腫瘤壞死因子-alpha 與MPP+ 離子引發多巴胺神經元細胞死亡之機制探討; Mechanism of dopaminergic cell death induced by MPP+ ions and tumor necrosis factor-alpha
65
0
0
全文
(2) 致謝 首先感謝恩師林靜茹老師在學業研究上悉心指導,使本論文撰寫 得以順利完成。文稿始成之初,承蒙弘光科技大學楊齊生老師、中山 醫學大學盧敏吉老師以及本校李妙蓉老師和王慧如老師等口試委 員,在論文上的指導與建議,提供諸多寶貴意見,始論文內容得以更 臻完善,在此至上衷心的謝意。 回顧兩年研究期間,承蒙生理學科蔡主任宗統老師、陳卓昇老 師、王慧如老師、許明志老師、林靜茹老師、黃怡萍老師、龔志力老 師、閔明源老師、盧敏吉老師以及劉惠華小姐在研究及生活上的指導 及關心。此外感謝生理科怡均及其他同學在課業上和實驗上的幫助以 及慧淇學姊、文玲學妹在生活上的砥礪與幫助。更由衷感謝我的母親 及朋友們這兩年來的陪伴與鼓勵,最後僅將此論文所有關心、鼓勵和 協助我的師長、同學、家人以及朋友。. 吳曉雲謹致於 中國醫藥大學醫學研究所 中華民國九十四年七月二十七日. 2.
(3) 中文摘要 過去有研究指出腫瘤壞死因子-alpha(tumor necrosis factor-alpha; TNF-α)與帕金森氏症有關,然而,TNF-α 對於神經細胞是保護或 是傷害的作用及其機制仍是不明的。本實驗欲探討 TNF-α 在帕金森 氏症疾病中所扮演的角色,因此,我們以 1-methyl-4-phenyl-pyridinium +. +. +. (MPP ions;MPP )單獨處理及 MPP 合併 TNF-α 作用於 PC-12 細 +. 胞上,作為研究模式。PC-12 細胞給予不同濃度的 MPP (600,300, 150,75 或 0μM)或是合併 TNF-α(15 或 30 ng/ml)處理,至 24 小 +. 時後,再以 MTT Assay 分析存活率。MTT Assay 結果顯示 MPP 處理 +. 可以引發細胞死亡,且死亡率隨著劑量增加而提昇。且 MPP 合併 TNF-α 處理,可以引發更高的細胞死亡率。在細胞形態上,我們也 看到了相似的結果。另外,以 DAPI 染色的結果證明,此一細胞死亡 現象屬細胞凋亡。我們進一步探討此一細胞凋亡現象可能之機制,從 +. 西方墨點法的結果得知,MPP 處理可以引發 caspase-3 活化及 JNK 磷 +. 酸化,而且當 MPP 合併 TNF-α 處理時可以引發更多的 caspase-3 活 化及 JNK 磷酸化。但是磷酸化 c-Jun(p-c-Jun)的表現量並未有增加 的趨勢。我們進一步以 kinase assay 偵測磷酸化 JNK 的活性,結果發 +. 現 MPP 合併 TNF-α 處理引起 JNK 活性上升。接著,為了探討 caspase-3 +. 及 JNK 是否為 MPP 合併 TNF-α 處理導致細胞死亡的主要因子,我 3.
(4) 們使用 caspase-3 及 JNK 專一性抑制劑評估細胞凋亡的情形。由 TUNEL +. 實驗結果證實,抑制 caspase-3 可以有效緩解 MPP 合併 TNF-α 處理 +. 導致的細胞死亡;然而抑制 JNK 只能輕微緩解 MPP 合併 TNF-α 處 理導致的細胞死亡。 +. 從以上實驗結果得知,MPP 會經由活化 caspase-3 而誘導細胞凋 +. 亡,而 TNF-α 加劇 MPP 之毒性和提昇 caspase-3 活性有關。而且此 +. 一神經毒素引起可經由活化而導致細胞凋亡。MPP 合併 TNF-α 處理 也能增加 JNK 磷酸化,但此一 JNK 的磷酸化的功能並不全然與細胞 凋亡有關,而其詳細功能仍待進一步的實驗調查。. 4.
(5) 英文摘要 Recent studies indicate tumor necrosis factor-alpha(TNF-α)is associated with Parkinson's disease(PD). However whether TNF-α plays a neuron protective or neuron destructive role remains to be elucidated. addition, the mechanism is still not clear.. In. To determine the role of TNF-α. in PD, we studied neurotoxic effect on dopaminergic neurons(PC-12 cells) induced by 1-methyl-4-phenyl-pyridinium(MPP presence or absence of TNF-α.. +. +. ions;MPP )in the. PC-12 cells were either treated with MPP. +. alone(600, 300, 150, 75 and 0μM) or in combination with TNF-α(15 or 30 ng/ml) for 24 hours and then assayed for cell viability.. MTT Assay. +. showed MPP treatment increased cell death in a dose-dependent manner. However, death rates were statistically higher in MPP +. than those treated with either MPP or TNF-α only. also observed similar trends.. plus TNF-α groups Morphologically, we. In addition, results from DAPI staining. indicated the cell death was apoptotic. signal molecules involved.. +. We further investigated possible. Data from western blot analysis showed that. caspase-3 and JNK activation increased when treated with MPP the increases were even higher in MPP. +. +. alone; and. plus TNF-α treated cells.. However, the amount of phosphorylated c-Jun was not elevated in all treated 5.
(6) groups.. We then detected the activation of JNK.. showed JNK activity increased clearly in MPP. +. Data from kinase assay. plus TNF-α treatment cells.. We later used Z-VAD (a caspase inhibitor) and a JNK inhibitor (SP600125) to determine whether blockade of caspase-3 and JNK activation can inhibit apoptosis.. Data from TUNEL assay demonstrated. caspase inhibition prevented the cell death efficiently. However, JNK inhibition only slightly prevented the cell death slightly. +. Our data suggest that MPP caused cell apoptosis through caspase-3 activation, and the enhanced toxicity induced by the addition of TNF-α is aiso related to caspase-3 activation.. In addition, JNK activation was. involved in the intricate signaling pathways.. Nevertheless, JNK activation. may partially involved in the induction of apoptosis.. Further investigation. will be needed to elucidate the role of JNK activation in this apoptosis.. 6.
(7) 目錄 致謝……………………………………………………………………2 中文摘要………………………………………………………………3 英文摘要………………………………………………………………5 目錄……………………………………………………………………7 壹、前言………………………………………………………………9 貳、研究目的…………………………………………………………19 参、實驗材料與方法…………………………………………………20 肆、結果………………………………………………………………30 伍、討論………………………………………………………………41 陸、參考文獻…………………………………………………………45 柒、實驗結果附圖……………………………………………………51. 7.
(8) 圖目錄 +. 圖 A、MPP 代謝路徑…………………………………………………14 +. 圖 B、MPP 進入細胞後對細胞產生的影響……………………………16 +. 圖 C、MPP 對細胞內訊息傳遞分子的影響…………………………17 +. +. 圖一、TNF-α 合併 MPP 處理可加重 MPP 的毒性…………………51 圖二、不同處理時間點引發細胞形態改變……………………………52 + 圖三、TNF-α 可加劇 MPP 引發的細胞死亡…………………………55. +. +. 圖四、MPP 及 TNF-α 合併 MPP 處理引發細胞凋亡 ………………56 +. 圖五、MPP 處理可引發 cytochrome-c 釋放……………………………57 +. +. 圖六、MPP 及 MPP 合併 TNF-α 處理可導致 caspase-3 活化………58 +. +. 圖七、MPP 及 MPP 合併 TNF-α 處理可導致 JNK 磷酸化…………59 +. +. 圖八、MPP 及 MPP 合併 TNF-α 處理不導致 c-Jun 磷酸化…………60 +. 圖九、MPP 合併 TNF-α 處理可增加 JNK 活性………………………61 +. +. 圖十、抑制 caspase 活化可阻止 MPP 及 MPP 合併 TNF-α 導致的 PC-12 細胞形態改變……………………………………………62 +. 圖十一、抑制 caspase 可緩解 MPP 合併 TNF-α 引發的細胞凋亡……64 圖十二、TUNEL Assay 結果量化圖表…………………………………65. 8.
(9) 壹、前言 帕金森氏症 (Parkinson,s disease;PD) 帕金森氏症的名稱由來,是源自於一位名為詹姆士•帕金森 (James. Parkinson) 的英國醫生,在西元 1817 年首度描述了這個病症, 後人便將他當時所發表的「震顫麻痺」(the shaking palsy)稱作帕金 森氏症。該病好發率主要集中於中高年齡族群。在台灣,年紀四十歲 以上人口,帕金森氏症的盛行率為每十萬人中有 358 人,而疾病發生 率為每十萬人中有 28.7 人。 偶發性帕金森氏症(sporadic PD)佔所有帕金森氏症病人的 90% 以上,這種家族性的 PD 被懷疑可能與基因突變有關,例如:. α- synuclein 及 parkin;自發性帕金森氏症(idiopathic PD)通常發生 於 55 歲以上的年齡族群,病程可持續 10-20 年,且目前自發性帕金森 氏症的病因仍屬不明(Gao et al., 2002)。 帕金森氏症病人之臨床症狀表現通常是合併出現的運動問題,諸 如:動作遲緩(bradykinesia)、靜止時震顫(resting tremor)或震顫 (tremor)、肌肉僵硬 (Fahn and Sulzer, 2004)、以及運動不能 (Akinesia)(Guyton and Hall, 2002)。臨床上患者並非每個症狀都會 出現,但通常只要出現兩個以上之症狀,並且其中包含動作遲緩或是 靜止時震顫之一時,即可被診斷為帕金森氏症。帕金森氏症患者通常 9.
(10) 最先會出現的症狀是「靜止時震顫」,但有些時候亦有可能是從「動 作遲緩」症狀開始;但是也有部份病人可能不會有震顫的症狀發生 (Fahn and Sulzer, 2004)。 帕金森氏症是一種常見的基底核神經病變。基底核是指位於每一 個大腦半球內的灰質體集合(Snell, 2003),擁有許多運動控制的功 能(Guyton and Hall, 2002)。帕金森氏症的病因主要是在腦內黑質緻 密區(substantia nigra)處的多巴胺神經元(dopaminergic neuron)發生 漸行性退化,而導致在紋狀體(striatum)的多巴胺(dopamine)減少 (Schmidt and Ferger, 2001;Guyton and Hall, 2002)。研究中發現,帕 金森氏症病人腦黑質中多巴胺神經元比同年紀者減少了 60~86 %並且 被核(putamen)區的多巴胺神經纖維末梢亦減少了 90~99%(Gao et al., 2002)。 雖然目前引發帕金森氏症的原因仍然不明,但是一般認為可能與 週遭生活環境有關,例如:水質不良或是暴露在具有農藥或殺蟲劑存 在的環境下等等,都可能是引起自發性帕金森氏症的危險因子 (Gao et al., 2002)。目前亦有許多證據顯示,氧化壓力增加、自由基傷害 及神經膠樣變性(Gliosis)、慢性炎症反應、基因缺損及細胞粒線體 功能喪失等,都與黑質細胞的死亡以及帕金森氏症有關(McGeer and McGeer, 2004;Sriram et al., 2002)。. 10.
