發掘頑抗性急性血癌的免疫療法新標的:發現血癌表面蛋白質表現異常的可能機制
30
0
0
全文
(2) 目. 錄. 中文摘要 ..................................................................................................................... 3 英文摘要 ..................................................................................................................... 4 第一章 INTRODUCTION .......................................................................................... 6 第二章 METHODS AND MATERIALS 第一節 Overall Scheme ........................................................................................... 7 第二節 Clinical Samples, Cell Line and Sample Preparation ................................. 8 第三節 Membrane Protein Extraction and Digestion.............................................. 9 第四節 TMT 10 Plex Mass Tag Labeling ............................................................. 10 第五節 High pH Reversed-Phase Peptide Fractionation ....................................... 11 第三章 RESULTS 第一節 Membrane proteome of the undifferentiated acute leukemia cell line MV4-11 through biotin-streptavidin capture .......................................................... 12 第二節 Optimizing membrane protein extraction—substitution of biotin-streptavidin capture for sucrose gradient enrichment .................................. 14 第三節 Optimizing membrane protein extraction—substitution of DDMIC for SDS as the detergent in cell lysis ............................................................................ 14 第四節 Precise analysis of membrane proteome in clinical samples of refractory acute myeloid leukemia .......................................................................................... 15 第五節 Highly overlapped identified proteins between PBMC and refractory blasts ....................................................................................................................... 17 第六節 Aberrant protein abundance distribution in refractory leukemic blasts .... 18 第七節 Analysis of normal PBMC revealed the physiological representation ..... 19 第八節 Dysregulated membrane proteome in refractory leukemic cells shows multi-functinality of its components ....................................................................... 20 第九節 Imbalanced vesicle-trafficking system in refractory leukemic blasts might lead to a mechanism leukemia-mediated dysregulation of surface proteins .......... 22 第四章 DISCUSSION 第一節 Discovering a potential mechanism of leukemia-mediated dysregulation of membrane proteins .................................................................................................. 25 第二節 Membrane proteome workflow had limitation in membrane protein identification ........................................................................................................... 27 第三節 Cells displayed diverse functions and morphologies by having a different. 1.
(3) membrane protein composition .............................................................................. 27 第五章 CONCLUSION AND PERSPECTIVES ......................................................... 28 參考文獻 ................................................................................................................... 29 . 2.
(4) 中文摘要 血液惡性腫瘤在這數十年來一直維持在台灣癌症死因的前十名, 而即使血 液惡性腫瘤的治療方法在持續進展中, 頑抗性急性血癌仍然是最難以治療的疾 病之一。此篇研究使用量化的蛋白質體學, 分析未分化急性血癌細胞株(MV4-11) 和頑抗性急性血癌病人臨床檢體的表面蛋白質, 以嘗試幫助頑抗性急性血癌的 未來醫療治療需求。在本研究中, 我們建立一蛋白質體分析流程, 以探尋頑抗性 急性血癌病人檢體的表面蛋白質體, 包含以流式細胞儀協助純化血癌芽細胞 (leukemic blasts), 選擇與優化萃取膜蛋白的介面活性劑與過程, 和以質譜儀進行 蛋白質的量化分析。 研究結果有兩大發現。我們在頑抗性急性血癌臨床檢體的分析中, 發現頑抗 性急性血癌臨床檢體和未分化急性血癌細胞株與正常人的周邊血液單核細胞 (peripheral blood mononuclear cells)相比, 有兩大重要不同。第一個發現是頑抗性 急性血癌的表面蛋白質體與其他兩種細胞相比, 不是由數種主要的蛋白質構成, 而是有許多相異, 並少許的蛋白質共同構成, 更精確來說, 佔表面蛋白質總體量 的一半量的蛋白質種類, 在細胞株 MV4-11, 人類周邊血液單核細胞, 與頑抗性 急性血癌病人臨床檢體分別是 5,9 和 26。頑抗性急性血癌表面蛋白質的分布分 常特別。第二個發現是頑抗性急性血癌有個特別增量的表面蛋白質-囊泡相關 表面蛋白質 8 (vesicle-associated membrane protein 8), 簡稱 VAMP8。VAMP8 在 過去的研究中, 被認為正常細胞生理中的囊泡運輸系統有重要的角色, 其在細 胞膜與體上表現的不平衡, 會影響細胞的囊泡運輸系統, 進而影響血癌表面蛋 白質的表現與分布。我們認為這項發現會幫助理解頑抗性急性血癌中蛋白質從 製造到表現, 與膜蛋白分布異常的可能機制, 或許能夠更加理解頑抗性急性血 癌的癌化, 與難以治療的頑抗本質的原因和未來治療方向。 總結來說, 我們建立一個精簡的表面蛋白質體分析流程, 能夠將頑抗性急性 血癌從臨床檢體的純化, 濃縮, 萃取, 一路做到量化蛋白質體分析, 並發現血癌 表面蛋白質表現異常的可能機制。. 3.
