-20 天,植株存活率即下降至 50%以下 (Hassan et al., 1986)。作物於生長季淹水所受之影響較為嚴重 (Heinicke, 1932; Kozlowski, 1984)。Andersen 等(1984) 以各種梨屬作物為試驗材料於春季 (4 月) 及秋季 (9 月) 進行淹水處理,結果顯示春季淹水處理之氣孔導度降低速率較秋季淹水處理快。
Olien(1987) 以 6 年生 ‘Macspur’ / M26 蘋果樹為試驗材料,分別春、夏與秋季進行
長達6 週的淹水處理,探討土壤淹水與植株生長之關係,結果顯示夏季與秋季淹
水處理之營養生長受抑制程度大於秋季淹水處理。對黃色百香果 (Passiflora edulis f. flavicarpa Deg) 而言,夏季淹水處理對地上部生長之抑制高於冬季淹水 (阮及 鄭,1986)。3 年生兔眼種藍苺‘Woodard’ (Vaccinium ashei Reade ‘Woodard’) 植株於 夏季高溫下連續淹水5 日存活率僅剩 67%,然而於春季低溫連續淹水 25 日之植株 存活率為83% (Crane and Davies, 1988a)。3 年生兔眼種藍苺‘Tifblue’ (Vaccinium ashei Reade ‘Tifblue’)於春季連續淹水處理 106-117 之植株存活率為 83%,夏季淹水 78-90 日之植株存活率僅剩 33% (Crane and Davies, 1988b)。對於蔓越莓 (Cranberry;
Vaccinium macropcarpan Ait.) 植株而言,休眠期淹水不會造成植株損傷,但於生長 季淹水則會快速造成植株損傷 (Schaffer et al., 1992)。
淹水逆境對植物生理與型態之影響程度可因其他環境因子而有所差異
(Schaffer, 1992)。Hasegawa(2002) 認為環境溫度對淹水逆境的影響程度主要因低溫 與植物體休眠時,土壤氧氣消耗量低,故對作物影響力較低,但當環境溫度超過 20℃時,氧氣消耗量大,氧氣消耗主要由植物體根部與土壤生物呼吸所致,故影 響較低溫大。土壤淹水也會造成細菌、真菌、線蟲與疾病之傳播,Crane 和 Davies(1989) 認為對於越橘屬 (Vaccinium) 作物而言,淹水逆境造成植株死亡之主 要原因為疫病 (Phytophthora cinnamomi) 之傳播與抗性,淹水所造成之傷害則為次 要。
2. 淹水對果樹作物水分狀態與光合作用生理之影響
高(1994) 認為乾旱與淹水最終結果皆會造成植物體缺水死亡,不過短期淹水 時植物缺水的程度不如乾旱時嚴重。一般情況下,短期淹水逆境會影響果樹光合 作用率與水分利用率,但並不立即影響植物體水分狀態 (Francisco et al., 2007),但 長期淹水最終仍會影響作物水分狀態,造成失水等生理障害。淹水逆境對植物體 水分生理之影響程度可因作物對淹水之敏感性與嫁接砧木種類等因子而有差異 (Kozlowski, 1984; Schaffer, 1998; Schaffer et al., 1992)。Francisco 等(2007) 以柑橘砧 木品種Carrizo 與 Cleopatra 為試驗材料進行連續 9 天淹水或乾旱處理,利用中午莖 部水勢 (Midday stem water potential; Ψs)、葉片滲透勢 (leaf osmotic potential; Ψπ)、
葉片中午相對水分含量 (Midday relative leaf water content; RWC) 與滲透勢調節 (osmotic adjustment; OA) 等水分生理參數比較兩種砧木於乾旱或淹水逆境之水分 狀態,結果顯示對柑橘幼株Carrizo 與 Celopatra 而言,短期 (9 天) 淹水逆境並不 會影響Carrizo 與 Cleopatra 柑橘幼樹之水分狀態,水分生理參數與對照組相比無顯 著差異。以楊桃 (Averrhoa carambola L.) 植株而言,持續淹水須超過 15 日葉片水 勢(leaf water potential) 才會下降,作物才出現失水症狀 (Ismail and Noor, 1996)。
短期淹水逆境雖不會降低植株中午莖部水勢 (midday stem water potential),但 作物之水分利用率 (water use efficiency; μmol CO2.mmol-1 H2O) 仍受淹水逆境所 影響,主要是因為光合作用受抑制程度較蒸散作用嚴重所致 (Francisco et al., 2007)。
一般而言短期淹水逆境會降低作物的光合作用率 (photosynthetic rate) 與光化 學效率 (PSⅡ efficiency; Fv / Fm) (Childers and White, 1942; Francisco et al., 2007;
Ismail and Noor, 1996; Schaffer, et al., 1992)。作物短期淹水造成光合作用速率降低 主要是因為氣孔關閉所造成;而根部淹水導致氣孔關閉並非因為葉片缺水,而是 因為淹水時根部產生離層酸 (abscisic acid; ABA) 向地上部傳導與細胞分裂素
