本完整報告,以由此計畫支持而完成之三篇論文為主。
1. Liu, T.-A., Liu, M.-C. and *Yang, Y.-S. (2008) Immunohistochemical Analysis of a Novel Dehydroepiandrosterone Sulfotransferase-like Protein in Drosophila Neural Circuits, Biochem. Biophys. Res. Comm., 367, 14-20. (SCI)
2. Lu, L.-Y., Hsieh, Y.-C., Liu, M.-Y., Lin, Y.-H., Chen, C.-J. and *Yang, Y.-S. (2008) Identification and Characterization of Two Amino Acids Critical for the Substrate Inhibition of Human Dehydroepiandrosterone Sulfotransferase (SULT2A1) Molecular Pharmacology, 73, 660-668. (SCI)
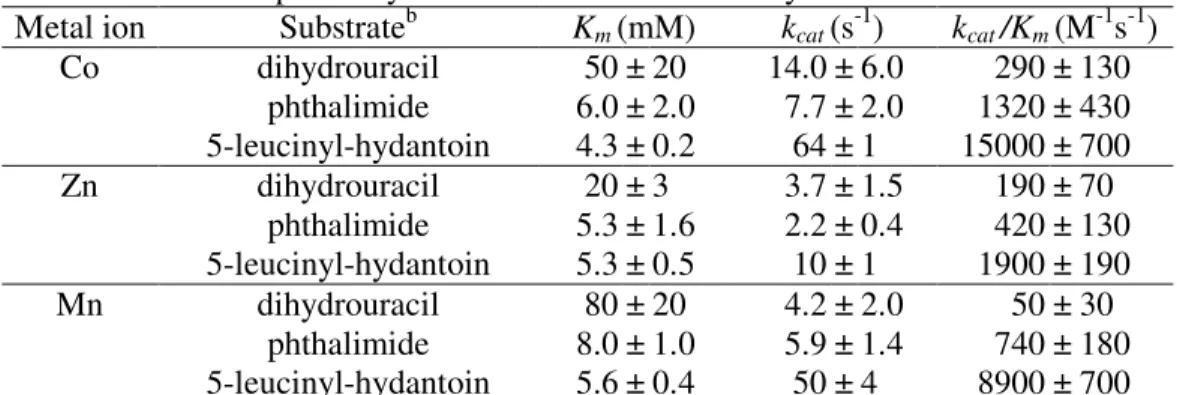
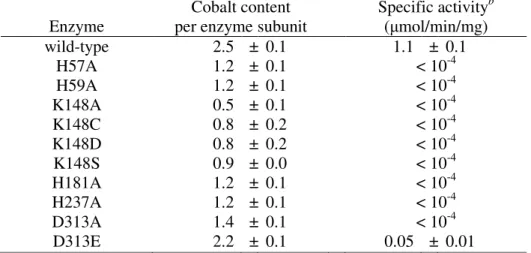
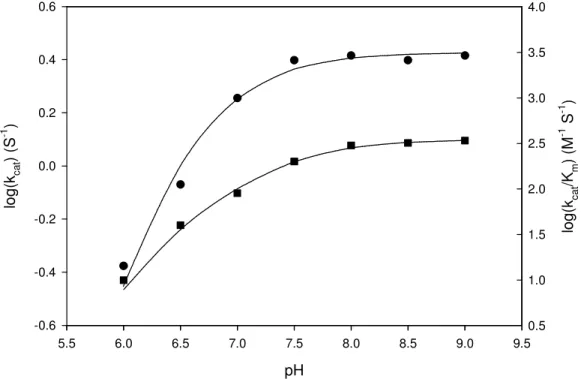
3. Huang, C.-Y., Hsu, C.-C., Chen, M.-C. and *Yang, Y.-S. (2008) Effect of metal binding and posttranslational lysine carboxylation on the activity of recombinant hydantoinase, J. Bio Inorg Chem, accepted. (SCI)
醯胺水解酵素超家族,包括醯亞胺水解酵素、尿囊素水解酵素、二氫乳清酸 水解酵素、海因水解酵素…等,這類酵素雖然在一級序列比對上的相似度可能只 有不到百分之三十,但都保留相同催化部份的結構。其中,被認為與酵素催化有 非常重要的關聯並高度保留的有四個組胺酸、一個天門冬胺酸、以及一個經過後 修飾的離胺酸與金屬離子螯合。此外,由序列與結構的比對也發現亞硫酸基轉移 酵素與核苷酸激酶雖然序列相似度在百分之二十以下,但結構上卻有一定程度的 相似,其特定結構的保留可能與特殊的受質/配體結合有關。此二類酵素之關連 性的研究亦十分值得進一步研究。本計劃的目的在於整合生物資訊、生物化學以 及分子生物學三種不同的途徑來更進一步了解這些酵素催化反應中的反應機制 與功能相關性。另一方面,我們也計畫對此酵素之催化相關後修飾胺基酸以點突 變方式進行解析。並嘗試解析這些胺基酸殘基對催化機制的影響。此外,針對亞 硫酸基轉移酵素的部分,我們也利用本計畫發展的工具進行分析,並進行不同類 別之酵素的單元融合進而創造出雙功能的人工酵素。 關鍵字:醯胺水解酵素超家族、醯亞胺水解酵素、亞硫酸基轉移酵素、核苷酸激 酶。
Amidohydrolase superfamily includes imidase, allantoinase, dihydroorotase, hydantoinase, and other related enzymes that were proposed based on the rigidly conserved structural domains in identical positions. Although the overall sequence homology within the enzyme superfamily can be lower than 30%, these conserved regions, compose of four histidines, one aspartate, and one modified lysine which is bridging one or two metal ions are involved in the catalysis. In addition, sulfotransferases and nucleotide kinases were also found to be similar in structure but different in sequence identity (<20%). The conservation of structural features may be correlated to specific functions such as substrate/ligand binding. The goal of this proposal is to understand the mechanism and functional properties of these enzyme families by using combined techniques of bioinformatics, biochemistry and molecular biology. In particular, we would like to distinguish the need for either a binuclear or mononuclear metal centre in imide hydrolysis. During the past year, we use site-directed mutagenesis to evaluate the role of the catalytic residues predicted by FAVA. In addition, we mutate residue of the oligomerization interface of sulfotransferase and successfully created a chimerical bi-functional sulfotransferase. Keywords: Amidohydrolase Superfamily, Imidase, Sulfotransferase, Nucleotide Kinase.
Immunohistochemical Analysis of a Novel
Dehydroepiandrosterone Sulfotransferase-like Protein in
Drosophila Neural Circuits
Tzu-An Liu1, Ming-Cheh Liu2 and Yuh-Shyong Yang1,3*
1 Department of Biological Science and Technology and Institute of Biochemical Engineering, National Chiao Tung University, Hsinchu, Taiwan
2 Department of Pharmacology, College of Pharmacy, The University of Toledo, Toledo, OH 43606 USA
3 Instrument Technology Research Center and National Nano Device Laboratories, NARL, Hsinchu, Taiwan
*Corresponding author:
Dr. Yuh-Shyong Yang Department of Biological Science and Technology, National Chiao Tung University,
75 Po-Ai Street,
Hsinchu30050, TAIWAN Fax: 886-3-5729288
E-mail: [email protected]
ABSTRACT
Sulfotransferase (ST)-catalyzed sulfation plays an important role in various neuronal functions such as homeostasis of catecholamine neurotransmitters and hormones.
Drosophila is a popular model for the study of memory and behavioral manifestations
because it is able to mimic the intricate neuroregulation and recognition in humans. However, there has been no evidence indicating that cytosolic ST(s) is(are) present in
Drosophila. The aim of this study is to investigate whether or not cytosolic ST(s)
is(are) expressed in the Drosophila nervous system. Immunoblot analysis demonstrated the presence of dehydroepiandrosterone (DHEA) ST-like protein in
Drosophila brain and a sensitive fluorometric assay revealed its sulfating activity
toward DHEA. Immunohistochemical staining demonstrated this DHEA ST-like protein to be abundant in specific neurons as well as in several bundles of nerve fibers in Drosophila. Clarification of a possible link between ST and a neurotransmitter-mediated effect may eventually aid in designing approaches for alleviating neuronal disorders in humans.
