The Journal of Nutrition
Biochemical, Molecular, and Genetic Mechanisms
Sulforaphane and
a-Lipoic Acid Upregulate
the Expression of the
p Class of Glutathione
S-Transferase through c-Jun and
Nrf2 Activation
1,2
Chong-Kuei Lii,
3Kai-Li Liu,
4Yi-Ping Cheng,
4Ai-Hsuan Lin,
4Haw-Wen Chen,
3* and Chia-Wen Tsai
3*
3Department of Nutrition, China Medical University, Taichung 404, Taiwan; and4Department of Nutrition, Chung Shan Medical
University, Taichung 404, Taiwan
Abstract
The anticarcinogenic effect of dietary organosulfur compounds has been partly attributed to their modulation of the activity and expression of phase II detoxification enzymes. Our previous studies indicated that garlic allyl sulfides upregulate the expression of thep class of glutathione S-transferase (GSTP) through the activator protein-1 pathway. Here, we examined the modulatory effect of sulforaphane (SFN) anda-Lipoic acid (LA) or dihydrolipoic acid (DHLA) on GSTP expression in rat Clone 9 liver cells. Cells were treated with LA or DHLA (50–600mmol/L) or SFN (0.2–5 mmol/L) for 24 h. Immunoblots and real-time PCR showed that SFN, LA, and DHLA dose dependently induced GSTP protein and mRNA expression. Compared with the induction by the garlic organosulfur compound diallyl trisulfide (DATS), the effectiveness was in the order of SFN. DATS . LA = DHLA. The increase in GSTP enzyme activity in cells treated with 5mmol/L SFN, 50 mmol/L DATS, and 600 mmol/L LA and DHLA was 172, 75, 122, and 117%, respectively (P , 0.05). A reporter assay showed that the GSTP enhancer I (GPEI) was required for GSTP induction by the organosulfur compounds. Electromobility gel shift assays showed that the DNA binding of GPEI to nuclear proteins reached a maximum at 0.5–1 h after SFN, LA, and DHLA treatment. Super-shift assay revealed that the transcription factors c-jun and nuclear factor erythroid-2 related factor 2 (Nrf2) were bound to GPEI. These results suggest that SFN and LA in either its oxidized or reduced form upregulate the transcription of the GSTP gene by activating c-jun and Nrf2 binding to the enhancer element GPEI. J. Nutr. doi: 10.3945/jn.110.121418.
Introduction
Epidemiologic studies have found that persons who consume a
high proportion of vegetables and fruits in their diet may
decrease their cancer risk (1,2). This can be partly attributed to
the rich content of numerous phytochemicals in vegetables and
fruits, including polyphenolic compounds, carotenoids, and
organosulfur compounds (2–5). The accumulated evidence
supports that garlic alliin-derived allyl sulfides and cruciferous
isothiocyanates protect animals against a variety of chemical
carcinogens (1,6). This chemoprevention can be partly
ex-plained by the potency of these phytochemicals in modulating
the activity and gene expression of phase II detoxification
enzymes (7–9).
Glutathione S-transferase (GST)
5is a phase II enzyme that
catalyzes the conjugation of glutathione with a variety of
electrophilic xenobiotics and facilitates their excretion. In
mammals, 8 GST isozymes, including A (a), M (m), O (v), P
(p), S (s), T (u), Z (z), and K (k), have been identified (10).
Recently, interest has grown in the physiologic properties of the
p class of GST (GSTP), not only because of its function in drug
detoxification but also because of its possible roles in cell
transformation and carcinogenesis (11,12). GSTP activity has
been used to evaluate the potency of chemoprevention agents in
1
Supported by National Science Council 97-2320-B-039-048-MY3.
2
C.-K. Lii, K.-L. Liu, Y.-P. Cheng, A.-H. Lin, H.-W. Chen, and C.-W. Tsai, no conflicts of interest.
* To whom correspondence should be addressed. E-mail: [email protected]. tw and [email protected].
5
Abbreviations used: AP-1, activator protein-1; ARE, antioxidant response element; DATS, diallyl trisulfide; DHLA, dihydrolipoic acid; DMSO, dimethylsulf-oxide; DTT, dithiothreitol; EMSA, electromobility gel shift assay; GAPDH, glycer-aldehyde 3-phosphate dehydrogenase; GPEI,p class of glutathione S-transferase enhancer I; GST, glutathione S-transferase; GSTA, a class of glutathione S-transferase; GSTM, m class of glutathione S-transferase; GSTP, p class of glutathioneS-transferase; LA, a-lipoic acid; NQO1, NAD(P)H-dependent quinone oxidoreductases 1; Nrf2, nuclear factor erythroid-2 related factor 2; SFN, sulforaphane; TRE, 12-O-tetradecanoylphorbol-13-acetate responsive element. 0022-3166/08 $8.00ã 2010 American Society for Nutrition.
Manuscript received January 18, 2010. Initial review completed February 2, 2010. Revision accepted February 15, 2010. 1 of 8 doi: 10.3945/jn.110.121418.
The Journal of Nutrition. First published ahead of print March 17, 2010 as doi: 10.3945/jn.110.121418.
benzo[a]pyrene-induced cancer (13). The importance of GSTP in
cancer prevention is further supported by the fact that
7,12-dimethylbenzanthracene-induced skin cancer is significantly
elevated in GSTP-null mice (14). Two enhancing elements were
identified in the 5
9 upstream region of the GSTP gene and were
named GSTP enhancer I (GPEI, –2.5 kb) and II (GPEII, –2.2 kb)
(15). GPEI has 2 phorbol-12-O-tetradecanoate-13-acetate
re-sponsive element (TRE)-like elements that are considered to be
required for basal and inducible expression of GSTP (16).
Enhancers of GSTP expression are regulated by multiple factors,
including activator protein-1 (AP-1), which is known to be a
heterodimer or homodimer composed of the products of c-Jun
and c-fos (17). Because the TRE-like elements in GPEI share
sequences similar to those of the antioxidant response element
(ARE), nuclear factor erythroid-2 related factor 2 (Nrf2) is also
regarded as a possible transcriptional factor that binds to
GPEI (18).