(11) 依據研究發現,帕金森氏症病人腦黑質區域中的微小膠細胞 (microglia)被活化,且分泌的TNF-α、干擾素-gamma(interferon-γ) 及淋巴素-1 beta(interleukin-1β)都有增加現象。這些被誘發的細胞 激素 (cytokine),可以誘導iNOS (inducible nitric oxide synthase)增 加,接著釋放NO(nitric oxide),使帕金森氏症病人腦部的氧化壓力 增加,導致細胞死亡(Rousselet et al., 2002)。MPTP引發多巴胺神經 元的死亡率在TNF-α基因缺損的剔除小鼠(knout out mice)中減少了 65%(Ferger et al., 2004)。亦有研究顯示,水楊酸類的抗炎症藥物可 +. 以緩解 MPP 引發的細胞死亡(Sairam et al., 2003);但是部份學者卻 認為NSAIDs(non-steroid anti-inflammatory drugs)藥物,例如: +. indomethacin、ibuprofen、ketoprofen與diclofenac 將會使MPP 引起的細 胞死亡率增加(Morioka et al., 2004)。這些研究結果顯示炎症反應在 PD中扮的角色仍未明確。. 腫瘤壞死因子-alpha(Tumor necrosis factor-alpha;TNF-α) TNF-α是一種多重功能的細胞激素,會誘發許多細胞內的反應。 TNF-α在正常腦組織的含量是很低的(Soeda et al., 2001),但在發炎 時或免疫反應被激發時,就會被誘發增加。在臨床上發現一些炎症疾 病,例如:類風濕關節炎(rheumatoid arthritis;RA)、炎症性腸症候. 11.
(12) 群(inflammatory bowel syndrome)會誘發細胞激素的增加(McGeer and McGeer, 2004)。在中樞神經系統,細胞激素可以促使炎症反應發生, 並且直接對細胞產生毒性反應而使細胞傾向死亡(Rousselet et al., 2002)或是藉由ROS(reactive oxygen species)的生成,使細胞粒線體 功能喪失進而導致細胞死亡(Sriram et al., 2002)。另外,細胞激素也 可能藉由抑制免疫反應並且促使細胞生長(Rousselet et al., 2002)。 上述這些與免疫相關的機制與許多中樞神經疾病如:多發性硬化 症(multiple sclerosis)、後天免疫不全症候群併發痴呆症候群 ,. (AIDS-dementia complex)、阿茲海默氏症(Alzheimer s disease)及 帕金森氏症等有關(Ferger et al., 2004)。 有研究證實,予以毒素(Sriram et al., 2002;Rousselet et al., 2002)、 缺氧、低血糖刺激、損傷(injury)(Rousselet et al., 2002)或是中風 (stroke)後(Sriram et al., 2002),TNF-α會被誘發且對神經細胞產 生毒性,然而在β-amyloid毒性刺激及癲癇(epilepsy)研究中卻發現 TNF-α具有神經保護的作用(Ferger et al., 2004)。 在帕金森氏症的研究中發現,MPTP引發多巴胺神經元的死亡在 TNF-α基因缺損的實驗鼠中減少了65%(Ferger et al., 2004);但是在 其它的研究中卻發現TNF-α基因缺損的剔除小鼠對於MPTP引發多巴 胺神經元的死亡並無影響(Rousselet et al., 2002)。因此,TNF-α在. 12.
(13) 神經細胞凋亡過程中雖然有著重要的影響(Soeda et al., 2001),但是 TNF-α對於神經細胞是保護或是促使死亡的角色仍然不甚清楚 (Ferger et al., 2004)。. 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine(MPTP) MPTP是一種存在海洛因(heroin)內的化學成分(Chun et al., 2001)。最初是在1979-1982年間被發現,當時在美國加利福尼亞州 (California)一群使用非法藥物的年輕人出現了類似帕金森氏症的症 狀:動作遲緩、肌肉的僵直、靜止時震顫,而這些症狀可被治療帕金 森氏症的藥物所緩解(Eberhardt and Schulz, 2003),之後發現這些海 洛因當中含有2.9% MPTP成分(Nicotra and Parvez, 2000)。 MPTP毒素會使得腦黑質內的多巴胺神經元喪失及紋狀體 (striatum)內的多巴胺含量減少。在人類、非人類靈長類動物及老鼠 的研究中發現,MPTP可引發帕金森氏症(Eberhardt and Schulz, 2003)。 因此,目前MPTP被廣泛利用於誘發帕金森氏症的動物模式,藉以研 究此病。 MPTP 屬於一種脂溶性物質(Prezedborski and Vila, 2003) ,本身對 腦部並沒有毒性(Schmidt and Ferger, 2001),但是 MPTP 會快速通過 腦血管障壁(blood-brain barrier;BBB),接著會被神經膠細胞中的單. 13.
(14) 胺氧化酶 B(momoamine oxidase B;MAOB)代謝成 MPDP. +. (1-methyl-4-phenyl-1,2,3-dihydropyridinium) ;之後可能是經由自我氧化 +. +. 作用而形成 MPP (1-methyl-4-phenypyridinium),MPP 即是一個具活 +. 性的毒素。(Prezedborski and Vila, 2003)。具毒性的 MPP 自神經膠細 胞內藉由單胺運輸器(monoamine transporter)被釋放至細胞外(Russ et al., 1996) ,然後再經由多巴胺運輸器(dopamine transporter;DAT)進 入多巴胺神經元(Schmidt and Ferger, 2001)。如圖 A 所示:. Blood Brain Barrier. MPTP. (MAO-B). Glial cells. MPDP+ MPP+. MPP+. Dopamine neuron. MPP+. DAT. MPP+. (圖 A:MPP+ 的代謝路徑;摘錄自 Prezedborski and Vila, 2003)。. 14.
(15) MPTP 毒性 +. MPTP或MPP 在動物實驗及各種不同的神經細胞株的研究已被 證實可導致如: primary cortical neuron (Sun and Chang., 2003)、 SH-SY5Y、MN9D(Nicotra and Parvez, 2002) 、PC-12(Park et al., 2003) +. 等細胞死亡。MPP 一旦進入細胞內之後除了與vesicular monoamine transporter (VMAT)結合,而進入vesicle,或是破壞粒線體中電子傳 遞鏈的complex I而傷害神經細胞之外,也可能藉由改變細胞內酵素 (enzyme)的功能而影響細胞存活(Prezedborski and Vila, 2003) 。如16 頁圖B所示。 +. 目前研究也已證實,MPP 導致的細胞死亡確實與許多細胞內訊 息傳遞有關,例如:Bcl-2、Bax、p53 (Nicotra and Parvez, 2002;Eberhardt and Schulz, 2003) 、JNK (Hunot et al., 2004) 、NF-κB (Cassarino et al., 2000)及caspases (Viswanath et al., 2001)等。其作用如第17頁圖C所 示。. JNK(c-Jun N-terminal kinases) JNK、ERK和p38合稱MAPK(mitogen-activated protein kinase)。其 中,JNK是一種SAPK(stress-activated protein kinase) (Johnson and Lapadat, 2002) ,並包含了三種類型:JNK1、JNK2及JNK3(Waetzig and Herdegen,. 15.
(16) 2004)。. Enzymes Mitochondria. MPP+ Synaptic vesicle. MPP+. MPP+ VMAT. DAT. MPP+ (圖B:MPP+ 進入細胞後對細胞產生的影響;摘錄自Prezedborski and Vila, 2003)。. JNK家族會被許多刺激所活化,例如:紫外線(ultraviolet rays)、 神經毒素及TNF-α(Tournier et al., 2000;Chun et al., 2001;Park et al., 2003) 。當JNK被活化後即可繼續活化其下游的受質(substrate)例如: c-Jun(Johnson, Lapadat et al., 2002)、Elk1、ATF2(Sugden and Clerk, 1998)、Bim及Bcl-2家族(Okuno et al., 2004)。 JNK目前被許多學者認為與控制細胞的死亡或凋亡有關。有研究 證明,抑制JNK活性可以增加化學治療藥物抑制腫瘤細胞生長的能力 (Johnson and Lapadat, 2002) 。有其他研究證明紫外線引發細胞凋亡的. 16.
(17) MPP+ DAT. MPP+ JNK pathway. Bax Bim. Oxidative stress sec. excitotoxicity. Oxidative phosphorylation Mitochondria. JNK pathway Cytochrome-c. ATP↓. Caspase-9. Apaf.1↑ caspases. Caspase-3. Bax↑etc. Cell death. (圖C:MPP+ 影響的蛋白質酵素路徑;摘錄自Eberhardt and Schulz, 2003)。. 過程需要JNK參與,並且JNK基因缺損的小鼠(JNK-deficient mice)可 對紫外線引發的細胞凋亡較具抗性,與其細胞內cytochrome-c 釋放及 caspase-3的活化都明顯被抑制有關(Tournier et al., 2000)。另外在腦 部局部缺血時也會刺激JNK活化,而使其直接與BimL結合,致使BimL 磷酸化,並引發下游Bax活化而使神經細胞凋亡(Okuno et al., 2004); 更有研究進一步證明在缺血-缺氧引發的神經死亡過程中以JNK3的活 化是重要的一個因子之一(Kuan et al., 2003)。. 17.
(18) 在帕金森氏症的研究中發現MPTP可以使腦黑質系統的JNK kinase(MKK4) 的表現量增加5倍,並使JNK的磷酸化增加2.5倍 (Nicotra and Parvez, 2002)。以JNK基因缺損剔除小鼠進行帕金森氏 症研究中發現,剔除JNK基因不但可以減少MPTP引發的神經死亡, 而且使得MPTP導致的多巴胺減少現象改善了39﹪(正常實驗鼠:減 少94﹪、JNK基因缺損剔除鼠:減少55﹪)(Hunot et al., 2004)。並 且,此神經毒素使JNK活化而導致的神經死亡可能與caspase-1及 caspase-3活化有關(Chun et al., 2001)。. PC-12細胞株 PC-12細胞株是大白鼠腎上腺髓質嗜鉻腫瘤細胞(rat adrenal medulla pheochromocytoma)所選出來的細胞株。PC-12細胞株經過神經 生長因子(nerve growth factor;NGF)刺激後會分化成類似交感神經 細胞的特性(Green et al., 1976)並具有分泌、儲存、合成及再吸收多 巴胺能力,所以,PC-12可視為一種多巴胺神經元,且具有多巴胺轉 +. 運器,因此常被用來進行MPP 的毒性研究(Nicotra and Parvez, 2000; Morioka et al., 2004)。. 18.
