行政院國家科學委員會專題研究計畫 期中進度報告
登革熱快速診斷、治療及致病機制--先天免疫和自噬現象
在登革病毒感染致病機制之探討(2/3)
期中進度報告(精簡版)
計 畫 類 別 : 整合型 計 畫 編 號 : NSC 98-2745-B-006-002- 執 行 期 間 : 98 年 12 月 01 日至 99 年 11 月 30 日 執 行 單 位 : 國立成功大學微生物學科(所) 計 畫 主 持 人 : 劉校生 共 同 主 持 人 : 凌斌、葉才明、林以行 報 告 附 件 : 出席國際會議研究心得報告及發表論文 公 開 資 訊 : 本計畫可公開查詢中 華 民 國 99 年 10 月 31 日
行政院國家科學委員會補助專題研究計畫
□ 成 果 報 告
█期中進度報告
先天免疫和自噬現象在登革病毒感染致病機制之探討(2/3)
The mechanism of innate immunity and autophagy in
dengue virus infection related
immunopathogenesis
計畫類別:□個別型計畫 ▓整合型計畫
計畫編號:NSC 98-2745-B-006 -002 -
執行期間:98 年 12 月 1 日至 98 年 11 月 30 日
計畫主持人:劉校生
計畫參與人員:林以行、凌斌、葉才明
成果報告類型(依經費核定清單規定繳交):█精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計
畫、列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年█二年後可公開查詢執行單位:國立成功大學微生物學科(所)
中 華 民 國 99 年 9 月 30 日
中文摘要:
關鍵詞:登革病毒、細胞自噬、MIF 登革病毒(DV) 之感染由輕微之登革熱到重症可能致死的登革出血熱以及登革 休克症候群(DHF/DSS)。先天免疫反應以及增加感染細胞內之病毒量可能是抗體依 賴性增強現象(ADE)造成登革病患重症發生之原因。本年度之研究重點:1. 登革病 毒感染肝癌 Huh7 細胞所誘發之自噬反應主要經由加速自噬反應流量(autophagic flux) 所 造 成 。 此 外 也 發 現 登 革 病 毒 非 結 構 蛋 白 NS1 並 不 會 被 自 噬 溶 酶 體 (Autophagolysome)中之酵素所降解,顯示自噬反應流量或許可協助病毒運送至胞 外;2. 登革病毒感染小鼠腦會誘發明顯之自噬反應以及明顯之病徵(包含體重下 降、肢體麻痺等),進一步以自噬反應抑制劑 3-MA 處理小鼠,於兩天內可見自噬反 應被抑制,小鼠之病徵亦減輕,同時病毒NS1 表現量亦下降,顯示病毒含量降低, 此結果顯示登革病毒誘發之自噬反應可經由促進病毒之複製而使病情加重; 3. 此外 我們也對ADE 狀態下可能被活化的寄主因子 HCK 以及 Mst-1 進行探討,初步並未 發現有顯著之影響;4. 登革病毒感染 Huh7 細胞誘發之巨細胞移動抑制因子(MIF) 對於血管內皮細胞通透性之影響。先前之研究已證實 MIF 與登革病人之病情正相 關,本年度集中於MIF DHF/DSS 之重要指標: 血管通透性之探討,結果顯示 DV 病 毒感染之肝癌細胞 Huh7 會產生大量之 MIF。無論是病毒感染後之條件性培養液或 重組MIF(rMIF)均會對內皮細胞(HMEC-1)造成膜通透性之改變,此現象是經由改變 細胞間結合蛋白ZO-1 分布之改變而造成。進一步之小鼠實驗證實 MIF 可造成皮下 微血管及腹腔血管通透性改變,以上結果顯示登革病患於 DHF/DSS 階段出血不止 現象可能是由MIF 造成血管通透性改變所造成。 本研究第三年將 1. 將進一步確認自噬現象在 ADE 所誘發之先天免疫反應以及 病毒量增加,所扮演之角色以及抗全長NS1 抗體與ΔC NS1 抗體對受感染細胞的自 噬作用可能的影響或其保護性; 2. 將進一步鑑定微矩陣晶片篩選之顯著基因尋找 DV 在 ADE 的情況下影響先天性免疫細胞的分子及其作用機制; 3. 進一步探討 MIF 如何與細胞接觸以及與其他因子(TNF-α,MCP-1 及 MMP 等)對血管通透性改變之 影響為何。本研究之發現將有助於臨床預測指標之建立及登革重症病患之治療。英文摘要:
Keywords:Dengue virus; Autophagy; MIFThe clinical syndromes dengue virus infection are from mild dengue fever to severe lethal symptoms including Dengue hemorrhagic fever (DHF)/dengue shock syndrome (DSS). Activation of innate immunity and increased virus load are responsible for severe DHF/DSS syndrome which is related to Antibody Dependent Enhancement (ADE). Autophagy plays an enhancing role in the replication of many viruses. DV 2 induces autophagy in many cells including monocytes and master cells. Dengue virus non-structure protein NS1 utilizes the double-membrane vesicle autophagosome as the docking platform for DV replication complex. It is intriguing to clarify whether ADE related increase of viral load is through autophagy. Moreover, whether single gene is in charge of triggering autophagy as well as whether DV induced autophagy in the cell also occurs in DV infected mice? In this study, we reveal that DV-2 replicon in the absence of structure proteins (prM, E, and core protein) replicates in the host cell and induces autophagy. Further, NS1 gene alone is sufficiently to induce autophagy. In master cells, under ADE conditions, DV infection further induces autophagy. This phenomenon was reversed by 3-MA treatment, and DV replication was also suppressed. In contrast, in DV infected monocyte THP-1, autophagy was not induced under ADE and DV-2 infection conditions. DV-2 infection induced autophagosome formation and no autophagic flux was detected. DV-2 infection of ICR suckling mice i.c. causes viral infection of mouse brain and induces autophagy. In the following two year, the underlying mechanism of DV as well as NS1 protein induced autophagy under ADE conditions as well as the gene possibly involved in DV related ADE will be explored. Our findings will shed the light on the establishment of DHF/DSS predication system; dengue virus vaccine development and effective treatment of the clinical severe dengue patients.