INTRODUCTION
Cytosolic STs are enzymes that catalyze the transfer of a sulfuryl group from the universal sulfate donor 3’-phophoadenosine 5’-phophosulfate (PAPS) to a variety of endogenous and exogenous compounds, such as steroids, amines, and various xenobiotic chemicals (1). The reaction, usually referred to as “sulfation”, occurs in many prokaryotic and vertebrate species and plays an important role in numerous biological processes including hormone regulation, homeostasis of neurotransmitters, as well as transport and metabolism of steroids in circulation. The sulfation of steroids decreases their biological activity, rendering them incapable of binding and activating steroid receptors. These sulfated steroids, nevertheless, may serve as prohormones, which can be reactivated by desulfation (2). Although most steroids are synthesized in steriodogenic organs, a few, such as progesterone (PROG), pregnenolone (PREG) and DHEA, are produced de novo in the central nervous system (CNS) and peripheral nervous system (PNS) (3). These neurosteroids regulate specific gene expression and protein synthesis, cellular development, neuroendocrine functioning, and behavioral pattern (4). Dehydroepiandrosterone sulfotransferase (DHEA ST) catalyzes the sulfation of DHEA, converting it to dehydroepiandrosterone sulfate (DHEAS) (5). Both DHEA and DHEAS are critical precursors for the production of several types of androgens and estrogens. DHEA can also be metabolized to form testosterone, estradiol and androstenediol (6). DHEA and DHEAS are involved in numerous neurophysiological processes, such as increasing neuronal excitability, and enhancing neural plasticity and neuroprotective properties. Previous studies have demonstrated that the concentrations of DHEA and DHEAS in blood decrease markedly with age in humans, and have been proposed to be the neuromodulators involved in age-related cognitive decline (7). These findings have led to the hypothesis that elevated concentrations of steroids may influence both physical and cognitive aging.
Drosophila melanogaster is a popular animal model for pathological and
neuropharmacological research. At the molecular level, the nervous system of the
Drosophila can mimic the intricate neuroregulation of the neuronal network in
humans. In Drosophila, several types of carbohydrate STs have been cloned and characterized, and the physical functions and biological regulations modulated by the sulfate conjugates have been extensively investigated (8). The sulfation of hexuronate and glucosamine units, usually N-acetylgalactosamine (GlaNAc) or N-acetylglucosamine (GlcNAc), has been shown to be essential for development and
embryogenesis, as well as differentiation and neuronal functions (9). The recent studies have demonstrated that sulfation is a critical regulator for developmental and neuronal functions in Drosophila, however, there has been no evidence indicating that cytosolic ST(s) is(are) present in this important animal model. In the present work, we used a specific DHEA ST antibody to determine the existence and distribution of DHEA ST-like protein in the nervous system of Drosophila. Moreover, we investigated and confirmed the presence of DHEA-sulfating activity of in Drosophila brain extracts by employing a continuous fluorometric assay.
MATERIALS AND METHODS
Preparation of recombinant STs
Recombinant human phenol-preferring phenol sulfotransferase (hP-PST), human catecholamine-preferring phenol sulfotransferase (hM-PST), and hDHEA ST were cloned into an expression vector, pGEX-2TK, respectively, and transformed into
Escherichia coli BL21 (DE3). These STs were expressed in the form of GST-fusion
protein and purified by glutathione (GSH)-bound sepharose. The methods of expression and purification of these three STs were described previously (10). Recombinant rat phenol sulfotransferase (rPST) was cloned into expression vector pET3c and transformed into E. coli BL21 (DE3). Briefly, DEAE, hydroxyapatite, and size-exclusion chromatography were used to purify the rPST (11). Putative
Drosophila cytosolic ST, dmCG5431, was found on the Flybase database (CG5431;
http://flybase.bio.indiana.edu/). By using RT-PCR, a full length cDNA encoding
dmCG5431 was cloned into pET-41b and transformed into E. coli BL21 (DE3).
GST-fusion and His-tagged ST was expressed and purified by GSH-bound sepharose and Ni-chelating column. All purified STs were in homogeneous form and determined by SDS-PAGE.
Indirected enzyme-linked immunosorbent assay (IELISA)
Approximately 1 µg of the recombinant STs, hP-PST, hM-PST, hDHEA ST, rPST and
dmCG5431 were coated on each well of an ELISA plate for screening using hDHEA
ST antibody. Following washes with PBS, 50 µl of serially diluted solutions of
hDHEA ST antibody were added to individual wells and incubated for 1 h.
Afterwards, each well was washed three times with PBST (PBS containing 0.1% BSA and 0.05% Tween-20). Bound antibodies were then detected using a goat anti-rabbit IgG conjugated with horseradish peroxidase (HRP) for 30 min in PBST. Finally, each well was washed and developed with 0.04% 2,2-azino-bis(3-ethylbenz- thiazoline-6-sulfonic acid) (ABTS) containing 0.01% H2O2 in PBS. For denatured conformation analysis, all steps were the same except that the recombinant STs were treated with 2% β-ME and heating prior to being coated on the wells.
Preparation of Drosophila brains homogenates
The Drosophila heads were freshly isolated by liquid nitrogen freezing and harvested through a sieve to separate the heads from the bodies. Approximately 0.2 g of frozen brain samples were homogenized using a mortar and pestle, dissolved in a lysis buffer (2 mM sucrose plus 3 mM β-ME, 0.2% Triton X-100 and 0.5% protease inhibitor cocktail in 10 mM HEPES buffer, pH 7.4) and then centrifuged to remove cell debris. The homogenate was centrifuged twice at 15,000 rpm for 20 min at 4°C. The supernatant was collected and the total protein concentration was estimated.
Immunoblot analysis
Approximately 5µg of five recombinant STs, hP-PST, hM-PST, hDHEA ST, rPST and
dmCG5431, respectively, were loaded onto individual wells of a 12% SDS-PAGE for
electrophoresis according to the method of Laemmli (12). After electrophoresis, the separated proteins were electroblotted onto a nitrocellulose membrane and blocked with 5% skimmed milk for 1 h. The membrane was incubated with hDHEA-ST antibody for 1 h and washed three times with PBST for 5 min. The membrane was then immersed in PBST containing antibody against rabbit IgG conjugated with HRP for 1 h. The bound antibodies were detected with an ECL western blotting reagents for chemiluminescent detection. The native immunoblot analysis procedure was similar to that described previously with minor modifications. All buffer solutions were free from the addition of SDS. Similarly, approximately 300 µg of Drosophila brain proteins and 20 µg of purified hDHEA ST were used for analyzing the protein expression of DHEA ST-like protein by native and SDS immunoblot. The steps were similar to those described previously.
Determination of sulfating activity of DHEA in Drosophila brain
DHEA-sulfating activity of Drosophila brain was determined by the continuous fluorometric assay developed by Chen et al. (13). By using DHEA as substrate, the
activity of DHEA ST was determined by monitoring the fluorescence intensity of MU. The standard assay mixture had a final volume of 1 ml, and contained 100 mM potassium phosphate buffer (pH 7.0), 5 mM β-ME, 20 µM PAPS, 4 mM MUS, 5 µM DHEA and 3.2 mU K65ER68G, the recombinant β-form of PST. For use in the assay, a partially purified DHEA ST-like protein fraction, located by cross-reactivity with antibody against hDHEA ST, was prepared from Drosophila brain homogenates by using native gel electrophoresis. The intensity of MU was monitored using a spectrofluorometer (Hitachi F-4500, Japan).
Immunohistochemistry
The Drosophila brain was perfused with 4% paraformaldehyde for fixation and then penetrated with 30% sucrose. After washing with PBS, the brain was blocked with 1% BSA overnight at 4°C to prevent nonspecific staining. The sample was then incubated with hDHEA ST antibody at 4°C for 72 h, rinsed with washing buffer (containing 0.1% BSA and 0.2% Triton X-100 in PBS, pH 7.4) for 20 min three times, and then stained with biotinylated goat anti-rabbit secondary antibody. After an overnight incubation at 4°C, the sample was washed with washing buffer for 20 min. Tertiary antibodies (streptavidin-Cy5 and biotinylated HRP) were also incubated with the brain sample overnight at 4°C. Afterwards, the brain sample was mounted using FocusClearTM and examined by confocal laser scanning microscope photomicrographs. The Drosophila carries gene trap Gal4 (12423) and UAS-GFP to expresses GFP in the DPM neurons (green) was used to analyze the colocalization of DHEA-ST like protein and DPM neuron. Steps were same as mentioned above (14).