(19) 貳、研究目地 帕金森氏病、中風及阿滋海默氏症併列為老年人三大疾病,影響 國人健康甚大,雖然此病沒有立即性的生命危險,但是對於日常生活 中所造成的困擾及不便卻是常見且嚴重的。目前醫學研究上對於此疾 病亦付出頗大心力,希望能找出病因來預防,或是尋找治療的方針來 降低此病對人類的威脅。許多學者的研究發現,免疫反應對於此病有 相當的關聯性。在帕金森氏症病人死後的腦組織中發現,患者腦部的 TNF-α含量較同年齡非患者的腦組織 TNF-α含量來得高,甚至其他 +. 研究者發現 TNF-α基因缺損的老鼠似乎可以影響 MPP /MPTP 對神經 細胞產生的傷害。但是,縱使有許多實驗及研究結果證實 TNF-α與 帕金森氏症的神經胞死亡有關,卻缺乏證據顯示 TNF-α在帕金森氏 症影響神經細胞死亡中所扮演的角色及機制為何。所以,在此我們以 +. MPP 引發 PC-12 細胞毒性作為一帕金森氏症的實驗模式,並且加入 TNF-α來探討以下幾點: +. 1、TNF-α是否會增加或緩解 MPP 對細胞產生的毒性? +. 2、探討 TNF-α與 MPP 導致的細胞死亡是否經由細胞凋亡? +. 3、TNF-α與 MPP 的暴露引發的細胞內訊息傳遞分子為何?. 19.
(20) 參、實驗材料與方法 (壹)實驗材料 一、實驗藥品 1、methyl-4-phenyl-pyridinium (Cat No:D-048 ):購自 Sigma。 2、Tumor necrosis factor-alpha (TNF-α) (Cat No:T5944 ):購自 Sigma。 3、Cytochrome-c antibody (Cat No:6598A):購自 Pharmingen。 4、Poly rabbit anti-caspase-3 (Cat No:556425 ) : 購自 Cell Signaling。 5、SAPK/JNK assay kit (Cat No:9810) :購自 Cell Signaling。 6、10 x TG-SDS buffer (Cat No:0783-2L): 購自 Amresco。 7、10 x TG buffer (Cat No:0307-2L): 購自 Amresco。 8、其他一般藥品購自 Bio-Rad、Biochemika、Fluka、GlBCO、Jackson、 MERCK 及 R&D SYSTEMS 等公司。. (貳)細胞實驗 細胞來源:PC-12 cell line 購自食品工業發展研究所,細胞株編號 為 60048。 一、細胞培養 細胞培養於 10 ㎝培養皿內,使用 RPMI 1640 medium 培養(內 含 10﹪fetal bovine serum、5﹪hourse serum 及 penicillin-streptomycin),. 20.
(21) 細胞置於 5% CO2,90%相對溼度的 37℃培養箱內。當細胞約 9 分滿 時再做繼代培養。 二、繼代培養 當細胞成長至約 9 分滿時,收集細胞於離心管內在 4℃環境下以 700 rpm 離心 1 分鐘;抽掉上清液後,再行加入 3 ml 培養液,以 23 號針頭上下抽吸打散細胞,最後將細胞分配到 10 ㎝培養皿加入 10 ml 新鮮培養液培養。 三、細胞蛋白質抽取 所需試劑:lysis buffer 50 ml 成分如下: 150 mM 的 Tris(pH=7.9). 16.6 ml. NaCl. 0.44 g. 100 mM EGTA. 0.5 ml. Na3Vo4. 0.009 g. 1%的 NP-40. 0.5 ml. Sodium deoxychlate. 0.13 g. 二次水. 33.4 ml. 1 tablets proteinase inhibitor 細胞培養於 6 孔培養盤內,以藥物處理至特定時間後,收集其培 養液後,再以 PBS 沖洗並收集之,之後以 2500 rpm、4℃下離心 5 分. 21.
(22) 鐘,倒掉上清液;離心後沉澱的細胞(cell pellet)放置於 -80℃冰箱 至隔天,取出回溫後加入 20μl 的 lysis buffer 並以超音波震盪 15 秒 x 4 次,最後細胞以 15000 rpm、4℃條件離心 30 分鐘。收取上清液並置 於 -80℃環境下保存。 +. 四、加藥(MPP 及 TNF-α處理方式如下) : +. +. MPP 溶液需於每次加藥前再配置。將 MPP 秤好所需重量後依 據所需濃度溶解於培養液。TNF-α(100 ng/ml)則依照所需之濃度分 +. 別加入培養皿中培養至特定時間。若需合併 TNF-α與 MPP 處理時, 則依所需濃度均勻混合兩種藥物後,再加入於細胞培養皿中培養至特 定時間。 五、MTT 試驗-(MTT Assay) 4. 將 PC-12 細胞培養至 96 孔盤內,每個孔盤約有 1×10 個 PC-12 細 胞。再經藥物處理至特定時間後,各加入 100μl MTT (0.5 mg/ml) 後 放置於培養箱內 4 小時,之後加入 100μl 10% SDS 以溶解藍紫色的 formazan,再放回培養箱內放置至隔天。最後用 ELISA reader 以 570 nm 波長讀取其吸光值。細胞存活率=(實驗組 O.D ÷ 對照組 O.D)× 100﹪。計算結果均以One-Way ANOVA分析,再以LSD test 比較特定 組別(selected groups)之間存活率之差異。當p值小於0.05時則判定為 有意義之差異。. 22.
(23) 六、4',6-Diamidino-2-phenylindole (DAPI) Staining 經藥物處理過後的細胞取出上清液,以 PBS 清洗,再以 4% paraformadehyde 固定 30 分鐘,以 PBS 清洗完成後,加入 DAPI 染劑, 於室溫避光環境下反應 30 分鐘再以 PBS 清洗。封片後以雷射掃描共 軛焦顯微鏡(laser scanning confocal microscopy)觀察細胞核是否斷裂 為小核(apoptotic nuclei)以判別細胞凋亡。 七、TUNEL staining TUNEL staining 使用購自Calbiochem的TdT-FragEL DNA Fragmentation Detection Kit(Cat. No.:QIA33)。內含藥品如下: 1、Proteinase K : 2 mg/ml proteinase K in 10 mM Tris, pH 8。 2、5X TdT equilibration buffer 1 M sodium cacodylate, 0.15 M Tris, 1.5 mg/ml BSA, 3.75 mM CoCl2, pH 6.6。 3、TdT labeling reaction mix : a mixture of labeled and unlabeled deoxynucleotides at a ratio optimized for DNA fragment end labeling with TdT。 4、TdT enzyme: terminal deoxynucleotidyl transferase。 5、Stop buffer : 0.5 M EDTA, pH 8。 6、Blocking buffer : 4% BSA in PBS。 7、50X conjugate : Peroxidase streptavidin conjugate 50-fold concentrated. 23.
(24) solution。 ,. 8、DAB tablets : 3,3 Diaminobenzidine (0.7 mg/tablet)。 9、H2O2/urea tablets : H2O2/urea (1.6 mg/tablet)。 10、Methyl green counterstain 0.3% methyl green。 11、TBS : 50 mM Tris(pH=7.6)內含0.9% NaCl。 4. 本實驗每個樣本使用5×10 顆細胞,將細胞養在1.8平方公分的蓋 玻片上,蓋玻片先以poly-lysine處理後,並以紫外線燈滅菌後使用。 +. 細胞分別給予下列處理:30 ng/ml TNF-α、300μM MPP 、30 ng/ml +. TNF-α合併300μM MPP 、10μM SP600125前處理15分鐘再給予30 +. ng/ml TNF-α合併 300μM MPP 及100μM Z-VAD前處理30分鐘後再 +. 給予30 ng/ml TNF-α合併300μM MPP 。以上各組細胞經藥物處理24 小時後,以PBS清洗再以4% paraformadehyde固定,之後蓋玻片以TBS 浸泡15分鐘,再以20μg/ml proteinase K作用10分鐘,然後經TBS清洗 後,再將細胞玻片浸在3% H2O2 5分鐘,再以TBS清洗玻片,並將細胞 浸泡在1倍的TdT Equilibration Buffer 20分鐘後,以TBS清洗後,再加入 60μl的TdT Labeling Reaction Mixture,小心蓋覆parafilm在樣本上,在 37℃環境下靜置反應90分鐘。反應後以TBS清洗細胞,每個細胞玻片 再加入100μl的stop solution,並靜置5分鐘。以TBS清洗後,再加入 100μl的blocking buffer且靜置10分鐘,之後以拭鏡紙將蓋玻片附近的. 24.
(25) blocking buffer吸乾,加入1倍的conjugate,在潮濕的環境下靜置30分 鐘。經TBS清洗後,玻片以1倍的DAB溶液(1 tablet/1ml 自來水)染 色細胞10分鐘。清洗後,再以methyl green作為對比染色。染色完畢的 細胞玻片以100﹪酒精及xylene浸漬後以Histomount將蓋玻片做封片處 理。將封片完成的玻片在400倍顯微鏡觀察計數細胞凋亡比例。各個 藥物處理組分別以隨機方式至少計數200~300顆以上細胞。細胞死亡 率計算如下:death ratio(﹪)=TUNEL positive cells ÷ total cells × 100﹪。計數結果均以One-Way ANOVA分析,再以LSD test 比較特定 組別(selected groups)之間存活率之差異。當p值小於0.05時則判定為 有意義之差異。 八、蛋白質定量 以 BSA ( bovine serum albumin ) 配 置 不 同 標 準 溶 液 ( BSA standard),其濃度如下:0.05 mg/ml;0.1 mg/ml;0.2 mg/ml;0.4 mg/ml 及 0.6 mg/ml。將細胞蛋白質抽取液做適當稀釋 (6 孔培養盤約 10 到 15 倍稀釋、10 cm 培養盤約 70 倍稀釋)。各取 BSA standard 及細胞蛋 白質抽取液 5μl 加入 96 孔盤內(各做二重複),最後加入 195μl 蛋 白質定量染劑 (Bio-Rad protein assay Dye Reagent Concentrate;Cat No. 500-0006),反應 10 分鐘後使用 ELISA reader 偵測 BSA standard 及蛋白 質樣本在 570 nm 波長下之吸光值。再將 BSA standard 之吸光值及濃度. 25.
(26) 之關係做出一標準曲線(standard curve)後,以 standard curve 推算出 各別細胞蛋白質樣本濃度。 九、西方轉漬法(Western Blot) SDS PAGE 電泳試劑成分如下: (1)10﹪Separating Gel 5.0225 ml 成分如下: 1.5 mM Tris-HCl (pH=8.8). 1.25 ml. 30﹪Acrylamide&0.8﹪Bis-acrylamid. 1.67 ml. 10﹪SDS. 0.05 ml. 10﹪ammonium persulfate. 0.05 ml. TEMED. 0.002.5 ml. 二次水. 2 ml. (2)Stacking Gel 成分如下: 1 M Tris-HCl (pH=6.8). 0.1875 ml. 30﹪Acrylamide&0.8﹪Bis-acrylamid. 0.25 ml. 10﹪SDS. 0.015 ml. 10﹪ammonium persulfate. 0.015 ml. TEMED. 0.0015 ml. 二次水. 1.01 ml. (3)4 倍 sample buffer 5 ml 成分如下:. 26.