(一)研究計畫之背景及目的
DV 為黃質病毒科具有單一正股 RNA 基因之病毒,隨著全球氣候暖化,未來在 台灣及其他地區的感染病例將會持續攀升。DV 在體內主要感染先天免疫細胞如巨 噬細胞等,大部分感染者可自行痊癒,其餘之感染者引起從較輕微的登革熱(dengue fever, DF)到較嚴重有致命危險的出血性登革熱(dengue hemorrhagic fever, DHF)以及 休克性登革熱(dengue shock syndrome, DSS),DHF 常伴隨血漿滲出及血小板減少等 症狀。DV 感染引發重症之出血性登革熱以及登革休克症候之可能原因有:1.由二次 異型登革病毒感染依附抗體增強現象(Antibody-Dependent-Enhancement; ADE)所引 起[1, 2];2. 病毒的變異性以及病毒量持續增加會影響病程的嚴重性 [3]; 3. DF, DHF 及DSS 為相類似之疾病,只是症狀輕重不同而已,而重症之 DHF 以及 DSS 可能為 DV 免疫複合體造成補體之活化而引起的一種過敏反應[4]; 4. ADE 造成 T 細胞之活 化,並進一步誘發細胞激素(cytokines)及趨化因子(chemokine 之大量表現,最後引發 內皮細胞膜通透性改變以及血漿的流失[5, 6]; 5. DV 感染引發發炎以及自體免疫反 應,因此促成DHF 以及 DSS [7]。 微血管之通透性主要受內皮細胞之微細結構,以及細胞與細胞間之緊密結合所影 響。微血管通透性之改變可能是血管內皮細胞受到過量表現之激素或一些媒介因 子,例如TNF-α 或 IL-8,MCP-1 及 MMP 等影響所造成,此作用機轉也可能是造 成DHF/DSS 病人出血不止之原因之一 [8-11]。
Macrophage migration inhibitory factor (MIF)為具有酵素 tautomerase 活性的細胞 激素,在發炎及免疫反應調控上扮演重要的角色[12-14]。MIF 最初被發現是 T 細胞 受到抗原刺激而產生的因子,可以去抑制巨噬細胞隨意移動。MIF 幾乎能在所有種 類的細胞中表現並且受到各種刺激而釋放[15]。一旦釋放,MIF 可以刺激單核球去 分 泌 其 他 proinflammatory 細 胞 激 素 , 像 是 TNF- α , 並 且 去 抵 抗 糖 皮 質 素 (glucocorticoids)抗發炎作用[16]。MIF 也可藉由促進單核球和內皮細胞上黏附 (adhesion)分子表現增加,而增加白血球-內皮細胞的交互作用[17, 18]。此外,登革
病 毒 感 染 MIF knockout (Mif-/-) 老 鼠 相 較 於 正 常 老 鼠 會 有 較 少 的 血 液 濃 縮 (hemoconcentration)以及致死情形發生[19]。然而,對於 MIF 在 DHF/DSS 中造成血 漿滲漏(plasma leakage)所扮演的角色目前仍然不清楚。目前已知在登革病毒感染情 況下,MIF 可以誘發不同細胞表現許多發炎因子,像是 TNF-α、IL-1β、MMP 以 及血管內皮生長因子(VEGF),而會造成血管通透性增加[14, 20, 21]。然而,對於 MIF 是否可以直接增加血管通透性目前仍不清楚。 為了調控細胞內生物合成之平衡,自噬現象是胞內長效蛋白及胞器分解所必要的 過程。在養份缺乏、病原體感染以及其他環境壓力的情況下均可引發細胞自噬現象。 在細胞自噬現象發生之初,細胞質中會形成一個雙層膜構造。此雙層膜逐漸延伸, 之後微管相關蛋白1-輕鏈 3 (LC3; microtubule-associated protein 1 light chain 3) 聚集 到此雙膜構造上,稱作LC3 螢光點(LC3 puncta),此延伸的雙膜構造最後形成自噬 泡(Autophagosome):自噬泡將胞內長效蛋白和受損胞器像是粒線體等包圍隔離。 自噬泡藉由和與溶酶體(Lysosome)之融合酸化成為自噬溶酶體(Autolysosome)。 最後自噬體內之物質被溶酶體水解酵素於酸性環境下水解以供細胞之再利用,如此 可以幫助細胞克服環境壓力[22]。LC3 蛋白保留於自噬泡雙層膜內外直至自噬體完 全成熟才會自自噬體分離並被分解。LC3 為一偵測細胞是否進行自噬現象的代表蛋 白質[23]。 自噬現象引發之先天性免疫反應來移除細胞內病菌,病毒也進展出不同的機制來 破壞此先天性免疫反應。HCV 感染是經由錯誤折疊蛋白質反應途徑(UPR)引發自噬 體之形成,並利用阻止自噬體和溶酶體融合來抑制自噬蛋白降解,此作用引起自噬 體堆積並且提高 HCV RNA 複製。HCV 感染所誘發之干擾自噬路徑進行,在 HCV 致病過程中可能扮演重要的角色[24, 25]。 DV2 型 能 夠 引 發 不 同 種 類 細 胞 株 中 的 自 噬 現 象 。 DV2 誘 導 的 自 噬 體 (autophagosome)進一步增強細胞外和細胞內的病毒力價,顯示自噬體會促進感染細 胞中病毒的複製。DV2 所誘導的細胞自噬現象需要 ATG5 蛋白之參與。我們及他人 證實登革病毒感染可活化細胞自噬機制,而所產生之自噬體有利於病毒複製[26,
27]。
