基質金屬蛋白酶基因多型性及酵素活性
在兒童心室中膈缺損的相關性
Genetic Variations and Circulating Levels of Matrix Metalloproteinases in
the Children with Ventricular Septal Defect
研 究 生 :鄭崑山
Student:Kun-Shan Cheng
指導教授:林志生 博士
Advisor:Chih-Sheng Lin, Ph.D.
國 立 交 通 大 學 生 物 科 技 學 院 生物資訊及系統生物研究所 博 士 論 文 A ThesisSubmitted to Institute of Bioinformatics and Systems Biology College of Biological Science and Technology
National Chiao Tung University for the Degree of Ph.D.
in February 2014
Hsinchu, Taiwan, Republic of China
i
Acknowledgement
口試完的當天,真的有鬆口氣的感覺。心裡想著終於不用抱電腦和無窮無盡的期刊 資料坐在咖啡廳內奮鬥了。回想六年的時光都在不知不覺,在電腦螢幕和咖啡苦澀味道 中度過。六年的時光並不算短而博士班學程的結束,得到除了是厚厚的論文和薄薄的一 紙畢業證書以外,更重要的是留在心底的值得去細細品味的經歷與回憶。總結一句:如 人飲水,冷暖自知。 學位口試的完成,要感謝陳銘仁 醫師、吳介信 老師、呂衍達 醫師、以及曲在雯 老 師於百忙之中抽空為我的博士論文進行指導,並給予寶貴的建議。由其是陳銘仁 醫師 不僅是在實驗題材給予我極大的幫忙,也在我做人處事方面給我非常珍貴的指導。而博 士學位的取得,則要感謝在我攻讀博士學位時,盡心指導我的指導教授林志生 老師。 林志生 老師是一位嚴以律己且肯為學生著想的老師,他以嚴格的言教與身教指導實驗 室學生。即使一開始是門外漢如我,老師也肯耐著性子對我諄諄善誘。老師可怕的效率 讓我不禁懷疑他是否平時真有好好休息(PS.半夜寄給他的信總是一下就有回信了),不 論如何老師作事的嚴謹和認真對我後來作事的態度應會起了很大的模範作用。 在攻讀博士學位的期間,因為在職身份的我,並非能一直在實驗室中,還好lab 中 一直都有一群認真聰明的學弟妹們。感謝認真的棠青學弟,他算是我的救星,實驗有問 題問他就沒錯了!再加上默默工作不需人操心的睦元,對於我整個實驗的流程幫助很大。 另外,聰明的千雅、燕秋、意涵、及芳沅等等學弟妹們全力的支持,及謝文郁 醫師跟 我相互打氣支持,我才能如此順利地完成此份論文。 感謝交大有這麼許許多多極富教學熱忱的老師如: 帥氣有型黃憲達 老師、高雅美麗 的王唯菁 老師以及年輕有為的林勇欣 老師等等;每個老師認真上課的態度讓我覺得能 在交大上課真的很幸福。 最後,要感謝的是一直守護我、支持我的父母和妻兒。感謝父母全心全意的支持和 從小以來的教誨,讓我有勇氣繼續走下去;感謝老婆大人支持我取得博士學位的心意, 讓我無窮盡在咖啡廳的日子裡無後顧之憂,令我相當窩心。感謝兩位胖兒子可愛的陪伴,ii 讓我在深夜著手論文時不曾喪失了信心而放棄。造就了此本論文需要感謝的人太多,雖 未能一一點名列出,但是都會記在我的心中。 鄭崑山 謹誌 國立交通大學 生物資訊及系統生物研究所 博士班 中華民國 一百零三年 二月
iii
基質金屬蛋白酶基因多型性及酵素活性在兒童
心室中膈缺損的相關性
研究生:鄭崑山 指導教授:林志生 博士
國 立 交 通 大 學 生 物 科 技 學 院 生物資訊及系統生物研究所 博士班中
文 摘 要
先天性心臟病佔了總出生人口比例約 1–3%,而其中心室中膈缺損(ventricular septal defect ,VSD)又是最為常見的先天性心臟病之一。比率可以佔所有先天性心臟病的 40%,部分病童的中膈缺損會隨著年紀增長而逐漸變小或完全閉合,其餘在臨床追蹤時 則依然保持缺陷。目前有許多因素被認為與VSD 形成有關,其中特定基因的單核苷酸基因多型性(single nucleotide polymorphisms, SNPs)被指出可能是造成 VSD 疾病主要原 因之一。
基質金屬蛋白酶 (matrix metalloproteinases, MMPs)是一類酵素, 其活性依賴於 Zn2+等金屬離子, 參與降解細胞外基質(extracellular matrix, ECM),並精密調控著組織中 ECM 的代謝平衡。MMPs 已被報導在胚胎發育、心肌分化,以及心臟組織的重塑扮演
著重要的角色,文獻探討中可以發現許多心臟疾病也與MMPs 的基因多型性有關。據此,
我們檢測VSD 的病患的 MMPs 基因 SNPs 與其血漿中 MMPs 活性,來探討 MMPs 基因 SNPs 和血液中 MMPs 活性與 VSD 之嚴重程度。
自2010 年 9 月起,我們收集了 95 位 VSD 病童的血液樣品,使用聚合酶連鎖反應-限制酶片段長度多型性(polymerase chain reaction- restriction fragment length
polymorphism, PCR-RFLP)來檢測這些病童的 MMP 基因 SNPs 包括:MMP -2 -735C>T, 以及 MMP-9 -1562C>T、R279Q 及 R574。並使用明膠酶電泳(gelatin zymography)用以測 定其血漿中MMP-2 和 MMP-9 活性。
iv
布比例有統計上的顯著差異性 (p < 0.05)。而檢測的 MMP-2/-9 的不同單核苷酸基因多型
性基因型別中的MMP-2 及 MMP-9 活性並無不同。另一方面,我們依病患超音波檢查
之VSD 直徑/主動脈主動脈直徑 (aortic root)比值分類病童 VSD 程度,將之分成輕微缺 損(VSD/Ao 0.2)、輕度缺損(0.2 < VSD/Ao 0.3)及中度缺損(0.3 < VSD/Ao 0.66) 三組。
明膠酶電泳結果顯示,特別在中度以上缺損病童分組中,皆可測得較高活性之MMP-2 與MMP-9 並顯著高於控制組。而分析結果也指出,隨著 VSD 趨於嚴重時,其 MMP-2 與MMP-9 的活性皆有增加的趨勢。經過一年的追蹤,我們發現 VSD 的病患癒合與其血 液中的MMP-9 活性有顯著相關 (p < 0.01)。 另外我們也研究了其血液中第三型金屬蛋白酶抑制因子(TIMP-3)以及 B 型利鈉肽 (BNP)的濃度和 VSD 病童血液中 MMP-2/-9 活性的關聯性。分析結果發現,TIMP-3 在 MMP-2 -735 C>T 的 CC 以及 MMP-9 R279Q 的 QQ 對偶基因型病人的濃度是較高,並 與其他對偶基因有統計的顯著性差異; 並且 TIMP-3 和 MMP-9 活性、BNP 血中濃度都 有正相關。此外,我們也發現MMP-9 活性、TIMP-3 及 BNP 血中濃度都與 VSD 的嚴 重度呈現正相關。 據本研究之結果,我們推測MMPs 的活性可能對於 VSD 病童其患病與否、及嚴重 程度、或其癒合比率上有一定的角色,因此值得未來進一步探討MMPs 與 VSD 病理機 轉之間的關係。 關鍵詞:基質金屬蛋白酶、金屬蛋白酶抑制因子、心室中膈缺損、單核苷酸基因多型性
v
Genetic variations and circulating levels of matrix metalloproteinases
in the children with ventricular septal defect
Graduate student: Kun-Shan Cheng Advisor: Chih-Sheng Lin, Ph.D.
Institute of Bioinformatics and Systems Biology College of Biological Science and Technology
National Chiao Tung University
Abstract
Congenital heart disease (CHD) account for 1–3% population, and ventricular septal defect (VSD) is the most common form of CHD that accounts for approximately 40% of total patients. Some of children with VSD can have spontaneous closure with aging, but the others don’t. Single nucleotide polymorphism (SNP) in specific gene is considered as one of the main factors of VSD formation.
Matrix metalloproteinases (MMPs) are a family of zinc-dependent endopeptidases involved in breakdown, and physiological homeostasis of extracellular matrix. MMPs play important roles in embryonic development, cardiomyocyte differentiation and causing cardiac tissue remodeling. The literature review shows many cardiac diseases are related to the SNPs of MMPs gene. Therefore, we aim to examine the SNPs of MMPs gene in patients with VSD and their plasma MMPs activities, in order to find out the relationship among gene SNPs, plasma MMPs activities and the severity of VSD.