(27) 1 M 的 Tris-HCl(pH=6.8). 1.25 ml. SDS. 0.4 g. bromophemol blue. 0.001 g. Glycerol. 1.5 ml. DTT. 0.38575 g. 二次水. 2.25 ml. Western Blot 西方轉漬法試劑如下 (1)10 mM PBS 1000 ml 成分如下: 0.5 MPB. 20 ml. NaCl. 9g. 二次水. 980 ml. (2)PBST:PBS 內含 0.05﹪Tween-20。 (3)Blocking buffer:PBST 內含 5﹪脫脂奶粉。 收集的蛋白質樣本以 SDS-PAGE 分離 (120V,1.5 hr) 後再將膠 片上的蛋白質轉漬至 PVDF 轉漬膜上。轉漬完畢的 PVDF 用 PBST 洗 3 分鐘 × 3 次後再用 blocking buffer 在室溫下震盪 1 小時,之後再用 PBST 洗 3 分鐘 × 3 次。加入一級抗體 (primary antibody)後置於 4℃冰箱震盪至隔天。將一級抗體去除後,用 PBST 洗 3 分鐘 × 3 次 再加入二級抗體 (secondary antibody) 在室溫下震盪 1 小時,再用. 27.
(28) PBS 洗 5 分鐘 × 6 次。之後把轉漬膜放到 ECL 溶液中反應 1 分鐘。 再將轉漬膜取出,以 X 光底片進行壓片,所得底片結果以數位相機 照相,並以電腦軟體(KODAK 1D3.6)分析量化。 十、JNK 活性分析(JNK activity assay) SAPK/JNK assay kit (Cat No:9810) :購自 Cell Signaling。內含 溶液成份如下: 1、. Phospho-c-Jun (Ser63) Antibody 100 μl (10 Western mini-blots)。. 2、. Kinase Buffer (10X)(Cat No:9802)15 ml。. 3、. Cell Lysis Buffer (10X)(Cat No:9803)15 ml。. 4、. ATP (10 mM)(Cat No:9804)50μl。. 5、. c-Jun Fusion Protein Beads (Cat No:9811) 800μl。. 6、. Anti-rabbit IgG, HRP-linked Antibody(cat No:7074)50μl。 細胞以藥物處理至特定時間後,收集培養液。並再以 PBS 收下. 細胞後,以 2500 rpm、4℃環境下離心 5 分鐘,離心後倒掉上清液, 以 PBS 清洗 2 次,加入 500μl cell lysis buffer,並存放冰上 5 分鐘, 把細胞以超音波震盪後再以微量離心機在 16000 rpm、4℃環境下離心 10 分鐘,取上清液,並定量蛋白質。 取 200μg 蛋白質加 20μl c-Jun fusion protein beads,混合均勻,置 於 4℃至隔天。以為量離心機 16000 rpm、4℃條件下離心 30 秒。倒掉. 28.
(29) 上清液,並以 SAPK/JNK assay kit 中的 cell lysis buffer 及 kinase buffer 各清洗兩次、每次清洗之間需以 16000 rpm、4℃條件離心 30 秒。最 後加入 45μl kinase buffer (內含 0.2 mM ATP) ,並置於 30℃水浴槽內 30 分鐘。加入 15μl 4 倍的 sample buffer. (內含 200 mM DTT),震盪. 後離心 30 秒。之後將樣本在 95℃中煮 4 分鐘,取 20μl 跑 SDS PAGE。 十一、統計分析 所有結果以 Mean ± S.D. (平均值 ± 標準誤) 來表示,並採用 One-Way ANOVA 統計法分析,再以 LSD 比較特定組別(selected groups)之間的差異性,當 p 值小於 0.05 時則判定為有意義之差異。. 29.
(30) 肆、結果 +. +. 實驗一、評估 MPP 、TNF-α或 MPP 合併 TNF-α對 PC-12 的作用 細胞凋亡的過程在形態學上具有一些特徵,例如:細胞萎縮、細 胞質濃縮、染色質凝聚並黏附在核膜形成隆起和出現凋亡小體;細胞 內的粒線體、高爾基體沒有腫脹現象;在相當長一段時間內,核膜保 持完整。這些細胞形態上的改變均可利用相位差顯微鏡(phase contrast microscope)的技術來觀察。 +. 圖一實驗使用未分化的 PC-12 細胞,並分別予以 150μM MPP 、 +. 30 ng/ml TNF-α 及 150μM MPP 合併 30 ng/ml TNF-α,經處理 24 小 時後,細胞以 400 倍相位差顯微鏡觀察細胞形態上改變。 由圖一實驗結果發現未經藥物處理組別(對照組)中部分 PC-12 細胞可觀察到神經軸生長(圖 A 白色箭號所指處) ,當單獨給予 TNF-α30 ng/ml 24 小時後,細胞在形態上與對照組比較沒有明顯差 異,且細胞仍然具有神經軸生長能力(圖 B) 。但是當給予 +. 150μM MPP 處理後,細胞開始出現細胞凋亡的特徵,例如:皺縮 (shrinkage) 、神經軸退化(neurite retraction)等(圖 C 黑色箭號所指 處) 。而且當合併加入 30 ng/ml TNF-α 處理時,細胞死亡的現象更是 明顯(圖 D)。 +. 為探討不同時間點 MPP 與 TNF-α 藥物處理對細胞形態的影. 30.
(31) +. 響。圖二實驗中分別給予細胞 300μM MPP 、30 ng/ml TNF-α 或 +. 150μM MPP 合併 30 ng/ml TNF-α 處理至不同時間點(1、2、6、12 及 24 小時) 。圖二實驗結果發現對照組在 1 小時(C1;圖 A)及 24 小時(C24;圖 C)細胞外觀成圓形、完整且部分細胞有神經軸的生 長表現(圖 A 及圖 C 白色箭號所指處) 。當單獨處理 30 ng/ml TNF-α 時細胞在各個時間點(TNF-α 處理 1 小時,T1,圖 B;TNF-α 處理 24 小時,T24,圖 D)其形態上與對照組比較沒有明顯差異。但是當 予以 300μM MPP. +. 處理 6 小時後(M6,圖 I) ,細胞開始出現皺縮、. 貼附性差或是漂浮的細胞碎片。且此死亡現象隨著處理時間增加而更 明顯。但是,當 300μM MPP. +. 合併加入 30 ng/ml TNF-α 處理時,細. 胞自 2 小時即開始出現上述特徵(TM2,圖 H),且此現象隨著時間 增加而增加。 由圖一、圖二中的結果顯示,在時間點上不但 TNF-α 與 MPP. +. 合併處理會引起較多細胞死亡,且細胞死亡現象隨著處理時間增加而 增加。. +. +. 實驗二、評估 MPP 、TNF-α或 MPP 合併 TNF-α處理對 PC-12 的存 活率(cell viability)影響 +. 為更進一步證實 MPP 合併 TNF-α同時處理能引起細胞死亡,在. 31.
(32) 此使用 MTT Assay 來評估細胞存活率。MTT Assay 被廣泛應用於細胞 存活率的檢測。MTT(3-(4,5dimethyl- thiazol- 2-yl) - 2,5-dip-henyltetrazolium bro- mide)是一種水溶性且黃色的物質,可以被粒線體 內的脫氫酶 (dehydrogenase)還原生成結晶狀的藍紫色產 formazan。 Formazan 以 10% SDS 溶解後,可利用 enzyme-link immunosorbent assay (ELISA)reader 在波長 570 nm 下測其吸光值並分析之。因為只有活 細胞的粒線體內才有脫氫酶,能將 MTT 還原成結晶狀的藍紫色物, 故所測得的吸光值與細胞存活率成正比。因此若吸光值愈大,則表示 細胞存活率愈高。所以我們可利用吸光值評估細胞存活率。 本實驗使用未分化的 PC-12 細胞來進行下列實驗,細胞培養在 96 +. 孔盤中且分為不同藥物處理組別: (1). 600μM MPP ; (2). 300μM +. +. +. MPP ; (3). 150μM MPP ; (4). 75μM MPP ; (5). 30 ng/ml +. +. TNF-α合併 600μM MPP ;(6). 30ng/ml TNF-α合併 300μM MPP ; +. (7). 30 ng/ml TNF-α合併 150μM MPP ; (8). 30 ng/ml TNF-α合併 +. +. 75μM MPP ; (9). 15 ng/ml TNF-α合併 600μM MPP ; (10) . 15 +. ng/ml TNF-α合併 300μM MPP ; (11). 15 ng/ml TNF-α合併 150μM +. +. MPP ; (12). 15 ng/ml TNF-α合併 75μM MPP ; (13). 30 ng/ml TNF-α;(14). 15 ng/ml TNF-α;(15). control。以上各組細胞經處 理 24 小時後細胞以 MTT Assay 評估細胞存活率。. 32.
(33) 圖三的實驗結果顯示,當分別給予 TNF-α 0、15 或 30 ng/ml 時, 並不影響細胞存活率(0 ng/ml TNF-α,100% ± 0.31;15 ng/ml TNF-α,95.12% ± 2.96;30 ng/ml TNF-α,99.75% ± 4.48) 。但是 MPP. +. +. 處理的細胞中,當濃度愈高時細胞的存活率就越低(75μM MPP , +. 93.09% ± 4.37;600μM MPP ,73.98% ± 2.33)。而且當合併加入 TNF-α15 或 30 ng/ml 時,細胞存活率更是明顯下降。例如:當細胞 給予 75μM MPP 處理 24 小時後,細胞存活率與對照組相較是沒有差 +. 異的(75μM MPP+,93.09% ± 4.37;對照組,100﹪± 0.31) 但是, 當 75μM MPP+ 合併 TNF-α 不論是 15 ng/ml 或是 30 ng/ml,細胞存活 率與分別與單一藥物處理組相較則呈現有統計意義的下降(圖三)。 而且二樣藥物合併處理所引發之細胞死亡率傾向大於此二種藥物分 別處理時引發的死亡率的總合。表示 MPP+ 對細胞的毒性因 TNF-α 的 存在而增加。 本實驗結果係以 mean ± S.D.來表示,以對照組當作 100﹪後以 One-Way ANOVA 分析其差異性,當 p 值小於 0.05 時則判定為有統計 意義之差異(每次實驗 3 重複,n =3) 。. 實驗三、利用 DAPI (4',6-diamidino-2-phenylindole)染色觀察藥物處 理是否引起細胞凋亡. 33.
(34) 4,6-聯脒-2-苯基吲哚(4',6-diamidino-2-phenylindole ;DAPI)是 常用的DNA特異性染料之一。它是一種半通透性染料,用於常規固定 細胞的染色。DAPI主要結合在DNA的A-T鹼基區,以螢光顯微鏡或共 軛焦顯微鏡用紫外光激發即可見明亮的藍色螢光。DAPI 染色是以細 胞核的形態改變作為評估細胞凋亡的指標。細胞凋亡時的特徵為細胞 核濃縮(condensation)、DNA斷裂 (DNA fragmentation) 及細胞皺縮 (shrinkage) ,若是有此現象可以做為細胞凋亡指標 (Schapira, 1999; Kim et al., 2003)。 +. +. 在前面實驗結果顯示 MPP 或 MPP 合併 TNF-α 處理可引發 +. PC-12 細胞死亡。並有文獻顯示 MPP 處理會引發細胞凋亡(Nicotra +. +. and Parvez, 2003) 。在此我們欲進一步探討 MPP 及 MPP 合併 TNF-α 處理引起的細胞死亡是否屬於細胞凋亡。之前實驗結果(圖 +. +. 一)顯示 150μM MPP 或 150μM MPP 合併 30 ng/ml TNF-α 處理 即可明顯引發 PC-12 細胞死亡。因此,本實驗將 PC-12 細胞培養在 +. 1.8 平方公分的蓋玻片上,並分別給予 150μM MPP 、30 ng/ml +. TNF-α、150μM MPP 合併 30 ng/ml TNF-α 處理 24 小時後,以 DAPI 染細胞核後並以雷射掃描共軛焦顯微鏡(laser scanning confocal microscopy)觀察細胞凋亡的情形。 實驗結果在圖四中發現在對照組或是 30 ng/mlTNF-α 處理的細. 34.