(cytokinins) 降低所表現之綜合結果所致 (高,1994; Jackson, 1991)。而光化學效率 降低的光化學損害則屬於淹水逆境降低光合作用率的非氣孔因子 (Francisco et al., 2007)。短期淹水影響作物光合作用之原因為氣孔關閉所造成之氣孔性因子,然而 於持續的淹水逆境下,由非氣孔因子抑制光合作用之能力則會逐漸上升 (Beckman et al., 1992)。
彙整前人之研究可知,淹水造成作物整體光合作用率降低之原因有:氣孔導 度降低、葉綠素含量降低、光化學效率降低、新生葉片面積減少、葉片提早老化、
早期落葉及新葉單葉面積減小等 (Childers and White, 1942; Francisco et al., 2007;
Ismail and Noor, 1996; Pezeshki et al., 1996; Save and Serrano, 1986; Schaffer, 1998;
Schaffer, et al., 1992)。
3. 淹水對果樹作物地上部營養生長與生殖生長之影響
許多研究顯示,淹水逆境會影響果樹地上部與根部之營養生長與生化反應,
影響程度依各作物對淹水之敏感度、砧木性狀、樹齡、淹水周期長短、淹水頻率、
季節與環境而有不同 (Hassan et al., 1986; Kozlowski, 1984; Schaffer, 1998; Schaffer et al., 1992)。
對多數作物而言,淹水逆境多會抑制地上部之營養生長,土壤淹水影響果樹 地上部形態之生長與變化一般可歸納出以下幾種特徵:
(1) 植株地上部營養生長受抑制
淹水逆境多會抑制果樹地上部營養生長,其中包括有:株高與枝條生長之抑 制、節間長度縮短、葉片與枝條之乾物重與鮮重降低、樹幹截面積生長受抑制、
葉片生長受抑制……等等 (DeWitt, 1978; Kozlowski, 1982;1984; Schaffer, 1998;
Schaffer et al., 1992)。地上部莖葉生長量下降與激勃素 (gibberellins; GAs) 合成受
抑制有關 (高,1994)。對 B17/B2 楊桃 (Averrhoa carambola L.) 植株而言,淹水超 過5 天即會影響植株之單葉與全樹葉面積生長及地上部與葉片之乾物重 (Ismail and Noor, 1996)。黃色百香果 (Passiflora edulis f. flavicarpa Deg) 實生苗進行淹水 處理4-6 日後,株高與節間長度明顯受抑制,葉片與莖之鮮、乾重皆低於正常灌溉 者;與正常灌溉植株相比,黃色百香果幼株於淹水20 天後之株高為正常灌溉的 84%
(阮及鄭,1986)。高叢藍苺 (Vaccinium corymbosum v. Bluecrop) 植株經淹水 4 個月 處理後植株地上部營養生長量明顯受抑制 (John and Gough, 1987)。淹水處理之奇 異果 (Actinidia deliciosa cv. Hayward) 植株乾物重與淹水日數成反比,而地上部之 落葉率則隨淹水日數增加而上升 (Smith et al., 1990)。
(2) 葉片生長受抑制、葉片老化加速、植株提早落葉
對淹水敏感之果樹而言,淹水逆境會抑制葉片展開、葉片生長量降低、加速 葉綠素分解、提早葉片老化,促使植株提早落葉 (Kozlowski, 1984; Schaffer, 1998;
Schaffer et al., 1992),例如:酪梨 (Ploetz and Shaffer, 1989)、桃、榅桲、梨 (Andersen, 1984) 與美洲胡桃 (Carya illinoinensis) (Wazir et al., 1988)……等等。對楊桃植株而 言,隨淹水日數增加,葉片萎凋率上升,淹水至第25 天時植株全樹落葉 (Ismail and Noor, 1996)。一般認為淹水促進葉片提早老化則與細胞分裂素 (cytokinins) 含量降 低與地上部乙烯含量增加有關 (高,1994)。
(3) 葉柄上偏性生長
當葉片近軸端側的葉柄生長速度快於遠軸端側時,葉片會向下彎曲,即稱做 上偏性 (epinasty)。一般作物於淹水或土壤缺氧之環境下,根部會累積乙烯前驅 物;氨基環丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid ; ACC),之後利用木 質部向地上部傳導ACC,造成葉柄上偏性 (高,1994)。一般而言乙烯 (ethylene) 和 高濃度的生長素 (auxins) 會誘發葉柄之上偏性生長。然而許多研究證明乙烯為葉
柄上偏性生長主要因子,而高濃度生長素造成葉片上偏性生長之主因為生長素會 促進乙烯形成,故生長素僅間接參與葉柄上偏生長之反應 (Kieber, 2006)。
(4) 莖幹基部肥大皮孔增生