Since September 2010, 95 children with perimembranous VSD were enrolled in this study and blood samples were collected. The SNPs of MMP-2 (-735C>T) and MMP-9 (-1562C>T, R279Q, and R574P) were determined using polymerase chain reaction followed by restriction fragment length polymorphism (PCR-RFLP) analysis. Plasma MMP-2 and MMP-9 activities were also confirmed by gelatin zymography.
vi
From the results of four SNPs assay in this study, there is a significant difference of MMP-9 R279Q SNP frequency between VSD patients and control group (p < 0.05). There is no difference between MMP-2 or MMP-9 genotypes among all the four polymorphisms. Children with VSD according to the ratio of VSD diameter/diameter of aortic root (Ao), were categorized into different groups: Trivial group (VSD/Ao ratio 0.2), Mild group (0.2 < VSD/Ao 0.3), and Median group (0.3< VSD/Ao 0.66). According to the results of gelatin zymography, there is significant higher MMP-2/-9 activity in each groups compared with control group. After one year follow-up, we also found that there are significant differences of MMP-9 activities between VSD spontaneous closure group and non-closure group (p < 0.01).
The concentrations of plasma tissue inhibitor of metalloproteinase-3 (TIMP-3) and B-type natriuretic peptide (BNP) in VSD children, and the correlations with MMP-2/-9 activities in VSD patients with different severities were also investigated. The data showed that the TIMP-3 concentration is significantly higher in carriers of the CC genotype of MMP-2 -735 C>T and QQ genotype of the MMP-9 R279Q polymorphisms than other genotypes. We also found that TIMP-3 correlated with MMP-9 activity and BNP
concentration. Overall, MMP-9, TIMP-3, and BNP all show a similar positive trend with regard to the severity of VSD.
In conclusion, we found the relation between the severity of VSD and activities of MMPs. Our data might hint that MMPs expression may play a role in the pathogenesis of VSD. It is worth to further investigate the correlation between MMPs and the mechanism of VSD formation or closure.
Keywords: Matrix metalloproteinase (MMP), Tissue inhibitors of metalloproteinase (TIMP), Ventricular septal defect (VSD), Single nucleotide polymorphisms (SNPs)
vii
Contents
Acknowledgment ... i
Chinese Abstract ... iii
English Abstract ... v Contents ... vii List of Tables ... x List of Figures ... xi I. Literature Review ... 1 1-1. Single-nucleotide polymorphisms ... 1
1-2. Ventricular septal defect ... 2
1-3. Matrix metalloproteinase ... 3
1-3-1. Structures and functions of matrix metalloproteinases ... 3
1-3-2. Gelatinase A (MMP-2, Type II collagenase) ... 5
1-3-3. Gelatinase B (MMP-9, Type V collagenase) ... 6
1-3-4. Roles of MMPs in cardiac development ... 6
1-4. Tissue inhibitor of metalloproteinases ... 8
1-4-1. Structures and functions of tissue inhibitor of metalloproteinases ... 8
1-4-2. Tissue inhibitor of metalloproteinase-3 ... 9
1-5. B-type natriuretic peptide ... 10
II. Research Strategy ... 18
III. Materials and Methods ... 20
3-1. Patients and sample collection ... 20
3-2. Patient grouping by VSD diameter/diameter of aortic root (Ao) ratio ... 20
3-3. Genomic DNA extraction ... 21
3-4. Genotyping ... 22
viii
3-4-2. MMP-9 -1562C>T polymorphism ... 22
3-4-3. MMP-9 R279Q polymorphism ... 23
3-4-4. MMP-9 R574P polymorphism ... 23
3-5. Gelatin zymography assay ... 23
3-6. Determination of the TIMP-3 and BNP concentrations by enzyme-linked immunosorbent assay (ELISA) ... 24
3-7. Statistical analysis ... 24
IV. Results ... 27
4-1. The information of patients and Patient grouping by VSD diameter/diameter of aortic root (Ao) ratio ... 27
4-2. Genotype distribution and allele frequency of MMP-2 -735C>T ... 27
4-3. Genotype distribution and allele frequency of MMP-9 -1562C>T, R279Q and R574P ... 28
4-3-1. MMP-9 -1562C>T polymorphism ... 28
4-3-2. MMP-9 R279Q polymorphism ... 29
4-3-3. MMP-9 R574P polymorphism ... 29
4-4. Plasma MMPs activity in the VSD children with different MMP polymorphisms ... 30
4-4-1. MMP-2 activity in gentoypes of -735C>T ... 30
4-4-2. MMP-9 activity in genotypes of -1562C>T, R279Q and R574P ... 30
4-5. Relationship between plasma MMPs activities and VSD severities ... 31
4-5-1. Relationships between MMPs activities and VSD defects ... 32
4-5-2. Relationships between MMPs activities and VSD defects grouping ... 32
4-5-3. MMP-9 activity in different sizes groups ……….. 32
4-6. Correlations between plasma MMP-2, MMP-9 activity, concentrations of TIMP-3 and BNP in VSD patients ... 33
4-6-1. TIMP-3 in different VSD patients and MMPs polymorphisms ... 33
4-6-2. BNP in different VSD patients and MMPs polymorphisms ... 33
4-7. Pearson’s correlation between MMP-2/-9, TIMP-3, and BNP ... 34
ix
V. Discussion ...51
5-1. Gene Polymorphisms of MMP-2/-9 in VSD patients ...52
5-2. Plasma activities of MMP-2/-9 in VSD patients and their relation with spontaneous closure rate of VSD ... 55 5-3. Correlations between plasma MMP-2/-9 activities, concentrations of TIMP-3 and BNP ...57
VI. Conclusions ...61
VII. References ...63
Appendix ...75
Appendix 1. Sketch and diagram of SNPs performed in the study ...75
Appendix 2. Database information of MMP-2 SNPs ...83
x
List of Tables
Table 1-1. Outlines of the information of MMPs and TIMPs SNPs defined and
relationship with particular diseases ...12 Table 3-1. Sequences of the primers used for MMP-2/-9 genotyping by PCR-RFLP ...26 Table 4-1. VSD patients’ age and control group’ age, gender, VSD sizes, and VSD/Ao
ratio ... 35 Table 4-2. Patient grouping by VSD diameter/diameter of aortic root (Ao) ratio ...36 Table 4-3. Genotype distribution and allele frequency of MMP-2 -735C>T
polymorphism ...37 Table 4-4. Genotype distribution and allele frequency of MMP-9 -1562C>T
polymorphism ...38 Table 4-5. Genotype distribution and allele frequency of MMP-9 R279Q
polymorphism ...39 Table 4-6. Genotype distribution and allele frequency of MMP-9 R574P
xi
List of Figures
Figure 1-1. Location of different VSDs ... 14
Figure 1-2. Domain structures of proMMP-2 are introduced here to stand for the general structures of MMP family ... 15
Figure 1-3. Molecular structures of hyaluronan and TIMP-2 ... 16
Figure 1-4. Schematic presentation of human atrial, B-type, and C-type natriuretic peptides ... 17
Figure 2-1. The flowchart of research strategies ... 19
Figure 4-1. Plasma MMP-2 activities in patients of different MMP-2 SNP alleles ... 41
Figure 4-2. Plasma MMP-9 activities in the VSD children of -1562C>T, R279Q and R574P ... 42
Figure 4-3. Echocardiography discerns normal and a patient of VSD ... 43
Figure 4-4. Correlations between circulating MMPs activities and VSD size or VSD/Ao ratio in the children patients ... 44
Figure 4-5. Circulating MMP-2 and MMP-9 activities in the patients with different levels of VSD severity ... 45
Figure 4-6. Plasma levels of MMP-9 (A); BNP (B); and TIMP-3 (C) in VSD patients with different VSD/Ao ratio ... 46
Figure 4-7. Plasma TIMP-3 concentrations in MMP-2/-9 polymorphisms ... 47
Figure 4-8. Plasma BNP concentrations in MMP-2/-9 polymorphisms ... 48
Figure 4-9. Pearson’s correlation between MMP-2/-9, TIMP-3, and BNP ... 49
Figure 4-10. Circulating MMPs activities in the patients received serial echocardiographic follow-up examinations ... 50
Figure 5-1. Pathways of regulating region specific cardiac morphogenesis ... 60
Figure A1. A sketch of human MMP-2 gene and SNP rs2285053 ... 75
Figure A2. Diagram of MMP-2 SNP rs2285053 ... 76
Figure A3. A sketch of human MMP-9 gene and SNP rs3918242 ... 77
Figure A4. Diagram of MMP-9 SNP rs391824 ... 78
Figure A5. A sketch of human MMP-9 gene and SNP rs17576 ... 79
xii
Figure A7. A sketch of human MMP-9 gene and SNP rs2250889 ... 81 Figure A8. Diagram of MMP-9 SNP rs2250889 ... 82
1
I. Literature Review
1-1. Single-nucleotide polymorphisms
Polymorphisms represent natural sequence variants (alleles), which may appear in at least 1% of a population and are considered biologically normal. Approximately 90% of DNA polymorphisms are single-nucleotide polymorphisms (SNPs) due to a single base exchange
(Collins et al., 1998). A SNP represents an alternative nucleotide in a given and defined genetic location. This definition does not include other types of genetic variability like insertions and deletions, and variability in copy number of repeated sequences. They occur one in every 300 nucleotides on average. Although the majority of DNA polymorphisms are probably functionally neutral, a proportion of them can exert allele-specific effects on the regulation of gene expression or function of the coded protein, then cause individual differences in various biological processes or in susceptibility to diseases (Brookes, 1999).
A SNP in which both alleles produce the same polypeptide sequence is called a
synonymous polymorphism (sometimes called a silent mutation); which produce a different polypeptide sequence is called a non-synonymous polymorphism (replacement
polymorphism). Mutations found in known diseases over half of all come from
non-synonymous polymorphisms (Stenson et al., 2009). When gene expression is affected by a SNP, this SNP type is referred to as an eSNP (expression SNP) and might be upstream or downstream from the gene.