Sulforaphane (SFN), an isothiocyanate compound rich in
cruciferous vegetables, has been demonstrated to be highly
effective in affording protection against chemically induced
cancer in animal models (6,9). This cytoprotection by SFN can
be attributed to its activation of apoptosis and also its effective
induction of the expression of phase II detoxification and
antioxidant enzymes, including the a class of GST (GSTA), m
class of GST (GSTM), NAD(P)H-dependent quinone
oxidore-ductases 1(NQO1), and g-glutamylcysteine synthase (9,19,20).
Recently, the upregulation of the gene expression of
cytopro-tective genes by SFN was shown to be dependent on Nrf2-ARE
(21,22).
a-Lipoic acid (LA) is a thiol antioxidant distributed in
vegetables, including broccoli, spinach, and tomatoes (23). LA
and its reduced form, dihydrolipoic acid (DHLA), not only act as
potent free radical scavengers and metal chelators (24) but also
participate in the recycling of other cellular antioxidants,
including vitamin C, vitamin E, and glutathione (25). Recently,
the expression of several phase II enzymes was reported to be
modulated by LA and DHLA. In human leukemia HL-60 cells
and neuroblastoma SH-SY5Y cells, LA is effective at
upregulat-ing NQO1 gene transcription (26,27). LA induction of GSTA2
expression is likely associated with the phosphatidylinositol
3-kinase pathway (8). Regarding GSTP, however, it is not clear
whether LA and DHLA induce the expression of this
detoxifi-cation enzyme.
Recently, we reported that garlic oil and 2 of its major
organosulfur components, diallyl disulfide and diallyl trisulfide
(DATS), can effectively upregulate GSTP mRNA and protein
expression. Moreover, GPEI is required for the induction of this
phase II enzyme (28–30). In addition to the garlic allyl sulfides,
we were also interested in examining whether organosulfur
compounds not derived from garlic are also effective at
upregulating GSTP expression and the possible transcription
factors involved. Therefore, in the present study, we examined
the modulatory effect of SFN, LA, and DHLA on GSTP
expression in rat liver Clone 9 cells. Moreover, we compared
the relative induction potency on GSTP of DATS, LA, DHLA,
and SFN.
Materials and Methods
Materials. All other chemicals were purchased from Sigma-Aldrich unless specified otherwise. SFN and DATS were obtained from LKT Laboratories. RPMI-1640 medium and penicillin-streptomycin solution were obtained from Gibco Laboratory. RNase inhibitor, oligo dT, and Moloney murine leukemia virus RT were purchased from Promega.
GSTP and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) primer were obtained from Applied Biosystems. Fetal bovine serum was purchased from Hyclone. Trizol and lipofectamine were ordered from Invitrogen.
Cell culture.Clone 9 cells, which were derived from normal rat livers, were obtained from Bioresources Collection and Research Center. They were grown in RPMI-1640 medium supplemented with 10 mmol/L HEPES, 100 kU/L penicillin, 100 mg/L streptomycin, and 10% fetal bovine serum at 378C in a humidified atmosphere of 5% CO2and 95% air. For all studies, cells between passages 4 and 10 were used. The cells were plated on 35-mm plastic tissue culture dishes (Falcon) at a density of 2.53 105cells/dish and were allowed to grow for 24 h. Fresh culture medium containing various concentrations of DATS, LA, DHLA, or SFN was then added and the cells were incubated for the indicated times. Cells treated with 0.1% dimethylsulfoxide (DMSO) alone were used as controls.
SDS-PAGE and Western blot.Cells were washed twice with cold PBS and were then harvested in 300 mL of 20 mmol/L potassium phosphate buffer (pH 7.0). Supernatants were centrifuged at 10,0003 g for 30 min at 48C. Protein concentrations were determined with the Coomassie Plus Protein Assay Reagent kit (Pierce Chemical). Four micrograms of cellular proteins from each sample was applied to 10% SDS-polyacry-lamide gels and electrophoretically transferred to polyvinylidene fluoride membranes (Millipore). The membranes were blocked at 48C overnight with 50 g/L nonfat dry milk solution and were then incubated with primary antibody against GSTP (Transduction Laboratories), GSTA, GSTM (all from Oxford Biomedical Research), NQO1, c-Jun, phospho-c-Jun, Nrf2 (all from Santa Cruz Biotechnology), or b-actin for 70 min at room temperature and were subsequently incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG, goat anti-mouse IgG (all from Perkinelmer Life Sciences), or rabbit anti-goat IgG (R&D Systems) secondary antibody. The bands were visualized by using an enhanced chemiluminescence kit (Perkin-Elmer Life Science).
Real-time PCR.Total RNA was extracted by using Trizol reagent. A total of 0.8 mg RNA was used for the synthesis of first-stand cDNA. RT was carried out in a programmable thermal cycler and was performed in 20 mL containing 25 mmol/L Tris-HCl (pH 8.3), 50 mmol/L (NH4)2SO4, 0.3% b-mercaptoethanol, 0.1 g/L bovine serum albumin, 5 mmol/L MgCl2, 1 mmol/L of each deoxynucleotide triphosphate, 2.5 U RNase inhibitor, and 2.5 mmol/L oligo dT and Moloney murine leukemia virus RT. The reaction mixture was incubated for 1 cycle at 428C for 15 min, 998C for 5 min, and 48C for 10 min. Real-time PCR was carried out in an ABI Prism 7000 sequence detector (Applied Biosystems) by adding 5 mL cDNA, 10 mL Master Mixture, 5 mL ddH2O, and 1 mL GSTP (Rn02770492_gh) and GAPDH primer (Mm99999915_gl) to each microwell. The reaction was run with the following program: 1 cycle at 508C for 2 min and 958C for 10 min, followed by 40 cycles at 958C for 15 s and 608C for 1 min. The comparative Ct (threshold cycle) method was used to determine the relative amount of GSTP mRNA (31). The DCt method was used for quantification of amplified gene targets ac-cording to the manufacturer’s protocol (Applied Biosystems). Briefly, the number of cycles required to reach a threshold level of log-based fluorescence (Ct value) was normalized to the Ct value of GAPDH gene in each sample. The relative expression value for GSTP gene was cal-culated as 22DDCt, where DDCt represents difference of Ct between the GSTP gene and the GAPDH gene (DCt = CtGSTP gene2 CtGAPDH gene; DDCt = DCtGSTP gene2 DCtcalibrator).