對許多較耐淹水之果樹而言,淹水與過度灌溉會誘導植株莖幹基部或靠近水 線上方增生肥大之皮孔 (hypertrophic lenticels),例如:桃、蘋果、梨、榅勃 (quince) (Andersen et al., 1984; Hook and Brown, 1973)、芒果 (Larson et al., 1993)、楊桃 (carambola) (Schaffer et al., 1992) 與高灌木藍苺 (highbush blueberry) (Abbott and Gough, 1987)。芒果植株土壤缺氧至第 4 天即開始增生肥大皮孔,淹水時間越長,
增生之皮孔數越多 (Larson et al., 1993)。皮孔之組成為木栓組織,莖幹基部增生肥 大皮孔可抗耐淹水逆境之主因推測可能因莖部肥大皮孔可讓氧氣擴散至根圈,使 根部能進行呼吸作用所致 (Hook and Scholtens, 1978; Schaffer, 1998; Schaffer et al., 1992)。而芒果植株於水耕環境下添加乙烯與氧氣也可促進肥大皮孔之增生,故推 測淹水處理所增生之肥大皮孔主要由乙烯誘導形成 (Larson et al., 1993)。
(5) 地上部不定根之形成
於長時間淹水處理下,許多果樹之莖基部或水線以上會形成不定根
(adventitious root) 以適應或降低淹水逆境,例如:芒果、蘋果與百香果植株 (阮及 鄭,1986; Hook and Brown, 1973; Schaffer, 1998),不定根結構上具較大之細胞空 隙,於長時間淹水之環境下兼具吸收與傳遞氧氣至淹水根部之功效,以減輕根部 缺氧 (anoxia) 之狀態 (Kozlowski, 1997)。一般而言,淹水誘導不定根形成通常較 容易出現在幼年期之作物,例如:芒果 (Schaffer, 1998)。
(6) 造成落花與落果、抑制生殖生長
一般而言,於土壤浸水與多雨之環境下均會造成植株落花與落果、降低果實
品質,抑制生殖生長 (小林,1985; Schaffer et al., 1992)。兔眼種藍苺‘Woodard’
(Vaccinium ashei Reade ‘Woodard’) 於春季至夏季進行淹水處理後恢復排水,秋季 花苞數僅為對照組的30-70% (Crane and Davies, 1988a)。然而對某些果樹而言,適 度淹水處理會抑制營養生長、促進植株開花,例如:楊桃 (Ismail and Noor, 1996) 與 蓮霧 (Lin, 1992)。
4. 淹水對果樹作物根系吸收水分與代謝之影響
對許多作物而言,過多的水或淹水 (waterlogging) 會影響根系對水分的吸 收、生長與分佈,主要是因為土壤缺氧所造成之缺氧症 (anoxia) (Buwalda, 1993)。
土壤浸水降低根部對水之通透性;增加根對水分吸收的阻力,根部由於缺氧,故 呼吸速率降低,也相對限制根系對養水分吸收之能力 (Kozlowski, 1984)。Andersen 等(1985) 以 2 年生之杜梨 (Pyrus betulaefolia Bunge)、豆梨 (Pyrus calleryana Decne)、西洋梨 (Pyrus communis L. ‘Old Home’ בFarmingdale 97’)、榲桲(Cydonia oblonga Miller ‘Provence BA 29’)、桃 (Prunus persica L. ‘Lovell’) 與褪色柳 (Salix discolor Muhl.) 為試驗材料,進行為期約 30 日有氧 (aerobic) 及無氧 (anaerobic) 之養液栽培處理,發現根系長期缺氧會降低根部呼吸速率與氣孔導度,根系缺氧 時間越長,根部呼吸速率與氣孔導度則越低。蘋果根系經浸水處理後,葉片中氮、
磷、鉀、鈣、鎂等大量元素的含量顯著降低 (Lee et al., 1982)。於根系代謝方面,
因淹水造成根系缺氧,故根系也進行無氧呼吸,造成乙醇及乙醛等有毒物質之累 積,可能導致根系組織之傷害 (Rowe and Beardsell, 1973)。
5. 淹水對果樹作物根系生長之影響
一般而言,植物體根系對淹水特別敏感 (Buwalda, 1993)。淹水對植物體之影 響程度依各作物對淹水之敏感度、樹齡、淹水周期長短、淹水頻率、季節與環境 而有不同 (Kozlowski, 1984; Schaffer, 1998; Schaffer et al., 1992)。
許多研究顯示,淹水或低氧均會減低根系之活力、降低根系乾物重、抑制果 樹新根生長與伸長,加速新根死亡 (turnover) (Buwalda, 1993; Kozlowski, 1984;
Olien, 1987; Schaffer, 1998; Schaffer et al., 1992)。平田(2000b) 認為梨樹根系於土壤 氧氣濃度20%時生長良好,土壤氧氣濃度降低至 10%以下根系生長即受影響。蘋
果於土壤氧氣含量12%以上之環境下可產生龐大偏黃褐色之鬚根系,而土壤氧氣
含量下降至2.5-11%時其鬚根系統減少,根系顏色較深,土壤氧氣濃度下降至 2.5%