RESULTS
Characterization of hDHEA ST antibody
The characterization of hDHEA ST antibody analyzed by IELISA and immunoblotting is shown in Fig. 1. The hDHEA ST antibody showed stronger affinity toward recombinant hDHEA ST and dmCG5431 in native conformations than the others co-tested (Fig. 1A). The dose-response curves showed that the hDHEA ST antibody was specific to hDHEA ST when the STs tested were first denatured by treating with β-ME and heating (Fig. 1B). The binding of hDHEA ST antibody to PST isoforms was barely discernible. The specificity of antibody for hDHEA ST was determined by probing five different types of recombinant STs by immunoblotting (Fig. 1C). The hDHEA ST antibody interacted with both hDHEA ST and dmCG5431 in their native forms, whereas neither hPST isoforms nor rPST was recognized. Furthermore, only hDHEA ST was identified by this specific antibody under the denatured conditions (Fig. 1D).
Expression of DHEA ST-like Protein in Drosophila brains
As shown in Fig. 2, the immunoblot of Drosophila brain extracts was analyzed by
hDHEA ST antibody. The DHEA ST-like protein expressed in soluble extracts of Drosophila brains was recognized in its native form (Fig. 2A). The SDS-PAGE
immunoblot showed that only hDHEA ST (positive control) was recognized when the native conformation was disrupted (Fig. 2B).
Localization of DHEA ST-like Protein in Drosophila brains
The distribution and relative abundance of DHEA ST-like protein in Drosophila brain are shown in Fig. 3. A total of six DHEA ST-like protein positive neurons were observed in the posterior section of Drosophila brain (Fig. 3A). Two DHEA ST-like protein positive neurons were also detected in the dorsal part of the brain (Fig. 3B). DHEA ST-like protein positive neuronal fibers, exhibiting the typical appearance of beaded nerve fibers, were seen throughout the entire Drosophila brain (Fig. 3C). Table
1 summarizes the relative localization and abundance of DHEA ST-like protein positive neurons and fibers. By using UAS-WGA as a trans-synaptic transmission marker and VAM-Gal4 as a driver, the colocalization of DHEA ST-like protein positive regions and DPM neuron were observed by transgenic fly that carries VAM-Gal4 and UAS-WGA and express WGA in the VAM neurons (Fig. 4).
Determination of the DHEA-sulfating activity in Drosophila brains
The sulfating activity of DHEA in partially purified Drosophila brain extract was assessed using a continuous fluormetric assay (Table 2). The complete system (I) could detect not only sulfating but also desulfating activities due to the presence of MUS. The DHEA-sulfating activity could not be observed in the absence of PAPS, and therefore reaction condition II gave a background activity exhibited mainly by arylsulfatase. As a result, the specific activity and total activity of DHEA-sulfating activities in Drosophila brain were determined to be 57.7 ± 12.1 pmole/min/mg and 0.7 ± 0.2 nmole/min/g, respectively. It is to be noted hat a high level of arylsulfatase activity (specific activity and total activity, 319.1 ± 7.5 pmol/min/mg and 6.8 ± 0.1 nmol/min/g, respectively) in the fraction was detected as well.
Discussion
To date twenty-three structures of cytosolic STs have been solved on eleven different isoforms. Crystal structures for the ST1A1 (phenol ST), ST1A3 (catecholamine ST), ST1E1 (estrogen ST), ST1B1 (thyronine ST), ST2A1 (DHEA ST), two isoenzymes of ST2B1 (pregnenolone ST and cholesterol ST), ST4A1 (neuronal ST) and three subfamilies of ST1C have been characterized. Structure-based sequence alignments indicate that the PAPS binding site, and structural fold, is highly conversed, albeit the homology of the amino acid sequences between different ST isoforms is not high (15). On the basis of the characteristic pattern of the STs, we demonstrated the possible presence of a cytosolic ST-like protein in Drosophila neural circuits by the specific recognition of the hDHEA ST antibody. The results obtained from immunoblot analysis and IELISA are close agreement with conserved nature of STs and indicate that STs may exert similar biological functions in various animals.
Drosophila is an excellent experimental model to systematically study the
neuroregulative mechanisms in human CNS. Many scientists have placed much effort into the molecular characterization and physical relevance of STs in this tiny creature, however, the biological significance of STs in Drosophila remains obscure. In general, sulfate conjugation is apparently involved in the metabolism of juvenile hormones and ecdysteroids in insects (16). In Prodenia eridania, sulfate conjugation of ecdysteroids seems to play a critical role in embryonic development and puparium formation (17). In the present study, the protein partially purified from Drosophila brain extracts was demonstrated to be capable of transferring a sulfuryl group from a sulfate donor, PAPS, to an analog of ecdysteroid, DHEA, and the result was consistent with that reported for Mosquito, Aedes togoi (18) suggested that sulfation in various insect species may exhibit similar biological functions in the metabolism of free hormones and post-stage embryogenesis. Earlier studies had also revealed the existence of cytosolic STs in flies of Diptera, Prodenia eridania (19). In addition to its activity on
p-nitrophenol, the ST prepared from Prodenia eridania gut was significantly active in
the sulfation of the steroids, such as DHEA, oestrone, and insect moulting hormones α-ecdysone and 22,25-bisdeoxyecdysone. It appears logical to suggest that STs comprehensively regulate the biochemical transformations for the purpose of detoxication, others may have important physiological implications in insects. Besides,
it is noted that DHEA-sulfating activity in Drosophila was significantly lower than that in rat and human (20, 21). This is to be expected because DHEA is a more common substrate for DHEA ST in mammalians than in insects. Furthermore, the validity of such comparison is always open to some question in consideration of differences in enzyme preparation and enzymatic assay. Nevertheless, It is noteworthy that the significant level of arylsulfatase activity was determined (Table 2). In agreement with these previous findings, the low or undetectable DHEA ST activities may be due to high levels of steroid sulfatase in soluble extracts, thereby interfering with the determination of DHEA ST in vitro (22).
Several neural regions of Drosophila brain were found to have DHEA ST-like immunoreactivity in this study. The immunoreactivity was selectively localized in the neurons of posterior and dorsal part of Drosophila brain, and nerve fibers indicating the relevant molecular and neuronal mechanism between this enzyme and its metabolites. In Drosophila, there are several types of projection neuron that forward information out of the antennal lobes. Medial and outer antennocerebral tracts (mACT and oACT, respectively) protrude into the ill-defined region and the lateral horn, while the others carry information along the inner antennocerebral tracts (iACT) to the mushroom body (MB). In our study, we observed an abundance of Drosophila DHEA ST-like protein expressed in the lateral horn and iACT (Table 1). The results implied that the DHEA ST-like protein may act as a neuromodulator of the ecdysteroids, which are involved in memory formation in Drosophila. The finding is also consistent with the result reported by Johnson et al (23). The sulfate-conjugated steroid is essential to the process of memory retention and significantly enhanced the cognition and learning in rats. Basically, they act through γ-aminobutyric acidA (GABAA) receptors, N-methyl-D-aspartate (NMDA)-type glutamatergic receptors and sigma receptors to induce excitatory cellular actions or inhibit cellular properties. Increasing evidences have suggested that DPM neuron may co-release amnesiac neuropeptide and acetylcholine in the amnesiac mutant flies (24). Transgenic expression of the amnesiac gene in the DPM neurons rescues the amnesiac memory phenotype, establishing a possible route between DPM neuron function and
amnesiac-dependent memory. Additionally, the paired conditioning of unconditioned
stimulus (US) (electric shock) and conditioned stimulus (CS) (odor stimulus) increases odor-evoked calcium signals and synaptic release from DPM neurons (25). These observations indicated that DPM neurons not only respond to the US pathway, but that they are also “odor generalists”, responding to all odors that were tested. The colocalization of DHEA ST-like protein and DPM neuron implied that DHEA ST-like protein may not merely play the role as a neuroregulator in the process of odor-specific memory trace in Drosophila but also involve in the modulation of specific memorial and behavioral formations.
ACKNOWLEDGMENT
This research was financially supported by National Science Foundation, under Grant NSC 96-96-2120-M-009-001 and 96-2627-B009-004. The authors deeply appreciate the technical contribution of Dr. Ann-Shyn Chiang, Institute of Biotechnology at National Tsing Hua University, and Ms. Pei-Chi Zheng, and Mr. Wei-Ti Chen, Institute of Biochemical Engineering at NCTU.
REFERENCES
[1] E. Chapman, M.D. Best, S.R. Hanson, C.H. Wong, Sulfotransferases: structure, mechanism, biological activity, inhibition and synthetic utility, Angew. Chem. Int. Ed.
43 (2004) 3526-3548.
[2] C.N. Falany, Enzymology of human cytosolic sulfotransferases, FESAB J. 11 (1997) 206-216.
[3] A.F. De Nicola, Steroid hormones and neuronal regeneration, Adv. Neurol. 59 (1993) 1199-1206.