Enzyme activity assays. GST activity was measured by using ethacrynic acid as the substrate because of its better selectivity of the p class isozyme (32). Briefly, the reaction mixture in a final volume of 1 mL contained 100 mmol/L potassium phosphate buffer (pH 6.5), 0.5 mmol/ L glutathione, 0.2 mmol/L ethacrynic acid, and an appropriate amount of the total proteins. The ethacrynate-glutathione conjugate formed was measured at 270 nm. The GST activity was measured with 1-chloro-2,4-dinitrobenzene, whereas NQO-1 activity was determined using 2,6-dichloroindophenol as the substrate (33).
Expression and reporter constructs.The pTA-GSTP Luc reporter with GSTP gene promoter region was constructed as described previ-ously (28). A 2.7-kb fragment of the gene for GSTP was inserted into the MluI and NheI site of pTA-SEAP/Luc vector (Clontech). In addition to the full-length construct (Luc-2713), 2 constructs with deletions from 22713 to 22605 bp 2604) and from 22713 to 22376 bp (Luc-2375) were generated. A reporter with the GPEI fragment was constructed by ligating the 22713 to 22605 bp segment into pTA-SEAP/Luc vector and was designated as Luc-GPE.
Transient transfection and luciferase activity assay.Clone 9 cells were plated at a density of 2.53 105cells on 35-mm plastic tissue culture dishes and the dishes were incubated until 70% confluence was reached. Cells were transiently transfected for 5 h with 0.1 mg of the pTA-GSTP Luc vectors by lipofectamine reagent and were then exposed to each of the organosulfur compounds for an additional 15 h. Cells were then washed twice with PBS and were lysed in 100 mL of lysis buffer. Luciferase activity was measured by using Luciferase Assay reagent (Clontech) according to the manufacturer’s instructions. The luciferase activity of each sample was corrected on the basis of b-galactosidase activity, which was measured at 420 nm with O-nitrophenyl b-D-galactopyranoside as a substrate. The value for cells treated with DMSO vehicle alone was regarded as 1.
Electromobility gel shift assay.Electrophoretic mobility shift assay (EMSA) was performed according to our previous study (29). Cells were washed twice with cold PBS followed by scraping from the dishes with PBS. Cell homogenates were centrifuged at 20003 g for 5 min. The cell pellet was allowed to swell on ice for 15 min after the addition of 200 mL of hypotonic buffer containing 10 mmol/L HEPES, 10 mmol/L KCl, 1 mmol/L MgCl2, 1 mmol/L EDTA, 0.5 mmol/L dithiothreitol (DTT), 0.5% Nonidet P-40, 4 mg/L leupeptin, 20 mg/L aprotinin, and 0.2 mmol/L phenylmethylsulfonyl fluoride. After centrifugation at 60003 g for 15 min, pellets containing crude nuclei were resuspended in 50 mL of hypertonic buffer containing 10 mmol/L HEPES, 400 mmol/L KCl, 1 mmol/L MgCl2, 1 mmol/L EDTA, 0.5 mmol/L DTT, 10% glycerol, 4 mg/L leupeptin, 20 mg/L aprotinin, and 0.2 mmol/L phenylmethylsul-fonyl fluoride and were incubated for an additional 30 min on ice. The nuclear extracts were then obtained by centrifugation at 10,0003 g for 15 min and were frozen at2808C until the EMSA was performed.
The LightShift Chemiluminescent EMSA kit (Pierce Chemical) and synthetic biotin-labeled double-stranded GPEI consensus oligonucleo-tide (forward: 59-AGTAGTCAGTCACTATGATTCAGCAAC-39; re-verse: 59-GTTGCTGAATCATAGTGACTGACTACT-39) were used to measure the effect of organosulfur compounds on GPEI nuclear protein-DNA binding activity. Unlabeled double-stranded GPEI (200 ng) and a mutant double-stranded oligonucleotide were also used to confirm specific binding. Two micrograms of nuclear protein, poly(dI-dC), and biotin-labeled double-stranded GPEI oligonucleotide were mixed with the binding buffer to a final volume of 20 mL and were incubated at room temperature for 30 min. The nuclear protein-DNA complex was separated by electrophoresis on a 6% Tris-boric acid-EDTA-polyacry-lamide gel and was then electrotransferred to a Hybond-N+ nylon membrane (GE Healthcare). The membrane was treated with strepta-vidin-horseradish peroxidase and the nuclear protein-DNA bands were developed by using an enhanced chemiluminescence kit. In the super-shift assay, nuclear protein was incubated with 1 mg of monoclonal anti-c-Jun antibody for 30 min after the binding reactions and was subjected to electrophoresis as described above.
Immunoprecipitation.A total of 15 mg of nuclear proteins was first incubated with 1 mg anti-Nrf2 antibody overnight at 48C. The cells were mixed with 0.1 g/L Protein A-Sepharose beads for 1 h at 48C. Immunoprecipitated complexes were pelleted by centrifugation at 160003 g for 2 min at 48C. The pellet was washed 5 times with 1 mL IP buffer (40 mmol/L Tris-HCl, pH 7.4, 1% Nonidet P-40, 150 mmol/L NaCl, 5 mmol/L EGTA, 1 mmol/L DTT, 1 mmol/L phenylmethylsulfonyl fluoride, 1 mg/L aprotinin, 1 mg/L leupeptin, 20 mmol/L sodium fluoride, 1 mmol/L sodium orthovanadate) and was then subjected to electrophoresis followed by Western blot.
Statistical analysis.Statistical analysis was performed with commer-cially available software (SAS Institute). Data were analyzed using 1-way ANOVA, and the significant difference among treatment means was assessed using Tukey’s test. Different from the 2 deletion constructs in the same treatment was analyzed by a separate ANOVA. A value of P, 0.05 was considered significant.
Results
GSTP protein expression.
In this study, clone 9 cells were
incubated with 50 mmol/L DATS, 50–600 mmol/L LA or DHLA,
or 0.2–5 mmol/L SFN for 24 h. To ensure that no cytotoxicity
resulted by treatment with these organosulfur compounds, we
first performed a cell viability assay. The
3-(4,5-dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide method showed that each
of the organosulfur compounds tested at the concentrations
stated above resulted in cell viability
.95% (data not shown).
