ARTICLE NO.RC988226
Characterization, Cloning, and Expression
of Porcine
a
B Crystallin
Jiahn-Haur Liao,* Chin-Chun Hung,*
,† Jiahn-Shing Lee,‡ Shih-Hsiung Wu,*
,†
and Shyh-Horng Chiou*
,†
,1*Laboratory of Crystallin Research, Institute of Biochemical Sciences, National Taiwan University, and †Institute of Biological Chemistry, P.O. Box 23-106, Academia, and ‡Department of Ophthalmology,
Chang-Gung Memorial Hospital, Taipei, Taiwan
Received January 20, 1998
a-Crystallin is one of the best studied proteins with a-Crystallin is a major lens protein present in the respect to posttranslational modifications, including lenses of all vertebrate species. Recent studies have re- age-induced alterations due to its long life in the eye vealed that bovine a-crystallins possess genuine
chaper-lens (1). The a-crystallins of most vertebrate lenses one activity similar to small heat-shock proteins. In
or-consist ofaA andaB subunit chains showing about 55-der to compare this chaperone-like structural protein
60% sequence similarity. They were previously re-from the eye lenses of different mammalian species, we
garded as lens-specific proteins and of exclusively have cloned and expressed one of the main a-crystallin
structural nature (2). Ingolia and Craig (3) first re-subunits, i.e., aB crystallin, from the porcine lenses in
ported that heat shock proteins of Drosophila showed order to facilitate the structure-function evaluation and
sequence similarity to mammalian a-crystallin. Re-comparison of this chaperonin protein. cDNA encoding
cently they have been found to be expressed in a variety aB subunit chain was obtained using a new ‘‘Marathon
of extra-lenticular tissues suggesting that there may cDNA amplification’’ protocol of Polymerase Chain
Re-be a general cellular function for these proteins (4 – 6). action (PCR). PCR-amplified product corresponding to
This protein is believed to play an important role in aB subunit was then ligated into pGEM-T plasmid and
the maintenance of lens transparency (1).a-Crystallin prepared for nucleotide sequencing by the
dideoxy-nucleotide chain-termination method. Sequencing sev- shows structural and functional similarities to small eral positive clones containing DNA inserts coding for heat-shock proteins (7); similarly its expression can aB-crystallin subunit constructed only one complete also be induced by thermal (8) and osmotic stress (9). full-length reading frame of 525 base pairs similar to In vitro studies ofa-crystallin also indicate chaperone-human and bovine aB subunits, covering a deduced pro- like activity of this lens protein in preventing the aggre-tein sequence of 175 amino acids including the universal gation of other proteins (10,11). However, the detailed translation-initiating methionine. The porcine aB crys- molecular mechanism for its chaperone-like action re-tallin shows only 3 and 7 residues difference to bovine mains unknown.
and human aB crystallins respectively, revealing the In this report, a major interest has been focused on close relatedness among mammalian eye lens proteins. the cloning and characterization of porcine aB crys-The sequence differences between porcine and sub- tallin, which is not well studied both at the protein and mammalian species such as chicken and bullfrog are
gene levels when compared with that obtained from much greater, especially at the N- and C-terminal
re-bovine lenses. We have amplified cDNAs constructed gions of these aB crystallins. Expression of aB subunit
from the lenses of pigs employing a newer ‘‘Marathon chain in E. coli vector generated a polypeptide which
cDNA amplification’’ protocol of PCR methodology to can cross-react with the antiserum against the native
aid in the structural analysis ofaB crystallin subunit and purified aB subunit from the native porcine lenses
chains. cDNA clone encoding the aB-crystallin chain albeit with a much lower activity. q 1998 Academic Press
of this mammalian species has been sequenced success-fully, thus circumventing the need for time-consuming process of protein sequencing due to the presence of a blocking group at the amino-terminus (1 and the refer-ences therein). It is also deemed essential to determine
1Corresponding address: S.-H. Chiou, Laboratory of Crystallin
aB-crystallin sequence for interspecies
structure/func-Research, National Taiwan University, Taipei, Taiwan. Fax:
find-ings that variousa-crystallins possess genuine chaper-one activity similar to that of heat-shock proteins (10,12). Comparison of the chaperone activity ofaA and aB crystallin subunits isolated from native lenses and expressedaB crystallin product indicates that in con-trast to the previous reports, aB from both native or recombinant source possesses less activity than aA after in vitro refolding, which may reflect their struc-tural difference in the primary structure resulting in different crystallin aggregates responsible for chaper-one activity.