Analytical methods to discover novel SNPs and detect known SNPs include DNA sequencing, restriction fragment length polymorphism (RFLP), capillary electrophoresis, mass spectrometry, single-strand conformation polymorphism (SSCP), electrochemical analysis, denaturing HPLC, gel electrophoresis and hybridization analysis (Kwok and Chen,
2
2003). All of the background reference and information of SNPs in this study were obtained from National Center for Biotechnology Information (NCBI)-SNP database
(http://www.ncbi.nlm.nih.gov/snp/).
1-2. Ventricular septal defect
Ventricular septal defect (VSD) is the most common form of congenital heart diseases which account for 40% of the patients having congenital heart diseases (Hoffman, 1995). The defect can be in any portion of the ventricular septum, and the physiologic consequences can range from trivial to severe. Although historically the incidence of VSD is cited as
approximately 1.5 to 3.5 per 1,000 term infants and 4.5 to 7 per 1,000 premature infants (Moe and Guntheroth, 1987), recent studies demonstrated an incidence of VSD in newborns to be 5 to 50 per 1,000 (Tikanoja, 1995). Sometimes, VSD might be not only an isolated cardiac malformation but also an intrinsic component of several complex malformations such as tetralogy of Fallot. However, patients whose cardiac malformation is predominately caused by a VSD will be focus on in this proposal.
There are many systems to classify the different VSD types. One of the systems can classify VSD according to their location, either within the muscular septum (muscular defects) or at its margins. VSD at the margins of the muscular septum can be related to hinge-points of the leaflets of the atrioventricular valves (perimembranous type), those of the arterial valves (juxta-arterial or subarterial type), or both (Fig. 1-1) (Penny and Vick, 2011). Symptoms, natural history and management of VSD depend on size, anatomical characteristics, and patients’ variances.
Rates of spontaneous closure for membranous and muscular VSD in infant and children were 37% and 50%, respectively during a mean follow-up of 12 months (Moe and Guntheroth,
3
1987). The others have no such a fortunate clinical course and their VSD persist. The
development of echocardiography had provided a good tool to observe the natural processes of spontaneous VSD closure (Murphy et al., 1986); Perimembranous defects often close by the development of a saccular pouch or aneurysm derived from tissue from the septal leaflet of the tricuspid valve. Muscular defects appear to close by progressive growth of tissue from the right ventricular side of the circumference of the defect. Neither the detail molecular insight of spontaneous VSD closure nor the study of association between extracellular matrix (ECM) and spontaneous VSD closure has been clarified.
1-3. Matrix metalloproteinase
1-3-1. Structures and functions of matrix metalloproteinases
Matrix metalloproteinases (MMPs) is a family of extracellular zinc-dependent neutral endopeptidases (Lombard et al., 2005), capable of degrading essentially all ECM components including fibrillar and non-fibrillar collagens, fibronectin, laminin and basement membrane glycoproteins (Fedarko et al., 2004). MMPs usually play an important role in ECM
remodeling in physiologic situations, such as embryonal development, tissue regeneration, and wound repair. In addition, there are more and more researchers turn their focus on
relationships of MMP proteins function and different pathological conditions, because of their potent degradative capacities. Some articles indicate the change of nucleotides and the SNPs of MMPs and tissue inhibitors of matrix metalloproteinases (TIMPs) correlated with many different diseases like coronary artery disease (Renko et al., 2004), Kawasaki disease (Ikeda et al., 2008), and cancer (Liu et al., 2011) (Table 1-1).
4
considered as potential clinical markers of disease activity. For example, serum MMP-2 level has been reported to be increased in the patients with liver cirrhosis (El-Gindy et al., 2003), and endometriosis (Huang et al., 2004); Serum MMP-9 level appears to increase in patients with congestive heart failure (Abou-Raya et al., 2004), stroke (Lynch et al., 2004; Reynolds et al., 2003), or myocardial infarction (Renko et al., 2004).
The MMP family currently consists of 28 enzymes with somewhat different capacities. The members are generally divided into si× groups according to either structure or preferred substrates and referred to as interstitial collagenases (MMP-1, -8, and -13), gelatinases (MMP-2 and MMP-9), stromelysins (MMP-3, -10, -11, and -12), matrilysins (MMP-7 and MMP-26), membrane-type MMPs (MMP-14, -15, -16, -17, -24, and -25) and others (Bode, 2003; Maskos and Bode, 2003; Visse and Nagase, 2003). Although the groupings of MMPs are based on their abilities to degrade various proteins of the ECM, they also play other important roles such as the activation of cell surface receptors and chemokines (Stefanidakis and Koivunen, 2006). In addition, MMP-2 has proteolytic activity to specific targets within the cell to cause acute, reversible contractile dysfunction in cardiac disease (Schulz, 2007).
The basic structures of MMPs can be approximately divided into three structurally well-preserved domain motifs, including a catalytic domain, an N-terminal domain and a C-terminal domain (Fig. 1-2A). Zinc-dependent catalytic domain of MMP is similar with subtle structural differences among the substrate specific groups (Nagase and Woessner, 1999). The N-terminal domain (propeptide domain) contains a unique PRCG(V/N)PD sequence in which the cysteine residue interacts with the catalytic zinc atom in the active site, prohibiting activities of the MMPs. Thus, the interaction has to be disrupted to “open” the cysteine switch in the process of MMPs activation (Van Wart and Birkedal-Hansen, 1990), which is a critical step that leads to ECM breakdown (Carmeli et al., 2004). The C-terminal hemopexin domain of metalloproteinases has a four-bladed propeller structure and contributes to substrate
5
specificity (Fig. 1-2B) (Wallon and Overall, 1997). In membrane-type MMPs, the hemopexin domain contains a transmembrane domain for anchoring the protein in the membrane; besides, the hemopexin domain in MMP-2 also has a function in the activation of the enzyme
(Morgunova et al., 1999).
The regulations of MMPs occur at many levels, including transcription (the major one), post-transcriptional modulation of mRNA stability, secretion, localization, zymogen
(proenzyme) activation and inhibition of activity by natural inhibitors of MMPs, TIMPs
(Brew et al., 2000).
Cell migration and tissue remodeling are two fundamental processes of embryogenesis. One group of MMPs in particular, gelatinases, are implicated in tissue remodeling and in enabling cell migration and invasiveness. Two forms of gelatinases have been extensively studied in cells, and both can degrade type IV collagen and interstitial collagens as well as many other ECM molecules.
1-3-2. Gelatinase A (MMP-2, Type II collagenase)
In 1978, Sellers et al. was the first team to separate a gelatinase activity from collagenase and stromelysin in culture medium from rabbit bone (Sellers et al., 1978). Next year, a similar enzyme, acting on basement membrane type IV collagen, was reported by Liotta et al (1979).
Gelatinase was purified from human skin, mouse tumor cells, rabbit bone, and human gingiva. Human and mouse MMP-2 are secreted as 72 kDa proenzymes of 631 and 662 amino acids, respectively. The removal of the pro-domain can be initiated by serine proteases such as thrombin and activated protein C. The resulting mature and active enzyme consists of a catalytic domain which is interrupted by three contiguous fibronectin type II-like domains, participating in binding to the gelatin substrates of the enzyme, and a C-terminal,
6
feature of gelatinase. The complete sequence of the human MMP-2 was published by Collier et al. (2001)
1-3-3. Gelatinase B (MMP-9, Type V collagenase)
Krane (1994) detected a gelatinase activity in rheumatoid synovial fluid. Sopata and Wize (1979) described a gelatinase from human polymorphonuclear leukocytes . Purification was achieved in 1983 and sequencing of the cDNA in 1989. Human neutrophil MMP-9 commonly occurs as a complex with lipocalin, and this complex protects this extracellular matrix remodeling enzyme from autodegradation (Fernandez et al., 2005).
MMP-9 is secreted as a 92 kDa zymogen. Cleavage of pro-MMP-9 at near residue 87 results in activation of enzyme with a mass of approximately 82 kDa. Pro-MMP-9 is secreted by monocytes, macrophages, neutrophils, keratinocytes, fibroblasts, osteoclasts, chondrocytes, skeletal muscle satellite cells, endothelial cells, and various tumor cells. Pro-MMP-9 can be activated by MMP-3 or by certain bacterial proteinases. MMP-9 has prodomain, three
fibronectin type II motifs in catalytic domain, and a hemopexin-like domain just like MMP-2. MMP-9 is inhibited by α2-macroglobulin or by TIMP-1. An interesting phenomenon, still not fully understood, is the binding of TIMP-1 to proMMP-9 to form a complex before secretion out of the cell (Roderfeld et al., 2007).
1-3-4. Roles of MMPs in cardiac development
MMP-2 is the earliest MMP known to be expressed during heart development. The process of cardiac looping converts the single, straight tubular heart into a S-shaped tube and transforms the primitive heart chambers into their adult anatomical positions before cardiac septation is complete. During the process of building a single heart tube, MMP-2 is expressed in the endocardium, early differentiating cardiomyocytes, and dorsal mesocardium but is soon lost within the myocardium (Cai et al., 2000).
