探討Adiponectin 調節人類前列腺腫瘤細胞之轉移和α5β1 integrin的表現; Adiponectin mediates cell migration and α5β1 integrin up-regulation in human prostate cancer cells
65
0
0
全文
(2) 誌謝辭 日月如梭,回想兩年前,帶著滿心的期待與抱負來到中國醫藥大學基礎醫學研究 所。開始第一年的研究生涯並不想像中的順遂,兩位指導老師的相繼離職,孤立 無援的我,一度想放棄研究;但不忘初衷深造的理想,依然咬緊牙根繼續往前走。 很幸運地,我來到藥理科湯智昕老師的實驗室,在這裡誠摯的感謝湯老師的協助 與悉心指導,讓我對研究生涯燃起了以往的信心。湯老師是位年輕有為的好老 師,在他身上感受到他對研究及教學的熱誠,讓我對研究這塊有了新的體認。研 究不光是抱著興趣就可以,還要加上耐心與求知慾;也不是像個技術員,有結果 做出來就行,而是面對問題的思考與邏輯性。湯老師也給學生很大的時間和空間 自由運用,彼此的相互尊重,使得實驗室氣氛和樂融融。對這位亦師亦友的湯老 師之感謝,無法以三言兩語來形容;但學生由衷的感謝,讓我在這一年中獲益匪 淺。感謝我的口試委員,曾文培老師和盧大宇老師在口試過程中,對我研究上的 指教評點及建議往後實驗方向。 此外,還感謝實驗室的學長姐不厭其煩指出我實驗中的缺失,還有同學們與學弟 妹們,謝謝你們的幫忙和體諒,有你們的陪伴讓我這兩年的研究生活變得多采多 姿,讓我開心順利過這兩年。也要謝謝所辦的麗如姐協助研究生們的各項事務。 再來感謝父母一路的支持,包容我的任性;在我失意時給我的鼓勵,讓我無後顧 之憂地專心研究而得以順利畢業。 這兩年的研究生涯,雖然一開始非常不順利而備感挫折,但也因為這些壓力激勵 我成長而得到甜美的果實;現在回想起來,過去的種種經歷是個很值得珍藏的回 憶。我會帶著豐碩的果實與各位的祝福,繼續努力向上,充實自己。最後,再次 感謝這一路陪伴、關心、幫助我的人。謝謝你們。. 研究生 盧慕恩 謹識 民國九十八年六月十日 於台中.
(3) 總目錄 (Contents) 頁次 目錄.......................................................................................................................... i 圖表目錄................................................................................................................ iii 縮寫表.................................................................................................................... iv 中文摘要................................................................................................................. v 英文摘要............................................................................................................... vii 第一章 序論........................................................................................................... 1 第一節、文獻回顧......................................................................................... 2 一、前列腺癌................................................................................................ 2 二、癌轉移機制( metastasis ) ...................................................................... 3 三、整合蛋白 integrin 與細胞移行作用 ..................................................... 5 四、Adiponectin 脂聯素.............................................................................. 9 第二節、 研究架構與動機......................................................................... 14 第二章 實驗材料及方法..................................................................................... 15 第一節、材料.............................................................................................. 16 第二節、細胞株 ( Cell line )及細胞培養( Cell culture ).......................... 16 第三節、細胞蛋白質萃取 ( Protein extraction ) ...................................... 17 第四節、蛋白質定量 ( Protein amount assay )......................................... 18 第五節、西方點墨法 ( Western blotting )................................................. 18 第六節、細胞遷移能力分析 ( Migration assay ) ..................................... 20 第七節、轉染作用與報導基因分析 ( Transfection and reporter gene assay ) ...................................................................................................................... 21 第八節、p38 激酶活性偵測 ( p38 MAPK kinase assay ) ........................ 21 第九節、流式細胞儀分析 ( Flow cytometric analysis )........................... 22.
(4) 第十節、Total RNA 萃取 ( Total RNA isolation ) .................................... 23 第十一節、反轉錄聚合酶連鎖反應 ( RT-PCR ) ..................................... 23 第十二節、即時聚合酶連鎖反應 ( Real-time Polymerase Chain Reaction, Real-time PCR )...................................................................... 24 第十三節、統計分析.................................................................................. 24 第三章 實驗結果................................................................................................. 25 第四章 討論......................................................................................................... 30 第五章 結論......................................................................................................... 35 未來展望與建議.......................................................................................... 36 實驗結果表圖....................................................................................................... 37 參考文獻............................................................................................................... 49. ii.
(5) 圖表目錄 Fig.1 The invasion process 癌細胞侵入轉移過程 ................................................ 4 Fig.2 Integrin heterodimer composition ................................................................. 7 Fig.3 The metastatic cascade .................................................................................. 8 Fig.4 Adiponectin 的作用機轉 ........................................................................... 10 Fig.5 Adiponectin 接受器的訊息傳遞 ............................................................... 11 Fig.6 Adiponectin-directed migration of human prostate cancer cells ................. 37 Fig.7 Adiponectin-directed migration of human prostate cancer cells involves upregulation of α5β1 integrins......................................................................... 39 Fig.8 AdipoR receptor is mediated adiponectin-mediated cell migration ............ 40 Fig.9 P38 is involved in adiponectin-mediated migration and integrin up-regulation in human prostate cancer cells .................................................................... 42 Fig.10 AMPK pathway is involved in adiponectin-mediated integrin up-regulation in human prostate cancer cells .................................................................... 44 Fig.11 Adiponectin induces cell migration and integrins up-regulation through NF-κB ....................................................................................................... 46 Fig.12 P38, AMPK and NF-κB inhibitors antagonized the adiponectin-induced κB luciferase activity in human prostate cancer cells..................................... 47 Fig.13 Schematic diagram of the signaling pathways involved in adiponectininduced cell migration in human prostate cancer cell............................... 48. iii.
(6) 縮寫表 縮 寫. 全. 文. AICAR. 5'-aminoimidazole-4-carboxamide-1-β-D-ribofuranoside. AMPK. 5’-AMP-activated protein kinase. BSA. Bovine serum albumin. CAMs. Cell adhesion molecules. cDNA. Complementary DNA. DEPC. Diethylpyrocarbonate. ECM. Extracellular matrix. FBS. Fetal bovine serum. IL-6. Interleukin-6. MEM. Minimum essential medium eagle. NF-κB. Nuclear factor kappa-B. P/S. Penicillin/streptomycin. PBS. Phosphate-buffered saline. PDTC. Pyrrolidine dithiocarbamate. PIN. Prostatic intraepithelial neoplasia. PSA. Prostate specific antigen. PVDF. Polyvinylidene fluoride membrane. qPCR. Quantitative real-time PCR. RT-PCR. Real-time polymerase chain reaction. TNF-α. Tumor necrosis factor-alpha. iv.
(7) 中文摘要 (Abstract in Chinese). v.
(8) 摘要 探討 Adiponectin 調節人類前列腺腫瘤細胞之轉移和 α5β1 integrin 的表現 脂聯素 ( Adiponectin ) 是一種從脂肪組織所分泌的賀爾蒙蛋白,能調整機體 的脂質代謝和血糖穩定;曾經被報導在腫瘤發展和轉移中扮演一個不可或缺的角 色。前列腺癌在歐美的發病率極高,除皮膚癌外;前列腺癌是全世界第二因癌症 導致死亡的疾病,僅次於肺癌。Integrins 在哺乳類細胞上是主要的黏著因子,並 且與乳癌細胞轉移到骨頭有十分密切的相關;Integrins 也與肺癌、乳癌、膀胱和 大腸癌的轉移有著密切的關係。在這項研究中,我們發現脂聯素增加了人類前列 腺腫瘤細胞 (PC3) 的轉移以及腫瘤細胞表面上 α5 和 β1 integrin 的表現。給予 人 類 前 列 腺 腫 瘤 細 胞 α5 或 β1 integrin 的 單 株 抗 體 和 AdipoR 接 受 器 或 5’-AMP-activated protein kinase ( AMPK ) 之短干擾 RNA ( siRNA ) 能夠抑制由 脂聯素增加人類前列腺腫瘤細胞的轉移和 α5β1 integrin 的向上調節。我們給予 人類前列腺腫瘤細胞 p38 抑制劑 ( SB203580 ),AMPK 抑制劑 ( AraA and compound C ),Nuclear factor-kappaB ( NF-κB ) 抑制劑 ( PDTC ) 和 IκB 蛋白酶 抑制劑 ( TPCK ) 都能抑制由脂聯素誘導腫瘤細胞的轉移及 integrins 的表現。 將人類前列腺腫瘤細胞給予脂聯素誘導了 p38、AMPK、IκB kinase α/β ( IKK α/β )、IκB 和 p65 Ser536 磷酸化,以及 κB-luciferase 活性。此外,我們也發現在 其他的人類前列腺腫瘤細胞 ( DU145 與 LNCaP ) 有著相似的訊號路徑。這項研 究是第一次報告,脂聯素導致人類前列腺腫瘤細胞轉移的機轉是藉由透過與 AdipoR 接受器的結合,而依序活化了 p38、AMPK 和 NF-κB 的訊息傳導路徑, 造成了 α5 和 β1 integrins 的轉錄調節。. vi.
(9) 英文摘要 (Abstract in English). vii.
(10) Abstract Adiponectin mediates cell migration and α5β1 integrin up-regulation in human prostate cancer cells Adiponectin is an adipokine has been reported to play an essential role in tumor progression and metastasis. Integrins are the major adhesive molecules in mammalian cells, and has been associated with cancer cells metastasis. In this study, we found that adiponectin increased the migration and the expression of α5 and β1, but not α2, αν and β3 integrins of human prostate cancer cells (PC3). Adiponectin-mediated migration and α5β1 integrin up-regulation was attenuated by AdipoR receptor and 5’-AMP-activated protein kinase (AMPK) siRNA. Pretreatment of cells with p38 inhibitor (SB203580), AMPK inhibitor (AraA and compound C), NF-ĸB inhibitor (PDTC), and IĸB protease inhibitor (TPCK), suppressed the adiponectin-induced cell migration and integrins expression. Stimulation of PC3 cells with adiponectin induced p38 phosphorylation, AMPK phosphorylation, IĸB kinase α/β (IKK α/β) phosphorylation, IĸB phosphorylation, p65 Ser536 phosphorylation, and ĸB-luciferase activity. In addition, we also found that the similar signaling pathways involved in the other prostate cancer cells (DU145 and LNCaP). Finally, this study showed for the first time that the adiponectin mediates migration of human prostate cancer cells. One of the mechanisms underlying adiponectin directed migration was transcriptional up-regulation of α5β1 integrin and activation of AdipoR receptor, p38, AMPK, and NF-ĸB pathways.. viii.
(11) 第一章、序論 (Introduction). 1.