MATERIALS AND METHODS
Isolation and characterization of froga-crystallin. The pig (Sus
scrofa var. domestica) lenses used were obtained from a local
slaugh-terhouse. The isolation and purification of porcinea-crystallin was essentially as described before (13,14). Thea-crystallin fraction from gel filtration column was further purified into itsaA andaB subunits
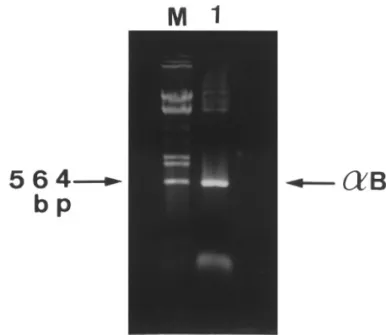
FIG. 1. Identification and size determination of PCR-amplified on reversed-phase HPLC according to the previous report (15).
porcineaB-crystallin cDNA. Electrophoresis was carried out in 1.2%
Preparation of mRNA from porcine lenses and cloning by PCR agarose gel. Lane M, DNA size markers of the EcoRI and HindIII amplification. Lens total poly(A)/RNA of porcine lenses was
puri-digestion products oflDNA, ranging from 564 to 21227 bp; Lane 1 fied using the QuickPrep mRNA preparation kit (Pharmacia, Upp- indicates the amplified PCR product of about 560 bp (arrow) encoding sala, Sweden). Double-strand cDNA synthesis was carried out with porcineaB-crystallin.
the Marathon cDNA amplification kit (Clontech Laboratories, Inc., Palo Alto, CA, USA) according to the manufacturer’s instructions. The PCR amplification was carried out using either a forward or
reverse primer corresponding to the ligated adaptor in combination debris. Soluble recombinant proteins were purified by gel filtration with anaB-specific forward primer, 5*-ATGGACATCGCCATCCAC- on Sepharose CL-6B, DEAE-Sephacel and followed by Sephacryl S-CAC-3* (3*-RACE) or anaB-specific reverse primer, 5*-CTTCTCTTC- 300HR. The purity of the recombinantaB-crystallin was confirmed ACGGGTGATGGG-3* (5*-RACE). The final amplification using a by SDS-PAGE and reversed-phase HPLC.
primer at the 5* end, i.e.
5*-C(A/T)(C/T)ACA(C/T)TCACCTAGC-Gel electrophoresis and immunoblotting. SDS-polyacrylamide CACCATG-3* within the 5*-noncoding region (with the initiator
co-slab gel (5% stacking/ 14% resolving gel) electrophoresis (SDS-PAGE) don underlined), and a primer with the stop codon at 3*-noncoding
was as described (16). For immunoblotting detection ofaB-crystallin, region, i.e. 5*-ATGCAGTTAGTGGAAGGGCATCTA-3* (with stop
co-the gels were subjected to electroblotting to a nitrocellulose mem-don underlined) were used to amplify the complete coding region
brane after SDS-PAGE followed by immunological analysis using from double-strand cDNA. The PCR products were purified on 1.2%
mouse antiserum against porcineaB-crystallin and reacted with per-agarose gel. The purified products were ligated into pGEM-T
(Pro-oxidase-conjugated AffiniPure goat anti-mouse IgG/IgM(H/L) mega, Madison, WI, USA) for sequence analysis by automatic
fluo-(Jackson ImmunoResearch Laboratories, Inc., West Grove, PA). A rescence-based sequencing using a model 373A DNA Sequencing
Sys-color development reaction was carried out using diaminobenzidine tem with a Taq DyeDeoxy terminator cycle sequencing kit (Applied
and hydrogen peroxide. Biosystems Inc., CA, U.S.A.).
