國立臺灣大學生物資源暨農學院昆蟲學系 碩士論文
Department of Entomology
College of Bioresources and Agriculture
National Taiwan University Master Thesis
大頭金蠅(雙翅目:麗蠅科)幼蟲於巴拉刈影響下 之發育變化及異速生長
Allometric growth and developmental changes of Chrysomya megacephala (Diptera: Calliphoridae) larvae under paraquat
ingestion treatment
郭羿宏 Yi-Hung Kuo
指導教授:蕭旭峰 博士 Advisor: Shiuh-Feng Shiao, Ph.D.
中華民國 103 年 6 月
June 2014
i
致謝
能夠完成這本碩士論文,真的要感謝很多人。首先當然是指導老師蕭旭峰老 師,從大三進入實驗室開始,老師在許多方面都給予了意見以及想法,並且無條 件的支持我們想做的實驗,並且花了許多時間修改我的論文。除此之外,也十分 謝謝口試委員:吳文哲老師、楊秋和老師、唐立正老師、以及黃文伯老師,針對 研究以及論文撰寫的部分給予了許多實用的建議。
感謝世綵學姐,不管是在一開始麗蠅飼養的方法、或是實驗種種瑣碎的難題 上,都給予了好多的幫忙,更是平實談天的好對象。名鍾學長也常常提出與法醫 昆蟲領域不同角度的觀點,以及改良實驗的方法,使我受惠良多。除此之外,實 驗的方面也要感謝奧山利規老師,在試驗設計以及統計學上提供了許多的建議以 及幫助。感謝宗學平常給予一些想法的建議以及在實驗上的支援,還可以一起講 垃圾話。謝謝朝盛在最後實驗的衝刺階段幫了我許多忙、以及口試的時候幫我擔 任記錄,內容清晰有條理。其他實驗室的夥伴:栴、孫哥等是平日分擔壓力的好 朋友。
最後要謝謝我的家人,爸爸、媽媽、還有爹地、媽咪,願意在各方面支持我。
兩位哥哥也在求學過程中給予了很多的幫助及建言。由於太多人要感謝,族繁不 及備載,總之再次謝謝大家。
中文摘要
最小死後間隔時間 (minimum post-mortem interval) 是犯罪調查中重要的線 索之一,通常可透過驗屍來估算,如:測量屍溫 (algor mortis) 及檢驗屍斑 (livor
mortis)。法醫昆蟲學也是常被用作最小死後間隔時間估算的工具之一,一般我們 使用節肢動物的發育及消長作為判定的指標。然而,不論驗屍或使用法醫昆蟲做 估算,結果時常會受到許多不同因子的影響。舉凡溫度、濕度或屍體狀態都有可 能造成不同。眾多因子當中,藥毒物對估算是頗具影響力的因子之一。我們在許 多狀況下必須針對藥毒物進行校正,因此我們需要有可信的校正方法。現階段的 化學分析常有再現性低落以及變異性高的問題,導致發生無法準確預測及校正的 狀況。為解決此問題,本研究旨在發展一新的校正方法。此新校正方法為利用大 頭金蠅 (Chrysomya megacephala)蛆蟲後氣孔 (posterior spiracle) 的異速生長
(allometric growth) 對受到不同巴拉刈濃度影響的蛆蟲進行區分。此新的方法可 解決舊有校正方法中存在的可能問題。本實驗使用混合巴拉刈 (paraquat) 的人 工飼料餵養大頭金蠅之幼蟲。每日將切下幼蟲之腹部末節進行形態變化的觀察,
並使用線性判別分析來檢測判定準確度。根據實驗結果,本方法的判定準確率於 二齡蟲中可達 75%,三齡蟲則為 43.48% (隨機猜測為 33.33%)。結果可證實此 一新的概念具有前瞻性,並可被用作替代性的校正方案。然目前三齡蟲後氣孔於 齡期內的不穩定度較高,使得我們在三齡的判定正確率低落,故往後仍需持續改 良此方法,找出其他更為穩定的特徵。在此之前我們建議本方法只針對二齡幼蟲
iii
進行校正。
關鍵詞:大頭金蠅、異速生長、後氣孔、死後間隔時間、法醫昆蟲毒理學、法醫 昆蟲學
Abstract
Minimum post-mortem interval (mPMI) is an important information in crime scene investigation. It can be estimated through autopsy, such as examining algor mortis or livor mortis. Forensic entomology is also one of our candidates of estimation tool. We usually estimate mPMI by examining the development or succession of arthropods in forensic entomology. However, no matter autopsy or forensic entomology, the estimation is often affected by many factors such as temperature, humidity, or the condition of carcass. Among these factors, drug and toxin are two most influential factors, which may have high impacts on our estimation.
Maggots grow slowly (or faster) with drug made the under-estimation (or over-estimation) of mPMI. As the consequence, a reliable calibration for drug and toxin’s impact is necessary. Chemical analysis is currently applied to detect the drug in maggots, but it is unreliable due to the low reproducibility and high variation. In this research, we introduce a new method which is calibrating the bias by checking the allometric growth of Chrysomya megacephala’s posterior spiracles. This new concept may solve the difficulties which exist in the current methods. Maggots of Chrysomya megacephala were reared on the artificial medium mixed with three
different concentrations (0, 500, 1000 ppm) of paraquat to inspect daily change of characters on its posterior spiracle. The results analyzed by linear discriminant
v
analysis showed the 75.00% and 43.48% maggots of 2nd and 3rd instar, respectively, were accurately predicted the concentration of paraquat provided during their developments. The accuracy, especially the estimation of 2nd instar larvae, is higher than 33.33% under randomly guessing. Despite it is not so high that can be directly applied, it is worth to pay more attention in the advantage of considering allometric growth in mPMI estimation, as well as its ecological significance of maggot's developmental response to the drug.
Keywords: Chrysomya megacephala, allometry, post spiracle, post-mortem interval, forensic entomotoxicology, forensic entomology
Table of Contents
致謝... i
中文摘要... ii
Abstract ... iv
Table of Contents ... vi
List of figures ... ix
List of tables ... x
1 Introduction ... 1
1.1 Post-mortem interval (PMI) estimation ... 1
1.2 Forensic entomology ... 2
1.2.1 Forensic entomology used in mPMI estimation ... 3
1.2.2 Chrysomya megacephala ... 4
1.2.3 Factors lead to misestimation... 5
1.3 Entomotoxicology ... 9
1.3.1 Difficulties in entomotoxicological analysis ... 9
1.3.2 Morphological changed caused by drugs / poisons ... 10
1.4 Spiracles ... 10
1.5 Paraquat (PQ) ... 11
1.6 Allometric growth ... 13
1.7 Aims ... 13
1.7.1 Constancy of characters on posterior spiracle within same instar stage .. 14
1.7.2 Influence of PQ on larval length ... 14
1.7.3 Influence of PQ on the structure of developmental profiles of larvae ... 14
1.7.4 Different population densities in poison treatments ... 14
1.7.5 Discriminating different allometry changes of spiracle shapes ... 15
1.7.6 Establishing an alternative calibrating method ... 15
2 Materials and Methods ... 17
2.1 Maintenance of Chrysomya megacephala ... 17
vii
2.1.1 Maintenance of adults ... 17
2.1.2 Maintenance of larvae ... 17
2.2 Medium for larval experiment ... 18
2.3 Experimental set-up ... 19
2.3.1 Test of characters’ constancy ... 19
2.3.2 Preparing medium of different PQ concentrations. ... 19
2.3.3 Assigning tested individuals ... 19
2.3.4 Experimental environment ... 20
2.4 Sample collection and preservation ... 20
2.4.1 Sample collection ... 20
2.4.2 Sample preservation ... 20
2.5 Morphological examination ... 20
2.5.1 Measurement of larval length ... 21
2.5.2 Measurement of posterior spiracle ... 21
2.6 Data analysis ... 28
2.6.1 Constancy of morphometric measurement in same instar ... 28
2.6.2 Influence of PQ on larval length ... 28
2.6.3 Influence of PQ on larval developmental profiles ... 29
2.6.4 Discriminating different allometry changes of spiracle shapes ... 29
3 Results ... 31
3.1 Size changes of characters on posterior spiracle within the same instar stage 31 3.2 Influence of PQ on larval length ... 35
3.3 Influence of PQ on structure of larval developmental profiles ... 38
3.4 Discriminating larvae of different spiracle shapes by linear discriminant analysis ... 41
4 Discussion ... 46
4.1 Constancy of characters ... 46
4.2 Effect of PQ on larvae of Chrysomya megacephala ... 46
4.2.1 Effect of PQ on larval length ... 46
4.2.2 Effect of PQ on developmental profiles... 47
4.3 Linear discriminant analysis ... 49
4.4 Application ... 51
References ... 53
Appendix ... 59
ix
List of figures
Fig. 1 Photos of posterior spiracles of Chrysomya megacephala. ... 16 Fig. 2 Measurement of larval length of Chrysomya megacephala. ... 23 Fig. 3 Measurements of posterior spiracles on 2nd instar maggot of Crysomya
megacephala. ... 24
Fig. 4 Measurements of posterior spiracles on 3rd instar maggot of Chrysomya
megacephala. ... 25
Fig. 5 Measurements of posterior spiracles on 2nd instar maggot of Chrysomya
megacephala. ... 26
Fig. 6 Measurements of posterior spiracles on 3rd instar maggot of Chrysomya
megacephala. ... 27
Fig. 10 Mean larval length of Chrysomya megacephala under different PQ treatments.