以下根系則會大量木栓化且死亡 (Childers and White, 1942)。田間土壤短暫的缺氧 會大量增加奇異果根系的死亡 (Buwalda, 1993),奇異果植株淹水 3 日後新根生長 量即明顯降低,以奇異果植株水耕供氧中斷試驗顯示,根部之生長與伸長明顯受 缺氧抑制 (Smith et al., 1989)。Ismail 和 Noor (1996) 以楊桃幼株為試驗材料,於 9 日之淹水處理下,發現淹水會抑制楊桃幼株根部之生長。與對照組相比,淹水處 理9 日之楊桃全株根部總乾物重約為對照組的 30%。與正常灌溉處理相比,黃色 百香果 (Passiflora edulis f. flavicarpa Deg) 幼株於浸水處理 24 小時後排水,雖處理 後根系外觀未受影響,但排水後根系之生長量即低於對照組 (阮及鄭,1986)。粗 皮檸檬與酸橙植株連續淹水處理21 日根系生長即受抑制 (Syvertsen et al., 1983)。
而比較植物體於淹水逆境下地上部與地下部比例可知,根系所受之影響較地 上部嚴重。淹水處理之奇異果 (Actinidia deliciosa cv. Hayward) 植株乾物重與淹水 日數成反比,而地上部與地下部乾物重比 (shoot : root ratio) 及落葉率則隨淹水日 數增加而上升 (Smith et al., 1990)。
6. 果樹作物適應淹水逆境之策略
許多耐淹水之作物於淹水狀態下,植物體內荷爾蒙相對含量也會發生明顯之 變化,導致植物體出現型態及生理上之變化以適應淹水逆境。一般於淹水逆境下 常見之外觀變化有:通氣組織形成、不定根生長及莖幹基部皮孔腫大等,這些外 觀形態上之變化均為植物適應缺氧的生存策略 (Kawase, 1981; Schaffer, 1998;
Yamamoto et al., 1995)。
根組織內部移動氧氣的能力通常與其多孔性有關。奇異果對於土壤缺氧逆境 極為敏感,主要可能因其根系內部之多孔性相對較其他物種低所造成 (Buwalda, 1993)。一般作物根域於缺氧狀況下多會形成通氣組織 (aerenchyma) 以維持組織之 含氧量;奇異果、蓮霧等作物於淹水後也會形成通氣組織以適應缺氧 (Lin, 1992;
Smith et al., 1990),降低缺氧對作物生長之傷害。一般而言,淹水誘導通氣組織形 成之機制為:淹水時根部誘導乙烯之生成,根部皮層接受乙烯訊息,誘發細胞死 亡,產生多孔性 (Kieber, 2006)。
7. 不同果樹作物對淹水逆境之耐性與敏感度
不同物種、品種之作物與砧木對淹水之敏感性各有差異,以果樹栽培而言,
於排水不良、易淹水之地區通常會篩選耐淹水 (flood tolerance) 砧木以抵抗淹水逆 境。砧木對淹水逆境之耐受性也因環境變化、樹齡與品種而有不同 (Schaffer et al., 1992)。一般而言,作為篩選砧木耐淹水之指標有:土壤浸水期間植株地上部生長 性狀之變化、淹水期間光合作用率與光化學效率變化、土壤浸水期間根系活力變 化與土壤浸水期間根部呼吸速率變化之敏感性等等,一般也多整合以上指標將作 物對淹水之相對耐受性 (relative tolerant) 評分成 8 種等級:極端耐淹水 (extremely tolerant)、非常耐淹水 (very tolerant)、中度耐淹水 (moderately tolerant)、中度敏感 (moderately sensitive) 、敏感 (sensitive)、非常敏感 (very sensitive)、極端敏感 (extremely sensitive) 與最敏感 (most sensitive) (Albrecht et al., 1997; Andersen et al., 1985; Beardsell, 1973; Mahbub et al., 2006; Roberto et al., 1999; Rowe and Hassan et al., 1986; Schaffer et al., 1992)。
(1) 極端耐淹水 (extremely tolerant)
例如:榅勃 (Quince; Cydonia oblonga Mill.) (Rowe and Beardsell, 1973) 與杜梨
(Birch-leaved pear; Pyrus betulaefolia Bunge) (Andersen et al., 1984; Mahbub et al., 2006)。然而榅勃植株因其抗淹水之特性,故常作用為梨樹栽培之抗淹水砧木 (Schaffer et al., 1992)。
(2) 非常耐淹水 (very tolerant)
Rowe 和 Beardsell(1973) 認為大多數梨屬 (Pyrus spp.) 作物均非常耐淹水 (very tolerance)。Andersen 等(1984) 於梨屬作物中細分其對淹水之抗性,分級中屬 於非常耐淹水 (very tolerance) 之梨屬作物依序有:豆梨 (Pyrus calleryana)、榅勃
‘Emla C’ (Pyrus oblonga ‘Emla C’)、榅勃‘Provence BA29’ (Pyrus oblonga ‘Provence BA29’)、傅氏梨 (Pyrus fauriei) 與西洋梨‘Bartlett’(Pyrus communis ‘Bartlett’)。