Amino acid and sequence analyses. Amino acid analysis and
N-Expression and purification ofaB-crystallin. The forward primer,
terminal sequence analysis by automated Edman degradation were
i.e. 5*-GCCATATGGACATCGCCATCCACCACC-3*) corresponding
carried out as described before (17). to the 5*-end, contains an ATG initiator codon (CATATG) and an
Assay for chaperone activity ofa-crystallin under thermal heating. NdeI restriction site. The reverse primer (5
*-GCAAGCTTCTACTT-PorcinebL-crystallin (195mg) each was incubated with 14mg a-crys-CTTGGGGGCTGCAG-3*) was complementarily overlapped with the
tallin, reversed-phase HPLC separatedaA,aB and 6 M urea-treated 3*-end of the coding region containing the HindIII site (AAGCTT)
aA,aB respectively. Six samples were all in phosphate-buffered saline linked to the translational stop codon. A nucleotide fragment for the
(PBS) in a final volume of 1.5 ml. Thermal aggregation experiments porcineaB-crystallin was synthesized by PCR amplification of the
were carried out at 607C and the relative scattering was measured at porcineaB-crystallin open-reading frame of the plasmid. The PCR
340 nm as described in the previous report (18). PorcinebL-crystallin product was purified, double-digested with NdeI and HindIII and
(195mg) in PBS heated alone was used as control. Concentrations of then ligated into NdeI/HindIII sites of the isopropyl
b-D-thiogalacto-proteins were determined by a protein assay (Bio-Rad). side (IPTG)-inducible E. coli expression vector pET21a(/) (Novogen
Inc., U.S.A.). The resulting plasmid, pET/aB, was transformed into
E. coli strain BL21(DE3) (Novogen Inc.). Transformants were
se-RESULTS AND DISCUSSION
lected on LB-agar plates supplemented with 100mg/ml ampicillin. For induction of gene expression, E. coli BL21 cells containing
recom-a-Crystallins consisting ofaA andaB subunit chains
binant plasmid were grown at 377C in 1 liter of LB medium
con-taining 100mg/ml ampicillin. When the A600of the growing culture are major water-soluble proteins of the transparent eye reached 0.7, IPTG was added to a final concentration of 1 mM. Four lens, which have previously been regarded as lens-spe-hours after induction, cells were harvested and resuspended in lysis
cific proteins (2). Recently they were found to be
ex-buffer (50 mM Tris-HCl, 1 mM EDTA, 100 mM NaCl) and lysed by
pressed in a variety of extra-lenticular tissues
sug-ultrasonication. After ultrasonication, the mixture was then
FIG. 2. Nucleotide and deduced amino acid sequences of the porcineaB-crystallin subunit. The nucleotide sequence of 570 base pairs including the designed primer sequences is shown above the amino-acid sequence of 175 residues, which includes the translation initiation methionine as the first amino acid. The asterisk (*) indicates the stop codon TAG. cDNA sequence is marked in every 10-nucleotide segment for easy tracing of sequence contents. Amino acids are denoted by one-letter symbols. The underlines point to the 5*- and 3*-end primers outside the coding region used in the amplification of cDNA encodingaB-crystallin chain. The first initiation Met residue is N-terminally acetylated for allaA andaB chains after protein synthesis in vivo.
