國立臺灣大學生命科學院生化科技學系 碩士論文
Department of Biochemical Science and Technology College of Life Science
National Taiwan University Master Thesis
嗜甲醇酵母菌 Pichia pastoris 醱酵生產重組 T4 endonuclease V
之研究
The production of recombinant T4 endonuclease V in Pichia pastoris fermentation
林俞均 Yu-Jiun Lin
指導教授:黃慶璨 博士 Advisor: Ching-Tsan Huang, PhD
中華民國 105 年 8 月
August, 2016
謝誌
終於也到了寫謝誌的一天,僅管碩班兩年以來三不五時幻想著謝誌內容,真 正面對螢幕與鍵盤時腦中仍如 Word 文件般一片空白。並非心中不懷感激,只是心 境竟與經典課文如出一轍:要感謝的人太多了,就感謝天吧。在此仍取寥寥數句,
盼能聊表內心無盡的感謝與祝福。
首先感謝我的指導老師黃慶璨老師,這幾年來不厭其煩軟硬兼施地指導,雖 然步伐緩慢而曲折,偶爾原地打轉,但我也在迷霧中逐漸看清人生的下一個目標。
最終位移或許不長,但我相信所有里程都是有所助益的,無論未來又有怎樣的變化,
我會謹記這段旅途帶給我的啟發。同時感謝吳亘承老師,熱心而誠懇地給予大家實 驗上的建議,改善 Lab106 的環境。以及楊啟伸老師,感謝您從大學面試以來就不 斷地給予我鼓勵,我始終銘記在心,難以忘懷。並且感謝李昆達老師,在答辯過程 中給予許多建議,在論文寫作上也指出了許多不足之處。
再來一一感謝 Lab106 的大家,Lab106 是碩班兩年下來待得最久的地方,就 連硬體我都想感謝一番(器材儀器需要敬重,醱酵槽需要乖乖),更不用說所有前人 今人和後人了,不但在實驗方面給予我各種建議,在日常生活方面也惠我良多。感 謝黃莉欣學姊哩哩,我不會忘記妳的名字,包紅包也絕不會手軟所以務必寄給我。
感謝昱伶學姊,一直以來處理實驗室大大小小的事情,真是辛苦了,祝妳接下來的 計畫都能順利進行。感謝巧青,相信妳到哪裡都能闖出一片天,有機會再去日本找 妳玩。感謝佳葳,一直很欣賞妳樂觀直爽的性格,我想我們還有不少機會碰面的。
感謝芝榕,好佳在我們兩個可以互相確認畢業相關事務,一起畢業也比較不孤單,
祝妳復健順利。感謝下一屆的蔡孟羲張景翔洪凱琳哲銘,希望你們都能早日口試完 畢,往下一個人生目標邁進。也感謝新進的學弟妹們,期待你們讓 Lab106 越來越 好。尤其鄭重感謝所有曾經幫我搬醱酵槽的勇士們,要是沒有你們,不是客套話,
我在物理上是絕對無法順利畢業的。
最後感謝我的家人,雖然兩年下來回家的次數不多,即使偶而回去也是沾沾 醬油,但台北到台南客運路程的那五個小時,或許是六年來最令我平靜的時光。
由衷感謝大家!
俞均 謹誌 105 年 8 月
摘要
T4 endonuclease V (T4N5)是 T4 bacteriophage 中的 DNA 修復酵素,為大小 16 kDa 的蛋白質,可專一性辨認因紫外線造成 DNA 損傷而形成的環丁烷嘧啶二聚體 (cyclobutane pyrimidine dimer, CPD)並切除,再由後續 DNA 修復機制進行修補。過 去研究利用大腸桿菌生產重組 T4N5,結合微脂體(Liposome)技術製成乳劑,針對 缺乏核苷酸切除修復(nucleotide excision repair)能力的患者進行臨床實驗,發現可 減少病灶生成,顯示 T4N5 移除紫外線造成的 DNA 損傷功能對人體亦有所幫助。
為使此技術能廣泛應用於防曬乳及保養品中,提升產品附加價值,須尋求低成本大 量生產重組 T4N5 的方法。現行的重組 T4N5 多以大腸桿菌進行生產,然而欲取得 目標蛋白質須經破菌及多道純化手續,且有內毒素的疑慮。嗜甲醇酵母菌 Pichia pastoris 為單細胞真菌表達系統,可進行高密度培養,生產基因重組蛋白質成本相
較低廉,同時沒有內毒素疑慮並可利用外泌訊息胜肽將目標蛋白質分泌至培養基 中以方便純化。本研究利用嗜甲醇酵母菌 P. pastoris KM71H 作為重組 T4N5 表達 系統,以搖瓶和醱酵槽進行培養,添加甲醇作為碳源並誘導異源蛋白質表現。搖瓶 培養可成功外泌表現出序列正確且具活性的 T4N5,活性可達 0.47±0.05 U/µL。但 以醱酵槽進行高細胞密度培養時,發現以 2.6 mL/L/h 的速率添加甲醇可能會對菌 體造成壓力使部分菌體死亡並裂解釋放胞內蛋白酶,但減低甲醇添加量又不利於 菌體生產異源蛋白質,產量皆不理想,須以其他方式減少菌體死亡裂解及抑制蛋白 酶作用。但當同樣以約 2.6 mL/L/h 的速率添加甲醇,將誘導溫度由 30oC 降低至 23oC,可減低培養基中的蛋白酶含量,並增進甲醇異化代謝路徑,大幅提升目標蛋 白質表現量。
關鍵字:重組 T4 endonuclease V、Pichia pastoris、醱酵培養、甲醇誘導、降溫培 養
II
Abstract
T4 endonuclease V (T4N5) is a DNA repair enzyme isolated from T4 bacteriophage.
It can specifically recognize and remove cyclobutane pyrimidine dimer which is resulted from the UV-induced DNA damage and might cause skin cancer. In previous reports, in conjunction of recombinant T4N5 expressed by Escherichia coli and liposome, T4N5 liposome lotion effectively penetrated into skin in vivo and clinical trials also indicated its potential for UV-induced DNA damage treatment. In order to extensive application of T4N5 in skin care products such as sunscreen, it’s important to develop a method for large-scale production recombinant T4N5 with low expenses. The major recombinant T4N5 are currently produced by E. coli. However, the procedures include cell lysis and protein purification, which may increase the production cost. Moreover, it is at risk due to endotoxins. Pichia pastoris is a well-defined expression system which can be cultivated at high cell density at low cost. In this study, P. pastoris KM71H was used as a heterologous expression system, and methanol was added to induce recombinant T4N5 expression. In flask, we have successfully expressed extracellular T4N5 with correct conformation and function. The T4N5 activity in supernatant of flask culture reached up to 0.47±0.05 U/µL. When using 5 L fermenter, methanol fed at 2.6 mL//L/h induced cell lysis and release of cellular proteases, while reducing methanol feeding rate led to low yield of target protein yield. Therefore, it’s necessary to modify other induction conditions.
In this study, we found that reduction of the induction temperature from 30oC to 23oC improved recombinant protein yield remarkably at the same methanol feeding rate. The amount of proteases in the culture decreased, and methanol dissimilation is improved.