(5) 英文摘要 Hematological malignancies have stayed as the top 10 death-causing neoplasms in Taiwan through decades. Even with the progress in therapy, refractory acute leukemia remains one of the most challenging diseases to treat. To address this unmet medical need, a quantitative proteomics approach was adopted to characterize the membrane proteome of an undifferentiated acute leukemia cell line and the refractory acute leukemia clinical sample. For the research, we have built up a workflow to interrogate the membrane proteome of refractory acute leukemia patient’s clinical sample, including flow cytometry-based purification of leukemic blasts, optimizing the choice of detergent to extract membrane proteins and downstream mass spectrometry-based quantitative proteomics analyses. Interestingly, from the clinical sample of refractory acute leukemia, we have two major discoveries that were not seen in undifferentiated acute leukemia cell line MV4-11 nor in normal peripheral blood mononuclear cells (PBMC). First, the membrane proteome of refractory acute leukemia was not predominated by a few abundant proteins. Specifically, half of the membrane proteome in MV4-11, normal PBMC and refractory acute leukemia were predominated by 5, 9 and 26, respectively. The distribution of protein abundance in refractory acute leukemia was unique. Second, we found the most upregulated protein in refractory acute leukemia sample was vesicle-associated membrane protein 8 (VAMP8) whose imbalance had been reported to undermine the physiological vesicle trafficking system in normal cells. This finding might pave the way to understanding more about how heavily treated refractory acute leukemia cells dysregulate their protein transport as well as the expression of surface proteins. In summary, we have established a streamlined workflow to enrich membrane proteins for refractory acute leukemia clinical samples for mass spectrometry-based quantitative proteomics analysis and have found a potential mechanism about how these cells dysregulate their surface protein expression. Key Words: refractory acute leukemia, membrane proteome, flow cytometry-based purification, mass spectrometry-based quantitative proteomics, vesicle-associated membrane protein 8, vesicle trafficking system. 4.
(6) 圖目錄 包括各章節之圖及其所在之頁數 若圖擷取自參考文獻,則須標註來源 Figure 1: The overall scheme of this project.....…………………………………... 8 Figure 2: The processing of refractory acute leukemia clinical sample and normal peripheral blood mononuclear cell control..………………………………………. 9 Figure 3: The composition of membrane proteome from MV4-11..……………… 13 Figure 4: The advantage from DDMIC for membrane protein extraction compared with SDS in terms of peptide and protein identifications.………………………… 15 Figure 5: The comparison of identifiable proteins between normal peripheral blood mononuclear cells and refractory leukemic blasts….……………………………... 16 Figure 6: The compositions of membrane proteome in refractory leukemic blasts, normal PBMC and MV4-11. ……………………………………………………... 19 Figure 7: A close-up analysis for normal PBMC membrane proteome…………... 20 Figure 8: A close-up analysis for refractory leukemic blast membrane proteome... 21 Figure 9: The quantification overview of the comparison between refractory leukemic blats and normal PBMC…………………………………………………………… 22 Figure 10: The comparison of VAMP8 expression between normal PBMC and refractory leukemic blasts.………………………………………………..……….. 24 Figure 11: The comparison of VAMP8 expression between normal PBMC and refractory leukemic blasts.………………………………………………..……….. 24. 5.
(7) 論文正文 INTRODUCTION Acute leukemia is the outgrowth of hematopoietic cells as a result of blocked differentiation and unchecked proliferation. Through decades in Taiwan, hematopoietic malignancies including leukemia and lymphoma stand as the top ten most fatal neoplasms. Acute myeloid leukemia (AML) is the most prevalent type of acute leukemia in adults. The current mainstay for treating AML is high dose cytotoxic chemotherapy and hematopoietic stem cell transplantation. With the advent of novel therapies such as target therapy and monoclonal antibodies, however, refractory/relapsed AML still occur in approximately half of the patient population. In the past five years, at National Taiwan University Hospital, among 200 patients receiving high dose chemotherapy followed by hematopoietic stem cell transplantation, more than 100 patients developed relapse, and the majority of them turned refractory to further therapy. This is a tremendous unmet medical need affecting many patients even only at their twenties. Therefore, it is mandatory to seek new therapeutic targets for this disease. The genetic aberrations of refractory AML patients differ vastly from normal cells, and hence it is not feasible to apply a universal treatment strategy in hope for covering all subtypes. In addition, only drugs with lipid-solubility or certain attributes can pass through cell membrane to target intracellular molecules. Membrane proteins therefore serve as hold promises for therapy development. Meanwhile, it is rational to conceive that, through rounds of chemotherapies, refractory acute leukemic cells have evolved a mechanism to escape attacks from cytotoxic agents as well as downplaying their immunogenicity to evade immunosurveillance. To harness the power of cancer immunotherapy which hinges on recognition of 6.
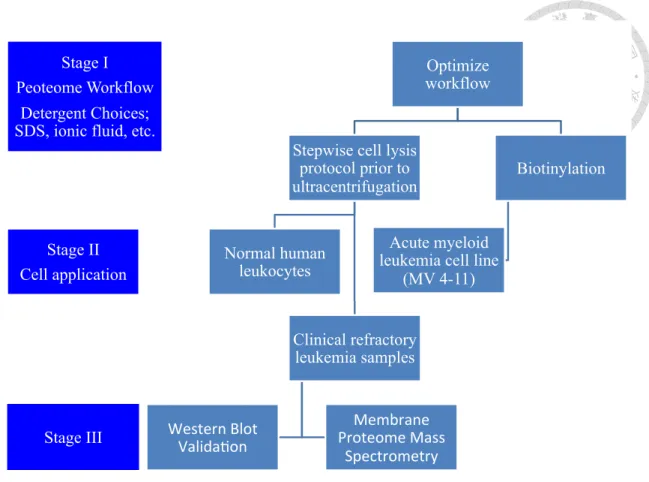
(8) surface antigens, we decided to characterize and identify novel surface proteins of refractory acute leukemia. This not only helps understand how these cells altered their surface protein expression but also potentially provides an avenue for future therapy to target novel leukemic antigens. The goal of this project is to employ an unbiased quantitative proteomics to measure and characterize the membrane proteome of refractory acute leukemia in hope for identifying dysregulated and targetable surface proteins.. METHODS & MATERIALS Overall Scheme The approach of this project was divided into 3 stages and is shown in Figure 1 below. Stage I is the optimization of the workflow. First, we sought to optimize the workflow to enrich membrane proteins. We set up biotin-streptavidin capturing method for AML cell-line MV4-11, and adopted a stepwise cell lysis protocol prior to centrifugation for normal human leukocytes -peripheral blood mononuclear cells and clinical samples. We also explored the possibility of an ionic-fluid compound as the alternative detergent in order to boost the identification rate of membrane proteins. Stage II is the application of the optimized workflow to not only cell line, PBMC, but also clinical refractory leukemia samples. We used flow cytometry to sort leukemic blasts from clinical samples for the downstream analyses. After successful identification of specific overexpressed proteins, we hope to validate the findings in much more clinical samples, and by western blot validation and membrane proteome analysis through mass spectrometry for further immunotherapy application as our Stage III approach.. 7.