... 36 Fig. 11 Diagram of developmental structure under different PQ treatments using
percentages of different Chrysomya megacephala stages recorded per time point.
... 39 Fig. 12 Diagram of discriminating 2nd instar larvae of Chrysomya megacephala under
different PQ treatments using posterior spiracle characters by LDA. ... 42 Fig. 13 Diagram of discriminating 3rd instar larvae of Chrysomya megacephala under
different PQ treatments using posterior spiracle characters by LDA. ... 43
List of tables
Table 1 Changes of characters' measurements (μm) with larval growth in 3rd instar (feeding stage) ... 32 Table 2 Changes of characters' measurements with larval growth in 3rd instar
(post-feeding stage) ... 33 Table 3 Daily measurement (sd) of characters from 3rd instar larvae of control groups
... 34 Table 4 Changes of larval length with PQ influence per ppm in each stage... 37 Table 5 Parameters of age, PQ concentration and population size that contribute to log odds ratio of proportion of each stage over proportion of 1st instar larvae ... 40 Table 6 Coefficients and the importance ranks of characters used in linear discriminant analysis for 2nd instars of Chrysomya megacephala under different PQ treatments ... 44 Table 7 Coefficients and the importance ranks of characters used in linear discriminant analysis for 3rd instars of Chrysomya megacephala under different PQ treatments ... 45
1
1 Introduction
1.1 Post-mortem interval (PMI) estimation
When investigating the homicide cases or the death cases without witness, many details should be aware of. One of the most important clues is post-mortem interval (PMI), which represents the time elapsed since death (Wells and Lamotte, 2010). By clarifying PMI, we eliminate and refining our lists of victim and suspect who does not match our prediction, which allow us to focus on those who are most suspected (Wells and Lamotte, 2010). However, we never know how long the real PMI is since there is no witness to tell when it exactly happened. In order to approximate the real PMI, we estimate the maximum PMI (MPMI) and minimum PMI (mPMI) to narrow down the estimating range for PMI. The definition of MPMI is the time elapsed since the victim was last seen alive; on the other hand, mPMI is the time elapsed we estimate the time of death by using many different methods (Villet et al., 2010).
Following an animal’s death, there comes many different changing signs. We
estimate mPMI based on the data which was built from the changing patterns of these signs. These signs act as many different “clocks” which can be used to estimate mPMI (Houck and Siegel, 2010). For instance, external examination, such as: algor mortis (the changes of body temperature), livor mortis (settlement of blood in different body parts) can be one of our choices (Houck and Siegel, 2010). Also, we may examine
changes of internal biochemical marker like concentration of potassium in vitreous humour (Tilstone et al., 2006). Except of these traditional methods, we have another option, which is, forensic entomology.
1.2 Forensic entomology
Forensic entomology is an application of arthropods on legal issue (Catts and Goff, 1992). In the medico-criminal investigation, thanks to the wide dispersion of insects, large amount, and the quick arrival of insects, forensic entomology becomes a critical tool (Bourel et al., 2003; Jien, 2004).
According to the book, The Washing Away of Wrongs (Chinese: 洗冤集錄), which written by Sung Tz’u (Chinese: 宋慈 ), the first application of forensic
entomology on homicide case can be traced back to 13th century in China. Later, in 18th - 19th century, some of the forensic doctors in Europe found many different assemblages of arthropod on corpse when exhuming the dead bodies; Forensic doctors like Bergeret use these details to predict the mPMI (Benecke, 2001). But the most important milestone should be the works of arthropods succession on dead bodies, which were done by Mégnin (Catts and Goff, 1992). Since the publication of Mégnin’s works, there comes a breakthrough of forensically related researches in many different aspects (Catts and Goff, 1992). Just like Mégnin, scholars all emphasize the importance of entomological application on investigation, and
3
spreading the concept of forensic entomology to the whole world.
1.2.1 Forensic entomology used in mPMI estimation
Forensic entomology could be used in examining the cause of death (e.g. drug abuse) (Beyer et al., 1980); or be used in locating the crime scene (Walsh-Haney et al., 2010); even could be provided as a source of DNA to identify the criminal (Curic et al., 2014). However, the most common usage is the mPMI estimation. Arthropods will
establish their colonies on carcass soon after the animals’ death, therefore, the developmental time of carrion-feeding arthropods can be considered as an index of mPMI (Villet et al., 2010).
Kashyap and Pillay (1989) suggested that prediction of forensic entomology can be more accurate and reliable than the results from the traditional / prevalent methods.
We can find several reasons to support his idea. First of all, arthropods can be found in almost all territorial landscape (Castner, 2010). Second, it only takes very short time for insects to arrive the dead bodies. Some studies even suggested flies will arrive and lay eggs within five minutes after the victims died (Bourel et al., 2003; Jien, 2004). This allows us to consider the time that flies arrive is very close to the time that victim died. Finally, the composition of arthropods is unique under different time, locations or the causes of death (Introna et al., 2001).
We usually use different methods to estimate mPMI in long term and short term
interval. In terms of long term interval (e.g. months or years), the mPMI estimation is based on the correlation between decomposition stages and different arthropods assemblage (Payne, 1965). According to the integrated data in Merritt and Wallace (2010), the decomposition for terrestrial corpse can roughly be subdivided into five stages: fresh, bloat, active decay, advance decay, dry (or remains). Each stage has different assemblage of arthropods, and these correlations will form a specific succession pattern which can be used on prediction (Payne, 1965). For instance, the larvae of blow flies can be found on corpse since the beginning of fresh stage, and disappear before early stage of dry, or larvae of dermestid beetles will establish their colonies on corpse at the end of decay stage or the start of dry stage (Kreitlow, 2010).
On the other hands, if we want to estimate short term interval (e.g. days to weeks), we usually use age of insect as an indicator instead of using succession pattern. For instance, the dipterans are usually the first one that arrive corpse, their developmental time should be the mPMI estimation which is most close to real PMI.
In practical cases, the size of maggots can be referred to their age. Investigators can collect the maggots from bodies, and measure the body length of maggots in order to
estimate their age. By knowing their age, we can have our mPMI prediction.