Keywords: Recombinant T4 endonuclease V, Pichia pastoris, Fermentation, Methanol induction, Low-temperature cultivation
目錄
摘要 ... I Abstract ... II 圖目錄 ... V 表目錄 ... VI
第一章、 前言 ... 1
一、 T4 endonuclease V ... 1
1. 結構及作用機制 ... 1
2. 應用價值 ... 2
3. 生產現況 ... 3
二、微生物異源表達系統 ... 4
1. 原核生物表達系統 ... 4
2. 真核生物表達系統 ... 4
三、Pichia pastoris 表達系統 ... 7
1. 酒精氧化酶及其啟動子 ... 7
2. 菌株與甲醇表現型 ... 8
3. 訊息胜肽 ... 8
4. 大規模表現 ... 9
四、研究動機與目的 ... 14
第二章、 材料與方法 ... 17
一、 實驗菌株與培養條件 ... 17
1. Escherichia coli ... 17
2. Pichia pastoris ... 17
二、培養基與藥品 ... 17
三、Pichia pastoris 表現載體建構 ... 20
四、Pichia pastoris 轉形與篩選 ... 23
1. 轉形 DNA 製備 ... 23
2. 勝任細胞製備 ... 23
3. 電穿孔轉形 ... 23
4. Pichia pastoris 轉形株篩選 ... 24
五、Pichia pastoris 誘導表現 T4 endonuclease V ... 25
IV
1. 搖瓶培養 ... 25
2. 醱酵槽培養 ... 25
六、蛋白質產物分析 ... 27
1. 西方墨點法 ... 27
2. LC-MS/MS ... 27
七、T4 endonuclease V 活性測定 ... 30
八、培養基胞外上清液蛋白酶活性分析 ... 31
第三章、 結果 ... 32
一、 轉形株篩選與確認 ... 32
1. 抗性濃度梯度培養基篩選轉形株 ... 32
2. 染色體 PCR 確認目標基因插入 ... 32
二、 搖瓶培養結果 ... 35
1. 以 SDS-PAGE 篩選表現量高轉形株 ... 35
2. 以西方墨點法確認目標蛋白質順利表現 ... 35
3. LC-MS/MS 確認目標蛋白質 ... 35
4. 搖瓶上清液 T4N5 活性測定 ... 35
三、 醱酵槽培養結果 ... 40
1. 以 BSM 培養 ... 40
2. 以 BMGY 置換到 BMMY 培養 ... 40
3. 蛋白酶活性分析 ... 41
第四章、 討論 ... 51
一、 培養基成分之探討 ... 51
1. BSM (Fermentation Basal Salts medium) ... 51
2. BMGY 培養,誘導前置換為 BMMY ... 52
二、 甲醇添加速率之探討 ... 52
1. 鹼添加量 ... 52
2. OD600及生菌數 ... 53
3. 胞外上清液蛋白酶活性 ... 53
4. SDS-PAGE 分析及胞外上清液總可溶蛋白質 ... 53
三、 其他培養條件之探討 ... 54
第五章、 結論 ... 56
第六章、 未來展望 ... 57
第七章、 參考文獻 ... 58
圖目錄
圖 一、T4N5 反應機制 ... 11
圖 二、T4N5 結構 ... 12
圖 三、嗜甲醇酵母菌之甲醇利用途徑。 ... 13
圖 四、本篇論文架構圖 ... 16
圖 五、本研究所使用表現載體圖譜 ... 21
圖 六、以不同 Zeocin 抗性濃度篩選轉形株 ... 33
圖 七、轉形株染色體 DNA 之聚合酶連鎖反應 ... 34
圖 八、搖瓶上清液 SDS-PAGE 與西方墨點分析-I ... 36
圖 九、搖瓶上清液 SDS-PAGE 與西方墨點分析-II ... 37
圖 十、以 LC-MS/MS 進行蛋白質鑑定,紅色字體為 T4N5 序列中辨認到的胺基 酸序列。 ... 38
圖 十一、T4N5 活性測定 DNA 電泳膠圖 ... 39
圖 十二、轉形株 His11 以 BSM 進行甲醇誘導之醱酵槽參數圖- ... 42
圖 十三、BSM 培養基醱酵上清液 SDS-PAGE 分析 ... 43
圖 十四、轉形株 His11 以 BMMY 進行甲醇誘導之醱酵參數圖 ... 44
圖 十五、轉形株 His11 以 BMMY 進行甲醇誘導之生菌數及吸光值 ... 45
圖 十六、BMMY 培養基醱酵上清液 SDS-PAGE 及西方墨點法分析 ... 46
圖 十七、不同甲醇添加速率醱酵上清液 SDS-PAGE 分析 ... 47
圖 十八、降溫誘導醱酵參數圖 ... 48
圖 十九、降溫誘導醱酵上清液 SDS-PAGE 分析 ... 49
圖 二十、醱酵槽上清液蛋白酶活性分析 ... 50
VI
表目錄
表 一、培養基組成 ... 18 表 二、PCR 使用引子 ... 22 表 三、西方墨點法分析所用試劑之組合成分 ... 28
第一章、 前言
一、 T4 endonuclease V
T4 endonuclease V (T4N5, EC 3.1.25.1)在 1975 年由 Tanaka 等人自受 bacteriophage T4 感染的大腸桿菌中分離出來[1],為 bacteriophage T4 denV gene 所表現的蛋白質,與修復紫外線造成的 DNA 損傷有關。紫外線會引發相鄰胸 腺嘧啶(Thymine)以共價鍵鍵結,產生環丁烷嘧啶二聚體(cyclobutane pyrimidine dimer, CPD)。CPD 會干擾 DNA 的複製,造成基因突變,而 T4N5 的功能便是 藉由靜電力差異專一性辨認出 CPD[2],並分兩個反應步驟將其切開,再由 DNA 光解酶或其他修復機制進行後續修復。
1. 結構及作用機制
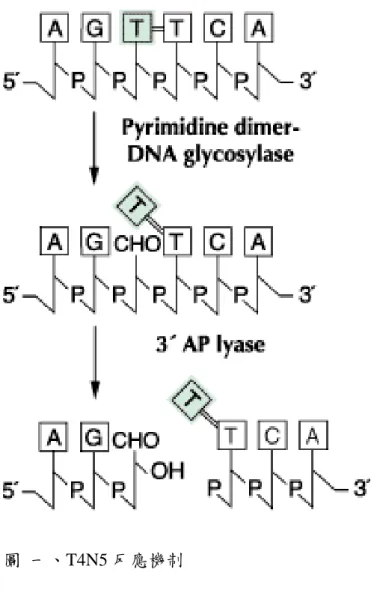
T4N5 序列全長 138 個胺基酸,大小約 16 kDa,由三個 a-helices 及連接 的 loops 所組成,其中 N 端穿過 H1 與 H3 helices 中間,並且在酵素作用上扮 演關鍵角色。T4N5 結構如圖一。T4N5 具有兩種催化活性,在雙股 DNA 上辨 認到 CPD 後會分兩步驟將其切開,首先會將 CPD 5’端鹼基和五碳糖間的 N- glycosyl bond 水解[3, 4],再藉由β-elimination 將脫鹼基位點(abasic site)五碳糖 間的磷酸二酯鍵切開[4]。反應簡圖如圖二。
其中以第一步的 DNA glycosylase 為速率決定步驟,藉由點突變實驗,發 現 Arg3、Glu23、Arg22、Arg26 為主要反應中心[5],會穩定 N 端的氨基使利 用 nucleopholic attack 水解 N-glycosyl bond,點突變上述反應中心的胺基酸或 N 端的 Threonine 皆會對活性造成影響[6]。
2
2. 應用價值
2.1 紫外線與皮膚癌
日光中的紫外線會造成相鄰的胸腺嘧啶鍵結,其中又分兩種鍵結形式,環 丁烷嘧啶二聚體(CPD)及 6-4 光產物((6 – 4) photoproducts)。根據估計人的皮膚 每天平均會產生一百萬次 DNA 突變,其中 CPD 產生的機率是 6-4 光產物的三 倍[7],且在哺乳類動物細胞中的移除速率遠比 6-4 光產物慢[8]。若突變點發生 在致癌基因(oncegenes)或 p53 及其他腫瘤抑制基因(tumor suppressor genes),便 有造成皮膚癌的風險。並且根據動物實驗,過度表現 CPD 光解酶的小鼠,相較 於過度表現 6-4 光產物的小鼠,對於日光誘發的腫瘤生成更具抵抗力,顯示 CPD 形式的突變是造成皮膚癌的主要成因[9]。
2.2 核 苷 酸 切 除 修 復 (Nucleotide excision repair) 與 著 色 性 乾 皮 症(хeroderma pigmentosum)
核甘酸切除修復(NER)為生物體修復紫外線造成的 DNA 損傷主要機制 [10],在辨識到 CPD 或 6-4 光產物後,會移除含突變嘧啶二聚體的一段 DNA,
留下另一股對應的 DNA 並由 DNA 聚合酶補回對應的序列,最後再由 DNA ligase 連接完成修復。在哺乳類細胞中,主要有九種蛋白質參與 NER 機制,這 些蛋白質的缺陷都會造成著色性乾皮症(хeroderma pigmentosum)[11]。
著色性乾皮症為一種罕見的遺傳性疾病,患者皮膚細胞被陽光中的紫外線破壞 後無法自行修復,對紫外線特別敏感,容易曬傷,產生黑斑,皮膚過早老化,
並在兒童期便誘發癌病變,患者平均壽命不到 20 歲[12]。
2.3 T4N5 liposome
由於 CPD 是造成皮膚癌的主要成因,又 T4N5 具有專一性辨識並切除 CPD 的能力,且此機制在人類細胞中依舊可行[13],在 1980 年代,Yarosh 等人便以 微脂體(liposome)包覆重組 T4N5 作為載體,使其得以送入細胞中,增強細胞對 紫外線的抗性並減少細胞死亡[14]。在動物實驗中,小鼠接受 UV 照射後,塗 抹 T4N5 微脂體乳劑可降低皮膚癌的發病率[15]。並且,在針對著色性乾皮症 患者的臨床試驗上,T4N5 微脂體乳劑亦可有效降低光化性角化病(actinic keratosis),一種上皮性癌前皮膚病變的病灶生成[16]。
在 2007 年,Yarosh 參與的 AGI Dermatics.將 T4N5 微脂體乳劑命名 為”Dimericine”,作為治療著色性乾皮症的藥物,向美國 FDA 取得了新藥快速 審查資格,然而目前仍未通過。
3. 生產現況
根據 Yarosh 於 1994 年透過 AGI Dermatics.申請的專利中(專利號 5,296,231),
利用 E. coli strain SR1268 表現重組 T4N5,兩升的大腸桿菌醱酵液經過破菌、
硫酸銨沉澱、單股 DNA 親和層析純化,最終可得到 15 mg 的純蛋白,濃度為 0.2 mg/mL。
4
二、微生物異源表達系統
異源表達(Heterologous expression)是利用特定宿主本身的蛋白質表現系統,
結合基因工程技術,將一段外源基因轉入宿主細胞中,藉此生產出重組蛋白質 (recombinant protein)。異源表達系統可降低成本,簡化操作程序,減少生產所需 時間等優點,現已廣泛應用於農業、學術、工業、醫療等各種領域上[17]。
其中微生物的異源表達系統具有生長快速,操作簡易,培養成本相對低廉 等優點。微生物異源表達系統可分為原核生物和真核生物,而真核生物表達系 統又可分為單細胞真菌如酵母菌及多細胞真菌如絲狀真菌等等。
1. 原核生物表達系統
大腸桿菌 (Escherichia coli)是最早的異源表達系統[18],細胞繁殖速度快,
操作簡便,成本低廉,且基因及生理特性研究透徹,大量生產技術發展成熟。
然而原核生物缺乏後轉譯修飾(post- translational modification)機制,無法進行 醣基化作用 (glycosylation) 與多種轉譯後修飾能力,不適合用於生產部分真核 生物蛋白質。且大腸桿菌外膜的脂多醣(lipopolysaccharides, LPS),又稱內毒素 (endotoxin),會在人體內引起強烈的發炎反應,甚至造成敗血症[19],由於不易 純化移除,不適合生產醫療用蛋白質。
2. 真核生物表達系統
對於需要後轉譯修飾來獲得生理功能的真核生物蛋白質而言,真核生物為 較合適的表達系統。常見的真核微生物異源表達系統又可分為單細胞酵母菌及 多細胞絲狀真菌。
2.1 絲狀真菌
絲狀真菌俗稱黴菌,具有生長快速,可分泌至胞外、具後轉譯修飾及生產 成本低等優點。常見的絲狀真菌表現系統有木黴菌屬 (Trichoderma) 和黑黴菌 屬(Aspergillus) 的真菌[20],應用於生產抗體、藥物、以及食品加工酵素等。但 相較於其他異源表達系統,絲狀真菌的異源蛋白質表現量低,在大量生產的應 用上仍有所限制[21]。
2.2 酵母菌
2.2.1 釀酒酵母菌
釀酒酵母菌 Saccharomyces cerevisiae 為單細胞的真核生物,和原核細菌 E.