and references therein). There are currently more than Isolation and Characterization of Porcine
60 determined sequences ofaA crystallin chains from aB-Crystallin
varied species of vertebrates by conventional protein
sequencing (19,20). However there have been feweraB Lens crystallins were similarly isolated by gel filtra-tion as described previously (14). Thea-crystallin frac-chains reported in the literature by conventional
pro-tein sequencing. In order to provide a means of correlat- tion from gel filtration column was purified into itsaA and aB subunits on C4-reverse phase HPLC column ing structure/function properties of a-crystallin with
chaperone activity, it is deemed essential to comple- (15). Purified aA and aB subunits were treated with the denaturing buffer (6 M urea, 50 mM Tris/HCl, pH ment the crystallin sequence information by the facile
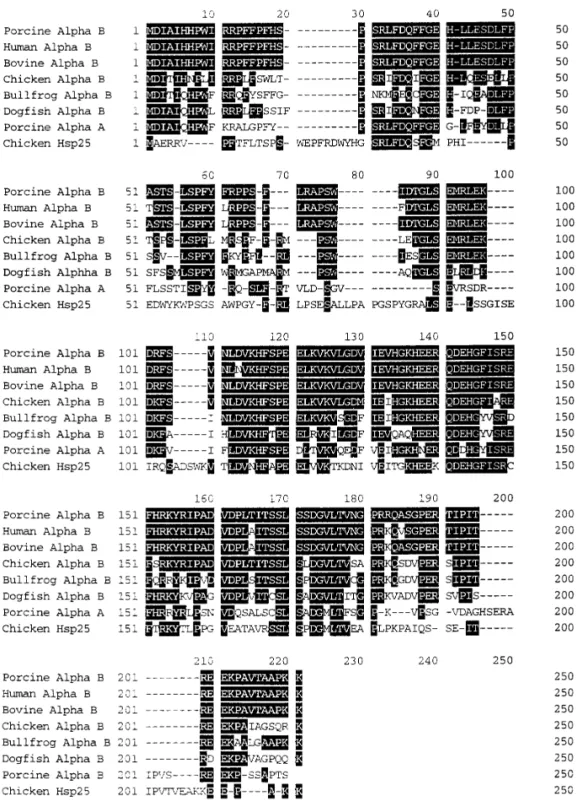
FIG. 3. Multiple sequence alignment and sequence comparison of sevena-crystallin subunits and chicken heat-shock protein (hsp25). The identical amino-acid residues among various sequences are expressed as white letters on black blocks. The gaps are introduced for optimal alignment and maximum homology among the sequences. The sequences listed are cited as follows: humanaB (24), bovineaB (25), chickenaB (26), bullfrogaB (23), dogfishaB (27), porcineaA (28), and chicken hsp25 (29). Note that the N-terminal segments (residues 1-100) are more variable among these compared sequences whereas C-terminal sequences are more conserved.
Tris/HCl, pH 8.0). Amino acid analysis and N-terminal primers based on the highly conserved mammaliana B-crystallin amino-acid sequence stretches PITREEK for sequence analysis by automated Edman degradation
5*-RACE and MDIAIHH for 3*-RACE (21,22). We have were used to check the purities and sequences of
sub-also used two primers within the 5* and 3* noncoding unit chains.
region with either initiator codon or stop codon to
Cloning and Sequence Analysis by the ‘‘Marathon’’ achieve the amplification of a 0.56 kb cDNA fragment Protocol of PCR cDNA Amplification (Fig. 1), a size corresponding to a polypeptide of about
PCR amplification of total lens cDNA mixtures pre- 170-180 amino-acid residues. After checking for nucleo-tide sequences by automatic DNA sequencing, a cDNA pared from two lenses of a single pig with the designed
sequence covering an open reading frame of 564-nucle-otides with the expected 5* and 3* primer sequences was obtained. The deduced amino acid sequence to-gether with its nucleotide coding sequence is shown in
Fig. 2. It is to be noted that we have sequenced several
positive clones containing DNA inserts coding for pig aB-crystallin subunit, all sequencing data pointing to only one complete full-length reading frame of 525 base pairs similar to human and bovine aB subunits, cov-ering a deduced protein sequence of 175 amino acids including the universal translation-initiating methio-nine. The use of ‘‘Marathon’’ PCR protocol for the am-plification of cDNA is actually a combination of 5*- and 3*-Race PCR with the advantage of avoiding using primers covering the N- or C-terminal protein coding regions which may lead to some uncertainty or ambigu-ity in the 5* and 3* nucleotide coding regions after cDNA sequencing.