(12) 第一節 文獻回顧 一、前列腺癌 前列腺癌的形成原因與症狀 前列腺癌(又名攝護腺癌)是出自前列腺的惡性腫瘤,前列腺是屬於男性生 殖系统的一種腺體,位在膀胱的下方,直腸的前方,製造濃稠的液體來構成精液 的一部份;幾乎所有前列腺癌都是發生在腺體細胞,這種癌症又稱為腺癌。根據 美國防癌協會 ( American Cancer Society ) 資料顯示,前列腺癌是美國男性中第 二大致命癌症 ( Cancer Reference Information ) ( Delongchamps NB et al.,2006 )。 根據2007年行政院衛生署十大癌症死亡率統計數據指出,台灣前列腺癌死亡 率躍升至男性癌症死亡十大死因的第七位。在台灣前列腺的發生率雖然不像美國 這麼高,但隨著年齡的老化,飲食習慣的西化,其罹患率已有增加的趨勢。前列 腺癌的真正致病原目前尚未完全了解,有報告推論前列腺癌可能和基因體質遺 傳、種族和荷爾蒙失調,而環境因素與飲食佔有很重要的角色 ( Hsing AW et al., 2006 )。 早期的前列腺癌通常沒有症狀,因此難以從症狀來診斷或定義此疾病。最常 見到症狀為類似前列腺增生的症狀,在這期間,會感到頻尿、夜尿、解尿困難和 尿滯留或解尿疼痛等問題。大部分到了晚期才會出現更嚴重的情況,例如血尿、 尿失禁等的情形,而癌細胞已擴散到鄰近的淋巴腺,骨骼或其他器官,引起骨頭 疼痛、病理性骨折或是脊椎受到壓迫而引起神經方面的症狀。. 前列腺癌的分期 前列腺癌分期主要依據腫瘤侵犯程度來區分。多數醫生使用美國癌症聯合委 員會 American Joint Committee on Cancer ( AJCC ) 系統。phaseⅠ:潛伏前列腺 癌,未擴散至淋巴結或遠處器官。直腸指檢無法偵測。phaseⅡ: 攝護腺處摸到 腫塊且仍局限在攝護腺。 phaseⅢ: 癌細胞已侵犯到鄰近的組織器官,但尚未擴. 2.
(13) 散至淋巴結或身體其他部位。phase Ⅳ: 癌細胞已轉移到淋巴腺、骨、肺或其他 遠處器官。 二、癌轉移機制 ( metastasis ) 轉移是指癌细胞在擴散到整個機體的過程。當癌細胞失去正常增生控制時, 細胞移動就會失去控制,使得癌細胞能從原始發生的部位經過血管或淋巴移動到 其他部位擴散;惡性腫瘤不但可以經由直接蔓延的方式破壞鄰近的正常組織或器 官,進一步地經由擴散轉移的方式對身體其他地方或組織繼續生長,形成新的腫 瘤,即繼發性腫瘤。癌轉移為一多重步驟的過程 ( multiple-step process ),包含: 1、原生癌細胞生長並且移動; 2、黏附於血管周圍的組織; 3、侵襲周圍組織並穿 透血管和淋巴管; 4、於血管和淋巴管內形成小腫瘤並隨血液、淋巴液而移動; 5、 黏附在遠處組織器官的血管和淋巴管壁; 6、穿越黏附處的血管和淋巴管並侵入周 圍的組織; 7、癌細胞增生、生長、轉移腫瘤的形成 (Fig.1)。 癌轉移機制中最重要的一環,為癌細胞與週邊細胞環境間活化的作用,這過 程包含: 1、癌細胞通過細胞質膜表面的蛋白與細胞外基質 ( extracellular matrix,ECM ) 交互作用; 2、癌細胞產生基質分解酵素分解基底膜 ( basement membrane ) 並通過; 3、癌細胞從分解的基質中移動 ( Boudreau N et al., 1998; Fornaro M et al.,.2001),經過這些過程後構成了癌轉移系統。 骨骼是所有癌症發生轉移相當常見的部位之一,僅次於肺臟及肝臟而位居第 三位。較常發生骨骼轉移的癌症包括乳癌、攝護腺癌及肺癌等。台灣的前列腺癌 其侵犯轉移的情形發現以骨骼轉移占最多,其次為淋巴結侵犯,另有轉移到肺 部、肝臟。骨轉移好發之部位為脊柱 (腰椎及胸椎)、骨盆,肋骨及胸骨 ( Mundy et al., 2002 )。對前列腺癌病人而言,淋巴結轉移是預後不好的指標。. 3.
(14) Fig.1 The invasion process. 癌細胞侵入轉移過程 The invasion process. Tumour invasion can be described by seven consecutive steps. By loosing homotypic cell–cell adhesion, malignant proliferating cells detach from the primary tumour mass and migrate through surrounding tissues towards blood- or lymphatic vessels (step 1). Rearrangements in the actin cytoskeleton as well as matrix degradation by proteolytic enzymes secreted by the tumour cells, will accompany this migratory step. The cells subsequently attach to and breach the basement membrane (step 2 and 3), gain access to the vessel and spread (step 4). A reverse process, again involving cytoskeletal rearrangements, occurs upon extravasation. Tumour cells adhere to the capillary endotheliae (step 5), penetrate them (step 6) and migrate through the extravascular tissue where they create a suitable microenvironment and newly attract blood supply, finally resulting in a secundary lesion or metastasis (step 7) ( Lambrechts et al., 2004 ).. 4.
(15) 三、整合蛋白 integrin 與前列腺腫瘤轉移的關係 整合蛋白 ( Integrins ) 細胞與細胞之間的黏附分子 ( cell adhesion molecules ; CAMs ) 與癌細胞轉移 有關 ( Okegawa T et al., 2004 )。Integrin 屬於其中一種,是細胞與細胞外基質之 間的黏附分子。當癌細胞產生局部侵犯或轉移的過程中,它所接觸到的細胞外基 質是相當多變的,因此 integrin 的次單位群必須不斷地產生變化並重新組合, 以利該細胞與外界環境之黏附,有利於細胞產生移動。細胞移行是細胞生物體發 展與維持的一個中間過程;若是這過程期間發生了錯誤會造成血管疾病、腫瘤的 形成和轉移。細胞表面的 integrins 是細胞移行的一個重要角色 ( Huttenlocher et al., 1996 )。 Integrins 由 非 共 價 地 相 互 作 用 方 式 去 形 成 24 種 不 同 的 異 質 雙 體 ( heterodimeric ),是一群位於細胞表面具有穿透黏著性的接受器,包含了 19 個 α 和 8 個 β 次單位 ( Fig.2 )。Integrins 主要連結細胞外基質與細胞內的細胞骨架 的結構和訊息傳遞分子,例如纖維連接蛋白、層連接蛋白、玻璃連接蛋白、膠原 蛋白等,調節著細胞存活、基因表現、增生、轉移、生長與分化,和組織重新塑 造 (Giancotti FG et al.,1999)。 Integrins 與許多細胞組成的過程有關,包含了血管增生、細胞凋亡、細胞的 轉移和腫瘤的生長。另一方面,顯示 integrins 於肺癌、胰臟癌和皮膚黑色素瘤 的表現有關,有著較差的預後 ( Böttger TC et al., 1999;Müller-Klingspor V et al., 2001;Nikkola J et al., 2004;Oshita et al., 2004;Yao ES et al., 2007 )。Integrins 也與肺癌、乳癌、膀胱和大腸癌的轉移有著密切的關係 ( Miyazaki T et al., 2005; Bub JD et al., 2006;Dieudonne MN et al., 2006 )。過去十年研究顯示,integrins 藉 由活化不同細胞內的訊息路徑影響了腫瘤的生長過程 ( Guo W et al., 2004; Giancotti FG et al., 2004 )。此外,ανβ3 integrin 與前列腺腫瘤生長、血管增生、 存活和侵襲有關。 前列腺癌的發展經由一連串步驟:前列腺表皮層新生 ( PIN ),進一步表皮層 5.
(16) 損傷,腫瘤侵襲,非雄性激素依賴階段 ( Scher HI et al., 2000;Culig Z et al., 2006;Dehm SM et al., 2006 )。這些時期的定義是由正常細胞多樣變化而來的 (Mimeault M et al., 2006 )。在前列腺癌的改變描述中,integrin 的功能表現異常 和它們的細胞外基質 ( ECM ) 有關 ( Boudreau N et al., 1998;Fornaro M et al., 2001 ) 。這些結果指出異常細胞間的交互作用與細胞外基質促進細胞增生、移行 和分化,經由上述的狀態造成癌症發展 ( Boudreau N et al., 1998;Fornaro M et al., 2001;Knudsen BS et al., 2006 )。在細胞實驗中,發現 integrins 透過許多細胞外 基質 ( ECM substrates ) 促進前列腺腫瘤細胞黏著和移行並且穿過內膜轉移 ( Cooper CR et al., 2002;Kumar CC et al., 2003;Wang X et al., 2005 )。Integrins 於 調控的癌症表現過程中扮演一個重要角色 ( Fig.3 )。. 6.
(17) Fig.2 Integrin heterodimer composition. Integrin α- and β-subunits form 24 heterodimers that recognize distinct but overlapping ligands. Half of the α-subunits contain I domains (asterisked). ( Shimaoka M et al., 2003). 7.
(18) Fig.3 The metastatic cascade. Changes in adhesion are prominent during the metastatic journey. At the onset of this process, cancer cells lose E-cadherin-dependent intercellular adhesions, acquire a migratory phenotype, penetrate the basement membrane, and invade the interstitial matrix. Tumour angiogenesis then allows cancer cells to enter the bloodstream, either directly or through the lymphatic system, by a process called intravasation. In the circulation, tumour cells form small aggregates with platelets and leukocytes. Finally, after stopping in the microcirculation of the target organ, tumour cells exit the bloodstream, by a process called extravasation, and undergo local expansion. (Guo W et al., 2004 ). 8.
(19) 四、Adiponectin (脂聯素) Adiponectin 亦稱 adipocyte complement-related protein ( Acrp30 ),AdipoQ 和 gelatin-binding protein-28 ( GBP28 ),由APM 1 ( AdiPose Most abundant gene transcript 1) 轉錄而來 ( Vasseur F et al., 2003 ),是脂肪細胞所分泌的一種內生性 生物活性多肽或蛋白質。人體內的 adiponectin 由 244 個氨基酸組成,分子量 為 30KDa ( Maeda K et al., 1996;Nakano Y et al., 1996 )。Adiponectin 作為一種 胰島素增敏激素 ( Insulin-sensitizing Hormone ) ,可以促進骨骼肌細胞的脂肪酸 氧化和葡萄糖再回收,於肝臟細胞明顯減少糖質新生作用,抑制肝糖分解,在體 內是血糖穩定和脂質代謝重要的調節因子 ( Scherer PE et al., 1995;Hu E et al., 1996 ) (Fig.3)。Adiponectin 可能和肥胖與癌症有著密切的關係。曾經有文獻提出 血清中 adiponectin 的減少,會增加肥胖相關癌症的風險,例如:大腸癌、乳癌、 內皮細胞,子宮內膜癌和前列腺癌 ( Miyoshi Y et al., 2003;Goktas S et al., 2005; Freedland SJ et al., 2005 )。有些研究對於經由 adipose tissue 所分泌的 adipokines 分子訊息和在前列腺腫瘤發展的關係感到相當興趣。許多 adipokines,尤其是 leptin、adiponectin、TNFα 和 IL-6 影響了腫瘤生長的過程,包含了前列腺癌。 Adiponectin 可分為兩種形式: globular adiponectin ( gAd ) 與 full-length adiponectin ( fAd ),gAd 與 AdipoR1 有很高的親合力;fAd 則與 AdipoR2 具有 較高的親合力 ( Yamauchi T et al., 2003 )。目前已知 adiponectin 接受器有兩種形 式: AdipoR1 大量表現於骨骼肌,另一個為 AdipoR2 表現於肝臟 ( Yamauchi T et al., 2003 )。最近研究指出,adiponectin 接受器表現在人類乳癌 ( Takahata C et al., 2007 ) 和前列腺癌 ( Michalakis K et al.,2007 ) 上。藉由增加或抑制 AdipoR1/R2 的表現顯示出,globular 和 full-length adiponectin 與 AdipoR1 和 AdipoR2 接受 器結合後,進一步地調控 p38、AMPK、PPARα ligand 活性、脂肪酸氧化和葡 萄糖回收 (Fig.4) ( Yamauchi T et al., 2003 ) 。然而 adiponectin 在前列腺腫瘤細 胞的移行能力大部分還尚未明瞭。. 9.