1.2.2 Chrysomya megacephala
Chrysomya megacephala, or oriental latrine fly, is the fly generally covered with
5
metallic green color and grayish pollen. In some rare individuals, it could be purplish or bluish. They are 8 - 10 mm in length, and ornamented with orange hair on its parafacials (Kano and Shinonaga, 1968). C. megacephala is an Old World fly (Greenberg, 1988). At first, it is only distributed in Australasian and Oriental regions (Kurahashi, 1982). In recent years, it was introduced into New World: through Brazil, further expanding into Mexico (Greenberg, 1988), and Texas, United State (Wells, 1991). Adults of C. megacephala are easily to be attracted by human food, feces, and carrion (Kano and Shinonaga, 1968). Because of this kind of endophilic behaviors, the eusynanthropic form of C. megacephala becomes one of the most important vectors of disease (Greenberg, 1973; Wells, 1991).
In Taiwan, C. megacephala is one of the predominant necrophagous species (Shiao and Yeh, 2008), and also one of the most earliest fly species which appear on carrion. As a consequence, O'Flynn and Moorhouse (1979) considered Chrysomya species to be primary flies, showing their significance in applications of forensic
entomology.
1.2.3 Factors lead to misestimation
Although there are many advantages of applying forensic entomology on mPMI estimation, but it also has disadvantages, which are the entomological evidences could easily be affected by other factors. Not only individual species-specific characters will
make the difference in prediction, but also lots of factors that may lead to misestimation. Those factors can be roughly divided into two categories: artificial or environmental factors. Sample preservation, or measuring methods could be the possible factors in the former category (Wells and Lamotte, 2010). According to Tantawi and Greenberg (1993) , larval length will be influenced by using different types of preserving fluid or killing methods (Wells and Lamotte, 2010). Adams and Hall (2003) preformed their research specifically toward blow flies, and suggested that by killing larva in boiling water and preserving them into 80% ethanol, the post-mortem changes of larval length can be reduced to minimum. Furthermore, Richards et al. (2013b) mentioned the preservation period also has effects on the larval length. For the latter one, we have known locations, food types, climate, maggot mass, drug and toxin may influence our estimation (Wells and Lamotte, 2010).
1.2.3.1 Body conditions
Different body conditions usually strongly affect the decomposition process. If the bodies are covered or wrapped by other object, the arrival time of primary flies will be postponed by several days (Benecke et al., 2004). Besides, it takes more time for flies to reach indoor carcass than outdoor ones (Reibe and Madea, 2010). For those cases, we may underestimate the mPMI. On the contrary, Avila and Goff (1998)
7
have reported that the fly arrival time will be shorten with those victims who were burned (Anderson, 2010).
1.2.3.2 Food types
There are lots of researches have been done for the influence of food types on fly development. By the reason of differences in nutrient contents, the consumption of different food will affect the developmental rate of larvae, which further influence mPMI estimation (Amendt et al., 2010). Take tissue from different organs as example, comparing the sizes of larvae that reared on brains, liver, lungs, heart or kidney, it is obvious that different developmental data from different food types will lead to totally different conclusions (Kaneshrajah and Turner, 2004). Even if we take same organ tissue into consideration, their difference conditions will also make different consequences (e.g. frozen, spoiled, or fresh liver) (Richards et al., 2013a). There were also some cases showed the effects of water content on corpse decomposition (Anderson, 2010; Moh, 2013).
1.2.3.3 Climate
Climate has its significant importance on mPMI estimation. Temperature contributes a lot to the developmental rate of larvae (Komar, 1998); activity of flies depends on weathers (e.g. sunny, rainy day), which influences the time they arrive.
1.2.3.4 Maggot mass
Many researches showed the importance of maggot mass might be more than we expected (Chuang, 2013; Deonier, 1940; Rivers et al., 2011). Generally, the maggots will accumulate together to form a maggot mass, increasing efficiency of feeding and digestion progress (Anderson, 2010; Greenberg and Kunich, 2002). Also, maggot mass can generate heat and maintain their body temperature (Rivers et al., 2010), which the heat can regulate their developmental rate when environmental temperature does not reach the idea condition (Chuang, 2013).
1.2.3.5 Drug and toxin
In addition to those mentioned above, drug, toxin, or other chemical compounds could have various impacts. Goff (2000) mentioned that when he was investigating a suicide case in 1987, he accidentally found cocaine has some effects on maggot development. Although they did realize the developmental rates will be different under abnormal conditions, such as being reared on food with mercury (Nuorteva and Nuorteva, 1982), not until then did they learn such “abnormal conditions” were that common. By the reason of this discovery, lots of related researches were published since then. Until now, the drug like Buscopan (Oliveira et al., 2009), insecticide malathion are known to have negative effects on developmental rate of maggots (Rashid et al., 2008). Contrarily, others like cocaine can accelerate maggots’ growth (Goff et al., 1989).
9
1.3 Entomotoxicology
Since 1980, entomologists have done lots of works, detecting the drug or toxin inside the insects, in order to find some useful tools which can be used in forensic investigation (Gosselin et al., 2011; Monthei, 2009). Beyer et al. (1980) accomplished one of first drug qualitative researches on maggots. They extracted drug from maggots of Cochliomyia macellaria, using gas chromatography to identify the drug is a kind of anticonvulsant (or antiepileptic) drug - phenobarbital. After that, many similar researches proposed one after the other. However, due to many limitations of applying laboratory data on the practical case, some scientists did not believe in its
potential prospects.
1.3.1 Difficulties in entomotoxicological analysis
Gosselin et al. (2011) pointed out several limitations and problems, such as the results of detection may be interfered by different sampling locations (e.g. brain, muscle, or liver). In general, most investigators randomly sample maggots from corpses in crime scenes (Definis-Gojanovic et al., 2007; Introna et al., 1990).
However, Gosselin et al. (2011) suggested that it is better to collect samples from internal organs. Moreover, the most serious problem is that almost all the pharmacokinetics of drug in insects is still unclear. According to Nolte et al. (1992) and Kharbouche et al. (2008), the pharmacokinetics may be different among different insect species or even different life stages in a same species (Gosselin et al., 2011).
Those issues make the prospects of entomotoxicology in forensic applications become
controversial.
1.3.2 Morphological changed caused by drugs / poisons
Some researches indicated that insects may have morphological changes due to the different environmental factors they live. For instance, Reynolds and Ferrington (2002) mentioned that larvae of Chironomidae have different morphological responses when exposure to different contamination level of heavy metal. Although the mutation rate may be variously depended on the species, the rate increased in all the studied species when they live in the seriously contaminated environment.
1.4 Spiracles
Basically, dipteran species have 10 pairs of spiracles, which are located on the prothorax, metathorax, and each abdominal segment (Stehr, 1991). The spiracles which located on prothorax is referred to as anterior spiracles, and those located on caudal segment of abdomen is known as posterior spiracles (Stehr, 1991). The others are named after the segment where they located. The structures of spiracles not only differ among species, but also among different instars or where they located (Stehr, 1991). Especially, the structures of posterior spiracles are much more different from that of the others. In calliphorid species, posterior spiracles arise from upper side of the posterior spiracular disc (Stehr, 1991). Posterior spiracle usually consists of an
11
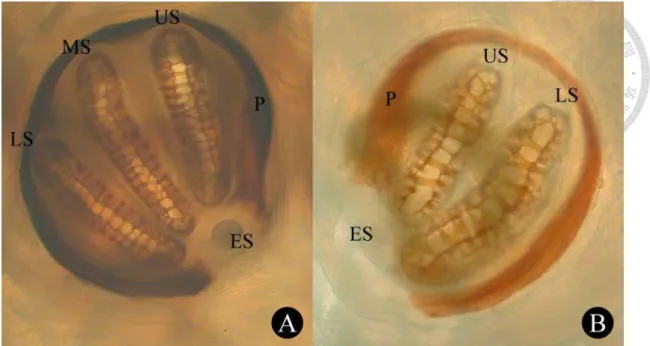
ecdysial scar, a broken peritreme that surrounding two or three spiracular splits in second and third instar larvae respectively (Stehr, 1991). Terminologies are depicted in Fig. 1.
According to Wang et al. (2008), the sizes of posterior spiracles increase dramatically from the younger instars to the older ones, however, it remains same size within the same instar stage. In other words, the structure of posterior spiracle is a constant indicator within the same instar stage.