(3) 中度耐淹水 (moderately tolerant)
Andersen 等(1984) 於梨屬作物中細分其對淹水之抗性,分級中屬於中等耐淹 水 (moderately tolerance) 之梨屬作物依序有:日本豆梨 (Pyrus dimorphophylla Makino)、秋子梨 (Ussurian pear; Pyrus ussuriensis Max.)、砂梨 (Sand pear; Pyrus pyrifolia Nakai)、川梨(Pyrus pashia)與西洋梨‘OH ×F97’(Pyrus communis ‘OH
×F97’)。Schaffer 等(1992) 整合前人之研究認為蘋果砧木 M.1、M.2、M.3、M.6、
M.7、M.13、M.14、M.15、M.16、M.26、‘Crab C sldg’與‘Jonathan sdlg’屬於中度耐 淹水之砧木。Rowe 和 Beardsell(1973) 認為蘋果 (Malus ×domestica L.)、柑橘屬作 物 (Citrus spp.)、歐洲李 (common plum; Prunus domestica L.) 與櫻桃李 (cherry plum; Prunus cerasifera L.) 均屬於中度耐淹水之作物。而藍苺、葡萄、芒果、楊桃 與番石榴也被分級為中度耐淹水之作物 (Crane and Davies, 1989; Schaffer et al., 1992)。而李屬 (Prunus species) 中的郁李(Prunus japonica)、櫻桃李‘S2544-2’
(Prunus cerasifera ‘S2544-2’)、櫻桃李‘GF8-1’ (Prunus cerasifera ‘GF8-1’)、歐洲李
‘Damas GF 1869’ (Prunus domestica ‘Damas GF 1869’)與歐洲李‘Damas de Toulouse’
(Prunus domestica ‘Damas de Toulouse’) 於分級上均屬於中度耐淹水之作物 (Schaffer et al., 1992)。柑橘砧木中的枳殼 (Trifoliate orange; Poncirus trifoliata) 與 粗皮檸檬 (Rough lemon; Citrus jambhiri) 於分級上也屬於中度耐淹水之作物 (Schaffer et al., 1992)。
(4) 中度敏感 (moderately sensitive)
蘋果砧木的M.4、M.9、M.26、M.111、MM.106 品系於分級上屬於中度敏感 之砧木 (Schaffer et al., 1992)。李屬中的歐洲李‘Cirule 43’ (Prunus domestica ‘Cirule 43’)、歐洲李‘Brompton’ (Prunus domestica ‘Brompton’)、歐洲李‘St. Julian A’ (Prunus domestica ‘St. Julian A’)、歐洲李‘GF355-2’ (Prunus domestica ‘GF355-2’)、櫻桃李
‘P34’ (Prunus cerasifera ‘P34’)、櫻桃李‘P855’ (Prunus cerasifera ‘P855’)、櫻桃李
‘P936’ (Prunus cerasifera ‘P936’)、櫻桃李‘P938’ (Prunus cerasifera ‘P938’) 與 Methley plum‘GF31’ (Prunus cerasifera ×P. salicina ‘GF31’) 於分級上屬於中度敏感 之作物 (Schaffer et al., 1992)。柑橘砧木的枳橙‘Carrizo’ (Citrus sinensis ×Poncirus trifoliata ‘Carrizo’)、椪柑 (Citrus reticulata) 與甜橙 (Citrus sinensis) 於分級上屬於 中度敏感之作物 (Schaffer et al., 1992)。而梨屬作物的 Pyrus amydaliformes
(Andersen et al., 1984)、中國李 (Chinese plum; Sour peach; Prunus salicina Lindl.) (Rowe and Beardsell, 1973)、無花果 (Ficus carica L.)、酪梨 (Persea americana Mill.)、枇杷 (Loquat; Eriobotrya japonica) 與番木瓜 (Carica papaya L.) (Schaffer et al., 1992) 於分級上也屬於中度敏感之作物。
(5) 敏感 (sensitive)
蘋果砧木的 M2、M104、MM106、MM109 品系,中國李 (Prunus salicina) 的
‘S37’、‘S300’、‘2540’、‘S2541’品系、酸櫻桃‘Stockton Morello’ (Prunus cerasus
‘Stockton Morello’)、苦橙 (Citrus aurantium) 與萊姆 (Citrus aurantiifolia) 於分級