(20) Fig.4 Adiponectin 的作用機轉 Adiponectin can activate AMPK and PPARα in the liver and skeletal muscle. In skeletal muscle, both globular and full-length adiponectin activate AMPK, thereby stimulating phosphorylation of ACC, fatty-acid oxidation, and glucose uptake. Adiponectin activates PPARα, thereby also stimulating fatty-acid oxidation and decreasing tissue TG content in muscle. In the liver, only full-length adiponectin activates AMPK, thereby reducing molecules involved in gluconeogenesis and increasing phosphorylation of ACC and fatty-acid oxidation. Adiponectin activates PPARα, thereby stimulating fatty-acid oxidation and decreasing tissue TG content in the liver. These alterations all increase insulin sensitivity in vivo. ( Yamauchi T et al., 2003 ). 10.
(21) Fig.5 Adiponectin 接受器的訊息傳遞. 11.
(22) Fig.5 Signal transduction by adiponectin receptors. Globular adiponectin exists as a trimer, whereas full-length adiponectin exists as at least 3 species of multimers: an LMW trimer, an MMW hexamer, and an HMW multimer. Suppression of AdipoR1 by RNA interference markedly reduces globular adiponectin binding, whereas suppression of AdipoR2 by RNA interference largely reduces full-length adiponectin–specific binding. The dotted line between AdipoR2 and globular adiponectin reflects that AdipoR2 is a relatively low-affinity receptor for globular adiponectin. AdipoR1 and AdipoR2 do not seem to be coupled with G proteins, since overexpression of AdipoR1/R2 has little effect on cAMP, cGMP, and intracellular calcium levels, but instead these receptors activate unique sets of signaling molecules such as PPARα, AMPK, and p38 MAPK. In C2C12 myocytes overexpressing AdipoR1/R2, adiponectin stimulates PPARα, AMPK, and p38 MAPK activation, glucose uptake, and fatty-acid oxidation .Suppression of AMPK or PPARα partially reduces adiponectin-stimulated fatty-acid oxidation, and suppression of AMPK or p38 MAPK partially reduces adiponectin-stimulated glucose uptake. In hepatocytes overexpressing AdipoR1/R2, adiponectin stimulates PPARα or AMPK and fattyacid oxidation. Suppression of AMPK or PPARα in these hepatocytes partially reduces adiponectin-stimulated fatty-acid oxidation. Moreover, treatment with adiponectin reduces plasma glucose levels and molecules involved in gluconeogenesis in the liver, and dominant-negative AMPK partly reduces these effects. These data support the conclusion that AdipoR1 and AdipoR2 serve as receptors for globular and full-length adiponectin and mediate increased AMPK, PPARα ligand activities, p38 MAPK, and adiponectin-induced biological functions. T-cadherin is capable of binding adiponectin but is thought to have no effect on adiponectin cellular signaling, since T-cadherin lacks an intracellular domain. Interaction of APPL1 (adaptor protein containing pleckstrin homology domain, 12.
(23) phosphotyrosine-binding domain, and leucine zipper motif 1) with AdipoR1 appears to play important roles in adiponectin signaling and adiponectin-mediated downstream events such as lipid oxidation and glucose uptake. S-S, disulfide bond. ( Kadowaki T et al., 2006 ). 13.
(24) 第二節、研究架構與動機 前列腺癌又名攝護腺癌,幾乎所有前列腺癌都是發生在腺體細胞。前列腺癌 細胞對於雄性激素相當敏感,而病患治療首選為荷爾蒙療法也就是阻斷雄性激素 -睪丸酮對前列腺癌細胞的作用,進而達到抑制癌細胞的增生及轉移的效果。但 仍有出現荷爾蒙療法失效的情況,稱為非雄性激素依賴型之前列腺癌 Androgen-independent prostate cancer ( AIPC )。 PC3 和 DU145是前列腺癌典型的人類前列腺腫瘤細胞株。文獻指出PC3 細 胞比中度轉移能力DU145細胞具有高度的轉移能力 ( Alimirah F et al., 2006 )。 PC3細胞株取得於具非雄性激素依賴型高度骨轉移的前列腺癌。PC3細胞有低 testosterone-5-alpha 並且表現 PSA (攝護腺特異性抗原)( Pulukuri SM et al., 2005 )。而LNCaP細胞源自於淋巴結轉移之雄性激素敏感的人類前列腺腫瘤細胞。 肥胖症與脂肪細胞所誘導的細胞激素與前列腺腫瘤生長有關。然而 adipokine 基因的變異性與前列腺癌的風險目前尚未十分徹底地研究;最近,令 人感到相當興趣的部分是基因與 adipokines 由脂肪組織所分泌的細胞與細胞之 間的訊息分子和它們在前列腺腫瘤發展的角色;而許多 adipokines 特別是 leptin 、 adiponectin 、 TNF-α 和 IL-6 影 響 了 腫 瘤 的 發 展 過 程 包 含 了 前 列 腺 ( Moore SC et al., 2009 )。 我們進一步地假設由 adipocyte 所分泌之 adiponectin 可能具有調節人類前 列 腺 癌 細 胞 的 移 動 能 力 及 integrin 參 與 轉 錄 調 控 的 表 現 。 也 進 一 步 探 討 adiponectin 對於不同之人類前列腺腫瘤細胞的移行能力。在本篇研究中,我們 發現 adiponectin 會增加人類前列腺癌細胞的移動能力及 α5β1 integrins 的表 現。此外,AdipoR 接受器、p38、AMPK、及 NF-κB 的訊號傳遞路徑參與在由 adiponectin 調控增加前列腺癌細胞的轉移過程當中。. 14.
(25) 第二章、材料與方法 (Materials and Methods). 15.
(26) 第一節、材料 化學藥品有機溶劑、實驗器材及設備大部分購自於以下廠商: Anti-mouse and anti-rabbit IgG-conjugated horseradish peroxidase、 rabbit polyclonal antibodies specific 如 β-actin、 p-p38、 p38、 IKK、 p-IκBα、 IκBα 和 p65 購 自於Santa Cruz Biotechnology ( Santa Cruz, CA, USA )。Rabbit polyclonal antibody specific 如 AMPKα phosphorylated Thr172 、IKKα/β phosphorylated Ser180/181 、p65 phosphorylated Ser536 與. AMPKα 購 自 於. Cell Signaling 與. Neuroscience. ( Danvers, MA )。PDTC、TPCK、SB203580、5-Aminoimidazole-4-carboxamide ribonucleoside (AICAR) 、 Compound C 和 adenosine-9-β-D-arabino-furanoside ( AraA ) 購自於 Calbiochem ( San Diego, CA )。Mouse monoclonal antibody specific 如 α2、 α5、 β1、β3、α2β1 以及 α5β1 integrin 購自於 Chemicon ( Temecula, CA )。Human full-length adiponectin ( Cat. No. 1065-AP ) 購自於 R&D Systems ( Minneapolis, MN ) NF-κB luciferase plasmid 購自於 Stratagene ( La Jolla, CA )。IKKα ( KM ) 和 IKKβ ( KM ) mutants 由 Dr. H. Nakano ( Juntendo University, Tokyo, Japan ) 提供。p38 dominant negative mutant 由 Dr. J. Han ( South-western Medical Center, Dallas, TX ) 提供。Luciferase assay kit 購自 於 Promega ( Madison, MA ) 。 其 他 化 學 藥 品 有 機 溶 劑 大 部 分 購 自 於 Sigma-Aldrich ( St. Louis, MO )。. 第二節、細胞株 ( Cell line )及細胞培養 ( Cell culture ) 一、細胞株 本實驗室所採用的人類正常前列腺細胞株 ( PZ-HPV-7 ) 與人類前列腺癌細 胞株 ( PC3 、DU145、LNCaP clone FGC ),購自食品工業發展研究所之生物資 源保存及研究中心 ( BCRC ),菌種中心編號 BCRC 60136 ( PZ-HPV-7 )、BCRC 60122 ( PC-3 ) 、BCRC 60348 ( DU 145 )、BCRC 60088 ( LNCaP clone FGC )。 人類正常前列腺細胞株 PZ-HPV-7 與人類前列腺癌細胞株 PC3 、DU145 16.
(27) 均屬於非雄性激素敏感之貼附型細胞,LNCaP clone FGC 則屬於雄性激素敏感之 半懸浮型細胞。人類正常前列腺細胞株 PZ-HPV-7 培養於含有 5 ng/ml human recombinant epidermal growth factor 和 50 μg/ml bovine pituitary extract 的 Keratinocyte-serum free medium。人類前列腺癌細胞株 PC3 、LNCaP clone FGC 培養於含有 10% 胎牛血清 ( FBS, Invitrogen Corpoation, California, USA ) 和 1 %. P/S ( penicillin/streptomycin ) ( Invitrogen Corpoation,California, USA ) 的. RPMI-1640 培養液。人類前列腺癌細胞株 DU145 培養於含有 10% 胎牛血清 ( FBS, Invitrogen Corpoation, California, USA ) 和 1 % P/S ( Invitrogen Corpoation, California, USA ) 之 MEM 培養液。 二、細胞培養 將細胞培養於 10 cm 培養皿 ( Corning ) 中,待細胞長至八分滿,吸掉培養 液並以 PBS ( phosphate-buffered saline;含有 137 mM NaCl 、2.7 mM KCl、10 mM NaHPO4 及 2 mM KH2PO4 ) 清洗,藉由 Trypsin-EDTA (含有 0.5 % Trypsin、EDTA-4Na 及 NaCl ) ( Invitrogen Corpoation, California, USA ) 作用使細 胞懸浮,吸掉 TE,加入 5 ml 培養液中和 TE,並將細胞打散,並取出適當量 之細胞液加入已含新鮮生長培養基之培養皿中,置於 37 0C 、5 % 二氧化碳 ( CO2 ) 培養箱中培養。. 第三節、細胞蛋白質萃取 ( Protein extraction ) 將人類前列腺癌細胞株 PC3 分盤培養於 6 孔盤內,依不同時間點給予細 胞 PC3 之 adiponectin 後,吸掉培養液,以冰 PBS 緩衝液輕輕沖洗,再吸乾殘 餘的 PBS 緩衝液。加入 150~200μl RIPA 緩衝液 ( 含1M Tris-Cl、0.5 M NaCl、 0.5M EDTA、10 % SDS、10 % DOS、10 % NP40 及 1 % proteinase inhibitors ), 置於冰上作用五分鐘,再以細胞刮勺 ( scraper ) 將細胞刮下,並把細胞懸浮液收 集至微量離心管,於 4 0C 下作用 30 分鐘 幫助裂解細胞。以 4 0C 13,200 rpm 離心 15 分鐘,吸取上清液至另一微量離心管,各取出 10 μl 做為定量用,其餘 17.