1.5 Paraquat (PQ)
Paraquat (1,1’-dimethyl-4,4’-bipyridylium dichloride, PQ), a non-selective and quick-acting contact herbicide, is widely used in the world (Melchiorri et al., 1996).
Davenport (1963) reported that bipyridylium quaternary compounds will react with NADPH-cytochrome c (or cytochrome P-450), and act as temporary sink for electrons.
PQ will collect the electrons and return it to NADPH-cytochrome P-450, inhibiting the electron transport chain by decreasing oxidative activity (Wilkinson and Murray, 1984). In addition, PQ has direct cellular toxicity which comes from its redox cycle (Dinis-Oliveira et al., 2006). When PQ is reduced by NADPH-cytochrome c reductase, the PQ radical will be generated. With the presence of oxygen, PQ radical has ability to produce superoxide radical (O2-
). The superoxide radical will further involve in a cascade, which the final product hydroxyl radical (OH-) is harmful to
cells (Dinis-Oliveira et al., 2006; Melchiorri et al., 1996).
PQ has also often been used as a poison for suicide, or involved in many accidental cases (Li et al., 2011). A high dose of PQ (over 30 ppm in human body) will lead to rapid death caused by the multiple organ dysfunction syndrome (MODS, also known as multiple organ failure), and MODS will accompany with lung damage (e.g. hemorrhage) (Dinis-Oliveira et al., 2006; Onyeama and Oehme, 1984). In addition, a lower dose (between 16 and 30 ppm in human body) of PQ can also be deadly, which results in lung fibrosis (Onyeama and Oehme, 1984). The expansion of fibrosis is followed by malfunction of lung, leading to the death at last. In the PQ related case, there is a great possibility for PQ remains in the corpse and has the chance to be consumed by maggots. Li et al. (2011) showed that PQ may be found in the serum of poisoned patients, and therefore this might influence the maggots which grow on the bodies.
Wilson and Hodgson (1971) firstly showed the existence of NADPH-cytochrome c in dipteran species. According to Wilkinson (1985), isozymes of cytochrome P-450 is involved in the regulation of hormone which contributed to the insects’ growth and development (e.g. biosynthesis of 20-OH ecdysone). Since PQ interrupts NADPH related systems, it is no doubt that PQ will influence the developmental rate of dipteran species. The effect of PQ on dipteran species has been reported by Darvas et
13
al. (1990). The weight of flies in that experiment decreased, and the mortality
increased when the PQ concentration rose.
In addition, Saigusa et al. (2005) reported that body length of two blow fly species became shorter when consuming food which was contaminated by PQ.
Therefore, investigators should be careful when they are dealing with PQ related cases.
1.6 Allometric growth
Allometry is a study that describes the relation between body size to shape (Small, 1996), and has usually been used in describing different growth rates of parts of living organism. In this research, we focus on the allometric growth of several characters in posterior spiracles and try to calibrate mPMI estimation by examining their differences.
We believe that PQ will have different effects on each characters of posterior spiracles, which causing the allometric growth. This difference may allow us to discriminate the effect levels and further calibrate the bias of mPMI estimation.
1.7 Aims
Due to the facts mentioned above, if the influence of possible factors is neglected, the estimation of mPMI may not meet our expectation. In order to calibrate the possible bias, several aspects were included in this research as follows:
1.7.1 Constancy of characters on posterior spiracle within same instar stage At the beginning, in order to take posterior spiracle characters into use, we need to make sure the maggots’ ages do not influence or have little effect to our study. That is because we can only calibrate the mPMI estimation with the indicators which their changes are only depended on different environments, but not influenced by time elapse. If the impact of aging is too large, it may conceal the influence by environmental factors. We mainly focus on 3rd instar, which maggots size changing
dramatically within.
1.7.2 Influence of PQ on larval length
Measuring larval length under different PQ treatment not only can examine the effect of PQ on larval length but also can provide as a reference data for mPMI estimation under PQ effect. However, if we want to predict the mPMI in the real case, we need to replace medium with animal tissue which is more related with the real
case.
1.7.3 Influence of PQ on the structure of developmental profiles of larvae The proportion of different developmental stages will change with the time passing. Therefore, the instar structure of a population can also reflect their developing time, and we can have the preliminary idea of mPMI by knowing their population structure.
1.7.4 Different population densities in poison treatments
15
We knew that maggot mass will change the feeding and developing patterns.
Therefore, by comparing PQ’s impact on different maggot density, we may know
when we can apply our lab data on real case.
1.7.5 Discriminating different allometry changes of spiracle shapes
By knowing the influence of PQ on shape of posterior spiracle, we may figure out the relationship of PQ concentration and the shape of maggots’ posterior spiracle.
Due to the allometric growth, different PQ concentration may cause different responses to the growth of spiracles. By using linear discriminant analysis, we may discriminate different larvae with different spiracle shapes, and thus knowing how much PQ they ingest. The results of discrimination analysis could be used in
calibrating process.
1.7.6 Establishing an alternative calibrating method
The main purpose of this research focuses on establishing a different calibrating method, which may provide an additional option for investigators when instrumental drug analysis is impossible or unavailable. By observing the allometric growth of posterior spiracle and body length, we suppose that some differences could be uncovered to help us separating different drug effects.
Fig. 1 Photos of posterior spiracles of Chrysomya megacephala. A, Posterior spiracle of 3rd instar larvae; B, Posterior spiracle of 2nd instar larvae; ES, ecdysial scar; LS, lower slit; MS, middle slit; P, peritreme; US, upper slit.
17
2 Materials and Methods
2.1 Maintenance of Chrysomya megacephala
The maintenance of blwo fly, C. megacephala, is referred to the method used by Yang and Shiao (2012). The blow flies have been reared in Insect Systematics Lab., Department of Entomology, National Taiwan University over 3 years. Both larvae and adults were reared under 27.5 ± 1°C, with a photoperiod of 12:12 (L:D) hrs, and
relative humidity 70 ± 10 %.
2.1.1 Maintenance of adults
The adults of C. megacephala were fed with the mixture of brown sugar and milk powder (Volume 1:1). Water was provided in a plastic container and induced to the flies with tissue paper. In order to stimulate the development of ovaries, pork liver will be provided in the first 3 days after the flies’ eclosion. The pork liver will be provided again to collect eggs on 5-7 days after eclosion, when the female become sexully mature. Several egg masses can be collected on pork liver, and mature eggs
can be produced by female about one week later.
2.1.2 Maintenance of larvae
The pork liver with the egg mass will be moved into cylindrical plastic container with tissue paper to prevent larvae from being drowned in the accumulated liquid on the bottom. Eggs hatched in one day and larvae will feed on the pork liver. The pork liver will be renewed after drying out. The whole container with 3rd instar larvae will
be moved into a larger plastic pot with sawdust inside. Larvae that enter post-feeding stage will creep out the container and pupate within the sawdust. Pupae will be separated and collected from sawdust and putted in a 30×30×30 cm net cage to wait
until the adult emerge..
2.2 Medium for larval experiment
Medium wes used to rear larvae of Chrysomya megacephala in our experiment, which is referred to Hong (1995). Medium was composed of: fish powder (300 g), yeast powder (120 g), agar (12 g), and water (1.8 L). Medium was prepared with following steps:
i. Soaking agar into the water.
ii. Boiling the water to dissolve agar completely. (Keep stirring during the heating.)
iii. Adding fish powder and yeast powder. (Sieving during adding to prevent the powder from caking.)
iv. Removing the fire immediately when medium is boiling again.
v. Cooling for 3 hours.
vi. After that, 300 g medium will be spooned into a cylindrical plastic container which is 14.3 cm in diameter and 5.5 cm in height, and covered with a cap.
These medium will be stored in the refrigerator at 4 ± 1°C before using.