Immunoblotting showed that LA, DHLA, and SFN dose
dependently induced GSTP protein expression in Clone 9 cells
(Fig. 1). LA and DHLA at 600 mmol/L caused a 5.4- and 4.8-fold
increase, respectively, in the GSTP level compared with the
control cells (P
, 0.05). This induction was similar to that noted
in cells treated with 50 mmol/L of DATS. It was interesting to
note that SFN showed the greatest potency in upregulating
GSTP expression among all the organosulfur compounds tested.
An 8.1-fold induction in GSTP expression was reached when
cells were exposed to 5 mmol/L SFN.
Organosulfur compounds affect GSTP mRNA level and
activity.
By real-time PCR, the increases in GSTP mRNA levels
were consistent with the changes noted in protein expression.
DATS caused a 1.1-fold increase in the GSTP mRNA level
compared with the control cells (P
, 0.05). There was a
dose-dependent induction of GSTP mRNA in cells treated with LA,
DHLA, and SFN. The increase in expression caused by SFN was
higher than that caused by LA or DHLA (Fig. 2A). Again,
enzyme activity toward ethacrynic acid was dose dependently
increased by LA, DHLA, and SFN (Fig. 2B).
FIGURE 1 Protein levels of GSTP induced by organosulfur com-pounds. Cells were cultured with 0.1% DMSO alone (2) or with various concentrations of DATS, LA, DHLA, or SFN for 24 h. GSTP protein was determined by immunoblot assay. A total of 4mg protein for each sample was applied for electrophoresis. Changes in GSTP protein expression were measured by densitometry. Data were normalized tob-actin expression. The level in control cells was set at 1. Each value represents the means6 SD, n = 4. Means without a common letter differ,P , 0.05.
GSTP promoter activity.
The different length constructs were
transiently transfected into Clone 9 cells to examine whether the
promoter activity of the GSTP gene was modulated by the
organosulfur compounds and to locate the possible responsive
sites. With the Luc-2713 reporter, 600 mmol/L of LA or DHLA
and 5 mmol/L of SFN resulted in 2.0-, 1.5-, and 3.7-fold higher
luciferase activity, respectively, than that in the control cells (P
,
0.05) (Fig. 3A). A 1.7-fold increase in reporter activity was noted
in cells treated with 50 mmol/L DATS. When the
22713- to
22605-bp region (GPEI) of the GSTP promoter was deleted
(Luc-2604), however, this increase in reporter activity was
completely abolished, and the activity was similar to that noted
in cells transfected with Luc-2375.
To further demonstrate the importance of GPEI in GSTP
expression in response to organosulfur compounds, a reporter
construct (Luc-GPE) was created by ligating the genomic 109-bp
GPEI segment (
22713 to 22605 bp) to the luciferase coding
region. The results clearly indicated that DATS, LA, DHLA, and
SFN increased the reporter activity by 243, 189, 143, and 352%,
respectively, compared with that in the control cells (P
, 0.05)
(Fig. 3B). These data establish that the GPEI bears the
organosulfur compound-responsive element and that this
ele-ment is essential for this stimulation of the promoter activity.
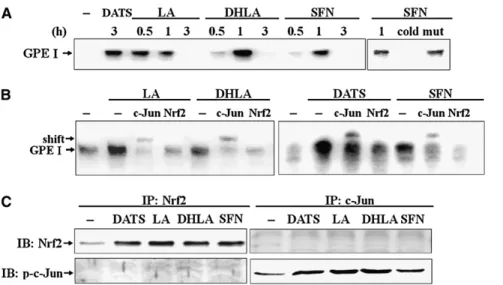
Protein binding activity on GPEI by EMSA.
An EMSA was
used to identify the transcription factors that were bound to
GPEI. In the presence of the organosulfur compounds, the DNA
binding activity reached a maximum at 0.5–1 h (Fig. 4A).
Specificity of the DNA-protein interaction for GPEI was
demonstrated by a competitive assay with 100-fold excess of
unlabeled double-stranded oligonucleotide (cold) and also with
a mutant double-stranded oligonucleotide (mut). Next, a
super-shift assay with highly specific antibodies directed against c-jun
and Nrf2 was performed. The GPEI nuclear protein band was
abrogated and super-shift occurred in the presence of anti-c-Jun
antibody (Fig. 4B). In addition, an immunoprecipitation with
anti-Nrf2 antibody was performed before EMSA. As noted,
anti-Nrf2 antibody diminished the binding of nuclear proteins to
GPEI oligonucleotides (Fig. 4B). Accompanied by the decrease
in Nrf2-GPEI binding, Nrf2 in the nuclear immunoprecipitates
was increased in cells treated with organosulfur compounds
(Fig. 4C). The activation of c-Jun after immunoprecipitation
with anti-c-Jun antibody was consistent with the result of
organosulfur compound-induced nuclear Nrf2 accumulation.
Following the immunoprecipitation, whether c-Jun interacts to
Nrf2 by serving as a partner was determined. No Nrf2 was
detected in the nuclear immunoprecipitates with c-Jun antibody
(Fig. 4C). Similarly, in the Nrf2 immunoprecipitates, there was
no p-c-Jun detected.
Expression of other phase II enzymes.
We also assessed the
expression of other detoxification enzymes that are known to be
upregulated by a Nrf2-dependent mechanism, including GSTA,
GATM, and NQO-1, by immunoblots. As indicated, various
concentrations of LA, DHLA, and SFN dose dependently
FIGURE 2 mRNA expression and enzyme activity of GSTP inducedby organosulfur compounds. Cells were treated with DMSO alone (2) or with various concentrations of DATS, LA, DHLA, or SFN for 24 h. (A) Real-time PCR of GSTP mRNA expression. The GSTP mRNA level in the control cells was regarded as 1. (B) GST activity determined by using ethacrynic acid as a substrate. Values are means6 SD, n = 3–4. Groups without a common letter differ,P , 0.05.