Sequence Alignment and Comparison of
aB-Crystallins from Different Species
Fig. 3 shows the optimal alignment by introducing
a minimum number of gaps along the entire lengths of eight determined sequences encompassing representa-tiveaB-crystallins from species of the major classes in vertebrates by multiple sequence alignment program. In the comparative analysis of sequence alignment, it is found that porcineaB crystallins show about 96-98% sequence homology to human and bovine aB crys-tallins. The porcine aB crystallin shows only 3 and 7 residues difference to bovine and humanaB crystallins respectively, revealing the close relatedness among mammalian eye lens proteins. The sequence differ-ences between porcine and submammalian species such as chicken (73% sequence homology) and bullfrog (68% sequence homology) are much greater, especially at the N- and C-terminal regions of these aB crys-tallins. It is noteworthy that there is only about 59% sequence homology between porcineaB crystallins and a more distantly related dogfishaB crystallin. The se-quence homology between porcine aA and aB crys-tallins is about 49% which is similar to that of bovine aA and aB (23), underlining the distinct differences found between these two subunits of a-crystallin as judged by the sequence variation of these two polypep-tide chains. One salient feature for the comparison be-tween mammalianaB and chicken heat-shock protein is that there is only about 35-38% sequence similarity, well below the homology found among aB crystallins
FIG. 4. Expression and immunoblotting analysis of porcine
aB-of various vertebrate species. In spite aB-of lower sequence
crystallin. The gel was subjected to electroblotting onto a
nitrocellu-lose membrane after SDS-PAGE followed by immunological analysis homology found between these two types of proteins,a
-using mouse antiserum against porcineaB-crystallin. Lanes 1 and crystallin has been shown to possess chaperone activity 2 represent expressed porcineaB-crystallin and purified native por- similar to small heat-shock protein (10).
cineaB-crystallin respectively. (A) Amido-black stained membranes,
standard protein markers (in kDa) are shown on the right lane: Expression and Immunological Comparison of Cloned phosphorylase b (94), bovine serum albumin (67), ovalbumin (43), and Native PorcineaB Crystallins
carbonic anhydrase (30), soybean trypsin inhibitor (20), and
lyso-Since there are so far several reports on the
success-zyme (14); (B) Immunoblot stained with horseradish peroxidase
FIG. 5. Thermal aggregation ofbL-crystallin at 607C in the presence ofa-,aA- oraB-crystallins. The concentration ofbL-crystallin used in these experiments was 0.13 mg/ml whereas that of variousa-crystallins was 0.0093 mg/ml. Concentrations of crystallin solutions used in the experiments were estimated by a protein assay (Bio-Rad). Curve 1,bL-crystallin control solution in the absence ofa-crystallin; Curve 2,bL-crystallin in the presence of refolded recombinantaB-crystallin; Curve 3,bL-crystallin in the presence of 6 M urea-treated and refolded HPLC-purifiedaA-crystallin; Curve 4,bL-crystallin in the presence of 6 M urea-treated and refolded HPLC-purified aB-crystallin; Curve 5,bL-crystallin in the presence of HPLC-purifiedaA-crystallin without urea denaturation; Curve 6,bL-crystallin in the presence of nativea-crystallin isolated from gel filtration.