上均屬於對淹水敏感之砧木 (Schaffer et al., 1992)。
(6) 對淹水非常敏感 (very sensitive; extremely sensitive)
蘋果砧木的 M26、M111、M779、M789、M793、‘Northern Spy sdlg’品系於分 級上屬於對淹水非常敏感之砧木 (Schaffer et al., 1992)。而 Rowe 和 Beardsell(1973) 指出甜櫻桃 (Prunus avium L.)、圓葉櫻桃 (St Lucie cherry; Mahaleb cherry; Prunus mahaleb L.)、桃 (Prunus persica (L.) Batsch)與山桃 (David’s peach; Prunus
davidiana L.) 對淹水非常敏感。而杏桃 (Apricot; Prunus armeniaca L.) (Rowe and Beardsell, 1973; Rowe and Catlin, 1971) 與奇異果植株 (Actinidia deliciosa)
(Buwalda, 1993; Smith et al, 1989; Smith et al., 1990) 也屬於對淹水非常敏感之作 物。
(7) 極端敏感 (extremely sensitive)
李屬中的山桃‘GF308’ (Prunus davidiana ‘GF308’)、桃 (Prunus persica)、梅 (Prunus mune)、綿毛櫻 (Prunus tomentosa) 於李屬作物中屬於對淹水相對極端敏感 之植株 (Schaffer et al., 1992)。
(8) 最敏感 (most sensitive)
扁桃 (Prunus dulcis Mill.) 與杏桃 (Prunus armeniaca) 於李屬作物中屬於對 淹水最敏感之作物 (Schaffer et al., 1992)。
(五) 梨樹根系之生長特性
梨屬 (Pyrus) 作物為喬木類,根系屬軸根系 (tap root system) 之深根性作物 (Atkinson, 1980; Jackson, 2003)。15 年生亞洲梨 (Asian pear) 根系於土壤中之垂直 分佈深度約為150 公分,90%以上之根系多集中於土壤下 60 公分內,根系之水平
分佈半徑為3.5 公尺,90%以上之根系多集中於距主幹 2 公尺內 (平田,2000b)。
40 年生鴨廣梨 (Pyrus ussuriensis Maxim. ‘Yaguang’) 根系之構成以 0-2 公釐粗之鬚 根為主(占 87.3%),根系垂直分佈小於樹高,69.1%的根系集中分佈於 0-30cm 之土 層內,88.3%的根系集中分佈於 0-50 公分深的土層中,根系水平分佈大於樹冠,91%
的根集中分佈於樹冠內,越近主幹的土壤根系密度越高,樹冠外根系分佈很少 (李 等,2007)。西洋梨根系於土壤中之垂直分佈範圍為 0.6-3.5 公尺,根系多集中於土 壤下60 公分內,52%之根系分佈於 0-20 公分近土表處,根系之水平分佈面積範圍 為7-28 平方公尺,62-80%之根系集中於距主幹 1 平方公尺內,90%之根系分佈於 距主幹4 平方公尺之範圍內 (Atkinson, 1980)。梨樹植株根系分佈之深度與範圍依 砧木、物種、品種、土壤性狀、整枝與栽培系統而有不同 (平田,2000b; Atkinson, 1980)。觀察日本地區 9 年生,主枝長 2.9 公尺、棚架式整枝 (trellis training) 之長 十郎 (Pyrus pyrifolia Nakai ‘Chojuro’) 梨樹根系分佈範圍可知,棚架式整枝梨樹之 根系於土壤內之垂直分佈為120 公分,90%以上之根系集中於土壤下 60 公分內,
分佈深度較淺,根系水平分佈半徑達3.5 公尺,分佈密度隨主幹距離而遞減。然而 於開心自然型 (open central leader training) 整枝系統下,根系於土壤內之垂直分佈 達150 公分,90%以上之根系集中於土壤下 90 公分內,根系垂直分佈較深,水平 分佈範圍為2.5 公尺 (平田,2000b)。
(六) 秋根對薔薇科落葉果樹生長之影響
梨樹於溫帶地區的日本,秋根之數量相對少於春根,但秋根之生長與維持對 植株隔年春季早期展葉、徒長枝之生長與隔年果實細胞分裂期營養之供應具重要 之生長意義。在日本溫帶地區,秋根於9-10 月間少量生產 1-2 次,大多數秋季生 長之新生根於10 至 11 月間木栓化,而少數之新生根則維持白色根之狀態越冬,
於隔年3 月轉色木栓化,相較於隔年 5 至 6 月春季長出的新生根壽命較長,一般 春根之壽命約10 至 20 日,更新速率較秋根快 (平田,2000b; 林,1960)。Christina
和Eissenstat(1996) 於美國賓州中部觀察 20 年生‘Red Delicious’/ ‘M.26’蘋果樹 10 月至隔年6 月植株根系之生長可知蘋果根系於初秋 (10 月) 果實採收後具 1 次小量 生長期,超過30%之秋根壽命達 200 日以上,然而春季萌芽期產生的春根壽命較 短,新生根於晚春時大量更新。