(35) 胞,細胞核完整、沒有核濃縮或裂解(apoptotic nuclei)的情形。在 +. MPP 處理的細胞,細胞出現凋亡的特徵,例如:細胞核斷裂及核 +. 濃縮。而 TNF-α 合併 MPP 處理後,細胞核斷裂或是濃縮的情形相 對增加。可知,本實驗中藥物處理可引發細胞凋亡。. 實驗四、以西方墨點法分析藥物處理引發 PC-12 細胞死亡的相關蛋白 質表現。 在MPTP的帕金森氏症的實驗模式中發現MPTP處理導致的細胞 死亡可能與凋亡的路徑有關,其相關蛋白質有cytochrome-c及caspase-3 +. (Fahn and Sulzer., 2004) 。另外也有其他學者發現1 mM的MPP 處理72 小時可引發SH-SY5Y細胞凋亡,且導致caspase-3的活化及JNK磷酸化 (Chen et al., 2003),另外,在其他神經細胞株例如:MN9D,也發現 +. 100μM的MPP 處理12小時也可引發細胞死亡,且可藉由抑制caspases 來緩解細胞死亡的比例(Choi et al., 1999)。在PC-12細胞也有研究發 +. 現經由MPP 處理後可降低細胞存活率,且引起caspase-3的活化(Park et al., 2003;Sheng et al., 2002;Viswanath et al., 2001;Chuenkova et al., +. 2003) 。除此之外,MPP 或MPTP也被證實會引起JNK磷酸化 (Hunot et al., 2004;Eberhardt and Schulz, 2003;Nicotra and Parvez, 2003) ,且有 +. 學者認為MPP 處理導致的神經細胞死亡可能經由JNK路徑而誘發. 35.
(36) +. caspase-3活化而致(Chun et al., 2001)。因此本實驗旨在探討MPP 或 +. MPP 合併TNF-α處理導致的PC-12細胞凋亡是否也與JNK及caspase-3 的路徑有關。 5. 實驗中將PC-12細胞培養在6孔盤培養皿內,每孔盤約有7×10 個細 +. +. 胞,並分別給予300μM MPP 、30 ng/ml TNF-α、300μM MPP 合併 30 ng/ml TNF-α處理至特定時間後,分別收取細胞蛋白質,再以西方 墨點法分析caspase-3、JNK及cytochrome-c 等相關路徑的蛋白質表現量 及JNK的活性分析。 +. 我們先測試MPP 是否能引發cytochrome-c 釋放,結果發現MPP. +. 不論是150或是300μM在處理4小時後即可引起cytochrome-c 釋放,且 +. 具有時間相依性的特徵(圖五) 。Viswanath等人曾證實MPP 處理引起 cytochrome-c 釋放之後,將導致caspase的活化(Viswanath et al., 2001) 。 +. +. 因此,我們便進一步探討MPP 及MPP 合併TNF-α處理導致的細胞 死亡是否會經由caspase的活化。在caspase家族中,caspase-3屬於細胞 凋亡的執行者(executioner) ,而且被認為是在帕金森氏症中導致神經 退化的主要因子(Viswanath et al., 2001) 。實驗結果如圖六所見,MPP +. 處理的確可引起caspase-3的活化,然而MPP 合併TNF-α處理所導致 +. 活化的caspase-3的表現量均高於單獨給予MPP 處理的組別。有文獻 +. 指出在MPP 處理後導致的神經細胞死亡機制與JNK活化caspase-3有. 36. +.
(37) +. 關(Chun et al., 2001) 。接著,我們欲探知MPP 合併TNF-α處理引起 的JNK磷酸化是否與此一凋亡現象有關?由四次實驗結果中觀察到 +. 不同時間點的TNF-α或MPP 單獨處理時,與對照組相較並不誘發 +. JNK磷酸化。但是,在TNF-α合併MPP 處理1小時後,則可觀察到磷 酸化的JNK表現量增加(TM1) ,而此現象在2、6、12及24小時均可觀 察到(TM2、TM6、TM12及TM24) 。然而,JNK磷酸化一旦被誘發後, 其表現量並不因為處理時間的增長而增加。 為進一步證實JNK的磷酸化後是否經由改變細胞內的c-Jun的活 性來影響細胞的存活,我們以西方墨點法,在實驗結果中(圖八)發 +. +. 現不論是單獨給予MPP 、TNF-α處理或是MPP 合併TNF-α處理 時,c-Jun的磷酸化卻不見增加(n=4) 。由圖七及圖八的實驗結果顯示, 或許圖七中觀察到的JNK的磷酸化並不是作用於改變細胞內的c-Jun 的活性。為進一步確認圖七所看到的結果,我們接著以kinase assay的 +. 方式來探討給予MPP 合併TNF-α處理後JNK活性的改變。結果如圖 +. 九所見,MPP 合併TNF-α處理時引發了JNK活性的增加,這與圖七 JNK磷酸化的結果相符。由圖七圖、八及圖九的實驗中,我們推測MPP. +. 合併TNF-α處理時可增加JNK的磷酸化及其活性,但此一活化的JNK 並非直接促使c-Jun的磷酸化,而是可能作用於其他下游的受質 (substrates)。. 37.
(38) +. 實驗五、評估 JNK 的活化與 caspase-3 的活化在 MPP 合併 TNF-α 引 發細胞凋亡之角色。 +. 經由以上實驗得知 TNF-α合併 MPP 處理導致細胞死亡且引發 了 caspase-3、JNK 磷酸化及 JNK 活性增加。因此本實驗使用 SP600125 (JNK 抑制劑) 及 Z-VAD (caspase 抑制劑)來觀察抑制 JNK 及 +. caspase-3 是否能緩解 TNF-α 合併 MPP 處理導致的細胞死亡。圖十 +. +. 實驗結果發現 MPP (圖 D)或是 300μM MPP 合併 30 ng/ml TNF-α 處理(圖 E)可導致 PC-12 細胞產生如:皺縮、神經軸退化且細胞呈 現懸浮或是貼附性差等現象。然而當細胞給予 100μM Z-VAD 前處理 +. 30 分鐘後,再給予 300μM MPP 合併 30 ng/ml TNF-α,細胞死亡的 +. 現象可以明顯的被緩解(圖 G) ,但是對於 300μM MPP 合併 30 ng/ml TNF-α 造成細胞懸浮或是貼附性差(poor adhesion)的情形卻無法改 善(圖 G)。當細胞給予 10μM SP600125 前處理 15 分鐘後再給予 +. 300μM MPP 合併 30 ng/ml TNF-α 處理,細胞死亡的現象則無明顯 改善,細胞仍然出現皺縮、神經軸退化、懸浮或是貼附性差(poor adhesion)等特徵(圖 F) 。實驗結果證明抑制 caspase 可以減少 300μM +. MPP 合併 30 ng/ml TNF-α 處理後引發的細胞死亡,但是抑制 JNK 對 +. 於 300μM MPP 合併 30 ng/ml TNF-α 處理而引發的細胞死亡則無明 顯的改善。. 38.
(39) 我們接著採用TUNEL assay來進一步確認圖十一得到的結果。細 胞凋亡中染色體DNA會形成斷裂,DNA雙鏈斷裂或只要一條鏈斷裂並 ,. 出現出現缺口而產生的一系列DNA的3 -OH末端,3'-OH末端可在去氧 核糖核苷酸末端轉移酶(TdT)的作用下,將去氧核糖核苷酸和螢光 素、過氧化物酶、鹼性磷酸化酶或生物素形成的衍生物標記到DNA 的3'-OH末端,從而可進行凋亡細胞的檢測,這類方法一般稱為去氧 核糖核苷酸末端轉移酶介導的缺口末端標記法(TUNEL)。由於正常 的或正在增殖的細胞幾乎沒有DNA的斷裂,因而沒有3'-OH形成,很 少能夠被染色。TUNEL檢測法,能對完整的單個凋亡細胞核或凋亡小 體進行染色,並能準確地反應細胞凋亡典型的形態特徵,即使極少量 的凋亡细胞亦能被檢測出。 圖十一中,細胞以藥物處理至 24 小時後以 TUNEL 染色法分析細 +. 胞凋亡情形。自實驗結果顯示給予 300μM MPP 單獨處理(圖 C) +. 或是 30 ng/ml TNF-α合併 300μM MPP 處理(圖 D)可導致 PC-12 細胞凋亡。然而當細胞給予 Z-VAD(100μM)前處理 30 分鐘後再給 +. 予 30 ng/ml TNF-α 合併 300μM MPP 處理(圖 F) ,細胞凋亡的比例 可以明顯的被緩解。圖十二則為此實驗量化結果。此實驗結果與圖十 相互應,證明抑制 caspase 可以有效緩解 30 ng/ml TNF-α合併 300μM +. MPP 處理而引發的細胞凋亡及形態上的改變,表示 caspase 在. 39.
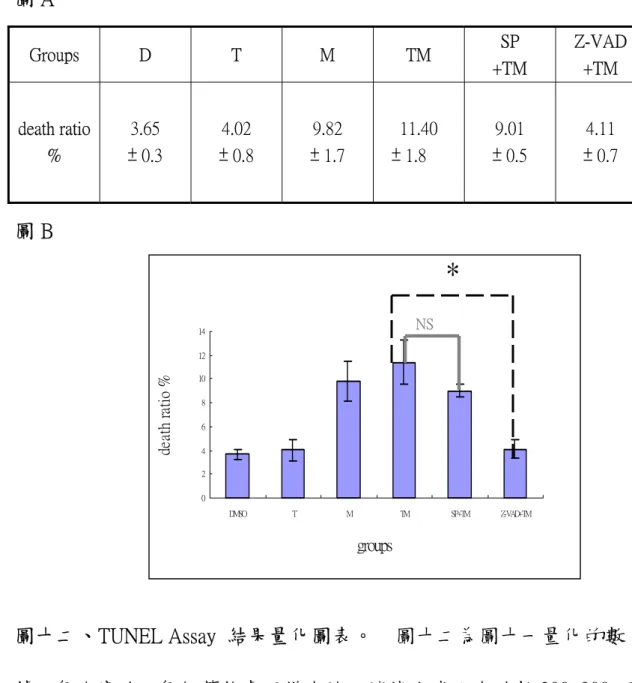
(40) +. TNF-α合併 MPP 處理產生的細胞死亡作用機制上扮演重要色。 +. TNF-α合併 MPP 可導致 PC-12 細胞凋亡,然而,當細胞給予 SP600125 (10μM)前處理 15 分鐘後再給予 30 ng/ml TNF-α合併 300μM +. MPP ,細胞死亡的現象雖有輕微緩解(TM:11.4% ± 1.8;SP + TM: 9.01% ± 0.5),但其效果不若抑制 caspase 作用有效 (TM:11.4% ± 1.8;Z-VAD + TM:4.11% ± 0.7)。實驗結果表示抑制 JNK 的活性對 +. 於 30 ng/ml TNF-α合併 300μM MPP 引發的細胞凋亡只有輕微抑制 的作用。. 40.