[4] M. Vallée, W. Mayo, M.L. Maol, Role of pregnenolone, dehydroepiandrosterone and their sulfate esters on learning and memory in cognitive aging, Brain Res. Rev. 37 (2001) 301-312.
[5] C. Longcope, Dehydroepiandrosterone metabolism, J. Endocrinol. 150 (1996) S125-S127.
[6] J. Young, B. Couzinet, K. Nauhoul, S. Brailly, P. Chanson, E.E. Baulieu, G. Schaison, Panhypopituitarism as a model to study the metabolism of dehydroepiandrosterone (DHEA) in humans, J. Clin. Endocrinol. Metab. 82 (1997) 2578-2585.
[7] S. Legrain, L. Girard, Pharmacology and therapeutic effects of dehydroepiandrosterone in older subjects, Drugs Aging 20 (2003) 949-967. [8] M. Kushe-Gullberg, L. Kjellén, Sulfotransferases in glycosaminoglycan biosynthesis, Curr. Opin. Struct. Biol. 13 (2003) 605-611.
[9] A.D. Lander, C.S. Stipp, J.K. Ivins, The glypican family of heparan sulfate proteoglycans: major cell-surface proteoglycans of the developing nervous system. Perspect. Dev. Neurobiol. 3 (1996) 347-358.
[10] Y. Sakakibara, Y. Takami, T. Nakayama, M. Suiko, M.C. Liu, Localization and functional analysis of the substrate specificity/catalytic domains of human M-form and P-form phenol sulfotransferases, J. Biol. Chem. 273 (1998) 6242-6247.
[11] T.M. Su, Y.S. Yang, Mechanism of posttranslational regulation of phenol sulfotransferase: expression of two enzyme forms through redox modification and nucleotide binding, Biochemistry 42 (2003) 6863-6870.
[12] U.K. Laemmli, Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature 227 (1970) 680-685.
[13] W.T. Chen, M.C. Liu, Y.S. Yang, Fluorometric assay for alcohol sulfotransferase, Ana. Biochem. 339 (2005) 54-60.
[14] Y. Wang, A.S. Chiang, S. Xia, T. Kitamoto, T. Tully, Y. Zhong, Blockade of neurotransmission in Drosophila mushroom bodies impairs odor attraction, Curr. Biol. 13 (2003) 1900-1904.
[15] V.L. Rath, D. Verdugo, S. Hemmerich, Sulfotransferase structural biology and inhibitor discovery, Drug Discov. Today 9 (2004) 1003-1011.
[16] A. Sannasi, P. Karlson, Metabolism of ecdysone: phosphate and sulphate esters as conjugates of ecdysone in Calliphora vicina, Zool. Jb. Physiol. 78 (1974) 378-386. [17] M. Slade, C.F. Wilkinson, Degradation and conjugation of Cecropia juvenile hormone by the southern armyworm (Predenia eridania), Comp. Biochem. Physiol. 49 (1974) 99-103.
[18] L. Shampengtong, K.P. Wong, An in vitro assay of 20-hydroxyecdysone sulfotransferase in the mosquito, Aedes togoi, Insect Biochem. 19 (1989) 191-196. [19] R.S. Yang, C.F. Wilkinson, Enzymatic sulphation of p-nitrophenol and steroids by larval gut tissues of the southern armyworm (Prodenia eridania cramer), Biochem. J. 130 (1972) 487-493.
[20] S. Aldred, R.H. Waring, Localization of dehydroepiandrosterone sulphotransferase in adult rat brain, Brain Res. Bull. 48 (1998) 291-296.
[21] T. Sugahara, Y.S. Yang, C.C. Liu, T.G. Pai, M.C. Liu, Sulphonation of dehydroepiandrosterone and neurosteroids: molecular cloning, expression, and
functional characterization of a novel zebrafish SULT2 cytosolic sulphotransferase, Biochem. J. 375 (2003) 785-791.
[22] G. Janer, S.
Mesia-Vela, F.C. Kauffman, C. Porte,
Sulfatase activity in the oyster Crassostrea virginica: its potential interference with sulfotransferase determination, Aquat. Toxicol. 74 (2005) 92-95.[23] D.A. Johnson, T.H. Wu, P.K. Li, T.J. Mather, The effect of steroid sulfatase inhibition on learning and spatial memory, Brain Res. 865 (2000) 286-290.
[24] A.C. Keene, M. Stratmann, A. Keller, P.N. Perrat, L.B. Vosshall, S. Waddell, Diverse odor-conditioned memories require uniquely times dorsal paired medial neuron output, Neuron 44 (2004) 521-533.
[25] D. Yu, A.C. Keene, A. Srivatsan, S. Waddell, R.L. Davis, Drosophila DPM neurons forma delayed and branch-specific memory trance after olfactory classical conditioning, Cell 123 (2005) 945-957.
LEGENDS FOR FIGURE AND TABLE
Figure 1 The characterization of hDHEA ST antibody using recombinant STs and putative cytosolic ST of Drosophila. (A) The hDHEA ST antibody titre determined by IELISA using recombinant proteins in native conformation. (B) The hDHEA ST antibody titre determined by IELISA using recombinant proteins denatured by treatment with β-ME and heating. (C) Native immunoblot of the five recombinant proteins with hDHEA ST antibody. (D) SDS-PAGE immunoblot of the five recombinant proteins with hDHEA ST antibody. Each lane contained approximately 5 µg purified proteins.
Figure 2 The expression of DHEA ST-like protein in Drosophila brain extracts. (A) The native immunoblot of Drosophila brain extracts with hDHEA ST antibody. (B) SDS-PAGE immunoblot of Drosophila brain extracts and purified hDHEA ST with
hDHEA ST. dmBE represented Drosophila brain extracts.
Figure 3 Distribution of the DHEA ST-like protein in Drosophila brain as illustrated by confocal laser scanning microscope. (A) DHEA ST-like protein selectively expressed in posterior region of brain. (B) DHEA ST-like protein expressed in dorsal part of the brain. (C) Expression of DHEA ST-like protein in neural fibers throughout the entire Drosophila brain. Granular staining suggests fibers labeled. 200X magnification.
Figure 4 Colocalization of the DHEA ST-like protein expression and DPM neuron in
Drosophila brain. The arrowhead indicates the colocalization (orange) of DHEA
ST-like protein (red) and DPM neuron (green) (400X magnification). Right-lower: the magnification of the colocalized region of DHEA ST-like protein and DPM neuron.
Table 1 Distribution and cellular colocalization of DHEA ST-like protein in Drosophila braina
Brain region Specific areasb Neural
cytoplasmc Fibers c Anterior brain MB’s vertical lobe ---- --- -superior medial PR ---- --- -median bundle ---- --- -MB satellite neuropil ---- ---
-anterior optic tubercle ---- +
ventrolateral PR ---- + MB’s medial lobe ---- --- -antennal lobe ---- --- -antennal nerve ---- --- -ventral body ---- --- -Middle brain pedunculus ---- + fan-shaped body ---- + superior lateral PR ---- ++ ventral body ---- ++ superior medial PR ---- + tritocerebrum ---- ++ superior arch ---- + antennoglomerular tract ---- ++ ellipsoid body ---- + nodulus ---- ++
inferior lateral deutocerebrum ---- +
Lo ---- ++ Medulla ---- + Posterior brain Lo ---- --- -LoP ---- ++ MB calyx ---- - PR bridge ++++++++++++ +
inner antennocerebral tract ---- +
lateral horn ++++++++++++ -
posterior lateral fascicle ---- ++
SOG ---- +
axons of vertical cells of the LoP ---- ++
axons of horizontal cells of the LoP ---- +
SOG nerves ---- ---
-aThe distribution of DHEA ST-like protein in Drosophila brain was investigated by the
continuous sections of the confocal laser scanning microscope photomicrographs. Detailed procedures were mentioned under Materials and Methods (14).
bAbbreviations: MB, mushroom body; PR, protocerebrum; Lo, lobula; LoP, lobula plate; SOG,
subesophaeal ganglis; SOG nerves, roots of nerves from the fused subesophageal ganliea.
cThe relative intensity of labeling was ranked by two independent observers. Ratings reflect
mainly the density of DHEA ST-like protein labeled cell and fibers. Negative, -; weak, +; moderate, ++; strong +++.