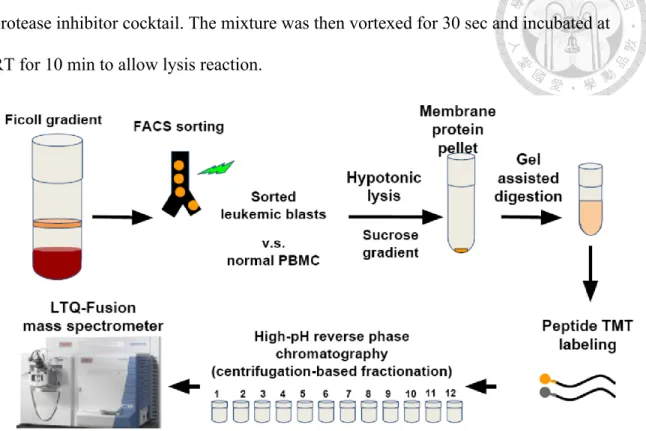
(9) Stage I Peoteome Workflow. Optimize workflow. Detergent Choices; SDS, ionic fluid, etc. Stepwise cell lysis protocol prior to ultracentrifugation. Stage II Cell application. Normal human leukocytes. Acute myeloid leukemia cell line (MV 4-11). Clinical refractory leukemia samples. Stage III Stage III. Biotinylation. Membrane( Proteome(Mass( Spectrometry. Western(Blot( Valida7on(. Figure 1. The overall scheme of this project. Three stages are planned to carry out in a stepwise fashion.. Clinical Samples, Cell Line and Sample Preparation The cell line used in the experiment was undifferentiated acute leukemia MV4-11 which bears myeloid and lymphoid markers at the same time, originally from ATCC and confirmed by STR analyses. The normal PBMC were from a healthy volunteer. The refractory acute leukemia sample was collected from a patient with acute myeloid leukemia with minimal differentiation (French American British classification M0), who had failed the induction chemotherapy idarubicin plus cytarabine, vincristine and clofarabine and signed the informed consent per IRB of NTUH. Leukemic blasts bear the surface expression of CD34 and were sorted using flow cytometry-based enrichment to attain >99% purity as the figure below. To prepare the sample, cells were lysed by 8.
(10) hypotonic buffer (10 mM HEPES, pH 7.5, 1.5 mM MgCl2, 10 mM KCl) 200 ul with the protease inhibitor cocktail. The mixture was then vortexed for 30 sec and incubated at RT for 10 min to allow lysis reaction.. Figure 2. The processing of refractory acute leukemia clinical sample and normal peripheral blood mononuclear cell control. Tryptic peptides were labeled in vitro using tandem mass tags (TMT) coupled with the downstream fractionation and LC-MS/MS analyses.. Membrane Protein Extraction and Digestion The protocol we used for membrane proteins extraction and digestion was modified from Dimayacyac-Esleta, et al. (2015) A two-step centrifugation process was used to isolate membrane proteins. Whole-cell proteins were centrifuged at 3000X g, at 4°C for 10 min. Supernatant, which contains membrane proteins, was then mixed with 1.8 M sucrose to sucrose concentration of 0.25M, and centrifuged at 13000X g, 4°C for 60 min. After centrifugation, membrane proteins were in pellet. Pellet was washed twice, with 1ml 0.1M NH4HCO3 and 13000X g, 4°C, 60 min centrifugation. After. 9.
(11) centrifugation, supernatant was removed and pellet was stored at -20°C. Most part of the protocol of membrane protein digestion was adopted from Dimayacyac-Esleta, et al. (2015) gel-assisted digestion method, with different types of digestion buffer (including SDS and ionic fluid DDMIC). Ionic fluid was modified based on Zhao, et al. (2014). The steps were as follows. 50ul of lysis buffer (6 M Urea, 0.1 M TEABC, 5mM EDTA, 2%SDS/ 6 M Urea, 0.1 M TEABC, 5mM EDTA, 2%UR-DDMIC/ Digitonin, Triton X100 buffer/ MemNS) was added respectively to membrane protein pellet. Lysis buffer and membrane proteins were then mixed and incubated for 30 min at 37°C. After incubation, 1.28 ul of 200 mM TCEP (tris(2-carboxyethyl)phosphine)and 0.52ul of 200mM MMTs were added to respectively chemically reduce and alkylate proteins, at room temperature for 30 min. Proteins were incorporated into a polyacrylamide gel directly in the sample tube by adding 2.5 ul of 10% APS, 24.6ul of 30% acrylamide solution, and 1ul of 100% TEMED. Cut the gel into small pieces, wash them with 1ml 25mM TEABC in 50%ACN twice, and lastly dehydrate with 100% acetonitrile (ACN). The gel with membrane proteins was incubated with trypsin in 25mM TEABC (protein: trypsin = 10:1 g/g) at 37°C for 16hr. 25 mM TEABC, 0.1% TFA, 0.1% TFA in 50% ACN and 100% ACN were sequentially used to wash the gel to extract tryptic peptides. The peptides were dried by SpeedVac machine.. TMT 10 Plex Mass Tag Labeling For peptides labeling, we used the TMT10plex Mass Tag Labeling Kits and Reagents from Thermo Scientific. Before use, TMT Label Reagents are equilibrated to room temperature. Add 41 uL of anhydrous acetonitrile to 0.8 mg vials and vortex occasionally for 5 min to let the reagent dissolve. 41uL of the TMT Label Reagent was 10.