(41) 伍、討論 本研究主要探討在帕金森氏症的神經細胞退化過程中,免疫反應 對於神經細胞是否有保護作用或是會加促神經細胞的死亡。並探討細 胞因這些傷害(cytotoxicity)所引發的死亡機制為何。過去已有許多 研究發現 TNF-α或是干擾素-gamma(interferon-γ)及淋巴素-1 beta (interleukin-1β)等免疫反應與神經細胞死亡或是帕金森氏症有關 +. (Rousselet et al., 2002;Ferger et al., 2004)。因此本論文使用 MPP 引 發 PC-12 細胞的死亡,並加入 TNF-α,作為實驗模式,來研究帕金 森氏症與 TNF-α的關係。 本實驗分別以 MTT Assay、DAPI 及 TUNEL 觀察細胞存活率、細 胞形態以及細胞凋亡情形,並以西方墨點法分析細胞死亡的相關蛋白 +. 質表現。MTT Assay 與細胞形態上觀察到 MPP 的處理在 PC-12 上可 +. 引起細胞凋亡,且 TNF-α的存在可以加劇 MPP 的細胞毒性。由細胞 +. 形態上的分析發現,TNF-α合併 MPP 處理 2 小時後即可觀察到細胞 +. 凋亡的現象,然而 MPP 處理在 6 小時後才能看到細胞有凋亡的情 +. 形。這結果表示在 MPP 引發 PC-12 細胞的死亡過程中 TNF-α會加速 神經細胞的死亡。過去已有研究指出 TNF-α在神經細胞凋亡過程中 雖有著重要的影響(Soeda et al., 2001)另外,也有學者提出 TNF-α基 因缺損的剔除鼠可以減少 MPTP 引發多巴胺神經元的死亡(Ferger et. 41.
(42) +. al., 2004)。這與本實驗觀察到的結果一致認為 MPP 或是 MPTP 引發 的神經毒素中 TNF-α的存在會促使神經細胞的死亡。 +. 目前熟知MPP 可藉由改變細胞內酵素(enzyme)的功能而影響 細胞存活(Prezedborski and Vila, 2003) ,有研究者認為其中JNK為一重 要影響因子(Hunot et al., 2004)。在本論文中,我們以西方點墨法也 +. +. 同樣觀察到MPP 或MPP 合併TNF-α的處理可以引發JNK磷酸化及 +. caspase-3活化的現象,且MPP 合併TNF-α時其JNK磷酸化現象較MPP. +. 單獨處理時明顯。之前有文獻顯示,JNK磷酸化後可以促使c-Jun磷酸 化,c-Jun磷酸化與細胞的死亡有關(Li et al., 2004) ,因此,本實驗接 著探討JNK磷酸化是否會引起c-Jun磷酸化,我們同樣以西方點墨法去 偵測c-Jun的磷酸化。實驗結果發現c-Jun的磷酸化並不因為藥物處理而 +. 增加。由實驗結果推測MPP 合併TNF-α能引發JNK磷酸化,但是在 此,磷酸化的JNK並未去影響其下游的受質:c-Jun。雖然這可能與一 般JNK磷酸化的作用不相符合,但是亦有學者認為JNK的受質除了 c-Jun外尚有Elk1、ATF2(Sugden and Clerk, 1998)、Bim及Bcl-2家族 (Okuno et al., 2004)。其中c-Jun、Elk1及ATF2被認為與細胞轉錄 (transcription)有關,而c-Jun除了和細胞轉錄有關外,也同樣和Bim 及Bcl-2家族被認為與細胞凋亡及存活有關(Sugden and Clerk, 1998; Okuno et al., 2004; Waetzig and Herdegen., 2004;Chang and Karin.,. 42.
(43) 2001) 。有研究發現在腦部局部缺血時也會刺激JNK活化而促使中樞神 經細胞死亡,但是卻同樣未引起c-Jun的磷酸化,而是經由BimL去促 使Bax影響粒線體功能而導致凋亡(Okuno et al., 2004)。 +. +. 接著,我們進一步證明JNK或caspase-3在MPP 或MPP 合併 TNF-α處理引起細胞的死亡是否有直接關係。於是,實驗中使用了 JNK及caspase抑制劑,並從細胞形態評估細胞凋亡的情形,從細胞形 +. 態結果發現MPP 合併TNF-α處理會導致細胞死亡、神經軸萎縮或是 使細胞無法貼附、呈現漂浮的狀態。當給予caspase抑制劑後可以有效 +. +. 的抑制MPP 合併TNF-α處理導致細胞死亡,但是對於MPP 合併 TNF-α處理引起細胞漂浮的狀態卻無法明顯改善。然而在給予JNK抑 制劑處理的細胞組別,在西方墨點法雖顯示SP600125能有效減少JNK 磷酸化(data not shown)。但是從細胞形態上來看,卻無法有效緩解 細胞凋亡。於是實驗中進一步以TUNEL方式評估細胞凋亡的比例,從 TUNEL的結果中發現caspase抑制劑可以顯著減少細胞的死亡(圖十 +. 二)。而JNK抑制劑只能輕微緩解MPP 合併TNF-α引發的細胞凋亡。 這個實驗結果證明抑制caspase時可以有效的緩解細胞的死亡;但是抑 +. 制JNK只能輕微的減少細胞死亡,表示MPP 合併TNF-α處理導致細 胞死亡主要是與caspase-3的路徑有關(caspase-dependent)。 目前熟知的細胞凋亡路徑中,caspase-3是其中一個重要的細胞死. 43.
(44) 亡訊息。然而,caspase-3可以經由被許多蛋白質直接或是間接活化例 如:caspase-9(Viswanath et al.,2001)、caspase-8與10(Budihardjo et +. al.,1999)、JNK(Chun et al.,2001)等。在本論文中發現MPP 合併 TNF-α會導致細胞的死亡,且引起了 JNK 及 caspase-3 的活化,然而 在此實驗顯示 JNK 活化似乎與 caspase-3 活化的凋亡路徑沒有相關 聯。因此,在後續實驗裡將探討其他可能活化 caspase-3 的細胞訊息, +. 例如: caspase-9 及 caspase-8。如此才能確認 MPP 合併 TNF-α導致 的細胞死亡,是否其他的細胞訊息來活化 caspase-3 而產生細胞的死亡 訊息。. 44.
(45) 陸、參考文獻 1.. Ballif BA, Blenis J (2001) Molecular mechanisms mediating mammalian mitogen-activated protein kinase (MAPK) kinase (MEK)-MAPK cell survival signals. Cell Growth Differ 12:397-408.. 2.. Budihardjo I, Oliver H, Lutter M, Luo X, Wang X (1999) Biochemical pathways of caspase activation during apoptosis. Annu Rev Cell Dev Biol 15:269-290.. 3.. Cassarino DS, Halvorsen EM, Swerdlow RH, Abramova NN, Parker WD, Jr., Sturgill TW, Bennett JP, Jr. (2000) Interaction among mitochondria, mitogen-activated protein kinases, and nuclear factor-kappaB in cellular models of Parkinson's disease. J Neurochem 74:1384-1392.. 4.. Chang L, Karin M (2001) Mammalian MAP kinase signalling cascades. Nature 410:37-40.. 5.. Chen XC, Fang F, Zhu YG, Chen LM, Zhou YC, Chen Y (2003) Protective effect of ginsenoside Rg1 on MPP+-induced apoptosis in SHSY5Y cells. J Neural Transm 110:835-845.. 6.. Choi WS, Yoon SY, Oh TH, Choi EJ, O'Malley KL, Oh YJ (1999) Two distinct mechanisms are involved in 6-hydroxydopamine- and MPP+-induced dopaminergic neuronal cell death: role of caspases, ROS, and JNK. J Neurosci Res 57:86-94.. 7.. Chuenkova MV, Pereira MA (2003) PDNF, a human parasite-derived mimic of neurotrophic factors, prevents caspase activation, free radical formation, and death of dopaminergic cells exposed to the Parkinsonism-inducing neurotoxin MPP+. Brain Res Mol Brain Res 119:50-61.. 8.. Chun HS, Gibson GE, DeGiorgio LA, Zhang H, Kidd VJ, Son JH (2001) Dopaminergic cell death induced by MPP(+), oxidant and specific neurotoxicants shares the common molecular mechanism. J Neurochem 76:1010-1021.. 9.. Crocker SJ, Smith PD, Jackson-Lewis V, Lamba WR, Hayley SP, Grimm E, Callaghan SM, Slack RS, Melloni E, Przedborski S, Robertson GS, Anisman H, Merali Z, Park DS (2003) Inhibition of calpains prevents neuronal and behavioral deficits in an MPTP mouse model of Parkinson's disease. J Neurosci 23:4081-4091.. 45.
(46) 10. Czlonkowska A, Kurkowska-Jastrzebska I, Czlonkowski A, Peter D, Stefano GB (2002) Immune processes in the pathogenesis of Parkinson's disease - a potential role for microglia and nitric oxide. Med Sci Monit 8:RA165-177. 11. Eberhardt O, Schulz JB (2003) Apoptotic mechanisms and antiapoptotic therapy in the MPTP model of Parkinson's disease. Toxicol Lett 139:135-151. 12. Fahn S, Sulzer D (2004) Neurodegeneration and Neuroprotection in Parkinson Disease. Neurorx 1:139-154. 13. Ferger B, Leng A, Mura A, Hengerer B, Feldon J (2004) Genetic ablation of tumor necrosis factor-alpha (TNF-alpha) and pharmacological inhibition of TNF-synthesis attenuates MPTP toxicity in mouse striatum. J Neurochem 89:822-833. 14. Gao HM, Hong JS, Zhang W, Liu B (2002) Distinct role for microglia in rotenone-induced degeneration of dopaminergic neurons. J Neurosci 22:782-790. 15. Gao HM, Liu B, Zhang W, Hong JS (2003) Synergistic dopaminergic neurotoxicity of MPTP and inflammogen lipopolysaccharide: relevance to the etiology of Parkinson's disease. Faseb J 17:1957-1959. Epub 2003 Aug 1915. 16. Gelderblom M, Eminel S, Herdegen T, Waetzig V (2004) c-Jun N-terminal kinases (JNKs) and the cytoskeleton--functions beyond neurodegeneration. Int J Dev Neurosci 22:559-564. 17. Gomez-Santos C, Ferrer I, Reiriz J, Vinals F, Barrachina M, Ambrosio S (2002) MPP+ increases alpha-synuclein expression and ERK/MAP-kinase phosphorylation in human neuroblastoma SH-SY5Y cells. Brain Res 935:32-39. 18. Greene LA, Tischler AS. (1976) Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc Natl Acad Sci U S A.;73(7):2424-8. 19. Guyton AC. and Hall JE.蓋統生理學第六版,賴亮全,林則彬,林富美譯 華杏, 台北,2002。 20. Halvorsen EM, Dennis J, Keeney P, Sturgill TW, Tuttle JB, Bennett JB, Jr. (2002) Methylpyridinium (MPP(+))- and nerve growth factor-induced changes in pro- and. 46.