FIGURE 3 GPEI is required for the upregulation of GSTP by phyto-derived organosulfur compounds. (A) Cells were transfected with different constructs and were then treated with 50mmol/L DATS, 600 mmol/L LA, 600 mmol/L DHLA, or 5 mmol/L of SFN for 15 h. The luciferase activity of cells transfected with pTA-2713 and treated with DMSO alone (2) was regarded as 1. (B) The GPEI-linked construct was transfected into cells and the cells were treated with DATS, LA, DHLA, or SFN for 15 h. Values are means6 SD, n = 3–4. Groups in the same construct without a common letter differ, P , 0.05. #Different from the 2 deletion constructs in the same treatment,P , 0.05.
stimulated GSTA, GSTM, and NQO-1 protein contents as well
as that noted for GSTP (Fig. 5A). In addition, enzyme activity
toward 1-chloro-2,4-dinitrobenzene (Fig. 5B) and
2,6-dichloro-indophenol (Fig. 5C) was increased by DATS, LA, DHLA, and
SFN.
Discussion
The importance of GSTP in cancer prevention is supported by
the finding that mice lacking this detoxification enzyme have a
significantly increased incidence of
7,12-dimethylbenzanthra-cene-induced skin cancer (14). A point mutation in the GSTP
gene that leads to a decrease in enzyme activity has also been
reported to be associated with increased cancer risk of the oral
cavity, bladder, lung, testicles, larynx, and breast (34).
More-over, because GSTP can be induced by numerous dietary factors,
it is accepted that enhancement of GSTP expression and activity
through a dietary regimen is a practical means of cancer
chemoprevention. In fact, studies have shown that the
suppres-sion of benzo[a]pyrene-induced forestomach neoplastic
forma-tion in mice by garlic is positively related to the potency on
modulating the expression of the GSTP enzyme (13,35). Garlic
oil and garlic allyl sulfides, including diallyl disulfide and DATS,
which are regarded as potent chemopreventive agents, are
FIGURE 4 Activation of GPEI binding ac-tivity by phytoderived organosulfur com-pounds. (A) Cells were treated with 200 mmol/L DATS, 600 mmol/L LA, 600 mmol/L DHLA, or 5 mmol/L SFN for the indicated times and nuclear extracts were prepared to measure GPEI binding activity by EMSA. Free probe at the bottom is not shown. (B) Nuclear proteins isolated from the cells treated with DATS for 3 h and LA, DHLA, SFN for 1 h were first added with GPEI oligonucleotides into each reaction for 30 min and were then incubated with antibodies to c-Jun for an additional 30 min at room temperature. The subsequent super-shift complexes were separated by 6% acrylam-ide gel electrophoresis. Aliquots of the su-pernatant after immunoprecipitation with anti-Nrf2 antibody were used for EMSA. (C) Nuclear extracts isolated from the cells treated with DATS for 3 h and LA, DHLA, and SFN for 1 h were subjected to immunoprecipitation (IP) with anti-Nrf2 or anti-c-Jun antibody. Aliquots of pellet after IP (15mg) were used for immunoblotting (IB) with anti-Nrf2 or anti-phospho-c-Jun antibody. The results shown are representative of 4 experiments.FIGURE 5 Protein expression and enzyme activity of the phase II detox-ification enzymes induced by organo-sul fu r compo unds. C ell s were cultured with 0.1% DMSO alone (2) or with various concentrations of DATS, LA, DHLA, or SFN for 24 h. (A) GSTA, GSTM, and NQO1 protein were determined by immunoblot as-say. A total of 8mg protein for each sample was applied for electrophore-sis. The protein was quantified by densitometry and the level in control cells was set 1. Values are means (SD),n = 3. Means without a common letter differ, P , 0.05. Cells were cultured with 50 mmol/L DATS, 600 mmol/L LA, 600 mmol/L DHLA, or 5 mmol/L SFN for 24 h to determine GST (B) and NQO1 (C) activity. Values are means6 SD, n = 3–4. Groups without a common letter differ,P , 0.05.
effective GSTP inducers in the small intestine, liver, and lung
(36). In this study, our results showed that organosulfur
compounds from vegetables other than garlic also act as GSTP
inducers with a differential potency. Moreover, we further
showed that such upregulation of GSTP gene transcription by
these organosulfur compounds is likely to be AP-1 and Nrf2
dependent.
In this study, LA, DHLA, and SFN dose dependently
increased GSTP protein in Clone 9 cells (Fig. 1). Of the
organosulfur compounds tested, SFN showed the greatest
potency in upregulating GSTP expression, followed by DATS,
whereas LA and DHLA were least effective. Such a discrepancy
among organosulfur compounds is consistent with their
differ-ential increase in GSTP mRNA and enzyme activity (Fig. 2).
Moreover, the LA, DHLA, and SFN treatments produced
relatively greater induction over the controls in the GSTP
protein than in the GSTP mRNA or enzyme activity. This might
be related to unique regulation of GSTP mRNA stability and (or)
posttranslational mechanisms involving proteasomal
degrada-tion of newly synthesized GSTP proteins by these compounds
(37). It is of interest to understand how these organosulfur
compounds differentially regulate GSTP gene expression.
Al-though no explanation for this finding is currently available, the
differential pharmacologic properties of these organosulfur
compounds in liver cells may be a possible explanation (38).
Our findings suggest that the upstream signaling activating
AP-1 and Nrf2 is likely to play a key role in the differential GSTP
gene transcription.
To demonstrate the working mechanism by which the
organosulfur compounds upregulate GSTP transcription, we
constructed Luc-reporters with serial deletion of the 59-flanking
region of the GSTP gene promoter. These results clearly showed
that the section from
22713 to 22605 bp is required for LA,
DHLA, and SFN induction of GSTP expression in Clone 9 cells
(Fig. 3). However, the second enhancer GPEII (22604 to 22376
bp), which is adjacent to the GPEI, had no influence on the
induction of the GSTP gene. This finding is consistent with the
work of Okuda et al. (16), who reported that GPEI is the main
regulatory element responsible for GSTP induction. The
published evidence suggests that AP-1 is the main transcription
factor that binds to the TRE-like element in GPEI (17). AP-1 is
mainly composed of c-Jun and c-Fos protein dimers. The results
of our super-shift assay in the present study clearly indicated that
c-Jun was involved in the formation of the nuclear protein-GPEI
complexes induced by LA, DHLA, and SFN (Fig. 4B).