19
2.3 Experimental set-up
In this research, we prepared media with 3 different PQ concentrations: 0 (control group), 500, and 1000 ppm. Each of them began with 20 and 200 eggs. Each of the six treatments were composed of 15 repeats and discarded one by one after the
number of alive maggots and maggot stages were recorded every day.
2.3.1 Test of characters’ constancy
Preliminary test was done to realize the constancy of characters. Relationships
between size of chosen characters and larval age will be examined.
2.3.2 Preparing medium of different PQ concentrations.
PQ (24% GAMOXONE®, Syngenta, Swiss) was used in treated groups. The concentrations were calculated with only effective PQ component. On the other hand, same amount of double distilled water will be used in control group instead of PQ. All
media were homogenized by using a hand blender (CSB-76TW, Cuisinart®, USA).
2.3.3 Assigning tested individuals
Pork liver will be provided in order to collect the eggs of Chrysomya megacephala. The liver was checked every hour until enough eggs were collected.
The collected egg mass was firstly sprayed with water and then artificially separated by pressing gently. Two hundred and 20 eggs were put on a small piece of tissue paper, and assigned for each concentration test group. Then each container was put into a net bag with sawdust inside. The net bag was made by breathe material. After the
container was set up, the opening of bags will be tied well with rubber band.
2.3.4 Experimental environment
The bags with container were put into growth chamber with temperature of 27.5
± 1°C, photoperiod of 12L:12D, and relative humidity 70 ± 10%.
2.4 Sample collection and preservation
2.4.1 Sample collection
All larvae were collected from the test groups, and their developmental stages were observed and recorded. There are six developmental stages in this experiment, which are egg + first instar, second instar, third instar, post-feeding, pupae and adult.
The instars of larval stages were identified by checking the number and size of spiracular split on posterior spiracular disc. Those larvae that creep outside the container or in the sawdust will be defined as in post-feeding stage. Pupae and adults
will also be recorded if present.
2.4.2 Sample preservation
Only the larvae of 1st to 3rd instar will be collected (without post-feeding stage).
The larvae were killed by boiling water for over 30 seconds in order to prevent it from browning (Niederegger et al., 2011). The collected larvae were preserved in 80%
ethanol for further examination.
2.5 Morphological examination
Samples from age one to 12 days were subdivided into several groups depend on
21
their instar stages. For each instar, at least 10 samples were picked out randomly. If total individual amount of an instar is less than 10, then all samples from that category
will be picked out. All measurements were done by using software ImageJ 1.47v.
2.5.1 Measurement of larval length
Digital camera (D700, Nikon, Japan) was used to take the photos of maggots’
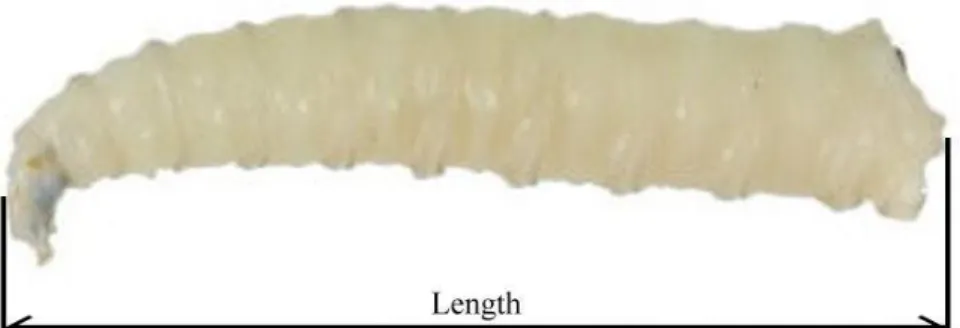
body. Photos of maggots’ body length were taken laterally along with a metric stage micrometer (No. 10310345, Leica, Germany). Definition of a maggot’s body length follows Day and Wallman (2006), which refers to the distance between the head to the
8th abdominal segment as in Fig. 2.
2.5.2 Measurement of posterior spiracle
The cutting method is based on Sukontason et al. (2010). The last segment of maggot’s abdomen will be cut down and mounted on slide in order to inspect the posterior spiracles. Compound microscope (BH2, Olympus, Japan) equipped with digital camera (Coolpix-995, Nikon, Japan) was used for examining all posterior spiracles. Posterior spiracle of 2nd instar larva was examined with 20x objective lens, and which of 3rd instar larva was examined with 10x objective lens.
2.5.2.1 Character selection
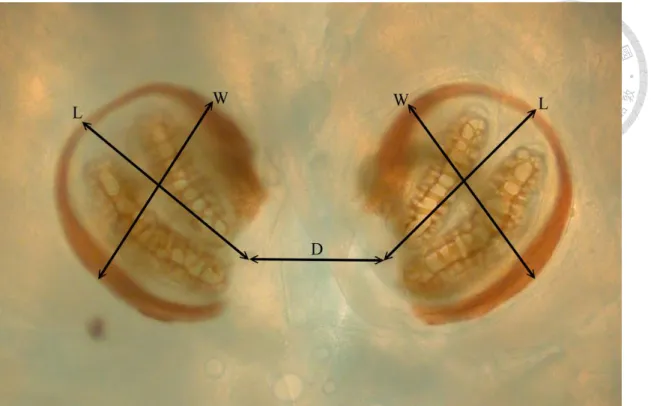
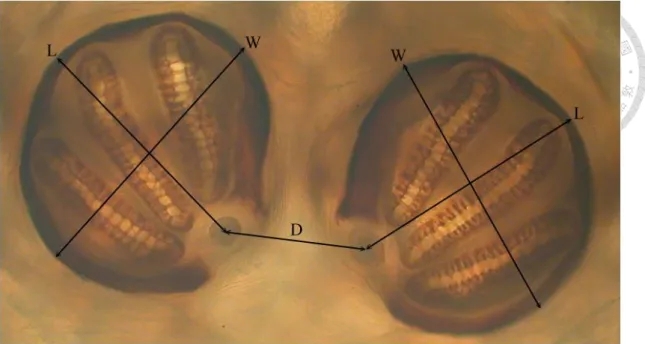
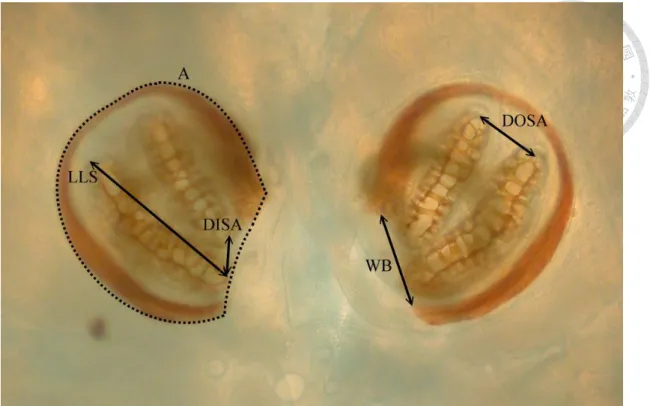
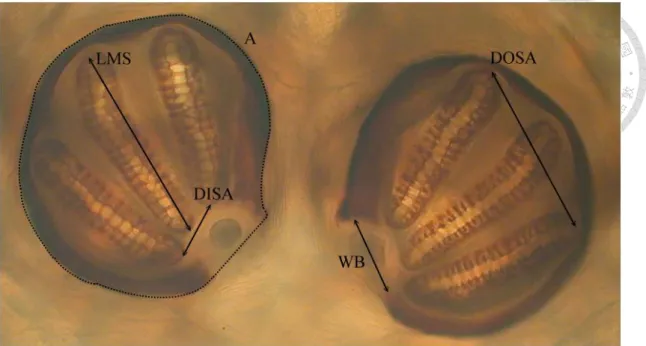
The character selection was mainly based on those in Sukontason et al. (2010), which including length (L), width of both right and left posterior spiracle (W) and the distance between two posterior spiracles (D) (Fig. 3 and Fig. 4). In addition to these
five characters, we further choose the distance from the inner apex of upper slit to the inner apex of lower slit (DISA), also the distance between outer apex (DOSA), length of middle slit (LMS) (lower slit (LLS) in 2nd instar), the width of peritreme broken (WB), and the area of posterior spiracles (A) (Fig. 5 and Fig. 6). We totally choose 15 characters in this research. (Table of characters can be found in Appendix I)
23
Fig. 2 Measurement of larval length of Chrysomya megacephala.