登革病毒的RNA 可轉譯出三個結構性蛋白 (structural protein),分別為核衣殼 鞘蛋白 (capsid protein, C)、前驅基質蛋白 (precursor membrane protein, prM) 及套膜 蛋白 (envelope protein, E) 與七個非結構性蛋白 (nonstructural protein) 包括 NS1、 NS2a、NS2b、NS3、NS4a、NS4b 及 NS5 。登革病毒 NS1 蛋白有釋出型與細胞附 著型兩種,釋出型的NS1 蛋白表現量與受登革病毒二次感染所造成的病毒血症具有 相關性,以抗體中和NS1 蛋白的作用,可保護宿主遭受病變或死亡的威脅 [5],細 胞附著型NS1 與病毒之 RF-RNA 形成複製複合體同時存在於細胞內胞器之雙層膜結 構中。老鼠中的NS1 蛋白抗體會與人類的血小板相互作用,導致出血性反應 [28]。 NS1 蛋白的抗體會形成自身抗體和內皮細胞相互作用,引發細胞外訊息傳遞,產生 NO 以及細胞凋亡現象 [29]。NS1 蛋白可能與自噬體促進病毒之複製有關,為本研 究探討之重點。未來亦將探討抗全長NS1 抗體與ΔC NS1 抗體對受感染細胞的自噬 作用可能的影響或其保護性,並評估ADE 的存在所造成的影響 登革病毒感染尚無有效疫苗預防,臨床上治療也以支持性療法為主。因此,本研 究探討DV 與先天免疫細胞的互動及其所引發的免疫致病機制,可提供重要資訊尋 找登革出血熱的預測指標,並提供未來臨床治療及疫苗研發的新方向。
(二) 研究方法
細胞株及病毒單核白血球細胞THP-1(Human monocytic cell line), HMEC-1及Huh7培養於含有 10%胎牛血清的RPMI或 MCDB 131 (Sigma-Aldrich, St. Louis, MO)培養液中。病毒 係自台灣分離出的PL0146品係。
質體轉殖
pRLucC124A-LC3wt from Dr. Marja Jaattela (Danish Cancer Society, Copenhagen, Denmark)[30]
Plasmid of tandem fluorescent-tagged LC3 (tfLC3) from Dr. Tamotsu Yoshimori (Research Institute for Microbial Diseases, Osaka University)[31]
免疫細胞螢光染色法[26]
利用慢病毒攜帶短夾干擾型核醣核酸系統 (lentivirus- based short hairpin RNA (shRNA) system) (中研院RNAiCore核心設施) 去建立MIF以及GFP基因減弱之穩定 Huh 7細胞株,所針對MIF序列為5'-GACAGGGTCTACATCAACTAT- 3',以及針對 GFP序列為5'-GCCACAACATCGAGGACGGCA-3'。慢病毒的製備方式為同時將 pMD.G, pCMVΔR8.91以及pLKO-based shRNA質體一起轉殖送入HEK293T細胞 中,在轉殖後四十八小時收集培養液中的病毒。受感染的細胞可藉由含有puromycin (2 μg/ml)的培養液進行篩選。 Permeability assays [8] 西方點墨法[8] 冷光酵素活性分析[26] 動物實驗 出生六天大的ICR 品系小鼠,以顱內注射方式感染登革病毒(2.5X105 PFU) ,感 染後0. 5、1 及 5 天犧牲小鼠,以生理食鹽水進行灌流後,收集腦組織進行西方點墨 法與冷凍包埋切片再以免疫組織染色、免疫螢光染色進行分析。
(三) 第二年研究結果與討論
I. 登革病毒感染可促進自噬通量(autophagy flux)之進行
A.
B.
D.
Figure 1. DV2 infection triggered autophagic flux in Huh7 cells.
A. Huh7 cells were transfected with plasmids of tandem fluorescent-tagged LC3 (tfLC3) and with or without DV2 infection (MOI=10) and NH4Cl treatment. The
fluorescent signals of images were detected using an Olympus FluoView 1000 confocal microscope. The images showed that mRFP merged with GFP and DAPI (nuclei). The result of quantification is B. Colocalization efficiency of mRFP with GFP signals of tfLC3 puncta was quantified and shown as the percentage of the total number of mRFP puncta. * * * : p<0.001. C. Huh7 cells were transfected with pRLucC124A-LC3wt and with or without DV2 infection (MOI=10) and NH4Cl treatment detected by Renilla
luciferase assay and Western blotting (D) to detect LC3, NS1, and p62 with specific antibodies. β-actin as an internal control. * * : p<0.01, * * * : p<0.001.