(47) anti-apoptotic signaling pathways in SH-SY5Y neuroblastoma cells. Brain Res 952:98-110. 21. Hayley S, Crocker SJ, Smith PD, Shree T, Jackson-Lewis V, Przedborski S, Mount M, Slack R, Anisman H, Park DS (2004) Regulation of dopaminergic loss by Fas in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson's disease. J Neurosci 24:2045-2053. 22. Russ H, Staust K, Martel F, Gliese M, and Schomig E (1996) The extraneuronal transporter for monoamine transmitters exists in cells derived from human central nervous system glia. Eur J Neurosci 8: 1256-64. 23. Hunot S, Vila M, Teismann P, Davis RJ, Hirsch EC, Przedborski S, Rakic P, Flavell RA (2004) JNK-mediated induction of cyclooxygenase 2 is required for neurodegeneration in a mouse model of Parkinson's disease. Proc Natl Acad Sci U S A 101:665-670. Epub 2004 Jan 2002. 24. Johnson GL, Lapadat R (2002) Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 298:1911-1912. 25. Kim EH, Jang MH, Shin MC, Shin MS, Kim CJ (2003) Protective effect of aqueous extract of Ginseng radix against 1-methyl-4-phenylpyridinium-induced apoptosis in PC12 cells. Biol Pharm Bull 26:1668-1673. 26. Kuan CY, Whitmarsh AJ, Yang DD, Liao G, Schloemer AJ, Dong C, Bao J, Banasiak KJ, Haddad GG, Flavell RA, Davis RJ, Rakic P (2003) A critical role of neural-specific JNK3 for ischemic apoptosis. Proc Natl Acad Sci U S A 100:15184-15189. Epub 12003 Dec 15181. 27. Leppa S, Saffrich R, Ansorge W, Bohmann D (1998) Differential regulation of c-Jun by ERK and JNK during PC12 cell differentiation. Embo J 17:4404-4413. 28. Li L, Feng Z, Porter AG (2004) JNK-dependent phosphorylation of c-Jun on serine 63 mediates nitric oxide-induced apoptosis of neuroblastoma cells. J Biol Chem 279:4058-4065. Epub 2003 Nov 4014. 29. Liu B, Hong JS (2003) Role of microglia in inflammation-mediated neurodegenerative diseases: mechanisms and strategies for therapeutic intervention. J. 47.
(48) Pharmacol Exp Ther 304:1-7. 30. Mathiasen JR, McKenna BA, Saporito MS, Ghadge GD, Roos RP, Holskin BP, Wu ZL, Trusko SP, Connors TC, Maroney AC, Thomas BA, Thomas JC, Bozyczko-Coyne D (2004) Inhibition of mixed lineage kinase 3 attenuates MPP+-induced neurotoxicity in SH-SY5Y cells. Brain Res 1003:86-97. 31. Mathiasen JR, McKenna BA, Saporito MS, Ghadge GD, Roos RP, Holskin BP, Wu ZL, Trusko SP, Connors TC, Maroney AC, Thomas BA, Thomas JC, Bozyczko-Coyne D (2004) Inhibition of mixed lineage kinase 3 attenuates MPP+-induced neurotoxicity in SH-SY5Y cells. Brain Res 1003:86-97. 32. McGeer PL, McGeer EG (2004) Inflammation and neurodegeneration in Parkinson's disease. Parkinsonism Relat Disord 10:S3-7. 33. Morioka N, Kumagai K, Morita K, Kitayama S, Dohi T (2004) Nonsteroidal anti-inflammatory drugs potentiate 1-methyl-4-phenylpyridinium (MPP+)-induced cell death by promoting the intracellular accumulation of MPP+ in PC12 cells. J Pharmacol Exp Ther 310:800-807. Epub 2004 May 2006. 34. Nicotra A, Parvez S (2002) Apoptotic molecules and MPTP-induced cell death. Neurotoxicol Teratol 24:599-605. 35. Nicotra A, Parvez SH (2000) Cell death induced by MPTP, a substrate for monoamine oxidase B. Toxicology 153:157-166. 36. Okuno S, Saito A, Hayashi T, Chan PH (2004) The c-Jun N-terminal protein kinase signaling pathway mediates Bax activation and subsequent neuronal apoptosis through interaction with Bim after transient focal cerebral ischemia. J Neurosci 24:7879-7887. 37. Park TH, Kwon OS, Park SY, Han ES, Lee CS (2003) N-methylated beta-carbolines protect PC12 cells from cytotoxic effect of MPP+ by attenuation of mitochondrial membrane permeability change. Neurosci Res 46:349-358. 38. Przedborski S, Vila M (2003) The 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model: a tool to explore the pathogenesis of Parkinson's disease. Ann N Y Acad Sci 991:189-198.. 48.
(49) 39. Rousselet E, Joubert C, Callebert J, Parain K, Tremblay L, G Orieux, Launay JM, Cohen-Salmon C, and Hirsch EC (2002) Behavioral changes are not directly related to striatal monoamine levels, number of nigral neurons, or dose of parkinsonian toxin MPTP in mice. Neurobiol Dis, 14: 218-28. 40. Rousselet E, Callebert J, Parain K, Joubert C, Hunot S, Hartmann A, Jacque C, Perez-Diaz F, Cohen-Salmon C, Launay JM, Hirsch EC (2002) Role of TNF-alpha receptors in mice intoxicated with the parkinsonian toxin MPTP. Exp Neurol 177:183-192. 41. Sairam K, Saravanan KS, Banerjee R, Mohanakumar KP (2003) Non-steroidal anti-inflammatory drug sodium salicylate, but not diclofenac or celecoxib, protects against 1-methyl-4-phenyl pyridinium-induced dopaminergic neurotoxicity in rats. Brain Res 966:245-252. 42. Saporito MS, Thomas BA, Scott RW (2000) MPTP activates c-Jun NH(2)-terminal kinase (JNK) and its upstream regulatory kinase MKK4 in nigrostriatal neurons in vivo. J Neurochem 75:1200-1208. 43. Schapira AH (1999) Science, medicine, and the future: Parkinson's disease. Bmj 318:311-314. 44. Schmidt N, Ferger B (2001) Neurochemical findings in the MPTP model of Parkinson's disease. J Neural Transm 108:1263-1282. 45. Sheng GQ, Zhang JR, Pu XP, Ma J, Li CL (2002) Protective effect of verbascoside on 1-methyl-4-phenylpyridinium ion-induced neurotoxicity in PC12 cells. Eur J Pharmacol 451:119-124. 46. Snell.臨床神經解剖學弟一版,劉亮廷譯,藝軒,台北,2003。 47. Soeda S, Ochiai T, Paopong L, Tanaka H, Shoyama Y, Shimeno H (2001) Crocin suppresses tumor necrosis factor-alpha-induced cell death of neuronally differentiated PC-12 cells. Life Sci 69:2887-2898. 48. Sriram K, Matheson JM, Benkovic SA, Miller DB, Luster MI, O'Callaghan JP (2002) Mice deficient in TNF receptors are protected against dopaminergic neurotoxicity: implications for Parkinson's disease. Faseb J 16:1474-1476. Epub 2002 Jul 1418.. 49.
(50) 49. Sugden PH, Clerk A (1998) "Stress-responsive" mitogen-activated protein kinases (c-Jun N-terminal kinases and p38 mitogen-activated protein kinases) in the myocardium. Circ Res 83:345-352. 50. Sun DS, Chang HH (2003) Differential regulation of JNK in caspase-3-mediated apoptosis of MPP(+)-treated primary cortical neurons. Cell Biol Int 27:769-777. 51. Teismann P, Tieu K, Choi DK, Wu DC, Naini A, Hunot S, Vila M, Jackson-Lewis V, Przedborski S (2003) Cyclooxygenase-2 is instrumental in Parkinson's disease neurodegeneration. Proc Natl Acad Sci U S A 100:5473-5478. Epub 2003 Apr 5417. 52. Tournier C, Hess P, Yang DD, Xu J, Turner TK, Nimnual A, Bar-Sagi D, Jones SN, Flavell RA, Davis RJ (2000) Requirement of JNK for stress-induced activation of the cytochrome c-mediated death pathway. Science 288:870-874. 53. Viswanath V, Wu Y, Boonplueang R, Chen S, Stevenson FF, Yantiri F, Yang L, Beal MF, Andersen JK (2001) Caspase-9 activation results in downstream caspase-8 activation and bid cleavage in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinson's disease. J Neurosci 21:9519-9528. 54. Waetzig V, Herdegen T (2004) Neurodegenerative and physiological actions of c-Jun N-terminal kinases in the mammalian brain. Neurosci Lett 361:64-67. 55. Xia XG, Harding T, Weller M, Bieneman A, Uney JB, Schulz JB (2001) Gene transfer of the JNK interacting protein-1 protects dopaminergic neurons in the MPTP model of Parkinson's disease. Proc Natl Acad Sci U S A 98:10433-10438. 56. Zhang W, Wang T, Qin L, Gao HM, Wilson B, Ali SF, Hong JS, Liu B (2004) Neuroprotective effect of dextromethorphan in the MPTP Parkinson's disease model: role of NADPH oxidase. Faseb J 18:589-591.. 50.
(51) A. C. C. M. B. D. TM. T +. +. 圖一、TNF-α 合併 MPP 處理可加重 MPP 的毒性。 PC-12 細胞培 養後分別予下列藥物處理:圖 A:對照組(C) ;圖 B:30 ng/ml TNF-α +. +. (T) ;圖 C:150μM MPP (M) ;圖 D:150μM MPP 合併 30 ng/ml TNF-α(TM),經處理 24 小時後,在 400 倍視野下以相位差顯微鏡 觀察細胞形態上改變。圖 A 對照組中白色箭頭所指處為神經軸。當 單獨給予 TNF-α 時細胞在形態上與對照組比較沒有明顯差異,白色 +. 箭號所指處為神經軸(圖 B) 。而 MPP 處理後,細胞開始出現皺縮、 +. 神經軸退化之現象(圖 C) 。當 MPP 合併加入 30 ng/ml TNF-α 處理 時,細胞死亡的現象更是明顯 (圖 D 黑色箭號所指處)。白色箭號 所指處為神經軸、黑色箭號所指處為貼附性差、死亡、皺縮的細胞或 是漂浮的細胞碎片(scale bar=10μM) 。. 51.