當其他大氣環境與土壤條件不為限制因子時,植物體地上部之生長、發育與 根部之生長、延長與老化幾乎與地溫相關,於10-30℃之範圍內,土溫越高,根系 延長範圍越廣,相對死亡 (mortality) 速率也會上升,主要因為溫度升高、促進根 系呼吸速率上升 (溫度係數 Q10上升)、加速碳水化合物與醣分之消耗,造成根系 提早老化更新,一般認為木本植物之根於土溫5-35℃範圍內才能生長,而以 15-30℃
最適宜 (Cox et al., 2000; Marshall and Waring, 1985; Pregitzer et al., 2000; Richards, 1983; Woodham and Alexander, 1966)。然而觀測 1 年生梨樹根系於土溫 10-40℃下
生長可知,根系生長量隨溫度上升而增加,至25℃以上時根系生長量則隨溫度上
升而降低,於40℃時幾乎停止生長 (林,1960)。根系之分佈受土溫影響。田間栽 培之蘋果根群於春季時分佈於近地表處,因該處於春季溫度最高,冬季少數有活 力之根系則繼續深入土中,以避開寒冷的地表 (Rogers, 1939)。梨樹秋季新根之壽 命與溫度有關,於土溫低 (20 ) ℃ 的時候,新生根組織較多汁、壽命較長。於土溫 高 (>30 ) ℃ 的時候,新生根木栓化的速度較快、壽命較短 (平田,2000b; 林,
1960)。
秋根之生長與二十世紀梨黑斑病 (Alternaria kikuchiana Tanaka) 之預防有 關。樹齡10 年以上之二十世紀梨樹勢較弱,幼葉與小果易受黑斑病之影響,若秋 根之生長良好,樹體可充分儲蓄養分,隔年春季展葉與果實發育速率較快,果實 較快脫離幼果期,可減輕黑斑病之威脅 (林,1960)。
對梨樹而言,秋根生長程度、樹體營養狀況及施肥之效果均有關連。果園於 秋季前土壤肥效持續的情況下,秋根大多會伸長,而於沒有施肥的年份,秋根的
成長是有限的。秋季施肥促進秋根生長,也增加隔年越冬之吸收根,可促使植株 持續吸收與儲蓄養分,供給隔年地上部之發育與生長 (林,1960)。
於園藝栽培而言,秋根於秋季禮肥之施用與隔年早期展葉及徒長枝之生長具 相關連之意義。一般於梨樹栽培上認為,禮肥的量越多越好,然而秋季過度施氮 肥會造成地上部延長生長,對於養分之儲存有負面之影響,特別是較弱的樹體,
而秋、冬季結霜也易造成葉片凍傷與枝枯病 (Phomopsis fukushii) 等傷害 (林,
1960)。然而秋季施氮肥過量也易使二十世紀梨出現黑班病 (林,1960)。
(七) 溫帶地區落葉果樹季節性根部與地上部生長之關係
於溫帶地區栽培之落葉果樹,地上部枝條與根之生長期相近或重疊,例如:
西洋梨 (Head, 1968)、二十世紀梨 (林,1960)、幸水梨 (平田,2000b)、長十郎梨 (平田,2000b)、桃 (Cockroft and Olsson, 1972)、蘋果 (橫田,2000)、奇異果 (Buwalda and Hutton, 1988) 與葡萄 (Freeman and Smart, 1976)……等等。
於英國溫帶地區觀察 3 年生西洋梨‘Conference’之枝條與根部季節性生長可 知,植株地上部枝條生長與根部之生長期相近 (Head, 1968)。一般而言,溫帶地區 梨樹枝條之伸長通常於果實快速發育時期停止 (Chalmers et al., 1984),於收穫時期 再重新開始小量生長 (Caspari et al., 1994),冬季休眠,具四季變化。日本鳥取地 區之二十世紀梨 (Pyrus pyrifolia ‘Nijisseiki’),根系生長較地上部早約 1 個月,根
系之生長速率隨地上部生長而增加,至6 月中旬新梢停止伸長、果實快速發育期
時停止,根部於9 月中旬果實採收後再小量生長 (林,1960)。
於紐西蘭北島地區觀察6 年生獼猴桃 (Actinidia deliciosa) 植株根部與枝條之 季節性生長可知,獼猴桃之根部生長較地上部早約10-14 日,地上部於 60 天內快 速生長後即停止,而根部之相對生長速率則隨地上部生長而增加,地上部停止生 長1 個月後根部之生長勢才逐漸下降 (Buwalda and Hutton, 1988)。
於紐約州觀察葡萄根部與枝條之季節性生長可知,根部之生長周期自春
天開始,生長速率隨溫度上升而增加,至仲夏達最大生長速率,之後出現2 次的
生長高峰,分別是:頂芽停止生長與果實採收後兩時期 (Freeman and Smart, 1976)。然而與其他落葉果樹不同的是,葡萄春季根部之生長約於萌芽(budburst)後 10 週開始,其它落葉果樹,例如:梨、蘋果、獼猴桃之發育即由根部或同時打破 休眠 (平田,2000a; 林,1960; Buwalda, 1988; Christina and Eissenstat, 1996; Richards, 1983)。
(八) 亞熱帶地區常綠果樹季節性根部與地上部生長之關係
於亞熱帶地區栽培之常綠果樹,地上部枝條與根部之生長期幾乎錯開,地上 部枝條與根系間呈間歇性生長之模式。