(28) 儲存於 -20 0C 備用。. 第四節、蛋白質定量 ( Protein amount assay ) 蛋白質濃度以 BCA ( bicinchoninic acid ) 方法來定量,利用蛋白質對兩價銅 離子 ( Cu2+ ) 進行還原,所生成的 Cu+ 可以 BCA 專一性地呈色而定量。分析 程序:1、製備一組系列 BSA ( 2 mg/ml ) 標準品 ( Pierce Biotechnology, Rockford, USA ),以去離子水稀釋一系列濃度:2、1、0.5、0.25、0.125、0.0625 mg/ml, 並作出標準曲線。 2、將已稀釋完成之標準溶液及待測樣品溶液,各取 10 μl 加 入 96 孔試盤。 3、每槽加 200 µl 之 BCA reagent [含 Reagent A (BCA) 及 Reagent B (CuSO4) ] 以 50:1 之比例均勻混合液後,於 37 0C 靜置 30 分鐘。4、 以 ELISA reader 測 562 nm (或 570 nm ) 吸光值,劃出標準曲線,以內插法計算 出樣品濃度 ( Y= A + BX ; Y:OD570 nm 讀值;X:待測樣品濃度)。. 第五節、西方點墨法 ( Western blotting ) 將定量後的蛋白質經 SDS-PAGE ( 90~120 voltage 的電壓進行電泳 1 小時) 分離後,切除集膠凝膠部分,將解析凝膠進浸入轉漬緩衝液 ( transfer buffer, 25 mM Tris , 192 mM Glycine )。取轉漬紙 PVDF 薄膜 ( Millipore ),裁切成膠體大 小, PVDF 需先以少量的甲醇 ( methanol ) 潤濕液一分鐘活化,再以去離子水 清洗數次,將 PVDF 浸入轉漬緩衝液。將膠體取起與轉印紙 PVDF 齊放,接著 於轉印槽中注滿 transfer buffer 後將電流以 4 0C 400 毫安培轉印 2 小時。轉漬 完 成 後 , 將 PVDF 浸 入 含 5 % 脫 脂 奶 粉 之 TBS-T ( TBS 含 有 0.05 % tween-20 ),置於室溫搖晃 1 小時,進行阻斷反應步驟 ( blocking step ) 以填塞 PVDF 上沒有蛋白質轉漬的空間。PVDF 以 TBS-T 清洗四次,每次 15 分鐘, 去除多餘的填塞物質。加入適量之一級抗體於 4 0C 搖晃反應隔夜或室溫一小 時,辨識目標蛋白質。PVDF 以 TBS-T 清洗 4 次,每次 15 分鐘,去除多於的 抗體。再加入二級抗體於室溫中搖晃 1 小時,放大目標蛋白質的訊號。 PVDF 18.
(29) 再以 TBS-T 洗 4 次,每次 15 分鐘。 取出 PVDF 稍微瀝乾,加上顯影劑 ( western blotting luminnol reagent,取 buffer A 及 buffer B 以 1:1 的比例混合) 均勻,再均勻加於轉印紙上,將轉印 紙放入底片夾,以感光底片感光至 Band 出現。. Western blotting所需配置之Buffer 組成如下: SDS-PAGE: 10% Resolving gel (5ml). 5% Stacking gel (3ml). 1.9ml H2O. 1.75ml H2O. 1.7ml 30% acrylamide mix. 0.65ml 30% acrylamide mix. 1.3ml 1.0 M Tris (pH6.8). 0.5ml 1.0 M Tris (pH6.8). 0.05ml 10% SDS. 0.01ml 10% SDS. 0.05ml 10% ammonium persulfate. 0.01ml 10% ammonium persulfate. 0.002ml TEMED. 0.002ml TEMED. Buffer 製備: sample buffer (5x): 7.81. ml. stacking. Lysis buffer: buffer. (0.5mM 50mM Tris pH7.5. Tris-HCl ,PH=6.8) 2.5 g SDS. 150mM NaCl. 14.36 ml glycerol. 10mM EDTA. 6.25 ml. 1% NP-40. 2-mecaptoethanol. 2 mg bromophenol blue. 0.25% deoxycholate. 加二次水至 50ml RIPA buffer:. TBS-T (20X): 19.
(30) 1ml lysis buffer. 48.46g Tris. 50mM PMSF. 0.5M EDTA. 50mM Na3Vo4. 58.44g Nacl (PH=7.5). 10μg/ml Aprotinin. 加去離子水至 1L。. 1mM NaF. Running buffer(10x):. Transfer buffer(10x):. 30.2g Tris-base. 30g Tris. 140g Glycine. 144g Glycine. 10g SDS. 加去離子水至 1L。. 加去離子水至 1L。. 第六節、細胞移行能力分析 ( Migration assay ) 利用雙層通透性的培養盤 transwell ( Costar, Corning Life Sciences, Acton, MA;pore size, 8-µm )。以細胞穿過多碳膜能力的不同,來分析比較藥物對細胞 遷移能力的影響。 在進行細胞移行實驗前,細胞先用不同濃度的抑制劑前處理 30 分鐘,包括 Compound C、 AraA、 SB203580、 PDTC 或 TPCK。於 lower chamber 中置 放 300µl 含 1% FBS 及 adiponectin 的培養液,再放入 upper chamber。將約 2 ×104 個細胞與 200 µl 無添加血清的 RPMI-1640 培養液注入 upper chamber , 置於 37 0C、5 % 二氧化碳 ( CO2 ) 培養箱中培養 16 個小時後,用 10 % formaldehyde 浸泡 10 分鐘,使細胞固定於 mesh 上,再以 PBS 清洗,並用 0.05 % crystal violet 染色 15 分鐘,並用 PBS 緩衝液清洗。在 upper chamber 的細胞 用棉花棒輕輕擦拭多餘的染劑及雜質。移行至 lower chamber 的細胞可以用顯微 鏡下計數數量。每次實驗皆為三重複,每個實驗在不同時間點再做一次。每個實 驗中,移行的細胞數量藉由細胞存活實驗 ( cell viability assay ) 結果去調整,以 20.
(31) 校正由 adiponectin 所引起的細胞增生效應。(正確的細胞移行數量 = 計數細胞 移行的數量 / 細胞存活百分比) ( Chu CY et al., 2007;Huang YC et al., 2007 )。. 第七節、轉染作用與報導基因分析 ( Transfection and reporter gene assay ) 利用 Lipofectamine 2000 ( LF2000 ; Invitrogen ) 轉染法,將 DNA 質體轉染 (transfection )至人類前列腺癌細胞株 PC3 中。欲轉染的 PC3 先以每孔 2×104 的細胞數種在 12 孔培養盤中, 370C 培養隔夜等待細胞貼附後,進行轉染。 將 buffer A:含有 1μg ĸB-luciferase plasmid 及 1μg DNA plasmid 的無血清 培養液混合 5 分鐘,再將其 buffer B:含有 2μg. LF2000 的無血清培養液混合,. buffer A+B 混合均勻後室溫靜置 30 分鐘。取出事先培養的 12 孔培養盤細胞, 吸掉培養液,加入 500 μl 無血清培養液 ,將混合均勻的 buffer A + B 一滴一 滴緩慢滴入培養盤中,輕輕搖晃均勻。將細胞放入 37 0C 培養箱中培養 3~4 小 時使轉染作用反應,將其上清液吸掉,加入 500 μl 含有血清的培養液使細胞回 復 24~36 小時後,吸掉培養液,加入 500 μl 無血清培養液,再將細胞與含有 Adiponectin 的培養液一同培養 24 小時後,移除上清液,並用冰的 PBS 緩衝溶 液清洗。每個孔盤加入 100 μl 的 reporter lysis buffer ( Promega ) 作用 10~15 分 鐘,再以細胞刮勺 ( scraper ) 將細胞刮下,並把細胞懸浮液收集至微量離心管, 以 4 0C 13,200 rpm 離心 15 分鐘,吸取上清液至另一微量離心管,取出 20 μl 的 細胞上清液置放於不透光的 96 孔盤中,每孔加入 80 μl 的 luciferase substrate 迅速均勻混合,以冷光儀測定測其 luciferase 活性,將得到之冷光產量讀值,以 百分率換算表示。. 第八節、p38 激酶活性偵測 ( p38 MAP kinase assay ) 利用p38 MAP kinase 基質 [activating transcription factor (ATF)-2] 來檢測 adiponctin 是否經由 p38 的磷酸化 ( Raingeaud J et al., 1995 )。將人類前列腺腫 瘤細胞 PC3 分盤培養於 6 孔盤內,吸掉培養液,以冰 PBS 緩衝液輕輕沖洗, 21.
(32) 再 吸 乾 殘 餘 的 PBS 緩 衝 液 。 加 入 含 有 1mM phenylmethylsulfonyl fluoride ( PMSF ) 之 1× cell lysis buffer ,置於冰上作用五分鐘,再以細胞刮勺 ( scraper ) 將細胞刮下,在冰上 sonicate 震盪細胞 5 秒/次,共四次。以 4 0C 13,200 rpm 離 心 10分鐘,吸取上清液至另一微量離心管,加入 20 μl Immobilized Phospho-p38 MAPK ( Thr180/Tyr182 ) Monoclonal Antibody Beads ,於 4 0C 搖晃隔夜。以 4 0. C 13,200 rpm 離心 30 秒,以 1× lysis buffer 清洗兩次,再用 1× kinase buffer. 清洗兩次後,加入 50 μl. 1× kinase buffer ,加入 1μl. ATF-2 fusion protein 與. 1μl ATP ,以上過程皆在冰上操作,之後於乾浴鍋上培養 30 0C 30分鐘,最後加 入 25 μl 3× SDS sample buffer,震盪均勻後,13,200 rpm 離心 30 秒,接著進行 Western Immunoblotting 實驗。. 第九節、流式細胞儀分析 ( Flow cytometric analysis ) 將人類前列腺癌細胞株 PC3 先分盤培養於 6 孔盤內,待細胞長約九分滿 後,吸掉培養液。以 PBS 緩衝液清洗,吸乾殘餘的 PBS 緩衝液,再以 37 0C 的 Trypsin 將細胞自培養盤中分離。接著使用含 75% 酒精溶液固定細胞至隔夜, 固定完畢後用 PBS 緩衝液清洗,並將細胞培養在 α2、α5、αν、β1、β3 integrin 的 mouse anti-human antibody 或 rabbit anti-human antibody ( 1:100 ) 之一級抗 體,室溫靜置培養 1 小時後。7,500 rpm 離心 5 分鐘,吸掉一級抗體,用 PBS 緩衝液清洗, 7,500 rpm 離心 5 分鐘,吸掉上清液,並將細胞與接有 fluorescein isothiocyanate-conjugated 的 goat anti-mouse secondary IgG 或 goat anti-rabbit secondary IgG ( 1:100; Leinco Tec., St Louis, MO ) 室溫下避光培養 45 分鐘後, 7,500 rpm 離心 5 分鐘,吸掉二級抗體,加入 500 μl PBS 緩衝液,然後進行流 式細胞技術進行實驗,使用 FACSCalibur and CellQuest software ( BD Biosciences, San Jose, CA ) 分析 ( Tang CH et al., 2006 )。. 22.