7
Cell proliferation is more pronounced within the left splanchnic mesoderm and left dorsal mesocardium. Blocking MMP-2 activity not only disrupts this asymmetric pattern of proliferation, it also randomizes the direction of cardiac looping and increases the incidence of dextrocardia (i.e., right-sided heart). Blocking MMP-2 activity also prevents midline fusion of the primitive heart tubes leading to cardiac bifida (Linask et al., 2005).
Some experiments support the postulate that MMP-2 has an important functional role in early cardiogenesis, cardiac cushion migration, and remodeling of the direction of cardiac looping (Cai et al., 2000; Linask et al., 2005).
Defects in cardiogenesis during the first three weeks of gestation are usually fatal;
however, embryo could survive with anomalous developmental events occurring in later stage. These anomalies often manifest as great vessels or cardiac septal defects in neonates. The septation of the atria and ventricles and division of the cardiac outflow tract into the aorta and pulmonary artery requires the migration, proliferation, and differentiation of two distinct mesenchymal populations, i.e. endocardial-derived cushion cells and cardiac neural crest (NC) cells (Creazzo et al., 1998).
MMPs have been implicated in regulating epithelial-to-mesenchymal transitions (EMTs) which responsible for forming both populations of cells. MMP-2 is expressed by endocardial cells prior to and during the EMT of the endocardium and the neural crest cells detach from the neural epithelium. Perturbed MMP-2 expressions in these studies disturb the migration and tissue remodeling of cushion cells and NC cells. Both of them are important and
participate in formation of atrioventricular and outflow tract regions of the developing heart (Alexander et al., 1997; Cai et al., 2000)..
Hyaluronan is abundant in the ECM of the pre-mesenchymal heart (Fig. 1-3A). Hyaluronan is an essential mediator of cell migration and invasion for proper heart
8
development. Hyaluronan relates to regulate the expression of multiple MMPs in several cell types and the transformation of endothelial cells to mesenchymal cells. Therefore, MMPs might play pivotal roles and involve in the endocardial and cardiac septation, and cardiac developing (Camenisch et al., 2002).
1-4. Tissue inhibitor of metalloproteinases
1-4-1. Structures and functions of tissue inhibitor of metalloproteinases
As the variable and complex functions of MMPs, one of the important mechanism for the regulation of the activity of MMPs is via binding to a family of homologous proteins referred to as the tissue inhibitors of metalloproteinases (TIMP-1 toTIMP-4). TIMPs can form 1:1 enzyme-inhibitor complexes to inhibit matrixins, and each member of TIMPs can inhibit almost every member of the MMP family. The two-domain TIMPs are of relatively small size, yet have been found to exhibit several biochemical and physiological/biological functions, including inhibition of active MMPs, proMMP activation, cell growth promotion, matrix binding, inhibition of angiogenesis and the induction of apoptosis (Woessner, 2001).The TIMPs have the shape of an elongated contiguous wedge consisting of an N-terminal segment, an all-β-structure left-hand part, an all-helical center, and a β-turn structure to the right. The N- and the C-terminal halves of the polypeptide chain form two opposing subdomains, each domain being stabilized by three disulfide bonds (Bode et al., 1999; Brew et al., 2000) (Fig. 1-3B).
The N-terminal subdomain exhibits a so-called OB-fold, known for a number of oligosaccharide/oligonucleotide binding proteins. This region consists of a five-stranded β-pleated sheet of Greek-key topology rolled into a closed β-barrel of elliptical cross-section.
9
The narrower opening of this barrel is bounded by the sB-sC loop, while its wider exit is (in contrast to other OB-fold proteins) covered by an extended segment connecting strands sC and sD, designated as “connector.” After leaving the barrel, the polypeptide chain passes two helices, forms a two-stranded β-sheet, runs through a wide multiple-turn loop, and terminates in a β-hairpin sheet. The last C-terminal residues do not exhibit a defined conformation and presumably form a flexible tail on the TIMP surface (Fig. 1-3B)
Although the TIMPs are similar to each other to the extent of 35-40% amino acids identity, these key similarities suggest a significant structural conservation. For example, all TIMPs contain 12 cysteine residues at conserved locations, and in the case of TIMP-1, it has been shown that these participate in the formation of six intrachain disulfide bonds and then stabilize the whole structure. The N-terminal halves of the TIMPs share the most amino acid identity, suggesting that this region may underlie the common property of MMP binding and inhibition; It is possible that the more variant C-terminal half of the TIMPs may subserve distinctive properties of each TIMP and in the case of TIMP-3 the C-terminal half contains the ECM-binding domain (Anand-Apte et al., 1996).
1-4-2. Tissue inhibitor of metalloproteinase-3
In 1983, TIMP-3 was first isolated as a transiently expressed 21 kDa protein in the ECM of transforming chick fibroblast cultures (Blenis and Hawkes, 1983). Peptide sequencing subsequently demonstrated homology to the TIMPs and its identity as a novel member of the family was confirmed by cloning of the chicken cDNA and demonstration of MMP inhibition
(Staskus et al., 1991). Subsequently, cDNAs of human TIMP-3 had been isolated (Apte et al., 1994).
TIMP-3 is an effective inhibitor of MMPS, proved by gelatin reverse zymography assays
10
These assays have shown that TIMP-3 and TIMP-1 are equipotent in their ability to inhibit MMP-1, MMP-2, MMP-3, and MMP-9. Otherwise, TIMP-3 has poor aqueous solubility and a specific localization in the ECM in contrast to TIMP-1,2,and 4 (Pavloff et al., 1992). The ECM ligand of TIMP-3 has been suggested being the hyaluronic acid (Yang and Hawkes, 1992).The affinity of TIMP-3 for ECM and its expression in a number of epithelia led some researchers to propose that TIMP-3 might be a component of basement membrane.
TIMP-3 also has unique function of playing an role in inhibiting the endothelial cell responses to physiologically relevant antigenic agents such as VEGF and bFGF, inhibiting migration and preventing invasion and tube formation by endothelial cells in a collagen matrix (Qi et al., 2003). TIMP-3 is the only TIMP known to be related to a disease: mutation of certain cysteine residues to serine results in early blindness, a condition known as Sorsby’s fundus dystrophy (Felbor et al., 1995).
1-5. B-type natriuretic peptide
Over the recent decades, one group of neurohormonal markers, including atrial
natriuretic peptide (ANP), N-terminal proBNP (NT-proBNP), and B-type natriuretic peptide (BNP), has generated much interest in the evaluation and management of cardiovascular disease. C-type natriuretic peptide is produced by endothelial cells and macrophages, whereas ANP and BNP are derived from cardiac muscle. ANP and BNP act through the natriuretic A-type receptor. Each of the natriuretic peptides is cleared by the natriuretic C-type receptor and degraded by neutral endopeptidases. ANP is produced from a 126 amino acid precursor (pro-ANP) whilst BNP is synthesized as a 134 amino acid pre-pro form that is secreted as a 108 amino acid pro-BNP precursor. Proteolytic cleavage of these pro-natriuretic peptide precursors produces active ANP and BNP (Fig 1-4) as well as other fragments. The
11
activity, but the cleavage products of pro-ANP are active (Goetze, 2012).
Both the ANP and BNP have beneficial compensatory actions including vasodilation, natriuresis, growth suppression and inhibition of both the sympathetic nervous system and the renin-angiotensin-aldosterone axis (Ruskoaho, 2003). ANP is derived from atrial as well as ventricular myocytes, and significant amounts can be released from cytoplasmic granules in response to relatively minor stimuli; on the other hand, nearly all circulating BNP is derived from ventricular myocytes, and its pulsatile synthesis and release is predominantly in
response to ventricular volume, pressure and wall tension (Sullivan et al., 2005). Besides the above advantage, BNP also has longer half-life than ANP and B-type natriuretic peptide (BNP) has better diagnostic, prognostic, and therapeutic values in heart failure and other heart
diseases (Ruskoaho, 2003).
As it is successfully applied in adult heart diseases, BNP has attracted increasing interests as a biomarker of VSD in the field of pediatric cardiology (Ozhan et al., 2007; Rademaker and Richards, 2005). Some researchers have stated that MMP-9 serves as potential biomarker of heart remodeling (Halade et al., 2013; Lopez et al., 2010). These findings also drew our attention to the correlations among levels of plasma BNP in VSD patients.
12
Table 1-1. Outlines of the information of MMPs and TIMPs SNPs defined and relationship with particular diseases Targeting gene (position, dbSNP
rs# cluster ID) Genotypes Relationship with particular diseases References
MMP-1 (-1607, rs1799750)
G GG
Patients who had the MMP-1 2G/2G genotype had a 1.71-fold increased risk of lung cancer (95% confidence interval,
1.22-fold to 2.41-fold increased risk) compared with patients who had the 1G/1G genotype.
(Liu et al., 2011)
MMP-2 (-1059, rs17859821)
GG AG AA
MMP-2 rs17859821 A allele carriers had lower all cause death rate, cardiac death rate and MACE rate than did GG genotype carriers (OR = 0.655, 0.580, 0.705; p = 0.030, 0.008, 0.011). (Hua et al., 2009b) MMP-13 (-77, rs2252070) AA AG GG
The allele and genotype frequencies of MMP-13-77A>G showed significant differences between Kawasaki disease patients with coronary artery lesions and without coronary artery lesions (p = 0.00989 and p = 0.00551, respectively).