coli 同樣具有容易培養和生長快速等優點,由於長久以來便應用於製造麵包和
酒類醱酵,因此被認為是 GRAS (generally regarded as safe) 的菌種。然而重組 蛋白質有過度醣基化的現象 [22],會影響重組蛋白質的活性與功能,且可能引 起過敏。此外,其重組蛋白質的產量較低,產量不易達到工業生產所需的規模。
2.2.2 嗜甲醇酵母菌
嗜甲醇酵母菌為一類可利用甲醇作為碳源和能量來源的酵母菌,這類酵母 菌皆具特別的甲醇利用途徑,會在特化的過氧化體 (peroxisome) 中進行初步 的甲醇代謝,接著產物進入細胞質進行下游的代謝反應(圖三)。甲醇進入過氧 化體後先由酒精氧化酶 (alcohol oxidase, AOX) 氧化產生甲醛和過氧化氫,過 氧化氫再由過氧化體中的過氧化氫酶 (catalase) 分解為水及氧分子。而甲醇代 謝所生成的甲醛則有兩種不同的代謝途徑:經由異化作用 (dissimilation) 來產 生能量,或者經由同化作用 (assimilation) 增加生物質量 (biomass)[23]。嗜甲 醇酵母菌可在高細胞密度進行培養,且當甲醇作為唯一碳源存在時,胞內會大 量表現酒精氧化酶,因此具有強力的啟動子。而當其他碳源存在時,則會抑制
6
啟動子的表現,為嚴謹的調控。且相較於釀酒酵母菌,嗜甲醇酵母菌所生產的 異源蛋白質的醣基化程度較低[24, 25]。藉由這些特性,可將嗜甲醇酵母菌進行 高密度培養,並藉由甲醇誘導調控,作為一套利用碳源控制大量表現目標蛋白 質的系統。
三、Pichia pastoris 表達系統
Pichia pastoris 為目前研究最為透徹的嗜甲醇酵母菌,具有生長快速、成本
低廉、表現量高等優點。1960 年代,P. pastoris 被用來生產作為牲畜飼料的單 細胞蛋白 (single-cell proteins, SCP)[26]。1980 年代Phillips Petroleum Company 和 Salk Institute Biotechnology/Industrial Associates, Inc. (SIBIA, La Jolla, CA,
USA)合作開發了P. pastoris 表現異源蛋白質的系統,包含了表現載體、菌種、
以及使用流程等[27],自此之後超過 1000 種以上的蛋白質使用此系統進行表 現[28]。
2006 年,以 P. pastoris 生產的重組磷脂酶 C,被 FDA 認可為 GRAS,用於 食用油的處理;並且,在 2009 年,以 P. pastoris 生產的生物製藥血管舒缓素 抑制劑” Kalbitor” (Dyax Corp.),亦得到了 FDA 的認可,顯示 P. pastoris 在未 來生物製藥表現重組蛋白質藥物的領域中,具有極大的潛力[29]。
1. 酒精氧化酶及其啟動子
酒精氧化酶(AOX, EC 1.1.3.13)為嗜甲醇酵母菌在甲醇代謝途徑中的第一 個酵素,在 P. pastoris 中有兩種酒精氧化酶參與甲醇代謝,AOX1 及 AOX2,
兩者的調控模式相同,而 AOX2 的 mRNA 表現量遠小於 AOX1,以 AOX1 為 主要參與甲醇代謝之酵素。當甲醇為唯一碳源時,由於酒精氧化酶代謝速率較 慢,需要大量表現酒精氧化酶以分解甲醇。在細胞高密度培養下,被誘導表現 的 AOX1 酵素可超過全部可溶蛋白質的 30%[26],所轉錄的 RNA 可達胞內 mRNA 約 5%之多。
除了酒精氧化酶啟動子,亦有其他參與代謝酵素的啟動子被分離出來,並 用以表現重組蛋白質,如其它甲醇代謝相關的啟動子 FLD1 (formaldehyde),醣 類代謝相關啟動子如 GAP (glyceraldehydes 3-phosphate dehydrogenase)[30-32]等,
使得利用 P. pastoris 表現異源蛋白質有更多的設計選擇。
8
2. 菌株與甲醇表現型
P. pastoris 可以 AOX 的基因修改後的差異分成以下三種不同的表現型: I,
Mut+ (methanol utilaiztion plus),兩種 AOX 基因皆保留功能,如野生型的 P.
pastoris、GS115 菌株等;II, MutS (methanol utilization slow),AOX1 基因失去功 能,只留有 AOX2 基因,故甲醇代謝速率較慢,如 KM71、KM71H 菌株等;
III, Mut- (methanol utilization minus),兩個 AOX 基因皆失去功能,此類菌株無 法生長在以甲醇為單一碳源的環境[33]。
另外, AOX 啟動子受其他碳源嚴格調控,當環境存在一定濃度的葡萄糖 或甘油時,Mut+和 Muts 菌株的甲醇代謝反應會受到抑制,只有在缺乏這類碳 源,並同時有甲醇存在時,方可啟動 AOX 啟動子使異源蛋白質大量表現,在 工業應用上有其限制。但目前已開發出其他改良的菌株,在低濃度的葡萄糖或 甘油條件下,也可經去抑制化啟動 AOX1 啟動子表現下游蛋白質[34]。
儘管 Mut+菌株甲醇代謝速率快,利用率高,以甲醇為唯一碳源生產異源 蛋白質時產量理應較高[24],但相較於 MutS菌株,其需氧量高,產熱快[35],
且甲醇代謝時會形成過氧化氫,過多的過氧化氫會對細胞的生長造成壓力甚至 導致菌體死亡,菌體破裂釋出的液胞內蛋白酶又會對培養基中的目標蛋白質造 成影響,因此 Mut+菌株較不利於進行大規模醱酵[36]。而 MutS甲醇代謝較慢,
易導致生長速率慢及誘導時間過長,但可藉由甘油與甲醇混合誘導的方式解決 此問題。
3. 訊息胜肽
P. pastoris 原生性外泌至胞外的蛋白質較少[33],因此若能將目標異源蛋白
質分泌到胞外,可大幅減少純化的步驟及成本。利用在外源基因前面加上一段 外泌訊息胜肽,可將異源蛋白質分泌到胞外。目前常使用的訊息胜肽是來自 S.
cerevisiae α 交配型的訊息胜肽 α-MF-prepro-sequence peptide (S. cerevisia 有兩種
交配型,a 和α),在 S. cerevisiae 行有性生殖時可將 α 交配型的交配信息素 (α- factor) 外泌至細胞外[37],其長度為 85 個胺基酸,帶有三個醣基化位點與兩個 KEX2 及一個 STE13 蛋白質內切酶切位,可藉由修改切位附近的胺基酸增加其 剪切效率[38]。
4. 大規模表現
P. pastoris 的大規模培養可以全合成培養基進行,添加葡萄糖、甘油等作為
碳源提高生物質量以利後續誘導期間異源蛋白質表現,又以甘油較為適合,避 免葡萄糖醱酵副產物乙醇對 AOX 啟動子造成抑制[39]。甲醇誘導期對於 P.
