嘉南藥理科技大學 生物科技系
碩士論文
6-shogaol藉由內質網壓力與氧化壓力誘發Hep - G2 細胞凋亡
6-shogaol Induces Apoptosis in Hep-G2 Cells through ER stress and Oxidative stress
指導老師:洪 瑞 祥 博士 研 究 生:黃 宏 彰
中華民國一百年九月七日
嘉南藥理科技大學生物科技系 Department of Biotechnology
Chia-Nan University of Pharmacy and Science
碩士論文
Thesis for the Degree of Master
6-shogaol藉由內質網壓力與氧化壓力誘發Hep - G2 細胞凋亡 6-shogaol Induces Apoptosis in Hep-G2 Cells through ER stress and
Oxidative stress
指導老師:洪瑞祥 博士(Dr. Jui-Hsiang Hung)
研 究 生:黃宏彰(Hung-Chang Huang)
中華民國一百年九月七日
Sep 7, 2011
誌謝
首先,我要感謝我的指導教授洪瑞祥老師,在老師細心與耐心的指導 與鼓勵下,讓我這個大學從來沒過跟過實驗的學生,學會了許多生物技術 領域的知識與專業技術,並且提供充分的研究資源,使我能順利的完成碩 士論文。此外,我要感謝嘉南藥理科技大學李冠漢老師和成功大學陳昌熙 老師,能夠在百忙中抽空前來擔任口試委員,在我口試期間給予許多寶貴 意見,讓我的論文更加完整流暢。
另外,也要感謝在念碩士班這段日子裡,幫助我的同學以及實驗室的 優子、納豆、奕融、尚一、易陽、志軒、子慧、奇樺、憶儒等等的學弟妹 們,謝謝你們幫我處理了許多大大小小的事情。還有教導我學會流式細胞 儀的騰達行業務蕙敏,沒有你我的實驗不會那麼的順暢。以及我的好同學 佳筠,感謝你常跟我一起預約儀器做實驗,和我分享實驗上的心得。
在來要感謝我的父母,因為您們辛苦的工作省吃儉用,給予了我衣食 無缺的學習環境,在我低落的那段時間你們沒有放棄過我,這些年來你們 辛苦了。還要感謝我最愛的男朋友:Jeff,認識你一路以來謝謝你不停的鼓 勵我、支持我,在我幾次面臨崩潰想放棄的時候,你都默默的看著我、守 護著我。沒有你,我不會完成這篇論文。最後我要感謝我和 Jeff 的愛犬小 特,雖然和你的緣分就這麼短短的十幾天,但在我最艱苦趕稿子那段時間,
摘要
肝癌,長久以來一直是國內癌症死亡率的第一或第二名。近年來的研 究顯示,B 型肝炎、C 型肝炎、肝硬化、酗酒、黃麴毒素等等,都是造成肝 癌的原因。根據統計,在台灣的肝癌患者約有百分之八十都是 B 型肝炎所 導致。目前治療肝癌的化學治療藥物有許多,然而因為肝癌的復發率很高,
化學治療藥物用量會逐漸上升導致產生抗藥性而失效,所以在肝癌的化學
治療藥物值得更進一步的去探討及開發。
先前研究發現,一種從薑中萃取出來的化合物 6-shogaol,在肝癌細胞 中可以透過大量的消耗 GSH,造成細胞內 ROS 含量的累積,進而誘發了細 胞的凋亡。本實驗中藉由添加抑制劑 NAC 抑制 ROS 後發現,確實能有效 的保護細胞因 6-shogaol 產生的 ROS 誘發的細胞凋死亡現象,但仍有一部分 的細胞還是會死亡,顯示 6-shogaol 仍會經由其他的機制造成肝癌細胞的死 亡。
初步結果顯示肝癌細胞經過不同濃度之薑萃取的化合物 6-shogaol 作用 後,肝癌細胞存活率隨 6-shogaol 濃度增加而遞減。進一步的,當藉由流式 細胞儀分析結果顯示,6-shogaol 作用下肝癌細胞內在 1-3 小時會產生大量 的 ROS,而隨 6-shogaol 濃度的增加可明顯的改變肝癌細胞的細胞週期,結 果顯示 6-shogaol 可誘導肝癌細胞的 sub-G1 和細胞凋亡族群比例也隨之提
網壓力,然而當加入 ROS 抑制劑 NAC 後,內質網壓力仍然存在。
綜合以上實驗結果顯示,薑之萃取化合物 6-shogaol 會藉由產生大量的 ROS 和誘發內質網壓力來誘導細胞的死亡,而 ROS 的產生及內質網壓力為 兩個獨立的訊息傳遞路徑,最後期待薑之萃取化合物 6-shogaol 的抗癌機制 能對在治療肝癌上有所幫助。
關鍵詞:肝癌、薑、6-shogaol、ROS、NAC、內質網壓力、細胞凋亡
Abstract
For long, the liver cancer always is in the first or the second place in the death rate in Taiwan. The studies in the few years shows, HBV, HCV, liver cirrhosis, drink excessively, and aflatoxin …etc.; the reason that above all causes liver cancer. According to the statistics, the patients of the liver cancer of the eighty per cent in Taiwan are all caused by HBV. At present, there are a lot of the chemistry treatment drugs for liver cancer treatment. However, because the rate of the recurrence is very high, the amount used of the chemistry treatment drugs will be higher and produce the drug resistance.
Therefore, the drug chemistry treatment in the liver cancer must be more explored and developed.
Formerly, the studies find, from the 6-shogaol compound, a kind of the ginger; it can cause the amount adding of the ROS in the cell with consuming GSH massive in the liver cancer cell, and causes the apoptosis. In the experimentation, by increasing the inhibitor of the NAC into the ROS, find that it could protect the cell effectively because the ROS from the 6-shogaol causes the apoptosis. But still, the part of the cell will died, and show the 6-shogaol still causes the death of the liver cancer cell from others.
In the first result, the cell of the liver cancer, after extracts the 6-shogaol
from the different density of the ginger; the survival rate of the liver cancer cell, can decrease because the consistency of the 6-shogaol increases. Further, from the data of the flow cytometry analysing, the 6-shogaol will produce the large ROS in the cell of the liver cancer, during the first to the third hour, and up to the consistency of the 6-shogaol increases, the cell cycle of the liver cancer cell will be changed; On the other hand, the result, tells that the 6-shogaol will induce obviously the cell strain of the live tumor to produce the ER stress; still is alive the ER stress after add the ROS into the NAC inhibitor.
Integrate above all the experiments and shows, the 6-shogaol compound from the ginger will induce the cell to die from producing the large ROS and the ER stress, and both of the ROS and the ER stress will become the two different alone ways for passing messages. Finally, look forward the Anti-cancer mechanisms of the 6-shogaol abstract compound from the ginger could be helpful for liver cancer treatment.