In addition to AP-1, several other transcription factors have
been reported to participate in the upregulation of GSTP
expression. In undifferentiated F9 embryonic stem cells, which
possess very low AP-1 activity, the GPEI element is active in an
AP-1-independent fashion (39). Nrf2 is one of the transcription
factors that attracts a lot of attention because of the sequence
homology between the TRE-like sequences on GPEI
(59-AGTCAGTCACTATGATTCAGCA-3
9) and the conserved
se-quences of the ARE (59-GTGACNNNGCA-39). Binding of
Nrf2/MafK to the GPEI and upregulation of rat GSTP
expres-sion were shown during hepatocarcinogenesis (18). However, by
treating RL34 liver epithelial cells with
15-deoxy-D-prostaglan-din j2 (12,14), Nrf2 was thought to not be an important
component responsible for transactivation of GPEI (40).
Al-though the role of Nrf2 in modulating GSTP expression in rats
is not well established, the importance of Nrf2 in regulating
human and mouse GSTP gene transcription has been well
documented (41,42). For instance, GSTP induction by
6-methylsulfinylhexyl isothiocyanate of wasabi, an analogue of
SFN, was shown to be completely abrogated in Nrf2-deficient
mice (43). To verify whether Nrf2 binds to the GPEI, we
performed an assay combining immunoprecipitation and
EMSA. Our results clearly showed that, in addition to AP-1,
Nrf2 is likely to bind to the GPEI. c-Jun has been shown to be a
binding factor in the activation of ARE-dependent transcription.
Nrf2 in association with Jun proteins regulate ARE-mediated
expression and coordinated induction of genes encoding
detox-ifying enzymes (44). The findings of a recent work by Levy et al.
(45) support that c-Jun seems to be a partner of Nrf2 in the
upregulation of ARE expression in human bronchial epithelial
cells exposed to 4-hydroxy-2-nonenal, although the response
varies with genes and cell types determined. In this study, the
immunoprecipitation result showed that Nrf2 may not bind
directly with c-Jun. Taken together, the EMSA results revealed
that the upregulation of this phase II detoxification enzyme by
LA and SFN is likely via multiple protein factors, at least c-Jun
and Nrf2, that may act in a complex manner.
In response to numerous prooxidants and electrophilics, Nrf2
dissociates from Keap protein and quickly translocates from the
cytosol into the nucleus, where it forms a heterodimer with small
Maf and binds to the ARE. This binding of Nrf2 to the ARE
upregulates the transcription of many cytoprotection enzymes.
These include glutamate cysteine ligase, heme oxygenase 1,
NQO1, and GST (21,27,43,46). In many types of cells, SFN has
been regarded as a potent Nrf2 activator that leads to
upregulation of NQO1 and GST isozymes including GSTA
and GSTM (43,47,48). This increase in the levels of those
detoxification enzymes accounts for, at least in part, the
protection by SFN against chemical carcinogens such as benzo
[a]pyrene-induced stomach and colon tumor formation (20,49).
In the present study, an increase in GSTA, GSTM, and NQO1
was also noted in cells treated with SFN, which suggests that the
Nrf2-ARE pathway was activated by treating Clone 9 cells with
SFN (Fig. 5).
LA, in addition to its well-recognized role in acting as a
coenzyme, is a natural antioxidant (24). LA is promptly taken up
by cells, where it can be reduced to DHLA by enzymes such as
dihydrolipoamide dehydrogenase, glutathione reductase, or
thioredoxin reductase. DHLA produced inside the cell is a
powerful reducing agent that can even reduce protein disulfides
to protein sulfhydryls and also reduce cystine to cysteine, which
is the limiting substrate for glutathione synthesis (50). Several in
vivo studies have further provided evidence that LA
supplemen-tation decreases oxidative stress and restores reduced levels of
other antioxidants under various physiologic and
pathophysio-logic conditions in brain and heart tissues and in RBC (51,52). In
addition to acting as a coenzyme and antioxidant, recent work
indicates that LA may also act as an inducer of several phase II
detoxification enzymes, including GSTA and NQO1 through a
CCAAT/enhancer binding protein and Nrf2-dependent pathway
(8). In this study, we further showed that LA and DHLA activate
AP-1 and Nrf2 translocation into the nucleus, where they bind
to GPEI and upregulate GSTP transcription.
In summary, SFN, LA, DHLA, and DATS are effective
inducers of GSTP gene transcription, and SFN shows the
greatest potency. Moreover, AP-1 and Nrf2 binding to the
enhancer element GPEI is essential for the induction of this
phase II detoxification enzyme.
Acknowledgments
C.K.L., H.W.C., and C.W.T. designed research; C.W.T., K.L.L.,
and C.K.L. conducted research; Y.P.C., and A.H.L. analyzed
data; C.K.L. and C.W.T. wrote the paper. C.W.T. had primary
responsibility for final content. All authors read and approved
the final manuscript.
Literature Cited
1. Moriarty RM, Naithani R, Surve B. Organosulfur compounds in cancer chemoprevention. Mini Rev Med Chem. 2007;7:827–38.
2. Higdon JV, Delage B, Williams DE, Dashwood RH. Cruciferous vegetables and human cancer risk: epidemiologic evidence and mech-anistic basis. Pharmacol Res. 2007;55:224–36.
3. Howard EW, Ling MT, Chua CW, Cheung HW, Wang X, Wong YC. Garlic-derived S-allylmercaptocysteine is a novel in vivo antimetastatic agent for androgen-independent prostate cancer. Clin Cancer Res. 2007; 13:1847–56.
4. Wang L, Gaziano JM, Norkus EP, Buring JE, Sesso HD. Associations of plasma carotenoids with risk factors and biomarkers related to cardiovascular disease in middle-aged and older women. Am J Clin Nutr. 2008;88:747–54.
5. Ogunleye AA, Xue F, Michels KB. Green tea consumption and breast cancer risk or recurrence: a meta-analysis. Breast Cancer Res Treat. 2010;119:477–84.