將 tandem fluorescent-tagged LC3 (tfLC3)質體轉染進入 Huh7 cells 中,觀察登革病 毒感染是否會促進自噬通量(autophagy flux)之進行。 tfLC3 質體是 LC3 蛋白同時帶 有RFP 以及 GFP 蛋白表現,因此,正常情況下利用特定波長螢光激發可觀察到紅 色螢光以及綠色螢光表現,而將影像重疊則可顯示有黃色點狀物 (puncta)。當此質 體進入到溶酶體中,因為環境酸鹼值改變,使得綠色螢光會受到遮蔽,因此只可觀 察到紅色螢光表現。藉此質體的特性,我們可用來觀察登革病毒感染是自噬體否會 與溶酶體融合,而完成自噬通量之過程。Figure 7A 及 7B 結果顯示,病毒感染 24 及36 小時主要觀察到黃色 puncta,感染 48 小時黃色 puncta 開始逐漸減少,72 小時 則觀察到較多的紅色puncta 形成,若有加入 NH4Cl 藥物去中和溶酶體的酸鹼值,阻 斷自噬通量過程,結果不論登革病毒感染時間,皆只可觀察到黃色puncta。因此, 登革病毒感染確實會驅動自噬通量發生。
pRLucC124A-LC3wt 質體為 LC3 質體前面建構帶有 Renilla luciferase 的基因,因 此,可藉由偵測 Renilla luciferase 的活性來了解 LC3 蛋白的表現。在登革病毒感染 情況下結果發現,Renilla luciferse 活性隨著感染時間增加而減少,顯示登革病毒感 染可驅使自噬通量過程而與溶酶體融合,使得LC3 蛋白被分解。登革病毒感染且加 入NH4Cl 藥物去中和溶酶體的酸鹼值,阻斷自噬通量過程,則 Renilla luciferase 活
性不會下降,顯示登革病毒感染造成的 Renilla luciferse 活性減少,確實是因為自噬 通量過程發生造成的。此實驗結果再次驗證登革病毒感染確實會驅動自噬通量發生。 此外,同時也以 Western blotting 方式進行分析。Figure 1D 結果顯示,在只有登 革病毒感染組別LC3 II 表現量比 mock 組別高,顯示確實登革病毒感染會引起自噬 現象,若病毒感染且加入NH4Cl 藥物則會造成 LC3 II 相較於只有病毒感染組別來得 高,顯示利用NH4Cl 藥物中和溶酶體的酸鹼值,阻斷自噬通量過程會使得 LC3 II 累積,也再次驗證登革病毒會完成自噬通量過程。然而,去偵測病毒蛋白NS1 卻發 現加入NH4Cl 藥物後,並沒有像 LC3 II 有累積的情形發生,是否意味著登革病毒 NS1 蛋白對於溶酶體具有抗性,這是我們未來進一步要去釐清的問題。 II. 登革病毒感染 ICR 小鼠之鼠腦並誘發自噬反應 A. B.
Figure 2. DV-2 infection of ICR suckling mice induced autophagy in the brain.
Six-day-old ICR suckling mice were intracranially inoculated with dengue virus type 2 PL046 strain (2.5X105 PFU/mouse). After virus infected for 5 days, mice were
sacrificed and the brain tissue was collected and cryosected. A. The frozen section was analyzed by immunohistochemistry (IHC) to detect viral protein, NS1 by specific
antibodies. B. The frozen section was analyzed by immunofluorescence (IFA) to detect viral protein, NS1 and autophagy marker, LC3 by specific antibodies. The images were taken by an Olympus FluoView 1000 confocal microscope.
登革病毒以顱內注射方式感染小鼠後,可於感染後第五天之鼠腦組織切片偵測到 病毒蛋白NS1,顯示病毒確實於小鼠中進行複製。同時,也可在鼠腦組織切片中偵 測到LC3 蛋白表現,並且與病毒蛋白 NS1 有 co-localization 的現象,顯示受到病毒 感染之小鼠腦中,登革病毒確實可誘發自噬反應發生。此實驗結果更加佐證先前研 究觀察到鼠腦組織中LC3 II 蛋白表現增加之情形。 A. B.
Figure 3. Inhibition of autophagy alleviates the DV2 infected clinical symptoms in suckling mice.
Six-day-old ICR suckling mice (n=2-6, each group) were intracranially treated with 3-MA (0.3mg/mouse) 2 hrs before inoculating dengue virus (2.5X105 PFU/mouse). A. The body weight (a), clinical symptoms (b), survival rate (c) were monitored daily after
inoculation. Clinical disease was scored as follows: 0: Healthy, 1: lightly sick, 2: moving slowly, 3: moving difficultly, 4: paralysis, 5: Death. B. After virus infection, mice were sacrificed and the brain tissue was harvested and total protein lysate was collected with different time and analyzed by Western blotting to detect NS1, LC3, and p62 with specific antibodies. β-actin as an internal control.
利用自噬反應抑制劑 3-MA 抑制小鼠腦中之自噬現象,再給予登革病毒感染,由 臨床症狀觀察結果可發現抑制自噬現象的組別(3-MA),其小鼠病程進行較緩慢。分 析鼠腦蛋白結果發現,抑制自噬現象之組別(3-MA) LC3-II 確實有減少,並且病毒蛋 白NS1 在自噬現象抑制組別(3-MA)也有減少之情形。
然而,在此研究中,自噬反應抑制劑 3-MA 抑制之情形能於小鼠感染後 0.5 天 及1 天觀察到,但當感染第三天及第五天結果發現,3-MA 抑制現象並不明顯(data not shown) ,顯示似乎 3-MA 藥效只有在三天內才具有效果。因此,未來對於感染較晚 期研究是否在第三天後再給予一次3-MA 處理,較能維持小鼠腦中自噬反應抑制之 現象,而能有助於我們觀察自噬反應對於登革病毒感染之影響。 總結以上,此實驗結果初步顯示,自噬反應在登革病毒感染小鼠模式中扮演幫 助病毒之角色,此現象與 in vitro 研究中相符合。然而,為了更進一步證實此現象, 未來研究中欲偵測了解是否給予自噬反應抑制劑3-MA 後,小鼠腦中之病毒效價相 較於正常病毒感染有減少之情形,而能更進一步確認自噬反應在登革病毒造成小鼠 病程中所扮演的角色。
Figure 4. Secretion of MIF in DENV-infected Huh 7 cells.
The supernatants of DENV-infected, UV- inactivated DENV-infected, or mock-infected Huh7 cells were collected at different time p.i. as indicated for the detection of MIF by ELISA. Data are presented as means ± SD from three independent experiments.