Keyword:liver cancer, Hep-G2, 6-shogaol, ROS, NAC, ER stress, apoptosis
目錄
頁次 中文摘要………I 英文摘要……….III 本文目錄………..V 圖目錄………..VIII 英文縮寫對照表………..X
第一章 緒論……….1
1.1 肝癌………1
1.2 薑………1
1.3 活性氧分子………2
1.4 內質網壓力………3
1.5 細胞凋亡………4
1.6 研究動機………5
第二章 材料與方法……….7
2.1 細胞培養………7
2.1.1 細胞培養條件...7
2.1.2 細胞繼代培養...7
2.1.4 冷凍細胞...8
2.1.5 解凍細胞...8
2.2 細胞存活率………8
2.3 細胞外型觀察………9
2.4 細胞內 ROS 含量的分析………...………..10
2.5 細胞週期的分析………..…………10
2.6 Caspase-3 分析………..11
2.7 Apoptosis 分析...12
2.8 西方墨點法………..13
2.8.1 細胞內蛋白質萃取………...13
2.8.2 蛋白質定量………...……14
2.8.3 蛋白質電泳………...14
2.8.4 蛋白質轉漬………...15
2.8.5 免疫墨點法………...15
第三章 結果………...16
3.1 6-shogaol 對肝癌細胞存活率的影響………..………16
3.2 6-shogaol 對 Hep-G2 細胞外形的變化………16
3.3 6-shogaol 對 Hep-G2 細胞內活性氧化物(ROS)的影響………..……17
3.5 6-shogaol 對 Caspase-3 的影響………18
3.6 6-shogaol 誘導 Hep-G2 細胞凋亡………..………..19
3.7 抑制 ROS 對 6-shogaol 誘導細胞死亡的影響………19
3.8 6-shogaol 誘導內質網壓力造成 Hep-G2 細胞凋亡………20
第四章 討論………...22
第五章 結論………...26
參考文獻……….27
附圖……….35
圖目錄
圖 1. MTT 試驗測試在藥物 6-shogaol 處理下不同肝癌細胞之存活率...35
圖 2. Hep-G2 細胞在 6-shogaol 半致死濃度(IC50)處理後細胞外形變化...36
圖 3a. Hep-G2 細胞在 6-shogaol 處理後 ROS 含量...37
圖 3b. ROS 增加含量統計圖...38
圖 4a. Hep-G2 細胞在 6-shogaol 處理後細胞週期...39
圖 4b. 6-shogaol 各濃度細胞週期分佈...40
圖 4c. 6-shogaol 各濃度細胞週期統計表...41
圖 5a. Hep-G2 細胞在 6-shogaol 處理後 Caspase-3 含量...42
圖 5b. Caspase-3 含量統計圖...43
圖 6a. Hep-G2 細胞在 6-shogaol 處理後細胞凋亡的變化...44
圖 6b. 細胞凋亡程度統計圖...45
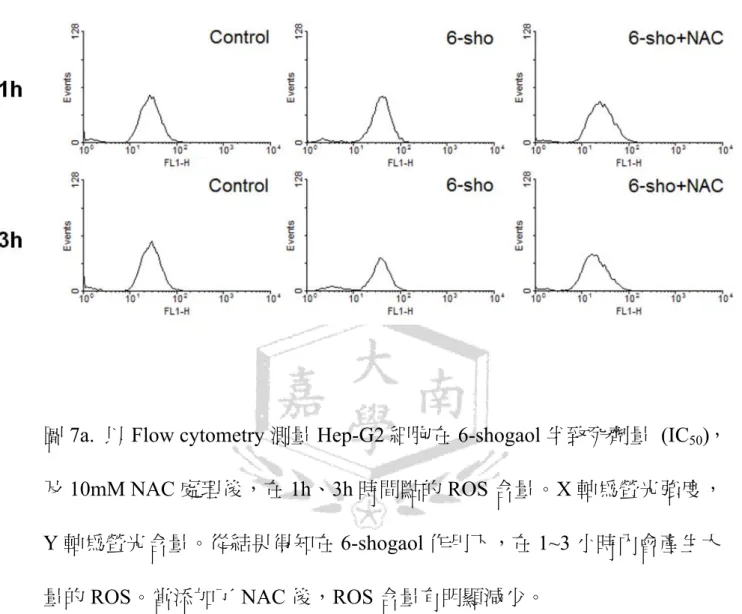
圖 7a. Hep-G2 細胞在 6-shogaol 和 NAC 處理後 ROS 含量...46
圖 7b. ROS 增加含量統計圖...47
圖 8a. Hep-G2 細胞在 6-shogaol & NAC 處理後細胞週期……...48
圖 8b. 6-shogaol & NAC 處理後細胞週期分佈…...49
圖 8c. 6-shogaol & NAC 處理後細胞週期統計表...50
圖 9. 6-shogaol 與 NAC 處理下 Hep-G2 細胞之存活率...51
圖 11a. Hep-G2 細胞在 6-shogaol 和 NAC 處理後蛋白質變化...53 圖 11b. 6-shogaol 誘導細胞凋亡信號的蛋白 GADD153 的變化...54
英文縮寫對照表 ER stress:Endoplasmic reticulum stress
PERK:protein kinase-like ER kinase ROS:Reactive oxygen species
GSH:Glutathione
P/S:penicillin & streptomycin FBS:fetal bovine serum
DMEM:Dulbecco’s modified Eagle’s medium PBS:Phosphate buffered saline
EDTA:Ethylenediaminetetraacetic acid
MTT:3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide ELISA:Enzyme-linked immunosorbent assay
PI:propidium iodide
DCFH-DA:2’, 7’- dichlorodihydrofluorescein - diacetate SDS:sodium dodecyl sulfate
BSA:Bovine serum albumin
SDS-PAGE:sodium dodecyl sulfate polyacrylamide gel electrophoresis TBST:Tris-Buffered Saline and Tween 20
NAC:N-Acetyl-Cysteine GSSG:Glutathione disulfide
NADPH:Nicotinamide adenine dinucleotide phosphate
第一章 緒論 1.1 肝癌
肝癌,是一種在全球常見的十大癌症之一(1)。根據研究觀察發現,在歐
洲地區較常造成肝癌的原因,多半是由 C 型肝炎病毒感染和肝硬化所造
成;在亞洲與非洲地區,大多數則和 B 型肝炎病毒較為有相關性(2)。在台 灣,根據行政院衛生署的統計,近幾年來「肝癌」一直蟬連全國十大癌症 死亡原因的前兩名中,台灣地區每年約有七千人因肝癌去世。
目前對於肝癌的治療方式,大致上有手術切除、肝動脈栓塞、局部酒 精注射治療、無線電頻燒灼術(Radiofrequency Ablation Therapy, RFA)、冷 凍治療等(3)。然而以上的方法不是完全適用在所有的肝癌患者身上,且也並 不能夠很完全根除肝癌細胞,防止復發的可能性,最終還是需藉由化學治 療。因為癌細胞會不斷的進行細胞分裂而增殖,其突變機率也較正常細胞 為高,再加上化療藥物在體內進行篩選,只要有一個經突變而產生抗藥性 的癌細胞存活,腫瘤又會再度復發。所以抗藥性的問題,是目前一個待解 決的問題。
1.2 薑
薑 (Zingiber officinale),是一種多年生草本科植物,原產於東南亞熱帶 區的植物,其根狀莖常被拿來食用。薑味道辛辣,中國人自古以來便常用
於調味或當做食材(4)。薑因含有一種名為薑辣醇 (gingerol),所以會有辛辣 且刺鼻的味道。從薑裡面萃取出來的化合物有 6-gingerol、8-gingerol、
10-gingerol,和 6-shogaol、8-shogaol、10-shogaol,其中屬 6-gingerol 最多。
6-gingerol 經過被脫水作用以後,便會形成 6-shogaol 的化合物(5)。
過去在治療癌症的研究發現,6-shogaol 會使人類肺癌細胞 A549 產生 自體吞噬,透過抑制 AKT/mTOR 這條路徑(6);在大腸癌方面,會透過產生 ROS (Rreactive oxygen species),caspase 的活化,以及 GADD153 的表現,
誘導癌細胞產生細胞凋亡(7)。6-shogaol 誘導細胞產生細胞凋亡,是因為 6-shogaol 進入細胞內在初期會產生大量的活性氧分子 (ROS),進一步消耗 掉了大量的穀胱甘肽 (GSH,glutathione),導致粒線體膜電位下降,活化了 caspase3/7,近而裂解了 DNA(8)。
1.3 活性氧分子 (Reactive oxygen species,ROS)
氧是生物體中一個很重要的的分子,可經由能量與電子的轉移,活化 為反應性較強的的氧分子稱為活性氧或自由基。自由基是一個帶有不成對 電子且獨立存在的物質,很容易去掠奪其他分子的電子,來使自己不穩定 的電子變的較穩固,在這過程中就會對一些中要的分子像核酸、蛋白質、
酵素等產生破壞。
在生物體中,氧分子會經由電子轉移而產生超氧陰離子,之後會形成
過氧化氫,接著轉變成羥基自由基,最後形成水。在這些過程中,就會產 生自由基,而身體裡面有一些可以清除這些自由基的酵素,例如過氧化氫 酶、穀胱甘肽過氧化酶 (GSH) 等等,會對這些自由基進行清除的動作,以 達到平衡狀態。但若這個平衡狀態被破壞以後,自由基過量的產生,便會 形成一種壓力,稱為氧化壓力 (oxidative stress)。