(33) 第十節、Total RNA 萃取 ( Total RNA isolation ) 將細胞培養於 6 孔培養皿中,待細胞長約九分滿後,吸掉培養液,以冰的 PBS 緩衝液清洗,再吸乾殘餘的 PBS 緩衝液,分別加入 500μl 的 TRIzol reagent ( Invitrogen Corpoation, California, USA ),搖晃均勻置於冰上 5 分鐘,待 細胞完全溶解,將細胞刮下,移至另一微量離心管中。室溫下靜置 5 分鐘再加 入100μl 氯仿 ( chloroform ) 劇烈振盪 1 分鐘,靜置於室溫 3 分鐘,於 4 0C 以 12,000 rpm 離心 15 分鐘,樣品會形成 phenol-chloroform,中間白色層以及上 面透明水層。 RNA 大部分留在水層,小心將水層轉移到新的微量離心管,加 入 等量之異丙醇 ( Isopropanol ) 沉澱核酸,靜置於室溫 10 分鐘後,於 4 0C 以 12,000 rpm 離心 15 分鐘,小心地去除上清液,可發現 RNA 在微量離心管底 部形成乳白色塊狀沉澱。加入 1ml 的 75 % 乙醇 ( 以 DEPC 水稀釋 ) 將沉澱 物沖離管壁,以 4 0C 7,500 rpm 離心 5 分鐘,去除上清液,將微量離心管倒放 置於室溫下風乾,最後加入 20μl 的 DEPC 去離子水回溶 RNA 並儲存於 -80 0. C。取 1μl RNA 加上 99μl 的 DEPC 水,利用分光光譜儀 ( OD260 /OD 280 nm ). 偵測 RNA 吸光值( UV/Visible spectrophotometer DU-800, Beckman Coulter )。. 第十一節、反轉錄聚合酶連鎖反應 ( RT-PCR ) 使用 SSⅢkit ( Invitrogen, Carlsbad, CA ) 將 RNA 轉成 cDNA,1、在微量 離心管內加入:DNTP ( 1μl )、Olgio ( 1μl )、RNA ( 2μg ),再加入 DEPC 水至 10μl 後,開始進行聚合反應。首先於 65 0C 作用 5 min 以 denature RNA,置冰上 1 分鐘。 2、每管再加入 10X RT buffer ( 2μl )、0.1M DTT ( 2μl )、 RNase out ( 1μl )、25 mM Mgcl2 ( 4μl ),混合好於 42℃ 作用 2 分鐘。 3、接著每管加入 SS Ⅲ ( 1μl ) 後放置 42 0C 作用 50 分鐘,然後於 70 0C 作用 15 分鐘,接著放入 冰上。 4、加入 RNAse H ( 1μl ) 後於 37 0C 作用 20 分鐘使反應完全,即是 cDNA,儲存於 -20 0C。. 23.
(34) 第十二節、即時定量聚合酶連鎖反應 ( Real-time Polymerase Chain Reaction, Real-time PCR ) 即時聚合酶連鎖反應 ( Real-time Polymerase Chain Reaction, Real-time PCR ) 的原理為在進行 PCR 時同步偵測 PCR 產物隨著反應增加的狀況 ( 即為“real time” ) 利用所偵測到的產物回推樣品中原始基因之表現量。Real-time PCR 可分 為兩大方式:TaqMan 及 SYBR® GreenERTM ( Invitrogen, Carlsbad, CA )。本研 究使用相對定量 SYBR® GreenERTM 的原理是先設計一個 primer,而此 primer 是設計在我們要的一段 DNA 序列中,可以用來與單股 DNA 黏合,藉由 SYBR® GreenERTM qPCR SuperMix 在 PCR 過程時,與 DNA 序列黏合的 primer,隨著 PCR 產物的增加,信號也隨著增強,因此可做相對定量的分析。 將 cDNA 取 10~100ng、SYBR® GreenERTM qPCR SuperMix 取 10 μl、10μM primer 取 0.8μl , 接 著 以 DEPC 水 加 至 20 μl 後 , 將 樣 本 放 置 Applied Biosystems 7900 ( ABI prism 7900, CA, USA ) 進行相對定量分析。TaqMan real-time PCR 是用帶有螢光標記及螢光接受體的探針 ( probes ) : α2、α5、αν、β1、 β3 ( ABI, CA, USA ) 加入 PCR 反應。將 cDNA 取 10~100ng、TaqMan ® Master Mix (Invitrogen, Carlsbad, CA ) 取 10 μl、10μM primer 取 0.8μl,接著以 DEPC 水加 至 20 μl 後,將樣本放置 Applied Biosystems 7900 ( ABI prism 7900, CA, USA ) 進行相對定量分析。 生工引子訂購 AdipoR1(F). CCTTTCCCCAAGCTGAAGCTGC. AdipoR1(R). CCTTGACAAAGCCCTCAGCGAT. AdipoR2(F). AACGAGCCAACAGAAAACCGATTG. AdipoR2(R). ATACACACAGAAACAGGCAACATTTG. 第十三節、統計分析 所有實驗均會進行三次重複,處理組與控制組的數據間,並以 student’s t-test, p ‹ 0.05,進行分析其結果是否有統計上差異。. 24.
(35) 第三章、實驗結果 (Result). 25.
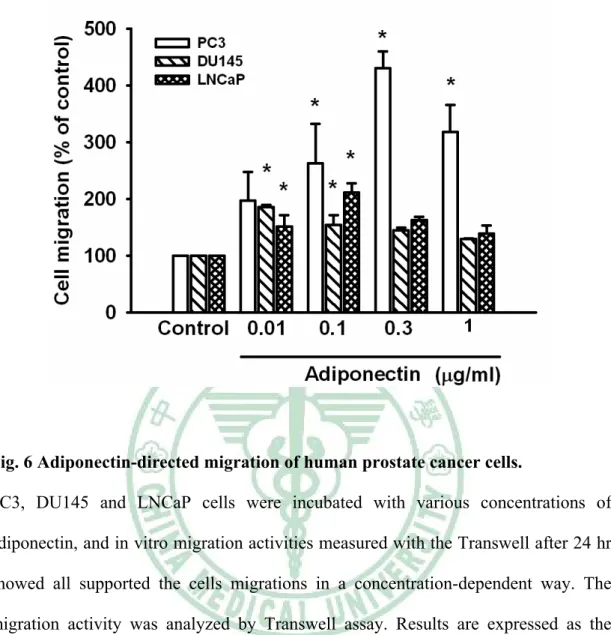
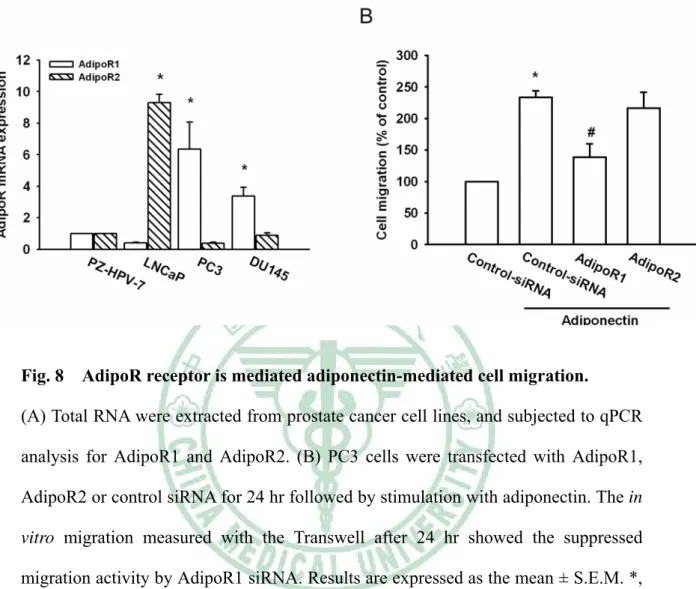
(36) Adiponectin 引發人類前列腺腫瘤細胞轉移 先前研究指出. adiponectin 能 刺 激 人 類 腫 瘤 細 胞 的 轉 移 和 侵 入. ( Spyridopoulos TN et al., 2007;Bråkenhielm E et al., 2004;Bub JD et al., 2006)。 本研究中,我們選用三株不同的人類前列腺腫瘤細胞株 ( PC3、DU145 及 LNCaP ) 進行 adiponectin 與人類前列腺腫瘤細胞轉移的關係。我們分別給予細 胞不同濃度之 adiponectin,而依濃度增加造成 androgen-independent 的前列腺腫 瘤細胞 ( PC3、DU145 ) 移行,也直接影響了 androgen-dependent 的前列腺腫瘤 細胞移行 ( LNCaP );在濃度 0.3 μg/ml 時,PC3 的移行能力表現有顯著的差異 (Fig.5)。. Adiponectin 造成人類前列腺腫瘤細胞移行與增加 α5β1 integirn 表現有關 先前的研究顯示,人類腫瘤細胞中有明顯的 integrins 表現 ( Wei Y et al., 2007 )。於是我們假設 integrins 可能參與在 adiponectin 影響前列腺腫瘤細胞的轉 移過程中。流式細胞技術分析 ( Flow cytometric analysis ) 與即時定量聚合酶連 鎖反應 ( qPCR ) 顯示,adiponectin 誘導了細胞表面上 α5β1 integrins 表現,而 在 α2、αν、β1、β3 及 α2β1 integrins 則沒有顯著的影響 ( Fig. 6A &6 B )。用 anti-α5β1 單株抗體 ( mAb ) ( 10 μg/ml ) 處理細胞 30 分鐘後進行細胞移行實驗 ( Migration assay ) ,實驗結果顯示抑制由 adiponectin 誘導的腫瘤細胞轉移 ( Fig.6C )。以上結果提出 adiponectin 是經由調控增加 α5β1 integrins 的表現來增 加前列腺腫瘤細胞的轉移。. AdipoR1/R2 接受器的參與和人類前列腺腫瘤細胞的轉移有關 之前報告指出,AdipoR1 大量表現於骨骼肌,AdipoR2 則主要表現於肝臟 ( Yamauchi T et al., 2003 )。然而 AdipoR1/R2 接受器在人類前列腺腫瘤細胞的表 現不是很明瞭。我們經由即時定量聚合酶連鎖反應來檢測前列腺腫瘤細胞之 AdipoR1 和 AdipoR2 接受器的表現。 PC3 及 DU145 之 AdipoR1 接受器的表現 26.