(Ikeda et al., 2008)
MMP-9 (-1562, rs3918242)
CC CT TT
The presence of MMP-9 1562C>T allele was found to be associated with early-onset coronary artery disease (OR = 3.2,
p = 0.001). The ECAD patients with MMP-9 1562C>T allele
had higher MMP-9 activity (p = 0.001).
13 MMP-9 (exon 10, rs2250889) CC CG GG
There are significant differences in the genotype distributions for rs2250889 between diffferant groups, suggest that the G allele of MMP-9 polymorphism rs2250889 is overrepresented in patients with histologically confirmed Giant cell arteritis.
(Rodriguez-Pla et al., 2008)
MMP-9 (-1562, rs3918242)
CC CT TT
The -1562C>T polymorphism of MMP-9 gene is significantly associated with atrial fibrillation risk in Chinese Han patients with hypertensive heart disease. The -1562T allele which is associated with increased expression of MMP-9 might be a genetic risk for the development of AF in this cohort.
(Gai et al., 2009)
MMP-9 (exon 6, rs17576)
GG AG AA
The MMP-9 rs17576 genotype AG and GG appeared to be significant ‘at-risk’ genotypes for Pelvic organ prolapse (OR: 5.41, 95% CI: 1.17– 25.04, p = 0.031; OR: 5.77, 95% CI: 1.29–25.86, p = 0.0219).
A.
A.
18
II. Research Strategy
Many researches documented the well-characterized MMP family intimately involved in normal tissue growth and ECM remodeling. Heart morphogenesis involves the characteristic mechanisms of cell proliferation, cell rearrangements, cell to cell, cell to matrix interactions, and tissue remodeling. MMPs family might regulate the coordination of these important processes, although little is known about the production, secretion, and clearance of these important proteinases throughout normal growth and development in human heart. VSD is the most popular and important congenital heart disease in pediatric cardiologic field.
Morphogenesis and tissue growth, remodeling, and repair are sentinel features of childhood and adolescence. The hypothesis derived from literature research, in this study is that the regulation of MMPs and TIMP in ECM might participate the mechanism of VSD occurrence, and MMPs and TIMP might have important roles in mediating events associated with heart remodeling and recovering. To the best of our knowledge, this is the first study to ascertain the relationships between MMPs family and VSD. The aim of this study is investigating the association among different polymorphic genotypes of MMP-2, and MMP-9 with VSD occurrence. VSD patients were followed up for one year to figure out if these genomic
polymorphisms related to the VSD spontaneous closure or not. Second, the VSD patients also were grouped according to different severities and tried to delineate the interaction of plasma MMP-2, MMP-9 and TIMP-3 activities involved in the occurrence of different VSD severities. Third, BNP was analyzed, which is the well-known cardiac biomarker, and was tested its relationship with MMP-2, MMP-9, and TIMP-3. This conducted study allowed us to realize the molecular interaction between ECM and congenital VSD, the most important pediatric cardiological problem. We also expected that this study could give us the insight of the role of MMPs and TIMPs in the heart morphological remodeling. The flowchart of reach strategies was present in Fig. 2-1.
20
III. Materials and Methods
3-1. Patients and sample collection
A total of 95 children (age range: 4.23 ± 2.51 years old) with perimembranous VSD were recruited from the outpatient base of the Taipei and Hsinchu Mackay Memorial Hospitals. There were 120 healthy individuals (age range: 5.53 ± 2.87 years old) were enrolled for gene polymorphism analysis. Age and gender distribution had no statistical difference between patient and control group. The study was performed with the approval of the Institutional Review Board of Mackay Memorial Hospital. Informed consent was obtained from the parents of all patients. The defects of all patients enrolled in this study were documented by 2-D echocardiography using a Phillips HP 4500 echocardiography machine with 5-8-MHz probe. Each patient received twice echocardiographys by two pediatric cardiologists for preventing interobserver variability and mean values of VSD sizes and aortic root diameters of patients were extracted. All the patients with defects enrolled in this study were
documented by 2-D echocardiography. The plasma of 12 healthy children donated blood samples as a control group in the MMPs activity assays.
Blood samples were collected by needle aspiration from the antecubital vein. Plasma was isolated by centrifuge 3,000 × g for 10 min, and stored at −80°C for further zymography assay.
3-2. Patient grouping by VSD diameter/diameter of aortic root
(Ao) ratio
Clinically, there is a wide range of parameters of a child’s heart that depend on the
patient’s body size and age. Therefore, the VSD size can be normalized by the aortic root (Ao) diameter and the VSD/Ao ratio usually correlated with patients’ clinical severities better than
21
VSD sizes did (Eroglu et al., 2003; Hornberger et al., 1989).Usually, VSD can be classified into three defect sizes: large, median and small sizes. The large defect is defined when the size is larger than two-thirds of the Ao; the median size is between two-thirds and one-third of the Ao; and small if the size is less than one-third of the Ao.
In our experimental design, we followed up the VSD patients regularly but very large VSD patients with poor medical control mandatorily undergo surgery. Therefore, patients in our study were exclusively those without extremely large defects. According to the definition of VSD/ Ao ratio, the patients were classified into three groups: VSD/Ao ratio ≦ 0.2 as Trivial group; 0.2 < VSD/Ao ≦ 0.3 as Small group; and 0.3 < VSD/Ao was considered as Median group.
3-3. Genomic DNA extraction
Genomic DNA for PCR-RFLP genotyping was isolated from the blood of 95 children by using the genomic DNA Mini kit. Up to 200 μL of whole blood was added with 30 μL of proteinase K (10 mg/mL) and mix briefly. After that, the mixture was incubated at 60°C for 15 min. Following then, 200 μL GB buffer was added and mixed by shaking vigorously, and incubated the mixture in a 60°C water bath for 15 min. A volume of 200 μL absolute ethanol was added to the sample lysate and immediately mixed by shaking vigorously. And then, the entire mixture (including any precipitate) was transferred to the GD Column and followed with centrifugation (at 14,000 × g for 5 min). The collections were added with 400 μL of W1 Buffer to the GD Column and centrifuged at 14,000 × g for 1 min. Then added 600 μL of Wash Buffer (ethanol added) to the GD Column and centrifuged at 14,000 × g for 1 min. The supernatant was displaced into new collection tube and centrifuged again at 14,000 × g for 3 min to dry the column matrix. Finally, 30 μL of pre-heated elution buffer (10 mM Tris-HCl,
22
pH 8.5) was added to the matrix and last for 5 min; after that, centrifuged at 14,000 × g for 1 min to elute the purified DNA. The maximum yield was up to 50 μg. Purified genomic DNA was stored in the −80°C.
3-4. Genotyping
3-4-1. MMP-2 -735C>T polymorphism
To analyze the -735C>T polymorphism, we amplified a region of the MMP-2 gene with the primers pair were shown in Table 3-1. The target sequence was amplified in a 50 µL reaction volume containing 1 μL of genomic DNA, 1 µL each primer (10 µM), 5 µL of 10X PCR buffer, 4 µL of 2.5 mM dNTPs, 0.5 µL of Taq Polymerase (Promega, Madison, WI, USA), and 37.5 µL distilled water. The PCR started with an initial activation step of 94°C for 2 min followed by 40 cycles of denaturation at 94°C for 30 sec, annealing at 60°C for 30 sec, and extension at 72°C for 30 sec, and a final step at 72°C for 7 min. The 297-bp fragment was purified with a purification kit (Invitrogen, Grand Island, NY, USA). Purified PCR products were then digested with Sau 96 I (New England BioLabs, Ipswich, MA, USA) at 37°C overnight and separated on a 2% agarose gel. Sau96 I does not digest the T allele but generates 202-bp and 95-bp fragments for the C allele.
3-4-2. MMP-9 -1562C>T polymorphism
The primers used to detect this polymorphism were shown in Table 3-1, which generated a 435-bp fragment containing the -1562C>T site in a 50 µL reaction volume. The annealing temperature was also shown in this table. The PCR product was then digested with Sph I (New England BioLabs) at 37°C overnight and separated on a 2% agarose gel. Sph I does not digest the C allele but generates 247-bp and 188-bp fragments for the T allele.
23
3-4-3. MMP-9 R279Q polymorphism
The primers used to detect this polymorphism were sense 5’-GGT GGG TGC TTC CTT TAA CA-3’ and antisense 5’ -GCG TTA GAG ACG TTG GAA CC-3’ (Ikeda et al., 2008), which generated a 467-bp fragment containing the R279Q site in a 50 µL reaction volume, and the annealing temperature was shown in Table 3-1. The PCR product was then digested with Sma I (New England BioLabs) at 25°C overnight and separated on a 2% agarose gel. Sma I does not digest the A allele but generates 296-bp and 171-bp fragments for the G allele.
3-4-4. MMP-9 R574P polymorphism
The primers used to detect this polymorphism were shown in Table 3-1, which
generated a 246-bp fragment containing the R574P site in a 50 µL reaction volume, and the annealing temperature was also shown in this table. The PCR product was then digested with Nla IV (New England BioLabs) at 25°C overnight and separated on a 2% agarose gel. Three genotypes could be distinguished: GG (246-bp), GC (246, 182 and 65-bp), and CC (182 and 65-bp).