Table 2 Sulfation and sulfatase activity in Drosophila braina
Fraction Reaction conditions Enzyme activity involved Specific activity (pmole/min/mg) Total activity (nmole/min/g) I. Completeb DHEA-sulfating protein + arylsulfatase 376.8 ± 9.5 7.5 ± 0.2
II. – PAPSc arylsulfatase 319.1 ± 7.5 6.8 ± 0.1
I – II DHEA-sulfating
protein 57.7 ± 12.1 0.7 ± 0.2
III. –Lysates K65ER68G 16.6 ± 1.7
aFor use in the assay, a partially purified fraction which cross-react with antibody against
hDHEA ST, was isolated from Drosophila brain homogenates by using native gel electrophoresis.
bDetailed procedures were described under Coupled-enzyme assay for alcohol sulfotransferase
(AST) in Materials and Method (13). Specific activity referred to MU produced following the
addition of extract whose protein concentration was determined by absorption at A280. Total
activity referred to MU produced with one gram of Drosophila brain extracts.
Identification and Characterization of Two Amino Acids Critical for the Substrate Inhibition of Human Dehydroepiandrosterone Sulfotransferase (SULT2A1)
Lu-Yi Lu, Yin-Cheng Hsieh, Ming-Yih Liu, Yih-Hung Lin, Chun-Jung Chen and Yuh-Shyong Yang
Department of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan, ROC
a) Running Title: Substrate inhibition of sulfotransferase
b) Corresponding author:
Dr. Yuh-Shyong Yang
Department of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan, ROC
75 Po-Ai Street, Hsinchu30050, TAIWAN, ROC Tel: +886-3-573-1983
Fax: +886-3-57-29288
E-mail: [email protected]
c) Numbering:
Number of text pages: 42 Number of Tables: 5 Number of Figures: 5 Number of references: 40
Number of words in the Abstract: 212 Number of words in the Introduction: 539 Number of words in the Discussion: 1728
d) Abbreviations: SULT, sulfotransferase; PAPS, 3′-phosphoadenosine 5′-phosphosulfate; PAP,
3′-phosphoadenosine 5′-phosphate; DHEA, dehydroepiandrosterone; ADT, androsterone; MUS, 4-methylumbelliferyl sulfate; MU, 4-methylumbelliferone; pNP, p-nitrophenol.
Abstract
Substrate inhibition is a characteristic feature of many cytosolic sulfotransferases. The differences between the complex structures of SULT2A1/DHEA and SULT2A1/PAP or SULT2A1/ADT (PDB codes are: 1J99, 1EFH, and 1OV4, respectively) have enabled us to elucidate the specific amino acids responsible for substrate inhibition. Based on the structural analyses, substitution of the smaller residue alanine for Tyr-238 (Y238A) significantly increases the Ki value for dehydroepiandrosterone (DHEA) and totally eliminates substrate inhibition for
androsterone (ADT). In addition, Met-137 is proposed to regulate the binding orientations of DHEA and ADT in SULT2A1. Complete elimination or regeneration of substrate inhibition for SULT2A1 with DHEA or ADT as substrate, respectively, was demonstrated with the mutations of Met-137 on Y238A mutant. Analysis of the Met-137 mutants and Met-137/Tyr-238 double mutants uncovered the relationship between substrate binding orientations and inhibition in SULT2A1. Our data indicate that, in the substrate inhibition mode, Tyr-238 regulates the release of bound substrate, and Met-137 controls substrate binding orientation of DHEA and ADT in SULT2A1. The proposed substrate inhibition mechanism is further confirmed by the crystal structures of SULT2A1 mutants at Met-137. We propose that both substrate binding orientations exhibit substrate inhibition. In addition, a corresponding residue in other cytosolic sulfotransferases was shown to have a function similar to that of Tyr-238 in SULT2A1.
Introduction
Sulfation is a widespread biological reaction catalyzed by members of the sulfotransferase (SULT) supergene family. These enzymes catalyze the transfer of a sulfuryl group from 3’-phosphoadenosine 5’-phosphosulfate (PAPS), the universal sulfuryl group donor molecule, to a substrate acceptor group. Cytosolic sulfotransferases catalyze sulfation of small molecules such as drugs, steroid hormones, chemical carcinogens, bile acids, and neurotransmitters (Mulder and Jakoby, 1990; Falany and Roth, 1993; Weinshilboum and Otterness, 1994; Glatt, 1997). Human SULT2A1 (dehydroepiandrosterone sulfotransferase, DHEA-ST) catalyzes the sulfation of various steroids and their derivatives, including hydroxysteroids such as dehydroepiandrosterone (DHEA), androsterone (ADT), testosterone, estradiol, and many other endogenous steroids (Falany, 1997; Chen et al., 1996; Kakuta et al., 1998). Steroid sulfation has been recognized as an important process for maintaining steroid hormone levels during their metabolism. In humans, dehydroepiandrosterone sulfate (DHEAS) is the most prevalent steroid precursor, and is one of the major secretory products of both adult and fetal adrenals (Chang et al., 2004).
Substrate inhibition is a common characteristic of sulfotransferases (Sugiyama et al., 1984; Marcus et al., 1980; Raftogianis et al., 1999; Ganguly et al., 1995). The catalysis and inhibition of DHEA and ADT by SULT2A1 have been reported to regulate the homeostasis and metabolism of these compounds and to maintain steroid levels (Chang et al., 2004). Previous studies have suggested that the onset of substrate inhibition is the formation of ternary dead-end complex (Duffel and Jakoby, 1981; Zhang et al., 1998). Furthermore, other kinetic studies also indicate that the dead-end complexes of E/PAP-SO3/RO-SO3 and E/PAP/ROH can be formed
through either sequential random or ordered Bi Bi kinetic mechanisms (Zhang et al., 1998; Varin and Ibrahim, 1992; Marshall et al., 2000). This indicates that the changes of PAP affinity to sulfotransferase, for example in the different redox states for rat SULT1A1 (Marshall et al., 1997; Marshall et al., 2000), can affect substrate inhibition and enzyme activity. Furthermore, the substrate inhibition in SULT1A1 has been studied from X-ray crystal structure and compared with that of SULT1A3 which shares 93% amino acid sequence identity. The presence of two p-nitrophenol (pNP) molecules in the crystal structure of SULT1A1 explained the cooperativity and inhibition at low and high substrate concentrations, respectively (Barnett et al., 2004; Gamage et al., 2003). The stoichiometry of estradiol binding with human SULT1E1 was shown to have two ligands with one enzyme homodimer. Two binding sites were proposed as the catalytic and allosteric sites, respectively (Zhang et al., 1998). In a separate study, the crystal structure of SULT2A1 shows the presence of two orientations of DHEA in SULT2A1 (Rehse et al., 2002). There are interesting similarity and difference between SULT1A1 and SULT2A1 regarding their substrate binding orientations. However, the substrate inhibition in SULT2A1 has not been studied in detail.
Structural alignments among the complex structures of SULT2A1/ADT, SULT2A1/DHEA, and SULT2A1/PAP (Chang et al., 2004; Rehse et al., 2002; Pedersen et al., 2000) reveal spatial variation of amino acids in different substrate and PAP binding states. We herein describe experimental considerations that lead us to propose two of the amino acids, Tyr-238 and Met-137, are residues responsible in regulating substrate inhibition. Finally, we give another experimental example to demonstrate that the corresponding Tyr-238 residues of other cytosolic sulfotransferases possess the similar function as that of SULT2A1.
Materials and Methods Materials
PfuTurbo DNA polymerase was purchased from Stratagene, and MUS, MU, DHEA, ADT, PAP,
PAPS, p-nitrophenol, dopamine, glutathione (reduced form), and dithiothreitol (DTT) were purchased from Sigma (St. Louis, MO). Potassium phosphate (dibasic) was obtained from J. T. Baker (U.S.A.). DEAE Sepharose fast flow, Glutathione S-transferase Sepharose fast flow and Sephacryl S-100 HR were obtained from Amersham Pharmacia Biotech Asia Pacific (Hong Kong). All other chemicals were obtained commercially at the highest purity possible.
Methods
Site-Directed Mutagenesis of the cDNA Encoding SULT2A1, SULT1A1, and SULT1A3 Site-directed mutagenesis was performed with PfuTurbo DNA polymerase using QuickChange (Stratagene, La Jolla, CA). All primers for mutagenesis were purchased from Mission Biotech Co., Ltd. (Taiwan). Wild-type SULT2A1, SULT1A1, and SULT1A3 cDNA incorporated in the pGEX-2TK expression vector were used as templates in conjunction with specific mutagenic primers. Mutated cDNA sequences were confirmed using an ABI Prism 377 DNA sequencer (Applied Biosystems, Foster City, CA) following the standard protocol.