登革病毒感染之Huh7 細胞 MIF 之釋放隨著時間之增加而增加,於 48 小時之釋 放量為3 小時之 6 倍。Mock 組及不活化之病毒 MIF 之釋放量甚低。
IV. 重組之 MIF(rMIF)可增加血管通透性
Figure 5. Biological functions of rMIF.
The HRP permeability of rMIF. HMEC-1 cells were incubated with rMIF at different doses with or without ISO-1 for 30 mins. The HRP permeability was determined as described in the Materials and Methods. Data are presented as means ± SD from three independent experiments. *p<0.05 (Student’s t test).
利用transwell 分析觀察 streptavidin-HRP 在重 rMIF 作用下對內皮細胞(HMEC-1) 通透性之影響,通過之HRP 會與受質 TMB 作用而呈色。當加入 20ng/mL 之重組 MIF 於 HMEC-1 細胞內,其 S-HRP 通透性為 BSA-對照組的 2.5 倍,此現象可被重 組MIF 之抑制劑 ISO-1 所抑制(2D)。
V. MIF 於 HMEC-1 細胞中調控 ZO-1 之角色
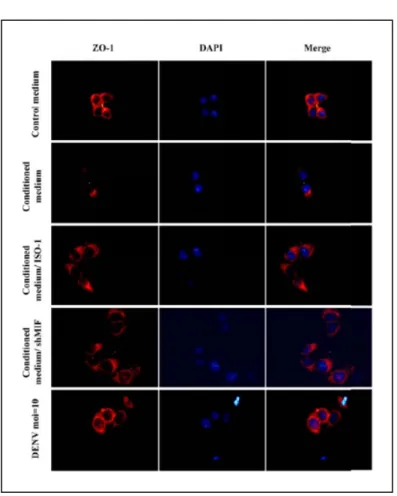
Figure 6. The effect of condition medium and rMIF on ZO-1 expression.
HMEC- 1 cells were incubated with conditioned medium or DENV (moi=10) for 4 h. The cells were fixed and stained with antibodies against ZO-1 followed by Alex-594 conjugated rabbit anti-mouse secondary antibody. Pictures were taken at magnification x200.
已知細胞與細胞間連接蛋白ZO-1 與內皮細胞之通透性有關。圖六顯示 HMEC-1 細胞若給予登革病毒感染後之條件性培養液(conditioned medium) 之處理,細胞表面 之ZO-1 蛋白表現減少,若加入 MIF 抑制劑,ZO-1 之表現量則恢復正常,此外登革 病毒感染以及以shRNA 抑制 MIF 之情況下 ZO-1 之表現均不受影響。
VI. MIF 於小鼠血管通透性所扮演之角色
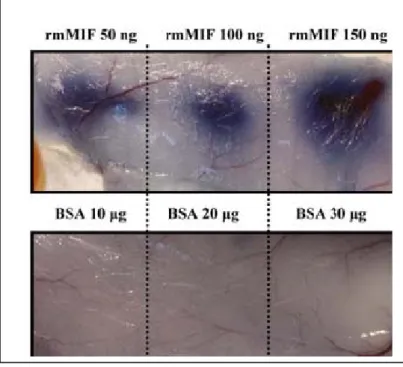
Figure 7. MIF increase the permeability of mice in vivo.
Mice were injected with 100 μl of 1% Evans blue intravenously. Different dosages of native or denatured rMIF, bovine serum albumin (BSA) were injected intradermally on the dorsum of mice. Mice were sacrificed 1 h post injection. Different doses of ISO- 1 as indicated were pre-incubated with 50 ng rMIF before injection on the dorsum of mice. Mice were sacrificed 1 h after injection.
由於沒有適當之動物模式研究登革病毒誘發之MIF 對於血管通透性改變之免疫 致病性。故以MIF 處理小鼠後,觀察腹腔內之體液與蛋白之滲漏情形。首先將染劑 Evan blue 注入小鼠血管內,再給予 rMIF 處理後,可見 Evan blue 滲出之程度隨 MIF 之劑量以及處理之時間而增加。以上結果顯示MIF 經由改變細胞間之結合蛋白 ZO-1 之排列進而影響內皮細胞通透性改變,因而造成血液滲漏之病徵。未來運用MIF 抑 制劑或抗體中和反應或許可避免登革病毒所造成支出血症狀及休克症候群。
目前已知MIF 之兩種受器為 CD74 及 CXC,MIF 可能經此受器並藉由 MAPK 及 JNK 訊息路徑傳送細胞通透性之訊息,因此 MIF 是否能與 CD74 及 CXC 受器接合 為未來探討之重點。此外除了MIF,其他之因子 TNF-α,IL-8,MCP-1 以及 MMP 等均會被登革病毒感染而大量表現並會影響細胞之通透性,它們與MIF 之關聯性及 整體對通透性之影響均有待進一步之釐清。 綜合上述結果,登革病毒誘發之MIF 除造成 TNF-α 以及 IL1β 等發炎激素之大量 產生外,並進一步活化PI3K/MEK-ERK/JNK 訊息傳遞路徑,並因此造成細胞間之 結合鬆動,因此於DHE/DSS 時病人之血漿滲漏、血球流失之病徵。
(四) 參考文獻
[1] S.B. Halstead, K. Larsen, S. Kliks, J.S. Peiris, J. Cardosa, J.S. Porterfield, Comparison of P388D1 mouse macrophage cell line and human monocytes for assay of dengue-2 infection-enhancing antibodies, The American journal of tropical medicine and hygiene 32 (1983) 157-163.
[2] R. Anderson, S. Wang, C. Osiowy, A.C. Issekutz, Activation of endothelial cells via antibody-enhanced dengue virus infection of peripheral blood monocytes, Journal of virology 71 (1997) 4226-4232.
[3] M.S. Diamond, D. Edgil, T.G. Roberts, B. Lu, E. Harris, Infection of human cells by dengue virus is modulated by different cell types and viral strains, Journal of virology 74 (2000) 7814-7823.