(12) added to sample and was incubated at RT for 1hr. To quench the reaction, 8ul of 5% hydroxylamine was incubated with the sample for 15 min. Lastly, samples were combined in equal amounts in a new microcentrifuge tube and stored at -80°C.. High pH Reversed-Phase Peptide Fractionation For peptide fractionation, we used the Pierce High pH Reversed-Phase Peptide Fractionation Kit from Thermo Scentific. The kit was divided into two parts: conditioning of the spin column and fractionation of digest samples. The spin column was first centrifuged at 5000X g for 2 min so that the resin material was packed and solution was removed. 100% ACN, 0.1% TFA solution were respectively used (0.1% TFA solution twice) to condition the column by centrifugation at 5000X g for 2 minutes and removal of the solutions. Elution solutions of different concentration (5,10,12.5,15,17.5,20,22.5,25,50%) of ACN in 0.1% triethylamine were prepared before fractionation. 10-100µg of digested sample was dissolved in 300µL of 0.1% TFA solution and loaded onto the column. After centrifugation at 3000X g for 2 min, eluate was retained as the “flow-through” fraction. Water was loaded onto the column and after centrifugation at 3000X g for 2 min, eluate was retained as the “washed” fraction. The column was washed by solution of 5% ACN and 0.1% TFA to remove unreacted TMT reagent. The column was placed in a new 2.0 mL sample tube, loaded with elution solution (ACN in TFA), and centrifuged at 3000X g for 2 min. Eluate was collected as peptide fraction. Repeat the previous step with different concentrations of elution solutions sequentially to collect different gradient fractions. The liquid contents of each sample tube were evaporated to dryness by SpeedVac concentrator. Before LC-MS analysis, dry samples were re-suspended in 0.1% formic acid (FA). 11.
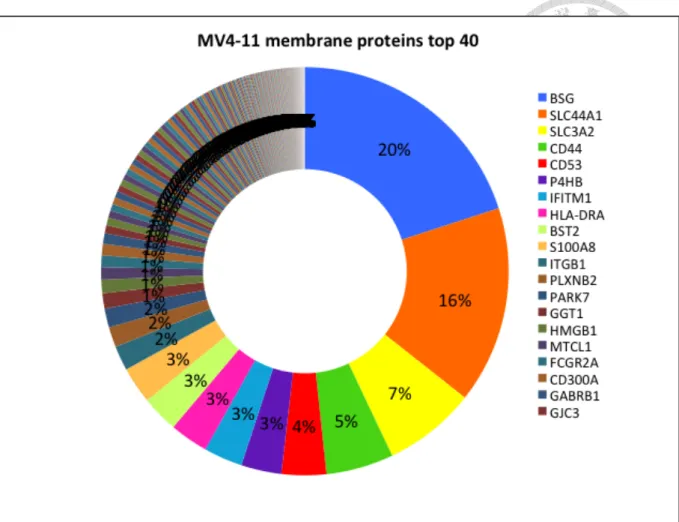
(13) RESULTS Membrane proteome of the undifferentiated acute leukemia cell line MV4-11 through biotin-streptavidin capture MV4-11 Cells were biotinylated by EZ-LinkTM Sulfo-NHS-SS-Biotin prior to cell lysis. After membrane were lysed, membrane proteins were separated from intracellular proteins and enriched through the biotin-streptavidin interaction. The average membrane protein from 20 million cells by biotinylation was 13.05µg, and about 272 kinds of proteins were identified. Figure3 showed the most abundant membrane proteins of MV4-11 cells. Five of the most abundant membrane proteins made up more than 50% of the total membrane protein abundance, while ten and twenty of the most abundant membrane proteins made up 60-70% and more than 75% of the total membrane proteins respectively. The abundance of different membrane proteins varied greatly, and membrane protein composition was dominated by several most abundant proteins. Several of the most abundant membrane proteins included BSG (CD147), SLC44A1 (CD92), SLC3A2 (CD98), CD44, CD53, etc.. 12.
(14) Figure 3. The composition of membrane proteome from MV4-11.. BSG (CD147) made. up 20% of the total membrane proteins. It bound to integrins and facilitated cell adhesion and activation, which participated in immunity and memory development. SLC44A1 (CD92), which made up 16%, assisted choline transport for membrane phospholipids synthesis and also had a cytoplasmic ITIM that might participate in inhibitory signaling. Other membrane proteins, such as SLC3A2 (CD98), CD44, CD53 had important roles in adhesion, including adhesion between immune cell-cell, lymphocytes- high endothelial venules (HEV), adhesion-activation functions, lymphoid–stroma interactions, etc, and other roles in signal transduction. Biotinylation workflow and Mass Spectrometry allowed us to probe into cell membrane proteome. We could learn about the membrane protein composition, abundant cells, their possible functions and mechanism from membrane proteome. 13.