除了本身身體內會產生活性氧外,文獻上指出 6-shogaol 本身含有 guaiacol compound,因此本身可以和氧化劑反應形成 phenolic radical,當 phenolic radical 在細胞內獲得穀胱甘肽 (GSH) 上的氫原子時,則形成最初 6-shogaol 的結構(8)。這個作用會持續的消耗掉細胞內大量的 GSH,一旦 GSH 被大量消耗,便無法清除身體內過多的自由基,同樣也會使得氧化壓 力增加。另也有文獻指出,內質網的cytochrome P450,也是細胞內產生 ROS 酵素的系統之一(9)。
1.4 內質網壓力 (Endoplasmic reticulum stress,ER stress)
內質網是一個在蛋白質合成中,扮演很重要角色的一個胞器,蛋白質 需要在這裡被正確的摺疊組合後,才能被送到細胞其他的部位使用。在內 質網裡有許多的酵素和蛋白,能夠幫助蛋白質正確的摺疊合成,再送出內 質網;而摺疊不正確的蛋白質則會繼續留在內質網內,進行修復或分解。
假如內質網被一些內在或外在因子影響,如病毒感染、胺基酸缺乏、缺氧、
基因突變等因素作用下,會造成摺疊不完全的蛋白累積在內質網內,進而 產生內質網壓力(10)。
不完全摺疊蛋白會利用 IRE1 及 ATF6 轉錄因子啟動一些相關基因,幫 助蛋白質正確摺疊。PERK 會自我磷酸化而活化,再去磷酸化 eIF-2α,使轉
譯作用減緩,減少新的蛋白質合成。如果還有未摺疊的蛋白質,則會被送 回細胞質由蛋白水解酶分解。
這些反應可以用來減少過多的蛋白質累積在內質網內,對內質往造成 的壓力。倘若蛋白質累積的情況沒有改善,內質網壓力過大的時後,就會 誘導凋亡信號啟動,導致細胞走向細胞凋亡。
1.5 細胞凋亡
細胞凋亡 (Apoptosis) 也稱做為有程序性的細胞死亡 (Programmed cell death),是在 1972 年由澳大利亞病理學家 John Kerr 首先提出的。細胞 凋亡是一種細胞自發性進行細胞自殺的行為,當細胞可能由於外在環境的 不適合、接受到死亡的訊號、內部 DNA 受到損傷、內質網壓力等等,以發 一連串程序性的細胞死亡過程。
細胞在進行凋亡的過程,包含了細胞膜會皺縮,細胞膜內的磷脂絲胺 酸 (Phosphatidyl Serine, PS) 會外翻到膜外;粒線體膜電位會下降,釋放出 細胞色素 C (Cytochrome C),進而活化了 caspaase,將 DNA 裂解為 200
basse-pair 的倍數片段 (DNA fragmentation),這些 DNA 片段會和胞器被胞 膜包成凋亡小體 (apoptosis body),最後被巨噬細胞 (phagocyte) 吞噬清除。
1.6 研究動機
到目前為止,雖然治療肝細胞癌的方式有許多,但是其效果非常有限,
且無法徹底根除癌細胞,使癌化的細胞不會再轉移到其他組織器官中,最 終都需要靠化學治療的方式。但是化學治療往往容易造成抗藥性的問題,
所以我們希望從天然的傳統藥物中,找尋有效又安全的藥物用在治療癌 症。然而,有些從傳統中藥材中提煉出來的化合物,譬如從冬蟲夏草萃取 的 FTY720,已被證實能誘導癌細胞凋亡(11);另外,牛樟芝中所含的成分 Antroquinonol,則可以抑制癌細胞的增值(12);而從辣椒中萃取出來的物質,
辣椒素 (capsaicin),也同樣具有抑制癌細胞增長,導致細胞凋亡的作用(13,
14)。故推測一種化學結構式與辣椒素類似,從薑中萃取出來的物質 6-shogaol 同樣具有抑制癌細胞增長,導致細胞凋亡的作用。
根據先前研究發現,6-shogaol 在進入細胞後會誘導 ROS 大量的增加,
導致細胞凋亡。另一方面也有發現,6-shogaol 會誘發一種讓細胞生長停滯 和造成 DNA 損傷的蛋白質 (Growth arrest- and DNA damage-inducible gene 153,GADD153) 表現(7)。而許多研究指出,GADD153 為內質網壓力誘導 細胞凋亡一個重要的蛋白質(15~19)。但目前尚無對於6-shogaol 和內質網壓力
對於肝癌之影響的相關研究,所以本篇要探討6-shogaol 是否同樣會透過氧 化壓力誘導肝癌細胞走向凋亡機制,另一方面也會透過內質網壓力這條路 徑造成細胞走向細胞凋亡。
第二章 材料與方法 2.1 細胞培養
2.1.1 細胞培養條件
將人類的肝癌細胞株 Huh-7 和 Hep-G2 培養在 10 cm 培養皿,內含 1%
抗生素 (10000 units/ml penicillin & streptomycin,P/S) 及 10% fetal bovine serum (FBS) 的 Dulbecco’s modified Eagle’s medium (DMEM) 培養液,置於 環境條件為含 5% CO2和溫度37℃的細胞培養箱中。
2.1.2 細胞繼代培養
細胞培養至約 80% ~ 90%滿時,先去除舊的培養液,再以 1×PBS (Phosphate buffered saline) buffer 沖洗兩次,加入 2ml 0.05% trypsin / EDTA,
放置於 37℃的細胞培養箱中反應 2 分鐘。取出後輕拍培養皿邊緣,使細胞 自培養皿盤底脫落,再加入 2ml 新鮮的培養液中和 trypsin 的活性,收集細 胞至 15ml 無菌離心管中,以 1500 rpm 離心 5 分鐘。離心後去除上清液,
並加入新鮮培養液來回吸打將細胞均勻沖散,平均分配細胞懸浮液至數盤 新的培養皿中,置於 5% CO2和溫度37℃的細胞培養箱中培養。
2.1.3 計數細胞
根據上述繼代培養方式,將離心後的細胞以適當體積的培養液回溶,
的 trypan blue 均勻的混合,在各取 10μl 的混合液放入血球計數器中,放置 顯微鏡下計算血球計數器上下兩個九宮格中左上、右上、左下、右下等八 格的細胞數量。
計算方式:N (八格區域細胞數的總和) N/8 (每格區域的細胞數量)
_______________________________________ × 8 (稀釋八倍) = N × 104 cells/ml 0.1×0.1×0.1 (每格區域的體積)
2.1.4 冷凍細胞
根據上述繼代培養方式,將離心後的細胞以適當體積的培養液回溶,
加入總體積 10%的 DMSO (Dimethyl sulfoxide) 混合均勻,將細胞混合液平 均分裝至冷凍小管中,放入-80℃冰箱儲存。
2.1.5 解凍細胞
將冷凍小管從-80℃冰箱取出後迅速以 37℃回溫,移至 15ml 離心管中,
加入 2ml 新鮮培養液混合均勻,以 1500 rpm 離心 5 分鐘。去除上清液,加 入適量培養液使細胞懸浮,平均分配細胞懸浮液至新的培養皿中,置於5%
CO2和溫度37℃的細胞培養箱中培養。
MTT (3 - (4,5 - Dimethylthiazol - 2 - yl ) - 2,5 - diphenyltetrazolium bromide) 為一種黃色水溶性化合物,是一種接受氫離子的染料,可作用於 活 細 胞 粒 線 體 中 的 呼 吸 鏈 , 在 琥 珀 酸 脫 氫 酶 (SDH) 和 細 胞 色 素 C (Cytochrome C) 的作用下 tetrazolium 環裂開,生成藍紫色的 formazan 結晶,
formazan 結晶的生成量與活細胞數目成正比,因為死細胞中琥珀酸脫氫酶 會消失,不能將 MTT 還原。之後再用 DMSO (Dimethyl Sulfoxide) 將細胞 內的結晶溶出。利用 ELISA reader 波長 570 nm 去測其紫色溶液的吸光值,
依吸光值的強弱便可以知道粒線體的活性,進而可以推算出細胞的存活率。
將肝癌細胞分別培養在 96-well 的培養皿,每個 well 裡含有 4000 顆細
胞,以及100μl 的培養液。待細胞貼壁後,將培養液更換為含不同濃度藥物 的培養液,培養 24 小時。去除含藥物的培養液,加入以培養液配製稀釋的 MTT 溶液 (0.5mg/ml),每個 well 20μl,置於 37℃的細胞培養箱中培養 4 小 時。4 小時後,去除含 MTT 的培養液,每個 well 加入 100μl 的 DMSO。用 連續波長分析儀 (ELISA reader) 以波長 570 nm 測定其吸光值,統計其細胞 存活率。
2.3 細胞外型觀察
將 3×106顆的Hep-G2 細胞培養在 10 cm 培養皿,待 12-14 小時細胞貼 壁後,加入濃度50 (μg/ml) 的 6-shogaol 至培養皿內,在不同的培養時間點