Expression, Purification and Characterization of Wild-type and Mutants of SULT2A1, SULT1A1, and SULT1A3
The expression and purification of SULT2A1, SULT1A1, and SULT1A3 were described previously (Sakakibara et al., 1998). Molecular weight of wild type (homodimer) and V260E mutant (monomer) of SULT2A1 was estimated by gel filtration chromatography. A homogeneous protein was obtained as determined by SDS-polyacrylamide gel electrophoresis (Laemmli, 1970).
The activities of wild-type and mutant SULT2A1 were determined according to the change of fluorescence based on a coupled-enzyme assay method (Chen et al., 2005). The fluorescence of MU at 460 nm was measured upon excitation at 355 nm. The reaction mixture with a final volume of 1 ml consisted of 100 mM potassium phosphate buffer at pH 7.0, 5 mM 2-mercaptoethanol, 20 µM PAPS, 2 mM MUS, 5.4 µg K65ER68G (Yang et al., 1996) of rat SULT1A1, SULT2A1 and 5 µM of DHEA or ADT at 37 °C. A linear response was obtained when 1.49 to 14.9 nM (0.1 to 1 µg) SULT2A1 was added in the standard assay condition. In the reaction condition, excess amount of K65ER68G of rat SULT1A1 was added to catalyze the regeneration of PAPS so that the production of PAP catalyzed by SULT2A1 could not accumulate (Chen et al., 2005). The reaction condition for SULT1A3 standard assay was similar to that for SULT2A1 assay except that the reaction mixture contained 1.5 to 7.3 nM (0.1 to 0.5 µg) SULT1A3 and 30 µM of dopamine at 37 °C to replace SULT2A1 and DHEA or ADT, respectively. The control experiments of the coupled-enzyme assay were performed in the absence of PAPS, dopamine, MUS, SULT1A3, and K65ER68G of rat SULT1A1, respectively. Only the complete reaction renders the fluorometric activity of SULT1A3. K65ER68G of rat SULT1A1 is inactive toward DHEA, ADT and dopamine in the conditions described above. Activity of SULT1A1 was determined according to the change of absorbency at 400 nm due to elimination of free p-nitrophenol (ε = 10500 M-1cm-1 at pH 7.0) as described previously (Yang et al., 1996). The reaction mixture consisted of 100 mM potassium phosphate buffer at pH 7.0, 1 to 100 nM (0.07 to 7 µg) SULT1A1, 50 µM PAPS, 5 mM 2-mercaptoethanol, and 5 µM p-nitrophenol at 37 °C.
Substrate Binding
The dissociation constants (Kd) of PAP, DHEA, ADT towards SULT2A1, respectively, were
determined with a spectrofluorimeter as described previously (Zhang et al., 1998). The decrease in intrinsic fluorescence of protein at 340 nm was observed upon excitation at 280 nm when an aliquot amount of PAP was added into the mixture consisted of 100 mM potassium phosphate buffer at pH 7.0, 100 nM wild-type or mutant SULT2A1 and varying concentrations of PAP at 25 °C with a final volume of 1.3 ml in a quartz cuvette of 1 cm square cross-section. In the formation of ternary dead-end complex, DHEA and ADT were added into the pre-incubated solution containing 0.5 µM wild type or mutants of SULT2A1, 100 mM potassium phosphate at pH 7.0, and 1 µM PAP at 25 °C. Each data point was duplicated, and the difference was within 10%.
Analysis of Kinetic Data
equation to the data. Kinetic data obtained from non-inhibitory experiments were individually fit to Michaelis-Menten Equation 1, and substrate inhibition data were fitted to substrate inhibition Equation 2 (Cornish-Bowden, 1995). The rate constants (Km, Vmax, and Ki) were obtained using
SigmaPlot 2001, V7.0 and Enzyme Kinetics Module, V1.1 (SPSS Inc., Chicago, IL). Data used represent mean values derived from two determinations.
v = V[S]/(Km + [S]) (Eq. 1)
v = V[S]/{Km + [S](1 + [S]/Ki)} (Eq. 2)
Protein Crystallization and Data Collection
Crystallization of M137I and M137W mutants was performed by the hanging-drop vapor-diffusion method at 18 °C. This condition was further refined to produce larger crystals using 2 µL hanging drops containing equal volumes of protein solution (1 µL) and a reservoir solution (1 µL) containing (NH4)2SO4 (1.6 M), NaCl (100 mM) and HEPES (0.1 M) at pH 7.5.
The crystals of M137I and M137W appeared within 3 days and grew to maximum dimensions after 7 days.
Crystallographic Data Collection and Processing
The crystals of diffraction quality were mounted on a Cryo-loop (0.1-0.2 mm), dipped briefly in 20 % glycerol as a cryo-protectant solution, and frozen in liquid nitrogen. X-ray diffraction data were collected at 110 K using the synchrotron radiation on the beamline BL13B1 at NSRRC (Hsinchu, Taiwan). The data were indexed and processed using the HKL2000 program (Otwinowski and Minor, 1997). Details of the data statistics are given in the Table 5.
Crystal Structure Determination and Refinement
The structures of the M137I and M137W were determined by molecular replacement as implemented in CNS v1.1 (Brunger et al., 1998) using the crystal structure of human dehydroepiandrosterone sulfotansferase (PDB code 1OV4) (Chang et al., 2004) as a search model. The M137I and M137W molecule was located in the asymmetry unit after rotation and translation function searches. All refinement procedures were performed using CNS v1.1. The composite omitted electron density maps with coefficients │2Fo -Fc│ were calculated and
visualized using O v11.0.4 (Jones et al., 1991), and the model was rebuilt and adjusted iteratively as required. Throughout the refinement, a random selection (10 %) of the data was placed aside as a “free data set”, and the model was refined against the rest of the data with F > 0 as a working set. The monomer protein model was initially refined by rigid-body refinement using the data from 30.0 to 3.0 Å-resolution, for which the group temperature B values were first restrained to 20 Å2. This refinement was followed by simulated annealing using a slow cooling
protocol with a starting temperature 2500 K, provided in CNS, applied to all data between 20.0 and 2.6 Å and 3.0 Å, respectively, for M137I and M137W. The bulk solvent correction was then applied, and group B factors were adjusted. After several cycles of positional and grouped B factor refinement interspersed with interactive modeling, the R-factors for the M137I and M137W complex decreased. The refinement then proceeded with another cycle of simulated annealing with a slow cooling with a starting temperature of 1000 K.
Model Validation
The correctness of stereochemistry of the model was verified using PROCHECK (Laskowski et al., 1993). The calculations of r.m.s. deviations from ideality for bonds, angles, and dihedral and improper angles performed in CNS showed satisfactory stereochemistry. In a Ramachandran plot (Ramachandran and Sasisekharan, 1968), all of main chain dihedral angles were in the most favored and additionally allowed regions. The refinement statistics are given in the Table 5. Coordinates
Atomic coordinates for the crystal structures of M137I and M137W mutants described in this work have been deposited in the PDB (access code 2QP3 and 2QP4, respectively).
Results
Predicting Critical Amino Acid Residues for Substrate Inhibition by the Comparison of the
Structure of SULT2A1 Complexes—Comparison between the SULT2A1/DHEA and
SULT2A1/PAP binary complex structures may reveal the alternative locations of the main regulatory amino acid residues when substrate is present or absent in the active site. The overall structure of the SULT2A1/PAP complex is very similar to that of the SULT2A1/DHEA complex, except for some residues and a loop from residues Tyr-231 to Gly-252 (Table 1). The root-mean-square deviation (RMSD) value of this loop between the two structures is 6.67 Å for the backbone (excluding Lys-242 and Thr-243, which were not resolved in the SULT2A1/PAP binary complex structure). As shown in Figure 1A, the main difference near the DHEA binding site between the two complexes is that the loop from residues Tyr-231 to Tyr-238 is closed in SULT2A1/PAP, while it is open in SULT2A1/DHEA (Rehse et al., 2002; Pedersen et al., 2000). Furthermore, in the SULT2A1/DHEA complex, several hydrophobic residues on this loop, including Tyr-231, Leu-234, and Tyr-238, are located in close proximity to the DHEA molecule; these residues contribute to the hydrophobic nature of the active site. Also, in the SULT2A1/DHEA binary complex structure, Tyr-238 acts likely as the gate toward the substrate-binding cavity, but moves away from the active site in the SULT2A1/PAP binary
complex structure (Figure 1A). It is therefore reasonable to speculate that Tyr-238 may play a critical role in regulating the release of substrate. As shown in Figure 1B, the ADT molecule shares the same location as that of the DHEA molecule in the previously proposed alternative orientation (Rehse et al., 2002). The SULT2A1/DHEA and SULT2A1/ADT complex structures are quite similar at this region, including the loop described above, and the location of Tyr-238 residue.