pastoris 生產異源蛋白質的表現量十分關鍵。由於甲醇為毒性物質,添加太多會
造成菌體的毒害[40],而添加太少則無法有效誘導 AOX 啟動子[33]。有學者利 用菌體代謝甲醇時,溶氧值(Dissolve oxygen, DO)的增減作為指標來監控甲醇的 添加量而發展出 DO spike 法[41],但此法不適用於 Muts菌株[42],亦有研究利 用甲醇監控儀偵測槽體內甲醇的濃度,藉此控制甲醇濃度維持恆定,常用於 Muts菌株培養[43]。
在高密度培養甲醇誘導期間,外泌的異源蛋白質可能會被培養基中的蛋白 酶降解,P. pastoris 本身外泌的蛋白酶較少[33],且研究指出培養基中的蛋白酶 主要是來自於菌體破裂釋出的液胞內蛋白酶,並且在甲醇誘導期間方可觀察到 [44]。菌體的壓力來源有二:甲醇代謝的第一步會產生甲醛及過氧化氫,兩者皆 對菌體有害,過氧化氣又會造成氧化壓力;同時目標蛋白質的大量表現可能引 發 Unfolded Protein Response (UPR),引發細胞凋亡[45]。
蛋白質降解現象會減低目標蛋白質產量,降低產物活性,並且不利於純化,
以下策略常用來減緩蛋白質降解現象。
10
4.1 培養條件
Pichia pastoris 可在 pH 3.0-7.0 的範圍生長,因此可針對不同異源蛋白質,
調整至最能降低蛋白酶活性並穩定異源蛋白質的 pH 值進行誘導培養[46-48]。
在培養基中添加額外的含胺基酸氮源,如 peptone 及 casamino acid 等,可 作為代替異源蛋白質的基質讓蛋白酶降解,並可補充額外氮源抑制外泌蛋白酶 表現[48, 49]。
亦有研究指出降溫誘導可增進蛋白質產量[50, 51],除了降低蛋白酶活性,
亦可減低細胞死亡及蛋白酶釋出,同時能提升 AOX 活性進而提高異源蛋白質 產量[52, 53]。
4.2 表現菌株
Pichia pastoris 有蛋白酶缺陷株如 SMD1163 (his4 pep4 prb1),SMD1165
(his4 pep4),SMD1168 (his4 pep4)可利用來進行異源蛋白質生產。PEP4 為 Proteinase A 的基因,PRB1 為 Proteinase B 的基因。Proteinase A 為液胞中的 aspartyl protease,可活化其他液胞蛋白酶如 carboxypeptidase Y 及 proteinase B 等。但蛋白酶缺陷株相較非缺陷株,有生長速率較慢,轉形較不易等問題[33]。
圖 一、T4N5 反應機制
Figure 1. The mechanism of T4N5
12
圖 二、T4N5 結構[54]
Figure 2. The structure of T4N5
圖 三、嗜甲醇酵母菌之甲醇利用途徑。[23]
Figure 3. Methanol utilization pathway in methylotrophic yeasts
14
四、研究動機與目的
過去的研究中證實,T4N5 結合微脂體技術後,可製成能被人體皮膚吸收 並進入皮膚細胞中幫助移除 CPD 的乳劑,有希望作為著色性乾皮症患者的藥 劑[16]。由於著色性乾皮症是僅 25 萬分之一的罕見遺傳性疾病[55],T4N5 微 脂體乳劑作為專門藥劑侷限性較大,但仍具有其他方面的應用潛力,例如,根 據近期研究統計,接受器官移植者,罹患黑色素瘤(melanoma)的機率是未接受 過器官移植者的兩倍[56]。儘管原因仍待詳細研究,對於器官移植患者,除了 避免日曬,或許也能使用 T4N5 微脂體乳劑減低致癌風險。並且,若能將 T4N5 微脂體乳劑應用於防曬乳或保養品上,製成兼具防曬美白且幫助修復 DNA 損 傷的產品,亦可大幅提升產品價值。
為達成商業應用的目標,工業化的大量生產有其必要性。相較於現今應用 於生產重組 T4N5 的大腸桿菌表達系統,P. pastoris 不具內涵體和內毒素疑慮,
易於外泌等優勢,且同樣可進行成本相對較低的高細胞密度培養。
因此,本研究利用 P. pastoris 為異源表達系統,以搖瓶和 5 L 醱酵槽進行 甲醇誘導,希望能大量生產出重組 T4N5,以作為工業化生產的第一步。
而在將生產規模由搖瓶放大到醱酵槽時,發現高密度培養生產效果不如 預期,因此調整甲醇添加速率及溫度等誘導條件,並從醱酵參數、生菌數、培 養液中蛋白酶活性等數據分析,探討造成產量不佳的原因並設法調整醱酵條件 以增加產量。
本篇論文具體目標
1. 建構以 P. pastoris 表現重組 T4N5 的表現載體 2. 確認重組 T4N5 的蛋白質基本性質
3. 以 P. pastoris 大量生產可外泌的重組 T4N5 4. 分析重組 T4N5 的產量、活性
5. 從醱酵參數及蛋白酶活性等探討適合生產重組 T4N5 的條件
16
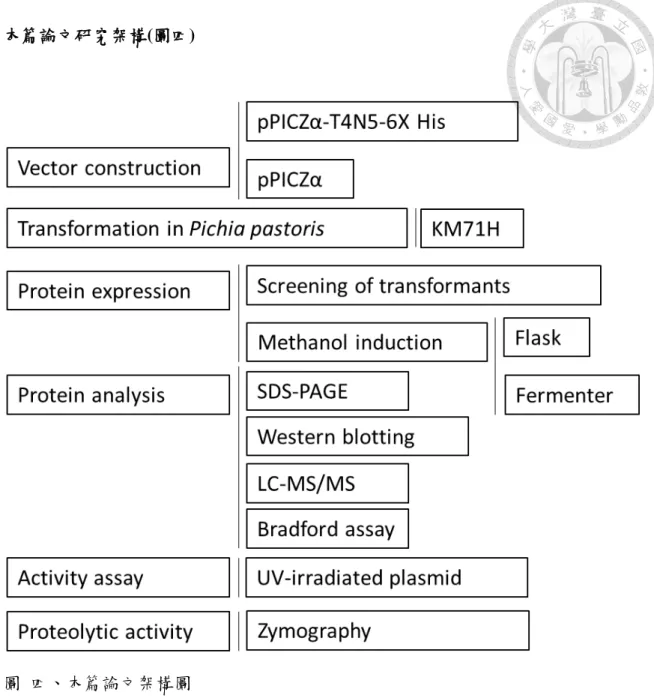
本篇論文研究架構(圖四)
圖 四、本篇論文架構圖
Figure 7. Schematic framework of this study
第二章、 材料與方法
一、 實驗菌株與培養條件
1. Escherichia coli
大腸桿菌 Escherichia coli EPI300 購自 Epicentre 公司(Madison, WI, USA),
用於表現載體建構與質體基因保存。以 Luria-Bertani (LB; Alpha Biosciences, USA) 液態培養基於 37oC、轉速 250 rpm 振盪培養,或添加 1.5 % 洋菜膠製 成固態培養基於 37oC 培養。
2. Pichia pastoris
本研究所選用的異源蛋白質表達宿主為嗜甲醇酵母菌 P. pastoris KM71H (MutS, his4, AOX1::ARG4, arg4),購自 Invitrogen 公司(Carlsbad, CA, USA)。以 YPD 液態培養基於 30oC、轉速 250 rpm 振盪培養,或添加 1.5 % 洋菜膠製成 固態培養基於 30oC 培養。
二、培養基與藥品
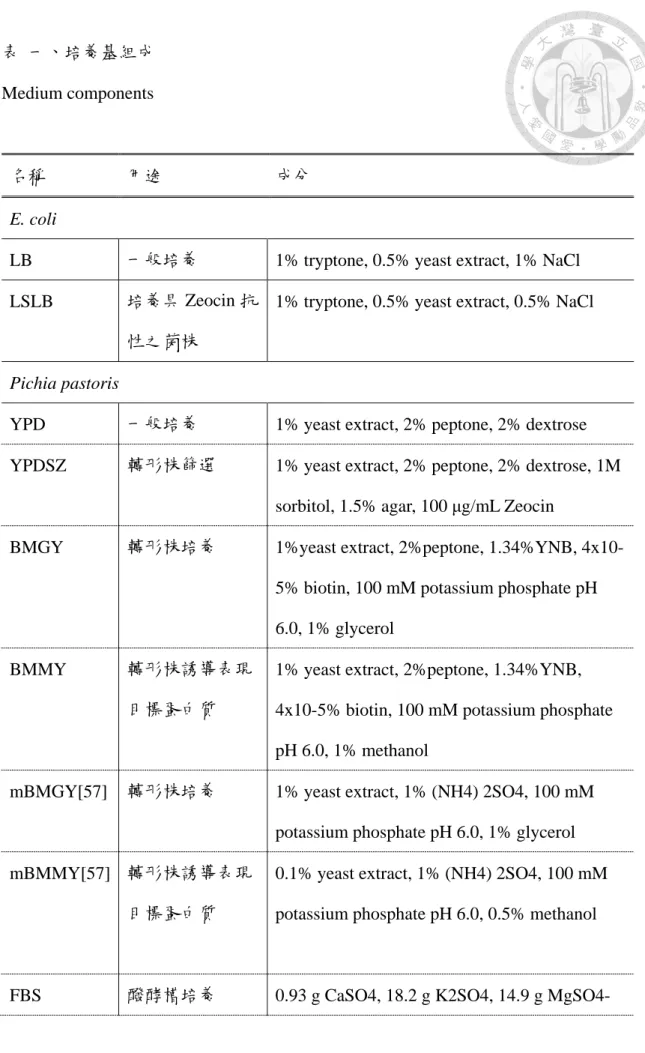
本研究所使用的培養基及其成分用途如表一所示。
18
表 一、培養基組成 Medium components
名稱 用途 成分
E. coli
LB 一般培養 1% tryptone, 0.5% yeast extract, 1% NaCl LSLB 培養具 Zeocin 抗
性之菌株
1% tryptone, 0.5% yeast extract, 0.5% NaCl
Pichia pastoris
YPD 一般培養 1% yeast extract, 2% peptone, 2% dextrose YPDSZ 轉形株篩選 1% yeast extract, 2% peptone, 2% dextrose, 1M
sorbitol, 1.5% agar, 100 μg/mL Zeocin
BMGY 轉形株培養 1%yeast extract, 2%peptone, 1.34%YNB, 4x10- 5% biotin, 100 mM potassium phosphate pH 6.0, 1% glycerol
BMMY 轉形株誘導表現
目標蛋白質
1% yeast extract, 2%peptone, 1.34%YNB, 4x10-5% biotin, 100 mM potassium phosphate pH 6.0, 1% methanol
mBMGY[57] 轉形株培養 1% yeast extract, 1% (NH4) 2SO4, 100 mM potassium phosphate pH 6.0, 1% glycerol mBMMY[57] 轉形株誘導表現
目標蛋白質
0.1% yeast extract, 1% (NH4) 2SO4, 100 mM potassium phosphate pH 6.0, 0.5% methanol
FBS 醱酵槽培養 0.93 g CaSO4, 18.2 g K2SO4, 14.9 g MgSO4-
7H2O, 4.13 g KOH, 40 ml Glycerol, 26.7 ml phosphoric acid (85%) 加水定量至 1 L PTM1 醱酵槽培養所需
微量元素
6.0 g CuSO4-5H2O, 0.08 g NaI, 3 g MgSO4- H2O, 0.2 g Na2MoO4-2H2O, 0.02 g H3BO3, 0.5 g CoCl2, 20 g ZnCl2, 65 g FeSO4-7H2O, 0.2 g biotin, 5 ml H2SO4 加水定量至 1 L
PTM4 醱酵槽培養所需
微量元素
2 g CuSO4·5H2O, 0.08 g NaI, 3 g MnSO4·H2O, 0.2 g Na2MoO4·2H2O, 0.02 g H3BO3, 0.5 of CaSO4·2H2O, 0.5 g CoCl2, 7 g ZnCl2, 22 g FeSO4·7H2O, 0.2 g biotin, 1mL of H2SO4加水 定量至 1 L
20
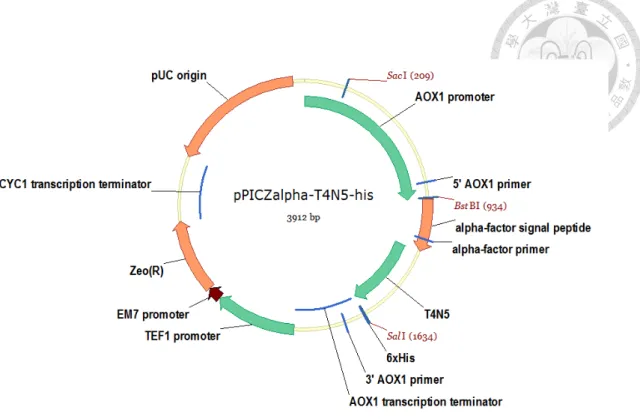
三、Pichia pastoris 表現載體建構
pPICZαB 質體骨架購自 Invitrogen 公司(Carlsbad, CA, USA),T4N5 基因序 列亦委託 Invitrogen 公司全基因合成,去掉頭尾的起始密碼子和終止密碼子,
並加上 Sal I 限制酶切位。
pPICZα-T4N5-his
由於前人研究中指出,T4N5 的 N 端蘇氨酸氨基對於其活性有重要影響,
增減或改動胺基酸皆會造成影響,故以 overlap PCR 的方式將合成的基因序列 直接接在 α factor 訊息胜肽 STE13 蛋白質內切酶切位後方,防止 N 端胺基酸 殘留。並以 overlap PCR 擴增出 5’AOX promoter 至 Sal I 切位的序列,以便利 用 5’AOX promoter 上的 BstBI 切位和 Sal I 切位將序列接進 pPICZαB 表現載體 中。建構好之表現載體如圖五。建構所使用引子參照表二。
圖 五、本研究所使用表現載體圖譜 α-T4N5-his
22
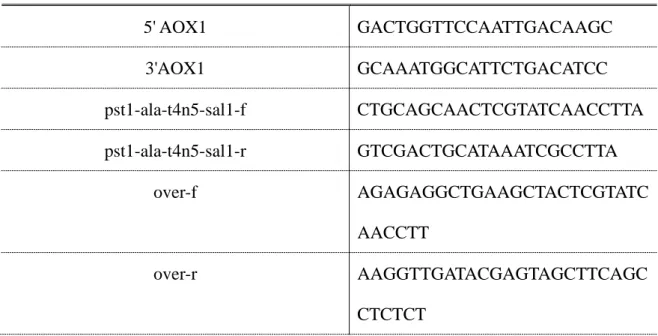
表 二、PCR 使用引子
Table 2. The primers used for plasmids construction
Name Sequence 5' to 3'
5' AOX1 GACTGGTTCCAATTGACAAGC 3'AOX1 GCAAATGGCATTCTGACATCC pst1-ala-t4n5-sal1-f CTGCAGCAACTCGTATCAACCTTA pst1-ala-t4n5-sal1-r GTCGACTGCATAAATCGCCTTA
over-f AGAGAGGCTGAAGCTACTCGTATC AACCTT
over-r AAGGTTGATACGAGTAGCTTCAGC CTCTCT
四、Pichia pastoris 轉形與篩選
1. 轉形 DNA 製備
帶有質體的 E. coli 轉形株以含25 μg/mL Zeocin 的 LSLB 培養基在 37℃、
250 rpm 條件下培養 16 小時,抽取質體,以限制酶 Sac I 將 pPICZαB 系統之質 體截切成線型(linear form)以利用同源重組(Homologous recombination)嵌入 P.