(15) Optimizing membrane protein extraction —substitution of biotin-streptavidin capture for sucrose gradient enrichment Biotinylation workflow yielded about 13.05µg membrane proteins from 20 million cells. Biotinylation could get relatively pure membrane proteins by the specific binding between Biotin and Streptavidin. However, the binding efficiency of biotin with membrane proteins, biotin with streptavidin beads, and elution process were more difficult to maintain stability and therefore resulted in experiment variants. It created not only more complication for carrying out experiments, but also difficulty in replicating experimental result. Another setback of biotinylation was its low protein yield. Although biotinylation might have higher specificity in protein yield, its binding process resulted in protein loss. For membrane proteins to be successfully processed through mass spectrometry, a minimal amount of proteins was required. Taking the limited and much less amount of cells from clinical samples into consideration, biotinylation was not the best option. To get more protein yield from cells, we turned to sugar gradient enrichment method. Sugar gradient enrichment method might have higher possibility in extracting non-cell membrane proteins, but it had much higher protein yield. Sucrose gradient enrichment had more membrane protein yield than biotinylation, and therefore was chosen as main membrane protein enrichment method for PBMCs and blasts from clinical sample.. Optimizing membrane protein extraction —substitution of DDMIC for SDS as the detergent in cell lysis In order to get the most abundant membrane proteins, we sought ways to optimize our membrane protein purification and extraction workflow. Zhao, et al. (2014). showed that an ionic fluid, 1-Dodecyl-3-Methylimidazolium Chloride (DDMIC), had 1.4 and 14.
(16) 3.5 times improvement on the integral membrane proteins and hydrophobic peptide number identification comparing to the SDS-assisted method. Therefore, we tried to compare the final peptides and proteins identification of SDS and DDMIC in Dimayacyac-Esleta, et al. (2015) gel-assisted digestion method. In gel-assisted digestion, PBMCs were digested by digestion buffer containing either SDS or DDMIC. All the other steps were the same for the samples. Figure4 showed the final result of peptides and proteins identification for SDS and DDMIC. Digestion buffer containing DDMIC had better identification than that containing SDS. DDMIC worked better than SDS as a detergent for membrane proteins lysis buffer and therefore DDMIC replaced SDS in digestion buffer for the following experiments in the PBMCs and clinical samples.. Figure 4.. The advantage from DDMIC for membrane protein extraction compared. with SDS in terms of peptide and protein identifications.. Precise analysis of membrane proteome in clinical samples of refractory acute myeloid leukemia 15.
(17) Our samples came from two different individuals. The normal PBNC were from a healthy volunteer, while the blasts came from a male refractory acute myeloid leukemia (AML) patient. Mononuclear cells from refractory AML patient’s clinical sample were separated and collected by Ficoll-based cell separation media and centrifugation. Then peripheral blood mononuclear cells and blasts were sorted by CD34 through flow cytometer. Blasts, which didn’t have CD34, were separated from lymphocytes, which expressed CD34 in their membrane. Initially, we planned to analyze the membrane proteins of the patient’s lymphocytes. After the flow cytometer, the amount of sorted lymphocytes did not meet the protein amount required for membrane proteome and therefore we had to abandon the original plan of analyzing the lymphocytes. PBMCs and blasts were processed through cell lysis, sucrose gradient enrichment, membrane protein extraction and digestion, TMT-labeling, and Hp-RP fractionation methods. Surprisingly, PBMCs had three times the amount of identified membrane protein as blasts as shown in Figure 5. Our postulation for this result is that refractory leukemic cells might modify their membrane proteins in a vast extent, such as extensive glycosylation, that has surpassed our current proteomics search algorithm.. 16.
(18) Figure 5.. The comparison of identifiable proteins between normal peripheral blood. mononuclear cells and refractory leukemic blasts.. The identified protein number and. abundance derived from processing the Mass Spectrometry data through UniProtKB/Swiss-Prot database. Identified protein abundance showed the abundance of proteins identified by the database. The mass difference of identified protein abundance between PBMCs and blasts might have two possible reasons. First was that the current database had limitation on whole-membrane protein identification. The second reason was the dysregulation of membrane proteins in blasts. It showed that the blasts only displayed about 30% of the amounts of membrane proteins in PBMCs.. Highly overlapped identified proteins between PBMC and refractory blasts As shown in Figure 5 above, identified protein number showed the total protein species identified by the database. PBMCs and blasts had similar protein species identification, with PBMCs having identification of only three species more than blasts. This was a really important fact, which pointed out a possible obstacle we had to overcome in our project. The identified protein abundance of PBMCs was three times more than that of blasts, whereas the identified protein number of PBMCs and blasts was almost the same. It was therefore reasonable to assume that even if we manage to increase protein abundance three times more in the future, it was very likely that the identified protein number would only increase slightly. From this data, we anticipated that if we want to identify much more protein species, we have to increase the amount of membrane proteins by ten times and possibly much more. The Venn Diagram in Figure 5 of identified protein species in PBMC and blasts. The total 195 protein species identified in blasts were all included in the 198 species identified in PBMCs. This indicated that if blasts or PBMCs had unique proteins that 17.