[4] L. Rosen, The Emperor's New Clothes revisited, or reflections on the
pathogenesis of dengue hemorrhagic fever, The American journal of tropical medicine and hygiene 26 (1977) 337-343.
[5] S. Green, A. Rothman, Immunopathological mechanisms in dengue and dengue hemorrhagic fever, Current opinion in infectious diseases 19 (2006) 429-436.
[6] A.L. Rothman, F.A. Ennis, Immunopathogenesis of Dengue hemorrhagic fever, Virology 257 (1999) 1-6.
[7] H.Y. Lei, T.M. Yeh, H.S. Liu, Y.S. Lin, S.H. Chen, C.C. Liu,
Immunopathogenesis of dengue virus infection, Journal of biomedical science 8 (2001) 377-388.
[8] Y.R. Lee, M.T. Liu, H.Y. Lei, C.C. Liu, J.M. Wu, Y.C. Tung, Y.S. Lin, T.M. Yeh, S.H. Chen, H.S. Liu, MCP-1, a highly expressed chemokine in dengue haemorrhagic fever/dengue shock syndrome patients, may cause
permeability change, possibly through reduced tight junctions of vascular endothelium cells, J Gen Virol 87 (2006) 3623-3630.
[9] N. Luplertlop, D. Misse, D. Bray, V. Deleuze, J.P. Gonzalez, V.
Leardkamolkarn, H. Yssel, F. Veas, Dengue-virus-infected dendritic cells trigger vascular leakage through metalloproteinase overproduction, EMBO Rep 7 (2006) 1176-1181.
[10] M. Khanna, U.C. Chaturvedi, M.C. Sharma, V.C. Pandey, A. Mathur, Increased capillary permeability mediated by a dengue virus-induced lymphokine, Immunology 69 (1990) 449-453.
[11] D. Talavera, A.M. Castillo, M.C. Dominguez, A.E. Gutierrez, I. Meza, IL8 release, tight junction and cytoskeleton dynamic reorganization conducive to permeability increase are induced by dengue virus infection of microvascular
endothelial monolayers, J Gen Virol 85 (2004) 1801-1813.
[12] J.A. Baugh, R. Bucala, Macrophage migration inhibitory factor, Crit Care Med 30 (2002) S27-S35.
[13] L. Leng, R. Bucala, Macrophage migration inhibitory factor, Crit Care Med 33 (2005) S475-477.
[14] T. Calandra, J. Bernhagen, C.N. Metz, L.A. Spiegel, M. Bacher, T. Donnelly, A. Cerami, R. Bucala, MIF as a glucocorticoid-induced modulator of
cytokine production, Nature 377 (1995) 68-71.
[15] T. Calandra, T. Roger, Macrophage migration inhibitory factor: a regulator of innate immunity, Nat Rev Immunol 3 (2003) 791-800.
[16] S.C. Donnelly, R. Bucala, Macrophage migration inhibitory factor: a regulator of glucocorticoid activity with a critical role in inflammatory disease, Mol Med Today 3 (1997) 502-507.
[17] Q. Cheng, S.J. McKeown, L. Santos, F.S. Santiago, L.M. Khachigian, E.F. Morand, M.J. Hickey, Macrophage migration inhibitory factor increases leukocyte-endothelial interactions in human endothelial cells via promotion of expression of adhesion molecules, J Immunol 185 1238-1247.
[18] M.A. Amin, C.S. Haas, K. Zhu, P.J. Mansfield, M.J. Kim, N.P. Lackowski, A.E. Koch, Migration inhibitory factor up-regulates vascular cell adhesion molecule-1 and intercellular adhesion molecule-1 via Src, PI3 kinase, and NFkappaB, Blood 107 (2006) 2252-2261.
[19] I. Assuncao-Miranda, F.A. Amaral, F.A. Bozza, C.T. Fagundes, L.P. Sousa, D.G. Souza, P. Pacheco, G. Barbosa-Lima, R.N. Gomes, P.T. Bozza, A.T. Da Poian, M.M. Teixeira, M.T. Bozza, Contribution of macrophage migration inhibitory factor to the pathogenesis of dengue virus infection, FASEB J 24 218-228.
[20] T. Calandra, J. Bernhagen, R.A. Mitchell, R. Bucala, The macrophage is an important and previously unrecognized source of macrophage migration inhibitory factor, J Exp Med 179 (1994) 1895-1902.
[21] H. Makita, M. Nishimura, K. Miyamoto, T. Nakano, Y. Tanino, J. Hirokawa, J. Nishihira, Y. Kawakami, Effect of anti-macrophage migration inhibitory factor antibody on lipopolysaccharide-induced pulmonary neutrophil accumulation, Am J Respir Crit Care Med 158 (1998) 573-579.
[22] H. Abeliovich, W.A. Dunn, Jr., J. Kim, D.J. Klionsky, Dissection of
autophagosome biogenesis into distinct nucleation and expansion steps, J Cell Biol 151 (2000) 1025-1034.
[23] Y. Kabeya, N. Mizushima, T. Ueno, A. Yamamoto, T. Kirisako, T. Noda, E. Kominami, Y. Ohsumi, T. Yoshimori, LC3, a mammalian homologue of yeast
Apg8p, is localized in autophagosome membranes after processing, Embo J 19 (2000) 5720-5728.
[24] D. Sir, C. Liang, W.L. Chen, J.U. Jung, J.H. Ou, Perturbation of autophagic pathway by hepatitis C virus, Autophagy 4 (2008) 830-831.
[25] M. Ait-Goughoulte, T. Kanda, K. Meyer, J.S. Ryerse, R.B. Ray, R. Ray, Hepatitis C virus genotype 1a growth and induction of autophagy, Journal of virology 82 (2008) 2241-2249.