pastoris 染色體基因中。
2. 勝任細胞製備
野生型 P. pastoris KM71H 之單一菌落接種至 3 mL YPD 液態培養基,於 30oC、轉速 250 rpm 振盪培養活化至 OD600約 15。取適量菌液接種至含 100 mL YPD 的 500 mL 三角瓶,使 OD600 約 0.1~0.2。於 30oC、轉速 250 rpm 振盪培 養至 OD600約 2.0 後,將菌體以 1500 g 4oC 離心 10 分鐘,以下步驟皆於冰上操 作。去除上清液,加入 50 mL Pre-treat solution (100 mM LiOAC, 10mM DTT, 0.6 M sorbitol, 10 mM Tris-HCl, pH7.5),重新懸浮菌體,於 30oC 靜置 30 分鐘,再 以 1500 g、4oC 離心 10 分鐘,去除上清液,以 2 mL 1 M 山梨糖醇 (sorbitol)懸 浮菌體,再以 1500 g、4oC 離心 10 分鐘,重複清洗三次,加入 1 M sorbitol 回 溶,使菌體濃度達到 1010 cells/mL,每管分裝 80 µL 置於冰上以備轉形。
3. 電穿孔轉形[58]
將 1 µg 線型質體與 80 µL 勝任細胞混合,移入已預冷之電擊管 (2 mm Gap cuvette , No.620 BTX, San Diego, USA),置冰上 5 分鐘,使用 Electro Cell Manipulator ECM630 Electroporation System (BTX, San Diego, CA, USA),以 1.5 kV、25 μF、200 Ω 條件給予電脈衝,完成後加入 1 mL 1 M sorbitol 回溶菌體,
並於 30oC 下靜置 1 小時,每 15 分鐘翻轉一次,最後取約 250 μL 菌液塗至含
24
Zeocin 的 YPDSZ 篩選培養基中 (1% yeast extract, 2% peptone, 2% dextrose, 1M sorbitol, 1.5% agar, 100 μg/mL Zeocin),於 30oC 下培養 2~3 天。
4. Pichia pastoris 轉形株篩選 4.1 抗藥性濃度梯度篩選
菌落長出後,將 YPDSZ 篩選培養基上之轉形株菌落以無菌牙籤挑起,並將 單一菌落點至含不同 Zeocin 濃度 (100、300、500 μg/mL ) 之 YPDZ 篩選培養 基,於 30oC 培養 2~3 天,觀察菌落大小,選取於高濃度 Zeocin 培養基上生長 較佳之轉形株進行後續分析。
4.2 轉形株染色體 DNA 分析
將篩選之目標轉形株接種於 3 mL YPDZ 培養基培養 24 小時,離心收取菌 體,抽取染色體 DNA 並以專一性引子進行聚合酶鏈鎖反應 (PCR),確認目標基 因是否成功插入染色體。
五、Pichia pastoris 誘導表現 T4 endonuclease V
1. 搖瓶培養
轉形株接種至 3 mL YPDZ 液態培養基,於 30℃、250 rpm 培養 24 小時活 化,取適量菌液,加入含 100 mL mBMGY 培養液之 500 mL 三角搖瓶中,使菌 體初始 OD600 為 0.1, 30℃、250 rpm 培養 24 小時,再以 3000 g 離心 10 分鐘,
去除上清液,以 20 mL (即原體積的 1/5)含 1%甲醇之 mBMMY 重新懸浮菌體,
於 30℃、250 rpm 條件下誘導培養,每天取樣 0.2 mL,離心取上清液保存於-20
℃,再加入 0.2 mL 甲醇使其濃度維持在 1%,胞外上清液進行 SDS-PAGE 膠體 分析、酵素活性測定與總蛋白質定量。
2. 醱酵槽培養
本實驗使用 5 L 醱酵槽 (FS-01-A05, Major Science, TW),分成三階段培養,
先後為批次培養(Batch),批次饋料培養(Fed-batch),甲醇誘導培養(Methanol induction)。批次培養階段,菌體利用槽內培養基養分大量生長,當碳源用盡時 菌體耗氧銳減使得溶氧值(DO)大幅提高,此時進入批次饋料培養,添加額外碳 源以達高細胞密度,隨後進入甲醇誘導階段。
2.1 前置培養
轉形株以試管小量活化後,取適量菌液接種至含100 μg/mL Zeocin 的 100 mL mBMGY 培養液中,以三角搖瓶 30℃、250 rpm 培養 22~24 小時,使 OD600 約為 16~20,作為種菌接種至醱酵槽中,槽內為含 4%甘油為碳源的 2 L FBS/BMGY 培養基,並添加 8.7 mL 的微量元素 PTM1/PTM4,培養溫度調控在 30oC,以 10%氨水及 2 N 硫酸控制 pH 於 6.0,通氣量 2 vvm (4 lpm),攪拌速率 為 800 rpm。,約 14-18 小時候甘油耗盡,溶氧值急遽跳升,以含 12 mL/L 微量 元素 PTM 的 250 mL 50% (v/v)甘油作為碳源進行批次饋料培養,過低的溶氧不
26
利於菌株生長,控制添加流速和時間使溶氧維持在 20-40%,約 6 小時內結束批 次饋料培養。利用槽內正壓將菌液注入滅過菌的收集瓶中,分裝至廣口離心瓶,
以 3000 g、4oC 離心 5 分鐘回收菌體,再以 2 公升 BMMY/mBMMY 培養基重 新懸浮菌體後注入醱酵槽,進入甲醇誘導培養。
2.2 甲醇誘導培養
含 12 mL/L 微量元素 PTM 的無菌甲醇為碳源誘導轉形株生產目標異源蛋 白質,為使菌體適應碳源轉換,逐步提高甲醇濃度,前兩個小時以 1 mL/h/L 的 速率添加甲醇,之後每半個小時提升 10%至最終流速為 3 mL/h/L 左右,每 24 小 時取樣測量 OD600吸光值、生菌數及蛋白質定量。
六、蛋白質產物分析
1. 西方墨點法
將含目標蛋白質的誘導後上清液與含 8% SDS 的 4X Protein sample buffer 混勻,在 99oC 下作用 10 分鐘。配置上層 5% SDS 聚丙烯胺焦集膠體,和下層 15% SDS 聚丙烯胺分離膠體,架設於直立式電泳槽 (HoeferTM Dual Gel Caster;
Amersham Biosciences, USA),並在內外槽加入適量 Tris-glycine SDS running buffer,將備好的蛋白質樣本,注入上層焦集膠體齒梳槽內,先通電 70 V,使 蛋白質在上下層膠體間聚集形成一直線,待焦集完成,即提高至 120 V 進行電 泳至追蹤染料跑出膠體。取出膠片浸泡於 Transfer buffer 中 5 分鐘,0.45 μm PVDF 轉印膜 (PerkinElmer, USA) 浸泡於 100%甲醇 10 分鐘,同時將濾紙及膠 片浸泡於 Transfer buffer。在轉漬夾片上,由負極至正極依序放置海綿、濾紙兩 片、電泳膠片、PVDF 轉印膜、濾紙兩片、海綿,扣上夾片並放置於濕式轉印 槽 (Bio-Red, USA) 內,於槽內加入適量 Transfer buffer,以 400 mA 及 150 V 條件轉印 100 分鐘。將轉印完成的 PVDF 轉印膜浸泡於 gelatin-NET 室溫振盪 1 小時,加入 gelatin-NET 稀釋 5000 倍之 rabbit anti-his 一次抗體 4oC 隔夜。
PBST buffer 振盪清洗三次,每次 10 分鐘。再以 gelatin-NET 稀釋 5000 倍之 goat anti-rabbit HRP-conjugated 二次抗體於室溫振盪 60 分鐘, PBST buffer 振 盪清洗三次,每次 10 分鐘。最後以冷光呈色劑 ECL 反應並照相。上述使用試 劑之配方如表三。
2. LC-MS/MS
委託台大基因體醫學中心進行蛋白質鑑定,以 Discoverer Daemon 1.3 比 對 Chihta_Mascot_NCBI_Viruses 資料庫進行分析。
28
表 三、西方墨點法分析所用試劑之組合成分 Table 3. The constituent of Western blotting reagent
Name Constituent
5% Western blot stacking gel 30% Acrylamide (29:1) 1.5M Tris-HCl (pH=6.8)
10% SDS (Sodium dodecyl sulfate) 10% APS (Ammonium persulfate) TEMED (Tetramethylethylenediamine) ddH2O
15% Western blot running gel 30% Acrylamide (29:1) 1.5M Tris-HCl (pH=8.8)
10% SDS (Sodium dodecyl sulfate) 10% APS (Ammonium persulfate) TEMED (Tetramethylethylenediamine) ddH2O
4x Protein sample buffer (pH=6.8) 8% SDS