於佛羅里達地區觀察甜橙‘Valencia’枝條與根部之季節性生長可知,植株地上 部枝條與根部之生長期幾乎錯開,地上部與根部呈交錯且間歇性之生長模式 (Bevington and Castle, 1985)。
於佛羅里達南部地區觀察兩種芒果‘Keitt’ 與‘Julie’植株季節性地上部與根部 之生長模式可知,兩種植株之地上部枝條與根部之生長模式完全不同,根部雖呈 間歇性生長之模式,但地上部生長有時與根部之生長重疊或交錯,兩者間呈不規 則之生長模式 (Willis and Marler, 1993)。
(九) 果樹作物根系與地上部生長之關係
根系生長與地上部光合產物之供應相關;根部全依賴葉片製造供應碳水化合 物,而地上部也全依賴根部供應礦物元素與水分,兩者間相互平衡與協調。而地 上部成熟葉 (供源器官; source) 之數量與功率與根系 (儲池;儲耗器官; sink) 之維 持,彼此間相互影響 (Richards, 1983; Buwalda, 1993)。
根系之生長與發育受植株葉果比 (Leaf : Fruit ratio) 之影響。Buwalda (1993) 奇異果植株葉果比與地下部生長調查可知,植株地上部葉果比上升,根系所分配 之養分越多,根生長量也上升。
(十) 果樹作物根系之觀測方法
一般作物根系之生長監測可分成破壞性 (destructiveness) 監測與非破壞性 (non-destructiveness) 監測 2 類。歸納常見的根系生長監測方式有以下 6 種:挖掘 法、剖面窗法、根窗、土柱法、土壤剖面法、微根窗與透地雷達技術 (Atkinson, 1980;
Bai et al.,2005; Bohm, 1979; Brown and Upchurch, 1987; Buwalda, 1993; Caldewll and Virginia, 1991; Hruska, 1999)。
1. 破壞性監測
(1) 挖掘法 (excavation)
挖掘法為過去傳統根系研究主要之技術,主要利用土壤挖掘之方式將作物根 系挖掘出土面,藉以分析研究植物體根系結構於土壤內之分布,主要可觀測果樹 作物整體主要根系 (>5mm diameter) 之結構 (Atkinson, 1980)。但挖掘法因需花費 大量之人力,且田間挖掘常造成鄰近作物根系之損壞 (Atkinson, 1980),對其根系 傷害極大。Dudney (1972) 統計挖掘法約有 22-37%的根系被保留於土壤內或因清 理時耗損而無法被測量。一般而言,挖掘法於每次測量時需將植株之根系挖出,
常造成根系損傷,故通常無法進行連續性之監測,限制根系研究之內容與範圍 (Atkinson, 1980)。
(2) 土柱法 (soil core; Auger method) 與土壤剖面法 (monolith method)
土柱法與土壤剖面法類似,主要差異點為土壤取樣之體積量。土柱法主要於 觀測植株周遭,於不同距離或位置採集抽樣一定體積之土壤,之後將採集之土壤
塊以不同之垂直距離分層採樣觀測土壤內根系之數量、顏色、根長度密度 (root length density; RLD)、根生物產量 (root biomass)、根系大小與重量,屬於破壞性監 測,而土柱法採樣之土壤體積多小於1000 立方公分(<1000cm3),而土壤剖面法獲 得之土壤體積較土柱法多,為1000-5000 立方公分,相對植物體所受之傷害較大 (Bohm, 1979; Buwalda, 1993)。
2. 非破壞性監測
(1) 根窗 (rhizotron) 與剖面窗法 (profile wall method)
根窗法利用根系生長箱自根系生長處附近製作一面透明的窗口,通常以塑膠 板或玻璃為材料,以觀察植株根系之生長。根窗法多應用於容器栽培,利用透明 的根系生長箱,直接觀察介質中植株根系之生長,但因介質容器大小之限制,根 窗法多使用於小型植株根系之觀測 (James et al., 1985; Richardson-Calfee et al., 2010)。而根窗也有應用於田間或森林作物,原理相似,也稱剖面窗法,通常於田 間進行,於預先挖掘於地下之根室觀測根系生長,但花費之成本與人力極高 (Bohm, 1979; USDA, 2007)。
(2) 微根窗 (minirhizotron)
Bates(1937) 認為過去根系生長之研究多以挖掘法 (excavation) 做為根系觀測 之技術,但因挖掘法為破壞性監測 (destructiveness) 且無法對同一根系進行連續性 之監測,故提出非破壞性 (non-destructiveness) 且可於多個時間段內原位重複觀測 根系之方法,即為微根窗 (minirhizotron) 之概念。Brown 和 Upchurch (1987) 對微 根窗之定義為:微根窗系統是由多組分組成的,在土壤中真實的或通過圖像記錄 一般難以接近的植物根系生長狀況。一般微根窗之基本構造是由微根窗管 (minirhizotron tube)、照相機 (camera) 或攝影機 (video camera)、標定手柄
(indexing handle)、控制器 (control box) 與電腦 (computer) 等儀器所組成 (Brown