3-5. Gelatin zymography assay
The activities of MMP-2 and MMP-9 in the plasma were measured by gelatin gel zymography as previously had described (Chen et al., 2008). Plasma was mixed with 2 X zymography sample buffer [0.125 M Tris-HCl, pH 6.8, 20% (v/v) glycerol, 4% (w/v) SDS, 0.005% bromophenol blue] incubated for 10 min at room temperature, and then loaded into SDS-PAGE which was performed in 7% acrylamide gels containing 0.1% (w/v) gelatin (Sigma-Aldrich, St. Louis, MO, USA). After electrophoresis under power supply of 100 V,
24
the gel was washed twice for 30 min in zymography renaturing buffer (2.5% Triton X-100) with gentle shake at room temperature to remove SDS, then incubated 18 h at 37°C in reaction buffer (50 mM Tris-HCl, pH 7.4, 200 mM NaCl, and 5 mM CaCl2). The gels were
then stained with coomassie blue for 30 min prior to destain with destain buffer (50%
methanol, 10% acetic acid, and 40% ddH2O). The presence of enzyme activity was evident by
clear or unstained zones, indicating the action of the enzyme on the gelatin substrate
(Stawowy et al., 2004). Gelatinase activity in the gel slab was quantified by Scion Image software (NIH, Bethesda, MD, USA), which quantifies the area of bands hydrolyzed by gelatinase. A MMP-2 or MMP-9 positive control (Chemicon, Temecula, CA, USA) was contained in each gel as a standard intensity value to normalize intensity of samples.
3-6. Determination of the TIMP-3 and BNP concentrations by
enzyme-linked immunosorbent assay (ELISA)
TIMP-3 concentrations were measured by using an ELISA kit (cat# MTM100 R&D Systems, Minneapolis, MN, USA). According to the manufacturer’s instructions, 100 μg of protein per sample was used. The plasma BNP concentration was determined by a
commercially available immunoassay kit (Shionoria BNP assay kit; Shionogi Ltd, Osaka, Japan). Results obtained via spectrophotometry were compared against serial dilutions of known concentrations of the respective standards.
3-7. Statistical analysis
The levels of various markers in the VSD samples are expressed as the mean ± standard deviation (SD) or shown as the median (interquartile range; IQR). Data were analyzed by using SPSS for Windows (SPSS statistics package, Chicago, IL). All polymorphisms were
25
tested separately for confirmation with Hardy-Weinberg expectations in both groups.
Differences between VSD patients and healthy control children were examined with the χ2 test for categorical variables. To compare the quantitative MMP activities between two groups, Student’s t-test was used for presumably normally distributed variables. One-way analysis of variance (ANOVA) method was used to test the differences in the MMPs enzyme activities among groups when the group numbers were more than two. A p value less than 0.05 was considered significant.
26
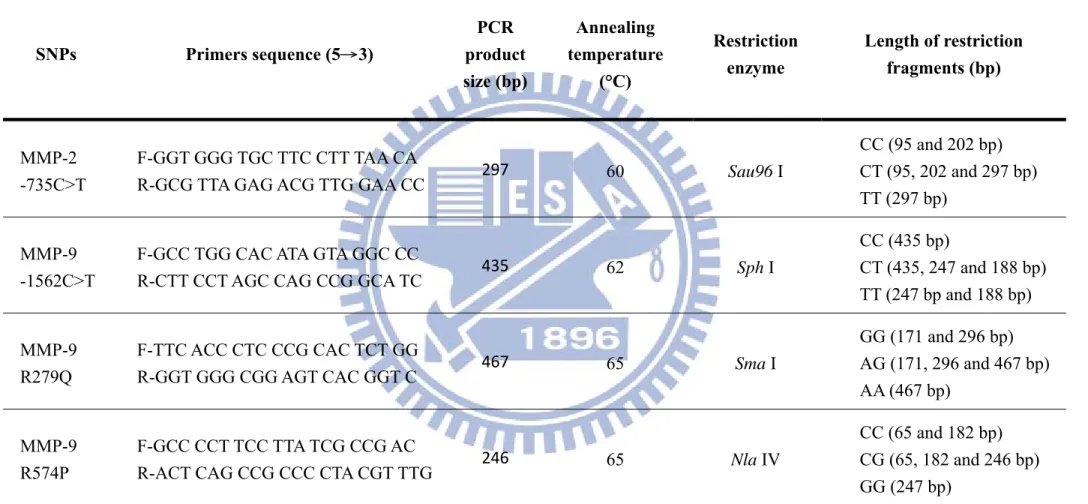
Table 3-1. Sequences of the primers used for MMP-2/-9 genotyping by PCR-RFLP
SNPs Primers sequence (5→3) PCR product size (bp) Annealing temperature (°C) Restriction enzyme Length of restriction fragments (bp) MMP-2 -735C>T F-GGT GGG TGC TTC CTT TAA CA
R-GCG TTA GAG ACG TTG GAA CC 297 60 Sau96 I
CC (95 and 202 bp) CT (95, 202 and 297 bp) TT (297 bp)
MMP-9 -1562C>T
F-GCC TGG CAC ATA GTA GGC CC
R-CTT CCT AGC CAG CCG GCA TC 435 62 Sph I
CC (435 bp) CT (435, 247 and 188 bp) TT (247 bp and 188 bp) MMP-9 R279Q F-TTC ACC CTC CCG CAC TCT GG
R-GGT GGG CGG AGT CAC GGT C 467 65 Sma I
GG (171 and 296 bp) AG (171, 296 and 467 bp) AA (467 bp) MMP-9 R574P F-GCC CCT TCC TTA TCG CCG AC
R-ACT CAG CCG CCC CTA CGT TTG 246 65 Nla IV
CC (65 and 182 bp) CG (65, 182 and 246 bp) GG (247 bp)
27
IV. Results
4-1. The information of patients and Patient grouping by VSD
diameter/diameter of aortic root (Ao) ratio
Age and gender distribution of the 95 patients and 120 control children having MMP-2/-9 genetic polymorphisms analysis in this study were presented in Table 2-1. The mean age of all patients is 4.23 ± 2.51 (mean ± SD) years old, with no significant difference from the control group (5.53 ± 2.87 years) (Table 4-1).
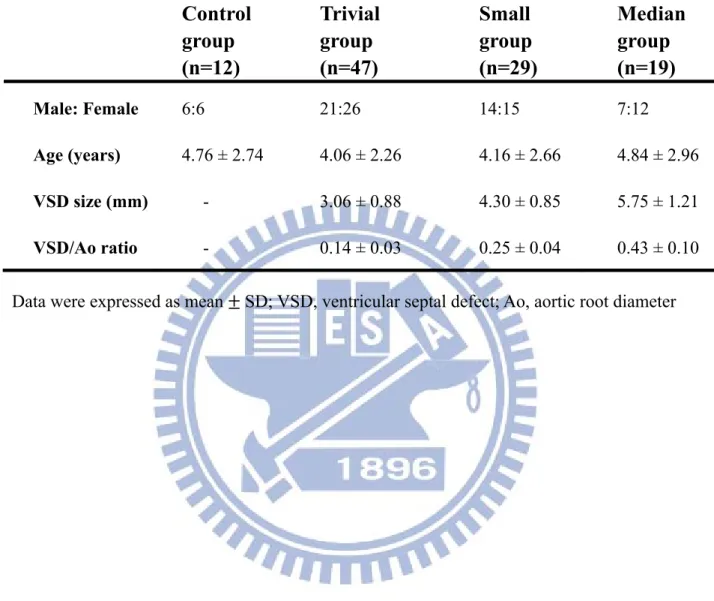
All patients enrolled in this study were examined by two cardiologists using 2-D and Doppler echocardiography and got the average. The diameter of VSD was defined as VSD size, and ratio of VSD diameter and diameter of aortic root (Ao) was measured. According to the VSD/ Ao ratio, patients were classified into three groups, Trivial (VSD/Ao ratio = 0.14 ± 0.03), Small (0.25 ± 0.04) and Median (0.43 ± 0.10). Twelve control children having plasma MMP-2/-9 activities analysis and each subgroup (Trivial, Small and Median) are comprised of 12, 47, 29 and 19 patients, respectively (Table 4-2).
4-2. Genotype distribution and allele frequency of MMP-2
-735C>T
The genomic organization of the human MMP-2 gene and localization of our target SNPs were depicted in Fig. A1A. In this study, -735C>T polymorphism of MMP-2 was analyzed by PCR-RFLP. The sequence of the human MMP-2 gene and the position of the PCR primers that were used to detect MMP-2 -735C>T was shown in Fig. A1B. Restriction enzyme digestion using the restriction enzyme Sau96 I generated three different length bands among different genotypes (Fig. A1A). We also used direct sequencing to detect MMP-2 polymorphism in some patients, and there are three genotypes of MMP-2 -735C>T shown in
28
Fig. A2B.
The genotypes and allele frequencies of MMP-2 -735C>T were recorded in Table 4-3. The frequencies of MMP-2 -735C>T genotypes and allele frequency in the VSD patients and control subjects should not conform to the Hardy-Weinberg equilibrium if there exist
differences. As showed in Table 4-3, the allele frequencies of C and T allele are 66.3% and 33.7%; however, the results were similar to control group (p = 0.919). The genotype frequencies of MMP-2 -735C>T were CC: 47.4%, CT 37.9%, TT 14.7%. No significant difference was found in the patient and control groups (p = 0.143).