(37) 比人類正常前列腺細胞 PZ-HPV-7 來的顯著;LNCaP 則是 AdipoR2 接受器表現 較為顯著 ( Fig.7A )。過渡性轉染 ( transient transfection ) AdipoR1 之短干擾 RNA 於 PC3,有效地抑制前列腺腫瘤細胞的移行 ( Fig.7B )。這些結果提出了 adiponectin/AdipoR 接受器的交互作用在前列腺腫瘤細胞轉移中扮演一個重要 的角色。 P38 和 AMPK 的訊息傳遞路徑與 adiponectin 的調控作用有關 最近文獻指出 adiponectin 除了在 C2C12 細胞中活化 AMPK 也活化了 p38 MAPK ( Tang CH et al., 2007 )。我們為了測試 p38 是否與 adiponectin 導致的細 胞移行能力有關;給予細胞 adiponectin 後,p38 磷酸化表現隨著時間增加 ( Fig.8A;上欄處)。接著我們直接檢測 p38 激酶活性對 adiponectin 的反應;藉 由偵測出 p38 激酶之其中一種基質 [activating transcription factor (ATF)-2] 的磷 酸化表現顯示 adiponectin 的確增加了 p38 的活性 ( Fig.8A;下欄處)。當給予 前列腺癌細胞 ( PC3、DU145 及 LNCaP ) p38 抑制劑 ( SB203580;10 μM ) ( Fig.8B );或是轉染 PC3 細胞之 p38 mutant 24 小時後 ( Fig.8C ),進行細胞移 行實驗、流式細胞技術分析 ( Fig.8D ) 及即時定量聚合酶連鎖反應 ( Fig.8E ), 可觀察到前列腺癌細胞株 PC3 的移行能力及 α5β1 的向上調節作用皆有顯著的 抑制現象。因此 p38 活化參與在 adiponectin 所調節細胞移行與 integrin 表現 中。研究顯示 adiponectin 會透過 AMPK 的活化增加脂肪酸的氧化作用 ( Tomas E et al., 2002 )。因此,接下來研究是否 AMPK 活化參與 adiponectin 作用中。 Adiponectin 隨著時間增加造成 AMPKα 在 Thr172 的磷酸化 ( Fig.9A )。更進一步 地,給予細胞 AMPK 抑制劑 ( AraA;0.5 mM ) 或 ( Compound C;10μM ) 30 分鐘後明顯地抑制了 adiponectin 所誘導的細胞移行能力;此外,還給予細胞 AMPK 活化劑 ( AICAR; 1mM ) 而增加了細胞移行能力 ( Fig.9B )。當給予前列 腺癌細胞 PC3 AMPK 抑制劑 ( AraA;0.5 mM ) 或 ( Compound C;10μM ),進 行流式細胞技術分析 ( Fig.9D ) 及即時定量聚合酶連鎖反應 ( Fig.9E ),可觀察 27.
(38) 到前列腺腫瘤細胞 PC3 的移行能力及 α5β1 的向上調節作用皆有顯著的抑制 現象。 為 了 確 定 哪 一 種 型 態 的 AMPKα1 或 AMPKα2 催 化 次 單 位 會 調 節 adiponectin 在前列腺腫瘤細胞的訊息傳遞;轉染 PC3 細胞之短干擾 RNA ( AMPK 1 或 AMPK 2 ) 24 小時後,進行細胞移行實驗;可觀察到 AMPKα1 明顯地抑制前列腺腫瘤細胞 PC3 的移行能力 ( Fig.9C ),因此 AMPKα1 參與 adiponectin 的作用之中。為了研究 p38 與 AMPK 上下游關係,給予細胞之 p38 抑制劑 ( SB203580;10 μM ),很明顯地抑制了 adiponectin 誘導的 AMPK 活性; 進一步地給予細胞之 AMPK 抑制劑 ( AraA;0.5 mM ) 或 ( Compound C; 10μM ),並不會影響 adiponectin 所增加 p38 kinase activity。因此,p38 可能作 為上游因子傳遞訊號給下游因子 AMPK。以上結果指出,p38 及 AMPK 訊號傳 遞路徑參與調控由 adiponectin 誘導的前列腺腫瘤細胞轉移。. NF-κB 訊號路徑參與調控由 adiponectin 誘導 integrin 的向上調節與細胞的移 行能力 之前研究指出,在人類腫瘤細胞的轉移和侵入中,與 NF-κB 轉錄因子的活 化參與有關 ( Boukerche H et al., 2007 )。為了進一步證明 NF-κB 訊號傳遞路徑 參與在由 adiponectin 調控增加前列腺腫瘤細胞的轉移中;我們給予細胞 NF-κB 抑制劑 ( PDTC;60μM ) 或 IκBα protease 抑制劑 ( TPCK;3μM ) 30 分鐘後。 (Fig.10A) 顯示由 adiponectin 誘導的前列腺腫瘤細胞的移行被抑制了;此外, 給予細胞 ( PDTC;60μM ) 或 ( TPCK;3μM ) 進行流式細胞技術分析及即時定 量聚合酶連鎖反應,顯示也拮抗了 adiponectin 誘導 α5β1 integrin 向上調節的作 用 ( Fig.10B & 10C )。以上結果指出 NF-κB 活化參與 adiponectin 誘導腫瘤細 胞移行以及 α5 和 β1 integrins 表現。. 28.
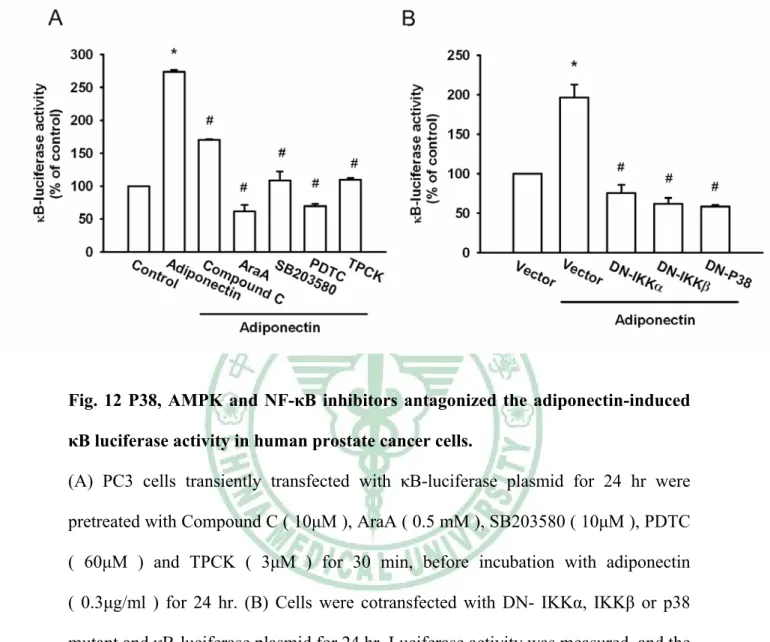
(39) 我們接下來研究 adiponectin 誘導 NF-κB 活化的上游因子與前列腺腫瘤細胞移 行的關係。首先依時間點給予 adiponectin 刺激細胞誘導了 IKKα/β 磷酸化 ( Fig.10D;上欄處)。另一方面依不同時間點給予 PC3 細胞之 adiponetin 後,也 使得 IκBα 的磷酸化隨著時間而增加 ( Fig.10D;中欄處)。先前的研究顯示 p65 Ser536 磷酸化增加 NF-κB 的轉錄活化能力 ( Madrid LV et al., 2001;Viatour P et al., 2005 ) 。並且 p65 Ser536 磷酸化專一性的抗體也用來驗證 p65 Ser536 的磷酸 化。因此,依不同時間點給予 PC3 細胞之 adiponetin 後,造成了 p65 Ser536 的磷酸化隨著時間而增加 ( Fig.10D;下欄處)。接著將 IKKα 或 IKKβ mutant 轉 染 PC3 細胞 24 小時後,進行細胞移行實驗;可觀察到 IKKα 或 IKKβ mutant 明 顯地抑制前列腺腫瘤細胞 PC3 的移行能力 ( Fig.10E )。這些結果顯示 IKKα 與 IKKβ 參與 adiponectin 作用之中。更進一步地,觀察 NF-κB 之活化作用,將 κB-luciferase 轉染入 PC3 細胞 24 小時,作為 NF-κB 活性的指示因子。給予 PC3 細 胞 之 adiponetin , κB-luciferase 的 活 性 會 增 加 ; 此 外 給 予 Compound C ( 10μM )、 AraA ( 0.5 mM )、 SB203580 ( 10μM )、 PDTC ( 60μM ) 及 TPCK ( 3μM ),皆能降低由 adiponetin 所誘導的 NF-κB 活性 ( Fig.11A )。將 PC3 細胞 一 同 轉 染 IKKα 、 IKKβ 或 p38 mutant 也 能 抑 制 NF-κB 促 進 因 子 的 活 性 ( Fig.11B )。綜合以上結果顯示,活化 p38 和 AMPK 參與在 adiponetin 所增加 NF-κB 的活化中。. 29.
(40) 第四章、討論 (Discussion). 30.
(41) 前列腺是男性生殖系統中的一個器官,屬外分泌腺,位在膀胱的下方,直腸 的前方。而主要功能是分泌和儲存前列腺液。前列腺液,含有抗菌因子保護尿道, 經前列腺腺管,排到後尿道,隨尿液排出體外。前列腺腺管内存有前列腺液,前 列腺液可與精子混合成精液。精液包含了精子和液狀物,此液狀物約有 10%-30% 是來自前列腺所製造 ( Haas GP et al., 1997)。 前列腺癌在歐美病率極高,在高齡男性中僅次於肺癌,但在台灣比較少見, 近年來發病率逐漸增加中。前列腺腫瘤細胞常常發生繼發性骨轉移 ( Greenlee RT et al., 2001;Mundy GR et al., 2002 )。除皮膚癌外,前列腺癌是全世界第二因 癌症導致死亡的疾病,僅次於肺癌 ( Mistry T et al., 2007 )。癌症最可怕的地方在 於病情進入轉移 ( metastasis ) 階段,癌細胞就會擴散到身體其他的器官組織, 而在癌症的死亡病例中,絕大部分元兇是癌細胞轉移至其他重要器官如:肺臟、 肝臟、骨骼與腦部等,只有不到 10% 是由最先長出的原發腫瘤引起。 前列腺腫瘤在先進國家中有廣泛多變的臨床病程,然而,近乎有八分之一案 例普遍有轉移情形。大多典型地皆轉移到骨頭 ( Mundy GR et al., 2002 )。前列腺 腫瘤生長和存活是依賴著雄性激素賀爾蒙。廣泛轉移的案例治療首選為荷爾蒙療 法,也就是阻斷雄性激素---睪丸酮對前列腺癌細胞的作用,進而達到抑制癌細 胞的增生及轉移的抗癌效果。 本文一開始,我們利用人類前列腺腫瘤細胞確認 adiponectin 是否會造成細 胞的移行能力。在這裡我們發現 adiponecin 促進 androgen-independent ( PC3 、 DU145 ) 和 androgen-dependent ( LNCaP ) 的前列腺腫瘤細胞的轉移。許多研究 初步的數據提出 adiponectin 具有抗癌的作用 ( Dieudonne MN et al., 2006 ),而 具有較低的 adiponectin 卻被指出具有增加胃癌、大腸癌、乳癌和內皮細胞癌的 風險 ( Dal Maso L et al., 2004 ; Ishikawa M et al., 2005 ; Kadowaki T et al., 2005) 。 因 此 , 由 我 們 的 結 果 發 現 , 不 論 是 androgen-dependent 或 androgen-independent 的細胞中,adiponectin 都可以促進細胞之移行。. 31.