or human sources in E. coli system, we have also quence determination for 15 cycles. The sequencing re-sult revealed that N-terminal sequence of this protein adopted a similar expression approach by cloning the
PCR-amplified fragment encoding aB-crystallin sub- (MDIAIHHPWIRRPFF) was identical to the deduced amino-acid sequence predicted from the aB cDNA se-unit in an E. coli expression vector. A nucleotide
frag-ment for the porcineaB-crystallin was synthesized by quence. Further confirmation of the expressed porcine aB-crystallin product was demonstrated by the West-PCR amplification of the porcine aB-crystallin
open-reading frame of the plasmid. The PCR product was ern immunoblotting analysis using antiserum against purified porcineaB-crystallin isolated from pig lenses purified, double-digested with NdeI and HindIII and
then ligated into NdeI/HindIII sites of the isopropylb- (Fig. 4). It is noteworthy that antiserum against native aB-crystallin did cross-react specifically only with ex-D-thiogalactoside (IPTG)-inducible E. coli expression
vector pET21a(/). The resulting plasmid, pET/aB, was pressed products and no reactions were detected with control lysate without IPTG induction.
transformed into E. coli strain BL21(DE3). For induc-tion of gene expression, E. coli BL21 cells containing
recombinant plasmid were grown at 377C. Soluble re- Comparison of Chaperone Activity of Native and combinant proteins were purified by gel filtration on RecombinantaA andaB Crystallins
Sepharose CL-6B, DEAE-Sephacel and followed by
Sephacryl S-300HR. The purity of the recombinantaB- In the present study we have used porcine b L-crys-tallin as the substrate for thermal heating experiments crystallin was confirmed by SDS-PAGE and large-pore
(C4) reversed-phase HPLC. To verify that the 22 kDa to measure and compare the chaperone-like activity of a-crystallins from native and recombinant sources. As band on SDS-PAGE indeed corresponded to porcine
aB-crystallin, the recombinant protein purified by re- shown in Fig. 5, the rapid aggregation ofbL-crystallin in the absence ofa-crystallin during incubation at 607C versed-phase HPLC was also subjected to Edman
se-is evident (curve 1) whereas the turbidity of bL-crys- REFERENCES tallin solution is almost completely inhibited in the
1. Groenen, P. J. T. A., Merck, K. B., de Jong, W. W., and
Bloemen-presence of nativea-crystallin (about 0.01 mg/ml, curve
dal, H. (1994) Eur. J. Biochem. 225, 1 – 19.
6). We have also compared the chaperone activity for
2. Harding, J. J., and Dilley, K. J. (1976) Exp. Eye Res. 22, 1 – 74. aA, aB directly obtained from reversed-phase HPLC
3. Ingolia, T. D., and Craig, E. A. (1982) Proc. Natl. Acad. Sci. USA
and refolded aA,aB after 6 M urea denaturation to- 79, 2360 – 2364.
gether with recombinant aB after purification and re- 4. Bhat, S. P., and Nagineni, C. N. (1989) Biochem. Biophys. Res. folding. It is noteworthy that the recombinant aB Commun. 158, 319 – 325.
(curve 2) and native and urea-treated refolded aB 5. Dubin, R. A., Wawrousek, E. F., and Piatigorsky, J. (1989) Mol.
(curve 4) all show less activity than nativea(curve 6) Cell. Biol. 9, 1083 – 1091.
andaA without urea denaturation (curve 5). The rea- 6. Iwaki, T., Kume-Iwaki, A., Liem, R. K. H., and Goldman, J. E. (1989) Cell 57, 71 – 78.
sons for this discrepancy are probably due to the
forma-7. Sax, C. M., and Piatigorsky, J. (1994) Adv. Enzymol. 69, 155 –
tion of improper foldingaB aggregates after expression
201.
or there may be some undetected contaminant proteins
8. Klemenz, R., Frohli, E., Steiger, R. H., Schafer, R., and Aoyama,
from E. coli expression system which may interfere
A. (1991) Proc. Natl. Acad. Sci. USA 88, 3652 – 3656.
with refolding or inhibit the chaperone activity per se.