2.4 細胞內 ROS 含量的分析 (Flow cytometry,流式細胞儀)
將 1×106顆的Hep-G2 細胞培養在 6 cm 培養皿,待 12-14 小時細胞貼壁 後,加入濃度 50 (μg/ml) 的 6-shogaol 至培養皿內。處理 1 小時與 3 小時後,
加入濃度為 10μM 2’, 7’- dichlorodihydrofluorescein – diacetate (DCFH-DA),
放回 37℃的培養箱反應 20 分鐘。DCFH-DA 本身會穿過細胞膜進入細胞 內,在細胞內和 ROS 發生反應。當 DCFH-DA 被 ROS 給氧化以及裂解成 DCF 的產物,在波長 488-530 nm 的光激發下,會產生綠色螢光。收集所有 的培養液至 15ml 無菌離心管中,以 1×PBS buffer 沖洗兩次,並將其沖洗液 收集至同管離心管中,加入 1ml 0.05% trypsin / EDTA,放置於 37℃的細胞 培養箱中反應30 秒。取出後拍擊培養皿邊緣,將細胞從培養皿盤底震落,
吸取一些 15ml 離心管中的培養液中和 trypsin 活性,將所有細胞液收集回離 心管中。以 1500 rpm 離心 5 分鐘,去除上清液,加入 1×PBS buffer 來回吸 打,在離心一次去除上清液。以 1ml PBS buffer 來回吸打,使細胞均勻分散,
放置在冰上避光。以flow cytometry 分析,觀察 ROS 的含量。
2.5 細胞週期的分析 (Flow cytometry,流式細胞儀)
將 3×106顆的Hep-G2 細胞培養在 10 cm 培養皿,待細胞貼壁後,去除 舊的培養液,以 1×PBS buffer 沖洗兩次,加入內含 1% P/S,以及 0.5% FBS
的 6-shogaol 至培養皿內,培養 12 小時並收集細胞。收集細胞時,將所有 的培養液收集至15ml 無菌離心管中,以 1×PBS buffer 沖洗兩次,並將其沖 洗液收集至同管離心管中,加入2ml 0.05% trypsin / EDTA,放置於 37℃的 細胞培養箱中反應 30 秒。取出後拍擊培養皿邊緣,將細胞從培養皿盤底震 落,吸取一些 15ml 離心管中的培養液中和 trypsin 活性,將所有細胞液收集 回離心管中。以 1500 rpm 離心 5 分鐘,用倒的去除上清液,並使其殘留一 些液體,用手拍擊離心管約 10 下震散細胞,加入 1×PBS buffer 來回吸打,
再離心一次,用倒的去除上清液,並使其殘留一些液體,用手拍擊離心管 約 10 下震散細胞。將離心管放置於震盪器上,以一滴一滴的方式,加入 1ml 70%的酒精,放置於 4℃冰箱至少 18 小時以上。以 1500 rpm 離心 5 分鐘,
用倒的去除上清液,並使其殘留一些液體,用手拍擊離心管約 10 下震散細 胞,加入PBS buffer 來回吸打沖散細胞使其均勻分散,加入濃度為 40μg/ml 的 propidium iodide (PI),放置室溫下避光反應 40 分鐘。PI 為一種螢光染劑,
細胞膜經過酒精固定後會產生孔洞,PI 便會透過膜上的洞進入細胞,結合 在 DNA 與 RNA 的鹼基對中間,在藉由 flow cytometry 以 493 nm ~ 630 nm 波長的光去激發,會產生紅色的螢光,透過分析螢光強弱便可以知道細胞 週期的變化。
2.6 Caspase-3 分析 (Flow cytometry,流式細胞儀)
將 1×106顆的Hep-G2 細胞培養在 6 cm 培養皿,待 12-14 小時細胞貼壁
後,加入濃度為0、10、25、50 (μg/ml) 的 6-shogaol 至培養皿內。培養 24 小時後,將所有的培養液收集至15ml 無菌離心管中,1×PBS buffer 沖洗兩 次,並將其沖洗液收集至同管離心管中,加入 2ml 0.05% trypsin / EDTA,
放置於 37℃的細胞培養箱中反應 30 秒。取出後拍擊培養皿邊緣,將細胞從 培養皿盤底震落,吸取一些 15ml 離心管中的培養液中和 trypsin 活性,將所 有細胞液收集回離心管中,以 1500 rpm 離心 5 分鐘。再用冷的 1×PBS buffer 沖洗兩次,離心並去除上清液。加入 0.5ml 的 BD cytofix/cytoperm slution 使細胞懸浮,放置冰上 20 分鐘。以 1500 rpm 離心 5 分鐘,去除 BD cytofix/cytoperm slution,在室溫下加入 0.5ml 1×BD Perm/wash buffer 清洗細 胞兩次,以1500 rpm 離心 5 分鐘去除清洗液。加入 100μl 1×BD Perm/wash buffer 懸浮細胞,並加入 20μl antibody,置室溫下 30 分鐘。以 1500 rpm 離
心 5 分鐘去除上清液,加入 1ml 1×BD Perm/wash buffer 清洗細胞,以 1500 rpm 離心 5 分鐘去除清洗液。加入 0.5ml 1×BD Perm/wash buffer 懸浮細胞,
並以 flow cytometry 分析,觀察 Caspase-3 的變化。
2.7 Apoptosis 分析 (Flow cytometry,流式細胞儀)
將 1×106顆的Hep-G2 細胞培養在 6 cm 培養皿,待 12-14 小時細胞貼壁 後,加入濃度為0、10、25、50 (μg/ml) 的 6-shogaol 至培養皿內。培養 24
小時後,將所有的培養液收集至15ml 無菌離心管中,1×PBS buffer 沖洗兩 次,並將其沖洗液收集至同管離心管中,加入 2ml 0.05% trypsin / EDTA,
放置於 37℃的細胞培養箱中反應 30 秒。取出後拍擊培養皿邊緣,將細胞從 培養皿盤底震落,吸取一些 15ml 離心管中的培養液中和 trypsin 活性,將所 有細胞液收集回離心管中,以 1500 rpm 離心 5 分鐘。再用冷的 1×PBS buffer 沖洗兩次,離心並去除上清液。以 1ml 1X binding buffer 使細胞懸浮,加入 體積各為 5μl 的 FITC annexin V 和 PI 至懸浮液中,輕輕震盪混合均勻,放 置反應 15 分鐘。反應完畢後在 1 小時內,以 flow cytometry 分析,觀察 apoptosis 的變化。
Annexin V 是一種會與磷脂絲胺酸 (Phosphatidyl) 做結合的螢光染 劑。細胞在走向凋亡的初期,細胞膜會開始外翻,原本只在細胞膜內的磷 脂絲胺酸會外翻到膜外。之後細胞膜會開始出現孔洞,此時PI 便會透過這 些孔洞進入細胞內與 DNA 結合。透過 flow cytometry 分析 Annexin V 與 PI 的螢光強度,便可了解 apoptosis 的程度。
2.8 西方墨點法
2.8.1 細胞內蛋白質萃取
將 3×106顆的Hep-G2 細胞培養在 10 cm 培養皿,待細胞貼壁後,加入 濃度為 50(μg/ml)的 6-shogaol 至培養皿內,在不同的培養時間點 0、3、6、
12、24 小時收集細胞。用刮棒將貼壁的細胞刮起,連同培養液收集至 15ml 離心管中,以 1500 rpm 離心 5 分鐘,去除上清液,加入 1×PBS buffer 來回 吸打,並將溶液移至eppendrof 中,以 1500 rpm 離心 5 分鐘,去除上清液。
每個樣品加入 200μl SDS lysis buffer,並充分的來回吸打,待溶液從黏稠狀 吸打至液體狀,以12000 rpm 離心 10 分鐘去除泡沫即可,其溶液便為細胞 總蛋白液。
2.8.2 蛋白質定量
將標準品 BSA 配至成濃度為 0、0.4、0.8、1.6、2 (mg/ml),備置在 eppendrof 中,體積為 150μl。預測的蛋白質取 5μl,與 145μl 的 ddH2O 混和,備置在 eppendrof 中。配製 protein assay reagent,並加進標準品與待測樣品的 eppendrof 中混合,每管加入 150μl,放進 60℃烘箱反應 1 小時。將每個標
準品與待測樣品各取 100μl 放置於 96-well 培養皿中,每孔 100μl,用連續 波長分析儀 (ELISA reader) 以波長 595 nm 測定其吸光值。以標準品的曲線 求出其斜率公式,將樣品吸光值代入公式中,計算出各樣品的蛋白質濃度。
2.8.3 蛋白質電泳
將蛋白萃取液取同濃度最大體積的量,用 6×loading dye 將體積補滿至 200μl,放進煮沸的水中加熱 5 分鐘,再放在冰上降溫待測。配置好 10% SDS
泳 100 分鐘。
2.8.4 蛋白質轉漬