200 mM Tris-HCl (pH=6.8) 0.4% Bromophenol blue 20% Glycerol
400 mM 2-ME (2-Mercaptoethanol)
Tris-glycine SDS running buffer (pH=8.4) 25 mM Tris
250 mM Glycine 0.01% SDS
Transfer buffer (pH 8.3) 192 mM Glycine
25 mM Tris-Base 20% methanol
PBS (pH 7.4) 137 mM NaCl
2.7 mM KCl 4.3 mM Na2HPO4 1.47 mM KH2PO4
PBST (pH 7.4) PBS with 0.2% Tween-20
30
七、T4 endonuclease V 活性測定
照射過 UV 的質體會產生 CPD,加入 T4N5 作用可專一性辨識到 CPD 並將 其切開,造成單股 DNA 斷裂,使質體由 supercoiled form 轉變成 nicked circular form,在洋菜膠中有肉眼可見的明顯泳動速率差異,便可利用這原理進行 T4N5 活性定量。
T4N5 的酵素活性單位較無統一標準,本研究採用 New England Biolabs (UK) 的活性單位定義:1 Unit 的 T4N5 可使 0.5 µg 受 UV 照射過的 pUC19 超過 95%由 supercoiled form 轉變成具單股 DNA 斷裂的 nicked circular form,反應在 37°C 下 進行 30 分鐘,總反應體積 20 µL,反應完的樣品以 DNA 洋菜膠電泳分析比例,
每個受 UV 照射過的質體上約有 3-5 個 CPD。
具體實驗方法如下:pUC19 利用 UV crosslinker (UVILink CL 508, Uvitec, Cambridge, UK) 照射 700 J/m2 254 nm 的 UVC,取 0.5 µg 於微量離心管中,加入 T4N5 reaction buffer (100 mM NaCl, 1 mM DTT, 1 mM EDTA, 25 mM Na2HPO4, pH 7.2)及 100 µg/ml BSA 進行反應,37°C 下反應 30 分鐘,反應完成後取 1 µL 進行 DNA 洋菜膠電泳,再藉由 Image J 分析條帶亮度比,求得胞外上清液中的酵素活 性。
八、培養基胞外上清液蛋白酶活性分析
搖瓶或醱酵槽培養的細胞上清液以酶譜法(Zymography)分析蛋白酶活性,以 利調整培養條件減低上清液中蛋白酶活性,防止目標異源蛋白質被降解。
製備10% SDS-PAGE,分離膠體含0.1%明膠,樣品加入Protein sample buffer,
於37℃反應10分鐘後,注入電泳樣品槽中,於4℃冷房中以100V 進行電泳。追蹤 染料跑出後,將膠體取出置於2.5% Triton X-100 溶液中,震盪反應40分鐘,重複 兩次,洗去膠片中之SDS。以二次水清洗膠片數次,加入胰蛋白酶之活性分析緩 衝液(100 mM Tris-HCl, pH 8.2),於37℃反應24小時,以CBR染劑染色30分鐘後退 染觀察結果。
32
第三章、 結果
一、 轉形株篩選與確認
1. 抗性濃度梯度培養基篩選轉形株
將表現載體以 SacI 切成線狀後,使用電穿孔轉形法送入 P. pastoris KM71H 勝任細胞,接著以含100 μg/mL Zeocin YPDSZ 抗性培養基篩選轉形株,待 2~3 天菌落長成後,再利用不同 100 和500 μg/mL Zeocin 濃度的抗性培養基篩選表 現可能較佳的轉形株 (圖六 a,b) 。
2. 染色體 PCR 確認目標基因插入
挑出數株在 Zeocin 濃度500 μg/mL 抗性培養基生長較快速之轉形株,再以 引子 pst1-ala-t4n5-sal1-r 和 over-f 進行染色體 PCR 分析,確認目標基因的確成 功插入 (圖七)。挑選 1 個轉入 pPICZα 的轉形株作為對照組,及 5 個轉入 pPICZα-T4N5-his 的轉形株 4、8、9、11、12 號,進行後續的異源蛋白質表現 分析。
圖 六、以不同 Zeocin 抗性濃度篩選轉形株
(a) 目標重組蛋白質表現載體 pPICZα-T4N5-His;(b) vector control pPICZα
Figure 6. Zeocin tolerance plate assay
(a) pPICZα-T4N5-His; (b) vector control pPICZα
34
圖 七、轉形株染色體 DNA 之聚合酶連鎖反應
將抽取出之染色體以 pst1-ala-t4n5-sal1-r 和 over-f 引子對擴增出大小為 400 bp 的 片段,確認載體成功插入染色體中。M 代表 1 kb DNA ladder;N 代表添加野生 型 P. pastoris 之染色體作為模板股之負控制組;P 代表添加質體為模板股之正控 制組。
Figure 7. Genomic PCR of transformants
The size of gene fragments amplified by pst1-ala-t4n5-sal1-r/over-f primer pairs are 400 bp. M: 1 kb DNA ladder; N: wild type, negative control; P: positive control.
二、 搖瓶培養結果
1. 以 SDS-PAGE 篩選表現量高轉形株
轉形株 4、8、9、11、12 號以搖瓶 1%甲醇誘導 120 小時,取上清液以 SDS-PAGE 比較胞外上清液中的目標蛋白質含量,如圖八所示,在目標蛋白質 大小的 16 kDa 附近可觀察到蛋白質條帶,最終選擇條帶較深的第 11 號轉形 株應用於後續實驗。
2. 以西方墨點法確認目標蛋白質順利表現
目標蛋白質重組 T4N5 的 C 端接有 6 個 Histidine,可用於蛋白質純化,並 可被 anti-his 抗體辨認,故轉形株 11 號(his11) 以搖瓶 1%甲醇誘導 120 小時,
取上清液進行西方墨點法,如圖九所示,確認大小約 16 kDa 的蛋白質條帶隨 誘導時間而增加,且能被 anti-his 辨認,即為目標重組蛋白質 T4N5 候選。
3. LC-MS/MS 確認目標蛋白質
將 SDS-PAGE 上可能是目標蛋白質的條帶委託台大基因體醫學中心進行 蛋白質鑑定,以 Discoverer Daemon 1.3 比對 Chihta_Mascot_NCBI_Viruses 資 料庫進行分析。如圖十所示,確認該條帶確實為重組 T4N5。
4. 搖瓶上清液 T4N5 活性測定
以 his11 搖瓶 1%甲醇誘導 120 小時,收取上清液並分析上清液中 T4N5 的 活性。如圖十一所示,取不同體積的上清液個別反應後再以 DNA 洋菜膠電泳 分析,經三重複誘導取樣並測定,計算出搖瓶上清液中的 T4N5 活性約為 0.47±0.05 U/µL。
36
圖 八、搖瓶上清液 SDS-PAGE 與西方墨點分析-I
T4N5 轉形株 his4、8、9、9、11、12 以搖瓶進行甲醇誘導 5 天,每天添加 1%
甲醇並取樣,每個樣品槽加入 20 µl 不同誘導時間之胞外上清液。跑 15%
SDS-PAGE 並以 CBR 染色。M 為 protein marker。
Figure 8. SDS-PAGE of flask culture supernatant of transformant his4, 8, 9, 11, 12 after methanol induction 0-5 day
圖 九、搖瓶上清液 SDS-PAGE 與西方墨點分析-II
T4N5 轉形株 His11 以搖瓶進行甲醇誘導 5 天,每天添加 1%甲醇並取樣,每 個樣品槽加入 20 µl 不同誘導時間之胞外上清液。(a)15% SDS-PAGE 以 CBR 染色。(b)西方墨點法分析,1 次抗體為 anti-his 5000 倍稀釋,2 次抗體為 anti- rabbit 5000 倍稀釋。V 為 vector control 採取同樣步驟進行甲醇誘導之第 5 天 上清液,M 為 protein marker。
Figure 9. SDS-PAGE of flask culture supernatant of transformant His11 after methanol induction 0-5 day (a) and Western blot analysis (b).
38
圖 十、以 LC-MS/MS 進行蛋白質鑑定,紅色字體為 T4N5 序列中辨認到的胺 基酸序列。
Figure 10. The assigned peptides sequences were matched to the T4 endonuclease V.
Red letters indicate the identified amino acids.