(42) Adiponecin 交互作用於兩個接受器 AdipoR1 和 AdipoR2。最近被定義出 adiponecin 調 節 生 物 特 性 透 過 adiponecin 之 接 受 器 AdipoR1 和 AdipoR2 ( Yamauchi T et al., 2003 )。AdipoR1 接受器對球狀脂聯素 ( globular adiponectin ) 具有高的親合力;AdipoR1 則是對長鏈脂聯素 ( full-length adiponectin ) 有較差 的親合力,而 AdipoR2 接受器對兩種形式的脂聯素皆具有親和性 ( Yamauchi T et al., 2003 ) 。 AdipoR1 大 量 表 現 於 骨 骼 肌 , AdipoR2 則 是 表 現 在 肝 臟 ( Kawaguchi T et al., 2005 )。然而,AdipoR1 和 AdipoR2 接受器表現在人類前列 腺腫瘤細胞還不是很明瞭。我們測試了 androgen-independent ( PC3、DU145 ), androgen-dependent ( LNCaP ) 和正常前列腺細胞,這四株細胞之 AdipoR1 和 AdipoR2 mRNA 的表現量。發現四株細胞皆有表現 AdipoR1 和 AdipoR2 。 PC3 和 DU145 的 AdipoR1 表 現 量 增 加 ; LNCaP 則 是 大 量 表 現 AdipoR2 。 將 AdipoR1 或 AdipoR2 之短干擾 RNA 轉染至細胞中,AdipoR1 之短干擾 RNA 明 顯地抑制細胞的移行。我們推測由 adiponecin 增加前列腺腫瘤細胞移行,必須 經由 AdipoR1 啟動下游的訊息傳遞。 Integrins 在細胞轉移和黏附上特別扮演了重要的角色。它 是 由 α 及 β 兩 種 穿膜次 單 位 以 非 共 價 鍵 的 方 式 所 組 成 的 。在哺乳動物細胞,19 個 α 和 8 個 β 次單位連結成多種組合體,相 互 配 對 組 合 形成 24 種的 integrins,可以結 合到不同種類的細胞外基質 ( Extracellular matrix,ECM ) 的配 位 體 上,發 生 作 用 ( Hynes RO et al., 2002 )。先前的研究顯示在數種腫瘤細胞上, integrins 調控 了細胞的轉移和侵襲 ( Albelda SM et al., 1990;Hieken TJ et al., 1996 )。然而,在 人類的前列腺腫瘤細胞藉由 adiponecin 使得 integrins 表現仍大部分是未知 的。在這裡我們利用流式細胞技術分析與即時定量聚合酶連鎖反應分析發現 adiponecin 增加 α5β1 integrins 表現而 α2、αν、β1、β3 及 α2β1 integrins 則沒有 顯著的影響,所以 integrins 的轉錄調控在腫瘤細胞轉移時扮演著重要的角色。 本研究中,我們也使用了 anti-α5β1 單株抗體去觀察 α5β1 integrins 所扮演的角 色,並且發現他能抑制由 adiponecin 誘導的細胞轉移,這表示了 α5β1 integrins 32.
(43) 可能參與在 adiponecin 誘導前列腺腫瘤細胞轉移以及 integrins 轉錄調控表現。 AMPK 是一種由催化作用的 α 與調節作用的 β 和 γ 三個次蛋白組成的 heterotrimeric serine/threonine 激酶 ( Carling D et al., 2004 )。過去研究顯示 AMPK 與 adiponectin 的代謝的作用的訊息路徑有關 ( Carling D et al., 2004 )。有 研究顯示 adiponecin 交互作用於兩個接受器 AdipoR1 和 AdipoR2,調節增加 AMPK、 PPARα 脂肪酸氧化的表現 ( Kawaguchi T et al., 2005 )。而在 C2C12 肌 肉細胞和肝臟細胞皆表現 AdipoR1/R2 ,adiponectin 刺激 PPARα、AMPK 和 p38 MAPK 的活化、葡萄糖再回收和脂肪酸的氧化作用。抑制 AMPK 或 PPARα 減少了由 adiponectin 刺激的脂肪酸氧化作用,並且部分減少了葡萄糖的再回收 ( Yamauchi T et al., 2003;Kadowaki T et al., 2005 )。我們利用 AMPK 抑制劑 ( AraA ) 或 ( Compound C ) 去抑制由 adiponectin 所誘導的細胞移行,這結果提 出了活化 AMPK 造成 adiponectin 誘導細胞的移行能力是必須的。我們也發現藉 由 AMPKα1 之短干擾 RNA 減少了由 adiponectin 所誘導的細胞移行。由文獻 中得知 AMPK 與 p38 交互作用進而調節了葡萄糖的代謝 ( Xi X et al., 2001 )。 進一步地藉由 p38 抑制劑 ( SB203580 ) 與 p38 mutant 減少了 adiponectin 所誘 導的細胞移行。確定了在由 adiponectin 調節的細胞移行和 integrin 的調節中是 經由 AdipoR-AMPK-dependent p38 的活化。 許 多 報 告 指 出 NF-κB 的 活 化 是 參 與 在 integrins 表 現 和 腫 瘤 轉 移 當 中 ( Huang YC et al., 2007 )。在本實驗中,我們證實 adiponecin 誘導的移行和 integrins 表現可被 NF-κB 抑制劑 ( PDTC ) 或 IκBα protease 抑制劑 ( TPCK ) 來抑制,因此 NF-κB 的活化可能參與了 adiponecin 誘導 integrins 表現之中。 NF-κB 轉錄因子在免疫與發炎反應中能調控許多基因的多向性。NF-κB 家族蛋 白包含 Rel 家族的同體或異體雙體次單位,包括 p50 和 p65 ( Baldwin AS Jr et al., 1996 )。在不活化的狀態下,NF-κB 藉由抑制蛋白 IκBα 的抑制,而正常地 保留在細胞質當中。例如像是 TNF-α 刺激後,IκBα 蛋白藉由 IKK 複合體將其 磷酸化,伴隨而來的是 IκB 被泛蛋白化 ( ubiquitination ),並且隨後被 26S 33.
(44) proteinase 降解。最後被釋放的 NF-κB 轉錄至細胞核內,而活化反應的基因 ( Vermeulen L et al., 2003 )。在本研究中,我們發現 給予 adiponectin 刺激下的 PC3 細胞造成了 IKK、IκB 以及 p65 的磷酸化。而利用過渡性轉染作 用 ( transient transfection ) κB-luciferase 來 反 應 NF-κB 活 性 , 我 們 也 發 現 adiponectin 可增加 NF-κB 活性。在這研究當中,也發現了 adiponectin 誘導 IKK、IκBα 以及 p65 的磷酸化可被 NF-κB 抑制劑 ( PDTC ) 或 IκBα protease 抑制劑 ( TPCK ) 抑制。 IKKα 或 IKKβ mutant 也減少了由 adiponectin 增加 IKK、 IκBα 和 p65 的磷酸化。. 34.
(45) 第五章、結論 (Conclusions). 35.
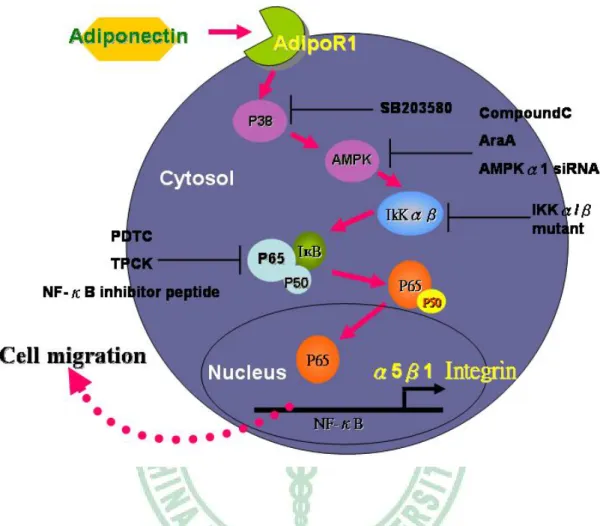
(46) 本研究首次表示了 adiponectin 調控前列腺腫瘤細胞的移行。幫助我們了解 人類前列腺細胞的轉移機制,強調其中一個機制是藉由活化 AdipoR1 receptor、 AMPK、p38 和 NF-κB 訊息傳導路徑,增加 α5β1 integrins 的轉錄調控 (Fig.9)。 以上推論能需要做進一步的研究與探討,以找出其真正的機制,在未來或許可以 應用在治療癌症上。. 未來展望與建議 台灣的前列腺癌發生率少於歐美國家,根據行政院衛生署癌症死亡統計數據 指出,國人罹患前列腺癌近年來有增加的趨勢。由於前列腺癌初期沒有明顯的症 狀, 腫瘤進展速度緩慢,所以在台灣無法得知較確切的發生率。近年來前列腺 癌之生物行徑 ( biologic behavior ) 慢慢被研究,基因治療逐漸進步成熟,渴望 這方面前瞻性的發展對台灣前列腺癌的治療與預後有實質上的幫助。. 36.
(47) Fig. 6 Adiponectin-directed migration of human prostate cancer cells. PC3, DU145 and LNCaP cells were incubated with various concentrations of adiponectin, and in vitro migration activities measured with the Transwell after 24 hr showed all supported the cells migrations in a concentration-dependent way. The migration activity was analyzed by Transwell assay. Results are expressed as the mean±S.E.M. *, p <0.05 compared with control. #, p <0.05 as compared with adiponectin -treated group.. 37.
(48) Fig. 7. 38.
(49) Fig. 7 Adiponectin-directed migration of human prostate cancer cells involves up-regulation of α5β1 integrin. (A) PC3 cells were incubated with adiponectin (0.3μg/ml) for 24 hr, and the cell surface expression of α2, α5, β1, β3, α2β1, or α5 integrins were determined using flow cytometry. (B) Cells were incubated with adiponectin (0.3μg/ml) for 24 hr, and the mRNA level of α2, α5, αv, β1 or β3 integrins were determined using qPCR. (C) Cells were pretreated for 30 min with α5β1 integrin followed by stimulation with adiponectin (0.3μg/ml) and in vitro migration activitie measured with the Transwell after 24 hr showed α5β1 integrin supported the cells migration. Results are expressed as the mean ± S.E.M. *, p < 0.05 compared with control;#, p < 0.05 compared with adiponectin-treated group.. 39.
(50) Fig. 8. AdipoR receptor is mediated adiponectin-mediated cell migration.. (A) Total RNA were extracted from prostate cancer cell lines, and subjected to qPCR analysis for AdipoR1 and AdipoR2. (B) PC3 cells were transfected with AdipoR1, AdipoR2 or control siRNA for 24 hr followed by stimulation with adiponectin. The in vitro migration measured with the Transwell after 24 hr showed the suppressed migration activity by AdipoR1 siRNA. Results are expressed as the mean ± S.E.M. *, p < 0.05 compared with control; #, p < 0.05 compared with adiponectin-treated group.. 40.
(51) Fig. 9. 41.