準備 6 張 3M paper 和 1 張 transfer membrane,將 3M paper 浸泡在 transfer buffer 備用,transfer membrane 浸泡在甲醇 (methanol) 備用。電泳結束後,
由上至下以 3 張 3M paper、gel、transfer membrane、3 張 3M paper 的順序,
疊好放置在半乾式碳板上,用小圓棒將空氣滾動排除,再以條件為 100 毫 安培 (mA) 進行轉漬 100 分鐘。
2.8.5 免疫墨點法
轉漬完成後,將 transfer membrane 依照想觀察的蛋白質分子量的不同,
切割剪裁不同位置並放到blocking buffer 中,並放置在 shaker 上搖晃 1 個半 小時。用 TBST 清洗 3 次,每次 10 分鐘,再依照蛋白質分子量位置加入 1 級抗體,放置在 4℃ shaker 上緩慢搖晃 overnight。用 TBST 清洗 3 次,每次 10 分鐘,再依照 1 級抗體萃取來源加入 2 級抗體,放置在室溫 shaker 上緩 慢搖晃 1 個半小時。用 TBST 清洗 3 次,每次 10 分鐘,最後以冷光試劑 (Western Chemiluminescent HRP substrate) 作用,再以冷光螢光影像分析系 統 (LAS-3000) 進行分析。
第三章 結果 3.1 6-shogaol 對肝癌細胞存活率的影響
將兩株不同之人類肝癌細胞 Huh-7 和 Hep-G2 經過濃度 0、5、10、25、
50、75、100、200 (μg/ml) 的 6-shogaol 處理 24 小時後,用 MTT 分析抑制 細胞生長的能力,以酵素免疫分析儀 (ELISA Reader) 偵測波長 570nm 吸收 峰的吸光值,吸光值越高則表示存活的細胞數量越多。實驗後發現,Huh-7 和 Hep-G2 在 6-shogaol 的刺激下,隨著處理的濃度增加,細胞的存活率與 沒處理藥物的對照組細胞相比,有逐漸下降的趨勢。由此可以證實,
6-shogaol 對於不同的肝癌細胞,同樣都具有抑制細胞生長及毒殺的作用,
並具有 does-dependent 的效果。而且從結果圖上明顯看的出來,Hep-G2 細 胞對於 6-shogaol 抑制細胞生長的效果,來的比 Huh-7 靈敏許多。另外可以 從 MTT assay 的結果中得到,Hep-G2 對於 6-shogaol 的半致死劑量 (LD50),
約為 50 (μg/ml) (圖 1a、1b)。
3.2 6-shogaol 對 Hep-G2 細胞外型的變化
培養中的癌細胞在顯微鏡下觀察,會呈現不規則、多觸角型態貼壁生 長。當癌細胞受到外在環境因素影響 (如藥物治療,細菌、黴菌汙染),細 胞走向凋亡的現象 (Apoptosis),觸角逐漸萎縮,最後癌細胞無法貼壁,而 漂浮在培養液中。
利用顯微鏡觀察經過 6-shogaol 處理後的 Hep-G2 細胞生長的情況,發
現經過 6-shogaol 50 μg/ml 處理後的細胞,細胞型態有逐漸產生皺縮的現 象,隨著6-shogaol 處理的時間逐漸增加,細胞皺縮、死亡的情形更加的明 顯,並同樣具有time-dependent 的效果 (圖 2)。
3.3 6-shogaol 對 Hep-G2 細胞內活性氧化物 (ROS) 的影響
由先前的 MTT assay 的實驗已知,6-shogaol 可以有效的抑制肝癌細胞 的存活率,而先前的研究指出在其他癌細胞中,6-shogaol 在進入細胞後會 大量誘導 ROS 的產生造成細胞死亡,因此我們進一步同樣也分析 6-shogaol 是不是在 Hep-G2 中,同樣也會誘導 ROS 的含量上升。
我們利用 DCFH-DA 會被 ROS 給氧化以及裂解成 DCF,在波長 488-530 nm 光激發下會產生綠色螢光的特性,利用流式細胞儀分析 ROS 含 量的情況。Hep-G2 細胞經過 50μg/ml 6-shogaol 處理 1 小時與 3 小時後,加
入 DCFH-DA 染色,在利用流式細胞儀分析,結果顯示當 Hep-G2 細胞處理 過 6-shogaol 和沒處理的對照組相比,ROS 的含量有明顯的增加,約增加了 三倍之多 (圖 3a、3b)。
3.4 6-shogaol 對 Hep-G2 細胞週期的影響
ROS 是一種活性極高的分子,因為它帶有未成對的電子,所以容易
damage,進而可能會改變 DNA 正常的複製週期。所以接著就來看看 Hep-G2 的細胞週期,受否受到6-shogaol 所誘導的 ROS 產生改變。
正常的細胞大多數會是停留在染色體未複製的單倍體時期 (G1),部 分細胞會在染色體複製期 (S),與染色體複製成二倍體時期 (G2)。當 DNA 發生 damage 時,DNA 無法正常的複製時,細胞週期便會停留在 Sub-G1 時 期。Hep-G2 細胞經過 0、10、25、50 (μg/ml) 6-shogaol 處理 12 小時後,收
集固定細胞,以PI 染色,經流式細胞儀分析發現,隨著 6-shogaol 濃度的上 升,其 Sub-G1 的細胞比例也跟著增加。根據數據量化圖得知,Sub-G1 時 期的細胞在濃度 25μg/ml 顯著的增加 13%左右;當濃度提高到 50μg/ml 時,
有近 90%的細胞都停留在 Sub-G1 時期 (圖 4a、4b、4c)。
3.5 6-shogaol 對 Caspase-3 的影響
從細胞週期得知,6-shogaol 會改變 Hep-G2 的細胞週期,隨著濃度增 加 Sub-G1 的比例也隨之增高。當 Sub-G1 增加,細胞便會開始誘導細胞走 向凋亡的路徑,一些凋亡因子便會開始啟動。Caspase-3 是一種凋亡蛋白,
當凋亡信號開始啟動時,Caspase-3 便會被活化。因以進一步我們要分析,
當 Sub-G1 的比例增加時,是否會啟動凋亡因子而誘導 Caspase-3 含量上升。
將 Hep-G2 細胞經過 0、10、25、50 (μg/ml) 6-shogaol 處理 24 小時後,以會 激發螢光的 antibody 染色,由流式細胞儀分析發現,隨著 6-shogaol 的濃度
增加Caspase-3 的含量也跟著增加,顯示細胞開始走向細胞凋亡 (圖 5a、5b)。
3.6 6-shogaol 誘導 Hep-G2 細胞凋亡
由先前 Caspase-3 的實驗已知,6-shogaol 會誘導細胞凋亡的訊號啟動,
導致細胞走向細胞凋亡。當細胞開始進行細胞凋亡時,原本只存在內膜細 胞膜上的磷脂絲胺酸會外翻到細胞膜外,而外翻的磷脂絲胺酸會被 Annexin V 辨識並結合。細胞凋亡到最後細胞會壞死,壞死的細胞細胞膜會出現孔 洞,此時染劑 PI 便會透過細胞膜上的孔洞進入細胞內與 DNA 做結合。利 用 Annexin V 與 PI 的螢光強度的搭配,可以用來區分細胞凋亡的程度。
我們以 0、10、25、50 (μg/ml) 6-shogaol 處理 Hep-G2 細胞 24 小時後,
以 Annexin V 與 PI 雙染法染色,經流式細胞儀偵測,可以明顯觀察到 6-shogaol 濃度增加時,細胞膜外翻的情形更加明顯,且有少部分的細胞壞 死的現象。經量化後結果顯示,在濃度 50μg/ml 時,有 50%的細胞開始走 向細胞凋亡 (圖 6a、6b)。
3.7 抑制 ROS 對 6-shogaol 誘導細胞死亡的影響
由前面的實驗得知了,6-shogaol 會使 Hep-G2 細胞產生大量 ROS,進 而改變了細胞週期,誘發凋亡信號啟動,導致細胞凋亡。接著我們想了解,
當ROS 被抑制後,是否能夠保護細胞受到 6-shogaol 造成的細胞損傷。利用
經流式細胞儀進行 ROS 量的分析後,從結果圖上可以看到,加入 6-shogaol 的 Hep-G2 細胞,ROS 的含量與沒添加的對照組相比,ROS 含量 確實有增加;當加入了 NAC 後,ROS 的含量明顯的下降了許多。經量化後 結果顯示,加入 NAC 在 3 小時的時後,ROS 的比例下降了一半以上,證實 NAC 確實能抑制 6-shogaol 誘導產生的 ROS (圖 7a、7b)。
透過細胞週期分析結果圖觀察到,抑制 ROS 確實能保護細胞,使細胞 週期不受影響。透過統計分析發現,添加 6-shogaol 的 Hep-G2 細胞 Sub-G1 期的細胞明顯的增加約四成;有加入NAC 抑制 ROS 的 Hep-G2 細胞,細胞 週期比例和為加藥的對照組相似,顯示細胞周期不受影響 (圖 8a、8b、8c)。
接著以 MTT assay 分析發現,加入了 50 μg/ml 6-shogaol 的細胞存活率
與未添加相比,細胞存活率有下降。但加入了 NAC 後發現,隨 NAC 的濃 度上升至 10mg/ml,細胞存活率有很明顯的上升至 80%左右,顯示 NAC 有 保護細胞,防止細胞受到6-shogaol 的毒殺 (圖 9)。