Kinetic Analysis of Tyr-238 Mutants of SULT2A1— The hypothesized gate residue of
Tyr-238 for regulating substrate inhibition, as revealed by the above structural comparison, was confirmed by site-directed mutagenesis. Vmax, Km and Ki values of wild type and Tyr-238 mutants
(Y238A, Y238F, and Y238W of SULT2A1) were determined using DHEA or ADT as substrates (Table 2). Y238A of SULT2A1 exhibited no substrate inhibition when ADT was used as substrate and the Ki value of DHEA increased 7.4 folds as compared to that of wild type
SULT2A1. The Ki values of DHEA and ADT for Y238F and Y238W of SULT2A1 also
increased for about 2 to 3 folds as compared to those of wild type. The changes of Km and Vmax
for Y238A, Y238F and Y238W ranged only from approximately 0.3 to 1.3 folds whenever DHEA or ADT was used as substrate (Table 2). This observation, consistent with the above prediction base on structural analysis, showed that Tyr-238 is regulating the substrate inhibition and demonstrated that the size of this amino acid side chain is important.
Predicting Amino Acid Residue Delineates the Orientations of DHEA and ADT in
SULT2A1—Mutating Tyr-238 to alanine completely and partially eliminate the substrate
inhibition for ADT and DHEA, respectively (Table 2), indicating that the additional amino acid(s) may be interacting with these two substrates in a different way since a single mutation (Tyr-238) is not capable of completely eliminating inhibition of both substrates. The complex structures of SULT2A1/DHEA and SULT2A1/ADT revealed two orientations for DHEA but only one for ADT as described earlier (Chang et al., 2004). Based on the available information, we might infer that the additional amino acid(s) that affect the substrate inhibition of DHEA may also affect its binding orientation in SULT2A1. To search for the possible amino acid(s), we superimposed the structures of SULT2A1/DHEA and SULT2A1/ADT on each other. Most of the amino acid residues hold the same positions in two complex structures, except for the side chains of Met-16, Ile-71, and Met-137 whose RMSD values (2.41, 2.50, and 2.40 Å, respectively) are noticeably higher than those of the others (Figure 2). Both Met-16 and Ile-71 of the SULT2A1/DHEA and SULT2A1/ADT complex structures form similar van der Waals interactions toward ADT and DHEA in the two orientations. By contrast, the Cε atom of the
Met-137 residue in SULT2A1/ADT complex structure points inward the substrate binding site and causes steric hindrance for ADT and prevents it from forming additional orientation observed only for DHEA. The shortest distance would have been only 1.11 Å between C-12 of
DHEA at this additional orientation and the Cε atom of Met-137 in SULT2A1/ADT complex
(Figure 2). From the structural analysis between SULT2A1/DHEA and SULT2A1/ADT complexes, it is reasonable to propose that, in addition to Tyr-238, Met 137 may serve as the amino acid that modulates the substrate inhibition and binding orientations of DHEA and ADT.
Kinetic Analysis of Met-137 and Met-137/Tyr-238 Mutants of SULT2A1—The proposed
effects of Met-137 on substrate inhibition were examined by mutational analysis of SULT2A1 at Met-137 and Tyr-238. When Met-137 of SULT2A1 was mutated to isoleucine (M137I) and valine (M137V) which contain smaller side chains, Ki values for ADT increase over one order of
magnitude (28.6 folds for M137I and 11.1 folds for M137V) (Table 2). The Ki value increases
only 5.4 folds when Met-137 was mutated to larger side chain, tryptophan (M137W). Changes of Ki values for DHEA were in a similar trend to those of ADT with smaller increases (5.9, 8.7, and
1.2 folds for M137I, M137V, and M137W, respectively) (Table 2). M137K was also prepared and exhibited no enzymatic activity, most probably due to the positive charge of lysine that interferes with the hydrophobicity of the binding site.
These observations are consistent with the above structural analysis that Met-137 side chain is a steric hindrance for ADT in SULT2A1/ADT complex but not for DHEA in SULT2A1/DHEA complex (Figure 2). By removing this steric hindrance, additional binding orientation for ADT may form and result in considerable changes (over one order of magnitude) of their Ki values
(M137I and M137V in Table 2). This proposal also agrees with our finding that the orientation of ADT may be disturbed when Met-137 is replaced with a smaller isoleucine (M137I/Y238A in Table 2) so that the substrate inhibition of ADT with M137I/Y238A can be observed. In contrast, by replacing a large side chain at Met-137 (M137W), additional steric hindrance may be created and one of the binding orientations of DHEA may be removed. This also predicts that, with additional mutation on Y238A at Met-137, the substrate inhibition for DHEA may be completely eliminated. Double mutations on SULT2A1 at Met-137 and Tyr-238 confirm our hypothesis (M137W/Y238A in Table 2). Two other double mutants, M137V/Y238A and M137K/Y238A (Table 2), also exhibited expected properties. The Ki values obtained with M137V/Y238A were
between those obtained with M137I/Y238A and M137W/Y238A. M137K/Y238A still exhibited no enzymatic activity as that of M137K.
Comparison of the Crystal Structure of Wild-type, M137I, and M137W of SULT2A1—The
data discussed above indicate that the additional substrate binding orientation modulated by Met-137 may be responsible for the substrate inhibition observed in Y238A only for DHEA but not for ADT (Table 2). The crystal structures of M137I and M137W in this study were determined and showed that the each global structure was similar to that of the wild-type, either the SULT2A1/DHEA or SULT2A1/ADT complex structure. In Figure 3, the structures of the residue Met-137 in SULT2A1 were compared to those of isoleucine (M137I) and tryptophan
(M137W) in SULT2A1 mutants. Replacing Met-137 with tryptophan (M137W) obviously contributes to steric hindrance and prevents the formation of the additional orientation of DHEA. This structural information is consistent with our hypothesis that the additional orientation of DHEA is responsible for the substrate inhibition in Y238A. As shown in Table 2, the DHEA substrate inhibition in Y238A can be eliminated in M137W/Y238A. On the contrary, replacing Met-137 with isoleucine (M137I) may remove the steric hindrance and provides a space for the formation of additional orientation for ADT in SULT2A1 (Figure 3). As shown in Table 2, the ADT substrate inhibition of Y238A can be regenerated with M137I/Y238A. Combination of the structural and kinetic data strongly supports our hypothesis that both Tyr-238 and Met-137 of SULT2A1 are responsible for the regulation of substrate inhibition as described above.
Dissociation Constants of PAP, DHEA and ADT in Binary and Ternary Complex Structures of SULT2A1 and Its Mutants—It is important to show that this ternary complex is intact in SULT2A1 mutants (Y238A, M137I, M137W, M137I/Y238A and M137W/Y238A) to support our proposal that Met-137 and Tyr-238 directly modulate substrate inhibition. As shown in Table 3, the dissociation constants of PAP of SULT2A1/PAP binary complexes are the same among wild type and all the mutants. Tight binding of PAP (in nM range as shown in Table 3) to the SULT2A1 and its mutants indicates that PAP binding site remains intact following such mutations. This information excludes the possibility that the change of Ki values reported in this
study is due to the change of PAP binding to SULT2A1 mutants.
Affinity of either DHEA or ADT to SULT2A1/PAP binary complex was noticeably decreased whenever there is an Y238A mutant (Y238A, M137I/Y238A, and M137W/Y238A) as compared to that with wild-type SULT2A1 (Table 3). Except for the DHEA with M137I, mutation at Met-137 (M137I or M137W) did not significantly affect the binding of either DHEA or ADT to the SULT2A1/PAP binary complex. The dissociation constants of ligands (PAP, DHEA and ADT) and the double mutants of SULT2A1, M137I/Y238A and M137W/Y238A, are very similar to those with single mutant, Y238A (Table 3). These observations are consistent with the proposed function of Tyr-238 and Met-137 that the former acts to prevent the release of bound substrate while the later modulates the orientations of bound substrate in the substrate inhibition mode.