9. DasGupta, S., Hohman, T. C., and Carper, D. (1992) Exp. Eye
We are currently refining the purification and refolding Res. 54, 461 – 470.
protocols for the recombinant aB crystallin in E. coli 10. Horwitz, J. (1992) Proc. Natl. Acad. Sci. USA 89, 10449 – 10453. expression system with the aim of achieving the correct 11. Wang, K., and Spector, A. (1994) J. Biol. Chem. 269, 13601 – folding aggregate with higher chaperone activity for 13608.
structure-function study ofaB crystallin. 12. Merck, K. B., Groenen, P. J. T. A., Voorter, C. E. M., de Haard-Hoekman, W. A., Horwitz, J., Bloemendal, H., and de Jong, W. W. (1993) J. Biol. Chem. 268, 1046 – 1052.
CONCLUSION 13. Chiou, S.-H. (1986) FEBS Lett. 201, 69 – 73.
14. Chiou, S.-H., and Azari, P. (1989) J. Protein Chem. 8, 1 – 17.
Insights into the mechanism of chaperone-like activ- 15. Perry, R. E., and Abraham, E. C. (1986) J. Chromatogr. 351, 103 – 110.
ity associated witha-crystallin and its subunitaA and
16. Laemmli, U. K. (1970) Nature 227, 680 – 685. aB chains may be revealed through the analysis and
17. Chiou, S.-H. (1988) FEBS Lett. 241, 261 – 264.
comparison of molecular sequences and structures
18. Horwitz, J. (1992) Proc. Natl. Acad. Sci. USA 89, 10449 – 10453.
from eye lenses of different animal species. In this
19. de Jong, W. W., Zweers, A., Versteeg, M., and Nuy-Terwindt,
study we have established the nucleotide sequence of
E. C. (1984) Eur. J. Biochem. 141, 131 – 140.
cDNA clone encoding the major aB crystallin subunit
20. Stapel, S. O., Leunissen, J. A. M., Versteeg, M., Wattel, J., and
from porcine eye lenses by a facile Marathon-Race
pro-de Jong, W. W. (1984) Nature 311, 257 – 259.
tocol of PCR methodology. In light of the recent intense
21. Dubin, R.-A., Ally, A.-H., Chung, S., and Piatigorsky, J. (1990)
interest in studies on the structural and functional sim- Genomics 7, 594 – 601.
ilarity between a-crystallin and heat shock proteins 22. Frederikse, P.-H., Dubin, R.-A., Haynes, J.-I. II., and Piatigor-and their associated chaperone activity, characteriza- sky, J. (1994) Nucleic Acids Res. 22, 5686 – 5694.
tion ofaA/aB crystallin from more diverse species both 23. Lu, S.-F., Pan, F.-M., and Chiou, S.-H. (1995) Biochem. Biophys. at the structural and functional levels may provide im- Res. Commun. 216, 881 – 891.
portant insights on the molecular origin and underly- 24. Kramps, J. A., de Man, B. M., and de Jong, W. W. (1977) FEBS
Lett. 74, 82 – 84.
ing mechanism for the evolution of chaperone activity
25. van der Ouderaa, F. J., de Jong, W. W., Hilderink, A., and
associated with this major class of crystallins in the
Bloemendal, H. (1974) Eur. J. Biochem. 49, 157 – 168.
animal kingdom.
26. Sawada, K., Agata, K., and Eguchi, G. (1992) Exp. Eye Res. 55, 879 – 887.
27. de Jong, W. W., Leunissen, J. A. M., Leenen, P. J. M., Zweers,
ACKNOWLEDGMENTS
A., and Versteeg, M. (1988) J. Biol. Chem. 263, 5141 – 5149. 28. de Jong, W. W., Terwindt, E. C., and Bloemendal, H. (1975) This work was supported by Academia Sinica and the National
FEBS Lett. 58, 310 – 313.
Science Council (NSC Grants 83-0203-B-001-086,
83-0418-B-001-020BA, 84-2311-B-001-050-BA & 86-2311-B-002-031-B15), Taipei, 29. Miron, T., Vancompernolle, K., Vandekerckhove, J., Wilchek, M., and Geiger, B. (1991) J. Cell Biol. 114, 255 – 261.