Fig. 3 Measurements of posterior spiracles on 2nd instar maggot of Crysomya megacephala. D, distance between two posterior spiracles; L, length of posterior spiracle; W, width of posterior spiracle.
25
Fig. 4 Measurements of posterior spiracles on 3rd instar maggot of Chrysomya megacephala. D, distance between two posterior spiracles; L, length of posterior spiracle; W, width of posterior spiracles.
Fig. 5 Measurements of posterior spiracles on 2nd instar maggot of Chrysomya megacephala. A, area of posterior spiracle (the area surrounded by dash line); DISA, distance between inner slit apex; DOSA, distance between outer slit apex; LLS, length of lower slit; WB, width of peritreme broken.
27
Fig. 6 Measurements of posterior spiracles on 3rd instar maggot of Chrysomya megacephala. A, area of posterior spiracle (the area surrounded by dash line); DISA, distance between inner slit apex; DOSA, distance between outer slit apex; LMS, length of middle slit; WB, width of peritreme broken.
2.6 Data analysis
RStudio Ver. 0.98.501 and R Ver. 3.0.2 were used in data analysis.
2.6.1 Constancy of morphometric measurement in same instar
The morphometric measurement of sclerotized characters on the insects is sometime not constant as it expected to be due to different level of sclerotization. The maggot's body length is that changes with time in a given instar. Thus, characters on the posterior spiracle with higher level of sclerotization might be more constant between two molting. Here we compared the slopes of the morphometric measurement from the body length and characters on the posterior spiracle against maggot's age (day of development within in a given instar) to find the relatively constant characters (with lower slope) to be adopted in the further analysis. To unify the units of measurement, we square root the area of both spiracles. Samples are the 3rd instar larvae that coming from the control groups, which its population size is 200 individual. Samples will be subdivided into 2 groups, which are above 6 day-old and below / equal to 6 day-old, to prevent from the impact of larval length decreasing
since the entering of post-feeding stage.
2.6.2 Influence of PQ on larval length
Generalized linear regression was used to test the differences of larval lengths, which is a dependent variable. In analysis, larval length behaves against covariate variable age. PQ concentration and population size act as independent variables to
29
compare the effects from the PQ under different population densities. If population size shows no effect on larval length, it will be removed from the model when analyzing the length of each instar respectively. Samples will be subdivided into 4 groups: 1st, 2nd, 3rd instar above 6 day-old, 3rd instar below / equal to 6 day-old, and
being analyzed separately.
2.6.3 Influence of PQ on larval developmental profiles
The effect of the PQ and population size on the fly development was analyzed by multinomial logistic regression with the six developmental stages as the dependent variables. The model choosing is based on the Akaike information criterion (AIC) value which developmental stages behave against covariate age. PQ concentration and population size act as independent variables to compare the effect from the PQ under different population densities. Output of multinomial logistic regression was the log odds ratio of proportion of each stage (except 1st instar) over proportion of 1st instar larvae, which also be referred as relative risk. The formula was:
log ( ).
The ratio was further calculated into daily proportion of each stage.
2.6.4 Discriminating different allometry changes of spiracle shapes
The shapes of posterior spiracles will be discriminated by analyzing total 15 characters with linear discriminant analysis (LDA). Measurement of area was unified as what we done in constancy test. Samples in analysis should be composed of data
from all of the three drug concentration categories in the same day while data with only one or two drug concentrations will be removed. The prediction power of LDA model was tested with additional samples.
31
3 Results
3.1 Size changes of characters on posterior spiracle within the same instar stage
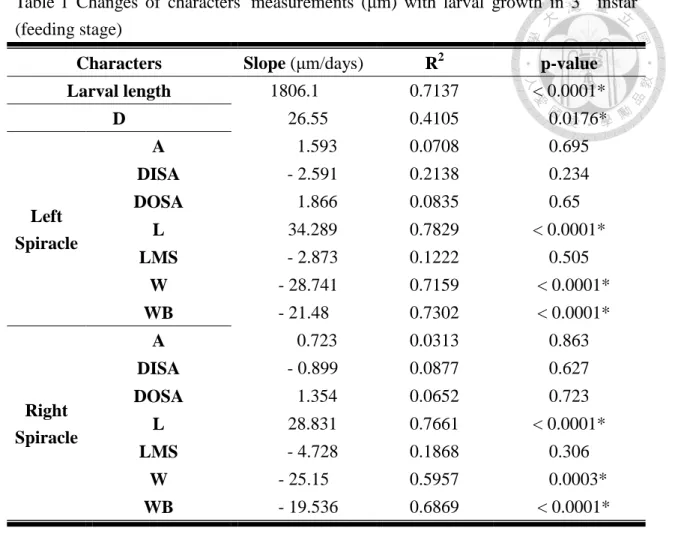
Slope of character sizes against ages are recorded in Table 1 and Table 2. Some of the characters on posterior spiracle were significantly influenced by their age (e.g.
D, L, W, WB). However, all slopes of characters on posterior spiracle are lower than the slope of larval length, no matter the larvae had entered post-feeding stage or not.
Table 1 Changes of characters' measurements (μm) with larval growth in 3rd instar (feeding stage)
Characters Slope (μm/days) R2 p-value
Larval length 1806.1 0.7137 < 0.0001*
D 26.55 0.4105 0.0176*
Left Spiracle
A 1.593 0.0708 0.695
DISA - 2.591 0.2138 0.234
DOSA 1.866 0.0835 0.65
L 34.289 0.7829 < 0.0001*
LMS - 2.873 0.1222 0.505
W - 28.741 0.7159 < 0.0001*
WB - 21.48 0.7302 < 0.0001*
Right Spiracle
A 0.723 0.0313 0.863
DISA - 0.899 0.0877 0.627
DOSA 1.354 0.0652 0.723
L 28.831 0.7661 < 0.0001*
LMS - 4.728 0.1868 0.306
W - 25.15 0.5957 0.0003*
WB - 19.536 0.6869 < 0.0001*
A, area of posterior spiracle; D, distance between two posterior spiracles; DISA, distance from the inner apex of upper slit to the inner apex of lower slit; DOSA, distance from the outer apex of upper slit to the inner apex of lower slit; L, length of spiracle; LMS, length of middle slit; W, width of spiracle; WB, width of peritreme broken. Those characters which significantly changed with aging were marked with asterisk.
33
Table 2 Changes of characters' measurements with larval growth in 3rd instar (post-feeding stage)
Characters Slope (μm/days) R2 p-value
Larval length - 939.7 0.5205 0.0005*
D - 7.22 0.2415 0.128
Left Spiracle
A - 3.089 0.2344 0.14
DISA - 1.047 0.0915 0.57
DOSA - 1.921 0.1329 0.408
L - 5.547 0.3438 0.0277*
LMS - 4.173 0.2903 0.0656
W - 3.304 0.2014 0.207
WB 3.485 0.2964 0.0632
Right Spiracle
A - 3.915 0.3014 0.0555
DISA - 0.1647 0.0173 0.914
DOSA - 0.942 0.0782 0.627
L - 6.206 0.3728 0.0164*
LMS - 4.16 0.3085 0.0497*
W - 4.95 0.3019 0.055
WB 2.355 0.1758 0.272
A, area of posterior spiracle; D, distance between two posterior spiracles; DISA, distance from the inner apex of upper slit to the inner apex of lower slit; DOSA, distance from the outer apex of upper slit to the inner apex of lower slit; L, length of spiracle; LMS, length of middle slit; W, width of spiracle; WB, width of peritreme broken. Those characters which significantly changed with aging were marked with asterisk.