3.8 6-shogaol 誘導內質網壓力造成 Hep-G2 細胞凋亡
根據上述 MTT assay 結果圖我們觀察到,NAC 確實有保護細胞加細胞 的存活率,但仍然無法回復細胞的存活率,仍有少部分的細胞死亡,顯示 可能 6-shogaol 會透過其他路徑造成細胞死亡。
利用西方墨點法分析,經濃度 50 μg/ml 6-shogaol 處理 0、3、6、12、
24 小時的細胞中萃取出來的蛋白質後發現,一些產生內質網壓力時,會誘 導增加的蛋白質如:IRE1α、IRE1β、ATF6α、GRP78、p-eIF2-α等等,隨 著處理的時間增加,都有明顯的增加的現象 (圖 10)。表示 6-shogaol 會造成 Hep-G2 細胞產生內質網壓力。
接著在加入 ROS 抑制劑 NAC 後,內質網壓力會誘導的蛋白質能隨著 處理時間的增加,仍有明顯得增加的趨勢 (圖 11a)。顯示 6-shogaol 除了 ROS 誘導的細胞凋亡外,可能同樣會透過內質網壓力造成細胞死亡。
在分析內質網壓力誘發凋亡信號的同源染色體 GADD153 後,從結果很 明顯看的出來 6-shogaol 誘導的內質網壓力會促使 GADD153 的表現量增 加,誘發細胞凋亡。抑制ROS 後,GADD153 表現量仍會增加,顯示 Hep-G2 細胞仍會透過氧化壓力誘導細胞凋亡 (圖 11b)。
第四章 討論
薑 (Zingiber officinale),在中國一直以來都是不可缺少的調味料與食 材。從薑中可以萃取出來的成分許多,近年來越來越多研究發現,從薑中 萃取出來的成分,許多都具有治療癌症抑制癌細胞生長,造成細胞凋亡的
作用(20~34)。這些成分中,以6-gingerol 和 6-shogaol 最多,而 6-shogaol 的效
果又來的比 6-gingerol 好。有研究指出,6-shogaol 會抑制癌細胞生長活性,
誘發一些凋亡信號的啟動,導致細胞走向凋亡的路徑(35)。而從我們的實驗 結果得知在 6-shogaol 的作用下,人類肝癌細胞 Hep-G2 會在 1 到 3 小時內 產生大量的 ROS。進一步的細胞週期受到影響,使細胞週期停滯在 Sub-G1 時期。當 Sub-G1 時期增加時,會導致凋亡蛋白 Caspase-3 活化,造成細胞 凋亡;另一方面,6-shogaol 也會產生內質網壓力,造成細胞凋亡。
過去的文獻指出,薑的萃取物造成細胞凋亡的原因,許多都和 ROS 的
生成有關(36~41)。6-shogaol 在進入細胞內後,會藉由消耗掉大量的抗氧化因
子穀胱甘肽,而使 ROS 無法被清除而累積。從實驗結果發現,Hep-G2 細 胞受到6-shogaol 影響,短時間內會產生二到三倍的 ROS。ROS 是一種本身 帶有未成對的電子的活性氧分子自由基,容易使 DNA 發生氧化性損傷 (Oxidative),造成 DNA damage 導致序列發生改變(41~44)。DNA damage 會造 成細胞週期的停滯,甚至造成細胞凋亡(45, 46)。
Caspase-3 是一個經常被激活的蛋白酶,具有催化裂解 DNA 的關鍵(47)。DNA 會被 Caspase-3 裂解成 180-200bp 的核小體,核小體會和其他細胞內的胞器 被胞膜包成凋亡小體 (Apoptosis body),接著被巨噬細胞清除。
細胞凋亡的初期,細胞膜上的磷脂會部分遺失開始呈現不對稱狀,而 原本只在細胞內膜的磷脂絲胺酸便會開始外翻到細胞外膜。Annexin V 是一 種可以用來標記磷脂絲胺酸的一種螢光染劑,透過流式細胞儀的分析可以 檢測細胞膜外翻的程度。當細胞凋亡程度增加,磷脂遺失的更多時,細胞 膜便會出現孔洞,此時DNA 螢光染劑可以透過這些孔洞進入細胞內與 DNA 做結合。
NAC 是含硫胺基酸 cysteine (半胱胺酸) 的前驅物,而半胱胺酸在體 內會參與合成穀胱甘肽 (glutathione)。穀胱甘肽在體內會以兩種形態存在在 體內,一是還原態的GSH,與氧化態 GSSG。還原態的 GSH 是身體內非常 重要的抗氧化劑,用來清除體內活性高的自由基。當 GSH 與自由基反應後,
會將自由基還原為較穩定的還原態,而 GSH 則氧化為氧化態的 GSSG。氧 化態的 GSSG,則會再藉由 NADPH 的作用,將其還原成還原態的 GSH,
以維持身體內的平衡。由於 6-shogaol 的作用產生了大量的 ROS 快速消耗了 細胞內的 GSH,而細胞內的 NADPH 還原 GSH 的速率有限,所以 ROS 無 法有效清除,對細胞造成氧化壓力。
力的蛋白 GRP78 和抑制生長造成 DNA 損傷基因 GADD153 的表現(48)。內 質網是細胞內摺疊蛋白質的地方,蛋白質會在內質網中正確的折疊再送出 內質網;而折疊不正確的蛋白質得會繼續留在內質網內進行修復或分解
(49)。當內質網受到一些外在環境因素影響,蛋白質發生不正確的摺疊,這
些折疊不正確的蛋白質過度的累積在內質網內時,會對內質網產生壓力。
而內質網壓力產生時會藉由 IRE1 及 ATF6 這兩個轉錄因子啟動,傳遞訊息 進入細胞核製造 GRP78 這個增加摺疊蛋白質能力的蛋白,來加強內質網摺 疊蛋白質的能力。所以氧化壓力既然誘導GRP78 的產生,這顯示 ROS 會影 響內質網折疊蛋白質的過程,使蛋白質無法正常摺疊而過度的累積,造成 內質網壓力的產生,才會誘導 GRP78 的表現來加強內質網摺疊蛋白質。由 此得知,ROS 是一種誘發內質網壓力的一個上游的調控因子。
然而又有許多的研究證據顯示,內質網在蛋白質折疊的過程中,所產 生的蛋白質氧化與氧化壓力的產生是具有相關性的(50)。當蛋白質在折疊過 程中,在粗糙的內質網內會行成雙硫鍵並進一步的進行修飾。雙硫鍵是由 兩個硫醇基透過氧化反應形成的,每形成一個雙硫鍵便會消耗一分子氧 氣,產生一分子的過氧化氫 (R-SH + SH-R + O2→ R-S-S-R + H2O2)。假如蛋 白質不正常的折疊,過多雙硫鍵的形成,會產生大量的副產物過氧化氫這 個活性氧分子,而造成了氧化壓力。
在本實驗中發現,以 NAC 去減少 ROS 的產生而抑制氧化壓力時,GRP78 和 IRE1、ATF6 等蛋白質仍然會隨時間表現,代表內質網壓力的現象仍然 是存在。這表示在 Hep-G2 細胞中,6-shogaol 所造成的氧化壓力並不會調 控內質網壓力的表現。所以依目前的結果可得知,6-shogaol 會同時使 Hep-G2 細胞產生內質網壓力與氧化壓力這兩條路徑造成細胞凋亡。
然而內質網壓力產生的氧化物,是否會加強氧化壓力所造成的細胞凋 亡仍值得進一步的做探討。未來可利用內質網壓力抑制劑 salubrinal,或是 siRNA 的方式抑制內質網壓力後,再分析細胞內 ROS 的含量是否有受到影 響,以證實 6-shogaol 是否會透過內質網壓力的路徑加強氧化壓力,造成 Hep-G2 細胞的細胞凋亡。
第五章 結論
綜合以上實驗結果得知,6-shogaol 能夠對人類肝癌細胞 Huh-7 和 Hep-G2 產生毒殺作用,有效的減少肝癌細胞的存活率,其原因是因為破壞 了細胞內 ROS 的平衡狀態。過多高活性的 ROS 的生成,影響了 Hep-G2 細 胞正常的細胞週期,而導致細胞凋亡的機制啟動。使用 ROS 的抑制劑 NAC 後,發現 Hep-G2 細胞存活率確實有回復的現象,但仍有部分死亡的細胞,
顯示 ROS 不是 6-shogaol 造成細胞毒殺的唯一途徑。
透過蛋白質的分析得知,6-shogaol 會誘導 Hep-G2 細胞產生內質網壓 力。內質網壓力是由於不正常摺疊蛋白的累積,而內質網又無法有效清除 這些蛋白質時,同樣也會誘導細胞產生凋亡的現象。在加入ROS 的抑制劑 NAC 後,並未改善內質網壓力的存在,顯示內質網壓力也是 6-shogaol 造成 細胞凋亡的機制之一。
所以本實驗證實了,6-shogaol 會使 ROS 大量產生,進而影響了 Hep-G2 細胞正常的細胞週期,誘導細胞凋亡。另一方面,同樣會造成不正常摺疊 蛋白質的累積,造成了內質網壓力,導致細胞凋亡。在兩者同時誘導細胞 凋亡的機制啟動,能夠有效的造成人類肝癌細胞Hep-G2 的死亡。
未來我們會利用 siRNA 的方式,抑制 6-shogaol 造成的內質網壓力,同 時在添加 NAC 抑制 ROS,看是否能夠回復九成以上甚至更高的細胞存活
參考文獻
1. Guan Ys, La Z, Yang L, He Q, Li P. p53 gene in treatment of hepatic carcinoma: Status quo.World J Gastroenterol. 2007;13:985-992.