4-3. Genotype distribution and allele frequency of MMP-9
-1562C>T, R279Q and R574P
Localizations of the studied SNPs of human MMP-9 gene were shown in Fig. A3, A5, A7. MMP-9 gene contains 13 exons. MMP-9 -1562C>T, R279Q and R574P polymorphisms of MMP-9 are located on promoter region, exon 6 and exon 10 of MMP-9 gene, respectively. PCR-restriction fragment length polymorphism analysis was used for the genotyping of the MMP-9 -1562C>T, R279Q and R574P polymorphism in this study
4-3-1. MMP-9 -1562C>T polymorphism
The sequence of the human MMP-9 gene and positions of the PCR primers that were used to detect MMP-9 -1562C>T was shown in Fig. A3B. PCR products were digested by restriction enzyme Sph I and generated two different length bands between different genotypes (Fig. A4A). There are two genotypes of direct sequencing maps for MMP-9 -1562C>T shown in Fig. A4B.
The distribution of the MMP-9 -1562C>T genotypes were described in Table 4-4. Only 2 genotypes, CC and CT, of the MMP-9 polymorphism -1562C>T were detected in both the
29
VSD and control groups. Genotype distribution and allele frequency were analyzed by using the chi-square test and the results showed that there was no significant difference in the genotypes and allele frequencies between the VSD and control groups.
4-3-2. MMP-9 R279Q polymorphism
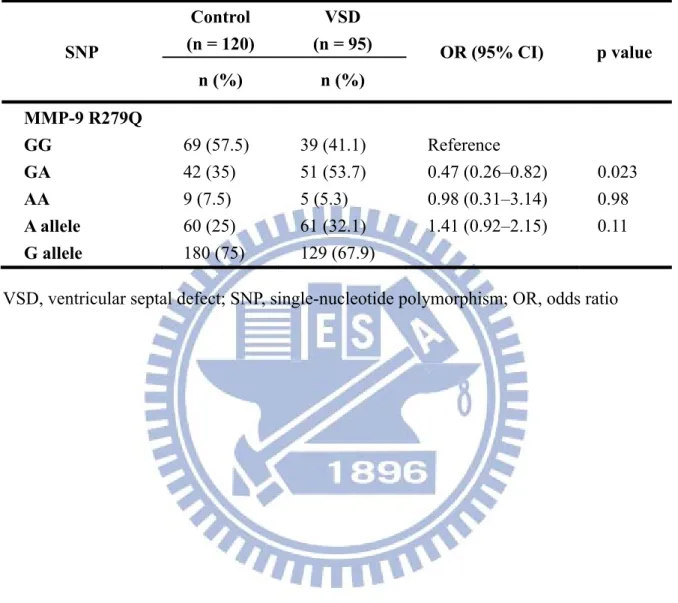
The sequence of the human MMP-9 gene and positions of the PCR primers for MMP-9 R279Q were shown in Fig. A5B. PCR products were digested by restriction enzyme Sma I and generated three different bands (AA genotype: 467-bp, AG genotype: 467-bp, 296-bp, and 171-bp, GG genotype: 296-bp and 171-bp) (Fig. A6A). Fig. A6B shows the direct sequencing map of three genotypes of MMP-9 R279Q.
The genotypes and allele frequencies of MMP-9 R279Q between VSD and control group were summarized in Table 4-5. With regard to the genotype distribution and allele frequency of the MMP-9 polymorphism R279Q, the genotypes G/G, G/A, and A/A were detected in 41.1%, 53.7%, and 5.3% of subjects in the VSD group, respectively, and in 57.5%, 35%, and 7.5% of subjects in the control group, respectively. The genotype distribution of the MMP-9 polymorphism R279Q in the VSD group was significantly different compared to the control group (p =0.023); however, there was no significant difference in the allele frequency between the VSD (G: 67.9% and A: 32.1%) and the control (G: 75% and A: 25%) groups.
4-3-3. MMP-9 R574P polymorphism
The sequence of the human MMP-9 gene and positions of the PCR primers that were used to detect MMP-9 R574P are shown in Fig. A7B. PCR products were digested by restriction enzyme Nla IV and generated three different length bands among different
genotypes (Fig. A8A). There are three genotypes of MMP-9 R574Q in the direct sequencing maps (Fig. A8B).
30
results show that the homozygous C/C, heterozygous C/G and homozygous G/G genotypes of MMP-9 R574P are 52.5%, 38.3% and 9.2% in the control group. Otherwise, the patient groups are 48.4 %, 47.4 % and 4.2 %, respectively. There are no significant differences in genotypes and allele frequencies between two conditions.
4-4. Plasma MMPs activity in the VSD children with different
MMP polymorphisms
In order to investigate the differences of MMPs activities in VSD patients of different MMP polymorphisms, we used the gelatin zymography to detect MMP-2 and MMP-9 activity in plasma.
4-4-1. MMP-2 activity in genotypes of -735C>T
The activities of MMP-2 enzyme in the different genotypes of MMP-2 -735C>T
polymorphism were compared with the Control group in Fig. 4-1 [CC, CT, TT genotypes vs. Control group: 808 (642–980), 806(617–996), 869 (581–1044) ng/mL vs. 554 (407–689) ng/mL]. CC; CT; and TT genotypes of MMP-2 -735 C>T alleles contain 45, 36, and 14 samples, respectively. We used standard MMP-2 as positive control (Chemicon, Temecula, CA, USA) to calibrate MMP-2 activity of each group. MMP-2 activity has no significant difference in three genotypes of MMP-2 -735 C>T polymorphism. All three genotypes have higher activities level than control group. (p = 0.01)
4-4-2. MMP-9 activity in genotypes of -1562C>T, R279Q and R574P
We also investigated the MMP-9 activities of VSD patients in different MMP-9 SNPs, including MMP-9 -1562C>T, MMP-9 R279Q and MMP-9 R574P.
The activities of MMP-9 enzyme in the different genotypes of MMP-9 -1562 C>T polymorphism were compared with the Control group in Fig. 4-2A [CC, CT genotypes vs.
31
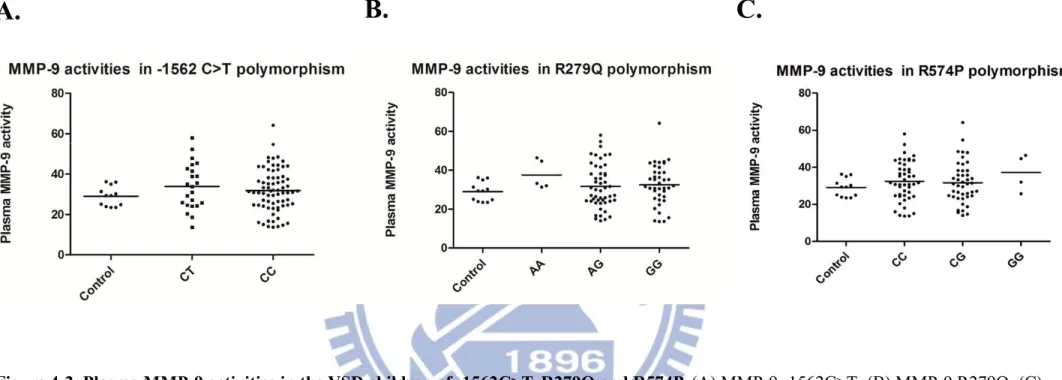
Control group: 31.8 (24.6–38.2), 33.9(24.6–42.8) ng/mL vs. 29 (25–32) ng/mL]. These two genotypes of MMP-9 -1562 C>T include 72 and 23 patients, respectively. We test the data through Student’s t-test and there is no significant difference of MMP-9 enzyme activities between different MMP-9 -1562C>T genotypes.
The results of MMP-9 polymorphism R279Q were shown in Fig. 4-2B. Each group of GG, GA, and AA has 40, 50, 5 samples and activities of them are 32.5 (25.8 – 39.0), 31.7 (24.1 - 39.1), 37.5(31.7– 45.6) ng/mL. Fig. 4-2C shows the part of MMP-9 R574P. Three groups of CC, CG, GG contain 47, 44, 4 samples and the results are 32.5 (24.6 – 41.5), 31.7 (24.2 – 38.2), 37.2(27.4 – 46.0) ng/mL, respectively. We used standard MMP-9 as positive control (Chemicon, Temecula, CA, USA) to calibrate MMP-9 activity of each group. As the data shown, MMP-9 activity had no significant difference in these MMP-9 genotypes of these two polymorphisms.
4-5. Relationship between plasma MMPs activity and VSD
severity
Fig. 4-3 shows the echocardiograms of a normal people and a perimembranous VSD patient. In clinically, large VSD patients with poor medical control are mandatory to surgery. So, our patients conducted in this study were brought into focus on the VSD patients without extreme large defects. First, we used VSD and VSD/Ao ration as the independent variables and determined the relationships between circulating MMP-2 and MMP-9 activities and VSD defects defined by VSD size and VSD/Ao ratio. According to the definition of VSD/aortic root (Ao) ratio, the patients were further classified into three groups: VSD/Ao ratio ≦ 0.2 is trivial group; 0.2 < VSD/Ao ≦ 0.3 is small group; and 0.3 < VSD/Ao ≦ 0.66 is considered to median group. Control group and each subgroup contain 12, 47, 29 and 19 patients,
32
next two paragraphs.