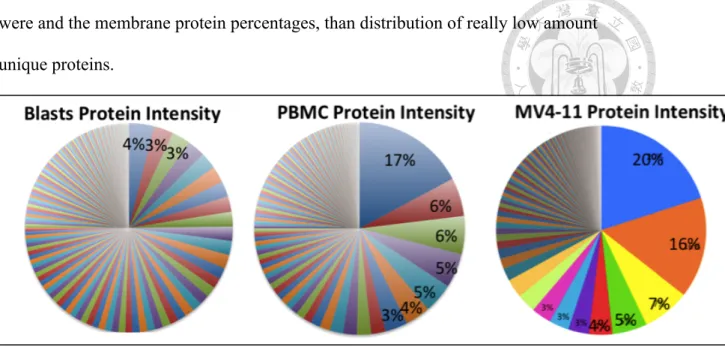
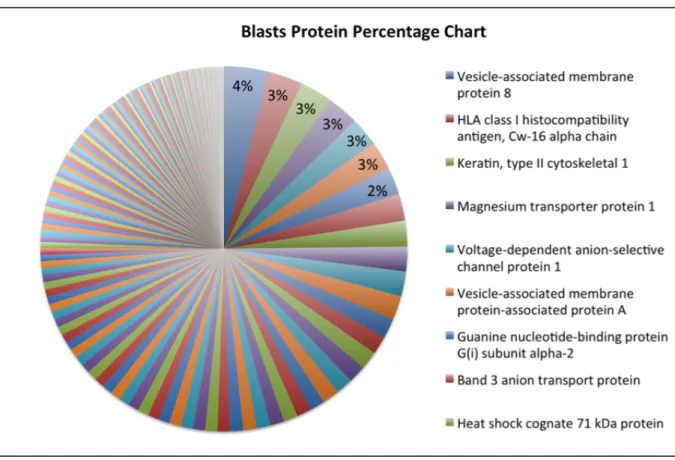
(19) the other didn't have, the amount of the unique proteins would be less than 1% or possibly much lower than 1% of the total membrane protein amount. The result of identified proteins abundance and identified protein number in PBMCs and blasts pointed out the obstacles in our research. If we were to identify unique proteins in blasts by our membrane proteome system, we had to increase protein amount significantly, to develop a much more efficient membrane proteome workflow than the currently used ones, or to improve the sophistication of protein identification by Mass Spectrometry or membrane protein database. All of the above approaches were widely regarded as the obstacle in the field of membrane proteome.. Aberrant protein abundance distribution in refractory leukemic blasts Even though blasts and PBMCs shared most of their protein species, the relative amount, or the percentage of the amounts of different proteins, differed greatly. The most abundant protein in PBMCs accounted for 17% of total membrane protein abundance, while the most abundant protein in blasts accounted for only 4%. In addition, the 10 most abundant proteins in PBMCs made up more than half of total membrane protein abundance, while half of the total blasts membrane protein abundance required the summation of top 25 abundant proteins. Most cells had its membrane protein abundance mainly dominated by several abundant proteins, a pattern similar to that of PBMCs and MV4-11. However, blasts showed an uncommon membrane protein composition without any dominant proteins. The result pointed out that refractory cancer cells, which had survived several rounds of harsh chemotherapy, might have escaped the treatments through drastically changing the composition of membrane proteins. Blasts’ ability to escape chemotherapy and immunosurveillance might have contributed to this pattern of composition, including what the most abundant proteins 18.
(20) were and the membrane protein percentages, than distribution of really low amount unique proteins.. Figure 6. The compositions of membrane proteome in refractory leukemic blasts, normal PBMC and MV4-11. This figure showed identified proteins percentage of blasts, PBMCs, MV4-11 cells, from the most abundant ones to the least abundant ones.. Analysis of normal PBMC revealed the physiological representation Several of the most abundant membrane proteins in PBMCs were important receptors that played roles in cell adhesion, platelet aggregation, connection of cytoskeleton to plasma membrane, and other signaling pathways (Figure 7). Integrin beta-3 was the most abundant membrane proteins in PBMCs, contributing to 17% of total membrane proteins. It had several roles from platelet aggregation, cell adhesion to host-virus interaction. The second most abundant proteins was Talin-1. Talin-1 connected cytoskeletons to plasma membrane and was especially important cell-cell contact of lymphocytes. Many abundant membrane proteins were receptors that binded to a broad range of ligands and had diverse involvement. Platelet glycoprotein 4(CD36) had involvement from inflammatory response, fat processing, to angiogenesis; Integrin alpha-6/beta-4. 19.
(21) assisted hemidesmosome activity; Platelet endothelial cell adhesion molecule was important for leukocyte transendothelial migration (TEM), apoptosis, and phagocyte ingestion prevention.. Figure 7.. A close-up analysis for normal PBMC membrane proteome.. Dysregulated membrane proteome in refractory leukemic cells shows multi-functinality of its components Leukemic blasts had a different membrane protein composition from PBMCs. Blasts didn't have dominant abundant proteins. Instead, ten of the most abundant membrane proteins had similar abundance of about 3% of total, and they made up only 20% of the total proteins (comparing to PBMCs, with the most abundant membrane protein making up 17% of the total). Besides membrane protein composition, blasts also had different abundant proteins with different functions from PBMCs. Vesicle-associated membrane protein 8(VAMP8) was the most abundant membrane proteins in blasts, but it only made up 4% of the total 20.
(22) membrane proteins. VAMP8 was a SNARE that played essentail role in vesicle-trafficking system. It helped early and late endosomes fusion, involved in midbody abscission of cell division, and fusion of autophagosome to lysososome in autophagy. HLA class I histocompatibility antigen involved in foreign antigens presentation in host-virus interaction. Magnesium transporter protein 1 associated woth N-oligosaccharyl transferase and therfore involved in N-glycosylation. Voltage-dependent anion-selective channel protein 1 formed channels on plasma membrane that helped small molecules diffusion and also on mitochondria that were associated to apoptosis. Vesicle-associated membrane protein-associated protein A, like VAMP8, was also a part of vesicle-trafficking system and it assisted plasma membrane recruitment of VAPA.. Figure 8.. A close-up analysis for refractory leukemic blast membrane proteome. 21.
(23) Imbalanced vesicle-trafficking system in refractory leukemic blasts might lead to a mechanism leukemia-mediated dysregulation of surface proteins After validating the processing in our TMT-based quantitative proteomics analysis (Figure 9), we focused on dysregulated surface proteins in refractory leukemic blasts. Surprisingly, as shown in Figure 10, the most upregulated protein in refractory leukemic blasts is VAMP8 (4-fold, p<0.01), a v-SNARE complex molecule mainly responsible for the formation of fused vesicles between membranes.. This molecule is playing a. critical role for protein trasport between endoplasmic reticulum, Golgi apparatus and plasma membrane.. 22.