[26] Y.R. Lee, H.Y. Lei, M.T. Liu, J.R. Wang, S.H. Chen, Y.F. Jiang-Shieh, Y.S. Lin, T.M. Yeh, C.C. Liu, H.S. Liu, Autophagic machinery activated by dengue virus enhances virus replication, Virology 374 (2008) 240-248. [27] A. Khakpoor, M. Panyasrivanit, N. Wikan, D.R. Smith, A role for
autophagolysosomes in dengue virus 3 production in HepG2 cells, The Journal of general virology 90 (2009) 1093-1103.
[28] C.F. Lin, S.W. Wan, H.J. Cheng, H.Y. Lei, Y.S. Lin, Autoimmune
pathogenesis in dengue virus infection, Viral Immunol 19 (2006) 127-132. [29] C.F. Lin, H.Y. Lei, A.L. Shiau, H.S. Liu, T.M. Yeh, S.H. Chen, C.C. Liu, S.C.
Chiu, Y.S. Lin, Endothelial cell apoptosis induced by antibodies against dengue virus nonstructural protein 1 via production of nitric oxide, J Immunol 169 (2002) 657-664.
[30] T. Farkas, M. Hoyer-Hansen, M. Jaattela, Identification of novel autophagy regulators by a luciferase-based assay for the kinetics of autophagic flux, Autophagy 5 (2009) 1018-1025.
[31] S. Kimura, T. Noda, T. Yoshimori, Dissection of the autophagosome
maturation process by a novel reporter protein, tandem fluorescent-tagged LC3, Autophagy 3 (2007) 452-460.
行政院國家科學委員會補助國內專家學者出席國際學
術會議報告
99 年 8 月 20 日報告人姓名
劉校生
服務機構
及職稱
國立成功大學微免所
教授
時間
會議
地點
99 年 7 月 31 日
99 年 8 月 3 日
韓國 釜山
本會核定
補助文號
NSC 98-2745-B-006 -002
會議
名稱
(中文)
BIT
第一屆世界病毒與感染症大會
(英文) BIT’s 1
stWorld congress of virus and
Infections-2010
發表論文
題目
(中文) 腸病毒 71 型
誘發之自體吞噬現象及其於致病
性中扮演之角色
(英文) Enterovirus 71 induced autophagy and its role in
2
會議內容
第一屆病毒與感染症世界大會由〝BIT〞組織所籌畫,開會地點為韓國第二大城 釜山(Busan),也是韓國南方第一大港,會議地點是在交通方便之 2 號地鐵站(Centum city)旁的國際會議中心(BEXCO Convention Center),會議的期間為 2010 年 7 月 31
至8 月 3 日。大會首日之上午為主題演講,下午六點為開幕晚宴,地點為 BEXCO 會議中心一樓會議廳。該晚宴約兩百位科學家參加,以Buffet 方式供應韓國傳統美 食,同時安排傳統之民俗表演,有舞蹈、打擊樂器表演等,十分精彩,尾聲亦有摸 彩活動。晚宴中遇見七、八位來自台灣不同單位之學者,包括陽明大學之榮譽教授 劉武哲教授等,顯示台灣學者參與國際會議十分踴躍。大會每天9:00 開始,中午於 會議中心三樓餐廳提供Buffet 午餐,下午由 1:30 至 6:00,所有之單元於 8 月 3 日下 午圓滿閉幕。
本大會由三大會所組成:1. WCVI-2010(World congress of virus and Infections-2010);2. SHV-2010(Symposium of Hepatitise Virus-2010);3. WSA-2010(World Summit of Anti-virus-2010)。
本人所參加的為第一屆 WCVI-2010 大會。總計有 12 個 track(主題),論文發表
約249 篇。本人受邀於 Track 2 General virology and Basic Sciences 之 Track-2-3:Viral
Replication Cycle 中口頭發表論文,題目為:Enterovirus 71 induced autophagy and its role in pathogenesis,時間為 8 月 1 日下午 5:00~5:20,地點是會議廳 201。本論文是 第一篇報導腸病毒71 型感染細胞為誘發細胞自噬反應,此自噬反應可進一步幫助病 毒之複製。本論文亦首度證實EV71 所誘發之自噬反應亦可在被感染之小鼠中發 生,自噬現象亦影響小鼠之病情,若抑制EV71 所誘發之自噬反應,則可減輕病情, 亦可降低病毒量。本文發表後受到極多之回響與熱烈之討論,收穫豐富。 二、與會心得 本 WCVI 大會統計口頭發表論文 250 篇,壁報論文 36 篇。摘錄精彩之壁報論 文如下:
enhancement or serotype and genotype differences in causative viruses.
本文發現四種血清型登革病毒均有ADE 之現象,此外病毒之基因型差異與病毒
引發之病情並無直接之關係。
2. Dr. Siew Pei Ong 之 Induction of high mobility group box (HMGB1 )release from K562 monocytic cell line upon Dengue virus infection causes vascular leakage in endothelial cells.