(52) Fig. 9. P38 is involved in adiponectin-mediated cell migration and integrin. up-regulation in human prostate cancer cells. (A) PC3 cells were incubated with adiponectin for indicated time intervals, and phosphor -p38 expression was determined by Western blot analysis (upper panel). The p38 activity was also examined by p38 activity kit (lower panel). Cells were pretreated for 30 min with SB203580 (10 μM) (B) or transfected with dominant negative (DN) mutant of p38 (C) for 24 hr followed by stimulation with adiponectin, and in vitro migration (B&C) cells surface α5β1 integrin (D) and mRNA expression of α5β1 integrin were measured with the Transwell and flow cytometry or qPCR (E) after 24 hr. Results are expressed as the mean ± S.E. *, p < 0.05 compared with control; #, p < 0.05 compared with adiponectin-treated group.. 42.
(53) Fig. 10. 43.
(54) Fig. 10 AMPK pathway is involved in adiponectin-mediated integrin up-regulation in human prostate cancer cells. (A) PC3 cells were incubated with adiponcetin (0.3μg/ml) for indicated time intervals, and phosphor-AMPK Thr172 (upper panel) or AMPK (lower panel) expression was determined by Western blot analysis. Cells were pretreated with AraA (0.5 mM) and compound C (10μM) for 30 min followed by stimulation with adiponectin or treated with 5-Aminoimidazole-4-caroxamide-1-β-D-ribofuranoside [AICAR (1 mM)] for 24 hr. The in vitro migration activity and migration expression was examined by Transwell (B) flow cytometry (D) and qPCR (E). (C) Cells were transfected with AMPKα1, AMPKα2 or control siRNA for 24 hr followed by stimulation with adiponectin. The in vitro migration measured with the Transwell after 24 hr. (F) Cells were pretreated with SB203580 (10μM) for 30 min followed by stimulation with adiponectin (0.3μg/ml) for 60 min, and phosphor-AMPK Thr172 expression was determine by Western Blot analysis (G) Cells were pretreated with AraA (0.5 mM) and Compound C (10μM) for 30 min followed by stimulation with adiponectin (0.3μg/ml) for 60 min, and p38 activity were then examined by using p38 activity kit. Results are expressed as the mean± S.E.M. *, p < 0.05 compared with control; #, p < 0.05 compared with adiponectin–treated group.. 44.
(55) Fig. 11. 45.
(56) Fig. 11 Adiponectin induces cell migration and integrins up-regulation through NF-κB. (A) Cells were pretreated for 30 min with PDTC (60μM) or TPCK (3μM) followed by stimulation with adiponectin (0.3μg/ml), and in vitro migration was measured with the Transwell after 24 hr. (B) PC3 cells were pretreated for 30min with PDTC (60μM) or TPCK (3μM) followed by stimulation with adiponectin (0.3μg/ml) for 24hr, and the cell surface α5β1 integrin was measured by flow cytometry. (C) PC3 cells were pretreated for 30min with PDTC (60μM) or TPCK (3μM) followed by stimulation with adiponectin (0.3μg/ml) for 24hr, and mRNA expression of α5 and β1 integrin was measured by qPCR. (D) PC3 cells were incubated with adiponectin (0.3μg/ml) for indicated time intervals, and p-IKKα/β, p-IκBα, and p-p65 expression was determined by Western blot analysis. (E) PC3 cells were transfected with dominant negative (DN) mutant of IKKα or IKKβ for 24 hr followed by stimulation with adiponectin (0.3μg/ml), and in vitro migration was measured with the Transwell after 24 hr. Results are expressed as the mean± S.E.M. *, p < 0.05 compared with control; #, p < 0.05 compared with adiponectin–treated group.. 46.
(57) Fig. 12 P38, AMPK and NF-κB inhibitors antagonized the adiponectin-induced κB luciferase activity in human prostate cancer cells. (A) PC3 cells transiently transfected with κB-luciferase plasmid for 24 hr were pretreated with Compound C ( 10μM ), AraA ( 0.5 mM ), SB203580 ( 10μM ), PDTC ( 60μM ) and TPCK ( 3μM ) for 30 min, before incubation with adiponectin ( 0.3μg/ml ) for 24 hr. (B) Cells were cotransfected with DN- IKKα, IKKβ or p38 mutant and κB-luciferase plasmid for 24 hr. Luciferase activity was measured, and the results were normalized to the β-galactosidase activity. Results are expressed as the mean± S.E.M. *, p < 0.05 compared with control;#, p < 0.05 compared with adiponectin -treated group.. 47.
(58) Fig. 13. Schematic diagram of the signaling pathways involved in adiponectin-. induced cell migration in human prostate cancer cell. Adiponectin increases migration activity by binding to the AdipoR1 receptor and activation of p38, AMPKα1, IKKα/β phosphorylation, which enhances binding of p65 to the NF-κB site, lead to α5β1 integrin expression and increases the migration of human prostate cancer cell.. 48.
(59) 參考文獻 Albelda SM et al. Integrin distribution in malignant melanoma: association of the beta 3 subunit with tumor progression.Cancer Res. 1990 Oct 15;50 (20):6757-64. Alimirah F et al. DU-145 and PC-3 human prostate cancer cell lines express androgen receptor: implications for the androgen receptor functions and regulation. FEBS Lett. 2006 Apr 17;580 (9):2294-300. Baldwin AS Jr et al. The NF-kappa B and I kappa B proteins: new discoveries and insights. Annu Rev Immunol. 1996 Apr ;14:649-83. Böttger TC et al. Prognostic value of immunohistochemical expression of beta-1 integrin in pancreatic carcinoma. Oncology. 1999 May ;56 (4):308-13. Boudreau N et al. Extracellular matrix signaling: integration of form and function in normal and malignant cells. Curr Opin Cell Biol. 1998 Oct ;10 (5):640-6. Boukerche H et al. mda-9/Syntenin regulates the metastatic phenotype in human melanoma cells by activating nuclear factor-kappaB. Cancer Res. 2007 Feb 15;67 (4):1812-22. Bråkenhielm E et al. Adiponectin-induced antiangiogenesis and antitumor activity involve caspase-mediated endothelial cell apoptosis. Proc Natl Acad Sci U S A. 2004 Feb 24;101 (8):2476-81. Bub JD et al. Adiponectin as a growth inhibitor in prostate cancer cells. Biochem Biophys Res Commun. 2006 Feb 24; 340 (4):1158-66. Carling D et al. The AMP-activated protein kinase cascade--a unifying system for energy control. Trends Biochem Sci. 2004 Jan;29 (1):18-24. Chu CY et al. Involvement of matrix metalloproteinase-13 in stromal-cell-derived factor 1 alpha-directed invasion of human basal cell carcinoma cells. Oncogene. 2007 Apr 12;26 (17):2491-501.. 49.
(60) Cooper CR et al. The role of alpha(v)beta(3) in prostate cancer progression.Neoplasia. 2002 May-Jun;4 (3):191-4. Culig Z et al. Androgen axis in prostate cancer. J Cell Biochem. 2006 Oct 1;99 (2):373-81. Dal Maso L et al. Circulating adiponectin and endometrial cancer risk. J Clin Endocrinol Metab. 2004 Mar;89(3):1160-3. Dehm SM et al. Molecular regulation of androgen action in prostate cancer. J Cell Biochem. 2006 Oct 1;99 (2):333-44. Delongchamps NB et al. The role of prevalence in the diagnosis of prostate cancer. Cancer Control. 2006 Jul;13 (3):158-68. Dieudonne MN et al. Adiponectin mediates antiproliferative and apoptotic responses in human MCF7 breast cancer cells. Biochem Biophys Res Commun. 2006 Jun 23;345 (1):271-9. Fornaro M et al. Integrins and prostate cancer metastases. Cancer Metastasis Rev. 2001 Nov ;20 (3-4):321-31. Freedland SJ et al. Association between serum adiponectin, and pathological stage and grade in men undergoing radical prostatectomy. J Urol. 2005 Oct;174 (4 Pt 1):1266-70. Giancotti FG et al. Integrin signaling. Science. 1999 Aug 13;285 (5430):1028-32. Goktas S et al. Prostate cancer and adiponectin. Urology. 2005 Jun;65 (6):1168-72. Greenlee RT et al. Cancer statistics, 2001. CA Cancer J Clin. 2001 Jan-Feb; 51(1):15-36 Guo W et al. Integrin signalling during tumour progression. Nat Rev Mol Cell Biol. 2004 Oct;5 (10):816-26. Haas GP et al. Epidemiology of prostate cancer. CA Cancer J Clin. 1997 Sep-Oct ; 47(5):273-87. 50.
(61) Hieken TJ et al. Beta3 integrin expression in melanoma predicts subsequent metastasis.J Surg Res. 1996 Jun;63 (1):169-73. Hsing AW et al. Prostate cancer epidemiology. Front Biosci. 2006 May 1;11:1388-413. Hu E et al. AdipoQ is a novel adipose-specific gene dysregulated in obesity. J Biol Chem. 1996 May 3;271 (18):10697-703. Huang YC et al. Stromal cell-derived factor-1 enhances motility and integrin up-regulation through CXCR4, ERK and NF-kappaB-dependent pathway in human lung cancer cells. Biochem Pharmacol. 2007 Dec 15;74 (12):1702-12. Huttenlocher A et al. Modulation of cell migration by integrin-mediated cytoskeletal linkages and ligand-binding affinity. J Cell Biol. 1996 Sep;134 (6):1551-62. Hynes RO et al. Integrins: bidirectional, allosteric signaling machines. Cell. 2002 Sep 20;110 (6):673-87. Ishikawa M et al. Plasma adiponectin and gastric cancer. Clin Cancer Res. 2005 Jan 15;11(2 Pt 1):466-72. Kadowaki T et al. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J Clin Invest. 2006 Jul;116 (7):1784-92. Kadowaki T et al. Adiponectin and adiponectin receptors. Endocr Rev. 2005 May;26(3):439-51. Knudsen BS et al. The impact of cell adhesion changes on proliferation and survival during prostate cancer development and progression. J Cell Biochem. 2006 Oct 1;99 (2):345-61. Kumar CC et al. Integrin alpha v beta 3 as a therapeutic target for blocking tumor-induced angiogenesis. Curr Drug Targets. 2003 Feb;4 (2):123-31. Madrid LV et al. Akt stimulates the transactivation potential of the RelA/p65 Subunit of NF-kappa B through utilization of the Ikappa B kinase and activation of the 51.
數據


+3
相關文件
Reading Task 6: Genre Structure and Language Features. • Now let’s look at how language features (e.g. sentence patterns) are connected to the structure
Promote project learning, mathematical modeling, and problem-based learning to strengthen the ability to integrate and apply knowledge and skills, and make. calculated
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
In BHJ solar cells using P3HT:PCBM, adjustment of surface energy and work function of ITO may lead to a tuneable morphology for the active layer and hole injection barrier
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
Most experimental reference values are collected from the NIST database, 1 while other publications 2-13 are adopted for the molecules marked..
In case of any disputes regarding any competition(s), and/or other related matters, the decision of the Organiser shall be final, binding and
Binding Warning message Binding Update message AAAO: the AAA server of the old foreign network to which the OFA belongs. AAAF: the AAA server of the new foreign network to which the