2. Czauderna P, Perilongo G. Hepatocellular Carcinoma. Orphanet Encyclopedia. 2004.
3. Bruix J, Sherman M. Management of Hepatocellular Carcinoma. Aasld Practice Guideline. 2005;42:1208-1236.
4. Anonymous. Zingiber officinale (ginger). Alternative Med Rev. 2003;
8:331–335.
5. Lantz RC, Chen GJ, Sarihan M, Solyom AM, Jolad SD, Timmermann BN.
The effect of extracts from ginger rhizome on inflammatory mediator production. Phytomedicine. 2007;14:123-128.
6. Hung JY, Hsu YL, Li CT, et al. 6-Shogaol, an active constituent of dietary ginger, induces autophagy by inhibiting the AKT/mTOR pathway in human non-small cell lung cancer A549 cells. Agric Food Chem.
2009;57:9809-9816.
7. Pan MH, Hsieh MC, Kuo JM, et al. 6-Shogaol induces apoptosis in human colorectal carcinoma cells via ROS production, caspase activation, and
8. Chen CY, Liu TZ, Liu YW, et al. 6-Shogaol (Alkanone from Ginger) Induces Apoptotic Cell Death of Human Hepatoma p53 Mutant Mahlavu Subline via an Oxidative Stress-Mediated Caspase-Dependent Mechanism. J Agric Food Chem. 2007;55:948-954.
9. Curtin JF, Donovan M, Cotter TG. Regulation and measurement of oxidative stress in apoptosis. J Immunol Methods. 2002;265:49-72.
10. Ryu EJ, Harding HP, Angelastro JM, Vitolo OV, Ron D, Greene LA.
Endoplasmic reticulum stress and the unfolded protein response in cellular models of Parkinson's disease. J Neurosci. 2002;22:10690-10698.
11. Hung JH, Lu YS, Wang YC, et al. FTY720 Induces Apoptosis in Hepatocellular Carcinoma Cells through Activation of Protein Kinase C d Signaling. Cancer Res. 2008;15: 1204-1212.
12. Kumar VB, Yuan TC, Liou JW, Yang CJ, Sung PJ, Weng CF.
Antroquinonol inhibits NSCLC proliferation by altering PI3K/mTOR proteins and miRNA expression profiles. Mutat Res. 2011;707:42-52.
13. Wang HM, Chuang SM, Su YC, Li YH, Chueh PJ. Down-Regulation of Tumor-Associated NADH Oxidase, tNOX (ENOX2), Enhances Capsaicin-Induced Inhibition of Gastric Cancer Cell Growth. Cell Biochem
14. Pramanik KC, Boreddy SR, Srivastava SK. Role of mitochondrial electron transport chain complexes in capsaicin mediated oxidative stress leading to apoptosis in pancreatic cancer cells. PLoS One. 2011;6:e20151.
15. Oyadomari S, Mori M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004;11:381-389.
16. Chhabra R, Dubey R, Saini N. Gene expression profiling indicate role of ER stress in miR-23a~27a~24-2 cluster induced apoptosis in HEK293T cells. RNA Biol. 2011;8.
17. Srinivasan K, Sharma SS. Edaravone offers Neuroprotection in a Diabetic Stroke Model via Inhibition of Endoplasmic Reticulum Stress. Basic Clin Pharmacol Toxicol. 2011:1742-7843.
18. Moriya S, Miyazawa K, Kawaguchi T, Che XF, Tomoda A. Involvement of endoplasmic reticulum stress-mediated CHOP (GADD153) induction in the cytotoxicity of 2-aminophenoxazine-3-one in cancer cells. Int J Oncol. 2011.
19. Quaglio E, Restelli E, Garofoli A, et al. Expression of mutant or cytosolic PrP in transgenic mice and cells is not associated with endoplasmic reticulum stress or proteasome dysfunction. PLoS One. 2011;6:e19339.
20. Lee C, Park GH, Kim CY, Jang JH. [6]-Gingerol attenuates
defense system. Food Chem Toxicol. 2011;49:1261-1269.
21. Butt MS, Sultan MT. Ginger and its health claims: molecular aspects. Crit Rev Food Sci Nutr. 2011;51:383-393.
22. Choudhury D, Das A, Bhattacharya A, Chakrabarti G. Aqueous extract of ginger shows antiproliferative activity through disruption of microtubule network of cancer cells. Food Chem Toxicol. 2010;48:2872-2880.
23. Oyagbemi AA, Saba AB, Azeez OI. Molecular targets of [6]-gingerol: Its potential roles in cancer chemoprevention. Biofactors. 2010;36:169-178.
24. Hsu YL, Chen CY, Hou MF, et al. 6-Dehydrogingerdione, an active constituent of dietary ginger, induces cell cycle arrest and apoptosis through reactive oxygen species/c-Jun N-terminal kinase pathways in human breast cancer cells. Mol Nutr Food Res. 2010;54:1307-1317.
25. Nigam N, George J, Srivastava S, et al. Induction of apoptosis by [6]-gingerol associated with the modulation of p53 and involvement of mitochondrial signaling pathway in B[a]P-induced mouse skin tumorigenesis.
Cancer Chemother Pharmacol. 2010;65:687-696.
26. Kundu JK, Na HK, Surh YJ. Ginger-derived phenolic substances with cancer preventive and therapeutic potential. Forum Nutr. 2009;61:182-192.
Zerumbone, a tropical ginger sesquiterpene, inhibits colon and lung carcinogenesis in mice. Int J Cancer. 2009;124:264-271.
28. Lee SH, Cekanova M, Baek SJ. Multiple mechanisms are involved in 6-gingerol-induced cell growth arrest and apoptosis in human colorectal cancer cells. Mol Carcinog. 2008;47:197-208.
29. Shukla Y, Prasad S, Tripathi C, Singh M, George J, Kalra N. In vitro and in vivo modulation of testosterone mediated alterations in apoptosis related proteins by [6]-gingerol. Mol Nutr Food Res. 2007;51:1492-1502.
30. Park YJ, Wen J, Bang S, Park SW, Song SY. [6]-Gingerol induces cell cycle arrest and cell death of mutant p53-expressing pancreatic cancer cells. Yonsei Med J. 2006;47:688-697.
31. Ihlaseh SM, de Oliveira ML, Teràn E, de Camargo JL, Barbisan LF.
Chemopreventive property of dietary ginger in rat urinary bladder chemical carcinogenesis. World J Urol. 2006;24:591-596.
32. Bidinotto LT, Spinardi-Barbisan AL, Rocha NS, Salvadori DM, Barbisan LF.
Effects of ginger (Zingiber officinale Roscoe) on DNA damage and development of urothelial tumors in a mouse bladder carcinogenesis model.
Environ Mol Mutagen. 2006;47:624-630.
of diarylheptanoids and gingerol-related compounds from the rhizome of Chinese ginger. J Ethnopharmacol. 2005;102:177-184.
34. Murakami A, Tanaka T, Lee JY, et al. Zerumbone, a sesquiterpene in subtropical ginger, suppresses skin tumor initiation and promotion stages in ICR mice. Int J Cancer. 2004;110:481-490.
35. Ishiguro K, Ando T, Maeda O, et al. Ginger ingredients reduce viability of gastric cancer cells via distinct mechanisms. Biochem Biophys Res Commun. 2007;362:218-223.
36. Shieh PC, Chen YO, Kuo DH, et al. Induction of apoptosis by [8]-shogaol via reactive oxygen species generation, glutathione depletion, and caspase activation in human leukemia cells. J Agric Food Chem. 2010;58:3847-3854.
37. Zeng HL, Han XA, Gu C, et al. Reactive oxygen species and mitochondrial membrane potential changes in leukemia cells during 6-gingerol inducedapoptosis. Zhong Yao Cai. 2010;33:584-587.
38. Taha MM, Abdul AB, Abdullah R, Ibrahim TA, Abdelwahab SI, Mohan S.
Potential chemoprevention of diethylnitrosamine-initiated and 2-acetylaminofluorene-promoted hepatocarcinogenesis by zerumbone from
Interact. 2010;186:295-305.
39. Nigam N, Bhui K, Prasad S, George J, Shukla Y. [6]-Gingerol induces reactive oxygen species regulated mitochondrial cell death pathway in human epidermoid carcinoma A431 cells. Chem Biol Interact. 2009;181:77-84.
40. Yagihashi S, Miura Y, Yagasaki K. Inhibitory effect of gingerol on the proliferation and invasion of hepatoma cells in culture.
Cytotechnology. 2008;57:129-136.
41. Vijaya Padma V, Arul Diana Christie S, Ramkuma KM. Induction of apoptosis by ginger in HEp-2 cell line is mediated by reactive oxygen species. Basic Clin Pharmacol Toxicol. 2007;100:302-307.
42. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Oxidative DNA damage: mechanisms, mutation, and disease. FASEB J. 2003 ;17:1195-214.
43. Rowe LA, Degtyareva N, Doetsch PW. DNA damage-induced reactive oxygen species (ROS) stress response in Saccharomyces cerevisiae. Free Radic Biol Med. 2008;45:1167-1177.
44. Bennett MR. Reactive Oxygen Species and Death, Oxidative DNA Damage in Atherosclerosis. Circulation Research. 2001;88:648.
45. Bartkova J, Horejsí Z, Koed K, et al. DNA damage response as a candidate
46. Zhou BB, Elledge SJ. The DNA damage response: putting checkpoints in perspective. Nature. 2000;408:433-439.
47. Porter AG, Jänicke RU. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999;6:99-104.
48. He S, Yaung J, Kim YH, Barron E, Ryan SJ, Hinton DR. Endoplasmic reticulum stress induced by oxidative stress in retinal pigment epithelial cells.
Graefes Arch Clin Exp Ophthalmol. 2008;246:677-83.
49. Kleizen B, Braakman I. Protein folding and quality control in the endoplasmic reticulum.
Current Opinion in Cell Biology. 2004;16:343–349.
50. Malhotra JD, Kaufman RJ. Endoplasmic reticulum stress and oxidative stress:
a vicious cycle or a double-edged sword? Antioxid Redox Signal.
2007;9:2277-2293.
a.
Huh-7
b.
圖 1. 以 MTT 試驗測試在藥物 6-shogaol 處理下不同肝癌細胞之存活率。X 軸為 6-shogao 的濃度,Y 軸為細胞存活率百分比。圖 A 為 Huh-7 肝癌細胞,
圖 B 為 Hep-G2 肝癌細胞。從結果得知,6-shogaol 在不同肝癌細胞下,同 樣都有抑制肝癌細胞的生長效果。*為與未加藥對照組相比,*p<0.05,** p
100 80 60 40 20 0
10 25 50 75 100
0 5
200 120
***
*
***
*** ***
***
6-shogaolμM
Cell viability %
Hep-G2
0 5 10 25 50 75 100
200 0
20 40 60 80 100 120
** ***
***
***
*** *** ***
6-shogaolμM
Cell viability %
圖 2. 以顯微鏡觀察 Hep-G2 細胞在 6-shogaol 半致死劑量 (IC50) 處理後,
細胞外形的變化 。從結果得知,Hep-G2 細胞隨著藥物處理時間的增加,
細胞有萎縮死亡的現象。
a.
圖 3a. 以 Flow cytometry 測量 Hep-G2 細胞在 6-shogaol 半致死劑量 (IC50) 處理後,在1h、3h 時間點的 ROS 含量。X 軸為螢光強度,Y 軸為螢光含量。
從結果得知在 6-shogaol 作用下,在 1~3 小時內會產生大量的 ROS。
b.
1h
圖 3b. ROS 增加含量統計圖,1h 與 3h 。結果顯示,處理 6-shogaol 的細胞 內在 1h 與 3h,ROS 含量增加約三倍。*為與未加藥對照組相比,***p<
0.0001。
3h
control μM
6-shogaol 50 0
1 2 3
***
ROS fold
2
control μM
6-shogaol 50 0
1 3
***
ROS fold
a.
圖 4a. Hep-G2 細胞以濃度為 0、10、25、50μM 的 6-shogaol 處理下, 12h 後經由 Flow cytometry 分析細胞週期。X 軸為螢光強度,Y 軸為螢光含量。
從結果得知,隨著 6-shogaol 的濃度增加,細胞週期產生改變,Sub-G1 的含 量隨6-shogaol 濃度增加而上升, 顯示 DNA 受到損傷。
b.
圖 4b. 6-shogaol 各濃度細胞週期分佈。隨濃度增加,Sub-G1 比例明顯的增 加。
0 10 25 50
0 20 40 60 80 100
120 G1
S G2
6-shogaolμM; 12h
Cell cycle %
Sub-G1
c.
圖 4c. 6-shogaol 各濃度細胞週期統計表。隨濃度增加,Sub-G1 佔總細胞週 期的百分比逐漸升高。
a.
圖 5a. 以 Flow cytometry 測量 Hep-G2 細胞在 6-shogaol 不同濃度處理後,
Caspase-3 的含量。X 軸為螢光強度,Y 軸為螢光含量。從結果得知,Caspase-3 的含量隨著 6-shogaol 的濃度增加而上升。
b.
圖 5b. Caspase-3 含量統計圖。結果顯示,處理 6-shogaol 的 Hep-G2 細胞,
Caspase-3 含量隨著 6-shogaol 的濃度增加而上升。*為與未加藥對照組相 比,** p<0.01,***p<0.0001。
2.0 1.5 1.0 0.5 2.5
***
***
C aspase 3 f o ld
0.0
Control 6sh
ogaol 10μM 6sh
ogaol 25μM 6shogao
l 50μM
a.
圖 6a. 以 Flow cytometry 測量 Hep-G2 細胞在 6-shogaol 不同濃度處理後,
細胞凋亡的變化。X 軸為 Annexin V 螢光,Y 軸為 PI 螢光。結果顯示,細 胞凋亡時的細胞膜外翻情形,隨6-shogaol 濃度逐漸增加而上升。
b.
50μg/ml 為明顯。*為與未加藥對照組相比,* p<0.05,***p<0.0001。
圖 6b. 細胞凋亡程度統計圖。結果顯示,細胞凋亡的比例,在濃度 最
80 60 40 20 100
*
***
A p o p to si s %
0
25 50
0 10
6-shogaol μM, 24h
a.
7a. 以 Flow cytometry 測量 Hep-G2 在 6-shogaol 死劑量 (IC ),
及 10mM NAC 處理後 1h、 間點的ROS 含量。X 軸為螢光強度,
軸為螢光含量。從結果得知在6-shogaol 作用下,在 1~3 小時內會產生大 ROS。當添加了 NAC 後,ROS 含量有明顯減少。
圖 細胞 半致 50
,在 3h 時 Y
量的
.
加了NAC 後,ROS 的含量明 的減少,3h 減少的最明顯。#為與未加藥對照組相比,###p<0.0001。*為
6-shogaol 50μM 組相比,***p<0.0001。
b
3h 1h
圖 7b. ROS 增加含量統計圖,1h 與 3h 。結果顯示,處理 6-shogaol 的細胞 內在 1h 與 3h,ROS 含量有大量的增加。在添
顯 與
Contro l
6shogaol 50μM
6s
aol 50 hog
μM+
NAC 1 0mM 0.0
0.5 1.0 1.5 2.0
2.5 ###
***
ROS fold
2.5 ###
2.0 1.5 1.0
***
ROS fold
0.5 0.0
Contro l
6shogaol 50μM
6s
aol 50 hog
μM+
NAC 1 0mM
a
圖 8a. Hep-G2 細胞以濃度為 0、50μM 的 6-shogaol 和 10mM NAC 處理下,
12h 後經由 Flow cytometry 分析細胞週期。X 軸為螢光強度,Y 軸為螢光含 量。加入 6-shogaol 後細胞週期明顯被改變,在添加了 NAC 後細胞週期不 受改變。
b
圖 8b. 6-shogaol 和 NAC 的添加後其細胞週期分佈。在加入 6-shogaol 後 Sub-G1 明顯增加,添加 NAC 後此現像消失。
0 50
50+NAC 10m
M 0
20 40 60 80 100
120 G1
S G2
6-shogaolμM; 12h
Cell cycle % Sub-G1
c
圖 8c. 6-shogaol 和 NAC 的添加後其細胞週期統計表。在加入 6-shogaol 後 Sub-G1 佔總細胞週期的百分比增加,添加 NAC 後此現像消失。
Hep-G2
圖9. 以 MTT 試驗測試在藥物 6-shogaol 與不同濃度 ROS 抑制劑(NAC)處理 下 Hep-G2 細胞之存活率。X 軸為 6-shogao (IC50) 與 NAC 的濃度,Y 軸為 細胞存活率百分比。結果顯示NAC 確實能保護細胞,但仍有部分的細胞死 亡,顯示有其他凋亡路徑存在。#為與未加藥對照組相比,###p<0.0001。*
為與 6-shogaol 50μM 組相比,*p<0.05,**p<0.01。
Control μM 6-sho 50
1mM NA C 5mM NA
C 10m
M N AC M+1mM NA
C
μ
6-sho 50 M+5mM NA C
μ
6-sho 50 M+10mM NAC μ
6-sho 50 0
50 100 150
200
**
###
*
Survival rate %
圖 10. 以 Western 分析 Hep-G2 細胞在 6-shogaol 半致死濃度 (IC50) 處理後 蛋白質變化。由左到右分別為處理藥物Oh、3h、6h、12h、24h。從結果得 知,6-shogaol 會使對 Hep-G2 產生內質網壓力的現象。