本文發現免疫細胞K562 在登革病毒感染後,其特定之結構與非結構蛋白可誘發
細胞產生HMGB1 蛋白,此蛋白進一步可造成內皮細胞之通透性改變,並因此
造成血管之通透性改變,與登革病毒造成出血性登革熱症狀有關 。
3. Dr. Peigang Wang 之 Development of Dengue Virus-like particle and its application in dengue egress study.
本文利用codon optimization 策略可製作出四種登革病毒血清型之 subviral 顆粒,
利用此構造,研究病毒之成熟過程。利用此系統配合siRNA 圖書館,篩選出與
登革病毒之釋放相關之寄主基因45 個。
二、重要演講摘要:
1. 大會主要演講題目:The Road to the Nobel Prize: From 2-4D to viruses and prions. Dr. Karl Maramorosch 介紹諾貝爾獎得主 Dr. Carleton Gajduselc 於 16 歲時以一位
暑期工讀生之程度開發合成出化合物2-4D(2-4-dichloro phenoxy acetic acid)用於
治療運動員之足部疼痛,卻意外發現可抑制草本植物之生長,為殺蟲劑之創始
人。後來Dr.Gajduselc 成為有名的病毒學家,他是第一位發現傳染病原造成 kuru
疾病發生於新納吉內亞,也就是現今所知的〝prion〞。
2. Dr. Guey Chuen perng 報導:Dengue virus pathogenesis-Role of progenitor cells.作 者發現登革病人感染初期骨髓細胞有被抑制的情形,本文探討骨髓之親源細胞在 DHF/DSS 病人中是否會被感染,觀察顯示骨髓細胞之感染性與病人之年齡有關。 3. Dr. Purwati Armand 報導:The role of cytokine in Dengue Hemorrhagic Fever
pathogenesis: focus on degrees of severity.
4
Thromboxome A2, Leucotnien 以及 MPTP,進一步以各種統計方法分析發現其中 之四項因子的表現於登革病毒感染之第一及第二期有上升的情形,而於第三期則 下降,上述之變化與病人之發炎反應過程或與補體, thromboxome 及 leucotnien 作用有關。
4. Dr. Ayae Honda 報導:Host protein Ebp1 induction mechanism by influenza virus infection. 流感病毒基因有八片段,每一片段基因負責一種以上病毒蛋白的製造病毒之 RNA polymerase 負責病毒之複製及轉錄,其由 PB2, PB1 及 PA 三蛋白所組成, 作者發覺寄主蛋白Ebp1 會與 PB1 蛋白結合,並進一步抑制病毒之 RNA 合成; Ep1 之表現當病毒感染時會增加。 三、本次大會帶回大會論文集一冊 四、這次第一屆病毒會議約兩百餘人參加,人數雖不多,但是卻來自世界各地負 責臨床與基礎研究之學者,對本身之問題多有深入之了解,利用此大會彼此 交流,並提供經驗之分享,對於全球崛起中之病毒因應極有幫助。
98 年度專題研究計畫研究成果彙整表
計畫主持人:劉校生 計畫編號: 98-2745-B-006-002-計畫名稱:登革熱快速診斷、治療及致病機制--先天免疫和自噬現象在登革病毒感染致病機制之探討 (2/3) 量化 成果項目 實際已達成 數(被接受 或已發表) 預期總達成 數(含實際已 達成數) 本計畫實 際貢獻百 分比 單位 備 註(質 化 說 明:如 數 個 計 畫 共 同 成 果、成 果 列 為 該 期 刊 之 封 面 故 事 ... 等) 期刊論文 0 0 0% 研究報告/技術報告 0 0 0% 研討會論文 0 0 0% 篇 論文著作 專書 0 0 0% 申請中件數 0 0 0% 專利 已獲得件數 0 0 0% 件 件數 0 0 0% 件 技術移轉 權利金 0 0 0% 千元 碩士生 2 2 100% 博士生 1 1 100% 博士後研究員 0 0 100% 國內 參與計畫人力 (本國籍) 專任助理 0 0 100% 人次 期刊論文 0 0 100% 研究報告/技術報告 0 0 100% 研討會論文 1 1 100% 篇 論文著作 專書 0 0 0% 章/本 申請中件數 0 0 0% 專利 已獲得件數 0 0 0% 件 件數 0 0 0% 件 技術移轉 權利金 0 0 0% 千元 碩士生 0 0 0% 博士生 0 0 0% 博士後研究員 0 0 0% 國外 參與計畫人力 (外國籍) 專任助理 0 0 0% 人次其他成果
(
無法以量化表達之成 果如辦理學術活動、獲 得獎項、重要國際合 作、研究成果國際影響 力及其他協助產業技 術發展之具體效益事 項等,請以文字敘述填 列。) 無 成果項目 量化 名稱或內容性質簡述 測驗工具(含質性與量性) 0 課程/模組 0 電腦及網路系統或工具 0 教材 0 舉辦之活動/競賽 0 研討會/工作坊 0 電子報、網站 0 科 教 處 計 畫 加 填 項 目 計畫成果推廣之參與(閱聽)人數 0國科會補助專題研究計畫成果報告自評表
請就研究內容與原計畫相符程度、達成預期目標情況、研究成果之學術或應用價
值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)
、是否適
合在學術期刊發表或申請專利、主要發現或其他有關價值等,作一綜合評估。
1. 請就研究內容與原計畫相符程度、達成預期目標情況作一綜合評估
■達成目標
□未達成目標(請說明,以 100 字為限)
□實驗失敗
□因故實驗中斷
□其他原因
說明:
2. 研究成果在學術期刊發表或申請專利等情形:
論文:□已發表 ■未發表之文稿 □撰寫中 □無
專利:□已獲得 □申請中 ■無
技轉:□已技轉 □洽談中 ■無
其他:
(以 100 字為限)
MIF 造成內皮細胞血管通透性改變與 DHF/DSS 出血症候之部分正在投稿之中3. 請依學術成就、技術創新、社會影響等方面,評估研究成果之學術或應用價
值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)(以
500 字為限)
1. 登革病毒誘發之自噬反應影響病毒複製之機制探討,除增進對病毒之瞭解外,亦開啟 利用自噬反應降低病毒感染之可能性。 2. 小鼠中自噬反應影響病毒相關之致病性之結果,開啟控制自噬反應治療登革病人之新 方向。3. MIF in vitro 及 in vivo 之改變血管通透性之研究,顯示其可能做為病情嚴重程度之 指標,並提供一新的治療方向。