(24) Figure 9.. The quantification overview of the comparison between refractory leukemic. blats and normal PBMC.. The left diagram represents the quantification of. peptide-spectrum matches in refractory leukemic blasts and normal PBMC. The midpoint of this log2 ratio curve sits at 0 means the these two samples were mixed 1:1. The right diagram points out that VAMP8 is the most upregulated proteins in refractory leukemic blasts.. As revealed in the quantitative proteomics analysis, VAMP8 protein abundance in blasts were over four times the abundance in PBMCs. Note that the abundance of total membrane proteins in blasts were already much lower than that in PBMCs, which displayed the possibly significant role of VAMP8 in blasts. Likewise, VAPA protein abundance in blasts were over two times the abundance in PBMCs. The much higher expression of VAMP8 and VAPA in blasts, comparing to that of PBMCs, pointed out that VAMP8 and VAPA were possibly the key players in determining blasts’ cancerous character and that digging into the vesicle-trafficking system of blasts might be an improtant issue in understanding the drug-risistent character of these refractory leukemia cells.. 23.
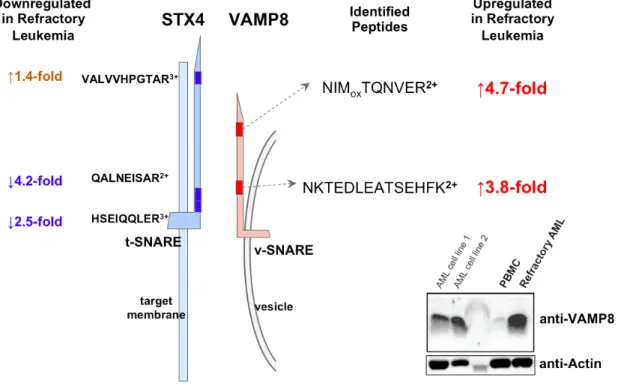
(25) Figure 10.. The comparison of VAMP8 expression between normal PBMC and. refractory leukemic blasts.. The protein abundance of VAMP8 in normal PBMC and. refractory leukemic blasts were shown, respectively.. Figure 11.. Imbalance between SNARE complex components measured by quantitative. proteomics.. 24.
(26) DISCUSSION Discovering a potential mechanism of leukemia-mediated dysregulation of membrane proteins VAMP8 and VAPA might indicate an interesting role vesicle-trafficking system played in refractory blasts and pointed out a new hypothesis of VAMP8 and VAPA involvement in refractory cancer drug resistance. VAMP and VAPA are important players in vesicle-trafficking system. Its relatively high expression in blasts implicates its association with the malignancy and cancerous property of refractory blasts. VAMP, together with syntaxin and SNAP25, comprises the SNARE [soluble NSF (N-ethylmaleimide-sensitive factor) accessory protein receptor] protein. SNARE is the main played in vesicle-trafficking system, which includes synaptic vesicle fusion with plasma membrane, release of neurotransmitters, cytokines, other important molecules, recycling, autophagy, etc. SNAREs are usually categorized into t-SNAREs (target membrane), and v-SNAREs (vesicle membrane). Syntaxin-1 and SNAP25 are usually on t-SNAREs while VAMPs on t-SNAREs. The interaction between t-SNAREs and v-SNAREs brings membrane close, and hence the mergence of lipid bilayers. Besides membrane dysregulation, VAMP8 may also play important roles in cancer progression, invasion, and drug resistance. Bilan, et al. (2008). reported that four endosomal SNARE proteins, syntaxin 8, syntaxin 7, vti1b and VAMP8, interfered with chloride channel activity by interacting and hence regulating CFTR protein trafficking to plasma membrane. Their experiments also showed that CFTR apical targeting were disturbed by endosomal SNARE overexpression. In Bilan, et al. (2008), overexpression of each of the four SNARE components led to CFTR channel number reduction at the cell surface and they suggested it was by cell surface delivery and/or recycling impairment. VAMP8, 25.
(27) according to studies, was expressed abundantly at both apical plasma membrane and submembrane vesicles, which backed the assumption that it cycled from plasma membrane to early endosome and recycling compartments. They hypothesized that overexpression of endosomal SNARE caused disturbance to the dynamic of the apical recycling endosome and therefore the channel number reduction. Bilan, et al. (2008). prompted us to think that the much lower identified membrane protein abundance we observed in blasts (Figure 5), which indicated dysregulation of membrane protein in blasts, was probably linked to the high membrane expression of VAMP8. We also assumed that the membrane protein dysregulation of blasts by expressing less membrane proteins, was possibly a tactic for blasts to avoid immune system and chemotherapy. Besides in membrane dysregulation, VAMP8 as a member of SNAREs may also play important roles in cancer progression, invasion, and drug resistance. In cancer biology, it is known that autophagy promotes malignant cell survival and tumorigenesis. VAMP8 mediates endosomes and autophagosomes’ fusion with lysosomes. Chen, et al. (2015). proved and pointed out directly that VAMP8 as a novel oncogene tightly associated with cancer cell proliferation and therapeutic resistance. They observed overexpression of VAMP8 in human glioma and proved that it promoted cell proliferation in vitro and in vivo. VAMP8 also elevated autophagy proteins expression and autophagosomes number and hence contributed to temozolomide (TMZ) resistance. These studies showed the overexpression of SNARE proteins can contribute towards cancer cell formation and cancer malignance.. 26.