圖 十一、T4N5 活性測定 DNA 電泳膠圖
在 20 µL 的反應體積內加入 0.5 µg 經 700 J/m2 254 nm UV 處理的 pUC19 質體,
及 0 到 2.8 µl 的 T4N5 轉形株甲醇誘導第 5 天上清液,37oC 反應 30 分鐘後取 2 µL 進行 DNA 電泳。N 為加入 vector control 甲醇誘導第 5 天上清液的控制 組,T4N5 為 T4N5 轉形株甲醇誘導第五天上清液與未經 UV 處理的 DNA 反 應。
Figure 11. T4N5 activity assay
0.5 µg of UV irradiated supercoiled pUC19 DNA in a total reaction volume of 20 µl.
Add T4N5 transformant methanol induction day 5 supernatant, react in 30 minutes at 37°C, load 2 µL in each well and run the gel electrophoresis. Lane N: add vector control methanol induction day 5 supernatant; T4N5: Add T4N5 transformant methanol induction day 5 supernatant to plasmid without UV irradiation.
40
三、 醱酵槽培養結果
1. 以 BSM 培養
轉形株 his11 以 Invitrogen 公司建議的醱酵槽培養基 basal salt medium (BSM)進行培養,由醱酵參數圖(圖十二)可觀察到進入甲醇誘導期後生菌數及 OD 皆持平,且在菌體適應甲醇後,鹼添加量和甲醇添加量成正比關係。進入 甲醇誘導其後定時取上清液,並以 SDS-PAGE 分析(圖十三),然而在 SDS- PAGE 上無法看到目標蛋白質的條帶。
2. 以 BMGY 置換到 BMMY 培養
由於 BSM 培養基目標蛋白質產量不如預期,於是先仿效搖瓶的培養條件,
以含 4% 甘油的 2 L mBMGY 批次培養,待槽體中碳源耗盡,DO spike 後再添 加甘油進行批次饋料培養,然而接菌後約 17 小時菌體便不再增長,溶氧值維 持在 60-70%左右,額外添加甘油亦無法使溶氧值下降,顯示實驗室前人修飾 過的培養基 mBMGY 應不適用於高細胞密度培養[57]。
故改用 BMGY 的原始配方,額外添加甘油置 4%批次培養,待碳源耗盡 DO spike 後將菌液在無菌的條件下取出回收,並以 2 L BMMY 培養基置換,以 2.6 mL/L/h、1.7 mL/L/h、1.3 mL/L/h 三種速率添加甲醇進行誘導,醱酵參數如圖 十四,生菌數、OD600、胞外上清液總可溶蛋白質濃度如圖十五,添加速率 2.6 mL/L/h 的誘導後 60 小時取樣以 SDS-PAGE 及西方墨點分析(圖十六),確認目 標異源蛋白質條帶,並以 SDS-PAGE 比較三種添加速率的異源蛋白質產量(圖 十七),然而其產量皆低於搖瓶誘導結果,且非上清液中主要的蛋白質。
參考過去文獻,降低溫度以 23oC 進行誘導[59, 60],進入甲醇誘導期後的 溶氧值起伏較大,且誘導前三天鹼添加量顯著提升(圖十八),胞外上清液以 SDS-PAGE 分析,誘導前三天目標蛋白質產量大幅提升(圖十九)。
3. 蛋白酶活性分析
以酶譜法(Zymography)分析醱酵上清液中的蛋白酶活性(圖二十),透明的 條帶代表膠體中的明膠被分解,無法以 CBR 染劑染色。由條帶粗細可觀察到,
蛋白酶活性隨誘導天數上升,並隨著甲醇添加速率上升有增加的趨勢。
42
圖 十二、轉形株 His11 以 BSM 進行甲醇誘導之醱酵槽參數圖-
His11 以含 4%甘油的 BSM 培養,再以 50%甘油批次饋料 5 到 6 小時,總誘導時間 120 小時,甲醇添加速率 2.5 mL/L/h。
Figure 12. His11 transformant induced by methanol fed with fermenter control. The batch culture is 2 L BSM medium.
圖 十三、BSM 培養基醱酵上清液 SDS-PAGE 分析
總誘導時間 120 小時,以 15%SDS-PAGE 進行蛋白質電泳,每個樣品槽加入 20 µl 不 同誘導時間之胞外上清液,再以 CBR 染色。M 為 Protein marker,P 為搖瓶誘導第 5 天之上清液。
Figure 13. SDS-PAGE of fermentation culture supernatant of transformant His11. M:
protein marker; P: flask culture supernatant of transformant His11.
44
圖 十四、轉形株 His11 以 BMMY 進行甲醇誘導之醱酵參數圖
His11 以含 4%甘油的 BMGY 培養,再以 50%甘油批次饋料 5 到 6 小時,回收菌體 並置換成 BMMY 培養基,總誘導時間 120 小時。甲醇添加速率(a) 1.3 mL/L/h,(b) 1.7 mL/L/h,(c) 2.6 mL/L/h。
Figure 14. His11 transformant induced by methanol fed with fermenter control. The batch culture is 2 L BMGY . After glycerol fed-batch, harvest the cells by centrifugation, resuspend cell pellet in 2 L BMMY. Methanol induction rate: (a) 1.3 mL/L/h, (b) 1.7 mL/L/h, (c) 2.6 mL/L/h.
圖 十五、轉形株 His11 以 BMMY 進行甲醇誘導之生菌數及吸光值
His11 以含 4%甘油的 BMGY 培養,回收菌體並置換成 BMMY 培養基,分別以(a) 2.6 (b) 1.7 (c) 1.3 mL/L/h 的速率添加甲醇,每 24 小時取樣測定生菌數、OD600、
胞外培養液總可溶蛋白質濃度。
Figure 15. His11 transformant induced by methanol fed with fermenter control. The batch culture is 2 L BMGY . After glycerol fed-batch, harvest the cells by centrifugation, resuspend cell pellet in 2 L BMMY. Methanol induction rate: (a) 1.3 mL/L/h, (b) 1.7 mL/L/h, (c) 2.6 mL/L/h.
46
圖 十六、BMMY 培養基醱酵上清液 SDS-PAGE 及西方墨點法分析
總誘導時間 120 小時,每個樣品槽加入 20 µl 不同誘導時間之胞外上清液進 行蛋白質電泳及西方墨點分析,1 次抗體為 anti-his 5000 倍稀釋,2 次抗體為 anti-rabbit 5000 倍稀釋。(a)誘導 0-60 小時上清液以 15% SDS-PAGE 進行 CBR 染色 (b)西方墨點分析
Figure 16. SDS-PAGE of fermentation culture supernatant of transformant His11 after methanol induction 0-60 h (a) and Western blot analysis (b). SDS-PAGE of supernatant after methanol induction 0-60 h
圖 十七、不同甲醇添加速率醱酵上清液 SDS-PAGE 分析
分別以 2.6、1.7、1.3 mL/L/h 的速率添加甲醇,並在誘導後第 3~5 天每 24 小時 取樣進行 SDS-PAGE 分析,P 為搖瓶誘導第 5 天之上清液。
Figure 17. SDS-PAGE of fermentation culture supernatant of transformant His11 in different methanol induction rate: 1.3, 1.7, 2.6 mL/L/h. M: protein marker; P: flask culture supernatant of transformant His11.
48
圖 十八、降溫誘導醱酵參數圖
His11 以含 4%甘油的 BMGY 培養,再以 50%甘油批次饋料 5 到 6 小時,回收菌體 並置換成 BMMY 培養基,同時將培養溫度由 30oC 降至 23oC,總誘導時間 120 小 時,甲醇添加速率 2.5 mL/L/h。
Figure 18. His11 transformant induced by methanol fed with fermenter control. The batch culture is 2 L BMGY . After glycerol fed-batch, harvest the cells by centrifugation, resuspend cell pellet in 2 L BMMY, culture at 23oC. Methanol induction rate: 2.5 mL/L/h.
圖 十九、降溫誘導醱酵上清液 SDS-PAGE 分析
His11 以含 4%甘油的 BMGY 培養,回收菌體並置換成 BMMY 培養基,以 23oC,
甲醇添加速率 2.5 mL/L/h 進行誘導,總誘導時間 120 小時,每個樣品槽加入 20 µl 不同誘導時間之胞外上清液進行 SDS-PAGE 分析。
Figure 19. SDS-PAGE of fermentation culture supernatant of transformant His11, methanol induction rate: 2.5 mL/L/h, culture at 23oC. M: protein marker; P: flask culture supernatant of transformant His11.
50
圖 二十、醱酵槽上清液蛋白酶活性分析
以酶譜法測試培養基上清液中的蛋白酶活性,透明條帶即代表礁體內的明膠被蛋 白酶分解。比較(a)醱酵槽培養基(甲醇添加速率 2.6 mL/L/h)與搖瓶培養基,(b)醱酵 槽甲醇添加速率 2.6 mL/L/h 與 1.3 mL/L/h,(c)醱酵槽甲醇添加速率 2.6 mL/L/h,誘 導溫度 30oC 與 23oC 的蛋白酶活性差異。
Figure 20. Zymography analysis of proteolytic activity in supernatant. Comparing of (a) fermenter and flask, (b) methanol feeding rate 2.6 mL/L/h and 1.3 mL/L/h, (c) induction temperature 30oC and 23oC at methanol feeding rate 2.6 mL/L/h.