Table 3 Daily measurement (sd) of characters from 3rd instar larvae of control groups
Age
Characters Larval
length D Left spiracle Right spiracle
DISA DOSA L LMS W WB DISA DOSA L LMS W WB
4 1341.03 (143.66)
193.15 (33.31)
364.21 (17.45)
111.09 (10.53)
269.94 (16.42)
362.35 (21.93)
299.05 (23.11)
434.49 (19.97)
152.66 (16.85)
362.24 (14.92)
106.48 (8.27)
266.60 (14.51)
368.33 (20.24)
298.01 (30.86)
422.04 (20.54)
145.36 (18.12) 5 1801.47
(50.85)
334.42 (54.04)
387.39 (11.90)
105.50 (11.38)
268.72 (14.72)
388.53 (19.12)
294.02 (7.72)
448.65 (21.53)
101.41 (8.98)
388.48 (12.30)
101.19 (9.68)
262.25 (18.32)
396.92 (6.29)
304.86 (16.39)
452.93 (5.38)
106.66 (16.80) 6 1720.51
(155.45)
253.74 (48.55)
368.80 (22.23)
105.71
(11.47) NA 430.40
(27.91) NA 379.80 (25.16)
107.76 (17.68)
365.35 (23.40)
104.39
(10.16) NA 425.98
(25.14) NA 375.40 (31.81)
105.04 (19.30) 7 1709.78
(97.58)
275.44 (22.11)
362.37 (13.91)
97.98 (15.59)
267.75 (11.39)
367.65 (21.08)
293.76 (16.85)
427.97 (16.64)
104.44 (13.47)
361.86 (9.92)
101.83 (10.38)
261.84 (11.86)
370.32 (15.07)
292.29 (9.34)
426.97 (11.01)
101.22 (8.57) 8 1626.99
(259.28)
249.66 (45.68)
352.67 (18.53)
105.30 (13.13)
262.79 (19.31)
343.99 (17.53)
283.58 (21.10)
422.78 (17.14)
121.09 (16.09)
350.78 (16.50)
102.72 (13.00)
252.74 (18.36)
346.29 (17.53)
284.38 (21.92)
418.78 (22.18)
123.44 (17.58) 9 1531.63
(289.22)
260.65 (40.27)
359.91 (20.53)
95.72 (12.74)
263.17 (24.43)
354.09 (25.01)
285.94 (17.86)
429.13 (28.03)
112.99 (14.62)
356.68 (22.93)
100.57 (11.30)
258.04 (17.40)
359.98 (23.20)
287.58 (12.51)
423.56 (31.93)
114.40 (11.81) 10 1396.86
(142.43)
228.60 (39.44)
351.76 (15.90)
106.88 (8.16)
256.38 (24.00)
345.14 (12.25)
286.33 (18.53)
416.67 (25.92)
116.28 (9.57)
348.63 (15.95)
110.88 (9.18)
251.81 (18.66)
244.41 (22.74)
278.59 (12.51)
408.51 (13.42)
119.01 (25.50) 11 1368.93
(93.25)
259.69 (34.33)
345.50 (10.64)
87.29 (24.28)
263.99 (8.64)
340.21 (9.24)
267.70 (12.51)
411.88
(13.57) NA 341.88 (8.53)
92.59 (18.61)
260.63 (2.74)
340.13 (17.64)
273.50 (13.99)
406.13 (4.84)
109.84 (15.87) A, area of posterior spiracle; D, distance between two posterior spiracles; DISA, distance from the inner apex of upper slit to the inner apex of lower slit; DOSA, distance from the outer apex of upper slit to the inner apex of lower slit; L, length of spiracle; LMS, length of middle slit; W, width of spiracle; WB, width of peritreme broken. Area of posterior spiracle was square rooted in this table. Larval length shows in millimeter (mm)
35
3.2 Influence of PQ on larval length
Both age and PQ concentration have significant effects on larval length (p-value
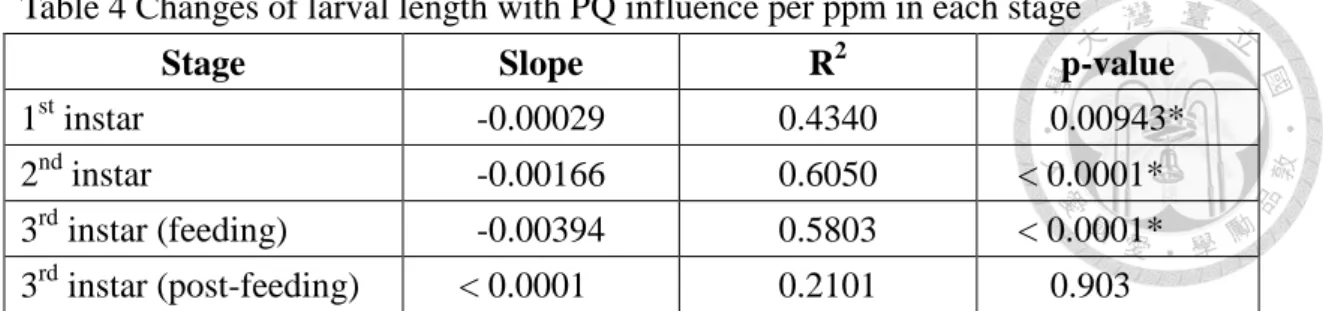
< 0.0001). Neither population density (p-value = 0.257), nor the interaction between PQ concentration and population size (p-value = 0.755) has significant effect on larval length. PQ concentration shorten 0.00029 (p-value = 0.00943), 0.00166 (p-value <
0.0001), and 0.00394 (p-value < 0.0001) mm per ppm of 1st, 2nd, feeding 3rd instar larval length, respectively (Table 4). The effect of PQ will disappear when they get into post-feeding stage (p-value = 0.903). Larval lengths in 500 ppm PQ treatment groups can also reach the maximum size (the larval lengths in control groups at six days old), when they become seven days old.
Fig. 7 Mean larval length of Chrysomya megacephala under different PQ treatments.
Larvae grew under 27.5 ± 1°C, photoperiod of 12L:12D, and relative humidity 70 ± 10%, lengths were measured every day.
37
Table 4 Changes of larval length with PQ influence per ppm in each stage
Stage Slope R2 p-value
1st instar -0.00029 0.4340 0.00943*
2nd instar -0.00166 0.6050 < 0.0001*
3rd instar (feeding) -0.00394 0.5803 < 0.0001*
3rd instar (post-feeding) < 0.0001 0.2101 0.903 Larval length which significantly changed with aging were marked with asterisk.
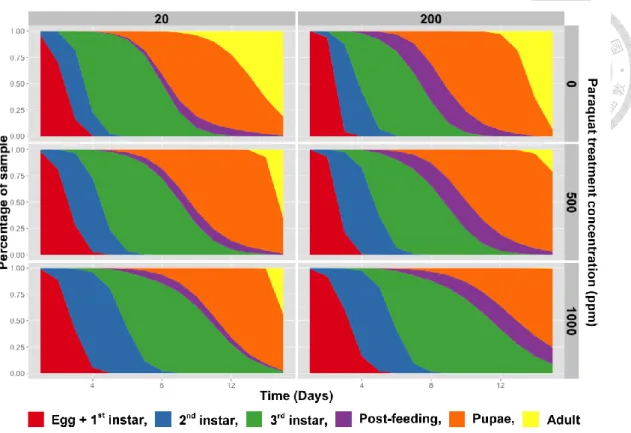
3.3 Influence of PQ on structure of larval developmental profiles
Table 5 shows the parameters of log odds ratio. The positive value of parameter means adding a unit of variable will increase log odds ratio, and vice versa. All parameters of population size in each stage are negative, and so are the parameters of PQ concentration except for that in post-feeding stage. The age has positive effect on log odds ratio in every stages from 2nd instar to adult. Except for the 3rd instar, post-feeding, pupae stage of interaction between age and PQ concentration, all parameter are positive.