(28) Membrane proteome workflow had limitation in membrane protein identification Continuous improvement in protein purification methods is the key to protein identification and therapeutic target discovery. The physicochemical properties of proteins are diverse, and therefore it is possible that different purification conditions favor respective subsets of proteins. Hence, it is imperative for proteomics analysis to gauge the efficiency of a certain purification method. The recognition of biases rising in the method is equally important: are a subset of proteins always lost during the purification? We should seek to improve our methods in order to extract as many diverse proteins as possible. Meanwhile, to identify proteins of low abundance is the other dimension for proteomics analysts to make progresses. This has always been a major obstacle. Furthermore, membrane proteins make up a small percentage of the total proteome. To get sufficient membrane proteins for experiments usually requires a large amount of cells or clinical samples. We thought lymphocyte membrane proteome might serve as a better experiment control for refractory myeloid leukemic blasts in the same patient, rather than using PBMC from normal donors. This experiment design, however, was hampered by the scarcity of the lymphocytes in refractory AML patients.. Cells displayed diverse functions and morphologies by having a different membrane protein composition As the comparison in this current project, PBMCs and leukemic blasts are different in function and morphology. Although they have many membrane proteins in common, the relative abundance of these proteins still varies. Correspondingly, this discrepancy results in different composition of membrane proteome. How membrane proteome associates with function and morphology of the cell remains the subject of future studies in coming years. 27.
(29) CONCLUSION & PROSPECTIVES We discovered that VAMP8 was overexpressed in refractory acute myeloid leukemia cells. In the future, we hope to further verify by analyzing more refractory leukemia patients’ clinical samples. If the overexpression of VAMP8 is a common phenomenon in myeloid leukemia, VAMP8 might become a novel prognostic factor with the clinical impact. We also hope to verify the assumption that VAMP8 promotes the proliferation of refractory AML cells. Overexpression and knockdown experiment of VAMP8 in vitro and in vivo would help clarify the clinical significance of VAMP8. Last but not least, we would also like to explore the role of VAMP8 in chemoresistance of refractory AML. It was reported in previous studies that VAMP8 elevated autophagy activity and therefore promoted malignant cell survival and tumorigenesis of malignant brain tumors. We could start by observing the expression levels and activities of autophagy proteins and autophagosomes in the leukemic blasts, and knock down autophagy-related proteins to see if chemoresistance can be reversed. This hopefully paves the way for more understanding of the mechanism of chemoresistance, which might, as a result, lead to better survival of patients.. 28.
(30) 參考文獻 1. Bilan, F., Nacfer, M., Fresquet, F., Norez, C., Melin, P., Martin-Berge, A., et al. (2008). Endosomal SNARE proteins regulate CFTR activity and trafficking in epithelial cells. Experimental Cell Research, 314(11-12), 2199-2211. 2. Chen, Y., Meng, D., Wang, H., Sun, R., Wang, D., Wang, S., et al. (2015). VAMP8 facilitates cellular proliferation and temozolomide resistance in human glioma cells. Neuro-Oncology, 17(3): 407–418. 3. Chen, Y. A., & Scheller, R. H. SNARE-mediated membrane fusion. Nature Reviews Molecular Cell Biology 2, 98-106 4. Dimayacyac-Esleta, B. R., Tsai, C. F., Kitata, R. B., Lin, P. Y., Choong, W. K., Lin, T. D., et al. (2015) Rapid High-pH Reverse Phase StageTip for Sensitive Small-Scale Membrane Proteomic Profiling. Analytical Chemistry, 87(24), 12016-23. 5. Mehdi, M., & Menon, M. K. C. (2016). SNARE Protein in Cellular Membrane Trafficking, Its Regulation and as a Potential Target for Cancer Treatment. Open Journal of Immunology, 06(01), 34-48. 6. Ramos, N. R., Mo, C. C., Karp, J. E., & Hourigan, C. S. (2015). Current Approaches in the Treatment of Relapsed and Refractory Acute Myeloid Leukemia. Journal of Clinical Medicine, 4(4), 665-695. 7. Zhao, Q., Fang, F., Liang, Y., Yuan, H., Yang, K., Wu, Q., et al. (2014). 1-Dodecyl-3-methylimidazolium chloride-assisted sample preparation method for efficient integral membrane proteome analysis. Analytical Chemistry, 86(15), 7544-7550. 8. Ziegler, Y. S., Moresco, J. J., Tu, P. G., Yates, J. R., 3rd, & Nardulli, A. M. (2014). Plasma membrane proteomics of human breast cancer cell lines identifies potential targets for breast cancer diagnosis and treatment. PLoS One, 9(7), e102341.. 29.
(31)
數據

+5

相關文件
式中 、 、 為隨物質而定的常數﹐表面張力隨液體性質不同可有很大差別。例 如 20 C 時有機液體苯的表面張力是 28.88
MR CLEAN: A Randomized Trial of Intra-arterial Treatment for Acute Ischemic Stroke. • Multicenter Randomized Clinical trial of Endovascular treatment for Acute ischemic stroke in
此為一項針對轉移性或復發性乳癌患者比較 NK105 與 Paclitaxel 的多國第 III 期臨床詴驗研究。此次修 正主要原因為根據 2016 年 1 月 23
10 SF12259A 黃文豊 一項隨機分配、開放性、多中心臨床 試驗,對於先前未接受過治療的 CD20 表面抗原陽性之瀰漫性大型 B 細胞淋 巴瘤或 CD20 表面抗原陽性之濾泡性
(現場取消未講授);三、 「中東地區的女性與發展—女性領導力(Women and Development in the Middle East—Feminizing Leadership) 」 、 「新網路世紀的創新與發展(Innovation and Growth
第一期至第三期 實體癌,經標準 治療無效;癌症 別:上皮性卵巢 癌、肺癌、肝 癌、乳癌、大腸 直腸癌、胰臟癌. 長聖國際生
提高免疫力上有良好的功效。可治 療糖尿病血管併發症及扁平疣、傳染性疣。在動物 試驗中,利用環磷醯胺製出免
coordinates consisting of the tilt and rotation angles with respect to a given crystallographic orientation A pole figure is measured at a fixed scattering angle (constant d