(52) 圖二、不同處理時間點引發細胞形態改變。 培養的 PC-12 細胞給予 +. +. 各種不同藥物(300μM MPP ,30 ng/ml TNF-α 或 300μM MPP 合 併 30 ng/ml TNF-α)及不同時間點(1,2,6,12 及 24 小時)處理後 以 400 倍相位差顯微鏡觀察細胞形態上改變。右圖中圖 A:對照組處 理 1 小時(C1);圖 B:TNF-α 處理 1 小時(T1);圖 C:對照組處 +. 理 24 小時(C24) ;圖 D:TNF-α 處理 24 小時(T24) ;圖 E:MPP 處 +. 理 1 小時(M1) ;圖 F:TNF-α 合併 MPP 處理 1 小時(TM1) ;圖 G: +. +. MPP 處理 2 小時(M2) ;圖 H:TNF-α 合併 MPP 處理 2 小時(TM2) ; +. +. 圖 I:MPP 處理 6 小時(M6) ;圖 J:TNF-α 合併 MPP 處理 6 小時 +. (TM6) ;圖 K:MPP 處理 12 小時(M12) ;圖 L:TNF-α 合併 MPP. +. +. 處理 12 小時(TM12) ;圖 M:MPP 處理 24 小時(M24) ;圖 N:TNF-α +. 合併 MPP 處理 24 小時(TM24)。白色箭號所指處為神經軸、黑色 箭號所指處為貼附性差、死亡、皺縮的細胞或是漂浮的細胞碎片(scale bar=10μM)。. 52.
(53) A. B. C1. T1. C. D. C24. T24. E. F. M1. TM1. G. M2. H. TM2. 53.
(54) I. J. M6. TM6. K. L. M12. TM12. M. N. M24. TM24. 54.
(55) MPP. 120. TNF-alpha 15ng/ml + MPP. & &. 100. *. # # de a t h ra t io. 80. TNF-alpha 30ng/ml + MPP. & &. # #. *. & &. # #. *. & &. # #. 60. 40. 20. 0 MPP 0 mM. MPP0.075 mM. MPP0.15 mM. MPP0.3 mM. MPP0.6 mM. groups. +. 圖三、TNF-α 可加劇 MPP 引發的細胞死亡。. 上圖顯示是以 MTT. Assay 來評估不同濃度的藥物處理 24 小時後對細胞存活率的影響。每 組數據均以對照組作為 100﹪後在以 One-Way ANOVA 分析,並以 LSD test 比較特定組別(selected groups)之間存活率之差異。所有數據均 以 mean ± S.D.來表示。當 p 值小於 0.05 時則判定為有統計意義之差 +. 異(每次實驗 3 重複,n =3) 。*:control vs. MPP ,p < 0.05;#:MPP +. +. 合併 TNF-α vs. 同濃度 MPP 單獨處理,p < 0.05;&:MPP 合併 TNF-α vs. 同濃度 TNF-α 單獨處理,p < 0.05。. 55. +.
(56) A. C. C. M. B. D. TM. T +. +. 圖四、MPP 及 TNF-α 合併 MPP 處理引發細胞凋亡。 細胞分別給 予以下藥物處理:圖 A:對照組(C) ;圖 B:30 ng/ml TNF-α(T) ; +. +. 圖 C:150μM MPP (M);圖 D:150μM MPP 合併 30 ng/ml TNF-α(TM),經處理 24 小時後,以 DAPI 染色並在雷射掃描共軛 焦顯微鏡(laser scanning confocal microscopy)觀察細胞凋亡的結果。 在對照組(圖 A)及 TNF-α 處理(圖 B)的細胞,細胞核完整、沒 +. 有核濃縮的情形。在 MPP 處理後(圖 C),細胞出現凋亡的特徵, +. +. 例如:細胞核斷裂及核濃縮。而在 MPP 及 TNF-α 合併 MPP 處理的 細胞 (圖 D) ,其細胞核斷裂或是濃縮的情況更是明顯。箭號所指處 為斷裂或是濃縮的細胞核。. 56.
(57) 4H. 8H. M300 M150. M300. M150. cytochrome-c. β-actin. +. 圖五、MPP 處理可引發 cytochrome-c 的釋放。. 上圖係以西方墨點. +. 法偵測 MPP 處理後 cytochrome-c 的表現量。實驗結果顯示 150μM +. MPP 可引發 cytochrome-c 的釋放,且 8 小時 cytochrome-c 的表現量多 +. 於 4 小時的表現量。在 300μM MPP 處理也看見同樣的趨勢。顯示 +. MPP 的處理可引發 cytochrome-c 的釋放,此一現象與 Viswanath 等人 +. 的實驗結果一致(Viswanath et al., 2001)。M300=300μM MPP ;M150 +. =150μM MPP ;4H=藥物處理 4 小時;8H=藥物處理 8 小時。. 57.
(58) C. T1. T2. T24. M2. M6. M12 M24 TM2 TM6 TM12 TM24. 32 kD Caspase-3. 17 kD Caspase-3 (cleaved form) β-actin. +. +. 圖六、MPP 及MPP 合併TNF-α處理可導致caspase-3活化。. PC-12. +. 細胞分別給予藥物處理(30 ng/ml TNF-α、300μM MPP 或是給予 +. 30 ng/ml TNF-α合併300μM MPP 或對照組)至特定時間點後,收取 細胞蛋白質,以西方墨點法偵測caspase-3的表現量。圖中cleaved caspase-3(17kD)代表活化的caspase-3。實驗顯示TNF-α單獨處理的 +. 細胞並未引起caspase-3活化。而MPP 處理的細胞在6小時後,活化的 caspase-3表現量即增加(M6),但此一增加的現象與處理時間的增長 +. 並無明顯關係(M12及M24)。而在TNF-α加MPP (TM)處理組別, +. 活化的caspase-3表現量高於MPP 單獨處理組別。但是,TM組的cleaved caspase-3的表現量也並未隨著處理時間增加而增加(n=3) 。C:對照 組處理24小時;T1:TNF-α處理1小時;T2:TNF-α處理2小時;T24: +. +. TNF-α處理24小時;M2:MPP 處理2小時;M6:MPP 處理6小時; +. +. M12:MPP 處理12小時;M24:MPP 處理24小時;TM2:TNF-α合 +. +. 併MPP 處理2小時;TM6:TNF-α合併MPP 處理6小時;TM12:TNF-α +. +. 合併MPP 處理12小時;TM24:TNF-α合併MPP 處理24小時。 58.
(59) C. T1. T24. M1 M2 M6 M12 M24. TM1 TM2 TM6 TM12 TM24. JNK. p-JNK. β-actin. +. +. 圖七、MPP 及MPP 合併TNF-α處理可導致JNK磷酸化。 PC-12細胞 +. 分別給予不同藥物處理(30 ng/ml TNF-α、300μM MPP 或是給予 +. 30 ng/ml TNF-α 合併 300μM MPP 或對照組)至不同時間點(1、2、 6、12、24 小時) 後收取細胞蛋白質,並且以西方墨點法偵測 JNK 及磷酸化 JNK(p-JNK)的表現量。圖中顯示 4 次實驗中的一次結果, JNK、p-JNK 與β-actin 的表現量。由實驗結果顯示,JNK 磷酸化的表 +. 現量,在 TNF-α合併 MPP 處理組別(TM1、TM2、TM6、TM12 及 TM24)有增加傾向(n=4) 。C:對照組處理 24 小時;T1:TNF-α 處 +. 理 1 小時;T24:TNF-α 處理 24 小時;M1:MPP 處理 1 小時;M2: +. +. +. MPP 處理 2 小時;M6:MPP 處理 6 小時;M12:MPP 處理 12 小時; +. +. M24:MPP 處理 24 小時;TM1:TNF-α 合併 MPP 處理 1 小時;TM2: +. +. TNF-α 合併 MPP 處理 2 小時;TM6:TNF-α 合併 MPP 處理 6 小時; +. +. TM12:TNF-α 合併 MPP 處理 12 小時;TM24:TNF-α 合併 MPP 處 理 24 小時。箭號所指處為 JNK 或 p-JNK 所在之分子量(54kD 及 46kD)。 59.
(60) C. T24. M1. M2. M6. M12. M24 TM1 TM2. TM6 TM12 TM24. P-C-Jun. β-actin. +. +. 圖八、MPP 及MPP 合併TNF-α處理並不導致c-Jun 磷酸化 (p-c-Jun) 的增加。. 培養的PC-12細胞分別給予不同藥物處理(30 ng/ml +. +. TNF-α、300μM MPP 或是給予30 ng/ml TNF-α合併300μM MPP 或 對照組)至不同時間點(1、2、6、12、24小時)後收取細胞蛋白質, 並且以西方墨點法偵測p-c-Jun的表現量。上圖為四次實驗的結果之 一,結果顯示p-c-Jun表現量並不因為各組藥物處理而增加(n=4) 。C: +. 對照組處理24小時;T24:TNF-α處理24小時;M1:MPP 處理1小時; +. +. +. M2:MPP 處理2小時;M6:MPP 處理6小時;M12:MPP 處理12小 +. +. 時;M24:MPP 處理24小時;TM1:TNF-α合併MPP 處理1小時;TM2: +. +. TNF-α合併MPP 處理2小時;TM6:TNF-α合併MPP 處理6小時; +. +. TM12:TNF-α合併MPP 處理12小時;TM24:TNF-α合併MPP 處理 24小時。. 60.
(61) C. T. M. TM. p-c-Jun. +. 圖九、MPP 合併 TNF-α 處理可增加 JNK 活性。 培養的 PC-12 細胞 +. 分別給予不同藥物處理 (30 ng/ml TNF-α、300μM MPP 或是給予 +. 30 ng/ml TNF-α 合併 300μM MPP 或對照組)至 24 小時後收取細胞 蛋白質,並以自 Cell Signal 購得之 Kinase Assay Kit 分析 p-c-Jun 表現 量用以評估 JNK 的活性。此圖為四次實驗的結果之一。結果顯示 +. p-c-Jun 表現量在 MPP 合併 TNF-α 處理的細胞有增加的趨勢(n=4) 。 顯示 JNK 活性因藥物合併處理而上升。C:對照組處理 24 小時;T: +. TNF-α 處理 24 小時;M:MPP 處理 24 小時;TM:TNF-α 合併 MPP 處理 24 小時。. 61. +.
數據
相關文件
如:parathion、malathion,易與 choline esterase 結 合,使失其作用,造成 acetylcholine 之大量積 存,引起神經興奮、死亡。可用藥 Atropine 以 斷 choline estease 和
“ smyang zhing 'phres pa dang dga' ba dang gnyid dang glal ba dang mi dga' ba dangbza' ba dang btung ba la chags pa dang sems rmugs..
Growth fac- tors that regulate local bone metabolism include growth hormone (GH), insulin-like growth factor-I (IGF-I), epider- mal growth factor (EGF) and interleukin-1 alpha
(C) Differential diagnosis point between central giant cell granuloma and aneurysmal bone cyst is at the indentification of sinsusoidal blood spaces within the tumor mass1. (D)
No risk factor for necrosis could be identified except the realization of dental extraction before treat- ment initiation (P = .025) (Table 2): at 36 months, the incidence of
The mean salivary level of alpha-amylase and the mean scores of state and trait anxiety in geographic tongue patients were higher than those of healthy controls.. But these
第八週 種子的萌發 種子萌發的機制 第九週 種子的萌發 種子萌發的機制 第十週 種子的萌發 種子萌發的機制 第十一週 種子收集 實際收集種子與觀察 第十二週
To evaluate Buffett’s pure stock-selection ability, we used Berkshire Hathaway’s 13F filings to collect the portfolio of publicly held companies and constructed a monthly time