第四章、 討論
本論文利用 P. pastoris 生產重組 T4 endonuclease V,並結合 5 公升小型醱酵槽 進行高細胞密度培養。在搖瓶表現階段可成功得到大小正確,具功能性,且可藉由 訊息胜肽分泌至胞外,為胞外培養液中的主要蛋白質。胞外培養液離心後未濃縮上 清液依照 New England Biolabs 的活性單位定義,即可達 0.47±0.05 U/µL,總可溶 蛋白質濃度約 0.1 mg/mL,因此認為此轉形株具有生產潛力。
然而在放大至醱酵槽規模時,即便以同樣的培養基及培養溫度、pH 值誘導,
仍無法得到理想的結果,甚至產量較搖瓶更低。因此以各個醱酵參數及定時取樣的 樣品分析數據,希望能找出原因並且加以改善。
一、 培養基成分之探討
1. BSM (Fermentation Basal Salts medium)
一開始參考 Invitrogen 公司建議的醱酵槽培養基 basal salt medium (BSM) 進行培養,醱酵參數圖無異狀(圖十二),進入甲醇誘導期後鹼添加量隨甲醇添 加量增加而增加,但分析 SDS-PAGE 時無法看到目標蛋白質條帶(圖十三)。文 獻指出,在 BSM 培養基中添加 1 % casamino acid,可減少培養液中蛋白酶對 目標蛋白質進行降解[61],然而本實驗中添加 casamino acid 後無顯著差異,為 求重現搖瓶理想誘導結果,將培養基條件換成接近搖瓶的 BMGY,誘導前置換 為 BMMY。
52
2. BMGY 培養,誘導前置換為 BMMY
搖瓶培養時使用本實驗室前人依據 Invitrogen 公司建議配方改良的培養 基,將成本較高的 YNB 及 peptone 替換成硫酸銨,降低成本且有助於異源蛋 白質產量提升[57]。但在醱酵槽規模中嘗試,發現此改良培養基無法進行高密 度細胞培養,故改以原培養基配方 BMY,添加 4%甘油(glycerol)為碳源以 BMGY 進行醱酵。並在甘油批次饋料後取出醱酵液離心,再以 BMY 回溶,添 加甲醇(methanol)為唯一碳源以 BMMY 進行誘導。
二、 甲醇添加速率之探討
1. 鹼添加量
由醱酵參數圖(圖十四)中可觀察到,進入甲醇誘導期後菌體代謝甲醇產酸,
為維持 pH 值恆定,醱酵槽控制器會自動添加鹼,故鹼添加量與甲醇添加量成 正比關係。然而對照實驗室前人的實驗數據[57, 62],發現前人實驗結果中鹼添 加量除以甲醇添加量的比值較高。
根據 P. pastoris 之甲醇代謝路徑圖(圖三),甲醇在過氧化體內被代謝成甲 醛 後 , 可 經 由 異 化 作 用 (dissimilation) 來 產 生 能 量 , 或 者 經 由 同 化 作 用 (assimilation) 增加生物質量 (biomass)[23]。若走異化作用最終產生二氧化碳及 能量,二氧化碳溶於培養基中使 pH 值下降即產酸。因此,同樣的甲醇代謝量,
鹼添加量越少便代表同化作用比例越高,即添加進去的甲醇主要用以增加生物 質量。對照前人實驗結果可推測,本實驗醱酵過程中有甲醇被用來增加生物質 量的情形較顯著。
2. OD600及生菌數
比較不同甲醇添加速率的生菌數及 OD600可發現(圖十五),甲醇添加量越 高者誘導期間 OD600越高,而生菌數換算成對數值前即無明顯差異。研究指出 P. pastoris 在高細胞密度醱酵條件下進入甲醇誘導期時,會因甲醇代謝副產物
過氧化氫產生氧化壓力,造成菌體死亡[45, 63]。故推測較高的吸光值代表培養 液中有較多死亡的細胞,隨甲醇添加量增加而增加;生菌數無顯著差異則代表 培養基中的甲醇濃度、養分及代謝廢物濃度仍能維持一定量的菌體生長,死亡 後即由新的菌體補充,和上述生物質量的假設相符。
3. 胞外上清液蛋白酶活性
以酶譜法分析培養液中的蛋白酶活性(圖二十),發現蛋白酶活性確實隨甲 醇添加量增加而增加,並且隨誘導天數增加而增加,和上述菌體死亡並裂解的 假設相符。
4. SDS-PAGE 分析及胞外上清液總可溶蛋白質
將不同甲醇添加速率下醱酵培養收取的樣本以同一片 SDS-PAGE 進行分 析(圖十七),可發現甲醇添加速率最高者非目標蛋白質條帶最為明顯,儘管蛋 白酶活性最高,目標蛋白質條帶仍是最為明顯的,且降低甲醇添加速率雖可降 低蛋白酶活性,但無助於異源蛋白質生產。推測可能原因為誘導期間是以甲醇 做為唯一碳源,儘管降低甲醇添加量減緩菌體死亡,碳源減少仍會對異源蛋白 質生產造成影響。
54
三、 其他培養條件之探討
由上述結果可推測,同樣的甲醇添加模式下無論再增加或再減少,都無助 於目標蛋白質的生產,因此須以其他方式減少細胞裂解及減低培養液中蛋白酶 活性。
文獻指出[59, 60] P. pastoris 降溫誘導有助於生產異源蛋白質,並且可能牽 涉到許多因素,例如有助於防止蛋白質摺疊過快,減少菌體死亡,降低蛋白酶 活性,提升 AOX 活性等[53]。本研究從 30oC 降至 23oC 進行誘導,發現目標 蛋白質產量大幅提升,在誘導後兩到三天產量最高(圖十九),並且鹼添加量最 高,誘導後四到五天蛋白質產量明顯減少,且鹼添加量亦減少(圖十八)。
由於本研究所使用的醱酵槽無法自動計算液體添加體積,進入甲醇誘導後 的饋料方式是以固定時間 15 分鐘內蠕動幫浦轉動的秒數來估計大概添加量,
再以重量變化求得實際添加量並作圖。在降溫誘導過程中,溶氧值變化明顯加 劇,除了可能是溫度降低對溶氧值造成的影響,亦和研究中指出的 AOX 活性 增加結果相符[53]。
誘導過程中生菌數無明顯變化,因此鹼添加量的上升代表甲醇代謝走異化 路徑比例較高。由本實驗結果可發現,鹼添加量越高,異源蛋白質產量越高,
故可以鹼添加量作為醱酵生產的指標。
蛋白酶活性分析亦顯示誘導前三天醱酵液中蛋白酶含量較低,代表菌體較 少死亡破裂現象,並且由於溫度較低,進一步降低醱酵槽內蛋白酶作用現象(圖 二十)。
但在誘導後第三天開始,鹼添加量明顯減少,觀察醱酵過程的參數變化,
推測可能是隨著甲醇添加的間隔過大,代謝完畢溶氧值上升時菌體碳源受限,
分解培養基中蛋白質已取得碳因而產鹼,使得鹼添加量受到影響。蛋白酶活性 分析亦顯示,誘導後第三天開始蛋白酶活性顯著增加,並造成目標蛋白質在誘
導後第四天大幅減少。後續將縮短甲醇添加間隔時間,以減緩此情況。
56
第五章、 結論
本研究欲以嗜甲醇酵母菌 P. pastoris 生產重組 T4 endonuclease V,具有生長快 速,成本低廉,可進行高密度培養等優勢,且相較現行多採用的大腸桿菌而言,沒 有內毒素疑慮並可利用外泌訊息胜肽將目標蛋白質外泌至培養基中方便純化。
實驗結果如下:
1. 以搖瓶進行甲醇誘導培養,可得到大小正確具活性的重組 T4N5,胞外上清 液活性達 0.47±0.05 U/µL,總可溶蛋白質濃度約 0.1 mg/mL。
2. 以 5 公升醱酵槽進行甲醇誘導培養,在與搖瓶同樣培養基及溫度、pH 值情 況下,誘導結果不如預期,甚至較搖瓶更低。
3. 以不同甲醇添加速率進行醱酵培養,發現甲醇添加速率越高,培養液中蛋 白酶活性越高,死菌數越高,但生菌數持平,推測是菌體因甲醇造成的壓 力而死亡,部分裂解造成胞內蛋白酶釋出,而多餘的甲醇又不走異化路徑 而走同化路徑生成生物質量,種種因素皆影響到異源蛋白質的生產。
4. 降低誘導溫度,可使異源蛋白質產量大幅提升。分析樣本發現醱酵液中蛋 白酶含量降低,同時醱酵參數中的鹼添加量大幅提升,代表降低溫度不只 降低蛋白酶活性,亦可減低菌體死亡情形使培養基中蛋白酶含量降低,並 增進甲醇利用效率。
5. 甲醇誘導過程中,鹼添加量可作為異源蛋白質產量的指標。
第六章、 未來展望
本研究首次以嗜甲醇酵母菌 P. pastoris 生產重組 T4 endonuclease V,並成功生 產出具活性的異源蛋白質,保存下多株具有生產潛力的轉形株。但在進一步放大生 產規模時發現,異源蛋白質容易被蛋白酶降解,產量不如預期,因此有必要調整醱 酵條件來達到高產量。同時,胞外上清液中最終的產物是許多因素交叉影響下的結 果,因此仍需多方面多次嘗試才能得到最理想的產量。本研究現階段發現,降低誘 導溫度有助於大幅提升異源蛋白質產量,然而降低溫度在工業應用上會增加成本,
未來仍可再進一步探討提升產量的策略,例如:
醱酵槽培養方面。
1. 改變甲醇添加模式,如持續添加或調整添加間隔,或以甲醇監控儀控制槽 內甲醇濃度等。
2. 調整誘導 pH 值,降低蛋白酶活性。
3. 增加誘導前的生物質量,使誘導期間菌體細胞密度更高。
轉形株方面
4. 檢測菌體內是否有蛋白質未順利外泌,並以分生策略增加外泌效率。
5. 以蛋白酶缺陷株或 Mut+菌株建立轉形株。
另外,本研究主要使用的轉形株 His11 雖帶有 His tag 並可藉由西方墨點法偵測,
但以鎳管柱純化時嘗試多種 pH 值及鹽濃度組合仍無法結合上管柱,在流徙步驟即 被沖洗下來,推測可能是 C 端的 His tag 被包埋在蛋白質中影響結合。但 T4N5 的 N 端在蛋白質活性上扮演重要角色[6],不適合將 tag 加在 N 端上,因此本實驗亦 已建立起不帶有 His tag 的 T4N5 轉形株,由於 T4N5 的 pI 值為 9.83,相較一般蛋 白質高,可望以陽離子交換法進行有效的純化。

![圖 二、T4N5 結構[54]](https://thumb-ap.123doks.com/thumbv2/9libinfo/9608249.633853/20.892.127.786.117.609/圖二T4N5結構54.webp)
![圖 三、嗜甲醇酵母菌之甲醇利用途徑。[23]](https://thumb-ap.123doks.com/thumbv2/9libinfo/9608249.633853/21.892.173.730.130.810/圖三嗜甲醇酵母菌之甲醇利用途徑23.webp)