6. Jones SB, Brooks JD. Modest induction of phase 2 enzyme activity in the F-344 rat prostate. BMC Cancer. 2006;6:62.
7. Hanlon N, Okpara A, Coldham N, Sauer MJ, Ioannides C. Modulation of rat hepatic and pulmonary cytochromes P450 and phase II enzyme systems by erucin, an isothiocyanate structurally related to sulfora-phane. J Agric Food Chem. 2008;56:7866–71.
8. Ki SH, Kim SG. Phase II enzyme induction by alpha-lipoic acid through phosphatidylinositol 3-kinase-dependent C/EBPs activation. Xenobio-tica. 2008;38:587–604.
9. Villa-Cruz V, Davila J, Viana MT, Vazquez-Duhalt R. Effect of broccoli (Brassica oleracea) and its phytochemical sulforaphane in balanced diets on the detoxification enzymes levels of tilapia (Oreochromis niloticus) exposed to a carcinogenic and mutagenic pollutant. Chemosphere. 2009;74:1145–51.
10. Strange RC, Spiteri MA, Ramachandran S, Fryer AA. Glutathione-S-transferase family of enzymes. Mutat Res. 2001;482:21–6. 11. Satoh K, Kitahara A, Soma Y, Inaba Y, Hatayama I, Sato K.
Purification, induction, and distribution of placental glutathione trans-ferase: a new marker enzyme for preneoplastic cells in the rat chemical hepatocarcinogenesis. Proc Natl Acad Sci USA. 1985;82:3964–8. 12. Tsuchida S, Sato K. Glutathione transferases and cancer. Crit Rev
Biochem Mol Biol. 1992;27:337–84.
13. Hu X, Benson PJ, Srivastava SK, Xia H, Bleicher RJ, Zaren HA, Awasthi S, Awasthi YC, Singh SV. Induction of glutathione S-transferase pi as a bioassay for the evaluation of potency of inhibitors of benzo[a]pyrene-induced cancer in a murine model. Int J Cancer. 1997;73:897–902. 14. Henderson CJ, Smith AG, Ure J, Brown K, Bacon EJ, Wolf CR.
Increased skin tumorigenesis in mice lacking pi class glutathione S-transferases. Proc Natl Acad Sci USA. 1998;95:5275–80.
15. Sakai M, Okuda A, Muramatsu M. Multiple regulatory elements and phorbol 12-O-tetradecanoate 13-acetate responsiveness of the rat placental glutathione transferase gene. Proc Natl Acad Sci USA. 1988; 85:9456–60.
16. Okuda A, Imagawa M, Maeda Y, Sakai M, Muramatsu M. Structural and functional analysis of an enhancer GPEI having a phorbol 12-O-tetradecanoate 13-acetate responsive element-like sequence found in the rat glutathione transferase P gene. J Biol Chem. 1989;264:16919–26. 17. Angel P, Imagawa M, Chiu R, Stein B, Imbra RJ, Rahmsdorf HJ, Jonat C, Herrlich P, Karin M. Phorbol ester-inducible genes contain a common cis element recognized by a TPA-modulated trans-acting factor. Cell. 1987;49:729–39.
18. Ikeda H, Nishi S, Sakai M. Transcription factor Nrf2/MafK regulates rat placental glutathione S-transferase gene during hepatocarcino-genesis. Biochem J. 2004;380:515–21.
19. Zhu H, Jia Z, Strobl JS, Ehrich M, Misra HP, Li Y. Potent induction of total cellular and mitochondrial antioxidants and phase 2 enzymes by cruciferous sulforaphane in rat aortic smooth muscle cells: cytopro-tection against oxidative and electrophilic stress. Cardiovasc Toxicol. 2008;8:115–25.
20. Bonnesen C, Eggleston IM, Hayes JD. Dietary indoles and isothiocy-anates that are generated from cruciferous vegetables can both stimulate
apoptosis and confer protection against DNA damage in human colon cell lines. Cancer Res. 2001;61:6120–30.
21. Keum YS, Yu S, Chang PP, Yuan X, Kim JH, Xu C, Han J, Agarwal A, Kong AN. Mechanism of action of sulforaphane: inhibition of p38 mitogen-activated protein kinase isoforms contributing to the induction of antioxidant response element-mediated heme oxygenase-1 in human hepatoma HepG2 cells. Cancer Res. 2006;66:8804–13.
22. Wagner AE, Ernst I, Iori R, Desel C, Rimbach G. Sulforaphane but not ascorbigen, indole-3-carbinole and ascorbic acid activates the tran-scription factor Nrf2 and induces phase-2 and antioxidant enzymes in human keratinocytes in culture. Exp Dermatol. 2010;19:137–44. 23. Packer L, Witt EH, Tritschler HJ. alpha-Lipoic acid as a biological
antioxidant. Free Radic Biol Med. 1995;19:227–50.
24. Bustamante J, Lodge JK, Marcocci L, Tritschler HJ, Packer L, Rihn BH. Alpha-lipoic acid in liver metabolism and disease. Free Radic Biol Med. 1998;24:1023–39.
25. Biewenga GP, Haenen GR, Bast A. The pharmacology of the antiox-idant lipoic acid. Gen Pharmacol. 1997;29:315–31.
26. Jia Z, Hallur S, Zhu H, Li Y, Misra HP. Potent upregulation of glutathione and NAD(P)H:quinone oxidoreductase 1 by alpha-lipoic acid in human neuroblastoma SH-SY5Y cells: protection against neurotoxicant-elicited cytotoxicity. Neurochem Res. 2008;33:790–800. 27. Elangovan S, Hsieh TC. Control of cellular redox status and upregulation of quinone reductase NQO1 via Nrf2 activation by alpha-lipoic acid in human leukemia HL-60 cells. Int J Oncol. 2008; 33:833–8.
28. Tsai CW, Yang JJ, Chen HW, Sheen LY, Lii CK. Garlic organosulfur compounds upregulate the expression of the pi class of glutathione S-transferase in rat primary hepatocytes. J Nutr. 2005;135:2560–5. 29. Tsai CW, Chen HW, Yang JJ, Sheen LY, Lii CK. Diallyl disulfide and
diallyl trisulfide up-regulate the expression of the pi class of glutathione S-transferase via an AP-1-dependent pathway. J Agric Food Chem. 2007; 55:1019–26.