39
Fig. 8 Diagram of developmental structure under different PQ treatments using percentages of different Chrysomya megacephala stages recorded per time point (days after egg laying) under 27.5 ± 1°C, photoperiod of 12L:12D, and relative humidity 70 ± 10%.
Table 5 Parameters of age, PQ concentration and population size that contribute to log odds ratio of proportion of each stage over proportion of 1st instar larvae
Stage Age PQ Population size
2nd instar 1.883 -0.002535 -0.048927
3rd instar 4.422 -0.003487 -0.047501
Post-feeding 5.489 0.003220 -0.028015
Pupae 5.626 -0.003101 -0.061321
Adult 6.393 -0.079667 -0.149406
41
3.4 Discriminating larvae of different spiracle shapes by linear discriminant analysis
Larvae of 2nd instar can be clearly separated into three different PQ treatment groups by LDA. Control groups mainly accumulate at bottom-left, 500 ppm groups locate at bottom-right, and 1000 ppm groups are clustered at top side (Fig. 9). When using this model to discriminate samples, the prediction accuracy is 75%. Furthermore, the accuracy can be increased up to 85% if we define the sample into only two groups (with / without PQ ingestion). The separation of 3rd instar larvae are otherwise not clear, and the trend can barely be found (Fig. 10). The accuracy of prediction is 43.48%, and it can be increased to 52.17% when discriminating the 2 with / without PQ ingestion groups.
The importance ranks of characters are listed in Table 6 and Table 7.
Fig. 9 Diagram of discriminating 2nd instar larvae of Chrysomya megacephala under different PQ treatments using posterior spiracle characters by LDA.
43
Fig. 10 Diagram of discriminating 3rd instar larvae of Chrysomya megacephala under different PQ treatments using posterior spiracle characters by LDA.
Table 6 Coefficients and the importance ranks of characters used in linear discriminant analysis for 2nd instars of Chrysomya megacephala under different PQ treatments
Character
LDA 1 LDA 2
Coefficient Importance
rank Coefficient Importance rank
D 2.79 x 10-3 13 -6.07 x 10-3 11
Left spiracle
A -4.97 x 10-4 14 -2.65 x 10-4 14
DISA -4.41 x 10-2 6 -6.63 x 10-3 10
DOSA -2.73 x 10-2 9 1.03 x 10-1 2
L 6.23 x 10-2 3 -5.94 x 10-3 12
LLS -2.26 x 10-2 11 -2.43 x 10-2 8
W -2.28 x 10-2 10 3.13 x 10-2 6
WB 3.11 x 10-2 7 -2.11 x 10-3 13
Right spiracle
A -8.22 x 10-5 15 3.69 x 10-5 15
DISA -2.78 x 10-2 8 -1.41 x 10-2 9
DOSA -1.30 x 10-1 1 1.04 x 10-1 1
L 4.79 x 10-2 5 -6.84 x 10-2 3
LLS -5.13 x 10-2 4 6.56 x 10-2 4
W 7.41 x 10-2 2 -6.00 x 10-2 5
WB -1.06 x 10-2 12 -2.51 x 10-2 7
45
Table 7 Coefficients and the importance ranks of characters used in linear discriminant analysis for 3rd instars of Chrysomya megacephala under different PQ treatments
Character
LDA 1 LDA 2
Coefficient Importance
rank Coefficient Importance rank
D -5.32 x 10-3 8 3.08 x 10-3 13
Left spiracle
A -8.58 x 10-5 15 -8.00 x 10-5 14
DISA -1.36 x 10-3 12 -2.80 x 10-2 2
DOSA 5.40 x 10-3 7 1.11 x 10-2 8
L* 3.92 x 10-2 2 -1.28 x 10-2 6
LMS 5.41 x 10-3 6 -4.84 x 10-3 11
W* 4.69 x 10-2 1 1.66 x 10-2 4
WB 7.61 x 10-3 5 -9.75 x 10-3 9
Right spiracle
A -1.18 x 10-4 14 5.90 x 10-6 15
DISA -2.65 x 10-3 11 -5.06 x 10-3 10
DOSA* 3.65 x 10-2 3 1.60 x 10-2 5
L 3.78 x 10-3 9 -1.68 x 10-2 3
LMS -1.68 x 10-2 4 -1.20 x 10-2 7
W 3.41 x 10-4 13 -3.77 x 10-2 1
WB 2.80 x 10-3 10 3.13 x 10-3 12
4 Discussion
4.1 Constancy of characters
In order to discriminate the morphological difference caused by PQ, character constancy is important to our hypothesis. According to the result of linear regression analysis (Table 1 and Table 2), there are 8 characters that would change with age in feeding stage, and there are 4 characters that would change with age in post-feeding stage. There are some characters changing with increasing of age, but we still can found the slope of all characters is smaller than 100 (μm/days), except for the larval length. That means the changes of the characters on posterior spiracles are smaller than changes of larval length. Thanks to high consistency of posterior spiracle, our new concept might have a chance to be put into practice.
4.2 Effect of PQ on larvae of Chrysomya megacephala
4.2.1 Effect of PQ on larval length
Effect of PQ on dipteran larval length has been reported in Saigusa et al. (2005), which suggested that PQ will decrease mean larval length of Calliphora vicina, while it had no significant effect on larvae of Lucilia sericata. Our results show PQ has negative effect on larval growth of Chrysomya megacephala. No matter in 1st, 2nd, or feeding stage of 3rd instar, the larval length was significantly shorten by PQ. When larvae get into post-feeding stage, we can found the effect of PQ was disappear. The
47
concealing the difference between all PQ treatment.
4.2.2 Effect of PQ on developmental profiles
Saigusa et al. (2005) suggested the initiation of pupation will be postponed due to PQ ingestion in both C. vicina and L. sericata. However, Mahat et al. (2014) indicated that PQ will neither postpone the initial egg laying time of Chrysomya megacephala, nor prolong their developmental time. Our results agree with Saigusa et
al. (2005), which PQ does affect the development of calliphorid larvae. We can
realized that width of some stages increase with the increasing of PQ concentration (Fig. 8), that means, the interval of each stage was prolonged. This will also cause the postponement of next stage. By examining the coefficients of PQ in the formula of model, we also found PQ has negative relation with the odd ratio of each stage except for pupae (Table 5). This refers to the fact that when PQ concentration increases, the proportion of subjective stage will become lesser than those from groups with lower PQ concentration in the same age. In other words, to reach same proportion as control groups, the groups with PQ ingesting treatment will takes more time. Therefore, we may also consider this as a postponement of subjective stage.
. The reason why the results are various among Mahat et al. (2014), Saigusa et al.
(2005) and ours is probably because the different PQ concentrations used in experiments. Saigusa et al. (2005) applied 8.7 ppm PQ in their treatment groups, and
the highest PQ concentration in Mahat et al. (2014) was 200 ppm (five times of lethal dose). The difference between Mahat et al. (2009) and Saigusa et al. (2005) may come from the different PQ sensitivity in different species. Even Mahat et al. (2014) used higher concentration than Saigusa et al. (2005), it seems that Chrysomya megacephala is less PQ sensitive than the other two species. Comparing their
experiments to ours, Mahat et al. (2014) and Saigusa et al. (2005) considered that low concentration may simulate the concentration in victim’s body, but they neglected the practical reality. Generally, those who commit suicide by PQ ingestion will have high PQ concentrations in their mouth and gastrointestinal system, which the concentrations may be much higher than 200 ppm. In addition, flies usually lay eggs on the natural openings (e.g. eye, mouth, anus) (Byrd and Castner, 2010; Moh, 2013) where probably contain high drug concentrations. As the consequence, those maggots collected from mouth or gastrointestinal system will ingest higher PQ concentrations than 200 ppm.
We further compared the model’s coefficients to examine the effect of two different population densities. We can found that coefficient of all stages was negative and without exceptation. This represents that with the increasing of larval density, larvae will stay in the same stage for a longer time. We knew that maggots will show gregarious behavior, which tend to assemble and form a maggot mass to increase their