4-5-1. Relationships between MMPs activities and VSD defects
For the plasma MMP-2 activity, it did not show significant correlations with VSD size and VSD/Ao ratio (Fig. 4-4A and 4-4B). Whereas, positive correlations between MMP-9 activity and VSD size (r2 = 0.073, p < 0.05) as well as VSD/Ao ratio (r2 = 0.170, p < 0.05) were evaluated (Fig. 4-4C and 4-4D).
4-5-2. Relationships between MMPs activities and VSD defects grouping
MMP-2 activity in different VSD size groupsThe MMP-2 activities in children with normal and different VSD groups were
determined by gelatin zymography. The activities of MMP-2 enzyme in the VSD groups were compared with the Control group in Figure 13A (Median, Small, Trivial groups vs. Control group: 845 (710 – 945), 840 (618 – 1,050), 787 (604 – 935) ng/mL vs. 554 (407 – 689) ng/mL). The results revealed that the circulating MMP-2 activity was significantly increased in the VSD patients. Compared with the Control group, the MMP-2 activity of Trivial, Small and Median groups were enhanced by 1.4 (p < 0.01), 1.5 (p < 0.01) and 1.5 (p < 0.01) folds, respectively (Fig. 4-5A). However, differences in the plasma MMP-2 activity among the VSD groups were insignificantly.
4-5-3. MMP-9 activity in different size groups
The activities of plasma MMP-9 was also determined by gelatin zymography and shown in Fig. 4-5B (Median, Small, Trivial groups vs. Control group: 38 (32 – 43), 35 (26 – 43), 28 (19 – 35) ng/mL vs. 29 (25 – 32) ng/mL). A trend of increased MMP-9 activity in the VSD groups was found. However, only the MMP-9 activity in Median group was
33
4-6. Correlations between plasma MMP-2, MMP-9 activity,
concentrations of TIMP-3 and BNP in VSD patients
4-6-1. TIMP-3 in different VSD patients and MMPs polymorphisms
The concentrations of plasma TIMP-3 were also determined by ELISA, and similar to MMP-9 (Fig. 4-6A), there seemed to be a trend for a correlation between TIMP-3
concentration and VSD/Ao ratio; however, the result did not reach the significance threshold (r2 = 0.04, p = 0.054, shown in Fig. 4-6C). The TIMP-3 concentrations are no significant difference among VSD grouped by VSD/Ao ratio. [Median, Small, Trivial groups vs. Control group: 1853 (1222 – 2019), 1748 (1092 – 1827), 1800 (1135 – 1859) pg/mL vs. 1230 (1045 – 1462) pg/mL].
The TIMP-3 ELISA assay showed that the concentration of circulating TIMP-3 was higher concentration in subjects of the CC genotypes of MMP-2 -735 C>T and the QQ genotype than in those with other genotypes of the MMP-9 R279Q polymorphism (Fig. 4-7)
4-6-2. BNP in different VSD patients and MMPs polymorphisms
The plasma BNP concentration was determined by an immunoassay. There was a positive trend between plasma BNP concentration and VSD/Ao ratio (r2 = 0.36, p < 0.001 shown in Fig. 4-6B), similar to MMP-9. BNP concentrations were not significantly different among the different MMP-2, and MMP-9 polymorphisms investigated in this study, as shown in Fig. 4-8B, although each genotypes of MMP-2/-9 had higher BNP concentration than control group.
34
4-7. Pearson’s correlation between MMP-2/-9, TIMP-3, and BNP
Pearson’s correlation coefficient showed that there was correlation between plasma activity of MMP-9 and TIMP-3 concentration (r = 0.567, p < 0.001) (Fig. 4-9A) and also of MMP-9 and BNP (r = 0.463, p < 0.001) (Fig. 4-9B). In the contrast, there was no correlation between MMP-2 and TIMP-3 (r= 0.178, p = 0.139) (Fig. 4-9C); MMP-2 and BNP (r= 0.185, p = 0.07) (Fig. 4-9D). As shown in Fig.4-9E, there was no correlation between TIMP-3 and BNP concentration (r = 0.151, p = 0.130).4-8. Relationship between MMPs activity and spontaneous closure
rate of VSD
Patient groups were followed one year later by echocardiography. Totally 82 patients returned to the outpatient department for the programmed echocardiographic examinations. After one-year follow-up, a total of 14 spontaneous closures occurred (17%) and 68 patients remained their defects. The plasma MMP-2 activity of VSD patients in the spontaneous closure group was compared with non-closure group, and no significant difference was noted [791 (617 – 945) ng/mL vs. 808 (624 – 990) ng/mL, p = 0.825] (Fig. 4-10A). In contrast, the plasma MMP-9 activity of VSD patients in the spontaneous closure group was significantly higher than non-closure group [42 (32 – 48) ng/mL vs. 31 (25 – 38) ng/mL, p < 0.01] (Fig. 4-10B).
35
Table 4-1. VSD patients’ age and control group’ age, gender, VSD sizes, and VSD/Ao ratio VSD group (n = 95) Control group (n = 120) Male: Female 42 : 53 62 : 58 Age (years) 4.23 ± 2.51 5.53 ± 2.87 BNP (pg/mL) 35.5 ± 21.7 15.7 ± 9.5 VSD size (mm) 3.98 ± 1.40 Ao (mm) 18.92 ± 5.29 VSD/Ao ratio 0.23 ± 0.12
Data are expressed as mean ± SD; BNP, B-type natriuretic peptide; VSD, ventricular septal defect; VSD size, diameter of the VSD; Ao, diameter of the aortic root; VSD/Ao ratio, ratio of the diameter of the VSD and the diameter of the aortic root
36
Table 4-2. Patient grouping by VSD diameter/diameter of aortic root (Ao) ratio
Control
group
(n=12)
Trivial
group
(n=47)
Small
group
(n=29)
Median
group
(n=19)
Male: Female 6:6 21:26 14:15 7:12 Age (years) 4.76 ± 2.74 4.06 ± 2.26 4.16 ± 2.66 4.84 ± 2.96 VSD size (mm) - 3.06 ± 0.88 4.30 ± 0.85 5.75 ± 1.21 VSD/Ao ratio - 0.14 ± 0.03 0.25 ± 0.04 0.43 ± 0.10 Data were expressed as mean SD; VSD, ventricular septal defect; Ao, aortic root diameter
37
Table 4-3. Genotype distribution and allele frequency of MMP-2 -735C>T polymorphism SNP Control (n = 120) VSD (n = 95) OR (95% CI) p value n (%) n (%) MMP-2 -735C>T CC 48 (40.0) 45 (47.4) Reference CT 60 (50.0) 36 (37.9) 0.6 (0.16-1.14) 0.143 TT 12 (10.0) 14 (14.7) 1.24 (0.52-2.58) 0.662 C allele 156 (65.0) 126 (66.3) 0.96 (0.64–1.43) 0.919 T allele 84 (35.0) 64 (33.7)
38
Table 4-4. Genotype distribution and allele frequency of MMP-9 -1562C>T polymorphism SNP Control (n = 120) VSD (n = 95) OR (95% CI) p value n (%) n (%) MMP-9 -1562C>T CC 94 (78.3) 73 (76.8) 1.09 (0.57–2.08) 0.79 CT 26 (21.7) 22 (23.2) TT 0 0 C allele 214 (89.2) 166 (88.3) 1.09 (0.60–1.99) 0.78 T allele 26 (10.8) 22 (11.7)
39
Table 4-5. Genotype distribution and allele frequency of MMP-9 R279Q polymorphism
SNP Control (n = 120) VSD (n = 95) OR (95% CI) p value n (%) n (%) MMP-9 R279Q GG 69 (57.5) 39 (41.1) Reference GA 42 (35) 51 (53.7) 0.47 (0.26–0.82) 0.023 AA 9 (7.5) 5 (5.3) 0.98 (0.31–3.14) 0.98 A allele 60 (25) 61 (32.1) 1.41 (0.92–2.15) 0.11 G allele 180 (75) 129 (67.9)
40
Table 4-6. Genotype distribution and allele frequency of MMP-9 R574P polymorphism
SNP Control (n = 120) VSD (n = 95) OR (95% CI) p value n (%) n (%) MMP-9 R574P CC 63 (52.5) 46 (48.4) Reference CG 46 (38.3) 45 (47.4) 1.34 (0.77–2.35) 0.31 GG 11 (9.2) 4 (4.2) 0.50 (0.15–1.66) 0.25 C allele 172 (71.7) 136 (72.3) 0.97 (0.63–1.48) 0.89 G allele 68 (28.3) 52 (27.7)
A.
B.
C.
Figure 4-2. Plasma MMP-9 activities in the VSD children of -1562C>T, R279Q and R574P. (A) MMP-9 -1562C>T; (B) MMP-9 R279Q; (C)
MMP-9 574P. As data shown, MMP-9 activities have no significant difference between VSD patients in these three subgroups of MMP-9 polymorphisms.






![TraditionalMLCalgorithmsmainlytacklethebatchMLCproblem,wheretheinputdataarepresentedinabatch[24,28].Nevertheless,inmanyMLCapplicationssuchase-mailcategorization[22],multi-labelexamplesarriveasastream.Onlineanalysisistherefore dimensionreducermotivatedbyma](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)