Substrate Inhibition in SULT2A1 Monomer—To examine the previous hypothesis that
dimerization results in substrate blocking, the monomer mutant (V260E) of SULT2A1, designated in the KTVE motif (Petrotchenko et al., 2001) along with a double mutant, Y238A/V260E (Table 2), were constructed to comprehend the substrate inhibition of SULT2A1. The wild-type SULT2A1 (homodimer) was found to have similar kinetic constants as those of the mutant monomer. Furthermore, the pattern of substrate inhibition in monomer mutant (V260E in Table 2) is the same as that of wild-type SULT2A1. The mutation of monomer
mutant at Tyr-238 (Y238A/V260E in Table 2) also gives exactly the same substrate inhibition pattern as that of Y238A of SULT2A1 (Table 2). Data obtained from this study indicates that dimer or monomer of SULT2A1 does not play an important role for the exhibition of substrate inhibition.
Functional Analysis of Tyr-238 and Met-137 Corresponding Residues in Other
Sulfotransferases—The multiple sequence and structure alignments of some solved-structure
sulfotransferases are shown in Figure 4. In upper left of Figure 4, the residues corresponding to Tyr-238 of SULT2A1 have been highlighted, and they are as follows: Phe-247 of human SULT1A1, Leu-247 of human SULT1A3, Met-247 of mouse SULT1E1, Leu-249 of human SULT2B1_v1 (Gamage et al., 2003; Lu et al., 2005; Kakuta et al., 1997; Lee et al., 2003). This comparison leads us to hypothesize that the Tyr-238 corresponding residue in other sulfotransferases may play a similar role in substrate inhibition because these residues all demonstrate possible steric hindrance for the release of substrate from the substrate-binding cavity, and therefore they may modulate substrate inhibition for their own preferred substrates. To characterize the function of the SULT2A1 Tyr-238 corresponding residues in other sulfotransferases, further mutational analysis was conducted with SULT1A1 and SULT1A3 at Phe-247 and Leu-247 residue, respectively (Table 4). As compared to those of wild type SULT1A1, the Ki value of F247A (with p-nitrophenol as substrate) increased approximately
12-fold, the Vmax/Km value increased approximately 3-fold, while Km remains pretty much the
same. The Ki value of dopamine for SULT1A3 is much higher than that of pNP for SULT1A1
(Table 4). The positively charged dopamine at the assay condition may contribute to this difference. Mutations at Leu-247 of SULT1A3 give Ki values comparable with the size of the
side chain of the mutated amino acid (the larger the amino acid side chain, the lower the Ki value)
as shown in Table 4 for L247A and L247Y. The data from these investigations support our hypotheses that substrate inhibition in SULT2A1 is modulated by Tyr-238, and that analogous residues of Tyr-238 in other cytosolic sulfotransferases play similar roles with respect to substrate inhibition.
Discussion
Recently, the study in structure and function of enzymes has been greatly facilitated as a result of an expansion by means of their X-ray crystal structures. Crystal structures of SULT2A1 containing ADT, DHEA, and PAP, respectively, have been previously reported (Chang et al., 2004; Rehse et al., 2002; Pedersen et al., 2000). DHEA has been found to have two binding orientations in SULT2A1 (Rehse et al., 2002), however, bound ADT possesses only one, in which it is flipped over along the long axis of the DHEA relative to the proposed alternative
orientation (Chang et al., 2004). The previously proposed alternative orientation of DHEA was speculated to be a substrate-inhibition orientation owing to the fact that it placed O-3 hydroxyl group of DHEA with respect to His-99 for 2.9 Å away, and closer to the modeled PAP. Furthermore, it contained more van der Waals interactions with hydrophobic residues than the proposed catalytic orientation (Rehse et al., 2002). Prior study (Gamage et al., 2005) however, suggested that this would not result in substrate inhibition because this would require a relative increase in the proportion of the proposed alternative orientation while the substrate concentration increases. It might infer that both the orientations of DHEA could cause substrate inhibition while PAP remained in the active site to form ternary dead-end complex. In this report, we identified two amino acids (Met-137 and Tyr-238) responsible for modulating substrate inhibition of DHEA and ADT and were able to provide experimental evidences to show that both substrate binding orientations indeed cause substrate inhibition.
The bound orientations observed for DHEA and ADT might be in the substrate inhibition mode because the crystals were usually formed under high concentration of ligands although PAP was not present in the solved crystal structures of SULT2A1/DHEA and SULT2A1/ADT (Chang et al., 2004; Rehse et al., 2002). Dissociation constants of DHEA for SULT2A1 and its mutants at Tyr-238 and Met-137 (Table 3) also indicate that substrate inhibition may be induced by two possible binding orientations for DHEA because the substrate inhibition could be eliminated only after two specific mutations (M137W/Y238A in Table 2). According to the crystal structures of SULT2A1 (Figure 1B), Tyr-238 has weak interactions with ADT and the closest distance between Tyr-238 and the substrates is about 4.6 Å. Structural analyses among SULT2A1 complexes (Figure 1A and 1B) strongly suggest that Tyr-238 acts as a gate residue to regulate the release of substrate from the substrate-binding cavity in a ternary dead-end complex. The dissociation constants of ternary dead-end complex shown in Table 3 also support this implication. Kd values of both DHEA and ADT increase significantly when Tyr-238 was mutated
to alanine. However, mutation of Tyr-238 alone can not completely eliminate DHEA substrate inhibition (Y238A in Table 2) indicating the existence of an additional substrate inhibition mode for DHEA but not for ADT. Structural analysis showed that Met-137 is closely interacting with DHEA and ADT and may modulate the substrate binding orientation (Figure 2). This scheme agrees with the experimental data that further mutation on Y238A at Met-137 not only can completely eliminate substrate inhibition for DHEA (M137W/Y238A in Table 2) but can also create substrate inhibition for ADT (M137I/Y238A in Table 2). These data strongly suggest that DHEA exists in only one orientation in M137W/Y238A and ADT presents in two orientations in M137I/Y238A. Comparison of the solved crystal structures shown in Figure 3 gives additional support for the proposed substrate binding orientations in these mutants.
substrates with two binding orientations. It has been proposed that substrate inhibition in SULT1A1 by estradiol could occur from the mis-orientation of substrate in the complex (Gamage et al., 2005). The presence of two pNP molecules in the crystal structure of SULT1A1 was postulated to explain cooperativity at low and inhibition at high substrate concentrations, respectively (Barnett et al., 2004). For SULT2A1, the Ki values shown in Table 2 indicate that
Met-137 and Tyr-238 may modulate substrate inhibition at low and high Ki values, respectively.
For ADT, the Ki value is significantly increased when the additional substrate binding orientation
is proposed to present in the enzyme (M137I and M137I/Y238A in Table 2). The binding of substrates to Y238A is significantly loosened with PAP and enzyme complex (Table 3). This may either totally eliminate substrate inhibition (for ADT) or significantly decrease the substrate inhibition contribution from the substrate bound at the additional orientation (for DHEA). This scheme is consistent with the Ki value described previously for the reduced form of rat SULT1A1
that Ki isproposed to be determined by the binding constant of substrate and PAP/enzyme binary
complex (Marshall et al., 2000).
Other possible causes for substrate inhibition proposed previously (Petrotchenko et al., 2001) were also examined in this study. In a previous study on SULT2A1/PAP complex structure (Pedersen et al., 2000), the loop of residues 231-252 shown in Figure 1A was speculated to prevent substrate binding while being involved in the dimer interface. This implies that the dimer may be an inactive form, with the second subunit of the dimer contributing to the locking in the substrate-blocking loop. Dimerization of SULT2A1 is not important for the substrate inhibition for exactly the same substrate inhibition patterns were observed for enzyme monomer, dimer and their mutants (Table 2).
The cause of sulfotransferase substrate inhibition has been attributed to the formation of an enzyme, substrate and PAP ternary complex (Duffel and Jakoby, 1981; Zhang et al., 1998). The binding of PAP is critical for the formation of the ternary complex (Hsiao and Yang, 2002) and it has been shown that affinity of PAP had profound effects on the activity of sulfotransferases (Marshall et al., 2000; Yang et al., 1996). Binding of PAP may significantly affect the substrate binding (Hsiao and Yang, 2002) and in turn affect substrate inhibition. This indicates that the changes of PAP affinity to sulfotransferase can affect substrate inhibition and enzyme activity. For example, rat SULT1A1 has been shown to significantly alter the transfer and physiological reactions in various redox states (Marshall et al., 1997). Mutation at other amino acid may also alter the PAP affinity to sulfotransferase (Hsiao and Yang, 2002; Yang et al., 1996). Mutation at Met-137 and Tyr-238 did not affect the nucleotide binding affinity to SULT2A1 (Table 3). In addition, the variations of the dissociation constants of DHEA and ADT in Tyr-238 and Met-137 mutants(Table 3) is consistent with what would be expected for the proposed functions of these two amino acids. Tyr-238 is proposed to modulate the release of bound substrate and can