30. Wu CC, Sheen LY, Chen HW, Kuo WW, Tsai SJ, Lii CK. Differential effects of garlic oil and its three major organosulfur components on the hepatic detoxification system in rats. J Agric Food Chem. 2002;50: 378–83.
31. Lehmann U, Kreipe H. Real-time PCR analysis of DNA and RNA extracted from formalin-fixed and paraffin-embedded biopsies. Methods. 2001;25:409–18.
32. Mannervik B, Alin P, Guthenberg C, Jensson H, Tahir MK, Warholm M, Jornvall H. Identification of three classes of cytosolic glutathione transferase common to several mammalian species: correlation between structural data and enzymatic properties. Proc Natl Acad Sci USA. 1985; 82:7202–6.
33. Jamieson D, Wilson K, Pridgeon S, Margetts JP, Edmondson RJ, Leung HY, Knox R, Boddy AV. NAD(P)H:quinone oxidoreductase 1 and nrh: quinone oxidoreductase 2 activity and expression in bladder and ovarian cancer and lower NRH:quinone oxidoreductase 2 activity associated with an NQO2 exon 3 single-nucleotide polymorphism. Clin Cancer Res. 2007;13:1584–90.
34. Taningher M, Malacarne D, Izzotti A, Ugolini D, Parodi S. Drug metabolism polymorphisms as modulators of cancer susceptibility. Mutat Res. 1999;436:227–61.
35. Hu X, Benson PJ, Srivastava SK, Mack LM, Xia H, Gupta V, Zaren HA, Singh SV. Glutathione S-transferases of female A/J mouse liver and forestomach and their differential induction by anti-carcinogenic organosulfides from garlic. Arch Biochem Biophys. 1996;336:199–214. 36. Lii CK, Tsai CW, Wu CC. Garlic allyl sulfides display differential modulation of rat cytochrome P450 2B1 and the placental form glutathione S-transferase in various organs. J Agric Food Chem. 2006; 54:5191–6.
37. Cherng SH, Hsu SL, Yang JL, Yu CT, Lee H. Suppressive effect of 1-nitropyrene on benzo[a]pyrene-induced CYP1A1 protein expression in HepG2 cells. Toxicol lett. 2006;161:236–43.
38. Kasuga S, Uda N, Kyo E, Ushijima M, Morihara N, Itakura Y. Pharmacologic activities of aged garlic extract in comparison with other garlic preparations. J Nutr. 2001;131:S1080–4.
39. Okuda A, Imagawa M, Sakai M, Muramatsu M. Functional cooper-ativity between two TPA responsive elements in undifferentiated F9 embryonic stem cells. EMBO J. 1990;9:1131–5.
40. Kawamoto Y, Nakamura Y, Naito Y, Torii Y, Kumagai T, Osawa T, Ohigashi H, Satoh K, Imagawa M, et al. Cyclopentenone prostaglandins Organosulfur compounds and glutathioneS-transferase expression 7 of 8
as potential inducers of phase II detoxification enzymes. 15-deoxy-delta (12,14)-prostaglandin j2-induced expression of glutathione S-transfer-ases. J Biol Chem. 2000;275:11291–9.
41. Nishinaka T, Ichijo Y, Ito M, Kimura M, Katsuyama M, Iwata K, Miura T, Terada T, Yabe-Nishimura C. Curcumin activates human glutathione S-transferase P1 expression through antioxidant response element. Toxicol Lett. 2007;170:238–47.
42. Ikeda H, Serria MS, Kakizaki I, Hatayama I, Satoh K, Tsuchida S, Muramatsu M, Nishi S, Sakai M. Activation of mouse Pi-class glutathione S-transferase gene by Nrf2(NF-E2-related factor 2) and androgen. Biochem J. 2002;364:563–70.
43. Morimitsu Y, Nakagawa Y, Hayashi K, Fujii H, Kumagai T, Nakamura Y, Osawa T, Horio F, Itoh K. A sulforaphane analogue that potently activates the Nrf2-dependent detoxification pathway. J Biol Chem. 2002;277:3456–63.
44. Venugopal R, Jaiswal AK. Nrf2 and Nrf1 in association with Jun proteins regulate antioxidant response element-mediated expression and coordinated induction of genes encoding detoxifying enzymes. Onco-gene. 1998;17:3145–56.
45. Levy S, Jaiswal AK, Forman HJ. The role of c-Jun phosphorylation in EpRE activation of phase II genes. Free Radic Biol Med. 2009;47:1172–9. 46. Shenvi SV, Smith EJ, Hagen TM. Transcriptional regulation of rat gamma-glutamate cysteine ligase catalytic subunit gene is mediated through a distal antioxidant response element. Pharmacol Res. 2009; 60:229–36.
47. Gao X, Talalay P. Induction of phase 2 genes by sulforaphane protects retinal pigment epithelial cells against photooxidative damage. Proc Natl Acad Sci USA. 2004;101:10446–51.
48. Ritz SA, Wan J, Diaz-Sanchez D. Sulforaphane-stimulated phase II enzyme induction inhibits cytokine production by airway epithelial cells stimulated with diesel extract. Am J Physiol Lung Cell Mol Physiol. 2007;292:L33–9.
49. Fahey JW, Haristoy X, Dolan PM, Kensler TW, Scholtus I, Stephenson KK, Talalay P, Lozniewski A. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc Natl Acad Sci USA. 2002;99:7610–5.
50. Suh JH, Wang H, Liu RM, Liu J, Hagen TM. (R)-alpha-lipoic acid reverses the age-related loss in GSH redox status in post-mitotic tissues: evidence for increased cysteine requirement for GSH synthesis. Arch Biochem Biophys. 2004;423:126–35.
51. Ghibu S, Lauzier B, Delemasure S, Amoureux S, Sicard P, Vergely C, Muresan A, Mogosan C, Rochette L. Antioxidant properties of alpha-lipoic acid: effects on red blood membrane permeability and adaptation of isolated rat heart to reversible ischemia. Mol Cell Biochem. 2009;320:141–8.
52. Bilska A, Dubiel M, Sokolowska-Jezewicz M, Lorenc-Koci E, Wlodek L. Alpha-lipoic acid differently affects the reserpine-induced oxidative stress in the striatum and prefrontal cortex of rat brain. Neuroscience. 2007;146:1758–71.